Author Archives: Albert Bordons

Yeasts 3000-years-old are alive and other histories of dormant cells
18th August 2019
Translated from the original article in Catalan
A few months ago -April 2019- my friend Jordi Diloli, Professor and Archaeologist, shared a very surprising article (Aouizerat et al 2019) with me. It was echoed on the internet (Borschel-Dan 2019), and I will comment here.
“Resurrected” yeasts from 3,000 years ago
The group of researchers led by Ronen Hazan of the Hebrew University of Jerusalem took samples of 21 clay containers from various sites in present-day Israel from 2500 to 5000 years ago, from the Persian, Philistine and Egyptian (this is the oldest) periods. Archaeologists believed that these vessels contained fermented beverages such as beer or mead (Figure 1). The authors submerged the containers in a rich YPD medium, specific for growing yeasts and other fungi, and incubated them at room temperature for 7 days. Then, samples of this medium were spread on agar plates with the specific medium, and the resulting colonies were isolated for subsequent analyses (Aouizerat et al 2019).

Figure 1. Clay vessels from where the yeasts were isolated (Image of Judah Ari Gross, Times of Israel).
The isolates that were found were 6 strains of different yeast species, and one of which was Saccharomyces cerevisiae, specifically from a Philistine site dated 3000 years ago. Obviously, it is very surprising that living yeasts of such ancient remains have been isolated. For this reason, the authors of the work carried out a series of experiments that could confirm this unique fact and that the isolates were not a product of contamination.
Firstly Aouizerat et al (2019) showed that it is possible to isolate yeasts from clay vessels that have contained beer or wine after a certain time. They did so with containers with unfiltered beer buried for 3 weeks, and also with another vessel that had repeatedly contained wine but not used last 2 years. With these samples they developed the isolation methodology and in both cases they were able to isolate yeasts. No isolates were obtained from a control sample with filtered beer, therefore without yeasts.
To demonstrate that the isolates were originals of the old vessels because these had contained the fermented beverage, authors applied the same protocol with samples of other ceramics that were surely not for this purpose, and also with sediments near the containers. The result was clearly negative for these samples: only 2 isolated yeasts from 110 samples, while the mentioned 6 yeast strains were isolated from the 21 initial samples. That is, yeasts would be significantly more abundant in containers of alcoholic fermented beverages than in other related archaeological vessels or sediments around them.
Another argument that supports the hypothesis of this work was the identification of these 6 yeasts. Total DNA was obtained and processed to sequence the genomes and compare them with the databases. Two of them, from the Egyptian period, were identified as Saccharomyces delphensis, a species that has been isolated from African dried figs and is not at all common on soil. Therefore, this suggests the use of figs in the alcoholic beverages of these containers. Another isolate was identified as Rhodotorula, common pollutant yeast in African beers. Another was identified as Debaryomyces, a frequent yeast in traditional African sorghum beers. As said before, another isolate was identified as Saccharomyces cerevisiae, the yeast most used to make wine, beer or bread (Figure 2). In spite of this, the genetic sequence of this S. cerevisiae was clearly different from the strains most commonly used today, as commercial or laboratory strains, and therefore the possibility of contamination is excluded. And finally, the other isolate was identified as Hypopichia burtonii, previously isolated yeast from Ethiopian mead.
These genetic data, together with the phenotypic characterization -fermentative kinetics and other biochemical characteristics carried out with the isolates by Aouizerat et al (2019)- suggest that these yeasts actually come from an environment related to alcoholic beverages. These authors even elaborated beer with these isolates and some of them, especially the Saccharomyces, gave a very good analytical and sensory result.

Figure 2. Saccharomyces cerevisiae at the scanning electronic microscope (MD Murtey & P Ramasamy)
Aouizerat et al (2019) conclude that the isolates are descendants of yeasts that were originally used 3000 years ago, in large quantities and in repeated fermentations. This would have facilitated their survival in pore microenvironments of the ceramic matrix of these containers, and the microcolonies would have continued to grow minimally for millennia thanks to the humidity and residual nutrients. The authors make the analogy with some handmade beers where it is usual that the containers waste serve as starter for new productions.
Finally, the authors of this work speculate that it is possible to isolate microorganisms from archaeological remains, not only yeasts, and that in the case of bacteria it could even be easier, given the resistance characteristics of some of them, such as the sporulated ones.
Is there no previous similar work to that of Aouizerat et al (2019) ?
As we have seen, this is certainly a very surprising finding. Scientifically, the work is quite accurate and has been “approved” by the international community: the article is published in an open-access journal with prestige (mBio, high impact factor: 6.7), of the American Society for Microbiology, where all the articles are reviewed by a minimum of two experts, besides the editors. The results presented by the article seem very well worked, and the conclusions are well reasoned.
However, in my opinion it is still almost incredible, and it is strange that nothing like this has been found before. Maybe if someone else had previously tried to isolate such old microorganisms without getting them, perhaps it would not have been published ? Maybe nobody has previously tried to do something similar ? A “malicious” explanation might be that archaeologists have their own interests and microbiologists or molecular biologists have others, and that for this type of work the collaboration of both is needed. Well, it seems not being so, since there are a lot of studies on microorganisms from ancient remains, but they have been almost always focused on the detection and analysis of ancient DNA. These studies demonstrated the presence of certain microorganisms although they did not proceed to isolate them.
DNA gives evidence of microorganisms in ancient remains
In relation to yeasts, the oldest evidence is that ribosomal DNA of Saccharomyces cerevisiae has been obtained from residues found in Egyptian wine jars 5000 years old (Cavalieri et al 2003). It must be remembered that the oldest archaeological evidence of large-scale wine production has 7400 years, in north of the Zagros Mountains, in present-day Iran (McGovern et al 1986). As it is known, S. cerevisiae is also the bread and beer yeast, derived from cereals, but since neither S. cerevisiae nor its spores are aerial, surely the use of this yeast in fermented grape juice, as well as dates, figs or honey, historically preceded its use for brewing and bread (Cavalieri et al 2003). It is probable that the wine yeasts naturally occurring in damaged grapes (Mortimer & Polsinelli 1999) were used to ferment other cereal products such as cereals, and after centuries of selection for humans, they evolved into specific strains to ferment food and beverages from cereals.
The genomes of pathogenic microorganisms have also been studied in archaeological remains by means of new massive DNA sequencing techniques, in order to know to epidemic diseases of historical importance, such as black plague, tuberculosis, cholera or leprosy (Andam et al 2016). Logically, in these cases the archaeological remains are human ones, such as bones, teeth, coprolites or mummified tissues. In this way, for example, the phylogeny and evolution of Yersinia pestis strains causing the black plague have been recognized by remains of the Bronze Age (5000 years ago) and until the well-known epidemics of the 6th and 14th centuries (Bos et al 2011). Another well-known case is the Helicobacter pylori genome identified in the intestine of the Ötzi mummy, the iceman in the eastern Alps, 5300 years old (Maixner et al 2016).
DNA has also been isolated from specific bacteria of the human gut, such as Bifidobacterium and Bacteroides, to demonstrate the human presence in archaeological sediments 5000-12000 years old, in north east of Poland (Madeja et al 2009).
It should be remembered that DNA is degraded over time, and in fact it is more unstable than other cellular components. This macromolecule spontaneously suffers damage by oxidation, hydrolysis, and fragmentation in pieces that may be less than 100 bp. Most fossils or other biological remains of more than 100,000 years old no longer contain PCR-amplifiable DNA (Hofreiter et al 2001), although it seems that if the samples are extracted from frozen sediments, with constant temperatures below zero, DNA could be recovered from up to 400,000 years or a little longer (Willerslev et al 2003). In addition the tissues are colonized over time by fungi and bacteria that greatly reduce the relative amount of endogenous molecules and can contribute to giving false positives. The risk of contamination is very high and often this is not taken in account. Generally the DNA of the host that is analysed can be less than 1% of the total DNA found. All these factors complicate the DNA extraction, the construction of sequence libraries, the alignment of DNAs and the analysis of genomes (Andam et al 2016).
Surprisingly, there are a few published works where it is found old DNA of plants, animals and various microorganisms, some million years (My) old, even hundreds of My. The most remarkable are those obtained from amber samples of 20-40 My, and those obtained from a halite 250 My old. This would be comparable to the Jurassic Park fiction where almost non-degraded DNA from the dinosaurs of 100 My old “was recovered”.
Hebsgaard et al (2005) thoroughly reviewed all these more spectacular cases, with the conclusion that these works suffered from inadequate experimental approaches and inadequate authentication of the results. Therefore, there are great doubts as to whether DNA sequences and in some cases viable bacteria could survive such large geological times.
In addition, it is worrying that these works with so old DNA have not been replicated independently in order to confirm their authenticity, and that they did not show a relationship between the age of the sample and the persistence of DNA depending on the different types of bacteria (Willerslev et al 2004). In contrast, Willerslev et al studied the persistence of DNA in permafrost and they found a clear relationship of DNA degradation with time (Figure 3). As seen, DNA amount is very small beyond 100,000 years and it is hardly found beyond 1 My.
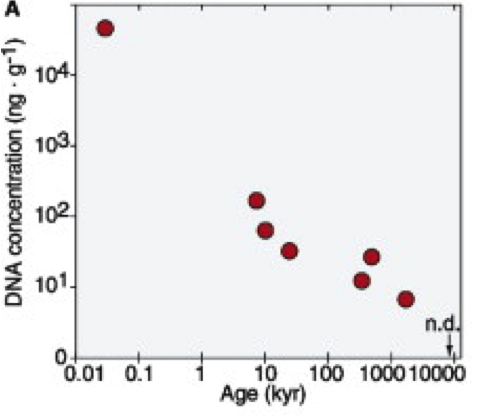
Figure 3. Persistence of not degraded bacterial DNA over time (kyr, thousands of years) maintained in permafrost, measured by fluorescence (Willerslev et al 2004).
When analysing the bacterial phyla of these DNA, Willerslev et al (2004) observed (Figure 4) that the most persistent are those of Arthrobacter, the main representative of Actinobacteria (high G+C gram-positive), followed by sporulated (Bacillaceae and Clostridiaceae), and finally the Gram-negative Proteobacteria.
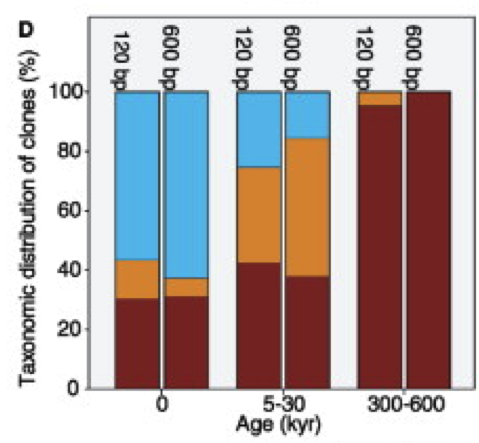
Figure 4. Proportions of the main bacterial phyla (Actinobacteria in brown, sporulated in orange and Proteobacteria in blue) based on DNA obtained from permafrost samples, along time (kyr, thousands of years) (Willerslev et al 2004).
This increased persistence of non-sporulated Actinobacteria is surprising because sporulated bacteria have always been considered the most resistant of all types of cells. Although endospores have special adaptations such as proteins binding DNA to reduce the rate of genetic modifications, they do not have active metabolism or repair and their DNA will degrade over time. The mechanism of greater resistance of Actinobacteria is unknown, but there may be some activity and repair of DNA at temperatures below zero, and/or adaptations related to the dormant cells state (Willerslev et al 2004).
Anyway, the limit for PCR-amplifying the DNA would be between 400,000 years and 1.5 My for samples kept below zero, but this is much more unlikely in non-frozen materials, such as the amber of halite samples of million years, and much less likely to find viable cells from these samples so old (Willerslev et al 2004).
“Resurrected” bacteria
The same commented works where DNA of some millions of years (My) was found, are the most surprising cases of having “resurrected” microorganisms, basically bacteria: viable cells of the sporulated Bacillus from amber samples of 30 My (Cano & Borucki 1995), Staphylococcus also from amber of about 30 My (Lambert et al 1998), and the most spectacular case of Bacillus from an halite of 250 My (Vreeland et al 2000 ). This sporulated bacterium would have been in a hyper-saline environment of the last Permian and trapped in a salt crystal, surviving until now. In the case of Staphylococcus isolated from amber, in spite of not being sporulated, they are bacteria very resistant to extreme conditions, and which have been isolated also from ancient permafrost and very dry environments (Lambert et al 1998).
In spite of this, the revision of these cases by Hebsgaard et al (2005) concludes that none of them fulfilled the relative rate of molecular distance test, which is the probable rate of mutations calculated in comparison to related lineages. Therefore, these isolations are arguable and not reproduced. In addition, in the case of the 250 My Bacillus, it has been argued that the inclusion of bacteria in the halite could be the consequence of a subsequent recrystallization (Lowenstein et al 2011).
Another review on microorganism preservation records (Kennedy et al 1994) comments published cases up to 600 My, indicating that it is curious that there are several cases with more than 1 My, and also cases with less than 10,000 years ago, but there are very few cases of intermediate periods. These authors also point out the doubts raised by works with surviving bacteria so old, which would surely be artefacts or contaminations.
On the other hand, the most credible works are those of Abyzov et al (2006) and Soina et al (2004), which demonstrated the presence of several living microorganisms, both prokaryotes and eukaryotes (especially yeasts, but also some microalgae), in Antarctic ice samples that have some thousands of years. These authors combined classical microbiological methods, such as enrichment and isolation of colonies, together with epifluorescence microscopy, electronic microscopy, and molecular techniques. The bacteria found were Gram-positive (Micrococcus) and gram-negative (Arthrobacter), which are not sporulated, but they have cist-shaped dormant cells, which can survive while maintaining viability at temperatures below 0ºC for some thousands of years.
When geologically ancient DNA findings are published as well as viable cultures of ancient samples, the independent reproduction of the results by another laboratory is fundamental, to exclude any contamination from the same laboratory. In the case of having recovered living cells, it is necessary to demonstrate the reproducibility of the isolation, sequencing the genomes of the cultures obtained in independent laboratories from the same sample, and checking that in both cases the genomes coincide (Hebsgaard et al 2005).
From the remains of the Roman fort of Vindolanda, in the north of England, viable endospores of Thermoactinomyces, member of Bacillales (Unsworth et al 1977) have been recovered. They are about 1900 years old and the remains were a mixture of clay with straw and other vegetable materials. The authors propose to use these sporulated bacteria as indicators in archaeological studies.
Besides sporulated bacteria, there are several groups of non-sporulated ones for which anabiosis resistance abilities have been demonstrated. In particular, they have been isolated from permafrost and the tundra soil of Siberia of about 1 My (Suzina et al., 2006), in the limit of what we mentioned earlier (Willerslev et al 2004), which is quite difficult to believe. In order to study experimentally the formation of these anabiosis forms, Suzina et al incubated several gram-positive and gram-negative bacteria, and some archaea, in poor media with limiting nitrogen, and after a few months they obtained their dormant cells. They had cist structures, with capsule and a thickened cell wall, intramembranous particles and a condensed nucleoid (Figure 5). They also observed that these cysts did not have metabolic activity and supported stress factors such as lack of nutrients or heating.
Studying the permafrost isolates, they confirmed that there are cist structures very similar to those obtained in the laboratory, with multi-layer wall structures of up to 0.4 μm. In fact, these authors believe that most of the bacteria present in the permafrost and the tundra are in the form of a cyst (Suzina et al 2006).
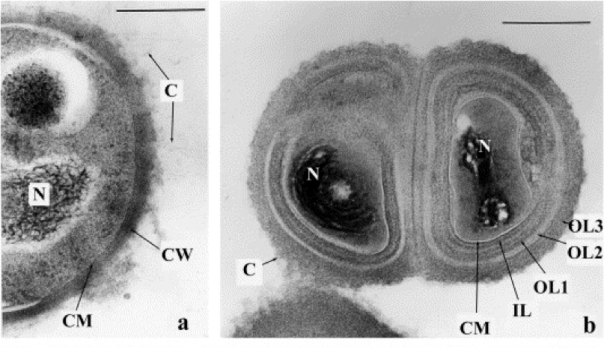
Figure 5. Sections of a vegetative cell (a) of Micrococcus luteus and of a cyst cell (b) of the same bacterium, obtained after 9 months of culture in a medium limiting in nitrogen. C, microcapsule; CW, cell wall; OL1, 2, 3, outer layers of the cell wall; IL, inner layer of the wall; CM, cytoplasmic membrane; N, nucleoid. The bar measures 0.3 μm (Suzina et al., 2006).
Other “resurrected” yeasts and fungi
Besides the surprising mentioned article by Aouizerat et al. (2019), there are other few published cases of yeasts and other “resurrected” fungi such as the following.
Chicha is a beer-like beverage from corn, yellowish and slightly effervescent, elaborated and consumed by Andean populations for some thousands of years, whose traditional process has the peculiarity of using amylase of saliva for convert the starch into fermentable sugars. Fermentation traditionally took place in clay containers called “pondos”. From the remains of the chicha pondos from the Hipia culture in Quito (2100-2800 years old), various yeast were isolated, especially Candida, Pichia and Cryptococcus (Gomes et al 2009). Interestingly, some of these yeasts have been confirmed molecularly as Candida theae, similar to those isolated from contaminated Asian tea (Chang et al 2012). It is worth mentioning the absence of Saccharomyces in these ancient chicha, although today it is the main yeast, coming probably from beer and wine fermentation that led the Spaniards (Gomes et al 2009).
From Greenland ice samples of about 100,000 years (Ma et al 1999), several microorganisms were revived, such as bacteria (Micrococcus, Rhodotorula, Sarcina) and yeasts (Candida, Cryptococcus) and other fungi (Penicillium, Aspergillus). The authors also isolated the DNAs and demonstrated the phylogenetic relationship of the isolates. Once again, we see how ice provides a stable environment that facilitates the conservation of microorganisms and their DNA.
Raghukumar et al (2004) have recovered living Aspergillus (sporulated Ascomycota fungus) and other fungi from sediment samples of the deep-sea, about 5900 m deep in the Chagos trench, south of the Maldives, in the Indian Ocean. Based on the depth in the sediment and the present Radiolaria, authors estimated that they correspond to a minimum of 180,000 years, and up to 430,000 years in some samples. From the isolates identified as A. sydowii they obtained spores that germinated and grew in hydrostatic pressure equivalent to the depth of 5000 m, and at a temperature of 5ºC. With microscopy of epifluorescence and bright field, the fungal hypha and their relation to the particles of the sediment are clearly observed (Figure 6). It seems that this Aspergillus found in the deep-sea is the oldest fungus recovered alive so far. The authors suggest that preservation would have been possible thanks to high hydrostatic pressure, along with low temperature.
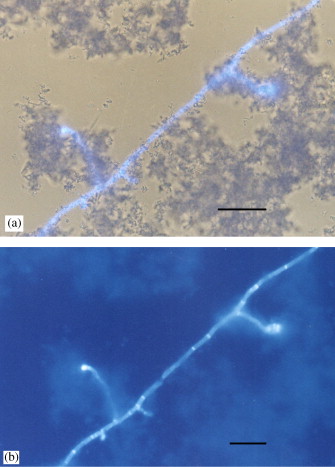
Figure 6. Photomicrographies of deep-sea sediment (5900 m) of the Indian Ocean with hyphae of Aspergillus sydowii and sediment particles. (a) epifluorescence microscopy combined with that of bright field; (b) epifluorescence (Raghukumar et al 2004).
One of the most surprising works, and hard to believe, is that of Kochkina et al (2001), where a lot of fungi of all kinds and bacteria, especially actinobacteria, were isolated from samples of permafrost from Russia, Canada and Antarctica reaching 3 My old. The authors even suggested that there is no limit of years to recover viable microorganisms. This article has had very little echo, and it is not even mentioned by later articles as Raghukumar et al. (2004).
Conclusions
As we have seen, evidence of DNA from no-living yeasts in ancient remains related to winemaking dates back to around 5000 years in ancient Egypt (Cavalieri et al 2003). Regarding other microorganisms, taking into account the natural degradation of DNA over time, it seems that the oldest samples would be about 400,000 years at most, in particular actinobacteria in frozen sediments such as permafrost (Willerslev et al 2003 ). Publications of bacterial DNA recovered from several millions years (up to 600 My) have many scientific concerns about their credibility and reliability (Kennedy et al 1994).
With regard to living yeast as those of 3000 years apparently isolated by Aouizerat et al (2019), it seems that Candida and others were isolated from containers to elaborate chicha about 2800 years old (Gomes et al 2009), although this reference is a review and the original work does not appear to have been published. Other authors (Abyzov et al 2006; Soina et al 2004) also find alive yeasts, without specifying which ones, in Antarctic ice samples of some thousands of years. More surprising are the isolated isolations of yeast and other fungi and bacteria from Greenland ice samples 100,000 years old (Ma et al 1999), as well as those of Aspergillus from the Indian Ocean seabed of about 180,000 years (Raghukumar et to 2004).
Regarding other “resurrected” microorganisms, some of the most reliable are the several Antarctic ice bacteria of some thousands of years (Abyzov et al 2006) and Thermoactinomyces spores of Roman remains 1900 years old (Unsworth et to 1977). Of the oldest, perhaps the anabiosis forms of bacteria conserved in permafrost a million years old (Suzina et al., 2006) would have certain likelihood. Curiously, these bacteria would be non-sporulated but they would have a cyst structure, with multi-layer walls and other intracellular modifications. The other findings of “resurrected” bacteria from more millions of years of amber or halite, just like their DNA and also because of this, are very hard to believe (Hebsgaard et al 2005).
Thinking in the cellular forms of resistance and anabiosis, as the bacterial endospores and the mentioned cysts, it must be remembered that yeasts, like many other fungi, have the ability to produce spores, in particular ascospores as they are Ascomycetes. Although these ascospores have a greater capacity for resistance than vegetative cells in dry conditions or other inhospitable environments and have a persistence in time, apparently there is no work (or I have not been found) related to the recovery of yeasts ascospores from ancient remains.
The work of Aouizerat et al (2019) makes no mention of the yeast spores, neither as a possible explanation of the yeast survival in these ancient remains. In fact, they propose that the microcolonies of yeasts on ceramics pores would have continued to grow minimally during these 3000 years thanks to the humidity and residual nutrients. Well, we do not know, and neither if the yeast ascospores have had any role.
Finally, we can believe the finding of Aouizerat et al (2019) is truth, but obviously further investigation in other similar archaeological samples must be done. This research should be done not only for yeasts, but also for bacteria of other fermented products. Besides considering the sporulated ones, other bacteria should be considered, that could survive thanks to the cell cysts or other forms of anabiosis.
Bibliography
Abyzov SS et al (2006) Super-long anabiosis of ancient microorganisms in ice and terrestrial models for development of methods to search for life on Mars, Europa and other planetary bodies. Adv Space Res 38, 1191-1197
Andam CP et al (2016) Microbial genomics of ancient plagues and outbreaks. Trends Microbiol 24, 978 –990
Aouizerat T et al (2019) Isolation and characterization of live yeast cells from ancient vessels as a tool on bio-archaeology. mBio 10, 2, 1-21
Borschel-Dan A (2019) Israeli scientists brew groundbreaking “ancient beer” from 5,000-year-old yeast. The Times of Israel, 22nd may 2019.
Bos KI et al (2011) A draft genome of Yersinia pestis from victims of the Black Death. Nature 478, 506–510
Cano, R.J. and Borucki, M.K. (1995) Revival and identification of bacterial spores in 25- to 40-million year-old Dominican amber. Science 268, 1060–1064
Cavalieri D et al (2003) Evidence for S. cerevisiae fermentation in ancient wine. J Mol Evol 57:S226-232
Chang CF et al (2012) Candida theae sp. nov., a new anamorphic beverage-associated member of the Lodderomyces clade. Int J Food Microbiol 153, 10-14.
Gomes FCO et al (2009) Traditional foods and beverages from South America: microbial communities and production strategies. Chapter 3 in Industrial Fermentation, ed. J Krause & O Fleischer, Nova Science Publishers.
Hofreiter M et al (2001) Ancient DNA. Nature Rev Genet 2, 353–359.
Kennedy MJ et al (1994) Preservation records of micro-organisms: evidence of the tenacity of life. Microbiology 140, 2513-2529.
Kochkina GA et al (2001) Survival of micromycetes and actinobacteria under conditions of long-term natural cryopreservation. Microbiology 70, 356-364
Lambert LH et al (1998) Staphylococcus succinus sp. nov., isolated from Dominican amber. Int J Syst Bacteriol 48, 511-518
Lowenstein TK et al (2011) Microbial communities in fluid inclusions and long-term survival in halite. GSA Today 21, 4-9
Ma L et al (1999) Revival and characterization of fungi from ancient polar ice. Mycologist 13, 70-73.
Madeja J et al (2009) Bacterial ancient DNA as an indicator of human presence in the past: its correlation with palynological and archaeological data. J Quaternary Sci 24, 317-321.
Maixner F et al. (2016) The 5300-year-old Helicobacter pylori genome of the Iceman. Science 351, 162–165
McGovern PE et al (1986) Neolithic resinated wine. Nature 381:480–481
Mortimer R & M Polsinelli (1999) On the origins of wine yeast. Res Microbiol 150, 199-204
Raghukumar C et al (2004) Buried in time: culturable fungi in a deep-sea sediment core from the Chagos Trench, Indian Ocean. Deep Sea Res Part I: Oceanog Res Papers 51, 1759-1768
Soina VS et al (2004) The structure of resting microbial populations in soil and subsoil permafrost. Astrobiology 4 (3), 345–358.
Suzina et al (2006) The structural bases of long-term anabiosis in non-spore-forming bacteria. Adv Space Res 38, 1209-1219.
Unsworth BA et al (1977) The Longevity of Thermoactinomycete Endospores in Natural Substrates. J Appl Microbiol 42, 45-52
Vreeland RH et al (2000) Isolation of a 250 milion-year-old halotolerant bacterium from a primary salt cristal. Nature 407, 897-900.
Willerslev E et al (2003) Diverse plant and animal DNA from Holocene and Pleistocene sedimentary records. Science 300, 791-795
Willerslev E et al (2004) Long-term persistence of bacterial DNA. Curr Biol 14, PR9-R10.

Bacteroides, our most abundant gram-negative bacteria
17th April 2019
Translated from the original article in Catalan.
Update on this topic: see the work of the Yolanda Sanz group, IATA-CSIC, such as Gómez del Pulgar et al., 2020.
What are Bacteroides ?
Bacteroides is the best-known genus of the most abundant gram-negative bacterial group within us, specifically in the intestine. They are up to 8·1010 per gram of stool. They are strict anaerobes, non-sporulated, non-mobile, with a form of rod with rounded tips (Figure 1). They are resistant to bile salts, at the concentration of 20% of the small intestine, and they have a good ability to use polysaccharides.
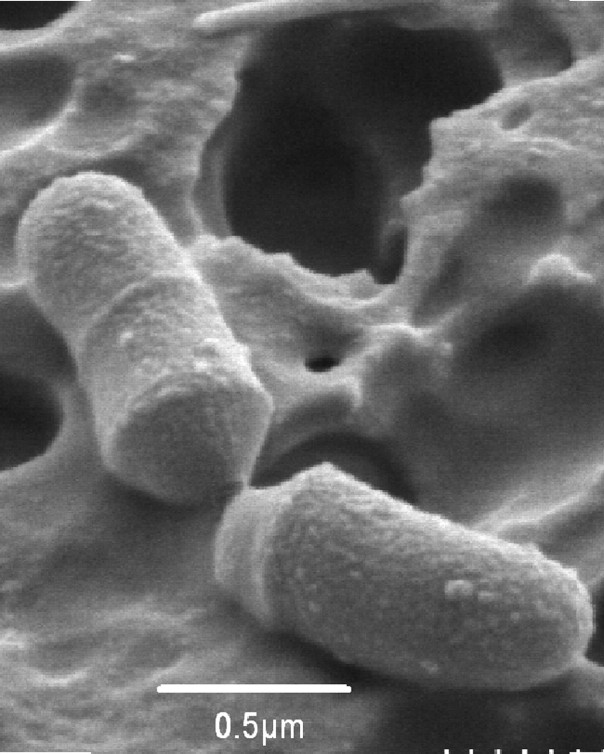
Figure 1. Electronic micrograph of cells of Bacteroides sp. D8 (Gerard et al 2007)
First of all, it should be noted that there are excellent revisions of Bacteroides, such as that of Wexler (2007), describing their beneficial aspects in the intestinal microbiota, which we will comment on here, as well as the toxic aspects and other characteristics.
Bacteroides live exclusively in the gastrointestinal tract of animals, and therefore they show great flexibility to adapt to the nutritional conditions of the intestinal environment. As commensals and mutualists, they establish long-term partnerships with the guests and provide them with benefits. The adaptation of these bacteria includes making modifications to this environment. For instance, many Bacteroides code for cytochrome bd oxidase, which can reduce oxygen concentrations, making it easier for them to grow as strict anaerobes, and at the same time, other bacteria of the usual microbiota also benefit from this (Wexler, Goodman 2017).
The most common substrates of these bacteria are the vegetable polysaccharides of the diet and of host’s mucus (Wexler, Goodman 2017). These carbohydrates are degraded and fermented, producing mainly short-chain fatty acids (SCFA). Bacteroides are the main producers of propionate in intestinal tract, and this acid is one of the beneficial SCFA, together with acetate and butyrate, because they are an energy source for colonocytes and contribute to maintenance of the correct glucose homeostasis and lipid metabolism (Ríos-Covián et al 2017). Bacteroides also remove side chains from bile salts, facilitating the return of bile acids to liver circulation. On the other hand, another beneficial aspect is that they exclude other possible pathogens as they colonize the intestinal tract and do not let others settle.
Due to the fact that the animal’s intestinal tract is the main habitat and environmental reservoir of Bacteroides, it is thought that there has been a symbiotic evolutionary relationship between these bacteria and the hosts (Troy, Kasper 2010). As in many other evolutionary cases, this mutual commensalism between microorganisms and hosts is almost a symbiosis, where virtually each of the organisms cannot live without the other.
As habitual residents of the intestine, the vast majority of Bacteroides are not harmful, on the contrary. Nevertheless, in conditions of metabolic imbalances such as diabetes or surgical patients, some of them are opportunistic and can be pathogens, and some have a certain resistance to antibiotics. In fact, B. fragilis, the most abundant species in the microbiota of healthy people, can give in these cases very serious infections and is the most important anaerobic pathogen bacterium in humans (Mancuso et al 2005). The abundance of B. fragilis is evident even because their bacteriophages are used as tracers of human faecal matter in water (Jofre et al 1995).
What kind of bacteria is Bacteroides ?
As detailed in the NCBI Taxonomy section, the genus Bacteroides is a bacterium of the Fibrobacter-Chlorobi-Bacteroidetes superphylum. We can see its phylogenetic relationship with other bacterial groups in Figure 2. Bacteroidetes phylum also includes Cytophaga, Flavobacter and Sphingobacter, in addition to the Bacteroidia class, which mainly includes the Bacteroidales order. This includes 2 families: the Bacteroidaceae and the Prevotellaceae. Besides Bacteroides, Prevotella is another of the best-known genera, which in fact was previously known as B. melaninogenicus.
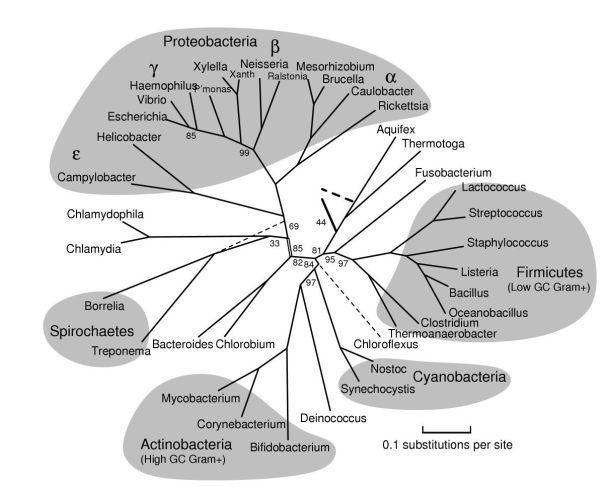
Figure 2. Phylogenetic tree of the bacterial groups (Bern, Goldberg 2005).
Bacteroides, some of the predominant in the human intestinal microbiota
The human intestinal microbiota, and from mammals in general, is very complex, but surprisingly, there are few phyla that predominate. Specifically, 98% of identified bacteria in humans (Figure 3) belong to 4 phyla: 64% Firmicutes, 23% Bacteroidetes, 8% Proteobacteria and 3% Actinobacteria. Therefore, Bacteroidetes are one of the most predominant bacteria in the intestinal microbiota. In fact, since Firmicutes are such a large and diverse phylum, which includes microbes as diverse as clostridial and lactic acid bacteria, it can be considered that Bacteroidetes, as a much more homogeneous group, are practically the predominant ones.
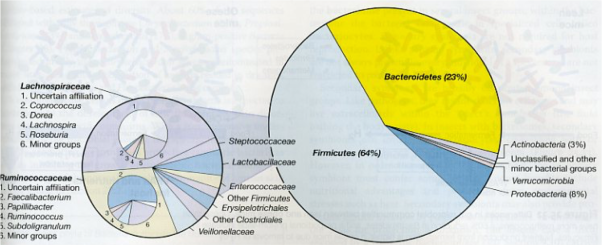
Figure 3. Bacterial composition of the human colon deduced from the 16S rRNA obtained from 17242 sequences of faecal samples (Madigan et al 2012)
To see in depth the predominant species of the intestinal microbiota, very recently, a metagenomic and functional study of 737 genomes sequenced from bacterial isolates of faecal samples from 20 British and American adults (Forster et al 2019) has been done. 273 bacterial species have been detected, of which 105 had not been found before. As we can see (Figure 4), among the 20 dominant species there are 8 Bacteroides, plus 2 Parabacteroides, that is 10 Bacteroidales, signalled in green. Therefore, they are half of the majority species. The other 10 are 6 clostridial (Firmicutes, in blue), 3 are Actinobacteria (in yellow) and 1 is Proteobacteria (in orange).
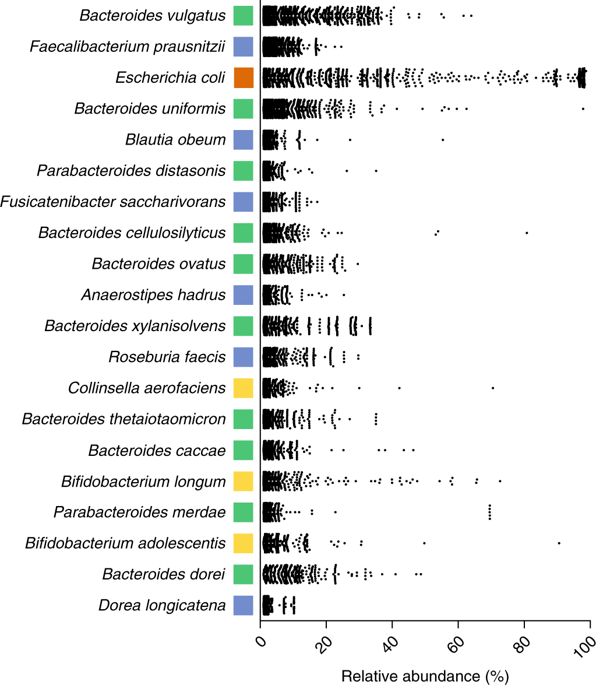
Figure 4. Major species of the human intestinal microbiota, detected with metagenomic data analyses (Forster et al 2019).
Although the microbiota is different in each person, at the strain level the individual microbiota is very stable. In a study with 37 healthy people (Faith et al 2013) about 200 strains of 100 different species have been found, and 60% of the strains remain for each person in a period of 5 years. Of those that remain, those of Bacteroidetes and Actinobacteria are the most stable.
In the same study (Faith et al 2013), gut microbiota of 6 people in the same family have been compared and it has been found that among the 75 most common bacterial species in the 6 persons, 18 are Bacteroidetes (24%): 11 Bacteroides, 3 Parabacteroides, Alistipes, Barnesiella, Odoribacter and Butyricimonas. The only species of the 75 found in everybody is a Bacteroides: B. vulgatus.
The microbiota that accompanies us is changing throughout life (Figure 5). In fact, there are relatively few Bacteroides in the babies. However, these bacteria are already present among the few microbes of the placenta, where Proteobacteria predominate (Aagard et al 2014). After the birth, Bacteroides are increasing over the first months and years, mainly with the weaning and diet changes, as microbial diversity increases. Then, in adults Bacteroides are ones of the most abundant microbes (Gómez-Gallego, Salminen 2016).
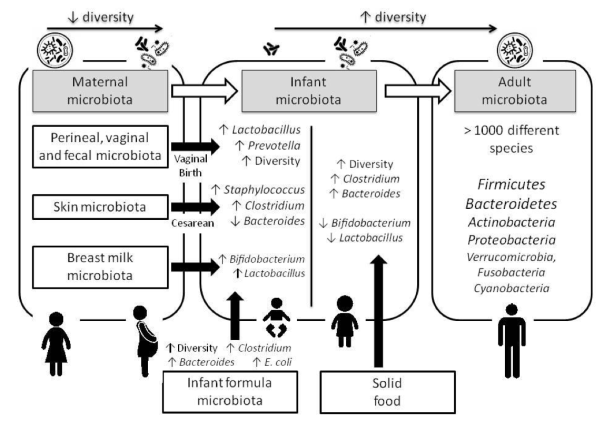
Figure 5. Changes in the human microbiota throughout life (Gómez-Gallego, Salminen 2016).
Solid food intake in children, between 4 months and 1 year, causes a significant increase in Bacteroidetes (Figure 6). We see the great difference in the microbial composition from 118 day to 370. It is a pity that in this study (Koenig et al 2011) no more intermediate samples were took between these days, where little by little children go from porridge and a bit of cereals, to the ingestion of peas and other legumes, carrots, potatoes, etc. This increase in Bacteroidetes with solid food is surely related to the fact that Bacteroidetes are specialists in the breakdown of complex polysaccharides, and at the same time these compounds promote their growth. At the same time, there is a clear increase in the levels of AGCC, an enrichment of microbial genes associated with the use of carbohydrates, a greater biosynthesis of vitamins, and also an increase of xenobiotic degradation. Therefore, the role of Bacteroidetes seems primordial in the establishment and maintenance of the adult’s microbiota. Even though there are differences between individuals, once adult, microbial composition is quite stable throughout life, with certain variations depending on changes in diet or habitat or medication.
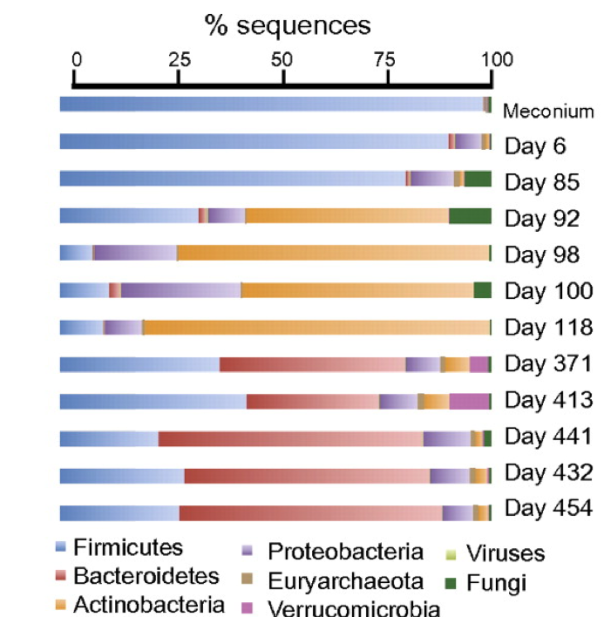
Figure 6. Metagenomic analysis of DNA sequences extracted from faecal samples of children (Koenig et al 2011).
Bacteroides in other mammals
The intestinal microbiota is present in all animals with a more or less developed digestive system. Apart from the insects, whose microbiota has been deeply studied (Engel, Moran 2013), the most studied in this aspect are mammals, of course. Their composition has been studied (Ley et al 2008), specifically in faecal samples of 106 individuals of 60 species of 13 different taxa, including human, other primates, herbivores, carnivores and omnivores.
Of the 17 bacterial phyla found, 65% were Firmicutes, 16% Bacteroidetes, 8% Proteobacteria and 5% Actinobacteria, among others. Therefore, the relevance of the Bacteroides is evident, and the proportions are similar to those mentioned above for humans. Regarding the majority group of Firmicutes, it is a pity that this work, like others, does not distinguish between different groups, especially among lactic acid bacteria and Clostridiales. Curiously in this work there is a greater presence of Bacteroides in primates and omnivores in general, and also in some herbivores, than in carnivores (Figure 7). In these there are very few Bacteroides, and instead there are more gamma-Proteobacteria, probably enterobacteria (Ley et al 2008).
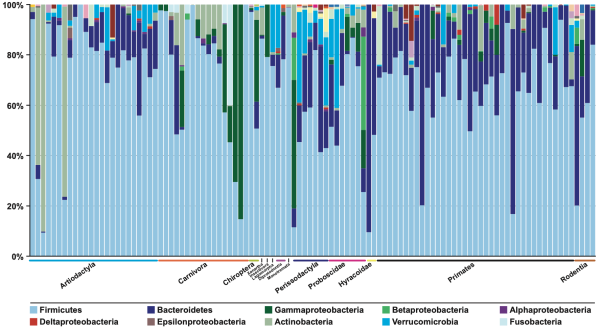
Figure 7. Percentage of faecal samples sequences of different mammals assigned to the main different bacterial phyla (Ley et al 2008)
Different Bacteroidales are biomarkers of lifestyles
In the search for microbial taxa that could be biomarkers of diets or lifestyles, it has been seen that the biomarker more clearly related with people from rich western countries is the genus Bacteroides, whereas to the sub-Saharan ones it is Prevotella, another one of the same phylum. These two genera, together with some from the clostridia group, are the most abundant ones.
If the long-term majority diet is rich in animal proteins and fats, as in Western countries, Bacteroides predominates, and if the diet is rich in carbohydrates like in sub-Saharan countries, Prevotella prevails (Gorvitovskaia et al 2016).
What about Bacteroides in cases of dysfunction?
The beneficial relevance of Bacteroides, or their group, Bacteroidetes,on health is obvious in cases of diseases or dysfunctions such as allergies or obesity (Figure 8), where the diversity of the microbiota is much lower, and the number of Bacteroidetes is low.
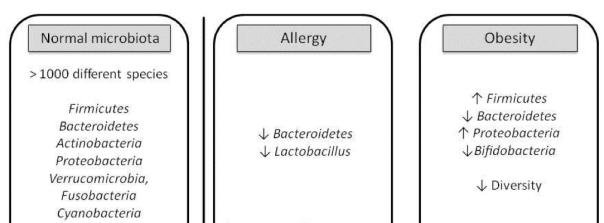
Figure 8. Changes in the microbiota in dysfunctional situations such as allergies and obesity. (Gómez-Gallego, Salminen 2016).
Bacteroides against obesity
Well-known experiments of intestinal microbiota in relation to obesity have been those carried out with mice without previous microbiota colonized with microbiota from human twins of which one was obese and the other lean (Ridaura et al 2013). The result was that the mice with obese twin microbiota (Ob) became obese, while those of lean twin microbiota remained lean (Ln) (Figure 9). In addition, in the lean mice a greater intestinal production of SCFA and a greater microbial transformation of the bile acids were observed, whereas in the obese there was a greater metabolism of branched amino acids.
As mentioned in the previous section, in the obese mice a reduction of 50% Bacteroidetes is observed, apart from an increase in Firmicutes and methanogens (Figure 10). And as we see the Archaea methanogens decrease the hydrogen, producing methane, and the lower level of hydrogen promotes fermentation of ingested food in excess by the Firmicutes.

Figure 9. Obese and lean mice resulting from colonization with gut microbiota from obese and lean human twins respectively (image of Kay Chernush / Getty Images).
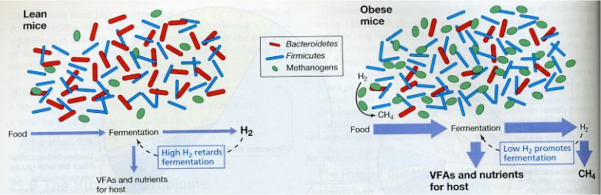
Figure 10. Differences in intestinal microbial communities between lean (left) and obese (right) mice (Madigan et al 2012).
The most surprising, however, of this work (Ridaura et al 2013) is the cohabitation experiment of the two types Ob and Ln mice, where it is observed that after 10 days of coexisting together, the obese have diminished their body fat (Figure 11), and when their microbiota have been studied by sequencing, a transfer of the microbiota from lean mice to obese is observed (Figure 12). As we can see, the main bacteria transferred are Bacteroidales, which strengthens the importance of these bacteria.
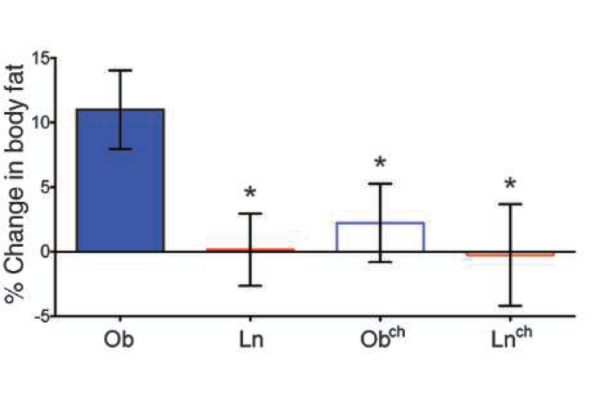
Figure 11. Adiposity (% body fat) of obese (Ob) and lean mice (Ln), and the same after 10 days of cohabitation in the same cage (Obch and Lnch) (Ridaura et al 2013).
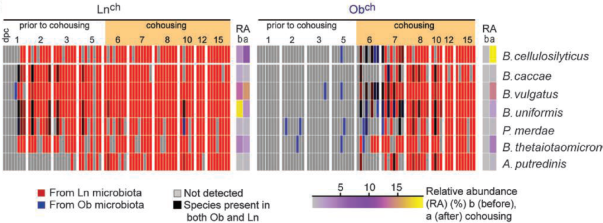
Figure 12. Demonstration of the transfer of Bacteroidales (7 species: 5 Bacteroides, 1 Parabacteroides and 1 Alistipes) of the intestinal microbiota of lean mice (Lnch) to the obese (Obch) after 10 days of cohabitation in the same cage. Each column corresponds to a mouse (Ridaura et al 2013).
Bacteroides against cholesterol
It has been known for many years that the intestinal microbiota is able to convert cholesterol in its saturated form, coprostanol (Figure 13). In other mammals some Eubacterium (belonging to the clostridial group) have been found to be responsible, but in humans we did not know what microorganisms could do it. Recently Gérard et al (2007) have isolated a strain of human stool that is able to do it and has been identified as Bacteroides, probably a species close to B. vulgatus.
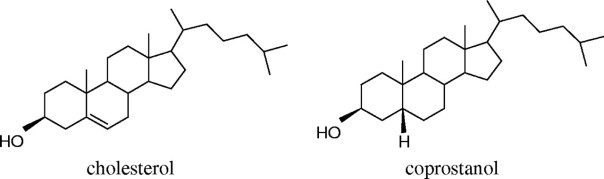
Figure 13. Formulas of cholesterol and coprostanol (Gerard et al 2007)
Glycans (polysaccharides), important for mutualism between Bacteroides and the human host
Most non-digested macromolecules that reach the colon are glycans (word virtually synonymous of polysaccharides), which are a very important part of the fibre. The only glycan that is practically digested previously in the small intestine is starch. The consortium of microorganisms that inhabit the colon produces a huge enzymatic repertoire with the ability to degrade a range of complex polysaccharides that the host cannot process. That’s why the intestinal microbiota is often referred to as a metabolic organ.
On the other hand, the abundant commensal microbes of the intestinal microbiota must resist the inhospitable conditions of the previous sections and to settle in the colon without affecting the host. Therefore, instead of interacting with the epithelial cells of the intestine, they remain in the external mucus layer on the epithelial surface. At the same time, this mucus protects resident microbes from attacks by other bacteria and bacteriophages, and it is a nutrient substrate. It has been shown that the ability to survive in this ecosystem is closely related to the use and production of glycans by resident bacteria (Comstock 2009).
Well, precisely this ability to interact with glycans is an important characteristic of Bacteroidales, which, as we have seen, are the most abundant microorganisms in the intestine, along with Firmicutes. In fact, Bacteroidales have an extensive enzymatic machinery to use the complex polysaccharides present in the colon, and use them as a source of carbon and energy. This great capacity has been proven by sequencing the genome of B. thetaiotaomicron (Xu et al 2003) where it has found containing more than 80 loci of polysaccharides that encode proteins related to the detection, importation and degradation of specific glycans of the colon.
As we can see (Figure 14), Bacteroides use both the glycans of the host’s diet and those produced by the intestinal epithelium, they metabolize them, and produce the beneficial SCFA, and on the other hand, they synthesize glycans that accumulate in the form of exopolysaccharide (EPS) contributing to form biofilms, and in capsules that give immune signals to the host (Comstock 2009). All in all, the relevance of the glycans in the mutual relations between Bacteroides and the human host is confirmed.
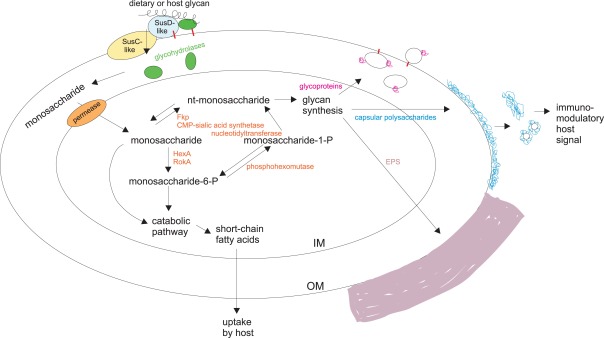
Figure 14. Use and production of glycans (polysaccharides) by Bacteroides. IM (inner membrane): cytoplasmic membrane; OM (outer membrane): external part of the gram-negative cell wall; EPS: exopolysaccharide of mucosal layers, not covalently linked, unlike the capsular polysaccharide (Comstock 2009).
In addition to the glycans produced by the host, some Bacteroides can also use those that produce other microorganisms of the microbiota, as shown by B. fragilis, the most frequent species on the surface of the intestinal mucosa, which can metabolize exopolysaccharides produced by bifidobacteria (Ríos-Covian et al 2016). EPS production for bifidobacteria is stimulated by bile. This ability of B. fragilis to use EPS of bifidobacteria gives them more survival capacity when nutrients are scarce. At the same time, the degradation of the EPS can affect the viability of the bifidobacteria, and therefore, Bacteroidales would have a regulatory role of the intestinal microbiota in general.
Some glycans produced by Bacteroidales have a beneficial effect on the host’s immune system. In particular, it has been seen that polysaccharide A (PSA) produced by B. fragilis is able to activate the immune response on T-cells dependent, which influences the development and homeostasis of the immune system (Troy, Kasper 2010). In fact, the colonization of germ-free mice (without microbiota) with B .fragilis is sufficient to correct the previous imbalance of cells Th1 and Th2 (T helper) (Figure 15). In addition, PSA can protect against colitis, such as those produced by Helicobacter, by repressing proinflammatory cytokines associated with another type of T cells -Th17- and other mechanisms (Mazmanian et al 2008).
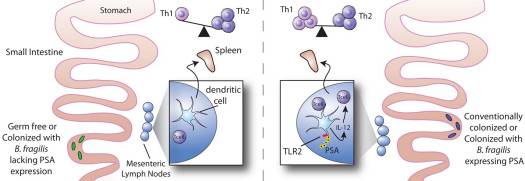
Figure 15. Impact of polysaccharide A (PSA) of Bacteroides fragilis in the development of the immune system by recovering the balance of Th1/Th2 cells (Troy, Kasper 2010).
The diet can make Bacteroides contribute to a good metabolic balance
In relation to said above about glycans such as EPS, it has been seen that if in the environment there is little organic nitrogen and an easily fermentable carbon source such as glucose, Bacteroides produce more lactate and less propionate, and instead with more organic nitrogen (yeast extract) and polysaccharides, these bacteria produce more propionate (Ríos-Covián et al 2017). When EPS are present, as more complex carbohydrates and slowly fermented, the carbon of the amino acids can be incorporated at the level of pyruvate, and then the path to succinate and propionate is enhanced and the redox equilibrium is maintained. Since a higher propionate production is beneficial to the host, these authors conclude that in cases of host metabolic dysfunctions, a good diet design (complex carbohydrates with organic nitrogen) would help to modify metabolic activity of Bacteroides, and these would help promote healthy effects to the host, in addition to interacting with the other beneficial bacteria.
Bacteroides as probiotics?
EFSA (European Food Safety Authority) has not accepted virtually any claim of positive effects of probiotics on health due to the restrictive requirements of studies with humans. The mechanism of probiotics action is strain-dependent and often is not well known. In addition, it could be that the incorporated bacteria did not produce sufficient measurable changes in healthy individuals to obtain a claim of health effect. Further studies at the genetic level, antibiotic resistance profile and probiotic selection criteria are required.
Traditional probiotics are mostly Lactobacillus and Bifidobacterium, but also some strains of other lactic acid bacteria, and from Bacillus, E. coli and Saccharomyces. Besides these, the so-called “next generation” probiotics are being introduced, thanks mainly to new culture and sequencing techniques. Among these new possible probiotics, there are the verrucomicrobial Akkermansia muciniphila, and some clostridia (see my post), like Faecalibacterium prausnitzii, the main producer of butyrate, but also some Bacteroidales. These ones also have a clear advantage over clostridia and other Firmicutes, because are much more stable in the intestinal tract throughout the life of the person (Faith et al 2013).
As we have seen, being some of the most abundant microorganisms in our intestinal microbiota, Bacteroides generally have clear benefits for the host, such as fighting against obesity, or cholesterol. Transplants of faecal microbiota for diarrhea associated with Clostridium difficile infections are being successful (Van Nood et al 2013) and therefore there is a clear possibility of using some specific strain or several ones, and in this way the Bacteroides are clear candidates due to their abundance in the samples of faecal microbiota.
In addition to those mentioned, other benefits of Bacteroides are those related to the immune system, at the level of cytokines and T cells and development of antibodies, in order to treat intestinal colitis, immune dysfunction, disorders of metabolism and even cancer prevention (Tan et al 2019).
Apart from the benefits shown to the host, a bacterial strain must have unambiguous security status in order to be considered probiotic. In the case of Bacteroides, recently, a strain (DSM 23964) of B. xylanisolvens isolated from stools of healthy humans has been studied and it has been shown to have no virulence determinants which have been found in some opportunistic Bacteroides, such as the enterotoxin bft and enzymatic biodegradative activities of extracellular matrix and PSA. This strain does not have resistance to antibiotics – although it is resistant to some – and no plasmids have been detected, which makes the transfer of possible resistance very unlikely. Therefore, this strain seems very safe (Ulsemer et al 2012a). It has also been seen that it does not adhere to the walls of the intestine, but it resists the action of gastric enzymes and low pH. In addition, as indicated by the name of the species, it degrades xylan and other pectins. These heteropolysaccharides are prebiotics, compounds that are beneficial for the gut microbiota.
Other basic probiotic characteristics found in this strain of B. xylanisolvens are the production of SCFA and immunomodulatory properties. These properties and the safety and good tolerance of this strain have been verified by incorporating it in fermented milk, after inactivation by heat. This milk has been ingested in trials by healthy humans, with safe effects (Ulsemer et al 2012b). Its safety has also been confirmed in studies of toxicity in mice, where high doses of the strain have not produced toxic or mutagenic effects, neither haematological nor histopathological damage (Ulsemer et al 2012c).
On the basis of these studies, the European Food Safety Authority has given the approval as a new food of the use of fermented milks with B. xylanisolvens DSM 23964 pasteurized (EFSA 2015). However, there is no claim to consider it as a probiotic, especially because bacteria are not viable as the product has been pasteurized, and by definition, probiotics should be living microorganisms.
Perspectives
We have seen the relevance of Bacteroides as one of the main components of the human intestinal microbiota and mammals in general. In addition to its fundamental role in the intestine and the possibilities of its use as a probiotic, it is an ideal model for the study of gut bacteria, because it is relatively easy of cultivating and has the potential to be genetically manipulated (Wexler, Goodman 2017). Therefore, it is necessary to deepen the knowledge of Bacteroidales, and in particular to know how they metabolize the host’s nutrients or mucus, or how they respond to changes in the host’s diet, or how they interact with the other microorganisms of the digestive tract. A better understanding of all these mechanisms will favour the design of therapeutics aimed at modifying the microbiota of patients suffering from various diseases and metabolic disorders linked to the intestinal microbiota (Wexler, Goodman 2017).
Bibliography
Aagaard K(2014) The placenta harbors a unique microbiome. Sci Transl Med 6, 237ra65
Bern M, Goldberg D (2005) Automatic selection of representative proteins for bacterial phylogeny. BMC Evolut Biol 5:34
Comstock LE (2009) Importance of glycans to the host – Bacteroides mutualism in the mammalian intestine. Cell Host & Microbe 5, 522-526
EFSA, European Food Safety Authority (2015) Scientific opinion on the safety of “heat-treated milk products fermented with Bacteroides xylanisolvens DSM 23964″ as a novel food. EFSA J 13(1):3956
Engel P, Moran NA (2013) The gut microbiota of insects – diversity in structure and function. FEMS Microbiol Rev 37, 699-735
Faith JJ et al (2013) The long-term stability of the human gut microbiota. Science 341, 1237439
Forster et al (2019) A human gut bacterial genome and culture collection for improved metagenomic analyses. Nature Biotechnol 37, 186-192
Gérard P et al (2007) Bacteroidessp. strain D8, the first cholesterol-reducing bacterium isolated from human feces. Appl Env Microbiol 73, 5742-5749
Gómez-Gallego C, Salminen S (2016) Novel probiotics and prebiotics: how can they help in human gut microbiota dysbiosis ? Appl Food Biotechnol 3, 72-81
Gómez del Pulgar EM, Bénitez-Páez A, Sanz Y (2020) Safety assessment of Bacteroides uniformis CECT 7771, a symbiont of the gut microbiota in infants. Nutrients 12, 551
Gorvitovskaia A et al (2016) Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 4:15, 1-12
Jofre J et al (1995) Potential usefulness of bacteriophages that infect Bacteroides fragilis as model organisms for monitoring virus removal in drinking water treatment plants. Appl Environ Microbiol 61, 3227-3231
Koenig JE et al (2011) Succession of microbial consortia in the developing infant gut microbiome. PNAS 108, 4578-4585
Ley RE et al (2008) Evolution of mammals and their gut microbes. Science 320, 1647-1651
Madigan MT, Martinko JM, Stahl DA, Clark DP (2012) Brock Biology of Microorganisms. 13th Ed. Pearson
Mancuso G et al (2005) Bacteroides fragilis – derived lipopolysaccharide produces cell activation and lethal toxicity via Toll-like receptor 4. Infect Immunity 73, 5620-5627
Mazmanian et al (2008) A microbial symbiosis factor prevents intestinal infammatory disease. Nature 453, 620-625
Ridaura VK et al (2013) Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 341, 1241214
Ríos-Covian et al (2016) Bacteroides fragilis metabolises exopolysaccharides produced by bifidobacteria. BMC Microbiol 16, 150
Ríos-Covian et al (2017) Shaping the metabolism of intestinal Bacteroides population through diet to improve human health. Front Microbiol 8, 376
Tan H et al (2019) Investigations of Bacteroides spp., towards next-generation probiotics. Food Res Internat 116, 637-644
Troy EB, Kasper DL (2010) Beneficial effects of Bacteroides fragilis polysaccharides on the immune system. Front Biosci 1, 15:25-34.
Ulsemer P et al (2012)a Preliminary safety evaluation of a new Bacteroides xylanisolvens isolate. Appl Env Microbiol 78, 528-535
Ulsemer P et al (2012)b Safety and tolerance of Bacteroides xylanisolvens DSM 23964 in healthy adults. Benef Microb 3, 99-111
Ulsemer P et al (2012)c Safety assesment of the commensal strain Bacteroides xylanisolvens DSM 23964. Regul Toxicol Pharmacol 62, 336-346
Van Nood E (2013) Duodenal infusion of donor feces for recurrent Clostridium difficile. New Eng J Medicine 368, 407-415
Wexler HA (2007) Bacteroides: the Good, the Bad, and the Nitty-Gritty. Clin Microbiol Rev 20, 593-621
Wexler AG, Goodman AL (2017) An insider’s perspective: Bacteroides as a window into the microbiome. Nat Microbiol 2, 17026
Wikipedia contributors. Bacteroides [Internet]. Wikipedia, The Free Encyclopedia, 2019 March 19
Xu J et al (2003) A genomic view of the human – Bacteroides thetaiotaomicron symbiosis. Science 299, 2074-2076

Plastic-eating bacteria
25th December 2018
Translated from the original article in Catalan.
Plastic ocean
We humans are destroying the planet Earth. Besides climate change (there are still ignorant people who do not believe it), the depletion of natural resources and the massive extinction of animal and plant species, one of the most visual effects is the coverage of the planet with rubbish. Since 71% of the surface is marine, most of the non-degrading waste finishes in the sea. In the oceans there are already large expansions covered by floating debris, especially plastics, called “plastic islands” (Figure 1). In the North Pacific area, where different sea currents come together, the “island” reaches 1500 km of radius, with plastics up to 200 meters deep, and continues to grow. There is more information of it, and also about the environmental consequences, in the Wikipedia article Great Pacific garbage patch.

Figure 1. Small portion of the Great Pacific Garbage Patch (From oceanandreserveconservationalliance.com)
PET plastics
Although there are many types of plastics, one of the most used and most abundant in waste and “plastic islands” is polyethylene terephthalate, known as PET or PETE (Figure 2). It is a type of thermoplastic polymer, vulgarly plastic, which belongs to the so-called polyesters, and is obtained by synthesis from petroleum. It is harmless, very resistant and lightweight and has multiple applications (Figure 3). Counting only bottles of PET for refreshing beverages, 1 million of them per minute are sold in the world. It is a recyclable material (see Pet bottle recycling in Wikipedia) but very resistant to biodegradation. In nature it can last some hundreds of years.
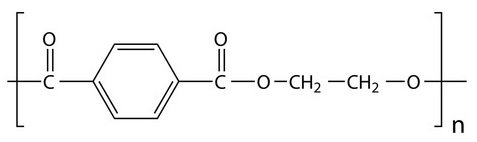
Figure 2. PET, polyethylene terephthalate.

Figure 3. Several applications of PET (From http://www.technologystudent.com).
PET is “eaten” by Ideonella sakaiensis
I. sakaiensis (Figure 4) are bacteria with rod shape, gram-negative, non esporulate aerobic heterotrophic, mobile with a flagellum, and catalase (+) and oxidase (+) (Tanasupawat et al 2016). They grow at neutral pH and are mesophilic, with optimum at 30-37°C. They belong to the phylogenetic group of betaproteobacteria, which include, besides many others, the known Neisseria (gonorrhoea and meningitis) and the nitrifying Nitrosomonas.

Figure 4. Scanning electron microscope images (false colour) of Ideonella sakaiensis cells grown on PET film for 60 h (From Yoshida et al 2016).
The 201-F6 strain, the first of the new species I. sakaiensis, was isolated from a landfill and identified in 2016 by a Japanese group of the Kyoto Institute of Technology that looked for bacteria using plastic as carbon source, from samples of remains of PET bottles (Yoshida et al 2016). They saw that these bacteria adhere to a low-grade PET film and can degrade it, by means of two enzymes characterized by these authors: a PETase and a MHETase, which produce terephthalic acid and ethylene glycol acid (Figure 5), which are benign environmental substances and that the bacteria can be metabolized. A colony of I. sakaiensis completely degraded a low-grade PET bottle in 6 weeks. High-grade PET products need to be heated to weaken them before the bacteria can degrade them. This is the first bacterium found as a PET degrader, and uses it as the only carbon source and energy source. Since PET has existed only for 70 years, these bacteria should have evolved in this short period until being able to degrade PET in a few weeks, instead of hundreds of years in nature (Sampedro 2016).
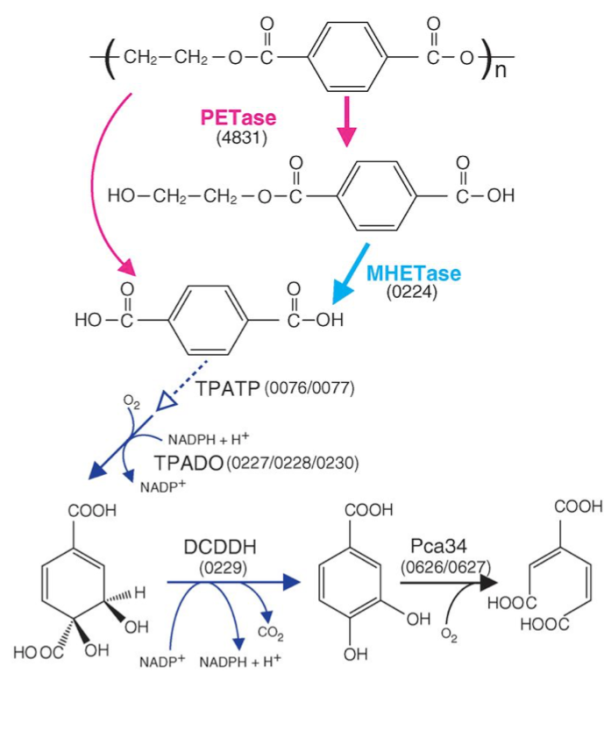
Figure 5. Predicted metabolic pathway of PET degradation by I. sakaiensis: extracellular PETase hydrolyses PET giving monohydroxyethyl terephthalic (MHET) and terephthalic acid (TPA). MHETase hydrolyses MHET to TPA and ethylene glycol (EG). The TPA is incorporated through a specific transporter (TPATP) and is catabolized to cyclohexadiene and this to protocatechuic acid (PCA) by the DCDDH. Finally, the PCA ring is cut by a PCA 3.4 dioxygenase with oxygen, as known for degradation of phenolic compounds and other xenobiotics. The numbers in parentheses are the ORF of the corresponding genes (From Yoshida et al 2016).
Previously, only some tropical microfungi (Fusarium solani) were known to degrade PET, and they also excreted esterases. In this case, Fusarium would be used to modify the polyester fabric, to achieve more hydrophilic and easier to work (Nimchua et al 2008). It is important to remember the structural similarity of synthetic PET fabrics (Figure 3) to those of natural fibre such as cotton, since these contain cutin, which is a polyester, a waxy polymer from the external parts of the plants. Therefore, the enzymes of Fusarium or Ideonella must be relatively similar to those that were already in nature long before the plastics were invented.
Recent genetic improvement of the enzyme PETase of Ideonella sakaiensis
In order to better understand the function and specificity of the PETase, a group of American and British researchers have recently characterized the structure of this enzyme (Austin et al 2018), mainly by high resolution X-ray crystallography, comparing it with a homologous cutinase obtained from actinobacteria Thermobifida fusca. The main differences between the two have been a greater polarization in the surface of the PETase (pI 9.6) than in the cutinase (pI 6.3), and on the other hand (Figure 6), a greater width of the active-site cleft in the case of PETase of I. sakaiensis. The cleft widening would be related with an easy accommodation of aromatic polyesters such as PET.
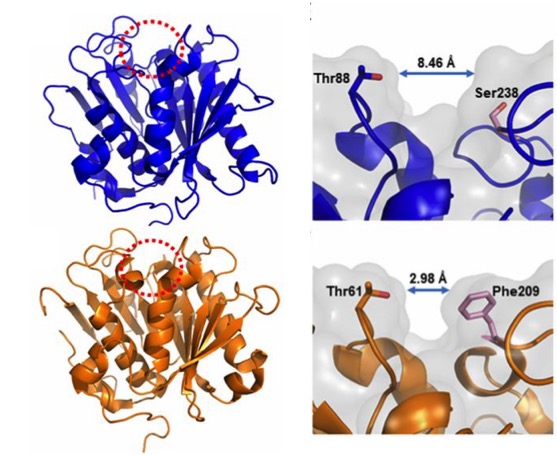
Figure 6. Compared structures (left) of the PETase of I. sakaiensis (above) and the cutinase of actinobacterium Thermobifida fusca (below), obtained by high resolution X-ray crystallography (0.92 Å). The active-site cleft is marked with a red dotted circle. Details (right) of the active site with different cleft widths in the PETase of I. sakaiensis (above) and the cutinase of T. fusca (below) are shown. (From Austin et al 2018).
Hypothesizing that the structure of the active site of the PETase would have resulted from a similar cutinase in an environment with PET, Austin et al (2018) proceeded to make mutations in the PETase active-site to make it more similar to cutinase and obtained a double mutant S238F/W159H which theoretically would make the entry of the active site closer (Figure 6). But their surprise was capital when they saw that the mutant degraded the PET better (an improvement of 20%), with an erosion of the PET film (Figure 7 C) even greater than the original PETase (Figure 7B). The explanation was that mutant changes in amino acid residues favoured PET intake in the active site, despite making a closest cleft (Austin et al 2018).
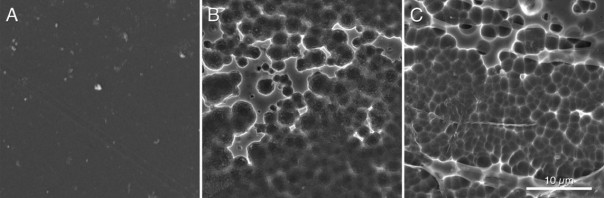
Figure 7. Scanning electronic microscopy images of a piece of PET without microorganisms (A), after incubating 96 h with PETase of the I. sakaiensis 201-F6 (B), and with PETase of the double-mutant S238F/W159H (C) (From Austin et al 2018).
In addition, these authors have shown that this PETase degrades also other similar semi aromatic polyesters, such as polyethylene-2,5-furonicarboxylate (PEF), and therefore this enzyme can be considered an aromatic polyesterase, but it does not degrade aliphatic ones.
The conclusion of their work is that protein engineering is feasible in order to improve the performance of PETase and that we must continue to deepen in the knowledge of their relationships between structure and activity for the biodegradation of synthetic polyesters (Austin et al 2018).
Other plastic-eating microbes ?
The discovery of I. sakaiensis has been very important for the possibility of establishing a rapid recycling process for PET, but it is not the first organism that has been found as plastic consumer. By the way, we can see the formulas of the main plastics derived from petroleum in Figure 8.
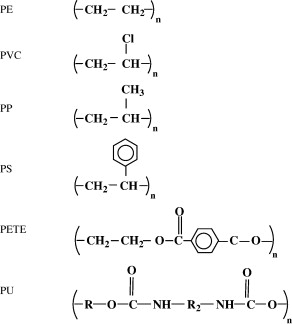
Figure 8. Formulas of the most common petroleum plastics: polyethylene (PE), polyvinyl chloride (PVC), polypropylene (PP), polystyrene (PS), polyethylene terephthalate (PET or PETE) and polyurethane (PU) (From Shah et al 2008).
Reviewing the bibliography, we see that many cases of plastic degrading microorganisms have been described (Shah et al 2008), especially polyethylene, polyurethane and PVC: various Pseudomonas, Rhodococcus and Comamonas among bacteria, and some Penicillium, Fusarium and Aspergillus between fungi.
Among the polyurethane consumers, mushrooms are highlighted (Howard 2002), and especially the plants endophyte Pestalotiopsis microspora, which can use polyurethane as the only source of carbon (Russell et al 2011).
On the other hand, the ability of the mealworms, the larval forma of the darkling beetle Tenebrio molitor, to chew and degrade the polystyrene foam is well known (Yang et al 2015). Fed only with the PS, these larvae degrade it completely in relatively short periods. As expected, the degradation of the PS is carried out by the intestinal bacteria of the animal (Figure 9). It has been demonstrated because degradation stops when administering antibiotics to the larva (Yang et al 2015). One of the isolated bacteria that has been shown to degrade PS is Exiguobacterium, from Bacillales group, but it is not the only one. In fact, when performing studies of metagenomics from gut of larvae eating PS, a large variety of bacteria have been found, and these vary depending on the kind of plastic, since the degradation of polyethylene has also been seen. Some of the bacteria with DNA found as predominant would be the enterobacteria Citrobacter and Kosakonia. It seems that the intestinal microbiota of Tenebrio is modified and adapted to the different ingested plastics (Brandon et al 2018).
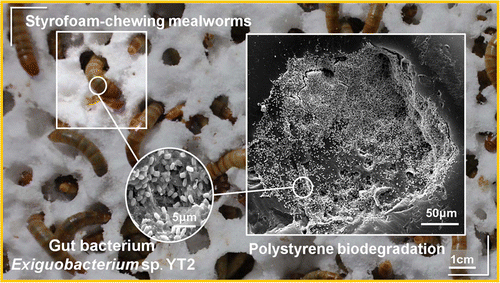
Figure 9. Biodegradation of polystyrene by the intestinal bacteria of Tenebrio, the mealworm (Yang et al 2015).
Finally, as we see the microbial biodegradation of non-biodegradable or recalcitrant plastics should not surprise us, since on the one hand, there are natural “plastics” such as polyhydroxybutyrate or polylactic acid that are easily degradable (Shah et in 2008), and on the other hand the adaptive capacity of the microorganisms to be able to break the most recalcitrant chemical bonds is very large. Microbes evolve rapidly, and acquire better strategies to break the plastics made by humans (Patel 2018). We have seen in this case the degradation of PET, which in less than 70 years some microbes have already found a way to take advantage of it.
The problem is that we are generating too much plastic waste in no time and the microorganisms have not had time yet to degrade them. It is clear that we will have to help our microbial partners, not generating more degrading polymers, and recycling and degrading them, by using these same degrading microbes, among other ways.
Bibliography
Austin HP et al (2018) Characterization and engineering of a plastic-degrading aromatic polyesterase. Proc Nat Acad Sci 115, 19, E4350-E4357
Brandon AM et al (2018) Biodegradation of Polyethylene and Plastic Mixtures in Mealworms (Larvae of Tenebrio molitor) and Effects on the Gut Microbiome. Environ Sci Technol 52, 6526-6533
Griggs MB (2017 april 24) These caterpillars chow down on plastic bags. Popular Science. http://www.popsci.com
Howard GT (2002) Biodegradation of polyurethane: a review. Int Biodeterior Biodegrad 42, 213-220
https://en.wikipedia.org/wiki/Great_Pacific_garbage_patch
https://en.wikipedia.org/wiki/PET_bottle_recycling
https://en.wikipedia.org/wiki/Polyethylene_terephthalate
Patel NV (2018 april 17) Scientists stumbled upon a plastic-eating bacterium – then accidentally made it stronger. Popular Science. http://www.popsci.com
Russell JR et al (2011) Biodegradation of polyester polyurethane by endophytic fungi. Appl Environ Microbiol 77, 17, 6076-6084
Sampedro J (2016 marzo 10) Descubierta una bacteria capaz de comerse un plástico muy común. El País
Shah AA et al (2008) Biological degradation of plastics: a comprehensive review. Biotechnol Adv 26, 246-265
Tanasupawat et al (2016) Ideonella sakaiensissp. nov., isolated from a microbial consortium that degrades poly(ethylene terephtalate). Int J Syst Evol Microbiol 66, 2813-2818
Yang et al (2015) Biodegradation and mineralization of polystyrene by plastic-eating mealworms: Part 2. Role of gut microorganisms. Environ Sci Technol 49, 12087-12093
Yoshida et al (2016) A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 351,1196–1199

Lactic acid bacteria of beers: the bad guys and the good ones
28th October 2018
It is not easy to “live” in the beer
In principle, lactic acid bacteria (LAB) and many other bacteria and generally most microorganisms, do not have it easy to survive in beer or other alcoholic beverages such as wine. This is one of the main reasons why wines and beers have been from ancient times the safest ways to drink hygienically something similar to water and that it was not contaminated, apart from boiled waters, such as tea and other herbal infusions.
The reasons for the difficult survival of microorganisms in beer are ethanol, the pH quite acidic (around 4), the lack of nutrients due to the fact that the yeasts have assimilated them, the little dissolved oxygen, the high concentration of carbon dioxide (0.5% by weight / volume) and the presence of humulone derived compounds (Figure 1) of hops: iso-alpha-acids, up to 50 ppm, which are microbiocides. All these obstacles make it very difficult for any microorganism to thrive. The most susceptible beers of unwanted microbial growth are those where some of the mentioned obstacles are dampened: beers with a higher pH of 4.5, or with little ethanol or little CO2, or with added sugars – which are nutrients -, or with little amount of compounds derived from hops (Vriesekoop et al 2012).
Figure 1. Humulone (left) of the hop is degraded during beer elaboration to isohumulone (right) and other iso-alpha-acids, which are compounds bitter and microbiocides (Wikipedia; Sakamoto & Konings 2003)
The acid pH of the beer (slightly higher than the wine) inhibits many of the best-known pathogens (Figure 2). And the cases we see that could grow at this pH near 4 are inhibited by other factors such as ethanol.
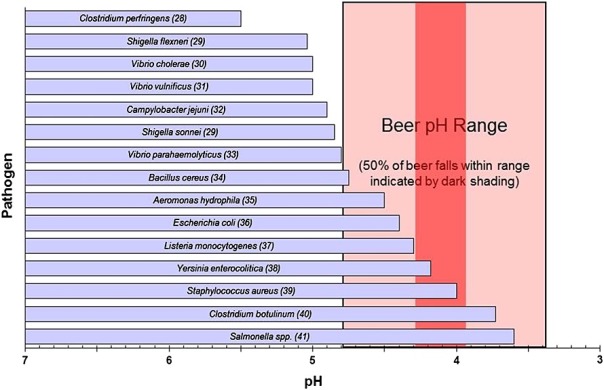
Figure 2. Range of acid pH for the growth of various bacteria, compared to the typical beer pH (Menz et al 2009).
The “bad” lactic acid bacteria of beer
Despite what we have just seen, some bacteria, particularly some LAB, have been able to adapt evolutionarily to the strict beer conditions, and they can survive and spoil them. In particular, the most frequent harmful species against the quality of beers are Lactobacillus brevis and Pediococcus damnosus (Figure 3). The first is the most frequent, and it can give tastes and undesired aromas, as well as turbidity to the final product. P. damnosus has the advantage of growing at low temperatures, and it can also produce undesired aromas, such as diacetyl (Vriesekoop et al 2012). Some Pediococcus and Lactobacillus may adhere to yeast, inducing them to sediment, which delays fermentation (Suzuki 2011).
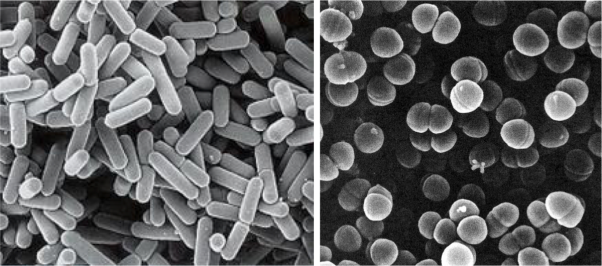
Figure 3. Lactobacillus brevis (left) and Pediococcus damnosus (right) at the electronic scanning microscope.
Some Pediococcus may also be responsible for the appearance of biological amines in some beers, at risk for the consumer. Amines in a certain concentration are toxic, they may be present in some fermented foods such as cheese, cold meat and alcoholic beverages such as wines and beers, and are produced by decarboxylation of amino acids by LAB. The level of tyramine and other amines has been used as a measure of quality in some Belgian beers made with LAB (Loret et al 2005).
Apart from these LAB, other bacteria related to problems of beer contamination are acetic acid bacteria such as Acetobacter, typically associated with oxygen intake in packaging or distribution. Other harmful bacteria are some enterobacteria, such as Shimwellia pseudoproteus or Citrobacter freundii, which proliferate in the early stages of fermentation, and produce butanediol, acetaldehyde and other unwanted aromatic compounds (Vriesekoop et al 2012). Other harmful bacteria for beer, especially when bottled, are Pectinatus and Megasphaera, which are strict anaerobes, of the clostridial family, and can produce hydrogen sulphide and short chain fatty acids, all of them unpleasant (Suzuki 2011 ).
The “good” lactic acid bacteria of beer
LAB are well known for being some of the microbes that most benefits contribute to the food production, on the one hand as an economic means of preserving food, and on the other hand to improve their quality and organoleptic characteristics. That’s why they are the main agents of fermented foods, along with yeasts. We have seen some of the LAB’s food benefits in other posts in this blog: prehistoric cheeses, or breast milk microbiota, and even wine bacteria.
Therefore, LAB also have a good role in the production of beers: in particular, as we will see below, in the production of acidified malt, and in some peculiar styles of beer such as the Belgian Lambic and the Berliner Weissbier.
As you know, malt is the raw material for making beer. The cereal is subjected to the malting process, where cereal grains, mainly barley, are germinated, the enzymes hydrolyse the starch into sugars, and all of this is then heated obtaining the must, the substrate solution which will be fermented by the yeasts ferment, producing ethanol and carbon dioxide.
The acidification of the malt, that is, with a lower pH, has the advantages of activating many important enzymes in malting, giving a lower viscosity to the malt and therefore to the final beer. Although adding mineral acids or commercial lactic acid can achieve acidification, it is often recommended or legislated a biological acidification, which is achieved by adding LAB. The use of LAB starter cultures is a relatively new process and in addition to the commented benefits on the quality of the malt, it has been shown to also inhibit unwanted molds that are a real problem in malting and that can give mycotoxins. The compounds produced by LAB that can inhibit the fungi are the same lactic acid and the consequent pH drop, bacteriocins, hydrogen peroxide, and other compounds not well known as perhaps some peptides (Lowe & Arendt 2004).
The most commonly LAB strains used to acidify malt are Lactobacillus amylolyticus previously isolated from the same malt. These strains are moderately thermophile, resistant to compounds derived from humulone, and they have the advantage of being amylolytic in addition to producing lactic acid, which lowers the pH (Vriersekoop et al 2012).
Beers with LAB participating in the fermentation, such as Lambic and Berliner Weissbier styles, belong to the type of spontaneous fermentation beers. The other types of controlled fermentation beers are the best-known Ale and Lager, both inoculated with specific yeasts. Ale beers are those of high fermentation, where Saccharomyces cerevisiae yeast used tends to remain on the surface and the fermentation temperature is above 15-20ºC. Lager ones are those of low fermentation, originally from Bavaria, where yeast S. pastorianus (S. carlsbergensis) tends to settle at the bottom of the fermenter and the temperature is between 7 and 13ºC.
Belgian Lambic beer
Traditional Belgian beers (in Dutch lambiek or lambik) are known for their sensorial characteristics due to LAB activity. They are traditional in Brussels itself and in the neighbouring region of Pajottenland, in the Zenne river valley, in the Flemish Brabant on the SW of the Belgian capital. One of the villages in this valley is Lembeek, which could be the origin of the name of this beer.
These beers of spontaneous fermentation represent the oldest style of making beer in the developed world, for some centuries. For a few years now (since around 2008), similar beers are made in the USA, called “American coolship ales” (Ray 2014).
Lambic beer is made with barley malt and a minimum of 30% of non-malted wheat. The cones of a special hops, completely dried and aged for 3 years, are added to the must. They are added not for their aroma or bitterness, but rather as antimicrobial, to prevent above all, the growth of gram-positive pathogenic bacteria in the fermentation broth.
Also to avoid these contaminants and to promote the microbiota typical of the Lambic fermentation, these beers are brewed only between October and May, since in summer there are too many harmful microorganisms in the air that could spoil the beer, and it is necessary to lower the temperature after boiling. Boiling of the must is done intensively, with an evaporation of 30%.
After boiling, the broth is left in open deposits, and in this way the microorganisms of the air present in the fermentation rooms of the brewery (usually at the top of the building) are acquired, and of the outside air, since the tradition says that the windows must be left open. It is assumed that the captured microbes are specific to the Zenne Valley. These open deposits are the koelschip in Dutch (coolship in English), like swimming pools (Figure 4). Being well open, with a lot of surface (about 6 x 6 m) and shallow depth (about 50 cm), they favour the collection of microbes from the room and from the outside. Another purpose of this form is the fastest cooling of boiled broth to start fermentation. They can be made of wood, copper, or stainless steel more recently.

Figure 4. Koelschip (in Dutch) or coolship in English, the open deposits, as swimming-pools, where the Lambic beer process begins (Brasserie Cantillon, Brussels).
The “inoculated” broth in this spontaneous way is left only one night in the coolship, and on the following day this must is pumped into fermentation tanks where there will stay a year, during which the sugar content will go down, up to about 30 g/L. Then it is transferred to oak barrels, previously used for sherry or port, and there it can be left for another two years, at temperatures of 15-25ºC. Some barrels are the same used since 100 years ago. The final product is a cloudy beer, with a pale yellow, very little CO2, dry, acidic, with about 6-8º of ethanol. It reminds a bit like the sherry and especially the cider, and with a slightly bitter taste (Jackson 1999).
In this long process of fermentation, up to 3 years, of course there is a diversity in the composition of the microbial population. In a first phase there is a certain predominance of Kloeckera yeasts and especially enterobacteria during the first month. After 2 months, Pediococcus damnosus and Saccharomyces spp. predominate, and alcoholic fermentation begins. After 6 months of fermentation the predominant yeast is Dekkera bruxellensis (Spitaels et al 2014), or what is the same, Brettanomyces (Kumara & Verachtert 1991), of which Dekkera is the sexual form.
Figure 5. Species of isolates in MRS and VRBG agar media, for lactic acid bacteria and enterobacteria respectively, during the process of making a Lambic beer. The number of isolates is given between brackets (Spitaels et al 2014).
We see (Figure 5) as in particular after 2 months the predominant bacterium is the LAB P. damnosus. It was appointed in the first studies as “P. cerevisiae“, but this name was finally not admitted because it included other species. The count of these in MRS is 104UFC per mL until the end of fermentation. Acidification seems to be rapidly taking place in the transition from the first stage to that of maturation, coinciding with the growth of P. damnosus, which produces lactic acid, although Dekkera/Brettanomyces and acetic acid bacteria also contribute to the acidification (Spitaels et to 2014).
In other trials with the American coolship ales (ACA) of Lambic style, Lactobacillus spp. have also been found, and in a metagenomic study (Bukolich et al 2012) of these ACA, DNA of several Lactobacillales has been detected. At the end of the process, a predominance of Pediococcus (Figure 6, panel C) was also observed. In the same figure in panel A we observe how the predominant unicellular fungus is also Dekkera/Brettanomyces.
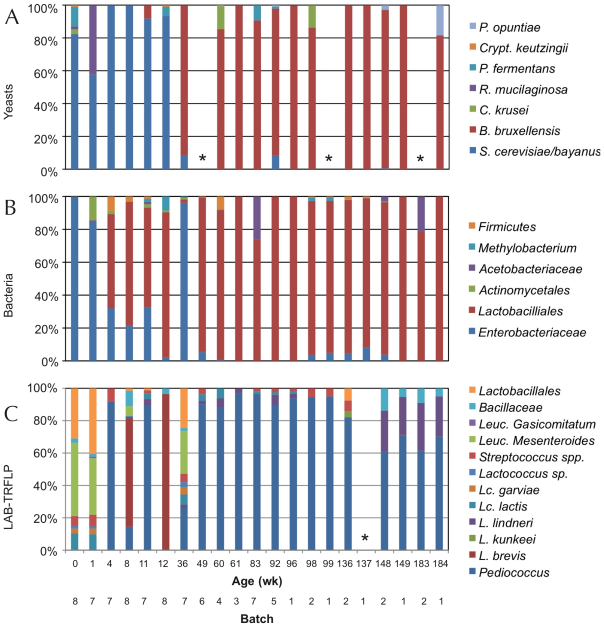
Figure 6. TRFLP analysis (polymorphisms of lengths of PCR-amplified terminal restriction fragments) of total DNA extracted from the fermentation samples of ACA beers (similar to Lambic) during 3 years, using primers for: ITS1/ITS4 of 26S rDNA for yeasts (panel A), 16S rDNA for bacteria (panel B), and specific ones for LAB (panel C). Samples marked with * did not give amplification (Bukolich et al 2012).
Lambic derived beers: Gueuze, Faro, fruity and others
The basic Lambic, which is difficult to purchase, is only found in a few Brussels cafes and the production area. In fact, Lambic is the basis for elaborating the others, much more common to consume:
The Faro is a Lambic sweetened with brown sugar and sometimes with spices.
The fruity Lambic are those that have been added whole fruits or fruit syrup. They can be with bitter cherry (kriek), which are the most traditional, or with raspberry, peach, grapes, strawberry, and sometimes also apple or pineapple or apricot or other.
And finally, the Gueuze, which are sparkling and easy to find. They are made by mixing young Lambics (from 6 months to 1 year) with other more mature ones (2-3 years) in thick glass bottles similar to those of champagne or cava and left for a second fermentation with the remaining sugars from the young Lambic. This would have been begun by a mayor of Lembeek in 1870 that owned a brewery and applied the fermentation techniques in the bottle that had been successful in the Champagne some years before (Cervesa en català 2012). The word Gueuze can have the same etymological origin as gist(yeast in Flemish) and it could also refer to the fact that it produces bubbles of CO2, that is, gas (Jackson 1999). However, another historical version would be that this beer was called “Lambic de chez le gueux” (Welsh from poor people) because the mentioned mayor of Lembeek had similar socialist ideas to those of the “Parti des Gueus” founded by the Calvinists from Flanders in the 16th century to fight against the Spanish empire. And since beer is feminine in French, the gueuxfeminine is gueuze, here it is.
In this refermentation in the bottle the populations of Dekkera/Brettanomycesand LAB are maintained, although other unicellular fungi such as Candida, Hansenula, Pichia or Cryptococcus (Verachtert & Debourg 1999) appear in limited numbers.

Figure 7. Several beer Gueuze and fruity Lambic, mostly Belgian (from www.swanbournecellars.com.au/).
The Berliner Weissbier (Figure 8) is another beer relatively similar to Lambic ones. It is also brewed with an important part of wheat must, it is cloudy, acidic and with 3% ethanol. It is traditional in Berlin and the north of Germany, made from the s. XVI and the most popular alcoholic beverage in Berlin until the end of the s. XIX. It was called the “northern champagne” by the Napoleon’s soldiers. Spontaneous fermentation of must involves a mixture of Dekkera/Brettanomyces, Saccharomycesand hetero-fermentative Lactobacillus.

Figure 8. Berliner Weisse beer (from G-LO, @boozedancing wordpress).
Beers similar to Lambic brewed in Spain
In the same way that the commented American Coolship Ales, Lambic style beers are also made in many other countries and, in the case of Spain, coinciding with the boom of artisanal beers, they are also elaborated, especially the fruity Lambic ones. According to the Birrapedia website, 6 of these are currently being processed, all of which are cherries. Two of them are made in Lleida, one in Barcelona, one in Alicante, one in the Jerte valley, and another in Asturias.
Resistance of lactic acid bacteria from beer to hop compounds
Lactobacillus and Pediococcus, both bad and good we have seen, and other contaminating bacteria of beers, have the ability to withstand hop compounds, which, as we have seen, are natural microbiocides. This resistance can be due to various defence systems, both active and passive (Sakamoto & Konings 2003). The active systems include efflux pumps, such as HorA and HorC, which carry the iso-alpha-acids (Figure 1) out the cell. HorA does it with ATP consumption, and HorC using the proton driving force (Figure 9). The corresponding genes horA and horC were originally found in L. brevis, but later they were also found in L. lindneri, L. paracollinoides and in the best known P. damnosus(Suzuki et al., 2006).
Curiously, HorA shows a resemblance of 54% to OmrA, a membrane transporter of Oenococcus oeni, related to the tolerance of this bacterium from wine to ethanol and other stressors (Bourdineaud et al 2004) (See some more about O. oeni in my post on the bacteria of the vine and the wine). Therefore, it is probable that HorA also has functions of exclusion of other compounds aside from those of the hops. It has been seen that these horAand horC resistance genes and their flanking regions are well preserved and have sequences almost identical to the different species that have them. Therefore, it is very likely that some have been acquired from others by means of horizontal gene transfer, by plasmids or transposons, as is usual in many other bacteria (Suzuki 2011).
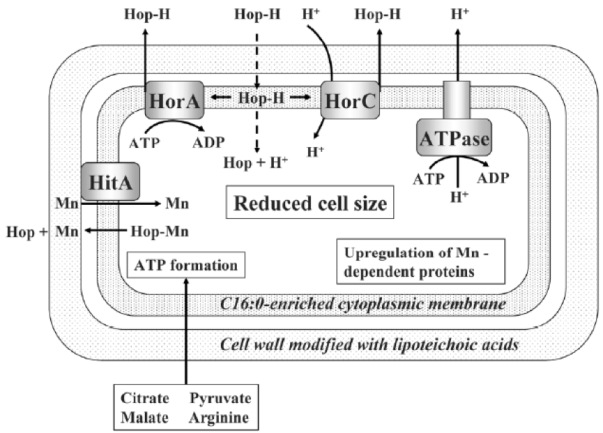
Figure 9. Mechanisms of resistance to hop compounds in Lactobacillus brevis (Suzuki 2011).
As we see in Figure 9, protons are pumped out by an ATPase, and the consumption of ATPs is compensated by forming it thanks to the consumption of substrates such as citrate, malate, pyruvate or arginine. Another mechanism of resistance, passive in this case, is the modification of the composition of membrane fatty acids, with the addition of more saturated ones, such as C16:0, which reduces the membrane fluidity and makes it difficult the entrance of the hop compounds. This also reminds us of the changes in membrane of O. oeni related to the resistance to ethanol (Margalef-Català et al 2016). The cell wall also changes its composition in the presence of the hop alpha-iso-acids, increasing the amount of high molecular weight lipoteichoic acid, which would also be a barrier. We also see (Figure 9) how hop compounds can lower the intracellular levels of Mn2+, and then a greater synthesis of Mn-dependent proteins is observed, and a greater capture of Mn2+ from outside. Finally, cells of L. brevis reduce their size when they are in beer (Figure 10), probably in order to decrease the extracellular surface, thus minimizing the effect of external toxic compounds (Suzuki 2011).
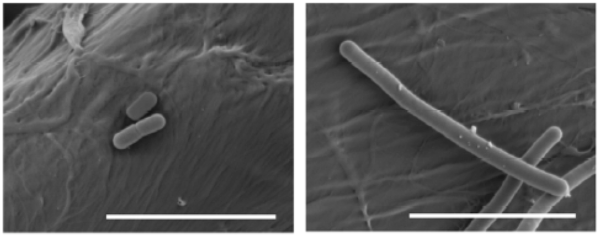
Figure 10. Effects of beer adaptation (left) in the size of Lactobacillus brevis cells compared to well grown cells in rich media MRS (right). The bars are 5 mm (Suzuki 2011).
All these mechanisms have been studied in L. brevis strains harmful to beer, but it is assumed that the resistance of beneficial bacteria from Lambic and others would be due to the same mechanisms, since they are of the same bacterial species.
As a conclusion to all said, we see that LAB have outstanding roles as beneficial in various aspects of brewery and malting, despite their most known role of harmful in the processing of the most common beers.
Bibliography
Birrapedia (seen 18 august 2018) Cervezas de tipo Fruit Lambic elaboradas en España. https://birrapedia.com/cervezas/del-tipo-fruit-lambic-elaboradas-en-espana
Bokulich NA et al (2012) Brewhouse resident microbiota are responsible for multi-stage fermentation of American Coolship Ale. PLoS One, 7, e35507
Bourdineaud J et al (2004) A bacterial gene homologous to ABC transporters protect Oenococcus oeni from ethanol and other stress factors in wine. Int J Food Microbiol 92, 1-14.
Cervesa en català (2012) Fitxes de degustació – Timmermans Gueuze Tradition http://cervesaencatala.blogspot.com.es/2012/06/fitxes-de-degustacio-timmermans-gueuze.html
Jackson, Michael (1999) Belgium’s great beers. Beer Hunter Online, July 30, 1999
Kumara HMCS & Verachtert H (1991) Identification of Lambic super attenuating micro-organisms by the use of selective antibiotics. J Inst Brew 97, 181-185
Loret S et al (2005) Levels of biogenic amines as a measure of the quality of the beer fermentation process: data from Belgian samples. Food Chem 89, 519-525
Lowe DP & Arendt EK (2004) The use and effects of lactic acid bacteria in malting and brewing with their relationships to antifungal activity, mycotoxins and gushing: a review. J Inst Brew 110, 163-180
Margalef-Català et al (2016) Protective role of glutathione addition against wine-related stress in Oenococcus oeni. Food Res Int 90, 8-15
Menz G et al (2009) Pathogens in beer, in Beer in Health and Disease Prevention, (Preedy, V. R. Ed.), 403–413, Academic Press, Amsterdam
Ray AL (2014) Coolships rising: the next frontier of sour beers in the U.S. First we feast 27 feb 2014
Sakamoto K & Konings WN (2003) Beer spoilage bacteria and hop resistance. Int J Food Microbiol 89, 105-124
Spitaels F et al (2014) The microbial diversity of traditional spontaneously fermented lambic beer. PLOS One 9, 4, e95384
Suzuki K et al (2006) A review of hop resistance in beer spoilage lactic acid bacteria. J Inst Brew 112, 173-191
Suzuki K (2011) 125th Anniversary Review: microbiological instability of beer caused by spoilage bacteria. J Inst Brew 117, 131-155
The Beer Wench (2008) My obsession with wild beers. Nov. 20, 2008 https://thecolumbuswench.wordpress.com/tag/lambic/
Verachtert H & Debourg A (1999) The production of gueuze and related refreshing acid beers. Cerevisia, 20, 37–41
Vriesekoop F et al (2012) 125th Anniversary review: Bacteria in brewing: the good, the bad and the ugly. J Inst Brew 118, 335-345

Bacillus as probiotics
12th August 2017
The probiotics
Probiotics are living microorganisms that, when ingested in adequate amounts, can have a positive effect on the health of guests (FAO / WHO 2006; World Gastroenterology Organization 2011, Fontana et al., 2013). Guests can be humans but also other animals. Lactic acid bacteria, especially the genus Lactobacillus and Bifidobacterium, both considered as GRAS (Generally recognized as safe), are the microbes most commonly used as probiotics, but other bacteria and some yeasts can also be useful. Apart from being able to be administered as medications, probiotics are commonly consumed for millennia as part of fermented foods, such as yoghurt and other dairy products (see my article “European cheese from 7400 years ago..” “December 26th, 2012). As medications, probiotics are generally sold without prescription, over-the-counter (OTC) in pharmacies.
I have already commented on the other posts of this blog the relevance of probiotics (“A new probiotic modulates microbiota against hepatocellular carcinoma” August 24th, 2016), as well as the microbiota that coexists with our body (“Bacteria in the gut controlling what we eat” October 12th, 2014; “The good bacteria of breast milk” February 3rd, 2013) and other animals (“Human skin microbiota … and our dog” December 25th, 2015; “The herbivore giant panda …. and its carnivore microbiota” September 30th, 2015).
Besides lactic acid bacteria and bifidobacteria, other microorganisms that are also used to a certain extent as probiotics are the yeast Saccharomyces cerevisiae, some strains of Escherichia coli, and some Bacillus, as we will see. Some clostridia are also used, related to what I commented in a previous post of this blog by March 21st, 2015 (“We have good clostridia in the gut ...”).
The Bacillus
In fact, Bacillus and clostridia have in common the ability to form endospores. And both groups are gram-positive bacteria, within the taxonomic phylum Firmicutes (Figure 1), which also includes lactic acid bacteria. However, bacilli (Bacillus and similar ones, but also Staphylococcus and Listeria) are more evolutionarily closer to lactobacillalles (lactic acid bacteria) than to clostridia ones. The main physiological difference between Clostridium and Bacillus is that the first are strict anaerobes while Bacillus are aerobic or facultative anaerobic.
Figure 1. Phylogenetic tree diagram of Gram-positive bacteria (Firmicutes and Actinobacteria). Own elaboration.
Bacterial endospores (Figure 2) are the most resistant biological structures, as they survive extreme harsh environments, such as UV and gamma radiation, dryness, lysozyme, high temperatures (they are the reference for thermal sterilization calculations), lack of nutrients and chemical disinfectants. They are found in the soil and in the water, where they can survive for very long periods of time.
Figure 2. Endospores (white parts) of Bacillus subtilis in formation (Image of Simon Cutting).
Bacillus in fermented foods, especially Asian
Several Bacillus are classically involved in food fermentation processes, especially due to their protease production capacity. During fermentation, this contributes to nutritional enrichment with amino acids resulting from enzymatic proteolysis.
Some of these foods are fermented rice flour noodles, typical of Thailand and Burma (nowadays officially Myanmar). It has been seen that a variety of microorganisms (lactic acid bacteria, yeasts and other fungi) are involved in this fermentation, but also aerobic bacteria such as B. subtilis. It has been found that their proteolytic activity digests and eliminates protein rice substrates that are allergenic, such as azocasein, and therefore they have a beneficial activity for the health of consumers (Phromraksa et al. 2009).
However, the best-known fermented foods with Bacillus are the alkaline fermented soybeans. As you know, soy (Glycine max) or soya beans are one of the most historically consumed nourishing vegetables, especially in Asian countries. From they are obtained “soy milk”, soybean meal, soybean oil, soybean concentrate, soy yogurt, tofu (soaked milk), and fermented products such as soy sauce, tempeh, miso and other ones. Most of them are made with the mushroom Rhizopus, whose growth is favoured by acidification or by direct inoculation of this fungus. On the other hand, if soy beans are left to ferment only with water, the predominant natural microbes fermenting soy are Bacillus, and in this way, among other things, the Korean “chongkukjang” is obtained, “Kinema” in India, the “thua nao” in northern Taiwan, the Chinese “douchi”, the “chine pepoke” from Burma, and the best known, the Japanese “natto” (Figure 3). Spontaneous fermentation with Bacillus gives ammonium as a by-product, and therefore is alkaline, which gives a smell not very good to many of these products. Nevertheless, natto is made with a selected strain of B. subtilis that gives a smoother and more pleasant smell (Chukeatirote 2015).
These foods are good from the nutritional point of view as they contain proteins, fibre, vitamins, and they are of vegetable or microbial origin. In addition, the advertising of the commercial natto emphasizes, besides being handmade and sold fresh (not frozen), its probiotic qualities, saying that B. subtilis (Figure 4) promotes health in gastrointestinal, immunologic, cardiovascular and osseous systems (www.nyrture.com). They say the taste and texture of natto are exquisite. It is eaten with rice or other ingredients and sauces, and also in the maki sushi. We must try it !

Figure 3. “Natto”, soybeans fermented with B. subtilis, in a typical Japanese breakfast with rice (Pinterest.com).
Figure 4. Coloured electronic micrograph of Bacillus subtilis (Nyrture.com).
Bacillus as probiotics
The endospores are the main advantage of Bacillus being used as probiotics, thanks to their thermal stability and to survive in the gastric conditions (Cutting 2011). Although Clostridium has also this advantage, its strict anaerobic condition makes its manipulation more complex, and moreover, for the “bad reputation” of this genus due to some well-known toxic species.
Unlike other probiotics such as Lactobacillus or Bifidobacterium, Bacillus endospores can be stored indefinitely without water. The commercial products are administered in doses of 10^9 spores per gram or per ml.
There are more and more commercial products of probiotics containing Bacillus, both for human consumption (Table 1) and for veterinary use (Table 2). In addition, there are also five specific products for aquaculture with several Bacillus, and also shrimp farms are often using products of human consumption (Cutting 2011).
For use in aquaculture, probiotic products of mixtures of Bacillus (B. thuringiensis, B. megaterium, B. polymixa, B. licheniformis and B. subtilis) have been obtained by isolating them from the bowel of the prawn Penaeus monodon infected with vibriosis. They have been selected based on nutrient biodegradation and the inhibitory capacity against the pathogen Vibrio harveyi (Vaseeharan & Ramasamy 2003). They are prepared freeze-dried or microencapsulated in sodium alginate, and it has been shown to significantly improve the growth and survival of shrimp (Nimrat et al., 2012).
As we see for human consumption products, almost half of the brands (10 of 25) are made in Vietnam. The use of probiotic Bacillus in this country is more developed than in any other, but the reasons are not clear. Curiously, as in other countries in Southeast Asia, there is no concept of dietary supplements and probiotics such as Bacillus are only sold as medications approved by the Ministry of Health. They are prescribed for rotavirus infection (childhood diarrhoea) or immune stimulation against poisoning, or are very commonly used as a therapy against enteric infections. However, it is not clear that clinical trials have been carried out, and they are easy-to-buy products (Cutting 2011).
Table 1. Commercial products of probiotics with Bacillus, for human consumption (modified from Cutting 2011).
| Product | Country where it is made | Species of Bacillus |
| Bactisubtil ® | France | B. cereus |
| Bibactyl ® | Vietnam | B. subtilis |
| Bidisubtilis ® | Vietnam | B. cereus |
| Bio-Acimin ® | Vietnam | B. cereus and 2 other |
| Biobaby ® | Vietnam | B. subtilis and 2 other |
| Bio-Kult ® | United Kingdom | B. subtilis and 13 other |
| Biosporin ® | Ukraine | B. subtilis + B. licheniformis |
| Biosubtyl ® | Vietnam | B. cereus |
| Biosubtyl DL ® | Vietnam | B. subtilis and 1 other |
| Biosubtyl I and II ® | Vietnam | B. pumilus |
| Biovicerin ® | Brazil | B. cereus |
| Bispan ® | South Korea | B. polyfermenticus |
| Domuvar ® | Italy | B. clausii |
| Enterogermina ® | Italy | B. clausii |
| Flora-Balance ® | United States | B. laterosporus * |
| Ildong Biovita ® | Vietnam | B. subtilis and 2 other |
| Lactipan Plus ® | Italy | B. subtilis * |
| Lactospore ® | United States | B. coagulans * |
| Medilac-Vita ® | China | B. subtilis |
| Nature’s First Food ® | United States | 42 strains, including 4 B. |
| Neolactoflorene ® | Italy | B. coagulans * and 2 other |
| Pastylbio ® | Vietnam | B. subtilis |
| Primal Defense ® | United States | B. subtilis |
| Subtyl ® | Vietnam | B. cereus |
| Sustenex ® | United States | B. coagulans |
* Some labelled as Lactobacillus or other bacteria are really Bacillus
Table 2. Commercial products of probiotics with Bacillus, for veterinary use (modified from Cutting 2011).
| Product | Animal | Country where it is made | Species of Bacillus |
| AlCare ® | Swine | Australia | B. licheniformis |
| BioGrow ® | Poultry, calves and swine | United Kingdom | B. licheniformis and B. subtilis |
| BioPlus 2B ® | Piglets, chickens, turkeys | Denmark | B. licheniformis and B. subtilis |
| Esporafeed Plus ® | Swine | Spain | B. cereus |
| Lactopure ® | Poultry, calves and swine | India | B. coagulans * |
| Neoferm BS 10 ® | Poultry, calves and swine | France | B. clausii |
| Toyocerin ® | Poultry, calves, rabbits and swine | Japan | B. cereus |
The Bacillus species that we see in these Tables are those that really are found, once the identification is made, since many of these products are poorly labelled as Bacillus subtilis or even as Lactobacillus (Green et al. 1999; Hoa et al. 2000). These labelling errors can be troubling for the consumer, and especially for security issues, since some of the strains found are Bacillus cereus, which has been shown to be related with gastrointestinal infections, since some of them produce enterotoxins (Granum & Lund 1997; Hong et al. 2005)
The probiotic Bacillus have been isolated from various origins. For example, some B. subtilis have been isolated from the aforementioned Korean chongkukjang, which have good characteristics of resistance to the gastrointestinal tract (GI) conditions and they have antimicrobial activity against Listeria, Staphylococcus, Escherichia and even against B. cereus (Lee et al. 2017).
One of the more known probiotics pharmaceuticals is Enterogermina ® (Figure 5), with B. subtilis spores, which is recommended for the treatment of intestinal disorders associated with microbial alterations (Mazza 1994).
Figure 5. Enterogermina ® with spores of Bacillus subtilis (Cutting 2011)
Bacillus in the gastrointestinal tract: can they survive there ?
It has been discussed whether administered spores can germinate in the GI tract. Working with mice, Casula & Cutting (2002) have used modified B. subtilis, with a chimeric gene ftsH-lacZ, which is expressed only in vegetative cells, which can be detected by RT-PCR up to only 100 bacteria. In this way they have seen that the spores germinate in significant numbers in the jejunum and in the ileum. That is, spores could colonize the small intestine, albeit temporarily.
Similarly, Duc et al. (2004) have concluded that B. subtilis spores can germinate in the gut because after the oral treatment of mice, in the faeces are excreted more spores that the swallowed ones, a sign that they have been able to proliferate. They have also detected, through RT-PCR, mRNA of vegetative bacilli after spore administration, and in addition, it has been observed that the mouse generates an IgG response against bacterial vegetative cells. That is, spores would not be only temporary stagers, but they would germinate into vegetative cells, which would have an active interaction with the host cells or the microbiota, increasing the probiotic effect.
With all this, perhaps it would be necessary to consider many Bacillus as not allochthonous of the GI tract, but as bacteria with a bimodal growth and sporulation life cycle, both in the environment and in the GI tract of many animals (Hong et al. 2005).
Regarding the normal presence of Bacillus in the intestine, when the different microorganisms inhabiting the human GI tract are studied for metagenomic DNA analysis of the microbiota, the genus Bacillus does not appear (Xiao et al., 2015). As we can see (Figure 6), the most common are Bacteroides and Clostridium, followed by various enterobacteria and others, including bifidobacteria.
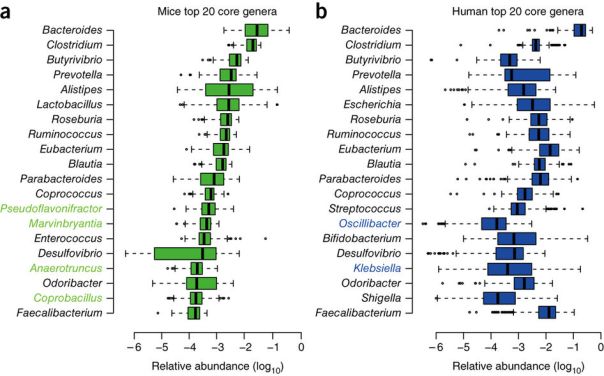
Figure 6. The 20 bacterial genera more abundant in the mice (left) and human (right) GI tract (Xiao et al. 2015).
In spite of this, several species of Bacillus have been isolated from the GI tract of chickens, treating faecal samples with heat and ethanol to select only the spores, followed by aerobic incubation (Barbosa et al. 2005). More specifically, the presence of B. subtilis in the human microbiota has been confirmed by selective isolation from biopsies of ileum and also from faecal samples (Hong et al. 2009). These strains of B. subtilis exhibited great diversity and had the ability to form biofilms, to sporulate in anaerobiosis and to secrete antimicrobials, thereby confirming the adaptation of these bacteria to the intestine. In this way, these bacteria can be considered intestinal commensals, and not only soil bacteria.
Security of Bacillus as probiotics
The oral consumption of important amounts of viable microorganisms that are not very usual in the GI treatment raises additional doubts about safety. Even more in the use of species that do not have a history of safe use in foods, as is the case of sporulated bacteria. Even normal bowel residents may sometimes act as opportunistic pathogens (Sanders et al. 2003).
With the exception of B. anthracis and B. cereus, the various species of Bacillus are generally not considered pathogenic. Of course, Bacillus spores are commonly consumed inadvertently with foods and in some fermented ones. Although Bacillus are recognized as GRAS for the production of enzymes, so far the FDA has not guaranteed the status of GRAS for any sporulated bacteria with application as a probiotic, neither Bacillus nor Clostridium. While Lactobacillus and Bifidobacterium have been the subject of numerous and rigorous tests of chronic and acute non-toxicity, and a lot of experts have reviewed data and have concluded that they are safe as probiotics, there is no toxicity data published on Bacillus in relation to their use as probiotics. When reviewing articles on Medline with the term “probiotic” and limited to clinical studies, 123 references appear, but Bacillus does not appear in any of them (Sanders et al. 2003).
Instead, there are some clinical studies where Bacillus strains have been detected as toxigenic. All this explains that some probiotic Bacillus producers refer to them with the misleading name of Lactobacillus sporogenes, a non-existent species, as can be seen from NCBI (https://www.ncbi.nlm.nih.gov/taxonomy/?term = Lactobacillus + sporogenes).
Finally, we should remember the joint report on probiotics of FAO (United Nations Food and Agriculture Organization) and WHO (World Health Organization) (FAO / WHO 2006), which suggests a set of Guidelines for a product to be used as a probiotic, alone or in the form of a new food supplement. These recommendations are:
- The microorganism should be well characterized at the species level, using phenotypic and genotypic methods (e.g. 16S rRNA).
- The strain in question should be deposited in an internationally recognized culture collection.
- To evaluate the strain in vitro to determine the absence of virulence factors: it should not be cytotoxic neither invades epithelial cells, and not produce enterotoxins or haemolysins or lecithinases.
- Determination of its antimicrobial activity, and the resistance profile, including the absence of resistance genes and the inability to transfer resistance factors.
- Preclinical evaluation of its safety in animal models.
- Confirmation in animals demonstrating its effectiveness.
- Human evaluation (Phase I) of its safety.
- Human evaluation (Phase II) of its effectiveness (if it does the expected effect) and efficiency (with minimal resources and minimum time).
- Correct labelling of the product, including genus and species, precise dosage and conservation conditions.

Conclusions
The use of Bacillus as probiotics, especially in the form of dietary supplements, is increasing very rapidly. More and more scientific studies show their benefits, such as immune stimulation, antimicrobial activities and exclusive competition. Their main advantage is that they can be produced easily and that the final product, the spores, is very stable, which can easily be incorporated into daily food. In addition, there are studies that suggest that these bacteria may multiply in GI treatment and may be considered as temporary stagers (Cutting 2011).
On the other hand, it is necessary to ask for greater rigor in the selection and control of the Bacillus used, since some, if not well identified, could be cause of intestinal disorders. In any case, since the number of products sold as probiotics that contain the sporulated Bacillus is increasing a lot, one must not assume that all are safe and they must be evaluated on a case-by-case basis (Hong et al. 2005).
Bibliography
Barbosa TM, Serra CR, La Ragione RM, Woodward MJ, Henriques AO (2005) Screening for Bacillus isolates in the broiler gastrointestinal tract. Appl Environ Microbiol 71, 968-978.
Casula G, Cutting SM (2002) Bacillus probiotics: Spore germination in the gastrointestinal tract. Appl Environ Microbiol 68, 2344-2352.
Chukeatirote E (2015) Thua nao: Thai fermented soybean. J Ethnic Foods 2, 115-118.
Cutting SM (2011) Bacillus probiotics. Food Microbiol 28, 214-220.
Duc LH, Hong HA, Barbosa TM, Henriques AO, Cutting SM (2004) Characterization of Bacillus probiotics available for human use. Appl Environ Microbiol 70, 2161-2171.
FAO/WHO (2006) Probiotics in food. Health and nutritional properties and guidelines for evaluation. Fao Food and Nutrition Paper 85. Reports of Joint FAO/WHO expert consultations.
Fontana L, Bermudez-Brito M, Plaza-Diaz J, Muñoz-Quezada S, Gil A (2013) Sources, isolation, characterization and evaluation of probiotics. Brit J Nutrition 109, S35-S50.
Granum, P. E., T. Lund (1997) Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Lett. 157:223–228.
Green, D. H., P. R. Wakeley, A. Page, A. Barnes, L. Baccigalupi, E. Ricca, S. M. Cutting (1999) Characterization of two Bacillus probiotics. Appl Environ Microbiol 65, 4288–4291.
Hoa, N. T., L. Baccigalupi, A. Huxham, A. Smertenko, P. H. Van, S. Ammendola, E. Ricca, A. S. Cutting (2000) Characterization of Bacillus species used for oral bacteriotherapy and bacterioprophylaxis of gastrointestinal disorders. Appl Environ Microbiol 66, 5241–5247.
Hong HA, Dic LH, Cutting SM (2005) The use of bacterial spore formers as probiotics. FEMS Microbiol Rev 29, 813-835.
Hong HA, Khaneja R, Tam NMK, Cazzato A, Tan S, Urdaci M, Brisson A, Gasbarrini A, Barnes I, Cutting SM (2009) Bacillus subtilis isolated from the human gastrointestinal tract. Res Microbiol 160, 134-143.
Lee S, Lee J, Jin YI, Jeong JC, Hyuk YH, Lee Y, Jeong Y, Kim M (2017) Probiotic characteristics of Bacillus strains isolated from Korean traditional soy sauce. LWT – Food Sci Technol 79, 518-524.
Mazza P (1994) The use of Bacillus subtilis as an antidiarrhoeal microorganism. Boll Chim. Farm. 133, 3-18.
Nimrat S, Suksawat S, Boonthai T, Vuthiphandchai V (2012) Potential Bacillus probiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeus vannamei). Veterinary Microbiol 159, 443-450.
Phromraksa P, Nagano H, Kanamaru Y, Izumi H, Yamada C, Khamboonruang C (2009) Characterization of Bacillus subtilis isolated from Asian fermented foods. Food Sci Technol Res 15, 659-666.
Sanders ME, Morelli L, Tompkins TA (2003) Sporeformers as human probiotics: Bacillus, Sporolactobacillus, and Brevibacillus. Compr Rev Food Sci Food Safety 2, 101-110
Vaseeharan, B., P. Ramasamy (2003) Control of pathogenic Vibrio spp. by Bacillus subtilis BT23, a possible probiotic treatment for black tiger shrimp Penaeus monodon. Lett Appl Microbiol 36, 83–87
World Gastroenterology Organisation Global Guidelines (2011) Probiotics and Prebiotics.
Xiao et al. (2015) A catalogue of the mouse gut metagenome. Nature Biotechnol 33, 1103-1108.


A new probiotic modulates gut microbiota against hepatocellular carcinoma
24th August 2016
Over the last years the beneficial effects of the human intestinal microbiota on various health markers have been displayed, such as inflammation, immune response, metabolic function and weight. The importance of these symbiotic bacteria of ours has been proved. You can see these other posts related with our microbiota: “The good clostridia avoid us from allergies“, “Gut bacteria controlling what we eat” or “Good bacteria of breast milk”
At the same time it has been seen that probiotics can be a good solution for many diseases with affected gut microbiota. Indeed, the beneficial role of probiotics to reduce gastrointestinal inflammation and prevent colorectal cancer has been proven.
However, recently it has been found that probiotics may have beneficial effects in other parts of the body beyond the gastrointestinal tract, particularly with immunomodulatory effects on an hepatocellular carcinoma (HCC). In this way, researchers at the University of Hong Kong, along with other from University of Eastern Finland, have published a study (Li et al, PNAS, 2016), where they have seen reductions of 40% in weight and size of HCC liver tumours in mice which were administered with a new mixture of probiotics, “Prohep.”
Hepatocellular carcinoma (HCC) is the most common type of liver cancer is the 2nd most deadly cancers, and it is quite abundant in areas with high rates of hepatitis. In addition, sorafenib, the drug most widely used to reduce the proliferation of tumour, is very expensive. The cost of this multikinase inhibitor is €3400 for 112 tablets of 200 mg, the recommended treatment of four pills a day for a month. Instead, any treatment with probiotics that would proved to be effective and could replace this drug would be much cheaper.
The new probiotics mix Prohep consists of several bacteria: Lactobacillus rhamnosus GG (LGG), Escherichia coli Nissle 1917 (ECN) and the whole inactivated by heat VSL#3 (1: 1: 1) containing Streptococcus thermophilus, Bifidobacterium breve, Bf. longum, Bf. infantis, Lb. acidophilus, Lb. plantarum, Lb. paracasei and Lb. delbrueckii.
In the mentioned work, Li et al. (2016) fed mice with Prohep for a week before inoculating them with a liver tumour, and observed a 40% reduction in tumour weight and size in comparison to control animals. As shown in Figure 1, the effect was significant at 35 days, and also for those who were given the Prohep the same day of tumour inoculation. Obviously, the effect of tumour reduction was much more evident when the antitumour compound Cisplatin was administered.
These researchers saw that tumour reduction was due to the inhibition of angiogenesis. This is the process that generates new blood vessels from existing ones, something essential for tumour growth. In relation to the tumour reduction, high levels of GLUT-1 + hypoxic were found. That meant that there was hypoxia caused by the lower blood flow to the tumour, since this was 54% lower in comparison to controls.
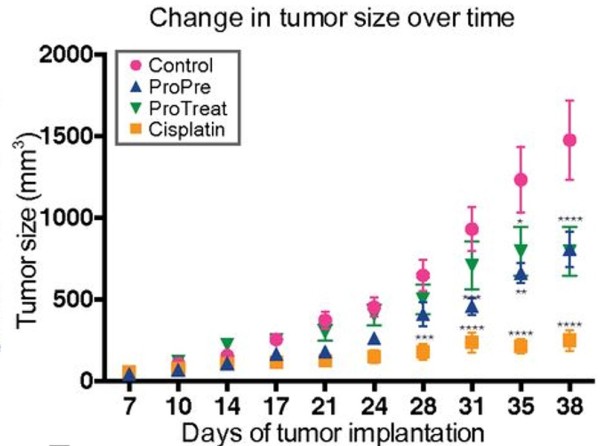
Figure 1. Change in tumour size. ProPre: administration of Prohep one week before tumour inoculation; ProTreat: administration of Prohep the same day of tumour inoculation; Cisplatin: administration of this antitumoral. (Fig 1B from Li et al, 2016).
These authors also determined that there was a smaller amount of pro-inflammatory angiogenic factor IL-17 and of Th17 cells of the immune system, cells also associated with cancer. The lower inflammation and angiogenesis could limit the tumour growth.
Moreover, these researchers established that the beneficial effects of probiotics administration were associated with the abundance of beneficial bacteria in the mice gut microbiota, analysed by metagenomics. So, probiotics modulate microbiota, favouring some gut bacteria, which produce anti-inflammatory metabolites such as cytokine IL-10 and which suppress the Th17 cell differentiation.
Figure 2. Bacteria of the human intestinal microbiota seen by scanning electron microscope (SEM) (coloured image of Eye of Science / Science Source)
Some of the bacteria identified by metagenomics in the microbiota of mice that were administered with Prohep were Prevotella and Oscillibacter. The first is a bacteroidal, gram-negative bacterium, which is abundant in the microbiota of rural African child with diets rich in carbohydrates. Oscillibacter is a gram-positive clostridial, known in humans as a producer of the neurotransmitter GABA. Both are an example of the importance of some clostridial and bacteroidals in the gut microbiota. In fact, they are majority there, and although they are not used as probiotics, are found increasingly more positive functions, such as avoiding allergies (see “The good clostridia avoid us from allergies“).
It is known that these bacteria produce anti-inflammatory metabolites and therefore they would be the main involved in regulating the activity of immune cells that cause tumour growth. The observed reduction of tumour in these experiments with mice would be the result of combined effect of these administered probiotic bacteria together with the microbiota itself favoured by them. We see a potential outline of these actions in Figure 3.
Figure 3. Simplified diagram of the possible mechanisms of gut bacteria influencing on the polarization of Th17 cells in the lamina propria of the intestinal mucosa. The microbiota bacteria activate dendritic cells, which secrete cytokines (IL-22, IL-23, IL-27). The bacteria can promote Th17 immunity inducing IL-23, which can be involved by means of TLR ligands signal or extracellular ATP or serum amyloid A (SAA). Meanwhile, some probiotic strains could inhibit the development of Th17 by means of the production of IL-12 and IL-27, in addition to promoting the growth and colonization of the bacteria that induce Th17 (Sung et al 2012, Fig. 2).
Although we know that the cancer progression is a very complex process and that in the tumour microenvironments there is an infiltration of many different types of immune system cells, such as T cells, neutrophils, killer cells, macrophages etc, the Th17 helper cell subpopulation appears to be prevailing in the tumour progression, and therefore these effects of probiotics and microbiota open good prospects.
It is still early to say whether these findings will contribute to the treatment of human liver cancer, and therefore research in humans is needed, in order to see if these probiotics could be used as such or in tandem with some drug, depending on the tumour stage and size. In any case, all this opens a new range of possibilities for research of the molecular mechanisms of the beneficial effects of probiotics beyond the intestinal tract.
Bibliography
El-Nezami H (2016 april 27) HKU develops novel probiotic mixture “Prohep” that may offer potential therapeutic effects on liver cancer. The University of Hong Kong (HKU) 27 Apr 2016
El-Nezamy H, Lee PY, Huang J, Sung YJ (2015) Method and compositions for treating cancer using probiotics. Patent WO 2015021936 A1
Li J, Sung CYJ, Lee N, Ni Y, Pihlajamäki J, Panagiotou G, El-Nezami H (2016) Probiotics modulated gut microbiota suppresses hepatocellular carcinoma growth in mice. PNAS E1306-E1315
Oelschlaeger TA (2010) Mechanisms of probiotic actions – A review. Int J Med Microbiol 300, 57-62
Packham C (2016) Probiotics dramatically modulate liver cancer growth in mice. Medical Press, Med Research 23 Feb 2016
Silgailis M (2016) Treating some cancers with probiotics in the future ? Probiotic Prohep. Lacto Bacto: Health, Microbes and More 23 Feb 2016
Sung CYJ, Lee NP, El-Nezami H (2012) Regulation of T helper by bacteria: an approach for the treatment of hepatocellular carcinoma. Int J Hepatology ID439024, doi:10.1155/2012/439024
UEF News and Events (2016) A novel probiotic mixture may offer potential therapeutic effects on hepatocellular carcinoma. University of Eastern Finland 1 Mar 2016

The effect of rapid climate change in the Arctic ecosystem
March 20th, 2016
The Arctic Ocean
Interestingly and coincidentally, “Arctic” comes from the Greek word αρκτος -arctos-, which means “bear” and is a reference to the constellation Ursa Minor, where is the North Star, which indicates the geographic North Pole .
The Arctic constitutes a unique ecosystem of the Earth, consisting of a large ice field, or ice-covered ocean, sometimes regarded as the northern part of the Atlantic Ocean, and it is surrounded by land, which is permafrost, with complete absence of trees. Life in the Arctic consists of organisms adapted to ice, including zooplankton and phytoplankton, fish, marine mammals, birds, land animals, plants and human societies fully adapted to the extreme conditions of the environment.
Due to global warming, isotherms are moving northward at a rate exceeding 50 km per decade over the past 30 years, so if we define the Arctic from a defined temperature or the tree line, its size is diminishing, being the reduction of sea ice the most visible effect.
Anthropogenic climate change: global warming, especially in the Arctic
Yes: climate change is here and it is generated by human activities, that is, it is anthropogenic. Previously there have been on Earth fluctuations in global temperature caused by natural phenomena, usually long-term and cyclical variations. For example, glaciations since about 2 million years are repeated every 100,000 years, and last ice age ended 15,000 years ago. So we are living now in an interglacial period and the next ice age could become not before 50,000 years. The cause of this cycle of glaciations seems to be orbital variations of the Earth, resulting in a lower insolation in high latitudes of the northern hemisphere during glacial periods.
Solar activity, like other stars, has cycles and roughly every 600 years there are periods of little activity (absence of very few solar spots and auroras), with lower energy output, which corresponds to cold periods in the Earth’s climate. The last minimal was in the period 1645-1715, and therefore from the middle of the eighteenth century we enjoy a maximum solar activity, with small cycles of minimum and maximum every 11 years.
Discounting these natural variations, it is clear that throughout the 20th century and especially since the 1960s there has been a steady increase in global average temperature (Figure 1), reaching almost 1ºC more than the beginning of the 20th century. In the early years of the current century the trend is worsening. The last 10 years have been the warmest since there are records, and the forecast is to continue increasing. Most experts agree that humans exert a direct impact on the heating process known as the greenhouse effect. The causes of this effect are some of gaseous components of the atmosphere, especially CO2, which has grown in parallel with rising temperatures, from about 300 ppm at the beginning of 20th century to nearly 400 ppm today. This CO2 and other gases as water vapour, methane and other exclusively anthropogenic absorb radiation and the result is that the atmosphere warms further.
Figure 1. Increase in average global temperature compared to the beginning of 20th century (from GISTEMP).
This global warming is particularly evident in the Arctic. The temperature increases are higher in northern latitudes, especially 60-70º N, where this past December 2015 (Figure 2) have raised to 9ºC above average in large areas of North America and Eurasia. This is called Polar Warming Amplification (PWA). The cause of this overheating in the Arctic respect of the rest of Earth is partly due to the loss of snow and ice (retroactive effect) because the largest area of land and water absorbs more solar energy than white ice (albedo effect), but also the PWA is partly due to the dynamic atmospheric transport, which transports heat energy from the clouds and subtropical regions to the north (Taylor et al 2013).
Figure 2. Thermal anomaly registered in December 2015 with respect to the average 1951-1980 (from GISTEMP).
Besides the consequences of this warming on the Arctic ice that we will comment below, another serious problem is the melting of permafrost, since then methane gas trapped under the frozen ground is released. This way, vast quantities of methane are released, and this greenhouse gas is contributing further to accelerate the global warming.
Less and less ice in the Arctic
Linear trends of sea ice extent and sea ice in the Arctic from 1979 to date are negative year after year, for any month is considered, but it is more clear by comparing Septembers, at the end of the summer when the ice is melting (Figure 3). Of the approximately 7 million km2 minimum in September (the maximum in March is about 16 million), about 100,000 km2 are melt per year, almost 9% every 10 years (Serreze et al 2007), so that there is now almost half ice than in 1979 (Figure 4).
Figure 3. Comparison of the extent of sea ice (in red): September 1979 and 2012 (from The Cryosphere Today).
Figure 4. Average monthly extension of Arctic sea ice since 1979 (Reeves et al 2013).
In addition to the reduction in surface ice, keep in mind the reduction in volume, representing now a third of what it was in September 1979.
There is a big difference between the different models for predicting the disappearance of Arctic sea ice. Half of them expect the total disappearance by September 2100. Predictions move since September 2040 the less optimistic until well past 2100 for the other (Serreze et al 2007).
Other problems resulting from the disappearance of sea ice are the ship traffic, which could shorten distances trips between the ports of northern countries, and on the other hand the exploitation of oilfields and other fossil fuels and minerals, since there is a large part of global reserves in the Arctic (Figure 5).
Figure 5. Left: forecast paths for open sea ships (blue) and for icebreakers (red) for 2040-2059. Right: Distribution of the potential major reserves of oil and gas (yellow) and licenses (red) and wells in operation or to operate (black). The dashed line indicates the limit of Conservation of Arctic Flora and Fauna (CAFF) declared by the Working Group of the Arctic Council (www.arctic-council.org). Figures from Reeves et al (2013).
Ecological consequences of the disappearance of the Arctic ice pack
There are many living beings linked to the ice. The polar bears roam on the Arctic ice, so we are feared for his fate. Many fish, seals and crustaceans (krill) form a food chain that starts from the algae that grow under the ice in a very consistent environment, rich in nutrients, especially favourable for marine life (Figure 6 A). Moreover, floating sea ice in summer is a good corridor for dispersion of terrestrial vertebrates (for instance arctic foxes) and plants.
The gradual disappearance of sea ice and warming in the Arctic coast involves a series of ecological imbalances (Figure 6 B). We see for example how walruses forced to remain grouped on the ground are more predisposed to disease transmission. The loss of sea ice diminishes dispersion by ice corridors and then the land populations are most isolated, thus gene flow is restricted. Polar bears and other predators that hunt on the sea ice have it much harder and their populations are at risk. Phytoplankton productivity decreases significantly, thereby reducing zooplankton, and then the whole food chain (fish, seals, etc.) is affected (Post et al 2013).
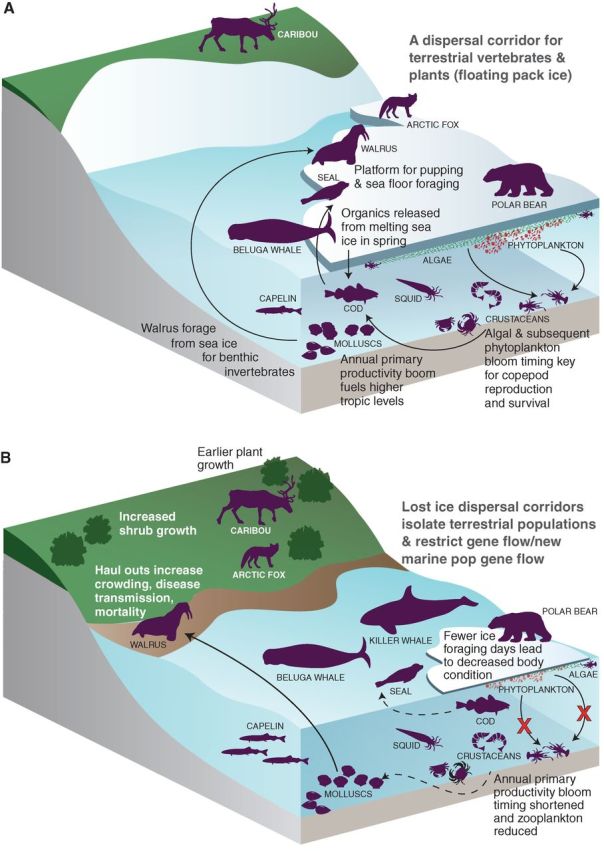
Figure 6. Ecological interactions influenced by sea ice. A: The distribution and seasonality of sea ice affects the abundance, distribution and interactions of the entire ecosystem in balance. B: The longest period without ice and less sea ice extent have disastrous consequences on the balance of the ecosystem (Post et al 2013).
The polar bear tries to survive
The polar bear (Ursus maritimus) is considered an endangered animal. There are only about 25,000 worldwide. The impact of climate change affects the exclusive habitat of polar regions and forecasts suggest that in a few years from now the ice of the Arctic will melt permanently and polar bears may become extinct because of warming area.
The polar bear is basically carnivorous, unlike others such as brown bears, and remains above the ice hunting seals. With the gradual disappearance of the ice it has more trouble finding preys, and some have begun to learn how to catch salmon rivers, as we see in the images (Figure 7).


Figure 7. White Bear dedicated to fishing salmons in order to survive (www.youtube.com/watch?v=9m_Q9Ojbcmw).
We have also seen groups of polar bears at sea fishing (see video) and dive emerging alternately as if they were dolphins or porpoises. Despite these small adaptations, the food is very low and it is clear that their populations are declining rapidly.
Orcas thrive north
The disappearance of the northern ice is a dramatic ecological change that is causing the disappearance of some species like the polar bear, but interestingly these imbalances benefit some other emerging species. This is the case of the killer whale (Orcinus orca), which is thriving more and more to the north (Figure 8).

Figure 8. Places (marked with numbers) of the Canadian Arctic where groups of orcas were repeatedly photographed between 2004 and 2009 (Young et al 2011).
Eskimo Inuit people live around the American Arctic (from Quebec to Alaska including Hudson Bay and adjacent islands) and the west coast of Greenland, and they are the first witnesses since the mid-twentieth century observing whales in their waters, unknown before. Moreover, in recent years scientists have made numerous orca’ sightings, they have been photographed individually (Young et al 2011), and their travels have been followed through bioacoustics (Ferguson et al 2010) and other techniques.


Figure 9. (Top): Narwhals with the characteristic great tusk, which gave rise to the myth of the unicorn. (Low): Group of orcas attacking narwhals cornered on the beach. Watch the video of PBS Nature.
For some years attacks by orcas on narwhals (as in Figure 9) have been observed repeatedly by Inuit Eskimos and studied in detail by several scientists. Laidre et al (2006) observed that before approaching whales, the narwhals tend to group, are more quiet and swim closer to the beach in shallow waters. During the attack, the narwhals disperse significantly but nevertheless mortality is very high. After predation, which can last several hours, oily stains are observed in sea surface, which come from fat of depredated narwhals (Figure 10).

Figure 10. Group of orcas surrounded by patches of oil on the sea surface from the fat of attacked narwhals (Laidre et al 2006).
Orcas’ attacks on narwhals are so common and effective that are beginning to affect the population. The effects are even worse in other cetaceans with smaller population such as whales of Greenland or bowhead (Balena mysticetus), which are now virtually extinct (Figure 11).
Figure 11. Scheme of preys’ proportions by a group of orcas from Hudson Bay (Ferguson et al 2010).
In conclusion, anthropogenic climate change is affecting the Arctic ecosystem severely (and all the other ecosystems), and although this problem is becoming known, effective policy measures to reduce emissions of CO2 and other greenhouse gases are so scarce that hardly will arrive in time. We are leading the planet Earth to a massive extinction of species and ecological changes ever seen in the history of humans.

The picture says it all: polar bear habitat is running out.
Bibliography
Arctic Council: http://www.arctic-council.org
Ferguson S.H., Higdon J.W. & Chmelnitsky E.G. (2010) The rise of killer whales as a major Arctic predator. In S.H. Ferguson, et al. (eds.): A little less Arctic: top predators in the world’s largest northern inland sea, Hudson Bay. Pp. 117–136. New York: Springer
GISTEMP, Goddard Institute for Space Studies Surface Temperature Analysis (NASA-GISS): http://data.giss.nasa.gov/gistemp/
Hawkings E (2014) nov 28: http://www.climate-lab-book.ac.uk/2014/hiatuses-in-the-rise-of-temperature/
Laidre KL, Heide-Jørgensen MP, Orr J (2006) Reactions of narwhals, Monodon monoceros, to killer whale, Orcinus orca, attacks in the Eastern Canadian Arctic. Can. Field Nat., 120, 457–465
Morell V (2012) Killer whale menu finally revealed. http://www.sciencemag.org/news/2012/01/killer-whale-menu-finally-revealed
PBS Nature: http://www.pbs.org/wnet/nature/invasion-killer-whales-killer-whales-attack-pod-narwhals/11165/
Post et al. (2013) Ecological Consequences of Sea-Ice Decline. Science. DOI: 10.1126/science.1235225: http://www.carbonbrief.org/knock-on-effects-for-wildlife-as-the-arctic-loses-ice
Reeves RR et al (2014) Distribution of endemic cetaceans in relation to hydrocarbon development and commercial shipping in a warming arctic. Marine Policy 44, 375-389
Serreze MC, Holland MM, Stroeve J (2007) Perspectives on the Arctic’s shrinking sea-ice cover. Science 315, 5818, 1533–6.
Taylor PC, Cai M, Hu A, Meehl J, Washington W, Zhang GJ (2013) Decomposition of feedback contributions to Polar Warming Amplification. J Climate 26, 7023-43
The Cryosphere Today: http://arctic.atmos.uiuc.edu/cryosphere/
Wikipedia: https://en.wikipedia.org/wiki/Climate_change
Wikipedia: https://en.wikipedia.org/wiki/Arctic
Young BG, Jeff W. Higdon JW, Steven H. Ferguson SH (2011) Killer whale (Orcinus orca) photo-identification in the eastern Canadian Arctic. Polar Research Vol 30

Human skin microbiota partly shared with our dog
December 25th, 2015
Diversity of the human microbiota in different parts of the body and between individuals
As I have commented in previous posts of this blog (Good Clostridia in our gut March 21st, 2015; Bacteria controlling what we eat October 12th, 2014; Bacteria of breast milk February 3rd, 2013), it becomes increasingly clear the importance of our microbiota, id est, all the micro-organisms, especially bacteria, with which we live.
The human microbiota varies from one individual to another, in relation to diet, age and the own genetic and phenotypic characteristics. Moreover, since we do not live isolated, there is also the influence of the environment, and of other people with we live, including our pets, dogs and others. They all have also their own microbiota.
The human body is home to many different microorganisms: bacteria (and archaea), fungi and viruses, that live on the skin, in the gut and in several other places in the body (Figure 1). While many of these microbes are beneficial to their human host, we know little about most of them. Early research focused on the comparison of the microorganisms found in healthy individuals with those found in people suffering from a particular disease. More recently, researchers have been interested in the more general issues, such as understanding how the microbiota is established and knowing the causes of the similarities and differences between the microbiota of different individuals.
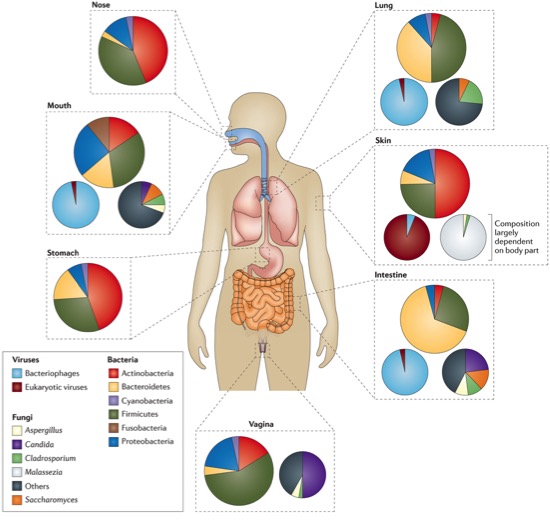
Figure 1. Types of microorganisms that live in different parts of the human body: bacteria (large circles), fungi (small circles right) and viruses (small circles left) (Marsland & Gollwitzer 2014)
Now we know that communities of microorganisms that are found in the gut of genetically related people tend to be more similar than those of people who are not related. Moreover, microbial communities found in the gut of unrelated adults living in the same household are more similar than those of unrelated adults living in different households (Yatsunenko et al 2012). However, these studies have focused on the intestine, and little is known about the effect of the relationship, cohabitation and age in microbiota of other parts of the body, such as skin.
Human skin microbiota
The skin is an ecosystem of about 1.8 m2 of various habitats, with folds, invaginations and specialized niches that hold many types of microorganisms. The main function of the skin is to act as a physical barrier, protecting the body from potential attacks by foreign organisms or toxic substances. Being also the interface with the external environment, skin is colonized by microorganisms, including bacteria, fungi, viruses and mites (Figure 2). On its surface there are proteobacteria, propionibacteria, staphylococci and some fungi such as Malassezia (an unicellular basidiomycetous). Mites such as Demodex folliculorum live around the hair follicles. Many of these microorganisms are harmless and often they provide vital functions that the human genome has not acquired by evolution. The symbiotic microorganisms protect human from other pathogenic or harmful microbes. (Grice & Segre 2011).
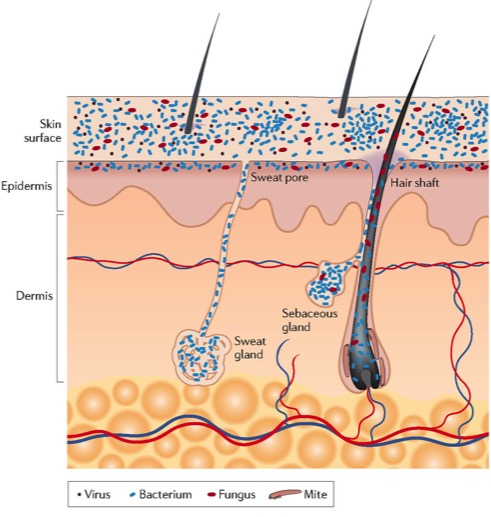
Figure 2. Schematic cross section of human skin with the different microorganisms (Grice & Segre 2011).
According to the commented diversity of microbiota, this is also very different depending on the region of skin (Figure 3), and therefore depending on the different microenvironments, that can be of three different characteristics: sebaceous or oily, wet and dry.
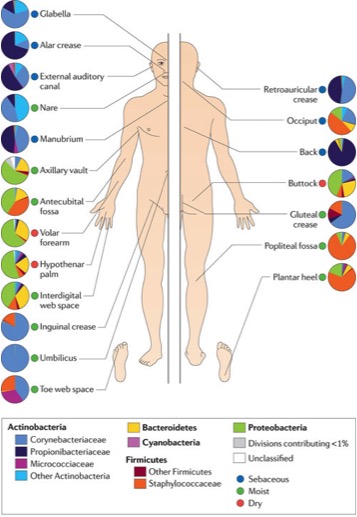
Figure 3. Topographic distribution of bacterial types in different parts of the skin (Grice & Segre 2011)
The skin is a complex network (structural, hormonal, nervous, immune and microbial) and in this sense it has been proven that the resident microbiota collaborates with the immune system, especially in the repair of wounds (Figure 4). As we see, particularly the lipopotheicoic acid (LTA), compound of the bacterial cell wall, can be released by Staphylococcus epidermidis and stimulates Toll-like receptors TLR2, which induce the production of antimicrobial peptides, and also stimulate epidermal keratinocytes via TLR3, which trigger the inflammation with production of interleukin and attracting leukocytes (Heath & Carbone 2013). All this to ensure the homeostatic protection and the defence against the potential pathogens. More information in the review of Belkaid & Segre (2014).
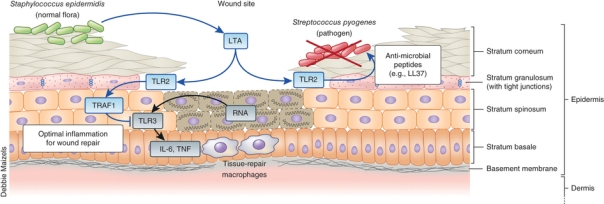
Figure 4. Contribution of the resident microbiota to the immunity and wound repair (Heath & Carbone 2013)
At home we share microbiota, and with the dog
As mentioned earlier, environment influences the microbiota of an individual, and therefore, individuals who live together tend to share some of the microbiota. Indeed, it was recently studied by Song et al (2013), with 159 people and 36 dogs from 60 families (couples with children and / or dogs). They study the microbiota of gut, tongue and skin. DNA was extracted from a total of 1076 samples, amplifying the V2 region of the 16S rRNA gene with specific primers, and then it was proceeded to multiplex sequencing of high performance (High-Throughput Sequencing) with an Illumina GA IIx equipment. Some 58 million sequences were obtained, with an average of 54,000 per sample, and they were analysed comparing with databases to find out what kind of bacteria and in what proportions.
The results were that the microbial communities were more similar to each other in individuals who live together, especially for the skin, rather than the bowel or the tongue. This was true for all comparisons, including pairs of human and dog-human pairs. As shown in Figure 5, the number of bacterial types shared between different parts was greater (front, palms and finger pulps dog) of the skin of humans and their own dog (blue bars) than the human with dogs of other families (red bars), or dogs with people without dogs (green bars). We also see that the number of shared bacterial types is much lower when compared faecal samples or the tongue (Song et al 2013).
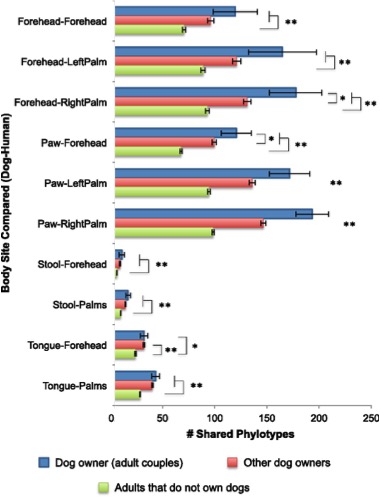
Figure 5. Numbers of bacterial phylotypes (phylogenetic types) shared between adults and their dogs (blue), adults with other dogs (red) and adults who do not have dogs with dogs. There are compared (dog-human) fronts, hands, legs pulps, and also faecal samples (stool) and tongues. Significance of being different: *p<0.05, **p<0.001 (Song et al 2013)
This suggests that humans probably take a lot of microorganisms on the skin by direct contact with the environment and that humans tend to share more microbes with individuals who are in frequent contact, including their pets. Song et al. (2013) also found that, unlike what happens in the gut, microbial communities in the skin and tongue of infants and children were relatively similar to those of adults. Overall, these findings suggest that microbial communities found in the intestine change with age in a way that differs significantly from those found in the skin and tongue.
Although is not the main reason for this post, briefly I can say that the overall intestinal microbiota of dogs is not very different from humans in numbers (1011 per gram) and diversity, although with a higher proportion of Gram-positive (approx. 60% clostridial, 12% lactobacilli, 3% bifidobacteria and 3% corynebacteria) in dogs, and less Gram-negative (2% Bacteroides, 2% proteobacteria) (García-Mazcorro Minamoto & 2013).
Less asthma in children living with dogs
Although the relationship with the microbiota has not fully been demonstrated, some evidence of the benefits of having a dog has been shown recently, and for the physical aspects, not just for the psychological ones. Swedish researchers (Fall et al 2015) have carried out a study of all new-borns (1 million) in Sweden since 2001 until 2010, counting those suffering asthma at age 6. As the Swedes also have registered all dogs since 2001, these researchers were able to link the presence of dogs at home during the first year of the baby with the onset of asthma or no in children, and have come to the conclusion that children have a lower risk of asthma (50% less) if they have grown in the presence of a dog.
Similar results were obtained for children raised on farms or in rural environments, and thus having contact with other animals. All this would agree with the “hygiene hypothesis”, according to which the lower incidence of infections in Western countries, especially in urban people, would be the cause for increased allergic and autoimmune diseases (Okada et al 2010). In line with the hypothesis, it is believed that the human immune system benefits from living with dogs or other animals due to the sharing of the microbiota. However, in these Swede children living with dogs and having less risk of asthma there was detected a slight risk of pneumococcal disease. This links to the aforementioned hypothesis: more infections and fewer allergies (Steward 2015), but with the advantage that infections are easily treated or prevented with vaccines.

References
Belkaid Y, Segre JA (2014) Dialogue between skin microbiota and immunity. Science 346, 954-959
Fall T, Lundholm C, Örtqvist AK, Fall K, Fang F, Hedhammar Å, et al (2015) Early Exposure to Dogs and Farm Animals and the Risk of Childhood Asthma. JAMA Pediatrics 69(11), e153219
García-Mazcorro JF, Minamoto Y (2013) Gastrointestinal microorganisms in cats and dogs: a brief review. Arch Med Vet 45, 111-124
Heath WR, Carbone FR (2013) The skin-resident and migratory immune system in steady state and memory: innate lymphocytes, dendritic cells and T cells. Nature Immunology 14, 978-985
Marsland BJ, Gollwitzer ES (2014) Host–microorganism interactions in lung diseases. Nature Reviews Immunology 14, 827-835
Okada H, Kuhn C, Feillet H, Bach JF (2010) The “hygiene hypothesis” for autoimmune and allergic diseases: an update. Clin Exp Immunol 160, 1-9
Song SJ, Lauber C, Costello EK, Lozupone, Humphrey G, Berg-Lyons D, et al (2013) Cohabiting family members share microbiota with one another and with their dogs. eLife 2, e00458, 1-22
Steward D (2015) Dogs, microbiomes, and asthma risk: do babies need a pet ? MD Magazine, Nov 03
Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, et al. 2012. Human gut microbiome viewed across age and geography. Nature 486, 222–7

The giant panda is herbivore but has the gut microbiota of a carnivore
September 30th, 2015
The giant panda (Ailuropoda melanoleuca, literally Greek for “white and black cat feet”) is one of the most intriguing evolutionary mammal species. Despite its exclusively herbivorous diet, phylogenetically it is like a bear because it belongs to Ursids family, order Carnivores. Its diet is 99% bamboo and the other 1% is honey, eggs, fish, oranges, bananas, yams and leaves of shrubs.
It lives in a mountain area in central China, mainly in Sichuan province, and also in provinces of Shaanxi and Gansu. Due to the construction of farms, deforestation and other development, the panda has been driven out of the lowland where he lived. It is an endangered species that needs protection. There are about 300 individuals in captivity and 3000 in freedom. Although the numbers are increasing, it is still endangered, particularly due to its limited space (20,000 km2) and its very specific habitat (bamboo forests).

Thus, the giant panda has an almost exclusive diet of different species of bamboo, mainly the very fibrous leaves and stems, and buds in spring and summer. It is therefore a poor quality -digestive diet, with little protein and plenty of fibre and lignin content. They spend about 14 hours a day eating and can ingest about 12 kg of bamboo a day.
Most herbivores have modifications of the digestive tract that help them to retain the food in digestion process and contain microbial populations that allow them to eat exclusively plant materials, rich in complex polysaccharides such as cellulose and hemicellulose. These specializations may be compartmentalization of the stomach of ruminants and other typical non-ruminants (kangaroos, hamster, hippopotamus and some primates) or enlargement of the large intestine, characteristic of equines, some rodents and lagomorphs (rabbits and hares).
However, despite his exclusively herbivorous diet, surprisingly the giant panda has a typical carnivorous gastrointestinal tract, anatomically similar to dog, cat or raccoon, with a simple stomach, a degenerated caecum and a very short colon. The gastrointestinal tract of pandas is about 4 times the size of the body, such as other carnivores, whereas herbivores have about 10-20 times the size of the body, to efficiently digest large amounts of forage. With this, the panda intestinal transit time is very short, less than 12 hours. This severely limits the ability of potential fermentation of plant materials (Williams et al. 2013).
For these reasons, the digestion of bamboo for panda is very inefficient, despite their dependency. Pandas consume the equivalent of 6% of their body weight per day, with a 20% digestibility of dry matter of bamboo. Of this, 10% corresponds to the low protein content of bamboo, and the rest are polysaccharides, particularly with coefficients of digestion of 27% for hemicellulose and 8% for the pulp.
It seems as if the giant panda would have specialized in the use of a plant with high fibre content without having modified the digestive system, by means of an efficient chewing, swallowing large quantities, digesting the contents of cells instead of plant cell walls, and quickly excreting undigested waste (Dierenfield et al. 1982).
In addition, having a dependency on one type of plant such as bamboo can lead to nutritional deficiencies depending on seasonal cycles of the plant. In this regard, recently Nie et al. (2015) have studied the concentrations of calcium, phosphorus and nitrogen from different parts of the bamboo that a population of free pandas eat. They have seen that pandas in their habitat have a seasonal migration in two areas of different altitudes throughout the year and that fed two different species of bamboo. Both species have more calcium in the leaves and more phosphorus and nitrogen in the stems. As the seasonal variation in appearance and fall of leaves of two species is different due to the different altitude, when pandas are in one of the areas eat the leaves of a species and stems of the other while they do the reverse when they are in the other zone. So, pandas synchronize their seasonal migrations in order to get nutritionally the most out of both species of bamboo.
Another drawback of the bamboo dependence is flowering. It is a natural phenomenon that happens every 40-100 years, and when bamboo flowers, it dies, reducing the availability of food for pandas. During 1970-1980 there were two large-scale blooms in the habitat of pandas, and there were more than 200 deaths for this reason. However, and given that probably pandas have found during their evolution with many other massive blooms, in these occasions they are looking for other species of bamboo or travel long distances to meet their food needs (Wei et al. 2015).
In return, and as adaptation to eat this so specific food, the giant panda has a number of unique morphological features, such as strong jaws and very powerful molars, and especially a pseudo-thumb, like a 6th finger, which is actually a modified enlarged sesamoid bone, as an opposable thumb, which serves to hold bamboo while eating (Figure 1).

Figure 1. The “pseudo-thumb” of giant panda. Image from Herron & Freeman (2014).
And how is that the panda became an herbivore ?
It has been estimated that the precursor of the giant panda, omnivorous as other Ursids, began to eat bamboo at least 7 million years ago (My), and became completely dependent on bamboo between 2 and 2.4 My. This dietary change was probably linked to mutations in the genome, leading to defects in the metabolism of dopamine in relation to the appetite for meat, and especially the pseudogenization of Tas1r1 gene (Figure 2) of umami taste receptor (Jin et al. 2011). The umami is one of the five basic tastes, along with sweet, salty, sour and bitter. Umami is like “pleasant savoury taste”, usually recalls meat, and is related to L-glutamic acid, abundant in meat. This mutation in pandas favoured the loss of appetite for meat and reinforced their herbivore lifestyle. However, other additional factors had probably been involved, since Tas1r1 gene is intact in herbivores such as horses and cows (Zhao et al. 2010).

Figure 2. Phylogenetic tree of some carnivores with data for giant panda deduced from fossils (in blue) and from the molecular study of TasTr1 gene made by Zhao et al. (2010).
The intestinal microbiota of giant panda
As expected, when sequencing the complete genome of the giant panda (Li et al. 2010), specific genes responsible for the digestion of cellulose and hemicellulose have not been found. Logically, these complex polysaccharides of bamboo fibres would be possibly digested by cellulolytic microorganisms of the intestinal tract. So, their presence in panda must be studied.
When studying the sequences of 16S ribosomal DNA from faecal microbiota of various mammals, an increase in bacterial diversity is generally observed in sense carnivores – omnivores – herbivores (Ley et al. 2008). This diversity is lower in the panda than in herbivores, and as shown in Figure 3, pandas are grouped with carnivores (red circles) despite being herbivorous from the diet point of view.

Figure 3. Principal component analysis (PC) of faecal bacterial communities from mammals with different colours according to the predominant diet (Law et al. 2008)
The intestinal microbiota of most herbivores contains anaerobic bacteria mainly from groups of Bacteroides, Clostridials, Spirochetes and Fibrobacterials, that have enzymatic ability to degrade fibrous plant material and thus provide nutrients for its guests. Instead, omnivores and carnivores have a particularly dominant microbiota of facultative anaerobes, such as Enterobacteriaceae, besides some Firmicutes, including lactobacilli and some Clostridials and Bacteroides.
As for the giant panda, the first studies made with culture-dependent methods and analysis of amplified 16S rRNA genes (Wii et al. 2007) identified Enterobacteriaceae and Streptococcus as predominant in the intestinal microbiota. Therefore, this study suggests that the microbiota of panda is very similar to that of carnivores, as we see in the mentioned comparative study with various mammals (Law et al. 2008), and therefore with little ability to use cellulose or hemicellulose.
However, a later study done with sequencing techniques of 16S (Zhu et al. 2011) from faecal samples of 15 giant pandas arrived at very different conclusions and it seemed that they found the first evidence of cellulose digestion by microbiota of giant panda. In 5500 sequences analysed, they found 85 different taxa, of which 83% were Firmicutes (Figure 4), and among these there were 13 taxa of Clostridium (7 of them exclusive of pandas) and some of these with ability to digest cellulose. In addition, in metagenomic analysis of some of the pandas some putative genes for enzymes to digest cellulose, xylans and beta-glucosidase-1,4-beta-xilosidase for these Clostridium were found. Altogether, they concluded that the microbiota of the giant panda had a moderate degradation capacity of cellulose materials.

Figure 4. Percentage of sequences of the main bacterial groups found in faecal samples from wild individuals of giant panda (W1-W7) and captive (C1-C8), according to Zhu et al. (2011). Under each individual the n. sequences analysed is indicated.
But just three months ago a work (Xue et al. 2015) has been published that seems to go back, concluding that the intestinal microbiota of the giant panda is very similar to that of carnivores and have little of herbivores. It is an exhaustive study of last-generation massive sequencing of 16S rRNA genes of faecal samples from 121 pandas of different ages over three seasons. They obtained some 93000 sequences corresponding to 781 different taxa.
They found a predominance of Enterobacteriaceae and Streptococcus (dark red and dark blue respectively, Figure 5A) and very few representatives of probable cellulolitics as Clostridials. Moreover, these are not increased when more leaves and stems of bamboo are available (stage T3). These results correspond with what was already known of the low number of genes of cellulases and hemicellulases (2%), even lower than in the human microbiome. This negligible contribution of microbial digestion of cellulose, together with the commented fact that the panda is quite inefficient digesting bamboo, contradicts the hypothetical importance of digestion by the microbiota that had suggested a few years earlier, as we have seen before.
In addition, in this work a lot of variety in composition of microbiota between individuals has been found (Figure 5 B).

Figure 5. Composition of the intestinal microbiota from 121 giant pandas, with (A) the dominant genera in all samples and (B) the relative contribution of each individual dominant genera, grouped by age and sampling time (Xue et al. 2015).
In this paper, a comparative analysis between the compositions of the intestinal microbiota of giant panda with other mammals has been made, and it has confirmed that the panda is grouped again with carnivores and is away from herbivores (Figure 6).

Figure 6. Principal component analysis (PCoA) of microbiota communities from faecal samples of 121 giant pandas (blank forms), compared with other herbivores (green), omnivores (blue) and carnivores (red). The different forms correspond to different works: the circles are from Xue et al. (2015), where this Figure has been obtained.
All in all, the peculiar characteristics of the giant panda microbiota contribute to the extinction danger of this animal. Unlike most other mammals that have evolved their microbiota and digestive anatomies optimizing them for their specific diets, the aberrant coevolution of panda, its microbiota and its particular diet is quite enigmatic. To clarify it and know how to preserve this threatened animal, studies must be continued, combining metagenomics, metatranscriptomics, metaproteomics and meta-metabolomics, in order to know well the structure and metabolism of gut microbiota and its relationship with digestive functions and the nutritional status of the giant panda (Xue et al. 2015).
References
Dierenfield ES, Hintz HF, Robertson JB, Van Soest PJ, Oftedal OT (1982) Utilization of bamboo by the giant panda. J Nutr 112, 636-641
Herron JC, Freeman S (2014) Evolutionary Analysis, 5th ed. Benjamin Cummings
Jin K, Xue C, Wu X, Qian J, Zhu Y et al. (2011) Why Does the Giant Panda Eat Bamboo? A Comparative Analysis of Appetite-Reward-Related Genes among Mammals. PLos One 6, e22602
Ley RE, Hamady M, Lozupone C, Turnbaugh PJ, Ramey RR et al. (2008) Evolution of Mammals and Their Gut Microbes. Science 320, 1647-1651
Li R, Fan W, Tian G, Zhu H, He L et 117 al. (2010) The sequence and de novo assembly of the giant panda genome. Nature 463, 311–317
Nie Y, Zhang Z, Raubenheimer D, Elser JJ, Wei W, Wei F (2015) Obligate herbivory in an ancestrally carnivorous lineage: the giant panda and bamboo from the perspective of nutritional geometry. Functional Ecology 29, 26–34
Rosen M (2015) Pandas’ gut bacteria resemble carnivores. Science News 19/05/2015
Wei G, Lu H, Zhou Z, Xie H, Wang A, Nelson K, Zhao L (2007) The microbial community in the feces of the giant panda (Ailuropoda melanoleuca) as determined by PCR-TGGE profiling and clone library analysis. Microb Ecol 54, 194–202
Wei F, Hu Y, Yan L, Nie Y, Wu Q, Zhang Z (2014) Giant Pandas Are Not an Evolutionary cul-de-sac: Evidence from Multidisciplinary Research. Mol Biol Evol 32, 4-12
Williams CL, Willard S, Kouba A, Sparks D, Holmes W et al. (2013) Dietary shifts affect the gastrointestinal microflora of the giant panda (Ailuropoda melanoleuca). J Anim Physiol Anim Nutr 97, 577-585
Xue Z, Zhang W, Wang L, Hou R, Zhang M et al. (2015) The bamboo-eating giant panda harbors a carnivore-like gut microbiota, with excessive seasonal variations. mBio 6(3), e00022-15
Zhao H, Yang JR, Xu H, Zhang J (2010) Pseudogenization of the Umami Taste Receptor Gene Tas1r1 in the Giant Panda Coincided with its Dietary Switch to Bamboo. Mol Biol Evol 27(12), 2669–2673
Zhu LF, Wu Q, Dai JY, Zhang SN, Wei FW (2011) Evidence of cellulose metabolism by the giant panda gut microbiome. Proc Natl Acad Sci USA 108, 17714–17719.
 grapes")
Bacteria of vineyard and terroir, and presence of Oenococcus in Priorat (South Catalonia) grapes
2nd May 2015
The vine growers believe that the land on which they grow vines gives the wines a unique quality, and that is called terroir. We can consider that the physiological response of the vines to the type of soil and climatic conditions, together with the characteristics of the variety and form of cultivation, result in a wine organoleptic properties that define their terroir (Zarraonaindia et al 2015 ). However, it is not known if there could be a very specific microbiota of each terroir, as this subject has been barely studied.
Wine microorganisms in the grapes? Saccharomyces is not there or it has not been found there
The main protagonists of wine fermentations, alcoholic one (yeast Saccharomyces cerevisiae) and malolactic one (lactic acid bacteria Oenococcus oeni) usually do not appear until the must grape is fermenting to wine, in the cellar. In normal healthy grapes, S. cerevisiae is hardly found.
Oenococcus oeni in the grapes ? We have found it !
Regarding O. oeni, so far very little has been published about its presence and isolation from the grapes. In some works, as Sieiro et al (1990), or more recently Bae et al (2006), some lactic acid bacteria (LAB) have been isolated from the surface of grapes, but not O. oeni. Only Garijo et al (2011) were able to isolate a colony (only one) of O. oeni from Rioja grapes. Moreover, DNA of O. oeni has been detected in a sample of grapes from Bordeaux (Renouf et al 2005, Renouf et al 2007) by PCR-DGGE of rpoB gene, although in these works no Oenococcus has been isolated.
I am pleased to mention that recently our team have managed to isolate O. oeni from grapes, and typify them, and we are now working on a publication about it (Franquès et al 2015). Indeed, our research team of lactic acid bacteria (BL-URV), together with colleagues working on yeasts from the same group “Oenological Biotechnology” (Faculty of Oenology at the Universitat Rovira i Virgili in Tarragona, Catalonia, Spain) is working on a European project, called “Wildwine “(FP7-SME-2012 -315065), which aims to analyse the autochthonous microorganisms of Priorat area (South Catalonia), and select strains with oenological potential. This project also involves the Priorat Appellation Council and the cellar Ferrer-Bobet, as well as research groups and associations wineries from Bordeaux, Piedmont and Greece. In the framework of this project we took samples of grapes (Grenache and Carignan) from several vineyards of Priorat (Figure 1), as well as samples of wines doing malolactic fermentation. From all them we got 1900 isolates of LAB. We optimized isolation from grapes from the pulp and juice with various methods of enrichment, and so we got 110 isolated bacteria from grapes, identified as O. oeni by specific molecular techniques. Once typified, we have found that the molecular profiles of these strains do not coincide with commercial strains and so they are autochthonous. In addition, some of these strains from grapes were also found in the corresponding wine cellars.

Figure 1. Taking samples of Grenache (left) and Carignan (right) in Priorat area to isolate lactic acid bacteria such as Oenococcus (Pictures Albert Bordons).
The microbiota of grapes
The grapes have a complex microbial ecology, including yeasts, mycelial fungi and bacteria. Some are found only in grapes, such as parasitic fungi and environmental bacteria, and others have the ability to survive and grow in wines: especially yeasts, lactic acid bacteria (LAB) and acetic acid bacteria. The proportion of all them depends on the maturation of the grapes and the availability of nutrients.
When the fruits are intact, the predominant microbiota are basidiomycetous yeasts as Cryptococcus and Rhodotorula, but when they are more mature, they begin to have micro fissures that facilitate the availability of nutrients and explain the predominance just before the harvest of slightly fermentative ascomycetes as Candida, Hanseniaspora, Metschnikowia and Pichia. When the skin is already damaged more damaging yeasts may appear, as Zygosaccharomyces and Torulaspora, and acetic acid bacteria. Among the filamentous fungi occasionally there may have some very harmful as Botrytis (bunch rot) or Aspergillus producing ochratoxin. Although they are active only in the vineyard, their products can affect wine quality.
On the other hand, environmentally ubiquitous bacteria have been isolated from the grapes skin, as various Enterobacteriaceae, Bacillus and Staphylococcus, but none of them can grow in wine (Barata et al 2012).
Coming back to the possible specific microbiota of terroir, it has been found that some volatile compounds contributing to the aroma of the wine, such as 2-methyl butanoic acid and 3-methyl butanol, are produced by microorganisms isolated in the vineyards, as Gram-positive bacterium Paenibacillus, or the basidiomycetous fungus Sporobolomyces or the ascomycetous Aureobasidium. Therefore, there could be a relationship between some of the microbial species found in grapes and some detected aromas in wine, coming from the must of course (Verginer et al 2010).
Metagenomics as analytical tool of microbiota from grapes
Since conventional methods of isolation and cultivation of microorganisms are slow, laborious and some microbes cannot be grown up in the usual isolation media, massive sequencing methods or metagenomics are currently used. These consist of analysing all the DNA of a sample, and deducing which are the present microorganisms by comparing the sequences found with those of the databases. For bacteria the amplified DNA of V4 fragment from 16S RNA gene is used (Caporaso et al 2012).
This technique has been used with samples of botrytized wines (Bokulich et al 2012) and various LAB have been found (but not Oenococcus), including some not normally associated with wine. It has also been used to see the resident microbiota in wineries and how it changes with the seasons, resulting that in the surfaces of tanks and machinery of the cellar there is a majority of microorganisms neither related with wine nor harmful (Bokulich et al 2013).
With this technique Bokulich et al (2014) have also analysed the grapes and they have seen clear differences between the proportions of bacterial groups (and fungi) from different places, different varieties, as well as environmental or bio geographical conditions. For example, when analysing 273 samples of grape musts from California, the 3 varieties (Cabernet, Chardonnay and Zinfandel) are quite discriminated in a principal components analysis with respect to the bacterial communities found in each sample (Figure 2).
Thus, the dominant bacterial taxa or groups in a variety or given environment could provide some specifics traits on those wines, and this could explain some regional or terroir patterns in the organoleptic properties of these wines (Bokulich et al 2014).

Figure 2. Principal component analysis of bacterial communities of grape musts samples of Sonoma (California) from 3 varieties (Cabernet in red, Chardonnay in green and Zinfandel in blue) (Bokulich et al 2014).
We have also carried out a massive sequencing study with the same grape samples from which we have obtained isolates of O. oeni, as said before (Franquès et al 2015), and in more than 600,000 analysed sequences of 16S rRNA, we have found mainly Proteobacteria and Firmicutes. Among these gram-positive, we have found sequences of lactic acid bacteria (15%) and from these we have successfully confirmed the presence of O. oeni in 5% of the sequences. Therefore, we have isolated O. oeni from grapes and we have detected their DNA in the samples.
The bacterial microbiota of the vineyards and soil
As we see, microbiota of grapes and wine has been studied a little, but the soil microbiota has not been characterized. This one can define more clearly the terroir, which is influenced by the local climate and characteristics of the vineyard.
In Figure 3 the main genera found in different parts of the vine and soil are summarized (Gilbert et al 2014).

Figure 3. Main bacteria and fungi associated with organs and soil of Vitis vinifera (Gilbert et al 2014)
Recently an interesting scientific work (Zarraonaindia et al 2015) has been published on this subject, with the aim to see if the soil could be the main original source of bacteria that colonize the grapes. These authors took samples of soil, roots, leaves, flowers and grapes from Merlot vines, from different areas and years, of Suffolk, New York, and they analysed the bacterial DNA by 16S rRNA sequencing. They found that 40% of the species found were present in all samples of soil and roots, while there was more variability in leaves and fruits, and moreover, 40% of those found in leaves and fruits were also found in soils. All this suggests that many bacteria originate in the soil.
Regarding the type of bacteria, they found that Proteobacteria (especially Pseudomonas and Methylobacterium) predominated (Figure 4), mainly in the aerial parts of the plant. There were also Firmicutes as expected, and Acidobacteria and Bacteroides.

Figure 4. Composition of the bacterial community, at Phylum level, in samples from different organs of the vine and its soil (Zarraonaindia et al 2015).
Although variations were observed in all samples depending on the year (there may be different climatic conditions) and according to different edaphic factors (pH, C: N, humidity), the principal-components analysis (Figure 5) showed that the main types of samples (soil, roots, leaves, grapes) differ quite well, and bacterial taxon composition in samples of grape juice before fermentation is similar to that of grapes.

Figure 5. Principal-components analysis showing the similarities in terms of the composition of bacterial taxonomic groups, among sample types, including musts (Zarraonaindia et al 2015).
This suggests that the bacterial community found in grapes remains relatively stable until the processing to musts, and that it is more stable than the differences between organs. At the same time, a large number of representatives of bacterial phyla of the grapes come from the soil. This can be explained because when grapes are harvested by hand, they are often placed in boxes that are left on the ground, or for mechanical harvest, the machinery used removes the soil and generates dust, which can colonize the grapes.
Therefore, the soil microbiota is a source of bacteria associated with vines and may play a role in the must and therefore in the wine, and potentially in the formation of the terroir characteristics. Some of these bacteria may have some roles not yet known in productivity or disease resistance of the plant, or contribute to the organoleptic characteristics of wine (Zarraonaindia et al 2015).
In addition, and thinking in wine microorganisms responsible for fermentations, as said, in our laboratory we have confirmed that there are some O. oeni strains in grapes and we have confirmed this by detecting their DNA in the same grapes.
References
Bae S, Fleet GH, Heard GM (2006) Lactic acid bacteria associated with wine grapes from several Australian vineyards. J Appl Microbiol 100, 712-727
Barata A, Malfeito-Ferreira M, Loureiro V (2012) The microbial ecology of wine grapes (Review). Int J Food Microbiol 153, 243-259
Bokulich NA, Joseph CML, Allen G, Benson AK, Mills DA (2012) Next-generation sequencing reveals significant bacterial diversity of botrytized wine. Plos One 7, e36357
Bokulich NA, Ohta M, Richardson PM, Mills DA (2013) Monitoring seasonal changes in winery-resident microbiota. Plos One 8, e66437
Bokulich NA, Thorngate JH, Richardson PM, Mills DA (2014) Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. PNAS nov 25, E139-E148
Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Huntley J, Fierer N, Owens SM, Betley J, Fraser L, Bauer M, Gormley N, Gilbert JA, Smith G, Knight R (2012) Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J 6, 1621–1624
Franquès J, Araque I, Portillo C, Reguant C, Bordons A (2015) Presence of autochthonous Oenococcus oeni in grapes and wines of Priorat in South Catalonia. Article in elaboration.
Garijo P, López R, Santamaría P, Ocón E, Olarte C, Sanz S, Gutiérrez AR (2011) Eur Food Res Technol 233, 359-365
Gilbert JA, van der Lelie D, Zarraonaindia I (2014) Microbial terroir for wine grapes. PNAS 111, 5-6
Renouf V, Claisse O, Lonvaud-Funel A (2005) Understanding the microbial ecosystem on the grape berry surface through numeration and identification of yeast and bacteria. Aust J Grape Wine Res 11, 316-327
Renouf V, Claisse O, Lonvaud-Funel A (2007) Inventory and monitoring of wine microbial consortia. Appl Microbiol Biotechnol 75, 149-164
Sieiro C, Cansado J, Agrelo D, Velázquez JB, Villa TG (1990) Isolation and enological characterization of malolactic bacteria from the vineyards of North-western Spain. Appl Environ Microbiol 56, 2936-2938
Verginer M, Leitner E, Berg G (2010) Production pf volatile metabolites by grape-associated microorganisms. J Agric Food Chem 58, 8344-8350
Zarraonaindia I, Owens SM, Weisenhorn P, West K, Hampton-Marcell J, Lax S, Bokulich NA, Mills DA, Martin G, Taghavi S, Van der Lelie D, Gilbert JA (2015) The soil microbiome influences grapevine-associated microbiota. mBio 6, e02527-14
