الأسماء في صفحات التنقل

The true grasses, family Poaceae (formerly Gramineae), is one of the most speciose plant families, comprising over 10,000 species with a Gondwanan origin approximated at about 80-100 million years ago (although there are fossil specimens that potentially push the origin earlier; Prasad et al. 2011; Vicentini et al. 2008; Stevens 2013).
Distributed world-wide, the true grasses are absent only in parts of Greenland and Antarctica, and are the most economically important group of monocots, as this family includes the true grains, pasture grasses, sugar cane, and bamboo.Species in this family have been domesticated for staple food crops (grains and sugar, for example), fodder for domesticated animals, biofuel, building materials, paper and ornamental landscaping, among other things.Grasslands cover at least 20% of the earth’s surface, although grasses also grow in biomes other than grasslands.
Grasses are primarily wind pollinated, most have dangling anthers.They have hollow stems and grow from the plant base, rather than the tip, as an evolutionary response to predation.Many also protect themselves from predation by secreting silica crystals in their leaves.There are two main kinds of grasses, cool-season (C3) and warm-season (C4) grasses, which are distinct in their means for fixing Carbon.The evolution of C4 fixation has arisen independently in 4 of the 12 currently recognized grass subfamilies; a combination of changes in paleoclimate including temperature, aridness, seasonality are thought to select for new origins of C4 lineages (Vicentini et al. 2008).
Genomic duplications are common in the true grasses, and thought to play important role in the evolution of the group as well as innovations leading to diversification of branches within Poaceae (for example, the evolution of flowers arranged as spikelets).
(The Plant List 2010; Prasad et al. 2011; Stevens 2013; Vicentini et al. 2008; Wikipedia 2013)
Van al die plante wat vir die mens nuttig is, is die lede van die grasfamilie (Poaceae of Gramineae) die belangrikste. Ons leef regstreeks en onregstreeks van gras: ons vernaamste stapelvoedsels word verskaf deur spesiaal gekweekte grasse (grane), bv. koring en rys, terwyl die meeste van die diere wat melk en vleis aan ons verskaf, hoofsaaklik gras vreet. Daar bestaan ook geen twyfel dat die grane die eerste plante was wat doelbewus deur die mens gekweek is nie.
Suikerriet (Saccharum officinarum) is inheems in Suidoos-Asië, en die produksie daarvan is beperk tot die subtropiese en tropiese klimaatstreke. Hierdie plant is die vernaamste bron van rietsuiker (sukrose). Die dik, sappige stamme van die plant bevat die suiker. By die bereiding van suiker word die stamme deur spesiale masjiene verpulp, waarna die sap ingedamp en geraffineer word. Suid-Afrika is 'n belangrike suikerproduserende land. Die suikerbedryf is feitlik uitsluitlik in KwaZulu-Natal gesetel.
Graansorghum (Sorghum vulgare) kom oorspronklik uit Noordoos-Afrika, maar het vandag 'n wye wêreldverspreiding. Verskeie tipes sorghum kom voor, nl. Graansorghum soetsorghum, grassorghum (lg. twee is voergewasse) en besemsorghum. Graansorghum is verreweg die belangrikste en die saad se voedingswaarde kom baie ooreen met die van mielies. Van die pluime van die besemsorghum word goeie, harde besems gemaak.
Graansorghum is deur die Bantoes tydens hul suidwaartse migrasie na Suid-Afrika gebring lank voor die koms van die blankes. Dit het 'n vername deel van hul dieet uitgemaak. Vandag word groot hoeveelhede graansorghum gebruik vir die maak van bier. 'n Aansienlike deel van die oes word egter ook as veevoer gebruik. Die jaarlikse oes het in die jare 1970–1977 gewissel tussen 220 000 en byna 700 000 t. Graansorghum is droogtebestand en sou op baie groter skaal gesaai word as die nodige aanvraag bestaan het.
Bamboes (subfamilie Bambusoidea): Hierdie plante vorm so 'n kenmerkende groep dat hulle as 'n subfamilie beskou word. Die meeste van hulle, by name die groot soorte, kom in tropiese gebiede voor. Party bereik 'n hoogte van 40 m, met stamme wat aan die onderpunt meer as 150 mm dik is. Die bamboessoorte is die lede van die grasfamilie wat gekenmerk word deur houtweefsels. In die tropiese lande van Asië speel bamboes 'n belangrike rol in die ekonomie; dit word onder meer gebruik vir die bou van huise en die maak van matte en meubels. Klein bamboessoorte word dikwels, ook in Suid-Afrika, as sierplante gekweek.
Die mielie (Zea mays) kom oorspronklik van Amerika en is eers aan die Europeërs bekend gestel na die ontdekking van Amerika in 1492. Dit word vandag in al die warmer dele van die wêreld verbou. Die graan dien as voedsel vir mens en dier. Die saad dien verder as basis vir talle nywerheidsprodukte, bv. kookolie, stysel, glukose en alkohol. Die mielie het 'n soliede stam en die manlike en vroulike blomme word in aparte bloeiwyses aan dieselfde plant gedra.
Die vroulike bloeiwyse (die kop) is 'n gewysigde aar, terwyl die manlike bloeiwyse 'n pluim is. Mielies is verreweg die vernaamste graansoort in Suid-Afrika en is op vier na die grootste mielieprodusent ter wêreld. Die grootste deel van die mielieoes kom uit die sg. Mieliedriehoek met punte Carolina, Ladybrand en Mafeking. Weens die wisselvalligheid van ons klimaat wissel die oes baie vanjaar tot jaar. Ons land kan egter reeds vir baie jare mielies jaarliks uitvoer, veral na Europa en Japan.
Koring (Triticum aestivum) is die belangrikste graansoort ter wêreld en die vernaamste stapelvoedsel in die Westerse lande. Dit word reeds sedert die vroegste tye deur die mens gekweek. Deur teling en seleksie, en selfs kruising van spesies, is duisende tipes en variëteite ontwikkel wat by 'n wye verskeidenheid klimaatstoestande aangepas is. Gewone koring kan verdeel word in harde korings, waarvan brood gebak word, en sagte korings, vir koek en beskuitjies.
Die sogenaamde Durumkoring (Triticum durum) word gebruik vir die maak van macaroni en verwante produkte. Die grootste deel van Suid-Afrika se jaarlikse koringoes van sowat 2 miljoen t kom uit die Vrystaat en uit die winterreënstreek van die Wes-Kaap, maar groot hoeveelhede koring word ook in Gauteng onder besproeiing verbou. Suid-Afrika was lank 'n koringinvoerland (veral uit die VSA, Kanada en Australië), maar vandag produseer ons genoeg koring vir eie gebruik.
Rys (Oryza saliva) kom oorspronklik uit Suidoos-Asië. Naas koring is dit die wêreld se vernaamste graan en dit dien as stapelvoedsel van die mensemassas van die Ooste. In teenstelling met ander graansoorte verkies rys vleierige grond. Gevolglik word die ryslande gedurende die groeityd met water oorstroom. China en Indië is die grootste produsente en verbruikers van rys. Die enigste Westerse lande waar noemenswaardige hoeveelhede rys gekweek word, is die Verenigde State en Brasilië. In Suid-Afrika was daar in die vroeë vyftigerjare aansienlike belangstelling in die verbouing van rys in die Gautengse Laeveld. Die huidige produksie is egter onbenullig. In Eswatini word ’n bietjie rys geproduseer, wat in die land verwerk word. Feitlik al ons rys word ingevoer, veral uit die Verenigde State.
Rog (Secale cereale) is die gehardste van al die wintergrane. Dit is na verwant aan koring, maar kan heelwat meer koue verdra en groei ook baie beter in skraal, sanderige grond. Vanweë sy bestandheid teen koue word rog in Noord-Europa en groot dele van Rusland as broodgraan in plaas van koring verbou. Rogbrood is egter donker van kleur en taamlik swaar. Die mout van die graan word ook gebruik vir die maak van sekere soorte whisky. In Suid-Afrika is in die verlede heelwat rog in die sg. Sandveld van die Wes-Kaap gesaai. Dit is egter mettertyd tot groot hoogte deur koring vervang. Tans beloop die totale produksie van die Republiek gewoonlik minder as 5 000 t en moet ons trouens rog invoer. Die meeste rog word nou in die Vrystaat geproduseer.
Gars (Hordeum-spesies): Daar is in hoofsaak twee garsspesies wat van mekaar onderskei word deur die aantal rye vrugbare syaartjies per aar. Hulle is onderskeidelik Hordeum vulgare, met ses rye en Hordeum distichum, met twee rye syaartjies. Ontkiemde gars (mout) word in die produksie van bier gebruik. Die tweerygars (Hordeum distichum) is veral geskik vir hierdie doel. Die graan word ook op groot skaal as veevoer gebruik. Die bekende gort is gars waarvan die doppies afgeskuur is. In Suid-Afrika word gars veral in die Suidwestelike dele van die Kaap verbou. Sesry- sowel as tweerygars word gesaai.
Daar is verskeie spesies wat as “mannas" geklassifiseer word. Die twee vernaamstes is egter Prosomanna (Panicum miliaceum) en Boermanna of gewone manna (Setaria italica). Hierdie twee-mannas kom oorspronklik uit die Ooste, waar veral Prosomanna steeds 'n belangrike graan is. Deurdat dit gou ryp word, kan dit met groot welslae in gebiede met 'n baie kort groeiseisoen gesaai word. In Westerse lande, onder meer ook in Suid-Afrika, word hierdie mannas feitlik uitsluitlik vir die produksie van saad vir kouvoëls gesaai.
(Avena-spesies): Hierdie graansoort kan maklik van koring onderskei word aangesien die bloeiwyse 'n los pluim is in teenstelling met die aar van koring. Dit is wel nie so ’n belangrike graansoort as koring nie, maar dien tog as menslike voedsel, veral as hawermoutpap. Hawergraan is 'n belangrike pitvoer vir diere, veral perde, maar die grootskeepse vervanging van perde deur trekkers het die belangrikheid daarvan sterk laat afneem. Hawer dien ook dikwels as weidingsgewas. Dit is ’n geharde plant wat beter as koring in onvrugbare grond aard.
Lede van die grasfamilie word dikwels vir verfraaiing in tuine aangeplant, hetsy as sierplante of vir grasperke. Van hierdie grasse kom die volgende in Suid-Afrikaanse tuine voor: Pampasgras (Ginerium argentium): Hierdie sterkgroeiende gras met pragtige wit pluime kom oorspronklik van die oop vlaktes of pampas van Suid-Amerika en dien as versiering in tuine deur ons land. Eulaliajaponica: Hierdie mooi polgras word veral ter wille van sy aantreklike groen blare met geel kolle geplant.
Fonteingras (Pennisetum ruppelii) word maklik gevestig, selfs in taamlik droë, skraal grond. Dit het baie dun halms en blare en aantreklike ligrooskleurige, donsagtige are. Die plante vorm digte polle wat ongeveer 'n meter hoog groei. Spaansriet (Arundo donax) is miskien nie ’n baie aantreklike plant nie, maar word dikwels in Suid-Afrikaanse tuine aangetref.
Van al die plante wat vir die mens nuttig is, is die lede van die grasfamilie (Poaceae of Gramineae) die belangrikste. Ons leef regstreeks en onregstreeks van gras: ons vernaamste stapelvoedsels word verskaf deur spesiaal gekweekte grasse (grane), bv. koring en rys, terwyl die meeste van die diere wat melk en vleis aan ons verskaf, hoofsaaklik gras vreet. Daar bestaan ook geen twyfel dat die grane die eerste plante was wat doelbewus deur die mens gekweek is nie.
Les gramínees o poacees (Poaceae) son una familia de plantes herbales, o bien raramente maderices, perteneciente al orde Poales de les monocotiledónees. Con más de 820 xéneros y cerca de 12 100 especies descrites,[2] les gramínees son la cuarta familia con mayor riqueza d'especies depués de les compuestes, les orquídees y les lleguminoses; pero, definitivamente, ye la primera n'importancia económica global.[3] Ello ye que la mayor parte de la dieta de los seres humanos provién de les gramínees, tantu en forma direuta —granos de ceberes y los sos derivaos, como farines y aceites— o indirecta —carne, lleche y güevos que provienen del ganáu y les aves de corrolada que s'alimenten de camperes o granos—. Ye una familia cosmopolita, que conquistó la mayoría de los nichos ecolóxicos del planeta, dende les zones desérticas hasta los ecosistemes d'agua salao, y dende les zones ablayaes y anegadizas hasta los sistemes montascosos más altos. Esta incomparable capacidá d'adaptación ta sofitada nuna enorme diversidá morfolóxica, fisiolóxica y reproductiva y en delles asociaciones mutualísticas con otros organismos, que converten a les gramínees nuna fascinante familia, non solo pola so importancia económica, sinón tamién pola so relevancia biolóxica.
Ente les especies más destacaes tán la caña d'azucre, el trigu, l'arroz, el maíz, el sorgo, la cebada, l'avena, el centenu o'l bambú.
Polo xeneral son yerbes, magar pueden ser maderices —como los bambúes tropicales—, cespitosas, rizomatusas o estoloníferas. Pola duración del so ciclu de vida pueden ser añales, bienal o perennes. Les gramínees añales, como ye lóxicu suponer, reprodúcense una sola vegada mientres el so ciclu vital —el casu del trigu o de l'avena, por casu—. Les especies perennes, sicasí, pueden reproducise delles vegaes —polo xeneral añalmente— o una sola vegada. Nel primer casu denominar iteróparas —la mayoría de les especies de camperes, por casu— y, nel segundu casu, semélparas —como ye'l casu de les distintes especies bambúes—.
Tienen tarmos cilíndricos a elípticos na so seición tresversal, articulaos, llamaos pelo normal cañes, polo xeneral con nuedos macizos y entrenudos buecos (pero pueden ser totalmente macizos como nel casu del maíz y dellos bambúes). Los nuedos son daqué más gruesos que los entrenudos y nellos nacen les fueyes y les yemes. Los entrenudos son dacuando daqué esnachaos na zona onde se desenvuelven les ramificaciones. Un pocu más arriba del nuedu esiste un meristema entrepolar en forma d'aníu que determina l'allargamientu del tarmu. En dellos xéneros esisten de dos a seis nudos bien próximos ente sigo (los cualos denominar nuedos compuestos), cada unu de los cualos lleva la so correspondiente fueya. En Cynodon dactylon, por casu, los nuedos tán en grupos d'a dos polos que les fueyes paecen opuestes. Polo xeneral los entrenudos basales son más curtios qu'el superiores; cuando hai dellos nuedos basales bien próximos, les fueyes paecen arrosetadas, esto ye, dispónense de forma tal qu'asemeyen formar una roseta basal de fueyes. Los principales tipos de tarmos nes gramínees son los siguientes:
Tienen fueyes de disposición alterna, dísticas, compuestes típicamente de vaina, lígula y llimbu. La vaina arrodia apretadamente al tarmu, los sos marxes se superponen pero nun se funden ente sigo (namái dacuando pueden ser atopaes formando un tubu). La lígula ye un pequeñu apéndiz membranosu, o raramente un grupu de pelos (tricomas), asitiáu na zona d'unión del llimbu cola vaina, na parte adaxial. El llimbu (o llámina) ye simple, usualmente llinial, con nerviación paralela. Puede ser esplanáu o dacuando endolcáu nun tubu, pue ser continuu cola vaina o tener peciolu. Amás d'esta descripción, ye necesariu encetar la variabilidá que puede topase pa cada unu d'éstos órganos:
Los macollos o macolles son la unidá estructural de la mayoría de les especies de gramínees. Formar a partir de les yemes axilares o secundaries del meristema basal de la exa principal. Cada unu d'estos biltos secundarios o macollos empecipien la so apaición cuando les plantes presenten ente dos y tres fueyes. Cada unu d'ellos, depués de producir les sos primeres fueyes, xenera'l so propiu sistema radicular. La suma o adición de macollos ye lo que conforma la estructura y la forma d'una planta de gramínea. Cuando les gramínees topar n'estáu vexetativu producen de cutio nuevos macollos y fueyes. Cada macollo, de la mesma, va empezar nel so momentu a producir nuevos macollos.
La inflorescencia elemental de les gramínees ye una pequeña espiga formada por una o más flores sentaes o sésiles sobre un raquis articuláu, de cutiu perbreve, llamáu raquilla y protexíu por bráctees maneros denominaes glumes. Esti tipu d'inflorescencia recibe'l nome d'espiguilla. Les flores pueden ser hermafrodites o unisexuales y presenten un periantu rudimentariu de dos o tres pieces, les lodículas o glumélulas. Estes lodículas son los órganos que, al ponese turgentes, determinen l'apertura del antecio o caxellu floral mientres el floriamientu, dexando que s'espongan los estigmes plumosos y los estames. Los antecios tán formaos pola lemma, xuntada a la raquilla, y la pálea ensertada sobre la exa floral que naz sobre la raquilla na axila de la lemma y soporta a los órganos florales puramente dichos. La lemma (o glumela inferior) tien forma de quilla, pue ser mútica o aristada y abraza a la pálea colos sos cantos. L'aresta naz na estremidá de la lemma o nel so envés. La pálea o glumela cimera ye llanceolada, binervada y ye como una tapa que zarra a la flor. Les glumas tán ensertaes sobre la raquilla, una más embaxo que la otra. Toos estos elementos son bien variables polo que ye conveniente analizalos por separáu.
Les espiguillas, de la mesma, tópense aconceyaes o arrexuntaes n'inflorescencies compuestes de tipu racimoso. Les más frecuentes son:
El frutu o granu de les gramínees ye un cariopse, frutu secu indehiscente, con una grana que la so testa ta soldada col pericarpiu formando una envoltura bien delgada. Esta envoltura zarra al embrión y al albume o endosperma. Esti frutu ye básicamente una variante del aqueniu, anque puede atopase una variedá de frutos na familia (ver por casu Werker 1997).[9] En dellos xéneros como Zizianopsis o Eleusine, el pericarpiu nun ta soldáu cola grana, de cuenta que'l frutu ye un aqueniu (o un utrículo según otru autores). En delles Bambúseas el frutu ye una nuez o una baga, mientres nel xéneru Sporobolus el pericarpiu ye mucilaginoso y dexa salir a la grana cuando s'enfiñe n'agua. Munchos xéneros, como Aristida, Stipa, Piptochaetium, Oryza y casi toles Paníceas tienen cariopses que s'esprenden de la planta envolubraos pola lemma y pola pálea. Nes Andropogóneas son les glumas les que persisten zarrando al cariopse. En Pennisetum y Cenchrus esprender tola espiguilla arrodiada d'un arreyo de goches o d'escayos. La forma del cariopse varia enforma según los xéneros, pudiendo ser casi circular como en Briza, oblongo como en Hordeum, llanceoláu como en Poa hasta casi llinial, como en Vulpia. Na parte inferior del cariopse vistu dorsalmente, vese l'embrión más o menos elípticu cubiertu pol pericarpiu tresparente. Del otru llau, correspondiente al riegu o sutura carpelar, estrémase, tamién por tresparencia, la mácula filar o filo (o zona d'unión de la grana col carpelu), que puede ser punctiforme, como en Poa y nes Paníceas, ovada, como en Briza subaristata, o llinial, como en Hordeum, Vulpia o Festuca.
L'embrión de les gramínees ye estructuralmente bien complicáu y consta de la plántula xunida a la so cotiledón llaminar, altamente modificáu, llamáu escudete. El cotiledón ye delgáu, parenquimatoso, llevando nel so parte esterior una capa de célules epiteliales que mientres la guañada secretan enzimes que hidrolizan les sustances de reserva alcontraes nel endosperma. La planta consta d'un nuedu cotiledonar, onde s'enserta'l cotiledón, una yémula cubierta d'un capuchón o coleoptilo y una radícula envuelta por otru capuchón o coleorriza. En munchos xéneros na parte esterna del nuedu cotiledonar hai una escama diminuta, el epiblasto, que pa dellos autores constitúi un restu d'un segundu cotiledón, ente qu'otros consideren que se trata d'un apéndiz de la coleorriza.[4][5]
El tamañu y el númberu de cromosomes tienen gran importancia na sistemática de les gramínees. Hai dos tipos cromosómicos estremos: el tipu festucoide caracterizáu por presentar cromosomes grandes y númberu básicu predominantemente x=7 y el tipu panicoide con cromosomes pequeños y númberos básicos predominantes x=9 y x=10. El tipu festucoide alcuéntrase en casi toles tribus de la subfamilia de les poóideas, con delles esceiciones. Por casu, la tribu Stipeae d'esta subfamilia tien cromosomes pequeños y númberos básicos x=9, 10, 11, 12, 14, 16 y 17. Les restantes subfamilies de les gramínees presenten el tipu cromosómico panicoide, con cromosomes pequeños y predominiu del númberu básicu x=9 y 10. Nes bambusóideas, erartóideas y arundinóideas los cromosomes son pequeños y el númberu básicu ye x=12. Na subfamilia Danthonioideae preséntense cromosomes de tamañu entemediu y númberu básicu x=6 y 7. La subfamilia Chloridoideae presenta cromosomes pequeños y dellos númberos básicos, x=7, 8, 9, 10, 11, 12 y 14. Les panicóideas siempres tienen cromosomes pequeños, con númberos básicos x=9 o x=10, anque esisten especies con otros númberos básicos, que varien dende x=4 a x=19.[5]
Les gramínees son morfolóxicamente distintes de cualesquier otra familia de plantes y, amás, son bien diverses tocantes a morfoloxía y vezu de crecedera. Les distintes especies de gramínees — como se describió na seición previa — difieren nos sos tamaños y númberos cromosómicos. Coles mesmes, difieren nel tamañu (o conteníu d'ADN) de los sos xenomes.
El xenoma del arroz, por casu, ye más de 11 vegaes más pequeñu que'l xenoma de la cebada, a pesar de que dambes especies son diploides y aparenten tener la mesma complexidá morfolóxica y fisiolóxica.[10]
El conteníu de xenes de les distintes especies de gramínees, sicasí, non varia tan llargamente como'l conteníu d'ADN total. L'arroz y la cebada, nuevamente, nun difieren más qu'en dos veces nel númberu promediu de fragmento de restricción qu'hibriden coles mesmes sondes.[11]
La mayor parte de les diferencies nel tamañu del xenoma ente especies de gramínees deber a diferencies nel ADN repetitivu. Los xenomes más grandes, como los de cebada o trigu, tán compuestos nun 75% d'ADN repetitivu, ente que los xenomes más pequeños, como'l del arroz, solo contienen menos del 50% d'ADN altamente repetitivu. Entá más, determinóse que bona parte d'esi ADN repetitivu ta compuestu de retrotransposones insertos ente los xenes.[12][13]
Los estudios de mapeo genómico en munches especies de gramínees utilizando les mesmes sondes d'ADN demostraron que non solo'l conteníu de xenes ta bien calteníu, sinón tamién l'orde de los xenes dientro de los cromosomes.
L'estensu caltenimientu nel conteníu de xenes y nel orde de los mesmos ente'l maíz y el sorgo nun ye inesperada una y bones dambes especies "namái" cunten con 15 a 20 millones d'años d'evolución independiente. Sicasí, similares observaciones pal arroz y el maíz, que diverxeron fai 60 a 80 millones d'años, indiquen que toles especies de la familia provienen d'un mesmu antepasáu común y que toes elles caltienen un mesmu repertoriu de xenes nel mesmu orde averáu.[14][15][16]
Los grandes rearreglos genómicos qu'estremen ente sigo a toles gramínees son la resultancia d'inversiones, translocaciones o duplicaciones cromosómicas qu'arreyen la mayor parte de los brazos cromosómicos.[17]
La mayoría, sinón toes, les gramínees son poliploides. Basaos nel supuestu que tolos xéneros y families que presenten un númberu cromosómico básicu x=12 son derivaos d'ancestros que sufrieron duplicaciones cromosómicas mientres la so evolución y que les subfamilies de gramínees más primitives (Anomochlooideae, Pharoideae, y Puelioideae) tienen un númberu cromosómico básicu x=12, puede deducise que l'ancestru de les gramínees yá yera un poliploide. Síguese, amás, que toles gramínees que se clasifiquen como diploides son, en realidá, paleopoliploides (esto ye, poliploides antiguos que presenten heriedu disómica[18]y que los sos proxenitores nun pueden ser identificaos por aciu ferramientes citogenéticas o marcadores moleculares).[19][20] La familia contién más del 60% d'especies, distribuyíes en tolos clados, que se clasifiquen como neopoliploides, esto ye que sufrieron un ciclu adicional de duplicación xenómica. Nestes especies, los xenomes doblaos nun diverxeron enforma del xenoma de los sos ancestros y el so númberu de cromosomes y comportamientu citolóxicu mientres la meiosis son indicativos de la duplicación cromosómica que los anició. La mayoría d'estos neopoliploides (más del 65%) derivaron de cruzamientos interespecíficos o intergenéricos polo que se-yos clasifica como alopoliploides.[19]
Les hemiceluloses y los polisacáridos de pectines de la paré celular primaria de les camperes son bien distintes de los de les demás espermatofites, tantu n'estructura como nes particularidaes de la composición de los xiloglucanos.[21] Los polisacáridos son menos ramificaos qu'en toles demás families de plantes, magar esta afirmación ta basada nun muestreo inda escasu d'especies. Les poacees pueden ser cianogenéticas o non. Cuando son cianogenéticas, los compuestos cianogenéticos son derivaos de la tirosina. Pueden presentar alcaloides (dacuando isoquinolina, pirrolizidina y indol). Raramente puede haber proantocianidines y cianidinas, en cantidaes traza, y namái en representantes de les subfamilies Panicoideae y Chloridoideae. Los flavonoides topáronse namái en dellos xéneros, Bouteloua Glyceria y Melica,[22] cuando tán presentes son quercetina, o kaempferol xuntu con quercetina. L'acedu elágico y l'arbutina nun s'atoparon en nengún miembru de la familia. Raramente atópense saponines y sapogenines, según tamién oxalatos llibres (por casu en Setaria).
Una xeneralización avera de la manera de reproducción de les gramínees ye que los miembros d'esta familia son plantes hermafrodites, que presenten fertilización cruciada (son alógamos) y se polinizan pol vientu. Obviamente, una familia con cerca de 10.000 especies cunta con munches esceiciones a esta regla, que descríbense de siguío.
Esti tipu de sistema reproductivu, nel cual esisten plantes femenines y plantes masculines, nun ye bien frecuente nes gramínees. Namái 18 xéneros son dioicos o presenten especies dioicas, siendo Poa el más conocíu d'ellos. Ello ye que les especies dioicas de Poa incluyir nun subxéneru separáu, Dioicopoa.[23]
Esti sistema reproductivu describe'l fechu de que nes poblaciones naturales d'una especie coesisten individuos femeninos ya individuos hermafrodites. Esta condición ye bastante rara nes gramínees. Bouteloua chondrosioides y delles especies del subxéneru Andinae de Poa son ginodioicas, magar Cortaderia ye l'exemplu más conspicuo.[24]
Nesti sistema los sexos tán separaos espacialmente pero nel mesmu individuu, esto ye, cada planta presenta inflorescencies femenines y masculines. Zea, Humbertochloa, Luziola, Ekmanochloa y Mniochloa son exemplos de xéneros con especies monoicas. Muncho más común ente les gramínees son les especies andromonoiques, una condición bien común nes Andropogóneas y Paníceas. Nes primeres, los dos sexos presentar en espiguillas distintos de pares heterógamos. Un par heterógamo d'espiguillas consiste usualmente nuna espiguilla sesil, con una flor neutra y otra hermafrodita, y una espiguilla pedicelada, con una flor neutra y otra masculina. Nes espiguillas bifloras de les Paniceas, sicasí, la flor inferior ye usualmente masculina o neutra, y la cimera ye hermafrodita. Dalgunos de los xéneros que ejemplifican esti tipu de sistema son Alloteropsis, Brachiaria, Cenchrus, Echinochloa, Melinis, Oplismenus, Panicum, Setaria, Whiteochloa, y Xyochlaena. Delles especies dientro d'estos xéneros pueden tener namái flores hermafrodites una y bones la flor inferior ye siempres neutra, raramente dambes flores son hermafrodites. Amás de les paníceas y les andropogóneas, Arundinelleae ye otra tribu con especies andromonoiques. Nel restu de la familia, les especies andromonoiques atópense bien esporádicamente, como por casu en Arrhenatherum, Hierochloe y Holcus.[24]
La gran mayoría de les especies de gramínees son hermafrodites, sicasí, frecuentemente son incapaces de producir granes cuando'l polen d'una planta poliniza los sos propios estigmes. Esto debe a qu'una gran parte de les especies de la familia presenten autoincompatibilidad, de tipu gametofítica y por cuenta de l'aición de dos xenes independientes (llamaos S y Z) con dellos apanfilos cada unu. Esti sistema d'autoincompatibilidad foi reparáu en dellos xéneros de la familia (Festuca, Secale, Lolium, Hordeum, Dactylis, ente munchos otros) y nun ye perfectamente eficiente. Ello ye que de la mayoría de les especies autoincompatibles puede llograse una proporción — magar amenorgada — de granes al autofecundar una planta.[24][25]
La autopolinización y l'autofecundación tán bien distribuyíes ente les gramínees. Polo xeneral, ye un mecanismu más común ente les especies añales qu'ente les perennes y, decididamente, muncho más frecuente ente les especies colonizadores. Esti mecanismu determinóse n'aprosimao 45 xéneros de gramínees, ente los cualos tópense xéneros económicamente bien importantes como Triticum, Oryza, Secale, Avena, Agropyron y Lolium. Una condición d'autogamia estrema ye la cleistogamia, na cual produzse la polinización y la fecundación dientro del antecio ensin que se produza la antesis. Esti últimu sistema ta distribuyíu en más de 70 xéneros pertenecientes a 20 tribus de gramínees.[24][26]
L'apomixis defínese como la reproducción asexual al traviés de granes. Nesti sistema reproductivu los embriones desenvolver por mitosis a partir d'una oósfera non amenorgada ensin que tenga llugar la fecundación. N'otres pallabres, cada embrión producíu ye genéticamente idénticu a llantar madre. Nes gramínees, la apomixis describióse per primer vegada en 1933 nuna especie de Poa. Dende aquel momentu identificóse esti mecanismu en cientos d'especies de poacees, particularmente nes Paníceas y nes Andropogóneas. Dalgunos de los xéneros que presenten especies apomícticas son Apluda, Capillipedium, Heteropogon, Themeda, Sorghum, Bothriochloa, Dichanthium, Cenchrus, Setaria y Paspalum.[24][27][28]
Les gramínees son una familia cosmopolita qu'habita dende los desiertos hasta los hábitats d'agua duce o marinos, y toles elevaciones salvo les más altes del planeta. Nel mundu desenvolviéronse estenses praderíes natives de gramínees onde hai seques periódiques, topografía plana o inclinada, quemes frecuentes, y en delles ocasiones onde hai llendo y so ciertes condiciones particulares de suelu. Les comunidaes apoderaes poles camperes suman el 24% de la vexetación del planeta, exemplos son les praderíes de Norteamérica, les pampes de Suramérica, el "veldt" o la sabana n'África, y les estepes euroasiáticas. Per fora de les praderíes herbales, los bambúes maderizos desempeñen un papel central na ecoloxía de los montes d'Asia tropical y templada.[3]
Les gramínees fueron ecológicamente esitoses y diversificáronse estensamente por cuenta de munches adaptaciones clave.[29] La espiguilla protexe a les flores pero, coles mesmes, dexa la polinización cuando les lodículas abren l'antecio. Coles mesmes, les espiguillas (lemmas con pelos o gabitos) tienen delles adaptaciones pa la dispersión del frutu. La versatilidad nos sistemes d'apareamiento, incluyendo l'autofecundación y l'apomixis, dexó a diverses especies de gramínees ser colonizadores esitoses de nuevos ambientes. L'anatomía de la fueya, que puede ser C3 o C4, dexa a estes especies esquizar y afaese a un ampliu rangu de hábitats. Los meristemas tópense allugaos na base de los entrenudos y na base de les vaines, protexíos por tola planta, lo que da como resultáu una adaptación al llendo y al fueu ensin igual ente toles plantes. El desenvolvimientu de les praderíes mientres el Miocenu (fai unos 25 a 5 millones d'años) pue fomentar la evolución de los grandes herbívoros, amás de representar una importante fonte d'alimentu y un estímulu pa la evolución del Homo sapiens.[3]
Les gramínees amás desenvolvieron ciertes carauterístiques fisiolóxiques que-yos dexaron conquistar hábitats onde prevalecen condiciones subóptimas pa la crecedera de les plantes. Una de tales carauterístiques ye la capacidá d'atropar betaínes de glicina y otros compuestos que se topen acomuñaos cola adaptación de les plantes a la crecedera en condiciones salines.[30] Per otru llau, les poóideas almacenen carbohidratos como fructanos, los que se topen en muncha menor concentración nes restantes especies de la familia. Esta carauterística ta acomuñada cola adaptación de tales especies a condiciones d'estrés hídricu (seques) y de baxes temperatures (xelaes).[31] Finalmente, otru mecanismu fisiolóxicu singular de les gramínees ye qu'aparentemente son la única familia d'anxospermes qu'adquier iones por quelación d'iones férricos con sideróforos que son absorbíos polos raigaños.[32]
Les micorrices y les micofiles son dos tipos de mutualismos nos cualos tópense arreyaos plantes cimeres y fungos. Les micorrizas son mutualismos ente fungos y los raigaños de les plantes. Les micofilas, entrín y non, son mutualismos ente fungos endófitos (aquellos que crecen dientro de les plantes) y la parte aérea de les plantes.[33] [34] Per otru llau, les gramínees tamién pueden acomuñóse con distintos xéneros de bacteries de vida llibre qu'afiten el nitróxenu atmosféricu.
La presencia de fungos endófitos puede modificar la supervivencia de les plantes de delles maneres yá que pueden producise tres tipos distintes d'asociaciones ente los simbiontes fúngicos y les plantes, que varien acordies col grupu taxonómicu del hospedante, coles estructures fúngicas y vexetales arreyaes y coles particularidaes intrínseques de la simbiosis.[35][36] Asina, el tipu I tien como hospedantes a miembros de la familia de les juncacees y a delles subfamilies de les gramínees. Estos fungos colonicen la totalidá de{ hospedante y producen les sos estructures sexuales n'estromes que suplanten a los frutos que tendríen de producir les plantes, polo qu'éstes pierden la capacidá de reproducise sexualmente. La interaición ye por demás agresiva pal hospedante y determina la so paulatina escayencia. El tipu II tien como hospedantes a les gramínees de la subfamilia de les poóideas. Non tolos individuos de la población colonizada presenten estromas en llugar de cariopses. Nos exemplares ensin síntomes esternos el miceliu endofítico llega a colonizar les cariopses ensin perxudicar la reproducción sexual de la planta. D'esta manera, el calter patóxenu de la interaición ye menor que nel casu anterior. A lo último, nel tipu III, que tamién se da na subfamilia de les poóideas, nunca apaecen estromas nos hospedantes; la colonización del fungu ye sistémica, algamando a les cariopses per mediu de les cualos arrobínase dicha asociación. Esti tipu d'interaición ye consideráu una simbiosis mutualista por cuenta de que los endófitos beneficien a los hospedantes aumentando la so crecedera, biomasa, tasa fotosintética, tolerancia a les xelaes y seques, resistencia a nematodos ya inseutos, polo qu'amonten la competitividá de les sos hospedantes. Coles mesmes, por cuenta de que producen alcaloides protexen a les plantes del ataque d'un ampliu espectru d'animales herbívoros, constituyendo asina parte del so sistema de defensa. La presencia d'endófitos afecta la palatabilidad de los camperes pa los herbívoros y tamién la palatabilidad de les granes pa los páxaros granívoros, los animales que comen material infectáu presenten diversos síntomes d'intoxicación.[37][38] El nivel de la infestación por áfidos y la de los sos parásitos y parasitoides, y entá el patrón y la tasa de descomposición de la campera muerta, tamién son afeutaos pol mutualismo.[39][40] Coles mesmes, los bárabos de mosques del xéneru Phorbia (o Botanophila) viven nel estroma de los fungos endofitos del xéneru Epichloë, y los adultos tresmiten los espermacios del fungu nuna forma análoga a la polinización por inseutos de les flores.[41] De la mesma, los endófitos ardíciense recibiendo l'apurra direutu de hidratos de carbonu producíos polos sos hospedantes. De les 232 micofilas conocíes nel mundu, 209 tienen como hospedantes a miembros de toles subfamilies de gramínees y representen a los trés tipos d'interaición. Les árees de distribución d'estes interaiciones tomen tantu zones fríes y templaes como tropicales. Nel Hemisferiu Norte son bien frecuentes les interaiciones tipu I, II ya III, ente que nel Hemisferiu Sur prevalecen les de tipu III.[42] Los fungos endofitos de la familia Clavicipitaceae tán llargamente distribuyíos ente les gramínees. Un xéneru d'esta familia de fungos, Epichloë, ye un endofito acutáu a la subfamilia Pooideae,[43][44][45][46][47] Neotyphodium ye l'estáu asexual o imperfectu de Epichloë.[48][49][50] Más del 30% de les especies de poóideas tán arreyaes en tales asociaciones, y esiste tresmisión d'estos fungos (la subfamilia Balansiae de les Clavicipitacees) tantu en forma vertical como horizontal.[51] Les micofilas son una asociación que paez datar de fai unos 40 millones d'años y una de les sos consecuencies ye la producción d'alcaloides como, por casu, la lolina.[52] Los alcaloides producíos, como la mentada lolina, son activos principalmente na defensa de les plantes contra inseutos.[53] Otres munches especies endófitas, aparentemente asintomáticas, pueden crecer conxuntamente nes gramínees, pero conozse bien pocu sobre les sos rellaciones. Márquez y collaboradores (2007), por casu, informaron que la gramínea Dicanthelium lanuginosum namái puede crecer en suelos calecíos por aición volcánica cuando el fungu endofítico Curvularia, col que ta acomuñada, tópase infectáu con un virus.[54] Esto indica que les rellaciones ente les gramínees y los fungos endófitos pueden ser desaxeradamente complexes y los sos efeutos insospechaos. Esisten llistaos d'especies de fungos endófitos acomuñaos con innumberables especies de gramínees.[55] Ello ye que namái nos bambúes hai siquier unes 1.933 especies de fungos descrites.
Los rexistros fósiles más antiguos indiquen qu'esta asociación tien unos 400 millones d'años, lo qu'indica la complexa coevolución ente les plantes y los sos fungos asociaos, que se manifiesta na amplia distribución del fenómenu (envaloróse qu'el 90% de les plantes terrestres tán micorrizadas) y na diversidá de mecanismos morfolóxicos, fisiolóxicos y ecolóxicos implicaos. Mientres la simbiosis, la planta hospedera recibe nutrientes minerales del suelu tomaos pol fungu (principalmente fósforu), ente que ésti llogra compuestos de carbonu deriváu de la fotosíntesis.[56] Los fungos formadores de micorrizas arbusculares constitúin micorrizas que colonicen el texíu interno de los raigaños de la planta hospedera, onde desenvuelven estructures carauterístiques de la simbiosis (arbúsculos y visícules), según miceliu extrarradical, el cuál interacciona col ecosistema de la rizósfera y ye l'encargáu d'estrayer nutrientes del suelu.[57] Nesti sentíu les gramínees nun son la esceición. Bona parte de les especies d'esta familia formen micorrizas, lo que favorez y optimiza la so adaptación a diversos tipos d'ambientes.[58]
El nitróxenu molecular (N2) ye la única reserva de nitróxenu accesible na biósfera. Práuticamente ilimitada, esta reserva nun ye direutamente utilizada polos vexetales y animales. El nitróxenu ye un constituyente esencial de molécules fundamentales de tolos seres vivos: aminoácidos, proteínes, acedos nucleicos, vitamines, ente les más importantes. Por que el nitróxenu atmosféricu pueda ser asimiláu, ye necesariu que sía amenorgáu. Les gramínees son capaces d'acomuñar con bacteries diazotróficas pertenecientes a los xéneros Azospirillum, Azotobacter, Azoarcus y Herbaspirillum les cualos realicen la fijación biolóxica del nitróxenu atmosféricu (N2). Estes bacteries son organismos de vida llibre capaces d'afitar nitróxenu dende la rizósfera, esto ye, dende la área circundante al sistema radicular de la planta ya incluyilo en compuestos (como'l amonio) fácilmente disponible y absorbible poles plantes. Amás d'afitar el nitróxenu atmosféricu, les bacteries diazotróficas favorecen el desenvolvimientu del sistema radicular de la planta cola cual conviven, al paecer al traviés de la producción de reguladores de crecedera o hormones. D'esta miente, favorecen una mayor absorción de nutrientes per parte de la planta. Informáronse medríes del orde de 5 % hasta 30 % nos rendimientos de gramínees como caña d'azucre, maíz, arroz, trigu y gramínees forrajeras como resultáu d'esta asociación. Estes asociaciones nun desenvuelven estructures estremaes nes que s'alluguen los microorganismos, como asocede nel casu de les lleguminoses y les bacteries del xéneru Rhizobium. En 1998 describióse otru tipu d'asociación de diazótrofos na cual la bacteria (llamada bacteria endófita) alcontrar nel interior del raigañu, el tarmu y les fueyes de la planta. Esta asociación foi afayada n'aislamientos de diazótrofos de plantes forrajeras de Paquistán, onde s'identificó una nueva bacteria fijadora de nitróxenu llamada Azoarcus. Esti microorganismu alcontrar nes capes esternes del córtex; una vegada nel interior de la planta tremar a los texíos aéreos probablemente per mediu de los vasos del xilema.[59][60]
Les gramínees son anemófiles, ye dicir qu'el polen ye tresportáu d'una planta a otra per mediu del vientu pa efeutuar la polinización. Nenguna poácea tien nectarios, magar delles camperes de montes tropicales —especialmente pequeñes bambusóideas— son polinizaes por inseutos.[61] La dispersión de les granes produzse principalmente por animales ya inclusive delles especies presenten estructures especializaes p'atraelos, como los elaiosomes.[62] Sicasí, la mayor parte de les especies presenten gabitos o aguyes por aciu los cualos los frutos o les diáspores xuntar a los animales que pasen. Munches especies se dispersn col vientu pa lo cual presenten llargos pelos nes arestes. Finalmente, Spinifex y dellos otros xéneros son plantes rodadoras, que son desarraigaes al maduror y tresportaes enteres pol vientu, esvalixando les sos granes mientres van rodando. Les arestes pueden ayudar tantu na dispersión por vientu como na dispersión por animales; la microestructura de la superficie de les arestes puede dar como resultáu qu'el cariopse sía direutamente "plantáu" nel suelu.[63][62]
Les especies maderices de bambúes son conocíes por floriar en forma sincronizada. Munches d'elles, amás, son perennes monocárpicas, ye dicir que vegetan bien d'años, florien una sola vegada y muerren depués de la dar les granes. Esta carauterística tamién s'atopa en dellos bambúes herbales. En plantes qu'amuesen esti tipu de reproducción, tolos miembros d'un mesmu clon florien simultáneamente, sía onde fora que fueren tresportaos en tol globu terrestre, y toles plantes, dempués d'un periodu reproductivu que puede dicise retrasáu, muerren.
Nos tarmos buecos de los bambúes munches vegaes atrópase agua, y vive nella una fauna distintiva. Les poacees aproven alimentu tanto pa los adultos (el polen) como pa les bárabos (los raigaños) de distintes especies d'escarabayos de la subfamilia Galerucinae de los crisomélidos.[64] Les gates de camparines de la familia Nymphalidaesobremanera les marrones Satyrinae y les emparentaes Morphinae, son comunes nos miembros d'esta familia (atopar n'alredor del 10% de los censos). Los inseutos del taxón Hemiptera-Lygaeidae-Blissinae reparar más comúnmente nes especies del cláu llamáu PACCMAD que nel cláu BEP.[65]
Son comunes nes poacees los fungos parásitos del orde Uredinales y los de la clase Ustilaginomycetes. Los qu'ataquen a les subfamilies Bambusoideae y Pooideae (incluyendo Stipa y parientes cercanos) son particularmente distintivos.[66] Los dos terceres partes de los Ustilaginales (unes 600 especies) alcuéntrase en Poaceae-[67][68]
Les gramínees y los sos parientes estinguíos daten de fai unos 89 millones d'años, el grupu principal diverxó fai unos 83 millones d'años.[69][70][71] Sacante los claos basales de la familia Anomochlooideae, Pharoideae y Puelioideae les espiguillas de les gramínees conocer na llende ente'l Paleocenu y l'Eocenu, fai unos 55 millones d'años,[72] y esta cifra ta a les traces en llinia con una estimación de la edá d'una duplicación del xenoma de les gramínees, asocedida fai unos 70-50 millones d'años.[73][74][75] Sicasí, el fósil d'una monocotiledónea (Programinis burmitis) perteneciente al Cretácicu tempranu (fai unos 100-110 millones d'años) ye similar a una gramínea bambusóidea. Magar esti fósil tien un númberu de calteres vexetativos que son comunes ente les poacees, la so identidá inda precisa confirmación.[76] Los texíos vexetales silicificaos (fitolitos) calteníos en fieces fosilizaes (coprolitos) de dinosaurios del Cretáceo tardíu topaes na India indiquen qu'a lo menos cinco taxones de gramínees escastaes taben presentes nel subcontinente indiu mientres esi periodu xeolóxicu (fai unos 71-65 millones d'años). Esta diversidá suxure que'l grupu basal de les gramínees se diversificaría y distribuyiría en Gondwana primero que la India quedara xeográficamente aisllada.[77]
La fotosíntesis C4 paez tar presente nes gramínees del Miocenu ceo a mediu, tantu nes Grandes Planicies de Norteamérica como n'África, fai unos 25-12,5 millones d'años. Quiciabes esti tipu de fotosíntesis tuvo primeramente acomuñada a cambeos adaptativos en respuesta a un amenorgamientu na concentración de CO2 na atmósfera, magar la gran espansión d'esti mecanismu fisiolóxicu asocedió fai namái unos 9-4 millones d'años. Entá nun ta claru si esti eventu tuvo amás favorecíu poles medríes na temperatura, l'amenorgamientu de les precipitaciones, l'aumentu de los vientos y la concomitante medría de quemes, que removeríen a los árboles de dellos hábitats nesi periodu.[78][79][80][81][82] Los detalles de los mecanismos de la fotosíntesis C4 y les morfoloxíes acomuñaes con ella son bien diversos y presenten una considerable variación, particularmente nel casu de la subfamilia de les panicóideas. Ello ye que la fotosíntesis C4 aparentemente anicióse y evolucionó independientemente hasta ocho veces nesta subfamilia.[83] Coles mesmes, esti mecanismu aniciar de manera independiente n'otres subfamilies, como Micrairoideae, Aristidoideae y Chloridoideae.[84] Independientemente de la so mayor eficiencia fotosintética, les gramínees C4 presenten menor conteníu de nitróxenu, mayor cantidá de fibres d'esclerénquima y pueden ser menos palatables que les gramínees C3.[85] A pesar d'estes carauterístiques, esistió una radiación de mamíferos herbívoros nel Miocenu,[86] que pudo tar acomuñada cola ampliación de les praderíes y sabanes, apoderaes por camperes. Sicasí, cuando les especies de gramínees de pradería espandir escontra Nebraska nel Miocenu ceo —fai unos 23 millones d'años— los ungulaos hipsodontos yá esistíen aquel día.[87]
La inflorescencia de les gramínees ye una estructura nueva nel repertoriu reproductivu de les plantes con flores. Ye entrevesgada dende'l puntu de vista de la bioloxía del desenvolvimientu, tien una importancia central na agronomía y, finalmente, ye una verdadera intriga evolutiva. La so arquiteutura controla'l tipu de polinización y la producción de granes, polo que ye una diana perimportante tantu pa la seleición natural como pal meyoramientu xenéticu y la bioteunoloxía. Ye de destacar que la diversidá d'estructures que presenten les espiguillas y espigues de les gramínees tean controlaes por xenes qu'afecten el desenvolvimientu y que nun se topen presentes en nenguna otra familia de plantes. Estos xenes aniciáronse depués d'estenses duplicaciones del xenoma y la posterior diversificación funcional de los mesmos.[88][89][90][91][92][93]
La pálea (y aparentemente tamién la lemma) quiciabes sía derivada de la mota, y les lodículas quiciabes sían derivaes de la corola (Ambrose et al. 2000[94]). Per otru llau un estudiu de morfoloxía comparativa suxure que la lemma ye una bráctea y que la pálea representa dos tépalos connaos del verticilu más esternu (Whipple y Schmidt 2006[95]). Teniendo en cuenta les rellaciones de parentescu cercanu ente Ecdeicoleaceae y Joinvilleaceae apocayá atopaes por Marchant y Briggs (2007[96]) y la probabilidá de que les flores de Anomochloa sían sui generis, la morfoloxía floral de Streptochaeta puede ser plesiomórfica (ancestral) na familia. Ye interesante que les flores de Ecdeicolea tamién son notablemente monosimétricas, colos dos tépalos adaxiales del verticilu esternu más llargos y aquillaos, y magar esto nun ye relevante en forma direuta, una diferenciación comparable nel verticilu esternu del periantu asocede en Xyridaceae, toos estos probablemente sían paralelismos. Una interpretación más común de la pálea ye que ye de naturaleza profilar/bracteolar, les monocotiledónees comúnmente tienen profilos bicarenaos, sicasí, paez que les bracteoles tuvieron que remanecer en Poaceae, una y bones los claos más emparentaos pero esternos a la familia ("outgroups" n'analís cladístico) nun les tener. Les lodículas paecen tar arreyaes na apertura de les flores estaminaes y les perfectes, ente que pueden tar ausentes nes flores pistilaes (Sajo et al. 2007[97]).
Los analises filoxenéticos utilizando secuencies espreses del ADN y l'estructura xeneral de los xenomes suxuren que les gramínees difieren muncho más d'otres monocotiledónees de lo que s'estremen éstes de les dicotiledónees.[98] Tales conclusiones alrodiu de les rellaciones de les poacees coles otres families de monocotiledónees y coles dicotiledónees nun son sorprendentes. Ello ye que les gramínees son fácilmente reconocibles y identificables de cualesquier otra familia, y la so monofilia tópase sofitada tantu pola morfoloxía como polos analises moleculares d'ADN. Los calteres fenotípicos que sofiten la monofilia de la familia son la inflorescencia con bráctees, el periantu amenorgáu, el tipu de frutu y los calteres del embrión y de la paré del granu de polen. Les semeyances coles ciperacees (Cyperaceae) nel vezu de cecimiento y nel tipu d'espiguillas representen una evolución converxente, y non una sinapomorfía. Ello ye que les ciperacees tán más emparentaes colos xuncos (Juncaceae) que coles gramínees, que pertenecen al nucleu de los Poales.
La importancia económico y ecolóxico de la familia motivó la realización d'una importante cantidá d'estudios sistemáticos. A principios del sieglu XIX, les diferencies ente les espiguillas de les poóideas y les panicóideas llevaron a Robert Brown a estremar a la familia nestos dos grupos básicos. A principios del sieglu XX, los calteres de la epidermis de les fueyes y el númberu de cromosomes llevó a la separación de les clorídeas de les poóideas. A mediaos del sieglu XX, l'anatomía interna de la fueya (en particular, la presencia o ausencia de l'anatomía Kranz y los calteres del embrión (presencia o ausencia d'epiblasto y ciertes carauterístiques del nuedu cotiledonar), llevaron a la reconocencia de cinco a ocho subfamilies. Dende fines del Sieglu XX, los estudios filoxenéticos basaos en delles secuencies de xenes demostraron ser congruentes con munches de les rellaciones filoxenétiques inferíes primeramente al traviés de calteres estructurales y fisiolóxicos. Los estudios moleculares más avanzaos sofiten la reconocencia de 13 subfamilies.[99][100][101][102][103]Los trés primeros llinaxes que diverxeron son Anomochlooideae (nativa de Brasil), Pharoideae (nativa de los trópicos del Viejo y Nuevu Mundu) y Puelioideae (nativa del oeste d'África). Los integrantes d'estos trés grupos solo son unes 25 de les casi 10.000 especies de la familia. El restu de les especies distribuyir en dos grandes grupos. El primeru, llamáu clado BEP, arrexunta a Bambusoideae, a Ehrharttoideae, y a Pooideae. El segundu, denomináu cláu PACCMAD, arrexunta a Panicoideae, Arundinoideae, Chloridoideae, Centothecoideae, Micrairoideae, Aristidoideae y Danthonioideae.[104] El cláu PACCMAD ta sofitáu por un calter del embrión: un llargu entrenudo nel mesocótilo. L'anatomía C3 ye l'estáu plesiomórfico o ancestral na familia. Toles especies C4 atopar nel cláu PACCMAD.
Toles subfamilies mentaes son monofiléticas, anque namái unes poques tienen sinapomorfías morfolóxiques que caractericen a tolos sos miembros. Más bien el so monofilia tópase sostenida por grupos de calteres morfolóxicos que tienen de reparase en xunto. Les rellaciones dientro de los grandes claos PACCMAD y BEP son nel so mayor parte pocu clares inda, de fechu la posición de les poóideas ye pocu clara en dellos analises.[105][106] El cladograma qu'amuesa les rellaciones ente los 13 claos mentaos ye'l siguiente:[103]
Poaceae
cláu BEP
Arundinoideae sensu stricto
La familia foi reconocida por tolos sistemes de clasificación de plantes, empezando pol de Carlos Linneo na so obra Systema naturae pasando polos d'Adolf Engler,[107] Arthur Cronquist,[108] Armen Takhtajan,[109] ente munchos otros. Los sistemes modernos de clasificación (sistema de clasificación APG III,[110] y los dos sistemes filoxenéticos previos, APGI y APGII)[111] tamién reconocen a esta familia.
Ente los calteres diagnósticos de la familia topen dellos de los órganos yá descriptos: les fueyes que tienen vaines llargues y abiertes y les lígulas na unión ente la vaina y la llámina, los tarmos redondos y usualmente buecos nos entrenudos, y les inflorescencies que la so unidá básica ye la espiguilla. Les flores individuales son pequeñes, con periantu inconspicuo y con un xinecéu que usualmente tien dos estigmes plumosos y un únicu óvulu. Nel frutu secu, de tipu aqueniu (cariopse), l'embrión —relativamente grande— ocupa una posición llateral y allúgase xuntu al tegumentu de la grana.
Son particularmente preseos pa la identificación de los xéneros los calteres de la espiguilla, como'l tamañu, el planu de compresión, la presencia o ausencia de glumas, el númberu de flores por espiguilla, la presencia de flores maneres o incompletes, el númberu de venes nes glumas y les glumelas, la presencia o ausencia d'arestes, y la forma de les inflorescencies secundaries.
De siguío bríndase una descripción de les subfamilies de les gramínees y de los xéneros más importantes o representativos dientro de caúna.
Les anomoclóideas son plantes herbales y presenten inflorescencies con una morfoloxía carauterística que nun se paecen a les espiguillas típiques o avezaes de les restantes gramínees. Paecen ser el cláu hermanu de too el restu de la familia,[99] Soreng y Davis 1998[100] lo cual suxure que la espiguilla carauterística de les camperes probablemente anicióse depués de que les anomoclóideas diverxeren del restu de les gramínees. Los integrantes d'esta subfamilia presenten pseudopecíolu con un pulvínulo apical, la lígula de les fueyes tresformada nuna guedeya de pelos y les cañes de la inflorescencia cimosas. Na inflorescencia presenten dos bráctees a lo llargo de cada ramificación y dos más debaxo de cada flor, o bien les flores disponer en forma espiralada a lo llargo d'exes racimosos, con delles bráctees debaxo de cada flor. Les anteres son centrifijas (en Anomochloa) o casi basifijas (Streptochaeta). Carauterísticamente, la primer fueya de la plántula escarez de llámina foliar. Los númberos cromosómicos básicos son x=11 y 18. Les especies pertenecientes a esta subfamilia fueron orixinalmente incluyíes dientro de les bambusóideas, pero anguaño reconozse que namái tienen un parentescu distante con elles. La subfamilia inclúi a dos xéneros —Anomochloa y Streptochaeta— con cuatro especies qu'habiten selves y distribúyense dende Centroamérica hasta'l sudeste de Brasil.[112]
Les faróideas caracterizar poles sos fueyes resupinaes, esto ye, coles llámines foliares cola cara abaxal escontra riba. Les espiguillas son unifloras y presenten seis estames coles anteres centrifijas. El coleoptile presenta llámina. Esta subfamilia inclúi cuatro xéneros y unos dolce especies pantropicales, que habiten nes selves.[112] Esta subfamilia, xuntu con Anomochlooideae y Puelioideae, foi tradicionalmente incluyida dientro de les bambusóideas, pero los estudios filoxenéticos sobre datos moleculares demostraron que los trés son los clados basales de tola familia de les gramínees. Les faróideas estremar de les puelióideas poles sos espiguillas unifloras y los sos estigmes trífidos.[112]
Les puelióideas xuntu coles demás subfamilies de gramínees, pero sacante les puelióideas y les anomoclóideas, presenten xineceos con dos estigmes y espiguillas que se desarticulen percima de les glumas al maduror. Caracterícense amás pol so androcéu con seis estames. Inclúi dos xéneros —Guaduella y Puelia— y aprosimao once especies distribuyíes per África tropical.[112]
Les bambusóideas, en sentíu acutáu, inclúin tantu especies herbales como maderices y son casi puramente tropicales na so distribución. Les fueyes son pseudopeciolaes. Les flores presenten trés lodículas y un androcéu con seis estames, raramente de dos a 14. L'ovariu lleva dos o tres estigmes, raramente unu solu. La primer fueya de les plántulas nun presenta llámina. Los númberos cromosómicos básicos son x=7 y x=9 a 12. Inclúi 112 xéneros con aprosimao 1647 especies tropicales a templaes.[2]
La diversificación dientro de los bambúes asocedió fai 30 a 40 millones d'años. Los bambúes maderizos formen un grupu monofiléticu hermanu del cláu que contién a les especies herbales. Los bambúes maderizos, colos sos tarmos d'hasta 40 metros d'altor, verdaderamente nun se paecen al verde. El floriamientu en munches d'estes especies tamién ye inusual, yá que asocede en ciclos d'hasta 120 años. Entá cuando los tarmos individuales viven por namái una o unes poques décades, dalguna forma de "reló" fai que los tarmos florien toos coles mesmes en tol rangu de distribución de la especie, causando dacuando sópitos cambeos ecolóxicos, como los que s'acomuñar a les rataes. Dellos xéneros de bambúes maderizos son Bambusa (120 especies), Chusquea (100 especies), Arundinaria (50 especies), Sasa (50 especies), y Phyllostachys (45 especies).[112]
Los miembros d'esta subfamilia presenten espiguillas con glumas bien amenorgaes y l'androcéu con seis estames, raramente con unu solu. Los númberos cromosómicos básicos d'esta subfamilia son x=10 y x=15. Entiende 21 xéneros y 111 especies, ente les que s'inclúin a los miembros de les tribus Ehrharteae del Hemisferiu Sur, según al cosmopolita Oryzeae.[2] Esti postreru ye acuáticu o de tierres húmedes. El representante más conocíu de la tribu Oryzeae ye l'arroz asiático Oryza sativa, unu de los cultivos más importantes del mundu. Nel norte d'África tamién se cultiva otra de les 22 especies de Oryza: O. glaberrima. N'Estaos Xuníos cobra importancia otra especie de la tribu, Zizania aquatica, l'arroz montés norteamericanu.[112]
Esta subfamilia ye la más grande de les gramínees. Consiste en 194 xéneros qu'integren unes 4200 especies.[2] Distribuyir nes rexones de clima templáu de too el globu. Ente los xéneros sobresalientes inclúyense importantes ceberes, como'l trigu, la cebada y l'avena, y tamién al centenu (Secale cereale), a les camperes utilizaes pa verde (como Poa, con 500 especies), pa segáu (Festuca, 450 especies), pa pasturas (como Phleum, Dactylis), y delles meruxes (como Agrostis, con 220 especies, y Poa). Otros xéneros importantes d'esta subfamilia son Stipa (300 especies), Calamagrostis (270 especies), Bromus (150 especies), y Elymus (150 especies).[2]Les poóideas estrémense porque les ramificaciones principales de la inflorescencia son dísticas, la lemma usualmente consta de cinco nervios. Amás, presenten oligosacáridos derivaos de la fructosa nel tarmu. Presenten cromosomes usualmente llongures y el númberu cromosómico básicu ye x=7, más raramente x=2, 4, 5 ó 6.[112]
Los integrantes de les cloridoideas presenten espiguillas que se desarticulen percima de les glumas y pelos bicelulares distintivos na epidermis de les fueyes. Sicasí, esti últimu calter puede ser una sinapomorfía de namái un subgrupu del cláu. Tol cláu salvu dos especies amuesa fotosíntesis pela vía del C4. Los númberos cromosómicos básicos prevalecientes na subfamilia son x=9 y x=19, anque esisten xéneros con x=7 y 8. La subfamilia distribúyese principalmente en rexones tropicales grebes y semiáridas, onde se supón que la fotosíntesis C4 ye ventaxosa. Los centros de distribución allugaos n'África y Australia suxuren un orixe nel Hemisferiu Norte. Dellos xéneros importantes son Eragrostis (350 especies), Muhlenbergia (160 especies), Sporobolus (160 especies), Chloris (55 especies), Spartina (15 especies) y Eustachys (10 especies).[112]
Les panicóideas fueron reconocíes taxonómicamente dende fai enforma tiempu, por cuenta de los sos espiguillas carauterístiques. Les cañes son usualmente sólides, les espiguillas tópense estruyíes dorsalmente, nun presenten raquilla y son bifloras. La desarticulación de la espiguilla al maduror producir por debaxo de les glumas. El tipu de fisioloxía de la fotosíntesis prevaleciente ye la C4. Los gránulos d'almidón nel endosperma son simples. Los númberos cromosómicos básicos más típicos son x=5, 9 y 10, anque tamién se topen especies con x=7, 12 y 14. La subfamilia ye principalmente tropical y contién dos grandes tribus, Andropogoneae y Paniceae, xuntu con un númberu de grupos pequeños. Les andropogóneas son relativamente fáciles de reconocer por cuenta de les sos espiguillas dispuestes d'a pares. Les paníceas nun son tan homoxénees como los miembros d'Andropogoneae. La subfamilia entiende 203 xéneros y 3.600 especies.[2] Ente los xéneros más importantes inclúyense Panicum (470 especies, polifilético), Paspalum (330 especies), Andropogon (100 especies), Setaria (100 especies), Sorghum (20 especies), y Zea (4 especies). El sorgo y el maíz son dos cultivos de gran importancia económica y dambos inclúyense nesta subfamilia.[112]
Les centotecoideas son una subfamilia ruinamente estudiada. Ta constituyida por unes 30 especies distribuyíes en 11 xéneros qu'habiten selves templaes templaes a tropicales. La so carauterística más distintiva ye la presencia d'estilu na flor y d'epiblasto nel embrión. El númberu cromosómico básicu más frecuente ye x=12, anque tamién hai xéneros con x=11.[112]
Con 14 xéneros y ente 20 a 38 especies, les arundinoideas son una subfamilia que la so esacta delimitación inda nun ye clara. Son gramínees hidrofíticas a xerofíticas qu'habiten rexones templaes a tropicales. Los sos númberos cromosómicos básicos son x=6, 9 y 12. Arundo (con 3 especies, Arundo donax ye la caña de Castiella) y Phragmites (2 especies) son los xéneros más conocíos d'esta subfamilia.[112]
Esta subfamilia monofilética perteneciente al cláu PACCMAD foi reinstalada y circunscripta nuevamente nel añu 2007. Los miembros de la mesma presenten estomas con célules subsidiarias en forma de domo, les lígulas con guedeyes de pelos, embriones pequeños, fotosíntesis C4, granos d'almidón simple nel endosperma. Entiende 8 xéneros y unes 170 especies na so mayoría tropicales. Dalgunos de los xéneros (como por casu Eriachne) nun taben asignaos a nenguna familia hasta apocayá tiempu y otros taben incluyíos n'otres subfamilies (exemplu Isachne nes Panicoideas).[104] Inclúi a los xéneros Isachne (100 especies), Eriachne (35 especies) y Micraira (8 especies).[112]
Les aristoideas inclúin 3 xéneros y de 300 a 385 especies de rexones templaes templaes, con arestes con una columna basal y fotosíntesis del tipu C4. Los númberos cromosómicos básicos son x=11 y x=12. Entiende al gran xéneru Aristida (230 a 330 especies) y Stipagrostis (50 especies).[112]
Danthonioideae ye una subfamilia bastante distribuyida en tol globu, especialmente nel Hemisferiu Sur. Presenten profilos bilobaos y les sinérgides del sacu embrionariu de tipu haustorial. Los númberos cromosómicos básicos son x=6, 7 y 9. Entiende 19 xéneros y unes 270 especies. Ente los xéneros con mayor númberu d'especies tópense Danthonia (100 especies) y Rytidosperma (90 especies). Cortaderia selloana ye la popular "cortadera" una gramínea ornamental que s'inclúi dientro d'esta subfamilia.[112]
El llistáu de tolos xéneros conocíos de gramínees, ordenaos alfabéticamente, aprovir nel anexu denomináu xéneros de Poaceae. En munchos casos se listan sinónimos. Los enllaces a sinónimos lleven al nome de xéneru preferíu.
La familia o dalguna de les sos subfamilies tienen los siguientes sinónimos:
Aegilopaceae, Agrostidaceae, Alopecuraceae, Andropogonaceae, Anomochloaceae, Arundinaceae, Arundinellaceae, Avenaceae, Bambusaceae, Chloridaceae, Eragrostidaceae, Festucaceae, Hordeaceae, Lepturaceae, Melicaceae, Miliaceae, Nardaceae, Oryzaceae, Panicaceae, Pappophoraceae, Parianaceae, Phalaridaceae, Pharaceae, Saccharaceae, Spartinaceae, Sporobolaceae, Stipaceae, Streptochaetaceae, Triticaceae, Zeaceae.[113]
La familia de les gramínees ye probablemente la que mayor importancia tien pa la economía humana.[4] Ello ye que alredor del 70% de la superficie cultivable del mundu ta semada con gramínees y el 50% de les caloríes consumíes pola humanidá provién de les numberoses especies de gramínees que son utilizaes direutamente na alimentación, o bien, indirectamente como forraxes pa los animales domésticos. En términos de la producción global, los 4 cultivos más importantes son gramínees: caña d'azucre (Saccharum officinarum), trigu, arroz y maíz. La cebada y el sorgo tán ente los primeres 12. Per otru llau, delles especies de gramínees utilizar na industria.
 Esta páxina forma parte del wikiproyeutu Botánica, un esfuerciu collaborativu col fin d'ameyorar y organizar tolos conteníos rellacionaos con esti tema. Visita la páxina d'alderique del proyeutu pa collaborar y facer entrugues o suxerencies.
Esta páxina forma parte del wikiproyeutu Botánica, un esfuerciu collaborativu col fin d'ameyorar y organizar tolos conteníos rellacionaos con esti tema. Visita la páxina d'alderique del proyeutu pa collaborar y facer entrugues o suxerencies. Les gramínees o poacees (Poaceae) son una familia de plantes herbales, o bien raramente maderices, perteneciente al orde Poales de les monocotiledónees. Con más de 820 xéneros y cerca de 12 100 especies descrites, les gramínees son la cuarta familia con mayor riqueza d'especies depués de les compuestes, les orquídees y les lleguminoses; pero, definitivamente, ye la primera n'importancia económica global. Ello ye que la mayor parte de la dieta de los seres humanos provién de les gramínees, tantu en forma direuta —granos de ceberes y los sos derivaos, como farines y aceites— o indirecta —carne, lleche y güevos que provienen del ganáu y les aves de corrolada que s'alimenten de camperes o granos—. Ye una familia cosmopolita, que conquistó la mayoría de los nichos ecolóxicos del planeta, dende les zones desérticas hasta los ecosistemes d'agua salao, y dende les zones ablayaes y anegadizas hasta los sistemes montascosos más altos. Esta incomparable capacidá d'adaptación ta sofitada nuna enorme diversidá morfolóxica, fisiolóxica y reproductiva y en delles asociaciones mutualísticas con otros organismos, que converten a les gramínees nuna fascinante familia, non solo pola so importancia económica, sinón tamién pola so relevancia biolóxica.
Ente les especies más destacaes tán la caña d'azucre, el trigu, l'arroz, el maíz, el sorgo, la cebada, l'avena, el centenu o'l bambú.
Qırtıckimilər və ya taxıllar (lat. Poaceae) — qırtıcçiçəklilər sırasına aid bitki fəsiləsi.[1]
Fəsilənin nümayəndələri birillik və ya çoxillik otlar, yerüstü hissələri bəzən odunlaşan, hətta ağacşəkilli bitkilərdir. Taxılkimilərin qədim ekoloji tiplərindən bambuk, qarğı, murquz kimi hündür gövdəli növləri də vardır. bu fəsiləyə daxil olan növlər saçaqlı, sürünən, kökümsov bitkilər olub, əksəriyyəti çim əmələ gətirəndir. Yarpaqları növbəli olmaqla iki cərgə üzrə düzülür, xətvarıdır, aşağısında buğum aralarını əhatə edən qın əmələ gətirir. Çiçəklər ikicinslidir, ziqomorfdur, mürəkkəb çiçək pulcuqları ilə əhatə olunaraq sünbülcüklərdə yerləşir. Çiçəkyanlığı iki dairədə yerləşmiş pərdəşəkilli pulcuqlardan təşkil olunmuşdur. Çiçəkyanlığının xarici dairəsində iki ədəd aşağı və yuxarı çiçək pulcuğu vardır. Erkəkcik 3 ədəddir. Yumurtalıq üst vəziyyətdədir. Bir yuvalı və bir intequmentli bir yumurtacığa malikdir. Ağızcıqlar oturaq olub, 2 ədəddir. Meyvə dən, nadir hallarda (bambuklarda) çəyirdək və ya giləmeyvədir. Toxum endospermlidir.[1]
Bu qrup bitkiləri A.A.Qrossheym (1939), V.D.Aleksandrovna (1941), İ.V.Larin (1950), Y.M.İsayev və b. (1965), Ə.İ.Mayılov (1965), S.H.Musayev (1980), V.S.Xəlilov (1971) və s. tədqiqatçılar öyrənmişlər. Azərbaycan florasında taxılların 306 növü məlumdur. Taxıl fəsiləsinin nümayəndələri yabanı halda səhralarda, yarımsəhralarda, bozqırlarda, meşələrdə, meşə kənarlarında, çəmənlərdə, xüsusilə dağ çəmən fitosenozlarının tərkibinə daxil olmaqla, özlərini fitosenozun komponenti, bəzi növləri isə edifikator olmaqla, fitosenozun formalaşmasında əsas iştirakçılarından sayılırlar. Taxıllar əksəriyyəti mal-qara tərəfindən istər yaş və istərsə də quru halında yaxşı yeyilir. Bu qrup bitkilərin yem əhəmiyyəti, kimyəvi tərkibləri tədqiqatçılar tərəfindən qismən öyrənilmişdir. Vitamin tərkibləri isə, xüsusilə "A" və "C" vitaminləri, yalnız bəzi mədəni yem bitkilərində öyrənilmişdir.[2]
Qırtıckimilər və ya taxıllar (lat. Poaceae) — qırtıcçiçəklilər sırasına aid bitki fəsiləsi.
Fəsilənin nümayəndələri birillik və ya çoxillik otlar, yerüstü hissələri bəzən odunlaşan, hətta ağacşəkilli bitkilərdir. Taxılkimilərin qədim ekoloji tiplərindən bambuk, qarğı, murquz kimi hündür gövdəli növləri də vardır. bu fəsiləyə daxil olan növlər saçaqlı, sürünən, kökümsov bitkilər olub, əksəriyyəti çim əmələ gətirəndir. Yarpaqları növbəli olmaqla iki cərgə üzrə düzülür, xətvarıdır, aşağısında buğum aralarını əhatə edən qın əmələ gətirir. Çiçəklər ikicinslidir, ziqomorfdur, mürəkkəb çiçək pulcuqları ilə əhatə olunaraq sünbülcüklərdə yerləşir. Çiçəkyanlığı iki dairədə yerləşmiş pərdəşəkilli pulcuqlardan təşkil olunmuşdur. Çiçəkyanlığının xarici dairəsində iki ədəd aşağı və yuxarı çiçək pulcuğu vardır. Erkəkcik 3 ədəddir. Yumurtalıq üst vəziyyətdədir. Bir yuvalı və bir intequmentli bir yumurtacığa malikdir. Ağızcıqlar oturaq olub, 2 ədəddir. Meyvə dən, nadir hallarda (bambuklarda) çəyirdək və ya giləmeyvədir. Toxum endospermlidir.
Bu qrup bitkiləri A.A.Qrossheym (1939), V.D.Aleksandrovna (1941), İ.V.Larin (1950), Y.M.İsayev və b. (1965), Ə.İ.Mayılov (1965), S.H.Musayev (1980), V.S.Xəlilov (1971) və s. tədqiqatçılar öyrənmişlər. Azərbaycan florasında taxılların 306 növü məlumdur. Taxıl fəsiləsinin nümayəndələri yabanı halda səhralarda, yarımsəhralarda, bozqırlarda, meşələrdə, meşə kənarlarında, çəmənlərdə, xüsusilə dağ çəmən fitosenozlarının tərkibinə daxil olmaqla, özlərini fitosenozun komponenti, bəzi növləri isə edifikator olmaqla, fitosenozun formalaşmasında əsas iştirakçılarından sayılırlar. Taxıllar əksəriyyəti mal-qara tərəfindən istər yaş və istərsə də quru halında yaxşı yeyilir. Bu qrup bitkilərin yem əhəmiyyəti, kimyəvi tərkibləri tədqiqatçılar tərəfindən qismən öyrənilmişdir. Vitamin tərkibləri isə, xüsusilə "A" və "C" vitaminləri, yalnız bəzi mədəni yem bitkilərində öyrənilmişdir.
Les poàcies o gramínies (Poaceae o Gramineae) són una família de plantes herbàcies o llenyoses de l'ordre de les Poales; constitueixen la branca arcaica de les angiospermes. Corresponen a la família de les gramínies en sistemes de classificació clàssics com el de Linné.
Tenen tiges cilíndriques anomenades palla si són herbàcies i canyes si són llenyoses, normalment amb nusos massissos i entrenusos balmats, així com fulles esparses, compostes típicament de beina, lígula i limbe. La beina envolta la tija, mentre que la lígula és un petit apèndix membranós, o em alguns casos pelut, situat a la zona d'unió del limbe amb la beina; el limbe sol ser allargat. Les flors són generalment hermafrodites; amb periant nul o, com a màxim, representat per dues o, de vegades, tres petites peces escamoses anomenades lodícules o glumèl·lules; androceu de tres estams; gineceu d'ovari unilocular i amb dos estigmes plomosos. Tancant la flor, hi ha dues bràctees anomenades glumel·les; la glumel·la inferior o lema està ben desenvolupada i sovint presenta una aresta; la glumel·la superior o pàlea és membranosa i de vegades molt reduïda. Una o comunament diverses flors constitueixen la inflorescència elemental de les gramínies: l'espícula o espigueta, que presenta dues bràctees a la base, anomenades glumes. Les espícules es reuneixen en panícules, raïms o espigues. Tenen els fruits en cariopsis. És una família cosmopolita, amb més de 700 gèneres i 12.000 espècies.
A aquesta família pertanyen tots els cereals, de gran importància en l'alimentació humana, les pastures, de gran importància en l'alimentació animal, les canyes i els bambús.
La família Poàcies inclou les dotze subfamílies següents.[1][2] S'hi indiquen també les tribus que hi pertanyen:[3]
Una tretzena subfamília, Micrairoideae, no gaudeix encara del suport general[1] i dues tribus no han estat assignades encara a cap subfamília (incertae sedis). Segons algunes fonts,[2] el nombre total de tribus i subtribus és de 40 i 76 respectivament, i de 45 tribus segons d'altres,[3] però sense concretar-ne el nombre de subtribus.
Pel que fa a les espècies, les poàcies es classifiquen en uns 700 gèneres que comprenen unes 11.000 espècies (dades de 2006).[4]
La següent llista de gèneres conté la majoria dels esmentats en la bibliografia botànica. En molts casos, alguns dels que hi ha llistats són sinònims. Els enllaços als sinònims porten al nom preferit del gènere.
Les poàcies o gramínies (Poaceae o Gramineae) són una família de plantes herbàcies o llenyoses de l'ordre de les Poales; constitueixen la branca arcaica de les angiospermes. Corresponen a la família de les gramínies en sistemes de classificació clàssics com el de Linné.
Tenen tiges cilíndriques anomenades palla si són herbàcies i canyes si són llenyoses, normalment amb nusos massissos i entrenusos balmats, així com fulles esparses, compostes típicament de beina, lígula i limbe. La beina envolta la tija, mentre que la lígula és un petit apèndix membranós, o em alguns casos pelut, situat a la zona d'unió del limbe amb la beina; el limbe sol ser allargat. Les flors són generalment hermafrodites; amb periant nul o, com a màxim, representat per dues o, de vegades, tres petites peces escamoses anomenades lodícules o glumèl·lules; androceu de tres estams; gineceu d'ovari unilocular i amb dos estigmes plomosos. Tancant la flor, hi ha dues bràctees anomenades glumel·les; la glumel·la inferior o lema està ben desenvolupada i sovint presenta una aresta; la glumel·la superior o pàlea és membranosa i de vegades molt reduïda. Una o comunament diverses flors constitueixen la inflorescència elemental de les gramínies: l'espícula o espigueta, que presenta dues bràctees a la base, anomenades glumes. Les espícules es reuneixen en panícules, raïms o espigues. Tenen els fruits en cariopsis. És una família cosmopolita, amb més de 700 gèneres i 12.000 espècies.
A aquesta família pertanyen tots els cereals, de gran importància en l'alimentació humana, les pastures, de gran importància en l'alimentació animal, les canyes i els bambús.
Teulu mawr o blanhigion blodeuol yw Poaceae neu Gramineae (teulu'r gwir weiriau/glaswelltau). Mae'n cynnwys tua 10,000 o rywogaethau mewn tua 675 genws a 12 is-deulu.[1][2] Mae ganddynt flodau bach wedi'u trefnu mewn "sbigolion" (spikelets). Mae'r blodau'n cael eu peillio gan y gwynt. Ceir y teulu ledled y byd ac mae glaswelltiroedd yn ffurfio tua 25% o lystyfiant y byd.[2] Mae'r teulu'n cynnwys y grawnfwydydd megis gwenith, indrawn a reis a chnydau eraill megis cansen siwgr.[2]
Teulu mawr o blanhigion blodeuol yw Poaceae neu Gramineae (teulu'r gwir weiriau/glaswelltau). Mae'n cynnwys tua 10,000 o rywogaethau mewn tua 675 genws a 12 is-deulu. Mae ganddynt flodau bach wedi'u trefnu mewn "sbigolion" (spikelets). Mae'r blodau'n cael eu peillio gan y gwynt. Ceir y teulu ledled y byd ac mae glaswelltiroedd yn ffurfio tua 25% o lystyfiant y byd. Mae'r teulu'n cynnwys y grawnfwydydd megis gwenith, indrawn a reis a chnydau eraill megis cansen siwgr.
Lipnicovité (Poaceae) je čeleď jednoděložných rostlin z řádu lipnicotvaré (Poales). Pro čeleď je povolen alternativní název Gramineae[1]. V češtině jsou obecně označovány jako trávy. Všechny pravé trávy patří do této čeledi, travám podobné rostliny z jiných čeledí, jako např. ostřice, lze označovat jako graminoidy, ale nejsou to pravé trávy.
Trávy jsou nejčastěji jednoleté, dvouleté či vytrvalé byliny, ale někdy to mohou být i netypické dřeviny, např. bambus, zřídka liány. Mohou být jen několik málo centimetrů vysoké až po více než 30 m u některých druhů bambusů. Mezi trávami je mnoho druhů výrazně xerofytických (suchomilných), rozšíření ve stepních až pouštních oblastech, je zde ale i mnoho druhů mezofilních, druhů lesů mírného pásma i tropů, ovšem na druhou stranu i hodně bahenních rostlin a rostlin vodních kořenících ve dně. Stébla jsou často dutá, v nodech bývají výrazná kolénka. Vytrvalé druhy mívají často oddenky, vzácně hlízy. Některé druhy jsou výrazně trsnaté, jiné rozvolněné s dlouhými výběžky.
Listy dosti často tvoří přízemní růžice, jsou jednoduché, střídavé, skoro vždy dvouřadě uspořádané, většinou přisedlé, řidčeji řapíkaté (nepravý řapík) s listovými pochvami. Čepele jsou celokrajné, nejčastěji čárkovité, někdy štětinovité či jehlovité, řidčeji až kopinaté nebo až obvejčité, žilnatina je převážně souběžná, zřídka dlanitá či zpeřená. Čepele mohou být normálně ploché, někdy jsou žlábkovitého tvaru či svinuté, zřídka oblé. Na rozhraní pochvy a čepele je skoro vždy přítomen jazýček různého tvaru či délky, který může být brvitý, chlupatý či lysý, roztřepený či celokrajný. Někdy může být redukovaný na věneček chlupů, např. rákos, nebo může být celkově zakrnělý, vzácně chybí.
Trávy jsou převážně jednodomé rostliny s oboupohlavnými květy, někdy mohou být květy jednopohlavné, v některých případech se mohou vytvářet jednopohlavné klásky, někdy se tvoří jednopohlavná květenství (např. kukuřice setá), řidčeji to jsou i dvoudomé rostliny.
Květy jsou v květenstvích. Pro čeleď je typickým základním květenstvím tzv. klásek, který může být jednokvětý (např. psineček) nebo vícekvětý, některé květy však mohou být sterilní. Klásek je v typickém případě podepřen 2 plevami, což je vlastně zvláštní typ listenu, které jsou buď přibližně stejné, nebo různé. Plevy jsou uspořádány střídavě (i když blízko sebe a mohou se zdát vstřícné), proto rozlišujeme dolní a horní plevou. Pleva může být osinatá či nikoliv. V některých případech je plev jiný počet, vzácně jsou zakrnělé nebo chybí (např. tajnička). Klásky většinou skládají další složená květenství, hlavně laty, hrozny, klasy, popř. stažené laty napodobující klas (licholas) (např. bojínek).
Okvětí je u trav velmi redukované a nenápadné. Dva vnější okvětní lístky jsou srostlé a přeměněny v plušku, ta je však někdy interpretována jako listencovitého původu. Vnitřní okvětní lístky jsou přeměněny v plenky (lodiculae), které jsou nejčastěji 2, vzácněji 3, někdy chybí. Květ je podepřen pluchou, což je útvar listenovitého původu. Plucha může být někdy osinatá, řidčeji může být osin i více. Tyčinky, gyneceum a plenky se nachází mezi pluchou a pluškou. Někdy může být pluška redukovaná nebo chybí.
Tyčinky jsou nejčastěji 3, méně často 1, 2 nebo 6, zřídka 4, u některých zástupců z podčeledi Bambusoideae dokonce mnoho (např. 120). Tyčinky jsou většinou volné, zřídka nitkami srostlé nebo srostlé s plenkami. Opylení probíhá skoro vždy větrem, (anemogamie). Gyneceum je srostlé ze 2, řidčeji 3 plodolistů, je synkarpní, semeník je svrchní. Blizny a čnělky jsou většinou 2, řidčeji 3 nebo 1.
Plodem je převážně obilka. Ta může být okoralá, za zralosti obalená pluchou a pluškou, nebo neokoralá (nahá). Obilka je v podstatě zvláštní typ nažky typický pro tuto čeleď. Zřídka zde může být i jiný typ plodu, např. bobule nebo oříšek.
Trávy jsou rozšířeny po celém světě. Snad s výjimkou zaledněných oblastí. Je známo asi 668 rodů a asi 10035 druhů[2] a patří tedy k nejpočetnějším čeledím krytosemenných rostlin.
Trávy nejsou příliš starou skupinou. Nejstarší doklady rostliny, která by mohla patřit k travám, jsou známy z křídy, asi před 100-110 milióny let, ale není to zcela jisté. Stáří genetického předchůdce dnešních trav (a obilnin) je odhadováno na 90 miliónů let.[3] Spory a další fosilní části travin z čeledi Poaceae byly objeveny u fosilní čelisti ornitopodního (hadrosauroidního) dinosaura z období spodní křídy, asi před 113 až 101 miliony let.[4][5] Koncem křídy snad už trávy začínaly nabývat svého dominantního postavení. Fosilní pozůstatky klásků trav jsou známy z třetihor, asi před 55 milióny let. Dnes to je na Zemi dominantní skupina, vegetace s dominancí trav zabírá v současnosti asi 20% zemské souše.[2] Asi před 8 milióny let došlo k rozšíření moderních trav C4 oproti dřívějším trávám C3.[6][7]
Travinami (místo listů a jiných „C3“ zdrojů) se předkové moderního člověka začali živit před 3,5 milióny let[8] (či dříve).[9] Jejich hospodářský význam pro člověka je naprosto zásadní (neolitická revoluce). Jednozrnná pšenice pochází pravděpodobně z Karacadağ[10] nedaleko Göbekli Tepe z doby před více než 10 tisíci lety.[11] Dvouzrnka byla nalezena v Nahal Oren. Mezi lipnicovité pak patří také všechny pravé obilniny, z nichž hlavní jsou: pšenice setá (Triticum aestivum), ječmen obecný (Hordeum vulgare, využívaný již 10 tisíc let v oblasti Úrodného půlměsíce)[12], žito seté (Secale cereale), oves setý (Avena sativa), rýže setá (Oryza sativa), kukuřice setá (Zea mays) a další. Z dalších užitkových rostlin je to cukrová třtina (Saccharum officinarum), bambus (bambusa) a další. Zásadní význam mají trávy také jako dominantní složka luk, pastvin, stepí aj.
Seznam a popis jednotlivých podčeledí, řazených do čeledi lipnicovité.[2][13]
Lipnicovité (Poaceae) je čeleď jednoděložných rostlin z řádu lipnicotvaré (Poales). Pro čeleď je povolen alternativní název Gramineae. V češtině jsou obecně označovány jako trávy. Všechny pravé trávy patří do této čeledi, travám podobné rostliny z jiných čeledí, jako např. ostřice, lze označovat jako graminoidy, ale nejsou to pravé trávy.
Græs-familien (Poaceae) er en plantefamilie i græs-ordenen. Med cirka 10.000 arter i mere end 650 slægter er den en af de største familier inden for blomsterplanterne. De er udbredt i alle verdens klimazoner.
Mange arter af græsser hører til de ældste nytteplanter og er siden tidernes morgen af livsvigtig betydning for mennesket. Alle kornsorter som hvede, rug, byg, havre, hirse, majs og ris medregnes til denne plantegruppe. Landskabsformer som enge, overdrev, stepper og savanner præger store dele af verdens landskaber.
Græs-familien omfatter både arter, der lever i kort og lang tid. De udviser en karakteristisk morfologi med en fælles "græsagtig" grundplan for de forskellige arter. Græsser har oftest en slank vækstform og har lange blade og ofte diskrete, enkle blomsterstande.
Mange arter er énårige og gennemlever hele deres livscyklus i en enkelt vækstsæson. De lever oftest kun få måneder og overlever den ugunstige årstid som frø i jorden. De såkaldte hemikryptofytter derimod har foryngelsesknopper nær jordoverfladen og overlever ugunstige perioder beskyttet af jord, løv eller sne. Dertil hører de toårige arter, der i løbet af sommeren eller efteråret danner kimplanter og først i det følgende år danner frø og frugter, ligesom flerårige arter, der lever få eller mange år. Disse besidder rosetter eller lignende, der er i stand til at overvintre. Individerne af en generation blandt længerelevende arter kan blive indtil 400 år gamle, f.eks. hos Rød Svingel.[1] Alm. Kvik er et eksempel på, at græsser kan formere sig ud fra fragmenter af udløbere.[2] De fleste arter er urteagtige; deres strå dør over jordoverfladen efter cirka et år. Undtagelser udgør træagtige Bambus-arter, hvis skud er tykvæggede og hårde og kan leve i flere årtier.
Nogle græsser er spinkelt bygget og bliver kun nogle få centimeter store (fx Enårig Rapgræs). Andre arter har forveddede strå og når højder indtil 40 meter eller mere, som eksempelvis bambus-arten Dendrocalamus giganteus. En- og toårige arter har sædvanligvis enkelte eller få skud i løse tuer med bløde blade. Hos disse græsser bærer alle eller de fleste stængler blomsterstande. De længerelevende arter danner i de fleste tilfælde fastere strå og bladplader og danner ud over blomstrende skud et større eller mindre antal ikke blomstrende skud. De vokser i løse eller tætte tuer eller plæneagtigt. Den sidstnævnte vækstform fremkommer, når planterne enten udbreder sig ved hjælp af mere eller mindre lange overjordiske, krybende, grønlige eller rødlige stængler, kaldet stoloner (fx Kryb-Hvene) eller ved hjælp af underjoriske, hvide eller brune rhizomer (fx Alm. Kvik). Ud over farven lader de to typer sig også adskille ved hjælp af knæene på udløberne med rødder: stoloner har på hvert knæ fuldstændige blade med bladskede, hvorimod rhizomer på disse steder blot har små, tynde skælformede lavblade. Hos tuedannende arter dannes kun meget korte udløbere, eller de unge sideskud udvikler sig indeni moderskuddets bladskeder, dette ses hos Fåre-Svingel. På denne måde opstår på grund af de tæt sammentrængte skud den typisk tueformede vækstform hos mange græsser. De fleste græsser har trævlerødder; de danner ingen hoved- eller pælerod. Ved stængelgrunden og ved udløbernes knæ dannes talrige rødder, der hver især kan udvikle siderødder af 1. og 2. orden. På denne måde kan der opstå rodsystemer med en betragtelig størrelse. Således kan en enkelt plante af Rød Svingel eksempelvis udbrede sig 250 meter i diameter.[1]
Græssernes stængler kaldes strå. De er oftest hule og trinde (runde i tværsnit). Kun få græsser har massive stængler. De er leddelt ved hjælp af faste, vævsfyldte knæ (nodier). Afsnittene mellem knæene kaldes internodier. Umiddelbart over knæene ligger vækstzonerne, stråene gror altså med indskudte meristemer (væv af små udifferentierede celler, der vedvarende deler sig). Stråene bliver på grund af disse vækstzoner bevægelige og smidige. De er på den måde i stand til atter at rette sig op efter påvirkning gennem vind eller regn.
Bladene hos græsserne består altid af to forskellige afsnit: bladskeden og bladpladen. Bladskeden er bladets grund og sidder fast på knæet og omslutter internodiet næsten indtil næste knæ på strået. Bladskederne er hos de fleste græsser åben i den ene side. Hos få græsarter er randene sammenvoksede og dermed bladskeden rørformet lukket, selvom de tidligt revner i den øvre del. Mens de basale bladskeder beskytter vækstpunkterne hos de unge skud, så opfylder stråenes bladskeder denne beskyttelsefunktion for vækstzonerne over knæene og sørger desuden for yderligere stabilitet.
Ved overgangen mellem bladskede og bladplade sidder hos de fleste arter et hindeagtig vedhæng, skedehinden. Den fremstår oftest som en farveløs, gennemskinnelig fortsættelse af hinden på indersiden af bladskeden og er en forlængelse af bladskedens indre epidermis. Den beskytter mod skader forårsaget af vindens gentagne bevægelser af strået samt mod indtrængen af snavs og parasitter i rummet mellem strå og bladskede. På grund af sit forskelligartede udseende er skedehinden nyttig ved artsbestemmelsen. Den er behåret eller ubehåret, kraveformet, tilspidset, udstrakt, meget kort eller meget lang. Til tider er skedehinden erstattet af en hårrække, i sjældne tilfælde mangler den helt.
Græssernes blomsterstande er opbygget af småaks, der kan være samlet i et aks, en top eller en dusk. Et aks har ustilkede eller ganske kortstilkede, rækkestillede småaks. En top har langstilkede småaks, hvor der normalt udgår flere grene fra samme etage, især de nedre grene er forlængede. En dusk har ganske korte grene. Småaksene består oftest af en eller flere tvekønnede blomster.[3] Såkaldte kolber er en variant af aks, hvor aksvævet er forstørret. Sjældnere består blomsterstanden af enkeltblomster.
Græsser udmærker sig ved karakteristisk reducerede blomster. Småaksene består først af et par yderavner, ydre og indre yderavne, der kan være sammenvoksede. Derover findes i småakset en eller flere blomster, der er omsluttet af såkaldte inderavner, nedre og øvre inderavne kaldet henholdsvis dækblad og forblad. Dækbladene kan opfattes som støtteblade for enkeltblomsterne. Avnerne varierer stærkt i deres form og størrelse. De to yderavner kan være ens eller forskellige af udseende. Dækbladet kan være spids eller stump i enden eller være tandet på forskellig vis. På rygsiden er dækbladet hvælvet, sammentrykt eller med køl. Dækbladets midtribbe kan være børsteformet forlænget i en såkaldt stak, der kan være ryg- eller spidsfæstet.
Blomsterne består af et forblad og to, sjældent tre, svulmeskæl, der undertiden har sammenvoksede rande. Det er svulmeskællene, der under blomstringen tvinger dækblad og forblad fra hinanden. I blomsten findes desuden oftest tre støvblade (sjældent seks, to eller kun et), der hver består af en tynd støvtråd og en todelt støvknap, der bærer pollenet. Endeligt findes der i hver blomst en rund, oversædig frugtknude, der i spidsen har to (sjældnere en eller tre) grifler med fjerformede støvfang.
Frugten er hos de fleste græsarter en tør skalfrugt, det vil sige en nød, hvor frugtknudens tynde væg og frøskallen er sammenvoksede. Sjældnere er frugterne bær eller stenfrugter med saftige eller kødede frugtvægge som det er tilfældet hos nogle bambus-arter.
Hos nogle græsser omsluttes nøden, kornet, mere eller mindre fast af avnerne. Skallen kan være helt sammenvokset med inderavnerne (som hos byg). Hos andre græsser er kornene nøgne, det vil sige at de ved modenhed ikke omsluttes af avnerne (som hos rug og hvede). Kornet har ofte en fure på langs, bugfuren. Inden i kornet ligger den stivelsesrige frøhvide og kimen. Det er græssernes korn, der på grund af deres rige indhold af stivelse, har givet disse planter deres store betydning som næringsplanter for mennesker.[4]
Cirka en femtedel af jordens plantedække indtages af græsser.[5] Savanner og stepper danner de største, naturlige græsområder på jorden i klimazoner, der ikke er egnet til skov. Dertil kommer de af mennesket skabte kulturlandskaber i især Centraleuropa. De er opstået i en lang proces efter istiden, hvor skove er blevet omdannet til enge og græsgange.
De varige, mere eller mindre tætte græsbestande gør gavn på mange måder. De forhindrer frem for alt ved hjælp af deres tætte rodsystem erosion af jord på grund af vind og vand. Desuden skaber de ved deres rodnet et højt indhold af organisk stof i jorden. Cirka en tredjedel af planternes primærproduktion forbliver tilbage under jorden i græsområder og fører dermed til humusdannelse. Dertil bidrager hvert år det døde overjordiske plantemateriale, der bliver tilbage som dækningsmateriale og kun langsomt nedbrydes. I flere græsområder spiller naturlige ildebrande en afgørende rolle. Lyn antænder ved afslutningen på vækstperioden den døde plantemasse. De uorganiske næringsstoffer i asken fremmer som gødning planternes nye vækst. Derudover bliver trævækst ødelagt. Brandene bidrager således til at holde græslandskabet åbent. Græsområder bebos af og ernærer en artrig og mangfoldig dyreverden af insekter (termitter og myrer), spindlere, fugle, småpattedyr og ikke mindst græssende flokke af store pattedyr som på de afrikanske savanner.[6]
De træløse stepper befinder sig i de halvtørre, tempererede zoner overvejende på den nordlige halvkugle. På den sydlige halvkugle svarer den argentinske pampas til de eurasiske stepper og de nordamerikanske prærier. Stepper er i årets løb underlagt to perioder med vegetatonshvile på grund af dels den strenge kulde om vinteren og dels den vedvarende tørke om sommeren. Nedbøren om foråret og i det sene efterår er tilstrækkelig for steppevegetationens vækst.[6][7]
Savannerne omfatter cirka 15 millioner kvadratkilometer. De opnår deres største udstrækning i den sydlige halvkugles troper med sommerregn. Analoge dannelser til savannen finder man i Venezuelas og Colombias Los Llanos, Brasiliens cerrado eller Nordaustraliens eukalyptus-stepper. Savanner har i modsætning til stepper og prærier træer og buske. Afrikas savanner ville uden brande og påvirkningen fra elefant- og hovdyrflokke på kort tid vokse til med trævækster. På savanner spiller udover de større dyr også termitter, myrer og græshopper en afgørende rolle som regulator i økosystemet. Planternes artsrigdom på savannen er forholdsvis lille. Især C4-græsser dominerer. Derimod er dyrenes artsrigdom yderst stor. Således lever cirka 1,5 millioner større dyr på jordens savanner, alene i Østafrikas Serengeti findes 98 store græssende dyr per kvadratkilometer.[6][7]
Kulturgræsland tjener mennesket økonomisk som grundlag for kvægavl. Enge og græsgange, der er domineret af græsser i det tempererede Europa er overvejende et resultat af et århundredlangt menneskeligt arbejde. Stadig for omkring 10.000 år siden var Centraleuropa et rent skovland. Udviklingen af bondekulturer, der udbredte sig fra Mellemøsten til Centraleuropa for cirka 6700 til 6400 år siden, muliggjorde at menneskene blev fastboende og førte til endnu stærkere indgreb i det naturlige plantedække. Der kom bebyggelser, de første agre og husdyr, der i første omgang søgte deres føde i skoven. Dyrenes fødesøgning, brand og skovhugst førte i løbet af en lang periode til en lysere skov. Med begyndelsen af jernalderen blev jordudnyttelsen forstærket, og med opfindelsen af leen blev indvinding af hø og halm mulig. På denne måde opstod de første større enge. I middelalderen blev landskabsudviklingen fuldendt til et åbent og meget differentieret kulturlandskab, hvor bebyggelser, agerland, levende hegn, krat og den tilbageværende skov vekslede med enge og græsgange.[6][8]
Græs-familien har et meget bredt spektrum af anvendelsesmuligheder. Det drejer sig dog kun om forholdsvis få slægter, der har betydning som nytteplanter for mennesker. Kun cirka 15 slægter (det er knap 2 procent, dog er bambus-slægter ikke medregnet) spiller en større rolle.[9]
Korn er af stor verdensøkonomisk betydning. Græsfrugter, det vil sige græskorn, tjener mennesker som basalt næringsmiddel. De leverer over 50 % af verdens fødevareforsyning. Hvede (Triticum-arter), majs (Zea mays) og ris (Oryza sativa) indtager en ledende rolle. Byg, rug, hirse og havre dækker cirka en tiendedel.
Hvede, byg og rug har deres oprindelse i den såkaldte frugtbare halvmåne, et område, der strækker sig fra Egypten over Palæstina til den Persiske Bugt. Her blev de vilde former udviklet gennem udvælgelses- og krydsningsprocesser til de nutidige kulturformer.
Ris har sin oprindelse i Kina eller Indien. Majs stammer fra Mexico. Græsser med små korn samles under betegnelsen hirse. De findes f.eks. i slægter som Digitaria, Echinochloa, Eragrostis, Panicum, Setaria, Sorghum.[9]
I 2012 blev følgende dyrkningsresultater opnået i verden (tabellen er sorteret efter udbytter):
Kornart Areal (ha) Udbytte per ha (ton) Samlet udbytte (ton) 1 Majs 176.991.927 4,94 875.098.631 2 Ris 163.463.010 4,39 718.345.380 3 Hvede 216.638.762 3,12 674.884.372 4 Byg 49.310.546 2,68 132.350.225 5 Hirse* 69.082.120 1,18 83.695.708 6 Havre 9.627.546 2,18 20.974.945 7 Rug 5.557.715 2,62 14.544.170 I alt 690.671.626 2.519.893.431 * Hirse omfatter her Sorghum og “Millet” (Alm. Hirse) Kilde: FAO: Faostat 2012[10]Ud over korn udnyttes stængler, blade og rødder. Græsser er en vigtig råstofkilde for stivelse, cellulose og sukker såvel som fedtstoffer og æteriske olier. De kan anvendes som råmaterialer, byggematerialer og fyldstoffer. Frem for alt bliver de forveddede strå af forskellige bambus-arter i Asiens tropiske og subtropiske områder anvendt til fremstilling af møbler, spiseredskaber, drikkekrus eller hegn, og ikke mindst anvendt i husbyggeri eller stilladser. I Nordvesteuropa bliver den her almindelige plante tagrør anvendt til tækning af huse.
Bambusskud bliver spist som grøntsag.
Citrongræsser (Cymbopogon) anvendes som krydderi eller lægeurter. Desuden tjener græsser til fremstilling af alkoholiske drikke som øl, rom eller snaps. Som selvfornyende råstoffer vinder græsser, især bambus og sukkerrør, tiltagende betydning indenfor fremstillingen af bioethanol som brændstof.
Hos følsomme mennesker kan pollen fra græsser udløse dannelsen af antistoffet immunoglobulin E (IgE), hvilket er kendt som høfeber. Såkaldte glykoproteiner, der er hæftet på pollenoverfladen, bliver let afgivet på slimhinderne og kan frembringe allergiske reaktioner. Desuden kan prolaminer i kornets såkaldte aleuronlag udløse sygdommen cøliaki på grund af en immunologisk overfølsomhedsreaktion.
Tidligere var 55 millioner år gammelt, fossilt græs fra Tennessee det ældste fund, man kendte af græs. 70 millioner år gammelt græspollen var imidlertid fundet i Egypten. Men i 2005 påviste undersøgelser af fossile ekskrementer efter dinosaurer bittesmå phytolitter (kiselceller), der kun findes i græs. Opdagelsen afslørede, at fem typer græs i slægt med moderne varianter fandtes i Gondwana-delen af det indiske subkontinent for 71-65 millioner år siden.[11]
Ordet "græs" er fælles for de germanske sprog og stammer fra det indoeuropæiske *ghre (= at gro, spire). Da det, som spirer og gror, har den samme farve, kommer "grønt" fra det samme ophavsord; det samme gælder "gran". Udtrykket "at bide i græsset", kommer fra tysk ins Gras beissen, mens det på engelsk hedder bite the dust. Udtrykket stammer fra den gamle iagttagelse, at sårede soldater bider i græs eller jord for at holde smerterne ud. Udtrykket findes allerede hos Homer, og på latin hed det humum ore mordere (= "i humus/jord med munden bide").[12]
Græsser er udbredt over hele verden. De forekommer fra havkysten til højfjeldet, fra ækvator til den anden side af polarkredsene i næsten alle terrestiske økosystemer og er altså udbredt vidt forskellige steder økologisk set. De vokser såvel på en konstant våd bund som på en ekstremt tør ligesom de både vokser meget varmt og i arktisk kulde.
Man finder græsser flydende i vandløb, bestanddannende i rørskove, som underbevoksning i skove, på både våd og tør bund, i vejkanter, på skråninger og klipper - selv skærver og murkroner koloniseres. Adskillige plantesamfund uden for skovene er hovedsageligt sammensat af græsser. De nordamerikanske prærier, Østeuropas stepper, Afrikas savanner og Sydamerikas pampas, men også landbrugets enge og græsgange i Europa, er naturlige såvel som af mennesket dannede landskabsprægende græsområder på jorden, hvor træer og buske er helt eller delvist manglende.
En liste over græsslægter med arter, der er vildtvoksende eller tilsyneladende vildtvoksende i Danmark:[13]
BEP-klade (Bambusoideae, Ehrhartoideae, Pooiedeae)
PACC-clade
Græs-familien (Poaceae) er en plantefamilie i græs-ordenen. Med cirka 10.000 arter i mere end 650 slægter er den en af de største familier inden for blomsterplanterne. De er udbredt i alle verdens klimazoner.
Mange arter af græsser hører til de ældste nytteplanter og er siden tidernes morgen af livsvigtig betydning for mennesket. Alle kornsorter som hvede, rug, byg, havre, hirse, majs og ris medregnes til denne plantegruppe. Landskabsformer som enge, overdrev, stepper og savanner præger store dele af verdens landskaber.
Die Süßgräser (Poaceae = Gramineae) sind eine Pflanzenfamilie in der Ordnung der Grasartigen (Poales). Mit etwa 12.000 Arten in rund 780 Gattungen[1] sind sie eine der größten Familien innerhalb der Blütenpflanzen. Sie sind weltweit in allen Klimazonen verbreitet und durch eine typische grasartige Gestalt gekennzeichnet.
Viele Arten der Süßgräser gehören zu den ältesten Nutzpflanzen und sind seit alters für den Menschen von lebenswichtiger Bedeutung. Alle Getreide wie Weizen, Roggen, Gerste, Hafer, Hirse, Mais und Reis zählen zu dieser Pflanzengruppe. Sie stellen in Form von Marktfrüchten oder als Viehfutter in der Veredelung heute die Basis für die Ernährung der Weltbevölkerung dar. Als Gras- oder Grünland wie Wiesen und Weiden, aber auch Steppen und Savannen prägen sie in weiten Teilen der Erde das Landschaftsbild.
Süßgräser umfassen sowohl kurzlebige als auch langlebige Arten. Sie weisen eine charakteristische Morphologie sowohl der vegetativen als auch der generativen Organe auf, mit einem gemeinsamen „grasförmigen“ Grundbauplan der verschiedenen Arten. Gräser sind meist schlankwüchsig und verfügen über lange, dünne, durch Knoten gegliederte Halme, parallelnervige, lange Blätter und oft unauffällige, einfache Blütenstände. Innerhalb der Unterfamilien, Tribus und Gattungen sind dagegen deutliche taxonspezifische Abwandlungen der Merkmale vorhanden.
Viele Arten sind einjährig und schließen ihren gesamten Lebenszyklus in einer Vegetationsperiode ab. Sie leben meist nur wenige Monate und überdauern die ungünstige Jahreszeit als Samen im Boden. Diese Formen, zu denen auch viele Getreidesorten gehören, werden Therophyten genannt.
Andere Arten sind mehrjährig und damit Hemikryptophyten. Sie verfügen über bodennahe Erneuerungsknospen und überdauern ungünstige Zeiten geschützt durch den Boden, Laubstreu oder Schnee. Dazu gehören zweijährige Arten, die im Laufe des Sommers oder Herbstes keimen und erst im folgenden Jahr Früchte und Samen bilden, ebenso wie ausdauernde und mehrjährige Arten, die wenige oder viele Jahre leben. Diese besitzen überwinterungsfähige Horste oder Rosetten. Die Individuen einer Generation ausdauernder Arten können bis zu 400 Jahre alt werden, so zum Beispiel der Rot-Schwingel (Festuca rubra).[2] Die Gemeine Quecke (Elymus repens) ist ein Beispiel dafür, dass sich Gräser aus Ausläuferfragmenten erneuern können (Rhizom-Geophyten).[3] Die meisten Hemikryptophyt-Arten sind krautig; deren Halme nach etwa einem Jahr Lebensdauer oberirdisch absterben. Ausnahmen bilden holzige Bambus-Arten (Bambuseae), deren Triebe dickwandig und fest sind und mehrere Jahrzehnte ausdauern können.
Etliche Süßgräser sind zart gebaut und werden nur wenige Zentimeter groß (z. B. Einjähriges Rispengras). Andere Arten haben verholzte Halme und erreichen Wuchshöhen bis zu 40 Metern und mehr, wie beispielsweise die Bambus-Art Dendrocalamus giganteus. Ein- und zweijährige Arten haben gewöhnlich einzelne oder wenige Triebe in lockeren Büscheln mit weicheren Blättern. Bei diesen Süßgräsern tragen alle oder die meisten der Sprossachsen Blütenstände. Die ausdauernden Arten bilden in den meisten Fällen festere Halme und Blattspreiten und neben blühenden Trieben eine größere oder kleinere Anzahl an nicht blühenden Trieben. Sie wachsen in lockeren oder dichten Horsten oder rasenförmig. Letztere Wuchsform ergibt sich, indem sich die Pflanzen entweder über mehr oder weniger lange, oberirdisch kriechende, grünliche oder rötliche Sprossachsen, namentlich Stolonen (z. B. das Weiße Straußgras) oder über unterirdische, weiße oder braune Rhizome ausbreiten (z. B. die Kriech-Quecke). Außer an der Farbe lassen sich die beiden Typen von sich an den Knoten bewurzelnden Ausläufern auch daran unterscheiden, dass Stolonen an jedem Knoten (Nodus) über vollständige Blätter mit Blattscheide und Blattspreite verfügen, Rhizome dagegen an diesen Punkten lediglich kleine, dünne, schuppenförmige Niederblätter entwickeln. Bei horstbildenden Arten bilden sich nur kurze Ausläufer, oder die jungen Seitentriebe entwickeln sich innerhalb der Blattscheiden des Muttertriebes (intravaginal), so beim Schaf-Schwingel (Festuca ovina). Auf diese Weise entsteht durch die gedrängt stehenden Triebe die typische büschelige, dicht horstige Wuchsform vieler Gräser. Wachsen die Triebe die untere Blattscheide durchstoßend (extravaginal), ist der Aufwuchs meist locker-horstig oder rasenförmig (z. B. Rot-Schwingel). Die meisten Süßgräser sind Flachwurzler; sie bilden keine Haupt- und Pfahlwurzeln. Am Stängelgrund und an den Knoten der Ausläufer werden zahlreiche sprossbürtige Wurzeln gebildet, die ihrerseits Seitenwurzeln 1. und 2. Ordnung entwickeln können. Auf diese Weise können Wurzelsysteme von beachtlicher Länge entstehen. So kann sich eine einzige Pflanze des Rot-Schwingels etwa 250 Meter im Durchmesser ausbreiten.[2]
Die Stängel der Süßgräser werden als Halme (über althochdeutsch halm ableitbar von germanisch halma, „Stroh, Getreidestängel, Grasstängel“[4]) bezeichnet. Sie sind meist hohl und rund. Nur wenige Grasarten besitzen markige Stängel. Sie sind durch feste, mit Gewebe gefüllte Knoten (Nodien) gegliedert. Die Abschnitte zwischen den Knoten werden als Internodien bezeichnet. Unmittelbar oberhalb der Knoten liegen die Wachstumszonen, die Halme wachsen also mit eingelagerten Meristemen. An diesen Stellen setzen die faserigen Verstärkungselemente, die den Halmen zusätzliche Stabilität und Zugfestigkeit verleihen, aus. Die Halme bleiben auf diese Weise beweglich und biegsam. Sie sind so in der Lage, sich nach Wind- und Regeneinwirkung wieder aufzurichten. Sie können entweder senkrecht hochwachsen, von einem gebogenen Grund aufsteigen oder gänzlich am Boden niederliegend wachsen. Grashalme variieren in Größe, Festigkeit und Zahl der Knoten. Sie sind meist im Querschnitt zylindrisch, selten etwas zusammengedrückt wie beim Zusammengedrückten Rispengras (Poa compressa). Bei einigen Süßgrasarten sind die untersten Internodien mehr oder weniger angeschwollen und verdickt. Die Halme etlicher Gräser sind unverzweigt, bei einigen Arten bilden sich von den Knospen in den Blattachseln ausgehende Seitenzweige. Die Beblätterung der Halme ist bei Süßgräsern immer wechselständig und fast ausnahmslos zweizeilig (distich) – im Gegensatz zur dreizeiligen Beblätterung der Sauergräser (Cyperaceae).
Die Blätter der Süßgräser bestehen immer aus zwei verschiedenen Abschnitten: der Blattscheide und der Blattspreite. Die Blattscheide entspricht dem Blattgrund, setzt am Knoten an und umschließt das Internodium bis fast zum nächsten Halmknoten. Die Scheiden sind bei der Mehrzahl der Gräser an einer Seite offen. Bei wenigen Grasarten sind die Ränder verwachsen und damit die Blattscheiden röhrig geschlossen, wenngleich sie früh im oberen Bereich aufreißen. Während die basalen Blattscheiden die Wachstumspunkte der jungen Triebe schützen, erfüllen diejenigen an den Halmen diese Schutzfunktion für die dortigen Wachstumszonen oberhalb der Knoten und sorgen außerdem für zusätzliche Stabilität. Der obere Teil der Blattscheiden kann bauchig aufgeblasen sein. Die Vorderseite des Blattscheidenendes kann in mehr oder weniger spitze, meist stängelumfassende „Öhrchen“ ausgezogen sein oder Büschel von Haaren tragen. Die Blattscheide geht am oberen Ende in die vom Halm abstehende Blattspreite über. Diese ist flach, gerollt oder gefaltet; stets länglich und mehr oder weniger spitz zulaufend. Sie zeigt die kennzeichnende Paralleladerung einkeimblättriger Pflanzen. Jede Blattader entspricht einem Leitbündel, der dem Stofftransport und der Aussteifung der Blattfläche dient.
Am plötzlichen Übergang von der Blattscheide zur Blattspreite sitzt bei den meisten Arten ein häutiges Anhängsel, das Blatthäutchen (Ligula). Es erscheint meistens als farbloser, durchscheinender Fortsatz der Oberhaut auf der Innenseite der Blattscheide und stellt eine Verlängerung der inneren Epidermis der Blattscheide dar. Es schützt vor Verletzungen durch Reibung des sich beim Wind hin und her bewegenden Halmgliedes sowie vor dem Eindringen von Schmutz und Parasiten in den Raum zwischen Halm und Scheide. Wegen seiner Gestaltungsvielfalt ist das Blatthäutchen für die Artbestimmung hilfreich. Es ist behaart oder unbehaart, kragenförmig, zugespitzt, langgezogen, sehr kurz oder sehr lang. Teilweise ist das Blatthäutchen durch eine Reihe von Haaren ersetzt, selten fehlt es ganz.
Die Blütenstände (Infloreszenzen) der Süßgräser bestehen aus einer Vielzahl von Teilblütenständen, seltener Einzelblüten, die in Ähren, Rispen und Trauben an einer Blütenstandsachse (Rhachis spicae) angeordnet sind. Die Teilblütenstände werden als Ährchen bezeichnet. Sie bestehen ihrerseits aus ein- bis mehreren, überwiegend zweigeschlechtigen Blüten. Sitzen die Ährchen ungestielt direkt an der Blütenstandsachse, handelt es sich um eine Ähre. Bei Fingergräsern befinden sich mehrere Ähren am Halmende in fingerartiger Anordnung. Sogenannte Kolben entstehen durch Abwandlungen von Ähren durch Vergrößerung des Achsengewebes. In Trauben befinden sich die Ährchen an unverzweigten Stielen. Die Ährchen können alle in die gleiche Richtung weisen (einseitswendig) oder sich in zwei Reihen an gegenüberliegenden Seiten der Achse befinden. Sind die Seitenäste einseits- oder allseitswendig verzweigt, handelt es sich um Rispen. In Ährenrispen oder Scheinähren sind die Seitenäste so kurz, dass die Blütenstände äußerlich wie Ähren erscheinen. Erst beim Umbiegen einer solchen Ährenrispe werden die tatsächlichen Verzweigungsmuster erkennbar.
Süßgräser zeichnen sich durch eine charakteristische Reduzierung der Blüten aus. Die Ährchen werden am Grunde von einer inneren und einer äußeren Hüllspelze (Gluma), die miteinander verwachsen sein können, eingefasst. Oberhalb davon stehen ein oder mehrere Blüten, jede mit einer Deck- sowie Vorspelze. Die Deckspelzen können als Tragblätter der Einzelblüten aufgefasst werden. Die Spelzen variieren in ihrer Form und Größe sehr. Die beiden Hüllspelzen können gleich oder verschieden gestaltet sein. Die Deckspelzen sind vielförmiger gestaltet. Sie können an den Enden spitz, stumpf oder verschiedenartig gezähnt sein. Auf dem Rücken sind sie gerundet, zusammengedrückt oder gekielt. Die Mittelrippe kann in einen Stachel oder eine Granne verlängert sein.
Die Blüten bestehen aus einer Vorspelze und zwei, selten drei, zuweilen an den Rändern verwachsenen Schwellkörperchen (Lodiculae), durch deren Anschwellen die Spelzen geöffnet werden. Es sind ferner meist drei Staubblätter (Stamina) vorhanden (selten sechs, zwei oder nur eines), von denen jedes einen Stiel (Filament) und einen den Pollen tragenden, zweiteiligen Staubbeutel (Anthere) aufweist. In jeder Blüte gibt es schließlich einen runden, aus zwei oder drei Fruchtblättern verwachsenen, oberständigen Fruchtknoten (Ovarium). Dieser verfügt an seiner Spitze über einen Stempel (Pistillum), der seinerseits auf kurzen Stielen ein, zwei oder selten drei fedrige Narbenäste (Stigmae) trägt. Der Fruchtknoten enthält die Samenanlage, die mit Fruchtknotenwänden zu einer Einheit, der Karyopse, verwächst.
Bei manchen Arten enthalten einige Blüten nur männliche Organe oder sind steril. Ferner sind etliche Arten verschiedenährig, das heißt, die Blüten mit nur weiblichen und nur männlichen Organen befinden sich getrennt in verschiedenen Blütenständen desselben Individuums (einhäusig), so beim Mais. Bei anderen Arten wie dem Pampasgras befinden sich die Geschlechter getrennt in den Blütenständen verschiedener Individuen einer Grasart. Sie sind zweihäusig.
Die Frucht ist bei den meisten Grasarten eine trockene Karyopse, eine Sonderform der Nussfrucht. Seltener sind die Früchte Beeren oder Steinfrüchte mit saftigen oder fleischigen Fruchtwänden, so wie bei einigen Bambus-Arten. Während der Reifezeit verwächst die Fruchtwand (Perikarp) mit der Samenschale (Testa) zu einer einsamigen, trockenen Schließfrucht. Die „Samenkörner“ stellen also keine Samen, sondern vielmehr Früchte dar. Unterhalb der Fruchtwand und der Samenschale liegt die eiweißreiche Aleuronschicht. Darunter folgt das den restlichen Samen ausfüllende stärkereiche Nährgewebe, das Endosperm. Gräser sind einkeimblättrig (monokotyl); bei ihnen ist das eine Keimblatt (Kotyledon) zu einem Scutellum (Schildchen) und zu einer Keimscheide (Koleoptile) umgestaltet. Das Scutellum liegt zwischen dem Endosperm und dem Embryo und spielt eine wichtige Rolle für den Stofftransport und die Hormonsynthese. Der Embryo verfügt bereits über deutlich erkennbare Wurzel- und Sprossanlagen. Die Koleoptile ist ein zylinderförmiges Schutzorgan, welches das Primärblatt des auskeimenden Embryos umgibt. Da die Koleoptile ein umgewandeltes Keimblatt darstellt, ist es als Organ ein Blatt. Wie alle Blätter besitzt es zwei Epidermen (außen und innen), Stomata und Leitbündel. Die Stärke und die Proteine dienen dem Embryo als Starthilfe für die Keimung, bevor es sich durch Photosynthese selbst versorgen kann. Die ausgereiften Früchte der Gräser sind in ihrer Gestalt und ihrem Aufbau charakteristisch. Die ehemalige Bauchnaht des Fruchtknotens erscheint auf einer Flanke des Korns als tiefe Furche.
An verschiedenen Stellen des Fruchtstandes bilden sich Zonen eines speziellen Gewebes, entlang dessen ein glatter Bruch entsteht, sobald der Samen reif ist. Bei den meisten Gräsern erfolgt dieser Bruch in der Ährchenachse unterhalb der Deckspelze. Die Karyopse ist in diesen Fällen meistens in Deckspelzen und Vorspelzen fest eingeschlossen und stellt als Gesamtheit die Ausbreitungseinheit (Diaspore) dar. Bei einigen Arten erfolgt der Bruch unterhalb der untersten Deckspelze des Ährchens (z. B. Perlgräser), unter dem einzelnen Ährchen oder in einem Büschel von Ährchen (Gerste), selten in der Hauptachse des Fruchtstandes (Dünnschwanz). Gräser mit nackten Früchten sind in den tropischen Gattungen Sporobolus und Eragrostis häufig. Bei diesen steht das Korn frei und wird ausgestreut, nachdem sich ein Bruch am Grunde der sie haltenden Deckspelze entwickelt hat.
Die Samen sind reich an Stärke. Diese kann aus einzelnen Stärkekörnern (Roggen, Weizen, Gerste) bestehen oder aus zu mehreren zusammengesetzten (Hafer). Auch in den Rhizomen und anderen vegetativen Organen speichern die Gräser Stärke, Saccharose und/oder Fructane. Bei den Fructanen kommt neben dem unverzweigten „Inulin-Typ“ der verzweigte „Phlein-Typ“ vor. Das Fructanmuster ist wie der Polymerisationsgrad oft kennzeichnend für die Art. Die äußere Endospermschicht (Aleuronschicht) der Karyopsen ist reich an Reserveproteinen. Sie enthält vor allem Albumine, Globuline, Gluteline (nur in verdünnten Säuren und Laugen löslich) und Prolamine (in 70–80%igem Ethanol löslich). Letztere sind beim Roggen- oder Weizenmehl Voraussetzung für die Backfähigkeit.
Einige Triben der Panicoideae bilden ätherische Öle in schlauchförmigen, verkorkten Zellen. Cymbopogon nardus liefert das Aetheroleum Citronella, das hauptsächlich aus Citronellal und Geraniol besteht und bei der Herstellung von Melissengeist oft das echte Melissenöl ersetzt. Weitere Cymbopogon-Arten werden angebaut, da sie Parfümöle wie Palmarosaöl und Lemongrasöl liefern. Diese Öle bestehen überwiegend aus Mono- und Sesqui-Terpenen, während Phenylpropanoide selten sind. Alkaloide sind selten. Es gibt Protoalkaloide und vereinzelt Pyrrolizidin- und β-Carbolintyp-Alkaloide. Cyanogene Glykoside (blausäure-produzierende Verbindungen) sind weit verbreitet, kommen aber immer nur in geringen Mengen vor. Cumarine kommen wahrscheinlich bei allen Vertretern vor, aber nur beim Gewöhnlichen Ruchgras (Anthoxanthum odoratum) und beim Duftenden Mariengras (Hierochloe odorata) in größeren Mengen. Polyphenole sind in geringeren Mengen enthalten.
In den Blattepidermen wird wie bei den Sauergräsern (Cyperaceae) häufig Kieselsäure in Form von Kieselsäurekörpern eingelagert. Oxalatkristalle scheinen vollkommen zu fehlen.
Etliche dieser Inhaltsstoffe zeigen als Bitterstoffe eine fraßhemmende Wirkung oder wirken toxisch auf Bakterien oder Pilze.
Bei ausdauernden Arten erfolgt die vegetative Ausbreitung überwiegend über Stolonen und Rhizome, die sich an den Knoten bewurzeln. Etliche Arten bedienen sich zusätzlich der unechten Viviparie, bei der keine Samen gebildet, sondern Brutknospen (Bulbillen), die erbgleiche Tochterpflanzen hervorbringen. Ein bekanntes Beispiel ist das Alpen-Rispengras (Poa alpina). Bei diesem Gras entwickeln sich im Blütenstand anstelle von Blüten grüne Pflänzchen, die an der Mutterpflanze verbleiben oder zu Boden fallen und als Diasporen dienen. Beim Zwiebel-Rispengras (Poa bulbosa) bilden sich basale, zwiebelartige Brutknospen, in denen Reservestoffe eingelagert sind. Jede Brutknospe bildet die Grundlage für eine neue Pflanze.
Gräser sind zur raschen Regeneration nach Verbiss oder Mahd befähigt. Dies liegt in der geschützten Lage ihrer Blattwachstumszonen (Meristeme) und Nebentriebknospen begründet. Die Wachstumszonen befinden sich an der Basis der Graspflanzen nahe der Erdoberfläche. Die Triebe bestehen aus unterschiedlich alten und gegenständig angeordneten Blättern. Junge Blätter wachsen an der Basis der Blattscheide (Interkalarmeristem). Ein erneutes Wachstum der Blätter nach Verlust durch Mahd oder Beweidung wird dadurch ermöglicht. Auch die einzelnen Blätter verfügen wie die Halme am oberen Ende der Blattscheiden im Übergang zu den Blattspreiten über teilungsfähiges Gewebe, das Nebentriebe bilden kann. Ferner sind die Halme durch das unterschiedliche Wachstum der teilungsfähigen Zonen oberhalb der Knoten zu einem Wiederaufrichten des Stängels nach Regen oder Tritt befähigt.
Die beschriebenen Wachstumsbereiche sind in verschiedene Zonen unterteilt: An der Basis findet die Zellteilung und damit eine Zellproduktion statt. Darauf folgt ein Bereich der Zellstreckung. In der folgenden Zone der Zelldifferenzierung erfolgt die Ausbildung der Blattzellen. Die Zellproduktion und Zellstreckung verschieben das ausdifferenzierte Blatt nach oben. Sobald das Blatt aus der Blattscheide ans Licht tritt, ist es photosynthetisch aktiv.[5]
Alle Süßgräser sind windblütig (anemogam). Die Blüten öffnen sich nur wenige Stunden am Tag, um Staubblätter und Narben dem Wind auszusetzen. Eine Selbstbestäubung wird durch die meist frühere Reife der Staubblätter verhindert (Proterandrie). Die starke Reduzierung der Blüten ist eine Anpassung an diese Form der Bestäubung. Gräser können auf auffällige Formen und Farben der Blüten und auf ein Nektarangebot zur Anlockung von Tieren verzichten. Die passive Pollenübertragung über den Wind und Luftströmungen erfolgt dabei weit weniger gezielt als bei der Tierbestäubung. Diesen Mangel gleichen die Windblüher mit der Massenproduktion von Blütenstaub aus. Dies führt während der Blütezeit zu regelrechten Staubwolken, die garantieren, dass zumindest ein kleiner Teil des weniger als einen Tag lebensfähigen Pollens seinen Bestimmungsort, die weiblichen Narben, erreicht. Beispielsweise bildet der Roggen (Secale cereale) pro Ähre etwa vier Millionen Pollenkörner; eine einzelne Blüte bis zu 57.000.[6] Eine große Blütenhülle wäre bei der Pollenverbreitung nur hinderlich. Die Lodiculae schwellen durch Wasseraufnahme an und drängen die Spelzen auseinander – die „Grasblüte“ öffnet sich. Die Filamente sind lang und dünn und lassen die Staubbeutel frei aus der Blüte heraushängen. So kann der Wind ungehindert den trockenen, nicht verklebten und leichten Pollen heraustragen. Die Fruchtknoten haben gefiederte und dadurch mit großer Oberfläche versehene Narben, die den Pollen gewissermaßen wieder aus Luft herauskämmen können. Die Effizienz dieser Form der Pollenverbreitung wird durch das Herausheben der Blütenstände über die Ebene des Blattwerkes sowie durch eine hohe Individuendichte der Graspflanzen verstärkt. Eine Sonderform der geschlechtlichen Ausbreitung ist die echte Viviparie, bei der die Samen schon auf der Mutterpflanze auskeimen. Die Samenausbreitung erfolgt auf vielfältige Weise; überwiegend durch den Wind (Anemochorie), über das Wasser (Hydrochorie) oder durch Tiere (Zoochorie).
Das Wurzelsystem der Wiesengräser bildet arbuskuläre Mykorrhiza (AM), eine Symbiose mit einem Pilz. Diese erleichtert der Graspflanze die Erschließung und Aufnahme von Nährstoffen aus dem Boden. Ein Pilzmycel verbindet mehrere Pflanzen derselben Art und andere Pflanzenarten, wodurch nicht nur das Gras selbst und der Pilz, sondern schließlich Wiese und Pilz eine Lebensgemeinschaft bilden.[7]
Unter den Gräsern gibt es sowohl C3- (die meisten heimischen Gräser wie Deutsches Weidelgras) als auch C4-Pflanzen (z. B. Mais, Hirse und Zuckerrohr), letztere mit effizienterer Photosynthese bei hohem Wärme- und Lichtangebot. C3-Pflanzen weisen dagegen bei kühleren Temperaturen und weniger Licht eine effizientere Photosynthese auf. Die Forschung hat gezeigt, dass der C4-Mechanismus zuerst bei den Gramineen, wahrscheinlich im Oligozän vor etwa 23 bis 34 Millionen Jahren entwickelt wurde, wobei es Hinweise auf über zehn unabhängige Entwicklungen gibt. Bei der geographischen Verbreitung ergibt sich eine auffällige klimatische Abhängigkeit der Photosynthesetypen. So ist der Anteil der C4-Pflanzen unter den Gräsern in kühlen und humiden Klimaten deutlich niedriger als in trockenen bis extrem ariden Regionen der Erde.[7]
Etwa ein Fünftel der Pflanzendecke der Erde wird von Gräsern eingenommen.[8] Savannen und Steppen bilden die großen, natürlichen Grasländer der Erde in Klimazonen, die für Wald nicht geeignet sind. Dem gegenüber stehen die durch menschliche Tätigkeit entstandenen Kulturgrasländer vor allem Mitteleuropas, die in einem langen nacheiszeitlichen Prozess vom Wald zur offenen, durch Wiesen und Weiden geprägten Landschaft entstanden.
Die dauerhaften, mehr oder weniger geschlossenen Grasbestände erfüllen vielfältige ökologische und biologische Aufgaben. Sie verhindern vor allem durch ihr dichtes und eng vernetztes Wurzelsystem die Abtragung der Bodenschicht durch Wind und Wasser (Erosion). Ferner erzeugen sie durch ihr Wurzelwerk einen hohen Gehalt an organischer Substanz im Boden. Etwa zwei Drittel der pflanzlichen Primärproduktion bleiben in Grasländern unterirdisch zurück und führen so zur Humusbildung. Dazu trägt das jährlich absterbende oberirdische Pflanzenmaterial zusätzlich bei, das als Mulch zurückbleibt und nur langsam zersetzt wird. In vielen Grasländern spielen natürliche Feuer eine Rolle. Blitze entzünden am Ende der Vegetationszeit die abgestorbene Pflanzenmasse. Die in der Asche enthaltenen anorganischen Nährstoffe fördern als Dünger den Neuaustrieb der Pflanzen. Darüber hinaus wird der Gehölzaufwuchs zerstört, die Brände tragen so zur Offenhaltung der Graslandschaft bei. Grasländer beherbergen und ernähren eine artenreiche und vielfältige Tierwelt: eine Vielzahl von Insekten (Termiten und Ameisen), Spinnen, Vögel, Kleinsäuger und zahlreiche im Boden lebende Tiere, nicht zuletzt Großsäuger wie jene der großen Tierherden in den afrikanischen Savannen. Letztere tragen wie die regelmäßigen Feuer dazu bei, die Verjüngung der Gehölze zu hemmen. Ihre Exkremente düngen den Boden. Durch den Fraß wird die Regeneration der Gräser so angeregt, dass die Primärproduktion um mehr als zwei Drittel zunimmt. Ferner sind sie für die Verbreitung der Früchte und Samen durch Epi-, Endo- oder Dysochorie von Bedeutung. Naturnahe Grasländer gehen heute weltweit zurück. Die Ursachen liegen in der Umwandlung in Acker- und Siedlungsland, der Aufgabe traditioneller Wiesen- oder Weidenutzungen sowie in der Intensivierung (Düngung) und Degradierung (Überweidung).[9]
Die baumfreien Steppen finden sich in den semiariden, gemäßigten Zonen vorwiegend auf der Nordhalbkugel. Auf der Südhalbkugel ist die argentinische Pampa eine zu den eurasischen Steppen und den nordamerikanischen Prärien analoge Vegetationsform. Es wird kontrovers diskutiert, ob sie auf natürliche Weise entstanden ist. Steppen unterliegen durch strenge Kälte im Winter und anhaltende Trockenheit im Sommer im Jahresverlauf zwei Perioden der Vegetationsruhe. Die im Frühjahr, Frühsommer und Spätherbst anfallenden Niederschläge reichen für das Wachstum der Steppenvegetation aus. Kennzeichnend für Steppen ist ihre hohe bodenbiologische Aktivität bei einem hohen Humusanteil (bis zu 10 %). Es können sich fruchtbare Schwarzerdeböden mit Humushorizonten bis zu einem Meter Mächtigkeit bilden. Die osteuropäischen Steppen lassen sich grob in etwa vier Vegetationstypen gliedern, die der zunehmenden Kontinentalität in Richtung Südost folgen. In Russland und der Ukraine finden sich Wiesensteppen mit Aufrechter Trespe (Bromus erectus), Flaumigem Wiesenhafer (Avenula pubescens), Schillergräsern (Koeleria) und vielen anderen Grasarten. Sie sind reich an einjährigen, und nicht grasartigen ausdauernde krautige Pflanzen. Darauf folgt die durch Pfriemengräser der Gattung Stipa dominierte Federgrassteppe mit schmalblättrigen „Horstgräsern“ und weniger Stauden, und schließlich die Kurzgrassteppe mit xerophytischen Festuca-Arten. Westliche Vorposten der osteuropäischen Steppen finden sich beispielsweise in der Pannonischen Tiefebene Ungarns. Im gemäßigten Nordamerika entspricht den Steppen die flächenmäßig deutlich kleinere Prärie zwischen Mississippi und den Rocky Mountains. Sie ist im Gegensatz zu den osteuropäischen Steppen weniger kontinental geprägt. Sie erreicht in West-Ost-Ausdehnung 1000 Kilometer, in Nord-Süd-Ausdehnung 2750 Kilometer. Im Osten findet sich die Hochgras-Prärie mit Wiesen-Rispengras (Poa pratensis), dem Pyramiden-Schillergras (Koeleria pyramidata), Prärie-Bartgras (Andropogon scoparius), Rutenhirse (Panicum virgatum) und vielen krautigen Pflanzen. Im Südteil folgt Richtung Westen die Mischgras-Prärie im Übergang zur am Fuße der Rocky-Mountains befindlichen Kurzgras-Prärie mit dem Moskitogras (Bouteloua gracilis) und Buchloe dactyloides. Die Bedeutung geschlossener Grasdecken als Schutz vor Erosion zeigen die verheerenden Sandstürme der 1930er Jahre in der danach benannten „Dust Bowl“ Nordamerikas durch großflächige Bodenzerstörungen im Zuge der Umwandlung in Ackerland. Noch im 17. und 18. Jahrhundert zogen Büffelherden mit einer geschätzten Bestandsgröße von 50 bis 70 Millionen Tieren über die Prärien. Heute sind es über 100 Millionen Hausrinder.[9][10]
Die Savannen umfassen etwa 15 Millionen Quadratkilometer. In den wechselfeuchten Sommerregengebieten der Tropen der Südhalbkugel erreichen sie ihre größte Ausdehnung in Afrika. Analoge Formationen der Savannen sind die Llanos Venezuelas und Kolumbiens, die Cerrados Brasiliens sowie die Eukalyptus-Steppen Nordaustraliens. Savannen sind im Gegensatz zu den Steppen und Prärien mit Bäumen und Sträuchern durchsetzt. Die Savannen Afrikas würden ohne Feuer und den Einfluss der großen Elefanten- und Huftierherden in kurzer Zeit mit Gehölzen zuwachsen. In den Savannen findet sich häufig ein kleinräumiges Oberflächenrelief aus flachen Hügeln und Senken mit Höhenunterschieden unter einem Meter. Dadurch unterscheiden sich die Standorte vor allem hinsichtlich der Wasserverfügbarkeit. Die unterschiedliche Wasserverfügbarkeit bestimmt schließlich die Nährstoffverfügbarkeit und die Vegetation. In Savannen spielen neben den Großtieren Termiten, Ameisen und Heuschrecken eine maßgebliche Rolle als Regulative im Ökosystem. Der Artenreichtum der Pflanzen der Savannen ist vergleichsweise gering. Die Hauptkomponenten sind C4-Gräser wie die Lampenputzergräser (Pennisetum) und Andropogon-Arten in Afrika. In Australien sind die Savannen, das sogenannte Spinifex- oder Hummock-Grasland, durch Igelkopfgräser der Gattungen Triodia und Plectrachne gekennzeichnet. Dagegen ist der Artenreichtum der Tiere ausgesprochen groß. So leben etwa 1,5 Millionen Großtiere in den Savannen der Erde, allein in der Serengeti Ostafrikas sind es 98 große Weidetiere pro Quadratkilometer. Die Zoomasse wird auf 150 bis 250 Kilogramm Trockenmasse pro Hektar geschätzt – jene der Wälder der gemäßigten Zone wird mit nur 10 Kilogramm Trockenmasse pro Hektar angegeben.[9][10]
In der Naturlandschaft Europas ist natürliches Grasland auf wenige Bereiche beschränkt. Nur in hohen Berglagen oberhalb der Waldgrenze, in Seemarschen, in oft überschwemmten Auenbereichen und im Randbereich von Hochmooren konnten sich kleinräumig natürliche, weitgehend baumfreie Grasländer, sogenannte Urwiesen, entwickeln. Sie sind heute stark vom Menschen überprägt. Kulturgrasland dient dem Menschen wirtschaftlich als Grundlage der Viehzucht. Die durch Gräser dominierten Wiesen- und Weidelandschaften des gemäßigten Europa sind im Wesentlichen das Ergebnis jahrhundertelangen menschlichen Wirkens. Noch vor etwa 10.000 Jahren war Mitteleuropa nahezu reines Waldland. Die Entwicklung bäuerlicher Kulturen, die sich vom Nahen Osten ausgehend vor etwa 6700 bis 6400 Jahren (Neolithikum) nach Mitteleuropa ausbreiteten, ermöglichte das Sesshaftwerden der Menschen und führte zu immer stärkeren Eingriffen in die natürliche Pflanzendecke. Es gab Siedlungen, erste Äcker und Nutztiere, die ihre erste Nahrung im Wald suchten. Der Fraß der Tiere, Brand und Holzeinschlag führten im Laufe längerer Zeit zu Auflichtungen in den Wäldern. Mit Beginn der Eisenzeit wurde die Landnutzung verstärkt, und mit Erfindung der Sense wurde die Gewinnung von Heu und Streu möglich. Auf diese Weise entstanden erste größere Wiesenareale. Im Mittelalter vollendete sich die Landschaftsentwicklung in einer offenen und differenzierten Kulturlandschaft aus Siedlungen, Waldresten, Feldgehölzen, Gebüschen, Äckern sowie artenreichen Wiesen und Weiden. Die typische Landschaft wird in der Literatur vielfach als „Parklandschaft“ oder „europäische Savanne“ beschrieben. Im Zuge der Agrarentwicklung in der Neuzeit wird die Landwirtschaft durch die Technisierung, Flurbereinigungen, Melioration sowie gezielte Ansaat ausgewählter Grasarten immer unabhängiger von natürlichen Gegebenheiten. Die artenreichen, extensiven Wiesen und Weiden wurden weitgehend von artenarmen, monotonen Wirtschaftswiesen und -weiden abgelöst. Zu den wichtigsten angebauten Futtergräsern des Dauergrünlandes gehören heute das Deutsche Weidelgras (Lolium perenne), das Wiesen-Lieschgras (Phleum pratense), das Gewöhnliche Knäuelgras (Dactylis glomerata) sowie Wiesen- und Rohr-Schwingel (Festuca pratensis, Festuca arundinacea).[9][11]
Etliche Gräser verhindern durch ihr dünnes Netzwerk aus Wurzeln und die Bedeckung des Bodens mit ihren oberirdischen Pflanzenteilen nicht nur dessen Abtragung durch Wind und Wasser, sondern sorgen zusätzlich für dessen Aufhöhung. Dafür sind besonders solche Arten geeignet, die unter vergleichsweise ungünstigen Standortbedingungen weitreichende Rhizome und Stolonen bilden können. So ist beispielsweise der Gewöhnliche Strandhafer (Ammophila arenaria) maßgeblich an der Festlegung der Treibsande sowie am Aufbau der Weißdünen auf den Inseln und an den Festlandsküsten beteiligt und erfüllt so eine wichtige Funktion im Küstenschutz. Auf regelmäßig überfluteten Schlickflächen der Küsten ist es der Strand-Salzschwaden (Puccinellia maritima), der mit seinen sich bewurzelnden Stolonen allmählich die kurzen dichten Rasen der Salzwiesen bildet und mit seinen kurzen steifen Halmen und Blättern den Schlamm gewissermaßen einfängt und die Oberfläche der Marsch langsam aufhöht. Dort, wo andere Süßgräser nicht mehr gedeihen können, übernimmt das Salz-Schlickgras (Spartina anglica) an ähnlichen Standorten die Funktion der Festlegung und Aufhöhung von Schlick der seewärtigen Seiten im Wattenmeer und entlang der Priele. Die Deiche der Küsten und Ströme werden schließlich mit einer Pflanzendecke ausgestattet, die von bodenhaltenden Süßgräsern dominiert wird.
In den Ebenen entlang der Flussufer des Binnenlands schützen Gräser den Boden vor Erosion und erfüllen eine ebenso wichtige Aufgabe im Hochwasserschutz. Beispielsweise bilden das Rohr-Glanzgras (Phalaris arundinacea) und der Wasser-Schwaden (Glyceria maxima) auf Schlammflächen und an Ufern dichte und hohe Aufwüchse mit kräftigen Rhizomen. Flussauen stellen nicht nur Retentionsflächen für Hochwässer dar, sondern sind aufgrund der Großgräser eine Senke („Falle“) für Sedimente, Nähr- und Schadstoffe.
Die Familie der Süßgräser bietet ein breites Spektrum an Nutzungsmöglichkeiten. Demgegenüber stehen jedoch nur relativ wenige Gattungen, die schließlich als Nutzpflanzen für den Menschen von Bedeutung sind. So werden lediglich 6 bis 7 Prozent der 600 bis 700 Gattungen als Nahrung oder als Werk- und Baustoffe verwendet. Nur etwa 15 Gattungen, das sind knapp 2 Prozent (ohne Berücksichtigung der Bambus-Gattungen), spielen dabei eine größere Rolle.[12]
Von großer weltwirtschaftlicher Bedeutung sind die Getreide. Grasfrüchte, beziehungsweise Getreidekörner, dienen dem Menschen als Grundnahrungsmittel. Sie liefern über 50 % der Welternährungsenergie. Weizen (Triticum-Arten), Mais (Zea mays) und Reis (Oryza sativa) nehmen dabei eine führende Rolle ein. Gerste, Roggen, Hirsen und Hafer decken etwa ein Zehntel ab. Weizen, Gerste und Roggen haben ihren Ursprung im sogenannten Fruchtbaren Halbmond, der sich von Ägypten über Palästina bis zum Persischen Golf erstreckt. Hier wurden die Wildformen in Kultur genommen, die über verschiedene Auslese- und Kreuzungsprozesse zu den heutigen Kulturformen entwickelt wurden. Reis hat seinen Ursprung in China oder Indien; Mais stammt aus Mexiko. Unter Hirsen werden Gräser verschiedener Gattungen mit kleinfrüchtigen Körnern zusammengefasst, wie Digitaria, Echinochloa, Eragrostis, Panicum, Setaria, Sorghum.[12]
Im Jahr 2019 wurden weltweit folgende Anbauergebnisse erzielt (Tabelle nach Erntemengen weltweit sortiert):
Neben den Grasfrüchten werden die Stängel, die Blätter und Wurzeln genutzt. Süßgräser sind eine wichtige Rohstoffquelle zur Gewinnung von Stärke, Zellulose, Zucker sowie Fetten und ätherischen Ölen. Sie können als Werk-, Bau- und Füllstoffe verwendet werden. Vor allem werden die verholzten Halme verschiedener Bambus-Arten in tropischen und subtropischen Gebieten Asiens zur Herstellung von Möbeln, Ess- und Trinkgefäßen oder Zäunen verwendet und nicht zuletzt im Haus- und Gerüstbau eingesetzt. In Nordwesteuropa wird das hier im Überfluss wachsende Schilf zu Eindeckung von Häusern verwendet.
Die Eismumie Ötzi soll einen Mantel aus Süßgräsern getragen haben.
Bambussprosse werden als Gemüse gegessen.
Zitronengräser (Cympopogon) werden als Gewürz- und Heilpflanzen verwendet. Ferner dienen Gräser zur Herstellung von alkoholischen Getränken wie Bier, Rum oder Korn. Als nachwachsende Rohstoffe gewinnen Süßgräser, vor allem Bambus und Zuckerrohr, zunehmende Bedeutung zur Herstellung von Bioalkohol als Treibstoff.
Schließlich sei noch die Verwendung zahlreicher Süßgrasarten mit auffälligen Blütenständen, wie beispielsweise das Pampasgras, als Ziergräser im Garten- und Landschaftsbau genannt.[12]
Die landwirtschaftliche Nutzung umfasst neben dem Getreideanbau die Nutzung zahlreicher Grasarten als Futterpflanzen für Rinder, Schafe oder Pferde in Form von Kulturgrasland wie Wiesen (Mahd zur Heugewinnung, Streunutzung, Silage) oder Weiden. Darüber hinaus werden geeignete Gräser für Rasen in privaten Gärten, in Parks, auf Golf- oder Sportplätzen eingesetzt, mit der Nutzungsart und -intensität angepassten Sortenmischungen.
Bei empfindlichen Menschen können Pollen von Süßgräsern die Bildung von Antikörpern Immunglobulin E (IgE) auslösen, was als Heuschnupfen bekannt ist. Sogenannte wasserlösliche I-Glykoproteine haften an der Pollenoberfläche, werden leicht an die Schleimhäute abgegeben und können allergische Reaktionen erzeugen. Ferner können in der Aleuronschicht der Getreidekörner enthaltene Prolamine durch eine immunologische Überempfindlichkeitsreaktion die als Zöliakie bezeichnete Krankheit auslösen.
Süßgräser sind weltweit verbreitet. Sie kommen von den Meeresküsten bis ins Hochgebirge, vom Äquator bis jenseits der Polarkreise in nahezu allen terrestrischen Ökosystemen vor und besiedeln dabei Standorte von großer ökologischer Bandbreite. Sie wachsen sowohl auf dauernassen bis extrem trockenen Böden als auch in sehr heißen bis arktisch kalten Klimaten.
Man findet Süßgräser flutend in Gewässern, bestandsbildend als Röhrichte, als Unterwuchs in Wäldern, auf wechselfeuchten wie auch trockenen Böden, an Straßenrändern, an Böschungen, auf Felsen – selbst Schotterflächen und Mauerkronen werden besiedelt. Die Familie der Süßgräser deckt nahezu alle denkbaren Standorttypen ab, wobei die einzelnen Arten und Populationen im Rahmen des Wettbewerbs um die Ressourcen (Konkurrenz) ihre jeweils eigenen Vorzugs- oder Existenzbereiche besiedeln. Etliche Pflanzenformationen außerhalb der Wälder werden im Wesentlichen durch Gräser aufgebaut. Die nordamerikanische Prärie, die Steppen Osteuropas, die Savannen Afrikas und die Pampa Südamerikas, aber auch die Wirtschaftswiesen und -weiden Europas sind die landschaftsprägenden natürlichen sowie unter menschlichen Einfluss entstandenen Grasländer der Erde, in denen Bäume und Sträucher zurücktreten oder ganz fehlen.
In der Erdneuzeit (Känozoikum) entstanden die modernen Familien der Blütenpflanzen, so auch die Gräser. Sie waren zunächst auf bewaldete und sumpfige Gebiete beschränkt. Mit der Entwicklung des kontinuierlichen Wachstumsprozesses und der Windbestäubung wurden ab dem Oligozän die offenen Länder erobert. Steppen und Grasländer breiteten sich vor allem im Miozän aus. Man nimmt an, dass die Evolution der Süßgräser mit jener der großen Weidetiere (Wiederkäuer, Pferde, Kamele etc.) parallel ging.[10]
Erkenntnisse indischer Wissenschaftler aus dem Jahr 2005 gehen einem Bericht der Zeitschrift Science zufolge davon aus, dass sich Gräser bereits in der Kreidezeit, dem letzten Abschnitt des Erdmittelalters (Mesozoikum), entwickelt haben. Diese Annahme geht auf Funde von Pflanzenbestandteilen zurück, die im fossilen Dung (Koprolith) von Dinosauriern gefunden wurden und auf reis- und bambusähnliche Gräser deuten.[14]
Süßgräser gehören zu den im Verlauf der Evolution sekundär entstandenen windblütigen Angiospermen. Spuren von Pollenkitt in Gräsern weisen darauf hin, dass die Vorgänger biotisch durch Vögel und Insekten bestäubt wurden. Pollenkitt verklebt die Pollenkörner zu größeren Übertragungseinheiten, was bei der Windbestäubung, die schwebfähige und leichte Pollen verlangt, störend wäre.[15]
Im Zuge des Übergangs zur Windblütigkeit wurden die Blüten reduziert. Entwicklungsgenetische Befunde deuten darauf hin, dass die Vorspelzen ein Verwachsungsprodukt zweier Blütenhüllblätter von ursprünglich drei und die Schwellkörperchen aus inneren Tepalen hervorgegangen sind. Die Gräserblüte lässt sich somit vom Grundtypus der dreizähligen Blüten einkeimblättriger Pflanzen ableiten mit zwei Kreisen à drei Blütenhüllblättern, zwei Kreisen à drei Staubblättern sowie drei Fruchtblättern. Der dreifächrige Fruchtknoten der Süßgräser wurde einfächrig und enthält nur noch eine Samenanlage. Vom äußeren und inneren Staubblattkreis blieb nur der äußere Ring erhalten. Vom inneren Hüllblattkreis blieben nur die zwei als Schwellkörperchen dienenden Schuppen, die dritte Tepale fiel aus. Nur bei einigen tropischen Arten sind noch drei Lodiculae vorhanden. Der äußere Hüllblattkreis besteht nur noch aus der Vorspelze, die aus zwei getrennten Blütenhüllblättern entstanden ist. Bei wenigen tropischen Gräsern sind zwei getrennte Vorspelzen erhalten. Die dritte Tepale fiel wiederum aus.
Etwa 80 % der Grasarten haben mehr als einen Chromosomensatz im Zellkern. Hybride, zum Teil auch fruchtbare, sind bei Süßgräsern selbst zwischen Gattungen nicht selten.[7] Viele der heutigen Gräser waren in der Naturlandschaft vermutlich nicht in der jetzigen Form vorhanden. Es wird angenommen, dass viele Graslandarten ihren Ursprung in diploiden Sippen haben, die während der Eiszeiten in südlichen Rückzugsgebieten überdauerten. In einem langen Prozess der Rückwanderung, der Anpassung an veränderte Standortbedingungen und verschärfter Konkurrenz sowie durch die vom Menschen seit dem Neolithikum neu geschaffenen Lebensräume konnten durch Kreuzungen diploider Elternarten tetra-, bis polyploide Sippen (Allopolyploidie) entstehen. So sind zum Beispiel Anthoxanthum odoratum, Agrostis stolonifera, Dactylis glomerata und Poa pratensis Hybride alter diploider Sippen.[11]
Die Typusgattung der Familie der Poaceae ist Poa. Der Gattungsname Poa ist vom griechischen Wort póa abgeleitet und bedeutet Kraut, Gras, Pflanze. Die veraltete Bezeichnung für die Familie lautet Gramineae. Nach dem Internationalen Code der Botanischen Nomenklatur Artikel 14 ist die weitere Verwendung des alten Begriffes als Ausnahme von den strengen Regeln erlaubt und damit legitim: Poaceae = Gramineae Jussieu nom. cons. (nomina conservanda) et nom. alt. (nomen alternativum).[16][17][18]
Süßgräser sind Bedecktsamer (Magnoliopsida). Im Gegensatz zu den Nacktsamern (Gymnospermae) ist bei ihnen die Samenanlage im Fruchtknoten eingeschlossen. Die Familie der Süßgräser gehört innerhalb der Einkeimblättrigen Pflanzen (Monokotyledonen) zur Ordnung der Süßgrasartigen (Poales). Die Familie umfasst etwa 10.000 Arten mit je nach verwendeter Systematik 600 bis 700 Gattungen. Die Poaceae sind in 13 Unterfamilien von ungleicher Größe unterteilt, die noch weiter in insgesamt 46 Tribus gegliedert sind. Die Unterfamilien können vom phylogenetischen Standpunkt aus zu zwei Hauptgruppen, „BEP-clade“ und „PACC-clade“ zusammengefasst werden.
Die allgemeinen Informationen dieses Artikels entstammen den unter Literatur und Weblinks aufgeführten Referenzen (Morphologie, Standorte, Verbreitung etc.). Darüber hinaus sind einzelne Aspekte, Spezialthemen, Zahlen usw. den aufgeführten Einzelpublikationen entnommen.
Die Süßgräser (Poaceae = Gramineae) sind eine Pflanzenfamilie in der Ordnung der Grasartigen (Poales). Mit etwa 12.000 Arten in rund 780 Gattungen sind sie eine der größten Familien innerhalb der Blütenpflanzen. Sie sind weltweit in allen Klimazonen verbreitet und durch eine typische grasartige Gestalt gekennzeichnet.
Viele Arten der Süßgräser gehören zu den ältesten Nutzpflanzen und sind seit alters für den Menschen von lebenswichtiger Bedeutung. Alle Getreide wie Weizen, Roggen, Gerste, Hafer, Hirse, Mais und Reis zählen zu dieser Pflanzengruppe. Sie stellen in Form von Marktfrüchten oder als Viehfutter in der Veredelung heute die Basis für die Ernährung der Weltbevölkerung dar. Als Gras- oder Grünland wie Wiesen und Weiden, aber auch Steppen und Savannen prägen sie in weiten Teilen der Erde das Landschaftsbild.
Bugʻdoydoshlar (gʻalladoshlar) (Poaceae yoki Gramineae) — bir urugʻ pallali oʻsimliklar oilasi. Bir yillik, ikki yillik yoki koʻp yillik oʻt, baʼzan buta yoki daraxtlardan tashkil topgan. Poyasi silindrsimon tik yoki yonboshlab koʻtarilib oʻsadi, boʻgʻimlarga boʻlingan, ichi gʻovak, shuning uchun u poxol yoki somon deb ham ataladi. Boʻyi 1 sm dan bir nechta oʻn m gacha; baʼzilarining poyasi yogʻochlanib baland boʻlib oʻsadi (mas, bambukning boʻyi 40 m, diametri 30 sm gacha). Barglari oddiy, ipsimon, qalami, bandsiz, navbatlashib joylashgan, qinli. Poyaning bir qismi shu barg qini ichida joylashadi. Gullari ikki jinsli, baʼzan bir jinsli, mayda, koʻrimsiz, oddiy toʻpgullarga (boshoqchalarga) yigʻilgan; bular esa murakkab toʻpgullar, shingil, roʻvak va boshqa hosil qiladi. Boshoqchalari bitta yoki bir nechta, baʼzan koʻp gulli. Mevasi don, baʼzan danaksimon, yongʻoqsimon yoki rezavor. Urugʻining koʻp qismi kraxmalli endospermdan iborat. B.ning koʻpchiligi uzun ildiz poyali va popuk ildizli. B.ning 600 turkumi, 10000 turi maʼlum. Oʻzbekistonda 91 turkumi (270 turi) usadi. Kupincha B. uchta kenja oilachaga bulinadi. B. yer sharining hamma qismida keng tarqalgan, koʻpchilik turlari tropik mamlakatlarda uchraydi. B. qalin oʻtzorlar, pichanzorlar, oʻtloqlar, savanna, preriy va boshqa hosil qiladi. B.ga muhim madaniy don oʻsimliklaridan — bugʻdoy, sholi, makka, suli, arpa, javdar, tariq, oq joʻxori va boshqa (qarang Donli ekinlar) hamda shakarqamish kiradi, shuning uchun ham kishilar hayotida ularning ahamiyati katta. Yovvoyi holda oʻsuvchi — oqsoʻxta, raygras, bugʻdoyiq va boshqa koʻpchilik turlari qishloq xoʻjaligi hayvonlari uchun yemxashak (qarang Yem-xashak ekinlari) hisoblanadi.
B.ning baʼzi turlaridan texnik oʻsimlik sifatida foydalanib, ulardan kraxmal, spirt, qogʻoz, xushboʻy va yeyiladigan moy olinadi va qurilish materiallari, arqon, chutka va boshqa tayyorlashda foydalaniladi. Koʻpchilik turlari dorivor, baʼzilari manzarali. Kuchma qumlarni mustahkamlash uchun ekiladigan turlari ham bor. Baʼzilari begona oʻt.
Oʻktam Pratov.

Bugʻdoydoshlar (gʻalladoshlar) (Poaceae yoki Gramineae) — bir urugʻ pallali oʻsimliklar oilasi. Bir yillik, ikki yillik yoki koʻp yillik oʻt, baʼzan buta yoki daraxtlardan tashkil topgan. Poyasi silindrsimon tik yoki yonboshlab koʻtarilib oʻsadi, boʻgʻimlarga boʻlingan, ichi gʻovak, shuning uchun u poxol yoki somon deb ham ataladi. Boʻyi 1 sm dan bir nechta oʻn m gacha; baʼzilarining poyasi yogʻochlanib baland boʻlib oʻsadi (mas, bambukning boʻyi 40 m, diametri 30 sm gacha). Barglari oddiy, ipsimon, qalami, bandsiz, navbatlashib joylashgan, qinli. Poyaning bir qismi shu barg qini ichida joylashadi. Gullari ikki jinsli, baʼzan bir jinsli, mayda, koʻrimsiz, oddiy toʻpgullarga (boshoqchalarga) yigʻilgan; bular esa murakkab toʻpgullar, shingil, roʻvak va boshqa hosil qiladi. Boshoqchalari bitta yoki bir nechta, baʼzan koʻp gulli. Mevasi don, baʼzan danaksimon, yongʻoqsimon yoki rezavor. Urugʻining koʻp qismi kraxmalli endospermdan iborat. B.ning koʻpchiligi uzun ildiz poyali va popuk ildizli. B.ning 600 turkumi, 10000 turi maʼlum. Oʻzbekistonda 91 turkumi (270 turi) usadi. Kupincha B. uchta kenja oilachaga bulinadi. B. yer sharining hamma qismida keng tarqalgan, koʻpchilik turlari tropik mamlakatlarda uchraydi. B. qalin oʻtzorlar, pichanzorlar, oʻtloqlar, savanna, preriy va boshqa hosil qiladi. B.ga muhim madaniy don oʻsimliklaridan — bugʻdoy, sholi, makka, suli, arpa, javdar, tariq, oq joʻxori va boshqa (qarang Donli ekinlar) hamda shakarqamish kiradi, shuning uchun ham kishilar hayotida ularning ahamiyati katta. Yovvoyi holda oʻsuvchi — oqsoʻxta, raygras, bugʻdoyiq va boshqa koʻpchilik turlari qishloq xoʻjaligi hayvonlari uchun yemxashak (qarang Yem-xashak ekinlari) hisoblanadi.
B.ning baʼzi turlaridan texnik oʻsimlik sifatida foydalanib, ulardan kraxmal, spirt, qogʻoz, xushboʻy va yeyiladigan moy olinadi va qurilish materiallari, arqon, chutka va boshqa tayyorlashda foydalaniladi. Koʻpchilik turlari dorivor, baʼzilari manzarali. Kuchma qumlarni mustahkamlash uchun ekiladigan turlari ham bor. Baʼzilari begona oʻt.
Oʻktam Pratov.
Famîleya genim û kurtîjan, famîleya genim, famîleya biqurfan (Poaceae = Gramineae) navê famîleyeke riwekan e ku li seranser erdê, di tevahiya rewşên avhewayê de endamên vê famîleyê kêm-zêde digihên. Di desteya genimiyan (genimkî) Poales de cih digire.
Çend ji endamên vê komê li Kurdistanê digihên, ne diyar e, lewra li ser floraya Kurdistanê xebateke zanistî nehatiye kirin. Li parêzgeha Dîlok, Riha, Amedê, li deşta Mûşê û Rewanê çandiniya genim, ceh pêşketî ye.
Giyayên bi hazaran salan in, ji aliyê mirovan ve çandiniya wan tên kirin in. Genim, ceh, tarîgan (şilêl), baçîk (xamkêsan), riz (birinc), garis (gilgil) ûêd di vê malbatê de cih digirin ku bo mirov û ajalanên xurdemeniyên sereke ne. 650 cins û bi qasî 10.000 endamên vê komê hene. Ji vê famîleyê re famîleya giyayên şîrîn ên mêrg û çîmenan jî tê gotin.
Les fenaesses, c' est ene famile di plantes ki dnèt del boune waide, u ki sont semêyes po do four u come dinrêyes.
Mins gn a eto des såvadjès fenaesses ki sont des crouwås.
No del famile e sincieus latén: Poaceae
Les fenaesses, c' est ene famile di plantes ki dnèt del boune waide, u ki sont semêyes po do four u come dinrêyes.
Mins gn a eto des såvadjès fenaesses ki sont des crouwås.
No del famile e sincieus latén: Poaceae
De gerzen (Gramineae of Poaceae; beide nammen binne tastien) binne in famylje fan it skift fan 'e gerseftigen (Poales). Ta dizze famylje hearre goed 8.000 plantesoarten. De gerzen binne ien fan de súksesfolste groepen planten op 'e ierde. It blêd wurdt troch weidzjende bisten opiten en de sieden, it nôt foarmet sa'n 80% fan it minsklik iten. In foarnaam part fan de rest fan ús iten wurdt foarme troch weidzjende dieren dy't ek wer fan gers ôfhinklik binne. Koartsein, ús ôfhinklikens fan de famylje fan de gerzen bedraacht sa'n 95% fan ús ferlet fan iten.
It beweide gers kin sa suksesfol wêze omdat it syn ynvestearring benammen ûnder de grûn hâldt. It bewarret syn fiedselfoarrie yn de woartels. Dat makket dat it alhiel net slim is, wannear't in weidzjend bist alle blêden mei sprietfoarm boppe de grûn opyt, want yn koarte tiid waakst der alwer nij blêd út de enerzjyreserves yn de woartels. Ek in steppebrân makket it gers neat út.
It gers foarmet in soartemint symbioaze mei de weidzjende bisten, troch in fan wjerskanten ôfhinklikens: De bisten binne ôfhinklik fan it gers, mar it gers is dat ek fan de bisten, omdat de bisten net allinnich it gers opite mar ek oare planten dy't oars heger groeie soene as it gers en it wichtige sinneljocht fuort helje soene.
De gerzen (Gramineae of Poaceae; beide nammen binne tastien) binne in famylje fan it skift fan 'e gerseftigen (Poales). Ta dizze famylje hearre goed 8.000 plantesoarten. De gerzen binne ien fan de súksesfolste groepen planten op 'e ierde. It blêd wurdt troch weidzjende bisten opiten en de sieden, it nôt foarmet sa'n 80% fan it minsklik iten. In foarnaam part fan de rest fan ús iten wurdt foarme troch weidzjende dieren dy't ek wer fan gers ôfhinklik binne. Koartsein, ús ôfhinklikens fan de famylje fan de gerzen bedraacht sa'n 95% fan ús ferlet fan iten.
It beweide gers kin sa suksesfol wêze omdat it syn ynvestearring benammen ûnder de grûn hâldt. It bewarret syn fiedselfoarrie yn de woartels. Dat makket dat it alhiel net slim is, wannear't in weidzjend bist alle blêden mei sprietfoarm boppe de grûn opyt, want yn koarte tiid waakst der alwer nij blêd út de enerzjyreserves yn de woartels. Ek in steppebrân makket it gers neat út.
It gers foarmet in soartemint symbioaze mei de weidzjende bisten, troch in fan wjerskanten ôfhinklikens: De bisten binne ôfhinklik fan it gers, mar it gers is dat ek fan de bisten, omdat de bisten net allinnich it gers opite mar ek oare planten dy't oars heger groeie soene as it gers en it wichtige sinneljocht fuort helje soene.
Graas (botanische naam: Gramineae of Poaceae, bei name zeen toegestange) is ein vanne saortriekste plantjefemielies oppe aerd. 't Guuef zoeaget 8000 saorte. Leje van dees femielie kómme op alle werrelddeiler veur. Op Antartica greutj beveurbeildj ouch 'n saort aan smeelj. Aafhenkelik vanne saort wuuerdj graas gebroek es voor veur vieë of luuj, veur touwe van te maken, veure sier of es bekleijing van parken of sportveljer. Ónger graas vallen ouch alle graansaorte en reet.
Toet recèntelik gove fossiel vónjste aan det graas zoeaget 55 miljoen jaor trögk evolueerdje. Recènte vónjste vanne graasechtige in coproliete (fossiel faecaelieje) van dinosauriërs oet 't kriet verweze nao 65 miljoen jaor trögk. Dateringe vanne aafkóms van ries suggerere 107 toet 129 miljoen jaor.
De relaties tösse de subfemielies Bambusoideae (bamboe-echtige), Ehrhartoideae (worónger ries) en Pooideae (ónger angere kaore) waeren es volg verklaordj: de Bambusoideae enne Pooideae zeen mieë aanein verwandj es ane Ehrhartoideae. De sjeijing vóltróg zich in 'n betrèkkelik kórte periode van 4 miljoen jaor.
Graas (botanische naam: Gramineae of Poaceae, bei name zeen toegestange) is ein vanne saortriekste plantjefemielies oppe aerd. 't Guuef zoeaget 8000 saorte. Leje van dees femielie kómme op alle werrelddeiler veur. Op Antartica greutj beveurbeildj ouch 'n saort aan smeelj. Aafhenkelik vanne saort wuuerdj graas gebroek es voor veur vieë of luuj, veur touwe van te maken, veure sier of es bekleijing van parken of sportveljer. Ónger graas vallen ouch alle graansaorte en reet.
De grasfamilie of in 't Zuudôostelik West-Vlams gesfamilie (Poaceae of Gramineae) is e familie van êenzoadlobbige blommeplantn in de orde Poales met oengeveer 12.000 sôortn, verdêeld over 780 geslachtn. Groangewassn, lyk terwe, rogge, gerst, hoaver, maïs en rys, zyn verteegnwoordigers van de familie, moar ook bouwmaterioaln (bamboe, riet, strôoi).
De stiengel van de plante is over 't algemêen hol tusschn de knoopn. De bloarn zyn lank, smal en parallelnervig. De bladbasis, 't oenderste dêel van 't blad, vormt e kokervormige bladschede roend de stiengel.
Grassn zyn te viendn over hêel de weireld ip al de continentn, zelfs ip Antarctica. Ze zyn de dominante vegetoasje van uutgestrekte gebiedn, lik steppe, savanne, pampa, Amerikoansche prairie, bus en toendra. Die graslandn èn d' oentwikkelienge van groaznde bêestn meugelik gemakt.
De grasfamilie is oenderverdêeld in 12 subfamilies:
De grasfamilie of in 't Zuudôostelik West-Vlams gesfamilie (Poaceae of Gramineae) is e familie van êenzoadlobbige blommeplantn in de orde Poales met oengeveer 12.000 sôortn, verdêeld over 780 geslachtn. Groangewassn, lyk terwe, rogge, gerst, hoaver, maïs en rys, zyn verteegnwoordigers van de familie, moar ook bouwmaterioaln (bamboe, riet, strôoi).
De stiengel van de plante is over 't algemêen hol tusschn de knoopn. De bloarn zyn lank, smal en parallelnervig. De bladbasis, 't oenderste dêel van 't blad, vormt e kokervormige bladschede roend de stiengel.
Grassn zyn te viendn over hêel de weireld ip al de continentn, zelfs ip Antarctica. Ze zyn de dominante vegetoasje van uutgestrekte gebiedn, lik steppe, savanne, pampa, Amerikoansche prairie, bus en toendra. Die graslandn èn d' oentwikkelienge van groaznde bêestn meugelik gemakt.
De grezenfemilie (Latien: Gramineae of Poaceae) is een van de succesvolste plaantefemilies op eerde. Der bestaon zo'n 8.000 soorten. Lejen van disse femilie koemen op alle werelddelen veur, zelfs op Antarctica greui nog een smele-soort. Vegetasies waor gressoorten domeneren wonnen greslanen eneumd.
Over 't algemeen verwies gres naor een greune plaante mit kerakteristieke dunne blaojen, dee rechstreeks uut de wortels onder de grond greuien.
't Blad (vake) mit een stukjen stengel wonnen deur grezende dieren egeten en de zaojen vormen zo'n 80% van 't voedsel van de meens. Een groot deel van de reste van ons voedsel wonnen evormd deur de grezende dieren dee oek weer van gres ofhankelijk bin. Kortumme, onze ofhankelijkheid van de femilie van de grezen is zo'n 95% van onze voedselbehoefte.
De meens kan gien gres verteren. Hij bezit namelijk gien verteringssappen dee de waand van een plaantaorige en de vaotbundels (narve) kunnen ofbreken. Disse bestaon veurnamelijk uut cellulose. Plaante-eters kunnen dit wel, deur de anwezigheid van micro-organismen in 't maagdarmkenaal. Bie nerekers, zoas de ko, 't schaap, de wisent, de bizon en de gnoe vien disse microbiële vertering (fermentasie) plaose in de veurmagen (netmaag, pens en boekmaag). Bie aandere plaante-eters zoas 't peerd en de olifant, vient disse fermentasie plaose in de dikke darm. De meens eet allinnig de zaojen van grezen. Dee bevatten gien cellulose, mar heufzakelijk zetmeel. Soms kunnen de zaojen tot wel 20% suker bevatten, zoas bie sukermaïs, waorvan de onriepe zaojen as greunte egeten wonnen.
Al eeuwen wonnen gres deur de meens gebruuk, en dan mit name in de vorm van gresvelden. Zoksoort gresvelden dienen uuteenlopende doelen: van dekorasie van de tune, (wonnen dan gezon eneumd) of park tot 't gebruuk as sportveld, zoas een vootbalveld. Op de leste meniere kump gres nogal es in 't niejs, denk daorbie bieveurbeeld an de meuizame totstandkoeming van een goeie gresmatte in de Amsterdam ArenA.










De grezenfemilie (Latien: Gramineae of Poaceae) is een van de succesvolste plaantefemilies op eerde. Der bestaon zo'n 8.000 soorten. Lejen van disse femilie koemen op alle werelddelen veur, zelfs op Antarctica greui nog een smele-soort. Vegetasies waor gressoorten domeneren wonnen greslanen eneumd.
Over 't algemeen verwies gres naor een greune plaante mit kerakteristieke dunne blaojen, dee rechstreeks uut de wortels onder de grond greuien.
Hô-pún-kho (la. Poaceae ia̍h Gramineae; ji̍t. イネ科 ia̍h 禾本科) sī tan-chí-io̍h khui-hoe si̍t-bu̍t ê chi̍t tōa hun-lūi. Pún lūi tang-tiong hâm kok-si̍t chháu-á-lūi, tek-á, kap it-poaⁿ chū-jiân chháu-po͘ ê chháu-á, lóng-chóng ū tāi-khài 780 sio̍k kap iok-lio̍k 12,000 chéng.
Hô-pún-kho (la. Poaceae ia̍h Gramineae; ji̍t. イネ科 ia̍h 禾本科) sī tan-chí-io̍h khui-hoe si̍t-bu̍t ê chi̍t tōa hun-lūi. Pún lūi tang-tiong hâm kok-si̍t chháu-á-lūi, tek-á, kap it-poaⁿ chū-jiân chháu-po͘ ê chháu-á, lóng-chóng ū tāi-khài 780 sio̍k kap iok-lio̍k 12,000 chéng.
Ko e mohuku ko e hingoa fakatonga ki he ngaahi fuʻu ʻakau mei he kāinga poaceae pe gramineae ʻi he lea fakasaienisi. ʻOku ʻi ai ʻa e ngaahi faʻahinga ʻe taha mano tupu ʻi he kāingá ni. ʻOku nau kau ki he holongā poales, tatau mo e kāinga ʻo e pako.
Kā tatau ai pē mo e grass ʻi he lea fakapilitānia ʻoku ʻikai tatau ʻa e ngaahi mohuku kotoa pē ki he poaceae, pea ʻoku ʻi ai ha poaceae ʻoku ʻikai ha mohuku. Ko e fakatātā mahuʻinga taha: ko e tō pe kaho pe pitu (kofe) ʻoku nau kau ki he poaceae, kā ʻikai ui ha mohuku.
ʻI he lea fakatonga fakatupunga ko e mohuku ko ngaahi fuʻu ʻakau kinautolu ʻoku ngāueʻaki ki he kai maʻa e fanga monumanu, ʻa ia pulu, hoosi, kosi, mo e hā fua. ʻOku lahi ʻene tupu ʻi he kaikaiʻanga lahi ʻi ʻuta.
Ko e foʻi lea musie, ʻoku meimei tatau, kā ʻoku ne ngāueʻaki maʻa e ngaahi musie (malaʻe) ʻi he ngoue ʻi kolo.
ʻOku ngāueʻaki ʻi heni ko e mohuku = poaceae, ʻoku lava te ke pehē: ko e mohuku moʻoni.
Ko e ngaahi faʻahinga mohuku:
Ko e mohuku ko e hingoa fakatonga ki he ngaahi fuʻu ʻakau mei he kāinga poaceae pe gramineae ʻi he lea fakasaienisi. ʻOku ʻi ai ʻa e ngaahi faʻahinga ʻe taha mano tupu ʻi he kāingá ni. ʻOku nau kau ki he holongā poales, tatau mo e kāinga ʻo e pako.
Kā tatau ai pē mo e grass ʻi he lea fakapilitānia ʻoku ʻikai tatau ʻa e ngaahi mohuku kotoa pē ki he poaceae, pea ʻoku ʻi ai ha poaceae ʻoku ʻikai ha mohuku. Ko e fakatātā mahuʻinga taha: ko e tō pe kaho pe pitu (kofe) ʻoku nau kau ki he poaceae, kā ʻikai ui ha mohuku.
ʻI he lea fakatonga fakatupunga ko e mohuku ko ngaahi fuʻu ʻakau kinautolu ʻoku ngāueʻaki ki he kai maʻa e fanga monumanu, ʻa ia pulu, hoosi, kosi, mo e hā fua. ʻOku lahi ʻene tupu ʻi he kaikaiʻanga lahi ʻi ʻuta.
Ko e foʻi lea musie, ʻoku meimei tatau, kā ʻoku ne ngāueʻaki maʻa e ngaahi musie (malaʻe) ʻi he ngoue ʻi kolo.
ʻOku ngāueʻaki ʻi heni ko e mohuku = poaceae, ʻoku lava te ke pehē: ko e mohuku moʻoni.
Ko e ngaahi faʻahinga mohuku:
ahisiaina fisihina hana hefa kaho (ʻikai tukufakaholo) kaho fōlalahi kefukefu koane kofe (ʻikai tukufakaholo) kola limuvai matalaʻulie matapekepeka moengālō mohuku mohuku ano pe misilaite mohuku siamane mohuku vai mohukuʻapōpoa musie musie liku musie matala pulū musie tonga musie vailima musie vai pitu (ʻikai tukufakaholo) puakatau saafa tō (ʻikai tukufakaholo) tolulu totoa totoatahi salapona sipilini takatakaʻaleala
The Poaceae (cried Gramineae or true grasses an aw) is a lairge an nearly ubiquitous faimily o monocotyledonous flouerin plants. Wi mair nor 10,000 domesticatit an wild species, the Poaceae represents the fift-lairgest plant faimily, follaein the Orchidaceae, Asteraceae, Fabaceae, an Rubiaceae.[4] Tho commonly cried "gresses", seagresses, rashes, an segs faw ootside this faimily. The rashes an segs is sib tae the Poaceae, bein members o the order Poales, but the seagresses is members o order Alismatales.
Gresslands is estimated tae compone 20% o the vegetation kiver o the Yird. Poaceae leeves in mony ither habitats, includin weetlands, wids, an tundra.
Domestication o poaceous cereal craps sic as maize (corn), wheat, rice, baurley, an millet lies at the foondation o sedentary leevin an ceevilization aroond the warld, an the Poaceae still constitutes the maist economically important plant faimily in modren times, providin forage, biggin materials (bamboo, thack) an fuel (ethanol), as weel as fuid.
|dead-url= (help) Poaceae (e quajtur edhe Gramineae ose barishte) është një familje e madhe e bimëve njëfarëshe të lulëzueshme. Me më shumë se 10.000 lloje të buta dhe të egra, Poaceae përfaqëson familjen e pestë më e madhe të bimëve, pas Orchidaceae, Asteraceae, Fabaceae, dhe Rubiaceae.
Drithërat të tilla si misri, gruri, orizi, elbi, dhe meli qëndron në themelin e jetesës dhe qytetërimit në botës. Poaceae ende përbëjnë familjen bimore ekonomikisht më të rëndësishme në kohët moderne, duke siguruar foragjere, materiale ndërtimi (bambu, kashtë) dhe lëndë djegëse (etanol), si dhe ushqim.
Poaceae (bos. trave) ili Gramineae su velika i gotovo sveprisutna porodica monokotiledonskih cvjetnica, poznatih kao trave[1][2]
U Poaceae se uključuju trave žitarice, bambusi i trave prirodnih i kultiviranih travnjaka i pašnjaka. Trave imaju šuplje stabljije, osim na čvorovima i uske naizmjenične listove u dva reda. Donji dio svakog lista obuhvata stabljiku, formirajući lisni omotač. Sa oko 12.000 vrsta, peti po brojnosti biljnih porodica, nakon Asteraceae, Orchidaceae, Fabaceae i Rubiaceae .[3]
Procjenjuje se da zatravljena područja, kao što su savane i prerije, gdje su dominantne trave, čine 20% od vegetacijskog pokrov na Zemlji. Trave su također važan dio vegetacije u mnogim drugim staništima, uključujući i močvare , šume i tundre.
Trave su ekonomski izuzetno značajna porodica, jer daju životne namirnice od pripitomljenih žitarica, kao što su kukuruz, pšenica, riža, ječam i proso, kao i stočnu hranu, građevinski materijal (bambus, slama I sl.) i gorivo (etanol). Iako se obično nazivaju "trave", morska trava, rogoz i oštrice ne spadaju u ovu porodicu. Trske su red Poales, u Poaceae, kao članice, ali su morske trave članovi reda Alismatales.
Trave uključuju neke od najsvestranijih oblika biljnog života. Postale su raširena pri kraju perioda kreda (period)krede i nađene su u fosiliziranom izmetu dinosaura i (coprolitima), koji je sadržavao razne fitolite, uključujući i trava koje su povezane sa suvremenim rižama i bambusima.[4] Trave su prilagođeni uvjetima u bujnim kišnim šumama, suhim pustinjama, hladnim planinama, pa čak i međuplimnim staništima, a važe kao danas najrasprostranjenije vrste biljaka; trave su vrijedan izvor hrane i energije za sve vrste divljači i organske hrane. Naredni kladogram pokazuje potporodice i približne brojeve vrsta (u zagradama):[5]
Chloridoideae (1600)
Danthonioideae (300)
Micrairoideae (200)
Arundinoideae (50)
Panicoideae (3250)
Aristidoideae (350)
Oryzoideae (110)
Bambusoideae – Bambusi (1450)
Pooideae (3850)
Puelioideae (11)
Pharoideae (13)
Anomochlooideae (4)
Prije 2005. godine, fosilni nalazi su pokazivali da su trava evoluirala od prije oko 55 miliona godina. Nedavni nalazi trava poput fitolita iz krede, dinosaurskim koprolitima su pomjerili datiranje na prije oko 66 miliona godina.[6][7] In 2011, revised dating of the origins of the rice tribe Oryzeae suggested a date as early as 107 to 129 Mya.[8] Odnosi između tri potfamilije: Bambusoideae, Oryzoideae i Pooideae u BOP kladusu su riješeni tako da su međusobno uže povezanije Bambusoideae i Pooideae, nego Oryzoideae .[9]
Stabljike trava su obično cilindrične (rjeđe spljoštene, ali ne i trouglste) i šuplje, povezane čvorovima ili koljencima, odakle rastu sjedeći listovi, čiji rukavac obavija stabljiku. Između stabljike i lista, nalazi se jezičak. Listovi trava su gotovo uvijek naizmjenični i jednostrani i imaju paralelne vene (provodne sudove). Svaki list je diferenciran u u donji omotač koji obavija stabljiku i i oštre, cjelovite (glatke) rubove. Liske mnogih trava su ojačane silikatnim fitolitima, koji obeshrabruju ispašu životinja; neke su dovoljno ošttre da zasijecaju I ljudsku kožu. Membranozni dodatak ili rub dlake zove se ligula, koja na raskrsnici između plašta i sječiva, sprečava vodu ili insekte da prodru u korice između stabljike I lista.
Cvjetovi Poaceae su neugledni i karakteristično raspoređeni u klasiće, od kojih svaki imaobično tri, ali općenito jedan ili više cvjetova. U klasića su dalje grupirani u klasove, a ovi i veće klasove ili metlice. Dio klasa koji nosi cvjetova zove se rahila. Klasić se sastoji od dvije brakteje (ili ponekad i manje) u bazi, pod nazivom pljeve, za jedan ili više cvjetova. Ocvijeće se sastoji od cvijeta okruženog sa dvije pljevice, jedn vanjska lema i jedns unutrašnja palea. Cvjetovi su obično hermafroditni , a kukuruz kao važan izuzetak je anemofilan , što znači da ga oprašuje vjetar.
Cvijetni omotač (periant) se svodi na dvije ljuske, pod nazivom lodikule, koje se proširuju i vežu za širenje lema i palea; oni se uglavnom tumače kao modificirane sepale. Ova kompleksna struktura se može vidjeti na slici na desnoj strani, posmatrajući klasić pšenice (Triticum aestivum). Plod trava je kariopsis ili pšeno, u kojem je sjemenski sloj spojen sa zidom ploda.
Postoje tri klasifikacije ukupnih oblika rasta trava:
Uspjeh trava u adaptivnoj radijaciji počiva na ulozi njihove morfologije u procesima rast u njihovom fiziološkoj različitosti. Većina trava se može podijeliti u dvije fiziološke grupe:
C3 trave se označavaju kao "hladnosezonske" trave, dok C4 biljke podrazumijevaju:
Porodica trava je jedna od najrasprostranjeniji i najobilnijig grupa biljaka na Zemlji. Trave se mogu naći na svim kontinentima, a nema ih samo na centralnom Grenlandu i većini Antarktika.[10][11]
Trave su dominantna vegetacija u mnogim staništima, uključujući i travnjake, pašnjake, livade i stepe. Također se javljaju kao manji dio vegetacije u gotovo svakom drugom kopnenom staništu. Biomi u kojima dominiraju trave su travnjaci. Ako se računaju samo velika, susjedna područja travnjaka, ti biomi pokrivaju 31% površine naše planete. Travnjaci uključuju pampase, stepe i prerije.
Trava osiguravba hranu za ispašu sisara, kao što su stoka, jeleni slonovi, kao kao i mnoge druge životinje, uključujući i insekte i ostale beskičmenjake. Mnoge vrste životinja jede travu kao glavni izvor hrane, a nazivaju se graminivori, koji uključuju goveda, ovce, konje, zečeve i mnoge. Trave također jedu i omnivori ili povremeno čak i oni svaštojedi, koji su prvenstveno mesožderne životinje.
Trave su neobične po tome da se meristem nalazi u blizini dna biljke, pa se brzo oporavie od obrezivanje na vrhu.
Evolucija velikih biljojednih životinja u kenoziku doprinijela je širenju trava. Bez velikih svaštojeda, opožarene površine brzo koloniziraju trave, a sa dovoljno kiše, i mladice drveća. Drveće na kraju istisne najveći dio trave. Gazeći svejedi uništavaju mlado drveće, ali ne i trave.[12][13]
Porodica trava daje vjerojatno najvažnije sirovine za ljudsku ekonomiju. U stvari, oko 70% obradivog zemljišta u svijetu je zasađeno travama, a 50% kalorija čovječanstvo konzumira iz brojnih vrsta trava koje se direktno koriste u hrani, ili indirektno kao stočna hrana za domaće životinje. U uvjetima globalne proizvodnje, četiri najvažnije kulture su trave: šećerna trska (Saccharum officinarum ), pšenica, riža i kukuruz, a ječam i sirak su među prvih 12. S druge strane, nekoliko vrsta trava se koristi u industriji.
Trave uključuju 759 poznatih rodova sa ukupno 11.554 priznatih vrste među kojima su I pšenica, kukuruz, riža, zob i druge žitarice.
|author= (pomoć) |accessdate= (pomoć) |journal= (pomoć) Le Poaceae Barnhart (ciamàde apò Gramineae) i è ìna famìa botànica de piànte erbàcee, o de rar fés lignùze, che le aparté al ùrden de le Poales, (Liliopsida). Con de piö de 670 zèner e quàze 10.000 spéci discriìde, le Poaceae i è la quàrta famìa con piö tàta richèsa de spéci dòpo de le Asteraceae, le Orchidaceae e le Fabaceae (leguminùze); però, i è la prìma per emportànsa económica globàl.[1] De fat, la diéta dei òm l'è furmàda per 'na gran part de derivàcc de le poaceae. En piö se se cunsìdera che apò a le bès•ce i è alimentàde suratöt co le granàie o col fé che i è ac per lo piö de la famìa de le poaceae, se capés che se tràta de 'na famìa de piànte de 'mportànsa capitàl.
L'è 'na famìa cosmopolìta, che la g'ha cunquistàt la gran part de i ambièncc ecològic dispunìbii söl nòst pianéta, de le zòne dezèrtiche 'nfìna ai ecosistémi de àiva salàda, e de le depresiù palüdùze 'nfìna ai sistémi muntagùs piö alcc. Chèsta braüra 'ndèl adatàs la g'ha dat come rezültàt 'na biodiversità enòrme che la s'è manifestàda ta endèla morfologìa, che 'ndèla fiziulugìa, che 'ndèi sistémi de reprudusiù e 'ndèle asociasiù condei óter organìsmi, isé che le graminàcee i è de cunsiderà 'na famìa 'nteresànte, m,ìa apéna per la sò 'mportànsa económica, ma apò a per la releànsa biológica.
En generàl i è èrbe, che de rar le pöl rià a véser lignùze compàgn dei bambú tropicài, cespitùze, rizomatùze o stulunìfere. El cìclo vitàl el pöl véser anuàl, bienàl o perène. Chèle anöàle, come che se pöl capì, le se reprodùce apéna 'na ólta endèl sò ciclo vitàl (compàgn che 'l formét o avena, per ezèmpe). Le spéci perèni 'nvéce le pöl reprodücìs divèrse ólte o 'na ólta apéna.
El gamp l'è cilìndric o condèna sesiù trasversàl elìtica, condèi gróp pié e la part tra 'n gróp e l'óter (internódo) öda (però quac spéci le g'ha 'l gamp töt pié compàgn del furmintù o 'n quac bambú). I gróp i è 'n bris piö gròs che i internòdi e söi gróp vé fò le fòie e i böcc. I internòdi àle ólte i è 'n pó schisàcc endóche se svelöpa le ramificasiù. En brizinì piö 'n alt del gróp gh'è 'n meristema intercalàr a fùrma de anèl che 'l detèrmina el slongamènt del gamp. Endèn quac zèner gh'è de 2 a 6 gróp bei tacàcc 'ntra de lur (che i se ciàma gróp compòsti), ognü co la sò fòia. Endèl Cynodon dactylon, per ezèmpe, i gróp i stà 'nsèma a dù a dù isé le fòie le sömèa opòste. En general i internòdi bazài i è piö cürcc de chèi ensìma; quan che gh'è divèrsi gróp bazài tacàcc giü a l'óter, le fòie le sömèa dispunìde endèna rözèta, che sömèa le rözète bazài tìpiche de 'n quac ótra famìa botànica. Le sórcc de gamp de le poaceae i è chèsti:
Le g'ha fòie a dispuzosiù alternàda, dìstiche, furmàde de sòlit de guàina, lígula e làmina. La làmina la stà bèla tacàda töt entùren al gamp, i sò òrli i se 'ncaàla però i se fónt mìa 'nsèma (l'è rar fés che se tróe spéci 'ndóche i se tàca onsèma a furmà 'n tübo). La lígula l'è 'na apendìce picinìna membranùza, o de rar n grüpèt de pilizì, sitüàt endèla zòna de üniù de la làmina co la guàina. La làmina l'è sèmpia, de sòlit lineàr, co la nervadüra paraléla. La pöl véser piàta o 'n quac vólte 'nridulàda sö a furmà come 'n tübo. La pöl véser cuntìnuasiù de la guàina o ìga 'n gambì. En piö, se pöl troà 'na sèrta variabilità per ognü de chèsti órghegn:
I ciöf i è l'unità strütüràla tìpica de quàze töte le spéci de poaceae. I se fùrma a pàrter dei böcc a le sèe o de chèi segondàre del meristéma bazàl del as principàl. Ognü de chèst böcc segondàre i àmbia a furmàs quan che la piànta la g'ha dò o tré fòie. Ognö de chèsti böcc, dòpo che i g'ha fat fò le prìme fòie, 'l fùrma 'l sò stès sistéma de raìs. La sóma de töcc chèsti böcc l'è chèl che dà strütüra e la la fùrma de ciöf de 'na piànta de le poaceae. Quan che la piànta l'è 'ndèl peìodo de vegetasiù la seghéta a fa sö ciöf e fòie nöe. Ògna ciöf, quan che l'è 'l sò momènt, el cuminciarà a fa sö ciöfècc nöf.
La nfiurescènsa elementàr de le gramineae l'è 'na spìga picinìna furmada de giü o piö fiùr sùra 'n ràchide articolàt (ciamàt rachìla), de spès cürt fés e prutigìt de le bràtee stérii ciamàde glöme. Chesto tìpo de 'nfiurescènsa la ciàpa 'l nòm de spighèta. I fiùr i pöl véser ermafrodìcc o unisesuài e i g'ha 'n pereànse rüdimentàl de 2 o 3 elemèncc, le glömète. Chèste glömète i è i órghegn che, quan che i vé gròs, i fà dèrver la cazèla fiuràla a la fiuridüra, per permèter de mèter fò 'l stìgma piümùs e i stam. Le cazèle fioràle i è furmàde de la lèma, tacàda a la rachìla, e de la pàlea 'nserìda sùra l'as fiuràl che 'l nas sùra a la rachìla a la sèa de la lèma e 'l té sö i órghegn fiurài véri e pròpri. La lèma (o glömèta inferiùra) la g'ha la fùrma de 'na chìglia e coi sò òrli la bràsa sà la pàlea. La arìsta la nàs ensìma a la lèma o söl sò dòrs. La pàlea o glömèta superiùra l'è lanceolàda, binervàda e l'è come 'n coèrcc che 'l cuàrcia zó 'l fiùr. Le glöme i è 'nserìde sùra la rachìla, giöna de sóta de l'ótra.
Le spighète i è a sò ólta reünìde en enfiurescènse compòste a la maniéra de 'na gratèla. Le sórcc piö frequènte i è:
El fröt o grà de le gramineae l'è cariòside, fröt sèc indeiscènt, condèna somésa co la capöcia saldàda al perecàrpe 'nfìna a furmà 'na copertüra sitìla. Chèsta copertüra la stà töta 'ntùren a l'embriù e a l'endospèrma. Chèsto fröt l'è 'n sostànsa la variasiù del achénio, aisebé che gh'è 'na bèla varietà de fröcc endèla famìa (éder apò Werker 1997). Endèn quac zèner, cóme Zizianopsis o Eleusine, el perecàrpe l'è mìa saldàt enséma a la somésa, isé che 'l fröt l'è de fat 'n achénio (o en utrícol a segónda del autùr). Endèn quac Bambùseae el fröt l'è 'na nus o 'na bàca, envéce 'ndèl zèner Sporobolus el perecàrpe l'è mucilaginùs e 'l làsa nà fò la somésa quan che 'l se 'mpiómba de àiva. Divèrse zèner, compàgn de Aristida, Stipa, Piptochaetium, Oryza e quàze töte le Paníceae lew g'ha i carióse che se destàca de la piànta 'nciurciàcc sö 'ndèla lèma e 'ndèla pàlea. Endèle Andropogoneae i è le glöme che rèsta 'nseràde dét endèl carióse. Endèl Pennisetum e 'ndèl Cenchrus se destàca töta la spighèta con entùren 'n invòlucro de spìne. La fùrma del carióse la vària fés a segónda del zèner, e 'l pöl véser quàze rotónt come 'ndèl Briza, bilónc come 'ndèl Hordeum, lanceolàt compàgn che chèl de la Poa o adiritüra quàze lineàr come 'ndèla Vulpia. Endèla part bàsa del carióse a vardàl de schéna, se èt l'embriù piö o méno elìtic cuarciàt zó del perecàrpe trasparènt. De l'ótra bànda, a l'altèsa de la sütüra carpelàr, se pöl véder, ac en trasparènsa, la mácula filar o fil (o zòna de uniù de la somésa col carpèl), che la pöl vése puntifórme, compàgn che 'ndèla Poa e 'ndèle Paniceae, a öf, compàgn che 'ndèla Briza subaristata, o lineàra, compàgn che 'ndèl Hordeum, Vulpia o Festuca.
Las graminèas o poacèas (Poaceae) son una familha de plantas erbacèas, o fòrça rarament lenhosas, apertenent a l'òrdre Poales de las monocotiledonèas (Liliopsida). Amb mai de 670 genres e près de 10.000 espècias descrichas, las gramineas son la quatrena familha amb màger riquesa d'espècias après de las compausadas (Asteraceae), las orquidèas (Orchidaceae) e las leguminosas (Fabaceae); mas, definitivament, es la primièra en importància economica globala. En fach, la màger part de la dièta dels èssers umans proven de las gramineas, tant en forma dirècta (granos de granatges e los sieus derivats, coma de farinas e d'òlis) o indirecta (carn, lach e d'uòus que provien del bestial e las ausèls de corral que s'alimentan de pasturas o granos). Es una familha cosmopolita, qu'a conquistat la majoritat dels cavòts ecologics de la planeta, dempuèi las zònas desertiques fins als ecosistèmas d'aiga salada, e dempuèi las zònas deprimidas e anegadizas fins als sistèmas montanhuts mai nautes. Aquesta incomparable capacitat d'adaptacion es sostenuda en una enòrma diversitat morfologica, fisiologica e reproductiva e en de divèrsas associacions mutualísticas amb d'autres organismes, que convertisson a las gramineas en una fascinanta familha, pas sonque per la siá importància economica, mas tanben per la siá importància biologica.
Ang mga damo o Poaceae at Gramineae ay isang pamilya sa Klase Liliopsida (ang mga monocot) ng mga namumulaklak na halaman. Ang mga halaman na napapaloob sa pamilyang ito ay kalimitang tinatawag na damo. Ang mga mukhang palumpong o kaya puno na halaman na kabilang sa pamilyang ito ay tinatawag na kawayan (meron din namang hindi makahoy at mala-yerba na kawayan). Mayroong mga 600 na sari at mga 9000–10,000 o mas marami pang uri ng damo (Kew Index of World Grass Species). Ang mga pamayanan ng mga halaman kung saan nakakarami ang mga Poaceae ay tinatawag na damuhan o grassland; sinasabing ang mga damuhan ay bumubuo sa 20% ng balot ng halaman ng mundo. Ang mga uri ng damo ay makikita sa iba't ibang habitat gaya ng mababasang lup, gubat at tundra. Sinasabing ang Poaceae ay ang pinakamahalagang pamilya ng halaman sa ekonomiya ng tao: kasama na dito ang mga pangunahing pagkain na butil at cereal na sinasaka sa buong mundo, damo sa damuhan at mga damong pinapakain sa hayop, at kawayan, na isang mahalagang gamit sa konstruksiyon sa silangang Asya at sub-Saharan Aprika.
Ang salitang "damo" ay ginagamit rin sa (maling) pagtukoy ng mga halamang mukhang damo na hindi napapabilang sa Poaceae lineage. Kasama na dito ang Juncaceae at Cyperaceae. Tinatawag na "totoong damo" ang mga Poaceae dahil sa pagggamit ng salitang damo sa maraming uri ng halaman.
![]() Ang lathalaing ito na tungkol sa Halaman ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Ang lathalaing ito na tungkol sa Halaman ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Poacéae, asalna mah disebut Graminae, nyaéta salah sahiji kulawarga tina tutuwuhan kembangan monokotil (Monocotyledonéae), nu mangrupa hiji sumber divisi tina urutan poales.[1][2] Tutuwuhan ieu sok disebut ogé salaku jujukutan.[1] Ieu mangrupa salah sahiji di antara lima kulawarga tutuwuhan kembangan nu pangluhurna dina hal jumlah spésés, ngan ieu jelas kulawarga flora pang lobana jeung pohara pentingna di bumi.[rujukan?] Diwangun ku leuwih ti 10.000 spésiés.[3]
Poacéae tumuwuh di sakabéh benua, di padang pasir boh habitat cai tawar atawa laut, nepi ka dina tempat pangluhurna.[1] Komunitas tutuwuhan nu didominasi ku jujukutan nyumbang antara 24 persén tina vegetasi bumi.[1] Poacéae sumebar di sakabéh dunya, iwal ti Greenland jeung antartika.[1]
Poacéae ogé mangrupa sumber nu pangpentingna dina dunia dahareun.[2] Jeung mangrupa kelompok pangpentingna sacara ekonomi, nu penting téh kaasup serealia, jukut, tiwu, jeung tutuwuhan bumbu.[2] Spésiés tina ieu tutuwuhan nepi ka geus diolah pikeun jadi tutuwuhan pangan pokok (seréalia jeung gula misalna), pakan pikeun sato ingon, biofuel, nepi ka bahan wangunan, kertas jeung lansekap hias.[2]
Suku suket-suketan utawa pari-parian (Poaceae utawa Graminae) iku siji suku saka tetuwuhan keping tunggal (Monocotyledoneae).
Poaceae (bos. trave) ili Gramineae su velika i gotovo sveprisutna porodica monokotiledonskih cvjetnica, poznatih kao trave
U Poaceae se uključuju trave žitarice, bambusi i trave prirodnih i kultiviranih travnjaka i pašnjaka. Trave imaju šuplje stabljije, osim na čvorovima i uske naizmjenične listove u dva reda. Donji dio svakog lista obuhvata stabljiku, formirajući lisni omotač. Sa oko 12.000 vrsta, peti po brojnosti biljnih porodica, nakon Asteraceae, Orchidaceae, Fabaceae i Rubiaceae .
Procjenjuje se da zatravljena područja, kao što su savane i prerije, gdje su dominantne trave, čine 20% od vegetacijskog pokrov na Zemlji. Trave su također važan dio vegetacije u mnogim drugim staništima, uključujući i močvare , šume i tundre.
Trave su ekonomski izuzetno značajna porodica, jer daju životne namirnice od pripitomljenih žitarica, kao što su kukuruz, pšenica, riža, ječam i proso, kao i stočnu hranu, građevinski materijal (bambus, slama I sl.) i gorivo (etanol). Iako se obično nazivaju "trave", morska trava, rogoz i oštrice ne spadaju u ovu porodicu. Trske su red Poales, u Poaceae, kao članice, ali su morske trave članovi reda Alismatales.
Ang mga damo o Poaceae at Gramineae ay isang pamilya sa Klase Liliopsida (ang mga monocot) ng mga namumulaklak na halaman. Ang mga halaman na napapaloob sa pamilyang ito ay kalimitang tinatawag na damo. Ang mga mukhang palumpong o kaya puno na halaman na kabilang sa pamilyang ito ay tinatawag na kawayan (meron din namang hindi makahoy at mala-yerba na kawayan). Mayroong mga 600 na sari at mga 9000–10,000 o mas marami pang uri ng damo (Kew Index of World Grass Species). Ang mga pamayanan ng mga halaman kung saan nakakarami ang mga Poaceae ay tinatawag na damuhan o grassland; sinasabing ang mga damuhan ay bumubuo sa 20% ng balot ng halaman ng mundo. Ang mga uri ng damo ay makikita sa iba't ibang habitat gaya ng mababasang lup, gubat at tundra. Sinasabing ang Poaceae ay ang pinakamahalagang pamilya ng halaman sa ekonomiya ng tao: kasama na dito ang mga pangunahing pagkain na butil at cereal na sinasaka sa buong mundo, damo sa damuhan at mga damong pinapakain sa hayop, at kawayan, na isang mahalagang gamit sa konstruksiyon sa silangang Asya at sub-Saharan Aprika.
Ang salitang "damo" ay ginagamit rin sa (maling) pagtukoy ng mga halamang mukhang damo na hindi napapabilang sa Poaceae lineage. Kasama na dito ang Juncaceae at Cyperaceae. Tinatawag na "totoong damo" ang mga Poaceae dahil sa pagggamit ng salitang damo sa maraming uri ng halaman.
The Poaceae (cried Gramineae or true grasses an aw) is a lairge an nearly ubiquitous faimily o monocotyledonous flouerin plants. Wi mair nor 10,000 domesticatit an wild species, the Poaceae represents the fift-lairgest plant faimily, follaein the Orchidaceae, Asteraceae, Fabaceae, an Rubiaceae. Tho commonly cried "gresses", seagresses, rashes, an segs faw ootside this faimily. The rashes an segs is sib tae the Poaceae, bein members o the order Poales, but the seagresses is members o order Alismatales.
Gresslands is estimated tae compone 20% o the vegetation kiver o the Yird. Poaceae leeves in mony ither habitats, includin weetlands, wids, an tundra.
Domestication o poaceous cereal craps sic as maize (corn), wheat, rice, baurley, an millet lies at the foondation o sedentary leevin an ceevilization aroond the warld, an the Poaceae still constitutes the maist economically important plant faimily in modren times, providin forage, biggin materials (bamboo, thack) an fuel (ethanol), as weel as fuid.
Poacéae, asalna mah disebut Graminae, nyaéta salah sahiji kulawarga tina tutuwuhan kembangan monokotil (Monocotyledonéae), nu mangrupa hiji sumber divisi tina urutan poales. Tutuwuhan ieu sok disebut ogé salaku jujukutan. Ieu mangrupa salah sahiji di antara lima kulawarga tutuwuhan kembangan nu pangluhurna dina hal jumlah spésés, ngan ieu jelas kulawarga flora pang lobana jeung pohara pentingna di bumi.[rujukan?] Diwangun ku leuwih ti 10.000 spésiés.
Le Poaceae Barnhart (ciamàde apò Gramineae) i è ìna famìa botànica de piànte erbàcee, o de rar fés lignùze, che le aparté al ùrden de le Poales, (Liliopsida). Con de piö de 670 zèner e quàze 10.000 spéci discriìde, le Poaceae i è la quàrta famìa con piö tàta richèsa de spéci dòpo de le Asteraceae, le Orchidaceae e le Fabaceae (leguminùze); però, i è la prìma per emportànsa económica globàl. De fat, la diéta dei òm l'è furmàda per 'na gran part de derivàcc de le poaceae. En piö se se cunsìdera che apò a le bès•ce i è alimentàde suratöt co le granàie o col fé che i è ac per lo piö de la famìa de le poaceae, se capés che se tràta de 'na famìa de piànte de 'mportànsa capitàl.
L'è 'na famìa cosmopolìta, che la g'ha cunquistàt la gran part de i ambièncc ecològic dispunìbii söl nòst pianéta, de le zòne dezèrtiche 'nfìna ai ecosistémi de àiva salàda, e de le depresiù palüdùze 'nfìna ai sistémi muntagùs piö alcc. Chèsta braüra 'ndèl adatàs la g'ha dat come rezültàt 'na biodiversità enòrme che la s'è manifestàda ta endèla morfologìa, che 'ndèla fiziulugìa, che 'ndèi sistémi de reprudusiù e 'ndèle asociasiù condei óter organìsmi, isé che le graminàcee i è de cunsiderà 'na famìa 'nteresànte, m,ìa apéna per la sò 'mportànsa económica, ma apò a per la releànsa biológica.
Swetgäärs (Poaceae = Gramineae) as en grat plaantenfamile üüb a hialer eerd. Flook plaantenslacher haa en grat bedüüdang üs iidj för a minsken.
At jaft wel amanbi 10.000 slacher üüb a eerd. Heer as en ütjwool:
Swetgäärs (Poaceae = Gramineae) as en grat plaantenfamile üüb a hialer eerd. Flook plaantenslacher haa en grat bedüüdang üs iidj för a minsken.
D Süessgreeser (Poaceae = Gramineae) sin e Pflanzefamilie, wo uf dr ganze Wält und in alle Klimazone verbreitet isch. D Arte, wo drzueghööre, häi die tüpisch grasartig Form.
D Süessgreeser ghööre zu de eltiste Nutzpflanze und sin sit langem für e Mensche vo lääbeswichdiger Bedütig. Alli Gedräid wie Wäize, Rogge, Gärste, Haaber, Hirse, Mais und Riis ghööre zu dere Pflanzegrubbe. As Määrtfrücht und as Veefueter sin si d Basis für d Ernäärig vo dr Bevälkerig vo dr ganze Wält. As Gras- oder Grüenland wie Matte und Wäide, aber au Steppe und Savanne brääge si s Landschaftsbild vo wite Däil vo dr Wält. Es git öbbe 10'000 Süessgreeserarte in mee as 650 Gattige und das macht sä zu äinere vo de grösste Familie vo de Blüetepflanze.
Morfologischi Merkmol vom Halm und vo de Bletter bim Fuggsschwanz
E schematischi Daarstellig vom ene Ääri mit zwäi Blüete vom ene Süegras und e Blüetediagramm
E schematische Lengsschnitt dur e Wäizechorn
E Rispe vom Fluug-Haaber (Avena fatua)
S Wurzlesüstem vom Gwöönlige Strandhaaber
E Bambus-„Wald“ mit Phyllostachys edulis
D Süessgreeser (Poaceae = Gramineae) sin e Pflanzefamilie, wo uf dr ganze Wält und in alle Klimazone verbreitet isch. D Arte, wo drzueghööre, häi die tüpisch grasartig Form.
D Süessgreeser ghööre zu de eltiste Nutzpflanze und sin sit langem für e Mensche vo lääbeswichdiger Bedütig. Alli Gedräid wie Wäize, Rogge, Gärste, Haaber, Hirse, Mais und Riis ghööre zu dere Pflanzegrubbe. As Määrtfrücht und as Veefueter sin si d Basis für d Ernäärig vo dr Bevälkerig vo dr ganze Wält. As Gras- oder Grüenland wie Matte und Wäide, aber au Steppe und Savanne brääge si s Landschaftsbild vo wite Däil vo dr Wält. Es git öbbe 10'000 Süessgreeserarte in mee as 650 Gattige und das macht sä zu äinere vo de grösste Familie vo de Blüetepflanze.
Varpėnē augalā (luotīnėškā: Poaceae) īr vėinskiltē, žėidėnē augalā, prī katrūm prėgol vėsuokės žuolės. Anūm stėibs īr stamens, tonkē toštievėdoris, šaknis īr koukštėnės, lapā ėlgi, sodarīti ėš vėina lakšta. Žėidā palėik sosikruovė̄ i varpas aba kekės, apdolkėnami viejė.
Varpėnē augalā īr nuognē paplitė̄ vėsom svietė, prisitaika pri bikuokės aplėnkuos. Dalės varpėniu augalū īr nuognē svarbūs maistėnē augalā (javā).
Puopolēriausi varpėnē augalā:
Wiru yura rikch'aq ayllu icha Qachu yurakuna (familia Gramineae icha Poaceae) nisqaqa huk yurakunap rikch'aq ayllunmi.
Kaymi huk qachu yurakuna:
Wiru yura rikch'aq ayllu icha Qachu yurakuna (familia Gramineae icha Poaceae) nisqaqa huk yurakunap rikch'aq ayllunmi.
Æфсирджынтæ, æфсирджын хортæ[1], хоргæрдæджытæ[1][2], тыллæг[3] (лат. Poaceae) у зайæгойты бинонтæ.
Уыцы зайæгойтæ тынг ахсджиаг сты адæмæн, систы калориты сæйраг ратæдзæн. Бæстæмæ гæсгæ ахсджиагдæр сты пырындз, нартхор, мæнæу, кæнынц дзы кастæ, ссад, сæкæр. Хъæддагæй зайгæ æфсирджынтæ (кæрдæг) у хизгæ фосы сæйрагдæр хæринаг.
Æфсирджынтæ, æфсирджын хортæ, хоргæрдæджытæ, тыллæг (лат. Poaceae) у зайæгойты бинонтæ.
Уыцы зайæгойтæ тынг ахсджиаг сты адæмæн, систы калориты сæйраг ратæдзæн. Бæстæмæ гæсгæ ахсджиагдæр сты пырындз, нартхор, мæнæу, кæнынц дзы кастæ, ссад, сæкæр. Хъæддагæй зайгæ æфсирджынтæ (кæрдæг) у хизгæ фосы сæйрагдæр хæринаг.
Με την ονομασία Αγρωστώδη (Gramineae), Αγρωστίδες ή Ποοειδή (Poaceae) φέρεται μεγάλη οικογένεια φυτών της ομοταξίας των Μονοκοτυληδόνων, που περιλαμβάνει ποώδη φυτά και σπανιότερα θαμνώδη ή δενδρώδη. Συνηθέστερα έχουν ρίζωμα και βλαστό καλαμοειδή.
Πρόκειται για πολύ σπουδαία οκογένεια φυτών αφού σ΄ αυτή περιλαμβάνονται τα δημητριακά καθώς και πολλά κτηνοτροφικά. Σύμφωνα με το ταξινομικό σύστημα του Έγκελς τα Αγρωστώδη ανήκουν στη τάξη των Λεπυρανθών (Glumiflorae), με βάση το σύστημα του Ταχτατζιάν (Takhtajian) ανήκουν στη τάξη των Ποωδών (Poales), ενώ σύμφωνα με το σύστημα του Cronquist στη τάξη των Κυπειρωδών (Cyperales).
Τα αγρωστώδη αναπτύσσονται τόσο σε υγρούς τόπους όσο και σε ξηρούς από τις υπερβόρειες μέχρι και τις τροπικές περιοχές, κυρίως όμως σε λιβάδια, στέπες, σαβάνες, πάμπας καθώς και σε παράμος.
Η οικογένεια των Αγρωστωδών περιλαμβάνει περίπου 600 - 700 γένη στα οποία περιλαμβάνονται από 6.000 μέχρι 10.000 είδη. Τα αγρωστώδη κατέχουν την 5η κατά σειρά θέση σε πλήθος ειδών ανάμεσα στις οικογένειες των αγγειόσπερμων.
Η επιστήμη που έχει ως κύριο αντικείμενο μελέτης τις αγρωστίδες ονομάζεται Αγρωστολογία
Με την ονομασία Αγρωστώδη (Gramineae), Αγρωστίδες ή Ποοειδή (Poaceae) φέρεται μεγάλη οικογένεια φυτών της ομοταξίας των Μονοκοτυληδόνων, που περιλαμβάνει ποώδη φυτά και σπανιότερα θαμνώδη ή δενδρώδη. Συνηθέστερα έχουν ρίζωμα και βλαστό καλαμοειδή.
Πρόκειται για πολύ σπουδαία οκογένεια φυτών αφού σ΄ αυτή περιλαμβάνονται τα δημητριακά καθώς και πολλά κτηνοτροφικά. Σύμφωνα με το ταξινομικό σύστημα του Έγκελς τα Αγρωστώδη ανήκουν στη τάξη των Λεπυρανθών (Glumiflorae), με βάση το σύστημα του Ταχτατζιάν (Takhtajian) ανήκουν στη τάξη των Ποωδών (Poales), ενώ σύμφωνα με το σύστημα του Cronquist στη τάξη των Κυπειρωδών (Cyperales).
Τα αγρωστώδη αναπτύσσονται τόσο σε υγρούς τόπους όσο και σε ξηρούς από τις υπερβόρειες μέχρι και τις τροπικές περιοχές, κυρίως όμως σε λιβάδια, στέπες, σαβάνες, πάμπας καθώς και σε παράμος.
Η οικογένεια των Αγρωστωδών περιλαμβάνει περίπου 600 - 700 γένη στα οποία περιλαμβάνονται από 6.000 μέχρι 10.000 είδη. Τα αγρωστώδη κατέχουν την 5η κατά σειρά θέση σε πλήθος ειδών ανάμεσα στις οικογένειες των αγγειόσπερμων.
Η επιστήμη που έχει ως κύριο αντικείμενο μελέτης τις αγρωστίδες ονομάζεται Αγρωστολογία
Кыяклылар (лат. Gramíneae, рус. Зла́ки), шулай ук Йончалылар[3]рус. Мя́тликовые (лат. Poáceae) — берөлешлеләр гаиләсе. 650 ыруг, якынча 10 мең төре билгеле, бар җир шарын буйлап таралган. Үлән, сирәгрәк агач сыман үсемлек. Идел-Урал буенда 39 төрдән (колынкойрык, кылган, айрык, йонча, бүтәгә, актамыр һ.б.) 139 төр үсә. Бер, ике, күп еллык үлән. Сабагы цилиндр формасында, гади яки тармаклы, төз яки җәйелүчән; төеннәрдән һәм төен араларыннан тора. Күпчелек кыяклылар төен араларының эче куыш (салам), сирәгрәк көпшәк паренхималы тукыма белән тутырылган (керпе үлән, мәккәй, тары һ.б.). Яфрагы кыяк, ланцет, җеп яки каеш сыман, ачык, сирәк очракларда кушылып үккән җиңсәле (күстерә, шаукамыш һ.б.) һәм кыяк (җепсә сыманнарга кадәр) яки ланцет рәүешле пластинкалы. Җиңсәнең пластинкага күчкән урынында гадәттә телчә була. Яфрагының урнашуы ике рәтле, сирәкләп спираль рәвешендә. Чәчкәсе вак, ике, сирәгрәк бер җенесле, чәчкә тәңкәләре һәм 1-2 чәчкә ярысы (лодикулалар) белән уратып алынган. Серкәчләре 1 дән алып 6 га кадәр (күпчелек төрләрдә 3). Җимешлеге 1, җимшәне өске. Чәчкәләре катлаулы чәчкәлекләр ясаучы башакларда 2 рәт булып урынлашкан: себеркеләр, башаклар, суклар, солтаннар яки башлар. Йонча, колынкойрык һ.б. ыруларга караган төрләргә апомиксис (аталандырылмыйча яралгы барлыкка килү) хас, кайсы берләрендә (йонча, солыбаш) вивипария (башаклар суганбаш булып үзгәрә) күзәтелә. Җимеше — бөртекчә, хайваннар яки җил белән тарала. Безнеж шартларда кыяклылар башлыча далада һәм болыннарда үсә, шулай ук урманнарда, дымлы урыннарда һәм биек тау башларында очрый. Ашарга яраклы үсемлекләр (тары, бодай, арыш, арпа һ.б.), мал азыгы үсемлекләре (айравык, күстерә, йонча, солыбаш, тимофеевка һ.б.), декоратив үсемлекләр (тарут, арышбаш, ялбыр кылган һ.б.), баллы үсемлекләр (бармакчалы үлән, камыш), сүсле үсемлекләр (сәрхәт кыягы, елан теле һ.б.). Кайсы бер кыяклылар (үгезут, исле башак һ.б.) аракы-ликёр һәм парфюмерия-косметика сәнәгатендә, айравык, керпе үләне, дөя кылганы һ.б. халык медицинасында кулланыла. Арыш күстерәсе, кара солы, сырылучан актамыр, керпе үләненең һәм шыртүләннең төрләре — чүп үләннәре. Йонча, солыбаш, иргылчык, арыш һәм тимофеевканың серкәсе аллергия авыралары тудыра. Үсемлекләрнең 15 сирәк төре (реликтлар сөялле солы үләне, Ледебур колерия; эндемиклар Коржинский кылганы, шырт колерия; мурт арышбаш, Лессинг кылганы, сарепта кылганы, Карелин таукыягы, чуклы таукыяк, актамыр, ялтыр очлыгылчык һ.б.) — БР ныңКызыл китабына, алар арасыннан 5 төр (Залесский кылганы, сылу кылган, төкле кылган, дөя кылганы, келерия) РФ ның Кызыл китабына кертелгән.
Кыяклылар (лат. Gramíneae, рус. Зла́ки), шулай ук Йончалыларрус. Мя́тликовые (лат. Poáceae) — берөлешлеләр гаиләсе. 650 ыруг, якынча 10 мең төре билгеле, бар җир шарын буйлап таралган. Үлән, сирәгрәк агач сыман үсемлек. Идел-Урал буенда 39 төрдән (колынкойрык, кылган, айрык, йонча, бүтәгә, актамыр һ.б.) 139 төр үсә. Бер, ике, күп еллык үлән. Сабагы цилиндр формасында, гади яки тармаклы, төз яки җәйелүчән; төеннәрдән һәм төен араларыннан тора. Күпчелек кыяклылар төен араларының эче куыш (салам), сирәгрәк көпшәк паренхималы тукыма белән тутырылган (керпе үлән, мәккәй, тары һ.б.). Яфрагы кыяк, ланцет, җеп яки каеш сыман, ачык, сирәк очракларда кушылып үккән җиңсәле (күстерә, шаукамыш һ.б.) һәм кыяк (җепсә сыманнарга кадәр) яки ланцет рәүешле пластинкалы. Җиңсәнең пластинкага күчкән урынында гадәттә телчә була. Яфрагының урнашуы ике рәтле, сирәкләп спираль рәвешендә. Чәчкәсе вак, ике, сирәгрәк бер җенесле, чәчкә тәңкәләре һәм 1-2 чәчкә ярысы (лодикулалар) белән уратып алынган. Серкәчләре 1 дән алып 6 га кадәр (күпчелек төрләрдә 3). Җимешлеге 1, җимшәне өске. Чәчкәләре катлаулы чәчкәлекләр ясаучы башакларда 2 рәт булып урынлашкан: себеркеләр, башаклар, суклар, солтаннар яки башлар. Йонча, колынкойрык һ.б. ыруларга караган төрләргә апомиксис (аталандырылмыйча яралгы барлыкка килү) хас, кайсы берләрендә (йонча, солыбаш) вивипария (башаклар суганбаш булып үзгәрә) күзәтелә. Җимеше — бөртекчә, хайваннар яки җил белән тарала. Безнеж шартларда кыяклылар башлыча далада һәм болыннарда үсә, шулай ук урманнарда, дымлы урыннарда һәм биек тау башларында очрый. Ашарга яраклы үсемлекләр (тары, бодай, арыш, арпа һ.б.), мал азыгы үсемлекләре (айравык, күстерә, йонча, солыбаш, тимофеевка һ.б.), декоратив үсемлекләр (тарут, арышбаш, ялбыр кылган һ.б.), баллы үсемлекләр (бармакчалы үлән, камыш), сүсле үсемлекләр (сәрхәт кыягы, елан теле һ.б.). Кайсы бер кыяклылар (үгезут, исле башак һ.б.) аракы-ликёр һәм парфюмерия-косметика сәнәгатендә, айравык, керпе үләне, дөя кылганы һ.б. халык медицинасында кулланыла. Арыш күстерәсе, кара солы, сырылучан актамыр, керпе үләненең һәм шыртүләннең төрләре — чүп үләннәре. Йонча, солыбаш, иргылчык, арыш һәм тимофеевканың серкәсе аллергия авыралары тудыра. Үсемлекләрнең 15 сирәк төре (реликтлар сөялле солы үләне, Ледебур колерия; эндемиклар Коржинский кылганы, шырт колерия; мурт арышбаш, Лессинг кылганы, сарепта кылганы, Карелин таукыягы, чуклы таукыяк, актамыр, ялтыр очлыгылчык һ.б.) — БР ныңКызыл китабына, алар арасыннан 5 төр (Залесский кылганы, сылу кылган, төкле кылган, дөя кылганы, келерия) РФ ның Кызыл китабына кертелгән.
Метлюжковыя (Poaceae), або злакі (Gramíneae) — сямейства аднадольных расьлінаў, да якога адносяцца такія вядомыя і здаўна выкарыстаныя ў гаспадарцы расьліны як пшаніца, жыта, авёс, рыс, кукуруза, ячмень, проса, бамбук, цукровы трысьнёг.
Метлюжковыя ў прыродзе расьселеныя па ўсіх кантынэнтах, уключаючы Антарктыду. Складаюць значную частку фітамасы ў многіх біяцэнозах, а ў стэпах і саванах — галоўную частку. Сямейства ўключае каля 600 родаў і 9-10 тысячаў відаў.
У большасьці прадстаўнікоў сямейства сьцеблы простыя часам галіністыя, цыліндрычныя ці пляскатыя, падзеленыя вузламі, полыя. Лісьце лінейнае ці лянцэнтнае. Кветкі жаўтлява-зялёныя, дробныя, сабраныя ў суквецьці каласкі, якія ўтвараюць колас або пэндзаль.
![]() — сховішча мультымэдыйных матэрыялаў
— сховішча мультымэдыйных матэрыялаў
Метлюжковыя (Poaceae), або злакі (Gramíneae) — сямейства аднадольных расьлінаў, да якога адносяцца такія вядомыя і здаўна выкарыстаныя ў гаспадарцы расьліны як пшаніца, жыта, авёс, рыс, кукуруза, ячмень, проса, бамбук, цукровы трысьнёг.
Метлюжковыя ў прыродзе расьселеныя па ўсіх кантынэнтах, уключаючы Антарктыду. Складаюць значную частку фітамасы ў многіх біяцэнозах, а ў стэпах і саванах — галоўную частку. Сямейства ўключае каля 600 родаў і 9-10 тысячаў відаў.
У большасьці прадстаўнікоў сямейства сьцеблы простыя часам галіністыя, цыліндрычныя ці пляскатыя, падзеленыя вузламі, полыя. Лісьце лінейнае ці лянцэнтнае. Кветкі жаўтлява-зялёныя, дробныя, сабраныя ў суквецьці каласкі, якія ўтвараюць колас або пэндзаль.
Тревите (науч. Poaceae) се фамилија на монокотиледонски растенија опфатени со научното име Poaceae или поранешното Gramineae. Ова е голема група на скриеносемени растенија кои се многу важни од економски и еколошки аспект. Фамилијата брои околу 635 родови и 9000 видови, правејќи ја четвртата најголема фамилија на растенија, по Fabaceae, Orchidaceae и Asteraceae.
Сите житарици припаѓаат на оваа фамилија. Најпознати култивирани растенија се оризот, пченицата, пченката и јачменот. Од шеќерната трска се добива најголемиот дел од светското производство на шеќер, додека од бамбусот се добиваат важни градежни материјали, а се користи и за производство на хартија и храна. Тревите се примарниот извор на храна за човекот, домашните и дивите животни.
Тревите се најшироко распространетите цветни растенија, кои можат да се сретнат од Арктикот, преку умерените и тропските региони, па до Антарктик. Тревите се единствените цветни растенија кои растат на Антарктикот. Овие растенија се најчести во отворените станишта, како што се прериите, тундрите, степите и саваните, но ги има и во шумите (особено во тропските области). Одделни видови се водни растенија, а некои се среќаваат и во пустините.
Тревите варираат во големината од тропските џиновски бамбуси (високи и повеќе од 100 m), до едногодишните ливадски треви високи само неколку сантиметри. Некои претставници немаат сабјести листови, но доколку постојат, тие можат да достигнат должина од 5 m.
Тревите се многу униформни по својата основна вегетативна структура, така што претставниците делат повеќе особености. Корењата се најчесто фиброзни, а постојат и дополнителни (адвентивни) корења, што може јасно да се забележи кај пченката. Стеблата се најчесто тревести (ливадски треви) или шупливи (бамбус), но постојат и исклучоци, како што се исполнетите со паренхим стебла на пченката или дрвенестите стебла на некои бамбуси.
Листовите се развиваат од нодиумите (членчињата) на стеблото и се поредени во два реда. Се состојат од долга и тесна ламина (лиска), како и од листен ракавец кој го обвива стебленцето. Лисниот ракавец дава потпора на регионот над нодиумот, бидејќи тој е изграден од меко ткиво наречено творно ткиво (меристем), чии клетки се делат и така растението расте во височина (за разлика од повеќето други растенија кои се елонгираат при врвот на стеблата и листовите). Значи, иако се пресече врвот на листот од тревата, тој повторно ќе биде во можност да расте, што е мошне значајно при косењето на тревата. Друга значајна особина на тревите е присуството на јазиче (ligula), што претставува кратка влакнеста или мембранозна проекција на точката каде лисниот ракавец се спојува со лисната ламина. Функцијата на лигулата сѐ уште не е позната, но се верува дека таа спречува навлегување на влага во регионот помеѓу лиската и стеблото.
Цветовите на тревите се неугледни и/или незабележливи, но тие најчесто се групираат во големи соцветија. На пример, кочанот кај пченката е всушност женското соцветие, додека метличката е машкото соцветие (пченката е еднодомно растение). Повеќето треви се опрашуваат со помош на ветрот (анемофилно), така што нивните цветови се силно редуцирани и многу прости и со невпечатливи бои (за разлика од растенијата кои се опрашуваат со помош на инсекти, кои се впечатливо обоени за да го привлечат самиот инсект).
Секој индивидуален цвет на тревите е со мала големина и кај него отсуствува околуцветник или перигон (чашкини и венечни ливчиња). Наместо тоа, тој поседува две ситни лушпи наречени lodiculi кои се наоѓаат во неговата основа. Над лодикулите се развиваат по 3 прашника (или 6 кај оризот). Единствениот плодник е натцветен и со две пердувести устенца. Тој созрева во одделен, едносеменски плод наречен кариопсис (caryopsis), кој се одликува со тоа што при неговото образување, семето се фузира со ѕидот на јајчникот (плодникот).
Цветовите кај тревите се групирани во мали класчиња. Секое класче се состои од оска наречена rachilla околу која се образуваат индивидуалните цветови. Секој цвет е затворен со две лушпести прицветници означени како lemma (надворешен прицветник) и palea (внатрешен прицветник). Во основата на рахилата се наоѓаат две лушпи наречени glumae (еднина gluma) кои се празни (не содржат цветови). Палеата е најчесто нежна и мембранозна, за разлика од глумите и лемите кои се груби по конзистенција. Постојат големи разновидности во деталната градба на цветовите на тревите, што е причина за големиот број на видови.
BEB-клад (разгранок)
PACC-клад (разгранок)
Соцветија на шеќерната трска (Saccharum officinarum), од Panicoideae.
Бамбусот (Bambusa oldhamii) припаѓа на фамилијата на треви.
Машкиот цвет на пченката (метличка).
Тревите (науч. Poaceae) се фамилија на монокотиледонски растенија опфатени со научното име Poaceae или поранешното Gramineae. Ова е голема група на скриеносемени растенија кои се многу важни од економски и еколошки аспект. Фамилијата брои околу 635 родови и 9000 видови, правејќи ја четвртата најголема фамилија на растенија, по Fabaceae, Orchidaceae и Asteraceae.
Сите житарици припаѓаат на оваа фамилија. Најпознати култивирани растенија се оризот, пченицата, пченката и јачменот. Од шеќерната трска се добива најголемиот дел од светското производство на шеќер, додека од бамбусот се добиваат важни градежни материјали, а се користи и за производство на хартија и храна. Тревите се примарниот извор на храна за човекот, домашните и дивите животни.
Тревите се најшироко распространетите цветни растенија, кои можат да се сретнат од Арктикот, преку умерените и тропските региони, па до Антарктик. Тревите се единствените цветни растенија кои растат на Антарктикот. Овие растенија се најчести во отворените станишта, како што се прериите, тундрите, степите и саваните, но ги има и во шумите (особено во тропските области). Одделни видови се водни растенија, а некои се среќаваат и во пустините.
பொவேசி (Poaceae,/poʊˈeɪsiaɪ/) அல்லது கிராமினே (Gramineae) ஒருவித்திலை பூக்கும் தாவரக் குடும்பத்தைச் சேர்ந்த பரந்த, எங்கும் காணுகின்ற, பொதுவாக புல்லினம் என அறியப்படும் தாவரங்களாகும். அரிசி, கோதுமை போன்ற தானியங்கள் இயற்கையான புல்வெளிகளிலும் செயற்கை புல்தரைகளிலும் காணப்படும் புல், மற்றும் மூங்கில் போன்ற உலகளவில் பொருளியல் முதன்மை வாய்ந்த்தாக இக்குடும்பம் விளங்குகின்றது. புல்லினங்களின் தண்டு கணுக்களில் தவிர, வெறுமையாகவும் குறுகிய மாறிவரும் இரண்டடுக்கு இலைகளையும் கொண்டுள்ளன. கீழுள்ள இலை தண்டைச் சுற்றிக்கொண்டு இலைமடலை உருவாக்குகின்றது. 780 பேரினங்களையும் 12000 இனங்களையும் கொண்டுள்ள புல்லினங்கள்[3] சூரியகாந்தி, ஆர்க்கிட், பபேசியே, காஃபி குடும்பங்களை அடுத்து ஐந்தாவது மிகப்பெரியத் தாவரக் குடும்பமாக பொவேசி விளங்குகிறது.[4] புல்லின் கிரேக்கப் பெயரான பொவே என்பதிலிருந்து இந்த தாவர இனம் பெயரிடப்பட்டுள்ளது.

Leaves of Poa trivialis showing the ligules
Bamboo stem and leaves, nodes are evident
A Chasmanthium latifolium spikelet
Wheat spike and spikelet
Spikelet opened to show caryopsis
Harestail grass
Grass
Sugarcane (Saccharum officinarum)
Roots of Bromus hordeaceus
Barley mature spikes (வாற்கோதுமை)
Illustration depicting both staminate and pistillate flowers of maize (மக்காச்சோளம்)
A grass flower head (meadow foxtail) showing the plain-coloured flowers with large anthers.
Anthers detached from a meadow foxtail flower
Setaria verticillata, bristly foxtail
Setaria verticillata, bristly foxtail
பொவேசி (Poaceae,/poʊˈeɪsiaɪ/) அல்லது கிராமினே (Gramineae) ஒருவித்திலை பூக்கும் தாவரக் குடும்பத்தைச் சேர்ந்த பரந்த, எங்கும் காணுகின்ற, பொதுவாக புல்லினம் என அறியப்படும் தாவரங்களாகும். அரிசி, கோதுமை போன்ற தானியங்கள் இயற்கையான புல்வெளிகளிலும் செயற்கை புல்தரைகளிலும் காணப்படும் புல், மற்றும் மூங்கில் போன்ற உலகளவில் பொருளியல் முதன்மை வாய்ந்த்தாக இக்குடும்பம் விளங்குகின்றது. புல்லினங்களின் தண்டு கணுக்களில் தவிர, வெறுமையாகவும் குறுகிய மாறிவரும் இரண்டடுக்கு இலைகளையும் கொண்டுள்ளன. கீழுள்ள இலை தண்டைச் சுற்றிக்கொண்டு இலைமடலை உருவாக்குகின்றது. 780 பேரினங்களையும் 12000 இனங்களையும் கொண்டுள்ள புல்லினங்கள் சூரியகாந்தி, ஆர்க்கிட், பபேசியே, காஃபி குடும்பங்களை அடுத்து ஐந்தாவது மிகப்பெரியத் தாவரக் குடும்பமாக பொவேசி விளங்குகிறது. புல்லின் கிரேக்கப் பெயரான பொவே என்பதிலிருந்து இந்த தாவர இனம் பெயரிடப்பட்டுள்ளது.
పోయేసి లేదా గ్రామినే (లాటిన్ Poaceae, Gramineae) పుష్పించే మొక్కలలో ఒక పెద్ద కుటుంబం. దీనిలో 620 ప్రజాతులు, 10,000 జాతులు ఉన్నాయి. ఇవి అన్ని ప్రాంతాలలోను, అన్ని రకాల నేలలలోను పెరుగుతాయి.
ಪೊಯೇಸಿಯಿ ಹುಲ್ಲುಗಳು ಎಂದು ಕರೆಯಲ್ಪಡುವ ಏಕದಳ ಹೂಬಿಡುವ ಸಸ್ಯಗಳ ಒಂದು ದೊಡ್ಡದಾದ ಮತ್ತು ಬಹುತೇಕ ಸರ್ವವ್ಯಾಪಿ ಕುಟುಂಬವಾಗಿದೆ. ಪೊಯೇಸಿಯಿ ಕುಟುಂಬದಲ್ಲಿ ಧಾನ್ಯ ಹುಲ್ಲುಗಳು, ಬಿದಿರುಗಳು ಮತ್ತು ನೈಸರ್ಗಿಕ ಹುಲ್ಲುಭೂಮಿ ಹಾಗೂ ಕೃಷಿಮಾಡಲಾದ ಹುಲ್ಲುಮೈದಾನಗಳು ಹಾಗೂ ಗೋಮಾಳದ ಹುಲ್ಲುಗಳು ಸೇರಿವೆ. ಹುಲ್ಲುಗಳು ಗೆಣ್ಣುಗಳ ಹೊರತಾಗಿ ಬೇರೆಡೆಗಳಲ್ಲಿ ಪೊಳ್ಳಾಗಿರುವ ಕಾಂಡಗಳು ಮತ್ತು ಎರಡು ದರ್ಜೆಗಳಲ್ಲಿ ಹುಟ್ಟಿರುವ ಕಿರಿದಾದ ಪರ್ಯಾಯ ಎಲೆಗಳನ್ನು ಹೊಂದಿರುತ್ತವೆ. ಪ್ರತಿ ಎಲೆಯ ಕೆಳಗಿನ ಭಾಗವು ಕಾಂಡವನ್ನು ಆವರಿಸಿರುತ್ತದೆ, ಮತ್ತು ಎಲೆ ಕೋಶವನ್ನು ರೂಪಿಸುತ್ತದೆ. ಈ ಕುಟುಂಬದಲ್ಲಿ ಸುಮಾರು ೭೮೦ ಕುಲಗಳು ಮತ್ತು ಸುಮಾರು ೧೨,೦೦೦ ಪ್ರಜಾತಿಗಳಿವೆ,[೧] ಮತ್ತು ಐದನೇ ಅತಿ ದೊಡ್ಡ ಸಸ್ಯ ಕುಟುಂಬವೆನಿಸಿಕೊಂಡಿದೆ.
|deadurl= (help) ಪೊಯೇಸಿಯಿ ಹುಲ್ಲುಗಳು ಎಂದು ಕರೆಯಲ್ಪಡುವ ಏಕದಳ ಹೂಬಿಡುವ ಸಸ್ಯಗಳ ಒಂದು ದೊಡ್ಡದಾದ ಮತ್ತು ಬಹುತೇಕ ಸರ್ವವ್ಯಾಪಿ ಕುಟುಂಬವಾಗಿದೆ. ಪೊಯೇಸಿಯಿ ಕುಟುಂಬದಲ್ಲಿ ಧಾನ್ಯ ಹುಲ್ಲುಗಳು, ಬಿದಿರುಗಳು ಮತ್ತು ನೈಸರ್ಗಿಕ ಹುಲ್ಲುಭೂಮಿ ಹಾಗೂ ಕೃಷಿಮಾಡಲಾದ ಹುಲ್ಲುಮೈದಾನಗಳು ಹಾಗೂ ಗೋಮಾಳದ ಹುಲ್ಲುಗಳು ಸೇರಿವೆ. ಹುಲ್ಲುಗಳು ಗೆಣ್ಣುಗಳ ಹೊರತಾಗಿ ಬೇರೆಡೆಗಳಲ್ಲಿ ಪೊಳ್ಳಾಗಿರುವ ಕಾಂಡಗಳು ಮತ್ತು ಎರಡು ದರ್ಜೆಗಳಲ್ಲಿ ಹುಟ್ಟಿರುವ ಕಿರಿದಾದ ಪರ್ಯಾಯ ಎಲೆಗಳನ್ನು ಹೊಂದಿರುತ್ತವೆ. ಪ್ರತಿ ಎಲೆಯ ಕೆಳಗಿನ ಭಾಗವು ಕಾಂಡವನ್ನು ಆವರಿಸಿರುತ್ತದೆ, ಮತ್ತು ಎಲೆ ಕೋಶವನ್ನು ರೂಪಿಸುತ್ತದೆ. ಈ ಕುಟುಂಬದಲ್ಲಿ ಸುಮಾರು ೭೮೦ ಕುಲಗಳು ಮತ್ತು ಸುಮಾರು ೧೨,೦೦೦ ಪ್ರಜಾತಿಗಳಿವೆ, ಮತ್ತು ಐದನೇ ಅತಿ ದೊಡ್ಡ ಸಸ್ಯ ಕುಟುಂಬವೆನಿಸಿಕೊಂಡಿದೆ.
Gors is 'n plant die gebruukt wordt om tuunen mee te verfraoiiene. 't Wor oak gebruukt as beneaming voor begroeide slikken (gorzen).
Gors is òk 'n belangriek voediengsmiddel vò graezers, zoas paerd'n en koei'n.
Poaceae (/poʊˈeɪsiaɪ, -siːiː/) or Gramineae (/ɡrəˈmɪniaɪ/) is a large and nearly ubiquitous family of monocotyledonous flowering plants commonly known as grasses. It includes the cereal grasses, bamboos and the grasses of natural grassland and species cultivated in lawns and pasture. The latter are commonly referred to collectively as grass.
With around 780 genera and around 12,000 species,[4] the Poaceae is the fifth-largest plant family, following the Asteraceae, Orchidaceae, Fabaceae and Rubiaceae.[5]
The Poaceae are the most economically important plant family, providing staple foods from domesticated cereal crops such as maize, wheat, rice, barley, and millet as well as feed for meat-producing animals. They provide, through direct human consumption, just over one-half (51%) of all dietary energy; rice provides 20%,[6] wheat supplies 20%, maize (corn) 5.5%, and other grains 6%. Some members of the Poaceae are used as building materials (bamboo, thatch, and straw); others can provide a source of biofuel, primarily via the conversion of maize to ethanol.
Grasses have stems that are hollow except at the nodes and narrow alternate leaves borne in two ranks. The lower part of each leaf encloses the stem, forming a leaf-sheath. The leaf grows from the base of the blade, an adaptation allowing it to cope with frequent grazing.
Grasslands such as savannah and prairie where grasses are dominant are estimated to constitute 40.5% of the land area of the Earth, excluding Greenland and Antarctica.[7] Grasses are also an important part of the vegetation in many other habitats, including wetlands, forests and tundra.
Though they are commonly called "grasses", groups such as the seagrasses, rushes and sedges fall outside this family. The rushes and sedges are related to the Poaceae, being members of the order Poales, but the seagrasses are members of order Alismatales. However, all of them belong to the monocot group of plants.

Grasses may be annual or perennial herbs,[8]: 10 generally with the following characteristics (the image gallery can be used for reference): The stems of grasses, called culms, are usually cylindrical (more rarely flattened, but not 3-angled) and are hollow, plugged at the nodes, where the leaves are attached.[8][9] Grass leaves are nearly always alternate and distichous (in one plane), and have parallel veins.[8]: 11 Each leaf is differentiated into a lower sheath hugging the stem and a blade with entire (i.e., smooth) margins.[8]: 11 The leaf blades of many grasses are hardened with silica phytoliths, which discourage grazing animals; some, such as sword grass, are sharp enough to cut human skin. A membranous appendage or fringe of hairs called the ligule lies at the junction between sheath and blade, preventing water or insects from penetrating into the sheath.[8]: 11
Flowers of Poaceae are characteristically arranged in spikelets, each having one or more florets.[8]: 12 The spikelets are further grouped into panicles or spikes. The part of the spikelet that bears the florets is called the rachilla. A spikelet consists of two (or sometimes fewer) bracts at the base, called glumes, followed by one or more florets.[8]: 13 A floret consists of the flower surrounded by two bracts, one external—the lemma—and one internal—the palea. The flowers are usually hermaphroditic—maize being an important exception—and mainly anemophilous or wind-pollinated, although insects occasionally play a role.[10] The perianth is reduced to two scales, called lodicules,[8]: 11 that expand and contract to spread the lemma and palea; these are generally interpreted to be modified sepals. The fruit of grasses is a caryopsis, in which the seed coat is fused to the fruit wall.[8]: 16 A tiller is a leafy shoot other than the first shoot produced from the seed.[8]: 11
Grass blades grow at the base of the blade and not from elongated stem tips. This low growth point evolved in response to grazing animals and allows grasses to be grazed or mown regularly without severe damage to the plant.[11]: 113–114
Three general classifications of growth habit present in grasses: bunch-type (also called caespitose), stoloniferous, and rhizomatous.[12] The success of the grasses lies in part in their morphology and growth processes and in part in their physiological diversity. There are both C3 and C4 grasses, referring to the photosynthetic pathway for carbon fixation. The C4 grasses have a photosynthetic pathway, linked to specialized Kranz leaf anatomy, which allows for increased water use efficiency, rendering them better adapted to hot, arid environments.[13]
The C3 grasses are referred to as "cool-season" grasses, while the C4 plants are considered "warm-season" grasses.[8]: 18–19
Although the C4 species are all in the PACMAD clade (see diagram above), it seems that various forms of C4 have arisen some twenty or more times, in various subfamilies or genera. In the Aristida genus for example, one species (A. longifolia) is C3 but the approximately 300 other species are C4. As another example, the whole tribe of Andropogoneae, which includes maize, sorghum, sugar cane, "Job's tears", and bluestem grasses, is C4.[14] Around 46 percent of grass species are C4 plants.[15]
The name Poaceae was given by John Hendley Barnhart in 1895,[16]: 7 based on the tribe Poeae described in 1814 by Robert Brown, and the type genus Poa described in 1753 by Carl Linnaeus. The term is derived from the Ancient Greek πόα (póa, "fodder").
Grasses include some of the most versatile plant life-forms. They became widespread toward the end of the Cretaceous period, and fossilized dinosaur dung (coprolites) have been found containing phytoliths of a variety that include grasses that are related to modern rice and bamboo.[17] Grasses have adapted to conditions in lush rain forests, dry deserts, cold mountains and even intertidal habitats, and are currently the most widespread plant type; grass is a valuable source of food and energy for all sorts of wildlife.
A cladogram shows subfamilies and approximate species numbers in brackets:[14]
PACMAD cladeChloridoideae (1600)
Danthonioideae (300)
Micrairoideae (200)
Arundinoideae (50)
Panicoideae (3250)
Aristidoideae (350)
BOP cladeOryzoideae (110)
Bambusoideae – bamboos (1450)
Pooideae (3850)
Puelioideae (11)
Pharoideae (13)
Anomochlooideae (4)

Before 2005, fossil findings indicated that grasses evolved around 55 million years ago. Finds of grass-like phytoliths in Cretaceous dinosaur coprolites from the latest Cretaceous (Maastrichtian) aged Lameta Formation of India have pushed this date back to 66 million years ago.[18][19] In 2011, fossils from the same deposit were found to belong to the modern rice tribe Oryzeae, suggesting substantial diversification of major lineages by this time.[20]
Wu, You & Li (2018) described grass microfossils extracted from the teeth of the hadrosauroid dinosaur Equijubus normani from northern China, dating to the Albian stage of the Early Cretaceous approximately 113–100 million years ago, which were found to belong to primitive lineages within Poaceae, similar in position to the Anomochlooideae. These are currently the oldest known grass fossils.[1]
The relationships among the three subfamilies Bambusoideae, Oryzoideae and Pooideae in the BOP clade have been resolved: Bambusoideae and Pooideae are more closely related to each other than to Oryzoideae.[21] This separation occurred within the relatively short time span of about 4 million years.
According to Lester Charles King the spread of grasses in the Late Cenozoic would have changed patterns of hillslope evolution favouring slopes that are convex upslope and concave downslope and lacking a free face were common. King argued that this was the result of more slowly acting surface wash caused by carpets of grass which in turn would have resulted in relatively more soil creep.[22][23]
There are about 12,000 grass species in about 771 genera that are classified into 12 subfamilies.[24] See the full list of Poaceae genera.
The grass family is one of the most widely distributed and abundant groups of plants on Earth. Grasses are found on every continent,[25][26] including Antarctica. The Antarctic hair grass, Deschampsia antarctica is one of only two plant species native to the western Antarctic Peninsula.

Grasses are the dominant vegetation in many habitats, including grassland, salt-marsh, reedswamp and steppes. They also occur as a smaller part of the vegetation in almost every other terrestrial habitat. Grass-dominated biomes are called grasslands. If only large, contiguous areas of grasslands are counted, these biomes cover 31% of the planet's land.[27] Grasslands include pampas, steppes, and prairies.[28] Grasses provide food to many grazing mammals,[29] as well as to many species of butterflies and moths.[30][31] Many types of animals eat grass as their main source of food, and are called graminivores – these include cattle, sheep, horses, rabbits and many invertebrates, such as grasshoppers and the caterpillars of many brown butterflies. Grasses are also eaten by omnivorous or even occasionally by primarily carnivorous animals.

Grasses dominate certain biomes, especially temperate grasslands, because many species are adapted to grazing and fire.[32]
Grasses are unusual in that the meristem is near the bottom of the plant; hence, grasses can quickly recover from cropping at the top.[33] The evolution of large grazing animals in the Cenozoic contributed to the spread of grasses. Without large grazers, fire-cleared areas are quickly colonized by grasses, and with enough rain, tree seedlings. Trees eventually outcompete most grasses. Trampling grazers kill seedling trees but not grasses.[11]: 137
Grasses are, in human terms, perhaps the most economically important plant family.[34] Their economic importance stems from several areas, including food production, industry, and lawns. They have been grown as food for domesticated animals for up to 6,000 years and the grains of grasses such as wheat, rice, maize (corn) and barley have been the most important human food crops. Grasses are also used in the manufacture of thatch, paper, fuel, clothing, insulation, timber for fencing, furniture, scaffolding and construction materials, floor matting, sports turf and baskets.

Of all crops grown, 70% are grasses.[35] Agricultural grasses grown for their edible seeds are called cereals or grains (although the latter term, when used agriculturally, refers to both cereals and similar seeds of other plant species, such as buckwheat and legumes). Three cereals—rice, wheat, and maize (corn)—provide more than half of all calories consumed by humans.[36] Cereals constitute the major source of carbohydrates for humans and perhaps the major source of protein; these include rice (in southern and eastern Asia), maize (in Central and South America), and wheat and barley (in Europe, northern Asia and the Americas).
Sugarcane is the major source of sugar production. Additional food uses of sugarcane include sprouted grain, shoots, and rhizomes, and in drink they include sugarcane juice and plant milk, as well as rum, beer, whisky, and vodka.
Bamboo shoots are used in numerous Asian dishes and broths, and are available in supermarkets in various sliced forms, in both fresh, fermented and canned versions.
Lemongrass is a grass used as a culinary herb for its citrus-like flavor and scent.
Many species of grass are grown as pasture for foraging or as fodder for prescribed livestock feeds, particularly in the case of cattle, horses, and sheep. Such grasses may be cut and stored for later feeding, especially for the winter, in the form of bales of hay or straw, or in silos as silage. Straw (and sometimes hay) may also be used as bedding for animals.
An example of a sod-forming perennial grass used in agriculture is Thinopyrum intermedium.
Grasses are used as raw material for a multitude of purposes, including construction and in the composition of building materials such as cob, for insulation, in the manufacture of paper and board such as oriented structural straw board. Grass fiber can be used for making paper, biofuel production,[37] nonwoven fabrics, and as replacement for glass fibers used in reinforced plastics.[38] Bamboo scaffolding is able to withstand typhoon-force winds that would break steel scaffolding.[27] Larger bamboos and Arundo donax have stout culms that can be used in a manner similar to timber, Arundo is used to make reeds for woodwind instruments, and bamboo is used for innumerable implements.[39]
Phragmites australis (common reed) is important for thatching and wall construction of homes in Africa.[40] Grasses are used in water treatment systems,[41] in wetland conservation and land reclamation, and used to lessen the erosional impact of urban storm water runoff.[42]
Grasses are the primary plant used in lawns, which themselves derive from grazed grasslands in Europe. They also provide an important means of erosion control (e.g., along roadsides), especially on sloping land. Grass lawns are an important covering of playing surfaces in many sports, including football (soccer), American football, tennis, golf, cricket, softball and baseball.
Ornamental grasses, such as perennial bunch grasses, are used in many styles of garden design for their foliage, inflorescences, seed heads. They are often used in natural landscaping, xeriscaping and slope and beach stabilization in contemporary landscaping, wildlife gardening, and native plant gardening. They are used as screens and hedges.[43]
Grass playing fields, courses and pitches are the traditional playing surfaces for many sports, including American football, association football, baseball, cricket, golf, and rugby. Grass surfaces are also sometimes used for horse racing and tennis. Type of maintenance and species of grass used may be important factors for some sports, less critical for others. In some sports facilities, including indoor domes and other places where maintenance of a grass field would be difficult, grass may be replaced with artificial turf, a synthetic grass-like substitute.[44]

In cricket, the pitch is the strip of carefully mowed and rolled grass where the bowler bowls. In the days leading up to the match it is repeatedly mowed and rolled to produce a very hard, flat surface for the ball to bounce off.[45]
Grass on golf courses is kept in three distinct conditions: that of the rough, the fairway, and the putting green. Grass on the fairway is mown short and even, allowing the player to strike the ball cleanly. Playing from the rough is a disadvantage because the long grass may affect the flight of the ball. Grass on the putting green is the shortest and most even, ideally allowing the ball to roll smoothly over the surface. An entire industry revolves around the development and marketing of turf grass varieties.[46]
In tennis, grass is grown on very hard-packed soil, and the bounce of a tennis ball may vary depending on the grass's health, how recently it has been mowed, and the wear and tear of recent play. The surface is softer than hard courts and clay (other tennis surfaces), so the ball bounces lower, and players must reach the ball faster resulting in a different style of play which may suit some players more than others. Among the world's most prestigious court for grass tennis is Centre Court at Wimbledon, London which hosts the final of the annual Wimbledon Championships in England, one of the four Grand Slam tournaments.
A number of grasses are invasive species that damage natural ecosystems, including forms of Phragmites australis which are native to Eurasia but has spread around the world.[47][48]

Grasses have long had significance in human society. They have been cultivated as feed for people and domesticated animals for thousands of years. The primary ingredient of beer is usually barley or wheat, both of which have been used for this purpose for over 4,000 years.[49]
In some places, particularly in suburban areas, the maintenance of a grass lawn is a sign of a homeowner's responsibility to the overall appearance of their neighborhood. One work credits lawn maintenance to:
...the desire for upward mobility and its manifestation in the lawn. As Virginia Jenkins, author of The Lawn, put it quite bluntly, 'Upper middle-class Americans emulated aristocratic society with their own small, semi-rural estates.' In general, the lawn was one of the primary selling points of these new suburban homes, as it shifted social class designations from the equity and ubiquity of urban homes connected to the streets with the upper-middle class designation of a "healthy" green space and the status symbol that is the front lawn.[50][51]
In communities with drought problems, watering of lawns may be restricted to certain times of day or days of the week.[52] Many US municipalities and homeowners' associations have rules which require lawns to be maintained to certain specifications, sanctioning those who allow the grass to grow too long.[53][54]
The smell of the freshly cut grass is produced mainly by cis-3-Hexenal.[55]
Some common aphorisms involve grass. For example:
A folk myth about grass is that it refuses to grow where any violent death has occurred.[56]

Leaves of Poa trivialis showing the ligules

Bamboo stem and leaves, nodes are evident

A Chasmanthium latifolium spikelet

Wheat spike and spikelet

Spikelet opened to show caryopsis

Harestail grass

Grass

Sugarcane (Saccharum officinarum)

Roots of Bromus hordeaceus

Barley mature spikes (Hordeum vulgare)

Illustration depicting both staminate and pistillate flowers of maize (Zea mays)

A grass flower head (meadow foxtail) showing the plain-coloured flowers with large anthers

Anthers detached from a meadow foxtail flower

Setaria verticillata, bristly foxtail

Setaria verticillata, bristly foxtail
Poaceae (/poʊˈeɪsiaɪ, -siːiː/) or Gramineae (/ɡrəˈmɪniaɪ/) is a large and nearly ubiquitous family of monocotyledonous flowering plants commonly known as grasses. It includes the cereal grasses, bamboos and the grasses of natural grassland and species cultivated in lawns and pasture. The latter are commonly referred to collectively as grass.
With around 780 genera and around 12,000 species, the Poaceae is the fifth-largest plant family, following the Asteraceae, Orchidaceae, Fabaceae and Rubiaceae.
The Poaceae are the most economically important plant family, providing staple foods from domesticated cereal crops such as maize, wheat, rice, barley, and millet as well as feed for meat-producing animals. They provide, through direct human consumption, just over one-half (51%) of all dietary energy; rice provides 20%, wheat supplies 20%, maize (corn) 5.5%, and other grains 6%. Some members of the Poaceae are used as building materials (bamboo, thatch, and straw); others can provide a source of biofuel, primarily via the conversion of maize to ethanol.
Grasses have stems that are hollow except at the nodes and narrow alternate leaves borne in two ranks. The lower part of each leaf encloses the stem, forming a leaf-sheath. The leaf grows from the base of the blade, an adaptation allowing it to cope with frequent grazing.
Grasslands such as savannah and prairie where grasses are dominant are estimated to constitute 40.5% of the land area of the Earth, excluding Greenland and Antarctica. Grasses are also an important part of the vegetation in many other habitats, including wetlands, forests and tundra.
Though they are commonly called "grasses", groups such as the seagrasses, rushes and sedges fall outside this family. The rushes and sedges are related to the Poaceae, being members of the order Poales, but the seagrasses are members of order Alismatales. However, all of them belong to the monocot group of plants.
La poacoj aŭ veraj herboj (Poaceae, pli frue konataj kiel Graminae) estas unukotiledonaj plantoj (klaso Liliopsida). En la familio estas ĉirkaŭ 600 genroj kaj 10 000 specioj. Tiuj plantoj ampleksas ĉirkaŭ 20% de la tera vegetaĵo. Tiu planta familio estas la plej grava por la homa ekonomio, vivo, entenas gazonajn kaj furaĝajn herbojn, la ĉefaj grenoj kreskas ĉie en la mondo. La bambuo en Azio ofte estas uzata por konstruado.
La agrikulturaj poacoj kulturataj por produktado de nutraĵoj nomiĝas cerealoj. Cerealoj signifas la ĉefan kaloriofonton por la homaro, ekzemple rizo en Barato kaj la Orienta Azio, maizo en Meksiko, kaj tritiko kaj hordeo en Eŭropo kaj Nord-Ameriko.
La familio aperis antaŭ proksimume 95 milionoj da jaroj en Gondvano.
La familio enhavas ĉirkaŭ 11 000 speciojn en ĉirkaŭ 700 genroj. Estas 13 subfamiloj kaj 2 ĉefaj kladoj (BEP kaj PACCMAD).
PoaceaeLa gresoj ĝenerale havas la sekvajn trajtojn:
Estas po du glumoj por unu spiketo, la malsupra (ĝenerale pli granda) kaj la supra. Estas po du glumetoj por unu floro. La malsupra, kiu estas pli eta estas ankaŭ nomita paleo.
Ene klado PACCMAD, evoluis plurfoje vojoj de C4-tipa fotosintezo. [1]
La poacoj aŭ veraj herboj (Poaceae, pli frue konataj kiel Graminae) estas unukotiledonaj plantoj (klaso Liliopsida). En la familio estas ĉirkaŭ 600 genroj kaj 10 000 specioj. Tiuj plantoj ampleksas ĉirkaŭ 20% de la tera vegetaĵo. Tiu planta familio estas la plej grava por la homa ekonomio, vivo, entenas gazonajn kaj furaĝajn herbojn, la ĉefaj grenoj kreskas ĉie en la mondo. La bambuo en Azio ofte estas uzata por konstruado.
La agrikulturaj poacoj kulturataj por produktado de nutraĵoj nomiĝas cerealoj. Cerealoj signifas la ĉefan kaloriofonton por la homaro, ekzemple rizo en Barato kaj la Orienta Azio, maizo en Meksiko, kaj tritiko kaj hordeo en Eŭropo kaj Nord-Ameriko.
Las poáceas (Poaceae) o gramíneas son una familia de plantas herbáceas, o muy raramente leñosas, perteneciente al orden Poales de las monocotiledóneas. Con más de 820 géneros y cerca de 12 100 especies descritas,[2] las gramíneas son la cuarta familia con mayor riqueza de especies luego de las compuestas, las orquídeas y las leguminosas; pero, definitivamente, es la primera en importancia económica mundial.[3] De hecho, la mayor parte de la dieta de los seres humanos proviene de las gramíneas, tanto en forma directa —granos de cereales y sus derivados, como harinas y aceites— o indirecta —carne, leche y huevos que provienen del ganado y las aves de corral que se alimentan de pastos o granos—. Es una familia cosmopolita, que ha conquistado la mayoría de los nichos ecológicos del planeta, desde las zonas desérticas hasta los ecosistemas de agua salada, y desde las zonas deprimidas y anegadizas hasta los sistemas montañosos más altos. Esta incomparable capacidad de adaptación está sustentada en una enorme diversidad morfológica, fisiológica y reproductiva y en varias asociaciones mutualísticas con otros organismos, que convierten a las gramíneas en una fascinante familia, no solo por su importancia económica, sino también por su relevancia biológica.
Entre las especies más destacadas están la caña de azúcar, el trigo, el arroz, el maíz, el sorgo, la cebada, la avena, el centeno o el bambú.
En general son hierbas, si bien pueden ser leñosas —como los bambúes tropicales—, cespitosas, rizomatosas o estoloníferas. Por la duración de su ciclo de vida pueden ser anuales, bienal o perennes. Las gramíneas anuales, como es lógico suponer, se reproducen una sola vez durante su ciclo vital —el caso del trigo o de la avena, por ejemplo—. Las especies perennes, en cambio, pueden reproducirse varias veces —en general anualmente— o una sola vez. En el primer caso se denominan iteróparas —la mayoría de las especies de pastos, por ejemplo— y, en el segundo caso, semélparas —como es el caso de las diferentes especies bambúes—.
Tienen tallos cilíndricos a elípticos en su sección transversal, articulados, llamados ordinariamente cañas, en general con nudos macizos y entrenudos huecos (pero pueden ser totalmente macizos como en el caso del maíz y algunos bambúes). Los nudos son algo más gruesos que los entrenudos y en ellos nacen las hojas y las yemas. Los entrenudos son a veces algo achatados en la zona donde se desarrollan las ramificaciones. Un poco más arriba del nudo existe un meristema intercalar en forma de anillo que determina el alargamiento del tallo. En algunos géneros existen de dos a seis nudos muy próximos entre sí (los cuales se denominan nudos compuestos), cada uno de los cuales lleva su correspondiente hoja. En Cynodon dactylon, por ejemplo, los nudos están en grupos de a dos por lo que las hojas parecen opuestas. En general los entrenudos basales son más cortos que los superiores; cuando hay varios nudos basales muy próximos, las hojas parecen arrosetadas, es decir, se disponen de forma tal que simulan formar una roseta basal de hojas. Los principales tipos de tallos en las gramíneas son los siguientes:
Poseen hojas de disposición alterna, dísticas, compuestas típicamente de vaina, lígula y limbo. La vaina rodea apretadamente al tallo, sus márgenes se superponen pero no se fusionan entre sí (solo ocasionalmente pueden ser encontradas formando un tubo). La lígula es un pequeño apéndice membranoso, o raramente un grupo de pelos (tricomas), situado en la zona de unión del limbo con la vaina, en la parte adaxial. El limbo (o lámina) es simple, usualmente lineal, con nerviación paralela. Puede ser aplanado o a veces enrollado en un tubo, puede ser continuo con la vaina o poseer pecíolo. Además de esta descripción, es necesario abordar la variabilidad que se puede hallar para cada uno de estos órganos:
Los macollos o macollas son la unidad estructural de la mayoría de las especies de gramíneas. Se forman a partir de las yemas axilares o secundarias del meristema basal del eje principal. Cada uno de estos brotes secundarios o macollos inician su aparición cuando las plantas presentan entre dos y tres hojas. Cada uno de ellos, luego de producir sus primeras hojas, genera su propio sistema radicular. La suma o adición de macollos es lo que conforma la estructura y la forma de una planta de gramínea. Cuando las gramíneas se hallan en estado vegetativo producen continuamente nuevos macollos y hojas. Cada macollo, a su vez, comenzará en su momento a producir nuevos macollos.
La inflorescencia elemental de las gramíneas se llama espiguilla y consiste en una pequeña espiga formada por una o más flores sentadas o sésiles sobre un raquis articulado, a menudo brevísimo, llamado raquilla y protegido por brácteas estériles denominadas glumas, que se insertan sobre la raquilla, una más abajo que la otra.
Las flores pueden ser unisexuales o hermafroditas y presentan un perianto rudimentario de dos o tres piezas llamadas lodículas o glumélulas, que son los órganos que determinan la apertura del antecio o casilla floral al ponerse turgentes durante la floración, permitiendo que se expongan los estigmas plumosos y los estambres. Los antecios están formados por dos glumelas:
Todos estos elementos son muy variables por lo que es conveniente analizarlos por separado.
Las espiguillas, a su vez, se hallan reunidas o agrupadas en inflorescencias compuestas de tipo racimoso. Las más frecuentes son:
El fruto o grano de las gramíneas es una cariópside, fruto seco indehiscente, con una semilla cuya testa está soldada con el pericarpio formando una envoltura muy delgada. Esta envoltura encierra el embrión y el albumen o endosperma. Este fruto es básicamente una variante del aquenio, aunque se puede encontrar cierta variedad de frutos en la familia (ver por ejemplo Werker 1997).[9] En algunos géneros como Zizianopsis o Eleusine, el pericarpio no está soldado con la semilla, de modo que el fruto es un aquenio (o un utrículo según otros autores). En algunas Bambúseas el fruto es una nuez o una baya, mientras en el género Sporobolus el pericarpio es mucilaginoso y deja salir a la semilla cuando se embebe en agua. Muchos géneros, como Aristida, Stipa, Piptochaetium, Oryza y casi todas las Paníceas poseen cariópsides que se desprenden de la planta envueltas por el lemma y por la pálea. En las Andropogóneas son las glumas las que persisten encerrando a la cariópside. En Pennisetum y Cenchrus se desprende toda la espiguilla rodeada de un involucro de cerdas o de espinas. La forma de la cariópside varía mucho según los géneros, pudiendo ser casi circular como en Briza, oblonga como en Hordeum, lanceolada como en Poa hasta casi linear, como en Vulpia. En la parte inferior de la cariópside, en visión dorsal, se aprecia el embrión más o menos elíptico cubierto por el pericarpio transparente. Por el otro lado, correspondiente al surco o sutura carpelar, se distingue, también por transparencia, la mácula hilar o hilo (o zona de unión de la semilla con el carpelo), que puede ser puntiforme, como en Poa y en las Paníceas, ovada, como en Briza subaristata, o linear, como en Hordeum, Vulpia o Festuca.
El embrión de las gramíneas es estructuralmente muy complejo y consta de la plántula unida a su cotiledón laminar, altamente modificado, llamado escudete. El cotiledón es delgado, parenquimatoso, llevando en su parte exterior una capa de células epiteliales que durante la germinación segregan enzimas que hidrolizan las sustancias de reserva localizadas en el endosperma. La planta consta de un nudo cotiledonar, donde se inserta el cotiledón, una yémula cubierta de un capuchón o coleoptilo y una radícula envuelta por otro capuchón o coleorriza. En muchos géneros en la parte externa del nudo cotiledonar hay una escama diminuta, el epiblasto, que para algunos autores constituye un resto de un segundo cotiledón, mientras que otros consideran que se trata de un apéndice de la coleorriza.[4][5]
El tamaño y el número de cromosomas tienen gran importancia en la sistemática de las gramíneas. Hay dos tipos cromosómicos extremos: el tipo festucoide caracterizado por presentar cromosomas grandes y número básico predominantemente x=7 y el tipo panicoide con cromosomas pequeños y números básicos predominantes x=9 y x=10. El tipo festucoide se encuentra en casi todas las tribus de la subfamilia de las poóideas, con algunas excepciones. Por ejemplo, la tribu Stipeae de esta subfamilia posee cromosomas pequeños y números básicos x=9, 10, 11, 12, 14, 16 y 17. Las restantes subfamilias de las gramíneas presentan el tipo cromosómico panicoide, con cromosomas pequeños y predominio del número básico x=9 y 10. En las bambusóideas, erartóideas y arundinóideas los cromosomas son pequeños y el número básico es x=12. En la subfamilia Danthonioideae se presentan cromosomas de tamaño intermedio y número básico x=6 y 7. La subfamilia Chloridoideae presenta cromosomas pequeños y varios números básicos, x=7, 8, 9, 10, 11, 12 y 14. Las panicóideas siempre tienen cromosomas pequeños, con números básicos x=9 o x=10, aunque existen especies con otros números básicos, que varían desde x=4 a x=19.[5]
Las gramíneas son morfológicamente distintas de cualquier otra familia de plantas y, además, son muy diversas en cuanto a morfología y hábito de crecimiento. Las diferentes especies de gramíneas —como se ha descrito en la sección previa— difieren en sus tamaños y números cromosómicos. Asimismo, difieren en el tamaño (o contenido de ADN) de sus genomas.
El genoma del arroz, por ejemplo, es más de 11 veces más pequeño que el genoma de la cebada, a pesar de que ambas especies son diploides y aparentan tener la misma complejidad morfológica y fisiológica.[10]
El contenido de genes de las diferentes especies de gramíneas, no obstante, no varia tan ampliamente como el contenido de ADN total. El arroz y la cebada, nuevamente, no difieren más que en dos veces en el número promedio de fragmentos de restricción que hibridan con las mismas sondas.[11]
La mayor parte de las diferencias en el tamaño del genoma entre especies de gramíneas se deben a diferencias en el ADN repetitivo. Los genomas más grandes, como los de cebada o trigo, están compuestos en un 75 % de ADN repetitivo, mientras que los genomas más pequeños, como el del arroz, solo contienen menos del 50 % de ADN altamente repetitivo. Más aún, se ha determinado que buena parte de ese ADN repetitivo está compuesto de retrotransposones insertos entre los genes.[12][13]
Los estudios de mapeo genómico en muchas especies de gramíneas utilizando las mismas sondas de ADN han demostrado que no solo el contenido de genes está muy conservado, sino también el orden de los genes dentro de los cromosomas.
La extensa conservación en el contenido de genes y en el orden de los mismos entre el maíz y el sorgo no es inesperada ya que ambas especies "sólo" cuentan con 15 a 20 millones de años de evolución independiente. No obstante, similares observaciones para el arroz y el maíz, las cuales divergieron hace 60 a 80 millones de años, indican que todas las especies de la familia provienen de un mismo antepasado común y que todas ellas conservan un mismo repertorio de genes en el mismo orden aproximado.[14][15][16]
Los grandes rearreglos genómicos que diferencian entre sí a todas las gramíneas son el resultado de inversiones, translocaciones o duplicaciones cromosómicas que involucran la mayor parte de los brazos cromosómicos.[17]
La mayoría, sino todas, las gramíneas son poliploides. Basados en el supuesto que todos los géneros y familias que presentan un número cromosómico básico x=12 son derivados de ancestros que sufrieron duplicaciones cromosómicas durante su evolución y que las subfamilias de gramíneas más primitivas (Anomochlooideae, Pharoideae, y Puelioideae) tienen un número cromosómico básico x=12, se puede deducir que el ancestro de las gramíneas ya era un poliploide. Se sigue, además, que todas las gramíneas que se clasifican como diploides son, en realidad, paleopoliploides (o sea, poliploides antiguos que presentan herencia disómica[18]y cuyos progenitores no pueden ser identificados mediante herramientas citogenéticas o marcadores moleculares).[19][20] La familia contiene más del 60 % de especies, distribuidas en todos los clados, que se clasifican como neopoliploides, o sea que han sufrido un ciclo adicional de duplicación genómica. En estas especies, los genomas duplicados no han divergido mucho del genoma de sus ancestros y su número de cromosomas y comportamiento citológico durante la meiosis son indicativos de la duplicación cromosómica que los ha originado. La mayoría de estos neopoliploides (más del 65 %) han derivado de cruzamientos interespecíficos o intergenéricos por lo que se les clasifica como alopoliploides.[19]
Las hemicelulosas y los polisacáridos de pectinas de la pared celular primaria de los pastos son muy diferentes de los de las demás espermatofitas, tanto en estructura como en las particularidades de la composición de los xiloglucanos.[21] Los polisacáridos son menos ramificados que en todas las demás familias de plantas, si bien esta afirmación está basada en un muestreo todavía escaso de especies. Las poáceas pueden ser cianogenéticas o no. Cuando son cianogenéticas, los compuestos cianogenéticos son derivados de la tirosina. Pueden presentar alcaloides (a veces isoquinolina, pirrolizidina e indol). Raramente puede haber proantocianidinas y cianidinas, en cantidades traza, y sólo en representantes de las subfamilias Panicoideae y Chloridoideae. Los flavonoides se han hallado solo en algunos géneros, Bouteloua Glyceria y Melica,[22] cuando están presentes son quercetina, o kaempferol junto con quercetina. El ácido elágico y la arbutina no se han encontrado en ningún miembro de la familia. Raramente se encuentran saponinas y sapogeninas, así como también oxalatos libres (por ejemplo en Setaria).
Una generalización acerca del modo de reproducción de las gramíneas es que los miembros de esta familia son plantas hermafroditas, que presentan fertilización cruzada (son alógamos) y se polinizan por el viento. Obviamente, una familia con cerca de 10 000 especies cuenta con muchas excepciones a esta regla, las cuales se describen a continuación.
Este tipo de sistema reproductivo, en el cual existen plantas femeninas y plantas masculinas, no es muy frecuente en las gramíneas. Solo 18 géneros son dioicos o presentan especies dioicas, siendo Poa el más conocido de ellos. De hecho, las especies dioicas de Poa se incluyen en un subgénero separado, Dioicopoa.[23]
Este sistema reproductivo describe el hecho de que en las poblaciones naturales de una especie coexisten individuos femeninos e individuos hermafroditas. Esta condición es bastante rara en las gramíneas. Bouteloua chondrosioides y algunas especies del subgénero Andinae de Poa son ginodioicas, si bien Cortaderia es el ejemplo más conspicuo.[24]
En este sistema los sexos están separados espacialmente pero en el mismo individuo, o sea, cada planta presenta inflorescencias femeninas y masculinas. Zea, Humbertochloa, Luziola, Ekmanochloa y Mniochloa son ejemplos de géneros con especies monoicas. Mucho más común entre las gramíneas son las especies andromonoicas, una condición muy común en las Andropogóneas y Paníceas. En las primeras, los dos sexos se presentan en espiguillas diferentes de pares heterógamos. Un par heterógamo de espiguillas consiste usualmente en una espiguilla sesil, con una flor neutra y otra hermafrodita, y una espiguilla pedicelada, con una flor neutra y otra masculina. En las espiguillas bifloras de las Paniceas, en cambio, la flor inferior es usualmente masculina o neutra, y la superior es hermafrodita. Algunos de los géneros que ejemplifican este tipo de sistema son Alloteropsis, Brachiaria, Cenchrus, Echinochloa, Melinis, Oplismenus, Panicum, Setaria, Whiteochloa, y Xyochlaena. Algunas especies dentro de estos géneros pueden tener solo flores hermafroditas ya que la flor inferior es siempre neutra, raramente ambas flores son hermafroditas. Aparte de las paníceas y las andropogóneas, Arundinelleae es otra tribu con especies andromonoicas. En el resto de la familia, las especies andromonoicas se encuentran muy esporádicamente, como por ejemplo en Arrhenatherum, Hierochloe y Holcus.[24]
La gran mayoría de las especies de gramíneas son hermafroditas, no obstante, frecuentemente son incapaces de producir semillas cuando el polen de una planta poliniza sus propios estigmas. Esto se debe a que una gran parte de las especies de la familia presentan autoincompatibilidad, de tipo gametofítica y debido a la acción de dos genes independientes (llamados S y Z) con varios alelos cada uno. Este sistema de autoincompatibilidad ha sido observado en varios géneros de la familia (Festuca, Secale, Lolium, Hordeum, Dactylis, entre muchos otros) y no es perfectamente eficiente. De hecho, de la mayoría de las especies autoincompatibles puede obtenerse una proporción —si bien reducida— de semillas al autofecundar una planta.[24][25]
La autopolinización y la autofecundación están muy distribuidas entre las gramíneas. En general, es un mecanismo más común entre las especies anuales que entre las perennes y, decididamente, mucho más frecuente entre las especies colonizadoras. Este mecanismo se ha determinado en aproximadamente 45 géneros de gramíneas, entre los cuales se hallan géneros económicamente muy importantes como Triticum, Oryza, Secale, Avena, Agropyron y Lolium. Una condición de autogamia extrema es la cleistogamia, en la cual se produce la polinización y la fecundación dentro del antecio sin que se produzca la antesis. Este último sistema está distribuido en más de 70 géneros pertenecientes a 20 tribus de gramíneas.[24][26]
La apomixis se define como la reproducción asexual a través de semillas. En este sistema reproductivo los embriones se desarrollan por mitosis a partir de una oósfera no reducida sin que tenga lugar la fecundación. En otras palabras, cada embrión producido es genéticamente idéntico a la planta madre. En las gramíneas, la apomixis fue descrita por primera vez en 1933 en una especie de Poa. Desde aquel momento se ha identificado este mecanismo en cientos de especies de poáceas, particularmente en las Paníceas y en las Andropogóneas. Algunos de los géneros que presentan especies apomícticas son Apluda, Capillipedium, Heteropogon, Themeda, Sorghum, Bothriochloa, Dichanthium, Cenchrus, Setaria y Paspalum.[24][27][28]
Las gramíneas son una familia cosmopolita que habita desde los desiertos hasta los hábitats de agua dulce o marinos, y todas las elevaciones salvo las más altas del planeta. En el mundo se han desarrollado extensos biomas nativos dominados por gramíneas donde hay sequías periódicas, topografía plana o inclinada, incendios frecuentes, y en algunas ocasiones donde hay pastoreo y bajo ciertas condiciones particulares de suelo. Las comunidades dominadas por los pastos suman el 24% de la vegetación del planeta, ejemplos son las praderas de Norteamérica, las pampas de Sudamérica, el "veldt" o la sabana en África, y las estepas euroasiáticas. Por fuera de las praderas herbáceas, los bambúes leñosos desempeñan un papel central en la ecología de los bosques de Asia tropical y templada.[3]
Las gramíneas han sido ecológicamente exitosas y se han diversificado extensamente debido a muchas adaptaciones clave.[29] La espiguilla protege a las flores pero, al mismo tiempo, permite la polinización cuando las lodículas abren el antecio. Asimismo, las espiguillas (lemmas con pelos o ganchos) poseen varias adaptaciones para la dispersión del fruto. La versatilidad en los sistemas de apareamiento, incluyendo la autofecundación y la apomixis, permitió a diversas especies de gramíneas ser colonizadoras exitosas de nuevos ambientes. La anatomía de la hoja, que puede ser C3 o C4, permite a estas especies explorar y adaptarse a un amplio rango de hábitats. Los meristemas se hallan ubicados en la base de los entrenudos y en la base de las vainas, protegidos por toda la planta, lo que da como resultado una adaptación al pastoreo y al fuego sin igual entre todas las plantas. El desarrollo de las praderas durante el Mioceno (hace unos 25 a 5 millones de años) puede haber fomentado la evolución de los grandes herbívoros, además de representar una importante fuente de alimento y un estímulo para la evolución del Homo sapiens.[3]
Las gramíneas además han desarrollado ciertas características fisiológicas que les han permitido conquistar hábitats donde prevalecen condiciones subóptimas para el crecimiento de las plantas. Una de tales características es la capacidad de acumular betaínas de glicina y otros compuestos que se hallan asociados con la adaptación de las plantas al crecimiento en condiciones salinas.[30] Por otro lado, las poóideas almacenan carbohidratos como fructanos, los que se hallan en mucha menor concentración en las restantes especies de la familia. Esta característica está asociada con la adaptación de tales especies a condiciones de estrés hídrico (sequías) y de bajas temperaturas (heladas).[31] Finalmente, otro mecanismo fisiológico singular de las gramíneas es que aparentemente son la única familia de angiospermas que adquiere iones por quelación de iones férricos con sideróforos que son absorbidos por las raíces.[32]
Las micorrizas y las micofilas son dos tipos de mutualismos en los cuales se hallan involucrados plantas superiores y hongos. Las micorrizas son mutualismos entre hongos y las raíces de las plantas. Las micofilas, entretanto, son mutualismos entre hongos endófitos (aquellos que crecen dentro de las plantas) y la parte aérea de las plantas.[33] [34] Por otro lado, las gramíneas también pueden asociarse con diferentes géneros de bacterias de vida libre que fijan el nitrógeno atmosférico.
La presencia de hongos endófitos puede modificar la supervivencia de las plantas de varios modos ya que se pueden producir tres tipos diferentes de asociaciones entre los simbiontes fúngicos y las plantas, las cuales varían de acuerdo con el grupo taxonómico del hospedante, con las estructuras fúngicas y vegetales involucradas y con las particularidades intrínsecas de la simbiosis.[35][36] Así, el tipo I tiene como hospedantes a miembros de la familia de las juncáceas y a varias subfamilias de las gramíneas. Estos hongos colonizan la totalidad del hospedante y producen sus estructuras sexuales en estromas que suplantan a los frutos que deberían producir las plantas, por lo que éstas pierden la capacidad de reproducirse sexualmente. La interacción es sumamente agresiva para el hospedante y determina su paulatino decaimiento. El tipo II tiene como hospedantes a las gramíneas de la subfamilia de las poóideas. No todos los individuos de la población colonizada presentan estromas en lugar de cariópsides. En los ejemplares sin síntomas externos el micelio endofítico llega a colonizar las cariópsides sin perjudicar la reproducción sexual de la planta. De esta manera, el carácter patógeno de la interacción es menor que en el caso anterior. Por último, en el tipo III, que también se da en la subfamilia de las poóideas, nunca aparecen estromas en los hospedantes; la colonización del hongo es sistémica, alcanzando a las cariópsides por medio de las cuales se propaga dicha asociación. Este tipo de interacción es considerado una simbiosis mutualista debido a que los endófitos benefician a los hospedantes aumentando su crecimiento, biomasa, tasa fotosintética, tolerancia a las heladas y sequías, resistencia a nematodos e insectos, por lo que incrementan la competitividad de sus hospedantes. Asimismo, debido a que producen alcaloides protegen a las plantas del ataque de un amplio espectro de animales herbívoros, constituyendo así parte de su sistema de defensa. La presencia de endófitos afecta la palatabilidad de los pastos para los herbívoros y también la palatabilidad de las semillas para los pájaros granívoros, los animales que comen material infectado presentan diversos síntomas de intoxicación.[37][38] El nivel de la infestación por áfidos y la de sus parásitos y parasitoides, y aún el patrón y la tasa de descomposición del pasto muerto, también son afectados por el mutualismo.[39][40] Asimismo, las larvas de moscas del género Phorbia (o Botanophila) viven en el estroma de los hongos endofitos del género Epichloë, y los adultos transmiten los espermacios del hongo en una forma análoga a la polinización por insectos de las flores.[41] A su vez, los endófitos se benefician recibiendo el aporte directo de hidratos de carbono producidos por sus hospedantes. De las 232 micofilas conocidas en el mundo, 209 tienen como hospedantes a miembros de todas las subfamilias de gramíneas y representan a los tres tipos de interacción. Las áreas de distribución de estas interacciones abarcan tanto zonas frías y templadas como tropicales. En el Hemisferio Norte son muy frecuentes las interacciones tipo I, II y III, mientras que en el Hemisferio Sur prevalecen las de tipo III.[42] Los hongos endofitos de la familia Clavicipitaceae están ampliamente distribuidos entre las gramíneas. Un género de esta familia de hongos, Epichloë, es un endofito restringido a la subfamilia Pooideae,[43][44][45][42][46] Neotyphodium es el estado asexual o imperfecto de Epichloë.[47][48][49] Más del 30% de las especies de poóideas están involucradas en tales asociaciones, y existe transmisión de estos hongos (la subfamilia Balansiae de las Clavicipitáceas) tanto en forma vertical como horizontal.[50] Las micofilas son una asociación que parece datar de hace unos 40 millones de años y una de sus consecuencias es la producción de alcaloides como, por ejemplo, la lolina.[51] Los alcaloides producidos, como la mencionada lolina, son activos principalmente en la defensa de las plantas contra insectos.[52] Otras muchas especies endófitas, aparentemente asintomáticas, pueden crecer conjuntamente en las gramíneas, pero se conoce muy poco sobre sus relaciones. Márquez y colaboradores (2007), por ejemplo, informaron que la gramínea Dicanthelium lanuginosum solo puede crecer en suelos calentados por acción volcánica cuando el hongo endofítico Curvularia, con el que está asociada, se halla infectado con un virus.[53] Esto indica que las relaciones entre las gramíneas y los hongos endófitos pueden ser extremadamente complejas y sus efectos insospechados. Existen listados de especies de hongos endófitos asociados con innumerables especies de gramíneas.[54] De hecho, solo en los bambúes hay al menos unas 1.933 especies de hongos descritas.
Los registros fósiles más antiguos indican que esta asociación tiene unos 400 millones de años, lo que indica la compleja coevolución entre las plantas y sus hongos asociados, que se manifiesta en la amplia distribución del fenómeno (se ha estimado que el 90% de las plantas terrestres están micorrizadas) y en la diversidad de mecanismos morfológicos, fisiológicos y ecológicos implicados. Durante la simbiosis, la planta hospedera recibe nutrientes minerales del suelo tomados por el hongo (principalmente fósforo), mientras que este obtiene compuestos de carbono derivados de la fotosíntesis.[55] Los hongos formadores de micorrizas arbusculares constituyen micorrizas que colonizan el tejido interno de las raíces de la planta hospedera, donde desarrollan estructuras características de la simbiosis (arbúsculos y vesículas), así como micelio extrarradical, el cual interacciona con el ecosistema de la rizósfera y es el encargado de extraer nutrientes del suelo.[56] En este sentido las gramíneas no son la excepción. Buena parte de las especies de esta familia forman micorrizas, lo que favorece y optimiza su adaptación a diversos tipos de ambientes.[57]
El nitrógeno molecular (N2) es la única reserva de nitrógeno accesible en la biósfera. Prácticamente ilimitada, esta reserva no es directamente utilizada por los vegetales y animales. El nitrógeno es un constituyente esencial de moléculas fundamentales de todos los seres vivos: aminoácidos, proteínas, ácidos nucleicos, vitaminas, entre las más importantes. Para que el nitrógeno atmosférico pueda ser asimilado, es necesario que sea reducido. Las gramíneas son capaces de asociarse con bacterias diazotróficas pertenecientes a los géneros Azospirillum, Azotobacter, Azoarcus y Herbaspirillum las cuales realizan la fijación biológica del nitrógeno atmosférico (N2). Estas bacterias son organismos de vida libre capaces de fijar nitrógeno desde la rizósfera, o sea, desde el área circundante al sistema radicular de la planta e incluirlo en compuestos (como el amonio) fácilmente disponible y absorbible por las plantas. Además de fijar el nitrógeno atmosférico, las bacterias diazotróficas favorecen el desarrollo del sistema radicular de la planta con la cual conviven, al parecer a través de la producción de reguladores de crecimiento u hormonas. De este modo, favorecen una mayor absorción de nutrientes por parte de la planta. Se han informado incrementos del orden de 5 % hasta 30 % en los rendimientos de gramíneas como caña de azúcar, maíz, arroz, trigo y gramíneas forrajeras como resultado de esta asociación. Estas asociaciones no desarrollan estructuras diferenciadas en las que se alberguen los microorganismos, como ocurre en el caso de las leguminosas y las bacterias del género Rhizobium. En 1998 se ha descrito otro tipo de asociación de diazótrofos en la cual la bacteria (llamada bacteria endófita) se localiza en el interior de la raíz, el tallo y las hojas de la planta. Esta asociación fue descubierta en aislamientos de diazótrofos de plantas forrajeras de Pakistán, en donde se identificó una nueva bacteria fijadora de nitrógeno llamada Azoarcus. Este microorganismo se localiza en las capas externas del córtex; una vez en el interior de la planta se disemina a los tejidos aéreos probablemente por medio de los vasos del xilema.[58][59]
Las gramíneas son anemófilas, es decir que el polen es transportado de una planta a otra por medio del viento para efectuar la polinización. Ninguna poácea tiene nectarios, si bien algunos pastos de bosques tropicales —especialmente pequeñas bambusóideas— son polinizadas por insectos.[60] La dispersión de las semillas se produce principalmente por animales e incluso algunas especies presentan estructuras especializadas para atraerlos, como los elaiosomas.[61] No obstante, la mayor parte de las especies presentan ganchos o agujas mediante los cuales los frutos o las diásporas se adhieren a los animales que pasan. Muchas especies se dispersn con el viento para lo cual presentan largos pelos en las aristas. Finalmente, Spinifex y algunos otros géneros son plantas rodadoras, las cuales son desarraigadas a la madurez y transportadas enteras por el viento, dispersando sus semillas mientras van rodando. Las aristas pueden ayudar tanto en la dispersión por viento como en la dispersión por animales; la microestructura de la superficie de las aristas puede dar como resultado que el cariopse sea directamente "plantado" en el suelo.[62][61]
Las especies leñosas de bambúes son conocidas por florecer en forma sincronizada. Muchas de ellas, además, son perennes monocárpicas, es decir que vegetan muchísimos años, florecen una sola vez y mueren luego de la dar las semillas. Esta característica también se encuentra en algunos bambúes herbáceos. En plantas que muestran este tipo de reproducción, todos los miembros de un mismo clon florecen simultáneamente, sea donde fuere que hayan sido transportados en todo el globo terrestre, y todas las plantas, después de un período reproductivo que se puede decir retrasado, mueren.
En los tallos huecos de los bambúes muchas veces se acumula agua, y vive en ella una fauna distintiva. Las poáceas proveen alimento tanto para los adultos (el polen) como para las larvas (las raíces) de distintas especies de escarabajos de la subfamilia Galerucinae de los crisomélidos.[63] Las orugas de mariposas de la familia Nymphalidae, en particular las marrones Satyrinae y las emparentadas Morphinae, son comunes en los miembros de esta familia (se encuentran en alrededor del 10 % de los censos). Los insectos del taxón Hemiptera-Lygaeidae-Blissinae se observan más comúnmente en las especies del clado llamado PACCMAD que en el clado BEP.[64]
Son comunes en las poáceas los hongos parásitos del orden Uredinales y los de la clase Ustilaginomycetes. Los que atacan a las subfamilias Bambusoideae y Pooideae (incluyendo Stipa y parientes cercanos) son particularmente distintivos.[65] Las dos terceras partes de los Ustilaginales (unas 600 especies) se encuentran en Poaceae.[66][67]
Las gramíneas y sus parientes extintos datan de hace unos 89 millones de años, el grupo principal divergió hace unos 83 millones de años.[68][69][70] A excepción de los clados basales de la familia Anomochlooideae, Pharoideae y Puelioideae las espiguillas de las gramíneas se conocen en el límite entre el Paleoceno y el Eoceno, hace unos 55 millones de años,[71] y esta cifra está a grandes rasgos en línea con una estimación de la edad de una duplicación del genoma de las gramíneas, ocurrida hace unos 70-50 millones de años.[72][73][74] Sin embargo, el fósil de una monocotiledónea (Programinis burmitis) perteneciente al Cretácico temprano (hace unos 100-110 millones de años) es similar a una gramínea bambusóidea. Si bien este fósil tiene un número de caracteres vegetativos que son comunes entre las poáceas, su identidad todavía necesita confirmación.[75] Los tejidos vegetales silicificados (fitolitos) preservados en heces fosilizadas (coprolitos) de dinosaurios del Cretáceo tardío halladas en la India indican que por lo menos cinco taxones de gramíneas extinguidas estaban presentes en el subcontinente indio durante ese período geológico (hace unos 71-65 millones de años). Esta diversidad sugiere que el grupo basal de las gramíneas se habría diversificado y distribuido en Gondwana antes de que la India quedara geográficamente aislada.[76]
La fotosíntesis C4 parece haber estado presente en las gramíneas del Mioceno temprano a medio, tanto en las Grandes Planicies de Norteamérica como en África, hace unos 25-12,5 millones de años. Quizás este tipo de fotosíntesis estuvo inicialmente asociada a cambios adaptativos en respuesta a una disminución en la concentración de CO2 en la atmósfera, si bien la gran expansión de este mecanismo fisiológico ocurrió hace solo unos 9-4 millones de años. Aún no está claro si este evento estuvo además favorecido por los incrementos en la temperatura, la disminución de las precipitaciones, el aumento de los vientos y el concomitante incremento de incendios, que habrían removido a los árboles de algunos hábitats en ese período.[77][78][79][80][81] Los detalles de los mecanismos de la fotosíntesis C4 y las morfologías asociadas con ella son muy diversos y presentan una considerable variación, particularmente en el caso de la subfamilia de las panicóideas. De hecho, la fotosíntesis C4 aparentemente se originó y evolucionó independientemente hasta ocho veces en esta subfamilia.[82] Asimismo, este mecanismo se originó de modo independiente en otras subfamilias, como Micrairoideae, Aristidoideae y Chloridoideae.[83] Independientemente de su mayor eficiencia fotosintética, las gramíneas C4 presentan menor contenido de nitrógeno, mayor cantidad de fibras de esclerénquima y pueden ser menos palatables que las gramíneas C3.[84] A pesar de estas características, existió una radiación de mamíferos herbívoros en el Mioceno,[85] que pudo estar asociada con la ampliación de las praderas y sabanas, dominadas por pastos. Sin embargo, cuando las especies de gramíneas de pradera se expandieron hacia Nebraska en el Mioceno temprano —hace unos 23 millones de años— los ungulados hipsodontos ya existían para entonces.[86]
La inflorescencia de las gramíneas es una estructura nueva en el repertorio reproductivo de las plantas con flores. Es intrincada desde el punto de vista de la biología del desarrollo, tiene una importancia central en la agronomía y, finalmente, es una verdadera intriga evolutiva. Su arquitectura controla el tipo de polinización y la producción de semillas, por lo que es una diana importantísima tanto para la selección natural como para el mejoramiento genético y la biotecnología. Es de destacar que la diversidad de estructuras que presentan las espiguillas y espigas de las gramíneas estén controladas por genes que afectan el desarrollo y que no se hallan presentes en ninguna otra familia de plantas. Estos genes se han originado luego de extensas duplicaciones del genoma y la posterior diversificación funcional de los mismos.[87][73][88][89][90][91]
La pálea (y aparentemente también la lemma) quizás sea derivada del cáliz, y las lodículas quizás sean derivadas de la corola (Ambrose et al. 2000[92]). Por otro lado un estudio de morfología comparativa sugiere que la lemma es una bráctea y que la pálea representa dos tépalos connados del verticilo más externo (Whipple y Schmidt 2006[93]). Teniendo en cuenta las relaciones de parentesco cercano entre Ecdeicoleaceae y Joinvilleaceae recientemente encontradas por Marchant y Briggs (2007[94]) y la probabilidad de que las flores de Anomochloa sean sui generis, la morfología floral de Streptochaeta puede ser plesiomórfica (ancestral) en la familia. Es interesante que las flores de Ecdeicolea también son notablemente monosimétricas, con los dos tépalos adaxiales del verticilo externo más largos y aquillados, y si bien esto no es relevante en forma directa, una diferenciación comparable en el verticilo externo del perianto ocurre en Xyridaceae, todos estos probablemente sean paralelismos. Una interpretación más común de la pálea es que es de naturaleza profilar/bracteolar, las monocotiledóneas comúnmente tienen profilos bicarenados, sin embargo, parece que las bracteolas tuvieron que reaparecer en Poaceae, ya que los clados más emparentados pero externos a la familia ("outgroups" en análisis cladístico) no las poseen. Las lodículas parecen estar involucradas en la apertura de las flores estaminadas y las perfectas, mientras que pueden estar ausentes en las flores pistiladas (Sajo et al. 2007[95]).
Los análisis filogenéticos utilizando secuencias expresas del ADN y la estructura general de los genomas sugieren que las gramíneas difieren mucho más de otras monocotiledóneas de lo que se diferencian éstas de las dicotiledóneas.[96] Tales conclusiones acerca de las relaciones de las poáceas con las otras familias de monocotiledóneas y con las dicotiledóneas no son sorprendentes. De hecho, las gramíneas son fácilmente reconocibles e identificables de cualquier otra familia, y su monofilia se halla sustentada tanto por la morfología como por los análisis moleculares de ADN. Los caracteres fenotípicos que apoyan la monofilia de la familia son la inflorescencia con brácteas, el perianto reducido, el tipo de fruto y los caracteres del embrión y de la pared del grano de polen. Las similitudes con las ciperáceas (Cyperaceae) en el hábito de cecimiento y en el tipo de espiguillas representan una evolución convergente, y no una sinapomorfía. De hecho, las ciperáceas están más emparentadas con los juncos (Juncaceae) que con las gramíneas, las cuales pertenecen al núcleo de los Poales.
La importancia económica y ecológica de la familia ha motivado la realización de una importante cantidad de estudios sistemáticos. A principios del siglo XIX, las diferencias entre las espiguillas de las poóideas y las panicóideas llevaron a Robert Brown a dividir a la familia en estos dos grupos básicos. A principios del siglo XX, los caracteres de la epidermis de las hojas y el número de cromosomas llevó a la separación de las clorídeas de las poóideas. A mediados del siglo XX, la anatomía interna de la hoja (en particular, la presencia o ausencia de la anatomía Kranz y los caracteres del embrión (presencia o ausencia de epiblasto y ciertas características del nudo cotiledonar), llevaron al reconocimiento de cinco a ocho subfamilias. Desde fines del Siglo XX, los estudios filogenéticos basados en varias secuencias de genes demostraron ser congruentes con muchas de las relaciones filogenéticas inferidas previamente a través de caracteres estructurales y fisiológicos. Los estudios moleculares más avanzados apoyan el reconocimiento de 13 subfamilias.[97][98][99][100][101]Los tres primeros linajes que divergieron son Anomochlooideae (nativa de Brasil), Pharoideae (nativa de los trópicos del Viejo y Nuevo Mundo) y Puelioideae (nativa del oeste de África). Los integrantes de estos tres grupos solo son unas 25 de las casi 10 000 especies de la familia. El resto de las especies se distribuyen en dos grandes grupos. El primero, llamado clado BEP, agrupa a Bambusoideae, a Ehrharttoideae, y a Pooideae. El segundo, denominado clado PACCMAD, agrupa a Panicoideae, Arundinoideae, Chloridoideae, Centothecoideae, Micrairoideae, Aristidoideae y Danthonioideae.[102] El clado PACCMAD está sustentado por un carácter del embrión: un largo entrenudo en el mesocótilo. La anatomía C3 es el estado plesiomórfico o ancestral en la familia. Todas las especies C4 se encuentran en el clado PACCMAD.
Todas las subfamilias mencionadas son monofiléticas, aunque sólo unas pocas poseen sinapomorfías morfológicas que caractericen a todos sus miembros. Más bien su monofilia se halla sostenida por grupos de caracteres morfológicos que deben observarse en conjunto. Las relaciones dentro de los grandes clados PACCMAD y BEP son en su mayor parte poco claras todavía, de hecho la posición de las poóideas es poco clara en algunos análisis.[103][104] El cladograma que muestra las relaciones entre los 13 clados mencionados es el siguiente:[101]
Arundinoideae sensu stricto
La familia fue reconocida por todos los sistemas de clasificación de plantas, comenzando por el de Carlos Linneo en su obra Systema naturae pasando por los de Adolf Engler,[105] Arthur Cronquist,[106] Armen Takhtajan,[107] entre muchos otros. Los sistemas modernos de clasificación (sistema de clasificación APG III,[108] y los dos sistemas filogenéticos previos, APGI y APGII)[109] también reconocen a esta familia.
Entre los caracteres diagnósticos de la familia se hallan varios de los órganos ya descriptos: las hojas que tienen vainas largas y abiertas y las lígulas en la unión entre la vaina y la lámina, los tallos redondos y usualmente huecos en los entrenudos, y las inflorescencias cuya unidad básica es la espiguilla. Las flores individuales son pequeñas, con perianto inconspicuo y con un gineceo que usualmente tiene dos estigmas plumosos y un único óvulo. En el fruto seco, de tipo aquenio (cariopse), el embrión —relativamente grande— ocupa una posición lateral y se ubica junto al tegumento de la semilla.
Son particularmente útiles para la identificación de los géneros los caracteres de la espiguilla, como el tamaño, el plano de compresión, la presencia o ausencia de glumas, el número de flores por espiguilla, la presencia de flores estériles o incompletas, el número de venas en las glumas y las glumelas, la presencia o ausencia de aristas, y la forma de las inflorescencias secundarias.
A continuación se brinda una descripción de las subfamilias de las gramíneas y de los géneros más importantes o representativos dentro de cada una.
Las anomoclóideas son plantas herbáceas y presentan inflorescencias con una morfología característica que no se parecen a las espiguillas típicas o usuales de las restantes gramíneas. Parecen ser el clado hermano de todo el resto de la familia,[97] Soreng y Davis 1998[98] lo cual sugiere que la espiguilla característica de los pastos probablemente se originó luego de que las anomoclóideas divergieran del resto de las gramíneas. Los integrantes de esta subfamilia presentan pseudopecíolo con un pulvínulo apical, la lígula de las hojas transformada en un mechón de pelos y las ramas de la inflorescencia cimosas. En la inflorescencia presentan dos brácteas a lo largo de cada ramificación y dos más debajo de cada flor, o bien las flores se disponen en forma espiralada a lo largo de ejes racimosos, con varias brácteas debajo de cada flor. Las anteras son centrifijas (en Anomochloa) o casi basifijas (Streptochaeta). Característicamente, la primera hoja de la plántula carece de lámina foliar. Los números cromosómicos básicos son x=11 y 18. Las especies pertenecientes a esta subfamilia fueron originalmente incluidas dentro de las bambusóideas, pero actualmente se reconoce que sólo poseen un parentesco distante con ellas. La subfamilia incluye a dos géneros —Anomochloa y Streptochaeta— con cuatro especies que habitan selvas y se distribuyen desde Centroamérica hasta el sudeste de Brasil.[110]
Las faróideas se caracterizan por sus hojas resupinadas, es decir, con las láminas foliares con la cara abaxial hacia arriba. Las espiguillas son unifloras y presentan seis estambres con las anteras centrifijas. El coleoptile presenta lámina. Esta subfamilia incluye cuatro géneros y unas doce especies pantropicales, las cuales habitan en las selvas.[110] Esta subfamilia, junto con Anomochlooideae y Puelioideae, fue tradicionalmente incluida dentro de las bambusóideas, pero los estudios filogenéticos sobre datos moleculares han demostrado que las tres son los clados basales de toda la familia de las gramíneas. Las faróideas se diferencian de las puelióideas por sus espiguillas unifloras y sus estigmas trífidos.[110]
Las puelióideas junto con las demás subfamilias de gramíneas, pero a excepción de las puelióideas y las anomoclóideas, presentan gineceos con dos estigmas y espiguillas que se desarticulan por encima de las glumas a la madurez. Se caracterizan además por su androceo con seis estambres. Incluye dos géneros —Guaduella y Puelia— y aproximadamente once especies distribuidas por África tropical.[110]
Las bambusóideas, en sentido restringido, incluyen tanto especies herbáceas como leñosas y son casi exclusivamente tropicales en su distribución. Las hojas son pseudopecioladas. Las flores presentan tres lodículas y un androceo con seis estambres, raramente de dos a 14. El ovario lleva dos o tres estigmas, raramente uno solo. La primera hoja de las plántulas no presenta lámina. Los números cromosómicos básicos son x=7 y x=9 a 12. Incluye 112 géneros con aproximadamente 1647 especies tropicales a templadas.[2]
La diversificación dentro de los bambúes ocurrió hace 30 a 40 millones de años. Los bambúes leñosos forman un grupo monofilético hermano del clado que contiene a las especies herbáceas. Los bambúes leñosos, con sus tallos de hasta 40 metros de altura, ciertamente no se parecen al césped. La floración en muchas de estas especies también es inusual, ya que ocurre en ciclos de hasta 120 años. Aun cuando los tallos individuales viven por solo una o unas pocas décadas, alguna forma de "reloj" hace que los tallos florezcan todos al mismo tiempo en todo el rango de distribución de la especie, causando a veces bruscos cambios ecológicos, como los que se asocian a las ratadas. Algunos géneros de bambúes leñosos son Bambusa (120 especies), Chusquea (100 especies), Arundinaria (50 especies), Sasa (50 especies), y Phyllostachys (45 especies).[110]
Los miembros de esta subfamilia presentan espiguillas con glumas muy reducidas y el androceo con seis estambres, raramente con uno solo. Los números cromosómicos básicos de esta subfamilia son x=10 y x=15. Comprende 21 géneros y 111 especies, entre las que se incluyen a los miembros de las tribus Ehrharteae del Hemisferio Sur, así como al cosmopolita Oryzeae.[2] Este último es acuático o de tierras húmedas. El representante más conocido de la tribu Oryzeae es el arroz asiático Oryza sativa, uno de los cultivos más importantes del mundo. En el norte de África también se cultiva otra de las 22 especies de Oryza: O. glaberrima. En Estados Unidos cobra importancia otra especie de la tribu, Zizania aquatica, el arroz silvestre norteamericano.[110]
Esta subfamilia es la más grande de las gramíneas. Consiste en 194 géneros que integran unas 4200 especies.[2] Se distribuyen en las regiones de clima templado de todo el globo. Entre los géneros sobresalientes se incluyen importantes cereales, como el trigo, la cebada y la avena, y también al centeno (Secale cereale), a los pastos utilizados para césped (como Poa, con 500 especies), para heno (Festuca, 450 especies), para pasturas (como Phleum, Dactylis), y algunas malas hierbas (como Agrostis, con 220 especies, y Poa). Otros géneros importantes de esta subfamilia son Stipa (300 especies), Calamagrostis (270 especies), Bromus (150 especies), y Elymus (150 especies).[2]Las poóideas se distinguen porque las ramificaciones principales de la inflorescencia son dísticas, la lemma usualmente consta de cinco nervios. Además, presentan oligosacáridos derivados de la fructosa en el tallo. Presentan cromosomas usualmente largos y el número cromosómico básico es x=7, más raramente x=2, 4, 5 o 6.[110]
Los integrantes de las cloridoideas presentan espiguillas que se desarticulan por encima de las glumas y pelos bicelulares distintivos en la epidermis de las hojas. No obstante, este último carácter puede ser una sinapomorfía de sólo un subgrupo del clado. Todo el clado salvo dos especies muestra fotosíntesis por la vía del C4. Los números cromosómicos básicos prevalecientes en la subfamilia son x=9 y x=19, aunque existen géneros con x=7 y 8. La subfamilia se distribuye principalmente en regiones tropicales áridas y semiáridas, donde se supone que la fotosíntesis C4 es ventajosa. Los centros de distribución ubicados en África y Australia sugieren un origen en el Hemisferio Norte. Algunos géneros importantes son Eragrostis (350 especies), Muhlenbergia (160 especies), Sporobolus (160 especies), Chloris (55 especies), Spartina (15 especies) y Eustachys (10 especies).[110]
Las panicóideas han sido reconocidas taxonómicamente desde hace mucho tiempo, debido a sus espiguillas características. Las cañas son usualmente sólidas, las espiguillas se hallan comprimidas dorsalmente, no presentan raquilla y son bifloras. La desarticulación de la espiguilla a la madurez se produce por debajo de las glumas. El tipo de fisiología de la fotosíntesis prevaleciente es la C4. Los gránulos de almidón en el endosperma son simples. Los números cromosómicos básicos más típicos son x=5, 9 y 10, aunque también se hallan especies con x=7, 12 y 14. La subfamilia es principalmente tropical y contiene dos grandes tribus, Andropogoneae y Paniceae, junto con un número de grupos pequeños. Las andropogóneas son relativamente fáciles de reconocer debido a sus espiguillas dispuestas de a pares. Las paníceas no son tan homogéneas como los miembros de Andropogoneae. La subfamilia comprende 203 géneros y 3.600 especies.[2] Entre los géneros más importantes se incluyen Panicum (470 especies, polifilético), Paspalum (330 especies), Andropogon (100 especies), Setaria (100 especies), Sorghum (20 especies), y Zea (4 especies). El sorgo y el maíz son dos cultivos de gran importancia económica y ambos se incluyen en esta subfamilia.[110]
Las centotecoideas son una subfamilia pobremente estudiada. Está constituida por unas 30 especies distribuidas en 11 géneros que habitan selvas templadas cálidas a tropicales. Su característica más distintiva es la presencia de estilo en la flor y de epiblasto en el embrión. El número cromosómico básico más frecuente es x=12, aunque también hay géneros con x=11.[110]
Con 14 géneros y entre 20 a 38 especies, las arundinoideas son una subfamilia cuya exacta delimitación todavía no es clara. Son gramíneas hidrofíticas a xerofíticas que habitan regiones templadas a tropicales. Sus números cromosómicos básicos son x=6, 9 y 12. Arundo (con 3 especies, Arundo donax es la caña de Castilla) y Phragmites (2 especies) son los géneros más conocidos de esta subfamilia.[110]
Esta subfamilia monofilética perteneciente al clado PACCMAD ha sido reinstalada y circunscripta nuevamente en el año 2007. Los miembros de la misma presentan estomas con células subsidiarias en forma de domo, las lígulas con mechones de pelos, embriones pequeños, fotosíntesis C4, granos de almidón simples en el endosperma. Comprende 8 géneros y unas 170 especies en su mayoría tropicales. Algunos de los géneros (como por ejemplo Eriachne) no estaban asignados a ninguna familia hasta hace poco tiempo y otros estaban incluidos en otras subfamilias (ejemplo Isachne en las Panicoideas).[102] Incluye a los géneros Isachne (100 especies), Eriachne (35 especies) y Micraira (8 especies).[110]
Las aristoideas incluyen 3 géneros y de 300 a 385 especies de regiones templadas cálidas, con aristas con una columna basal y fotosíntesis del tipo C4. Los números cromosómicos básicos son x=11 y x=12. Comprende al gran género Aristida (230 a 330 especies) y Stipagrostis (50 especies).[110]
Danthonioideae es una subfamilia bastante distribuida en todo el globo, especialmente en el Hemisferio Sur. Presentan profilos bilobados y las sinérgidas del saco embrionario de tipo haustorial. Los números cromosómicos básicos son x=6, 7 y 9. Comprende 19 géneros y unas 270 especies. Entre los géneros con mayor número de especies se hallan Danthonia (100 especies) y Rytidosperma (90 especies). Cortaderia selloana es la popular "cortadera" una gramínea ornamental que se incluye dentro de esta subfamilia.[110]
El listado de todos los géneros conocidos de gramíneas, ordenados alfabéticamente, se provee en el anexo denominado géneros de Poaceae. En muchos casos se listan sinónimos. Los enlaces a sinónimos llevan al nombre de género preferido.
La familia o alguna de sus subfamilias poseen los siguientes sinónimos:
Aegilopaceae, Agrostidaceae, Alopecuraceae, Andropogonaceae, Anomochloaceae, Arundinaceae, Arundinellaceae, Avenaceae, Bambusaceae, Chloridaceae, Eragrostidaceae, Festucaceae, Hordeaceae, Lepturaceae, Melicaceae, Miliaceae, Nardaceae, Oryzaceae, Panicaceae, Pappophoraceae, Parianaceae, Phalaridaceae, Pharaceae, Saccharaceae, Spartinaceae, Sporobolaceae, Stipaceae, Streptochaetaceae, Triticaceae, Zeaceae.[111]
La familia de las gramíneas es probablemente la que mayor importancia tiene para la economía humana.[4] De hecho, alrededor del 70 % de la superficie cultivable del mundo está sembrada con gramíneas y el 50 % de las calorías consumidas por la humanidad proviene de las numerosas especies de gramíneas que son utilizadas directamente en la alimentación, o bien, indirectamente como forrajes para los animales domésticos. En términos de la producción global, los 4 cultivos más importantes son gramíneas: caña de azúcar (Saccharum officinarum), trigo, arroz y maíz. La cebada y el sorgo están entre los primeros 12. Por otro lado, varias especies de gramíneas se utilizan en la industria.
|coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |issn= incorrecto (ayuda). La referencia utiliza el parámetro obsoleto |mes= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda)
|coautores= (ayuda)
|coautores= (ayuda)
|coautores= (ayuda) (no posee descripción de la familia, solo de los géneros).Las poáceas (Poaceae) o gramíneas son una familia de plantas herbáceas, o muy raramente leñosas, perteneciente al orden Poales de las monocotiledóneas. Con más de 820 géneros y cerca de 12 100 especies descritas, las gramíneas son la cuarta familia con mayor riqueza de especies luego de las compuestas, las orquídeas y las leguminosas; pero, definitivamente, es la primera en importancia económica mundial. De hecho, la mayor parte de la dieta de los seres humanos proviene de las gramíneas, tanto en forma directa —granos de cereales y sus derivados, como harinas y aceites— o indirecta —carne, leche y huevos que provienen del ganado y las aves de corral que se alimentan de pastos o granos—. Es una familia cosmopolita, que ha conquistado la mayoría de los nichos ecológicos del planeta, desde las zonas desérticas hasta los ecosistemas de agua salada, y desde las zonas deprimidas y anegadizas hasta los sistemas montañosos más altos. Esta incomparable capacidad de adaptación está sustentada en una enorme diversidad morfológica, fisiológica y reproductiva y en varias asociaciones mutualísticas con otros organismos, que convierten a las gramíneas en una fascinante familia, no solo por su importancia económica, sino también por su relevancia biológica.
Entre las especies más destacadas están la caña de azúcar, el trigo, el arroz, el maíz, el sorgo, la cebada, la avena, el centeno o el bambú.
Kõrrelised (Poaceae ehk Gramineae) on üheiduleheliste õistaimede sugukond.
Kõrrelisi on umbes 600 perekonda 9000–10 000 liigiga. On hinnatud, et taimekooslused, kus domineerivad kõrrelised, katavad umbes 20% maakera maismaast. Kõrreliste hulka kuuluvad tuntud teraviljad nisu, rukis, kaer, oder ning kõik muru- ja heintaimed. Teatud kõrrelised, näiteks aruheinad, kastikud, siidpöörised, helmikad ja stepirohud, on aianduses kasutusel dekoratiivtaimedena.
Kõrrelised on rohttaimed, kellele on omased seest õõnsad silindrikujulised varred. Lehed paiknevad varrel vahelduvalt. Lehel saab eristada lehetuppe, mis ümbritseb kõrt ja lehelaba. Eri liikidel võib lehetupp olla nii avatud kui ka kinnine, viimasel juhul on lehetupe servad tangentsiaalselt kokku kasvanud. Kõrrelistele on omane (ehkki esineb ka teistes taimesugukondades) lehelaba ja -tupe üleminekukohas olev keeleke (näiteks päiderool) või karvaring (näiteks pillirool). Lehelaba alusel võib teatud rühmadel eristada nn kõrvakesi, mis ümbritsevad mõlemalt poolt vart.
Kõrreliste õisik on harilikult tipmine ja see koosneb osaõisikutest, mida nimetatakse pähikuteks. Pähikud võivad koosneda ühest või sageli mitmest õiest (kuni 20[1]); botaaniliselt saab eristada tähki, pööristähki ja pöörist, esineb ka siirdetüüpe.
Pähiku alusel paiknevad libled, enamasti on neid 2, vahel ka 1 või 3[1]. Kõrreliste õit ümbritsevad sõklad, kusjuures eristatakse välissõkalt ja sisesõkalt. Esimene neist on suurem ja tugevam, kattes ja kaitstes ka sisesõkalt. Välissõkla tipuosast turritab mõnikord välja ohe. Välissõklalt (ka liblelt) võib leida ka nn anduri, mis on vastava struktuuri seljaosa (välimist osa) läbiv eenduv keskrood.
Nende õied on enamasti hüpogüünsed ehk ülemise sigimikuga. Kuna kõrrelised on tuultolmlejad, ei vaja nad tolmeldajate ligimeelitamiseks suuri ja värvilisi õisi. Kõrreliste õite värvivalik ulatub beežist pruunini ning oranžist kuldseni. Viljaks on enamasti teris.
Kõrrelised on välisehituselt sarnased lõikheinalistega.
Kõrrelistel on majanduslikult tähtis väärtus, näiteks bambust kasutatakse Ida-Aasias ja Aafrika Sahara piirkonnas ehitustöödel, söögiks kasvatatakse aga nisu, otra, rukist jne.
Alamsugukonda hirsilised kuuluvad näiteks perekonnad:
Alamsugukonda nurmikalised kuuluvad näiteks perekonnad:
Kõrrelised (Poaceae ehk Gramineae) on üheiduleheliste õistaimede sugukond.
Poaceae edo gramineak landare loredunen familia bat da. Landare belarkarak dira, urteko zikloa dutenak. Zurtoina lastozkoa da, oinaldetik zurtoina besarkatzen duten hosto luze, estu eta txandakatuak, buruxkatan bildutako loreak eta kariopsi-motako fruituak dituzte.
Bere baitan garia, artoa, arroza, banbu eta abar daude.
Poaceaek 12 azpifamilia ditu.[1][2] Leinuak ere:[3]
Poaceaek 700 genero eta 11.000 espezie ditu (2006an).[4]
Poaceae edo gramineak landare loredunen familia bat da. Landare belarkarak dira, urteko zikloa dutenak. Zurtoina lastozkoa da, oinaldetik zurtoina besarkatzen duten hosto luze, estu eta txandakatuak, buruxkatan bildutako loreak eta kariopsi-motako fruituak dituzte.
Heinäkasvit (Poaceae, aikaisemmin myös Graminaceae) on yksisirkkaisten luokkaan kuuluva kasviheimo. Heimoon kuuluu noin 600 sukua ja 10 000 lajia. Useimmat lajit elävät luonnostaan vain yhdellä mantereella, mutta ihmisen mukana ne ovat levinneet laajalle.[2] Arviolta 20 % maapallon kasvillisuuspinnasta on heinien peittämää. Heinäkasvit on taloudellisesti tärkein kasviheimo.
Heinien varsi on ontto korsi. Varressa on solmumaisia nivelkohtia, joista tapahtuu suurimmaksi osaksi pituuskasvu. Korsi voi olla suureksi osaksi puutunut tai kokonaan puutunut (kuten bambuilla).
Lehden tyvi on solmusta lähtevä vartta ympäröivä tuppi. Lehtilapa on pitkä ja kapea.

Tuulipölytteiset kukat voivat olla tähkissä, kerrannaistähkissä, tertuissa tai röyhyissä. Hedelmä on jyvä. Ulkokale ja sisäkale (kuvassa A:t) ovat heinäkasvien tähkylän uloimmaiset suomut, jotka suojaavat kukintoa. Ulkokale on sisäkaletta hieman lähempänä kukintoperää. Vihne (kuvassa C) on pieni sukanen, joka on tavallisesti kiinnittynyt heinäkasvien ulkohelpeen (kuvassa B) kärkeen tai selkään.[3] Viljakasveista vihneet näkyvät selvästi ohralla ja rukiilla.
Molekyyligeneettisiin tutkimuksiin perustuva fylogeneettinen luokittelujärjestelmä erottaa 12 alaheimoa. Niiden keskinäistä sukulaisuutta kuvataan kaavalla
Anomochloöideae {Pharoideae [Puelioideae (PACMAD + BEP -kladit)]}
PACMAD-kladi koostuu alaheimoista Panicoideae, Aristidoideae, Chloridoideae, Micrairoideae, Arundinoideae ja Danthonioideae. BEP-kladissa ovat alaheimot Bambusoideae, Ehrhardtoideae ja Pooideae.[4]
Kaksi sukua ja neljä lajia siellä täällä hajanaisesti Keski-Amerikasta Kaakkois-Brasiliaan ulottuvan alueen metsissä. Alaheimoon kuuluvat Anomochloa marantoidea, joka on sukunsa ainoa laji, ja kolme Streptochaeta-lajia.[4]
Neljä sukua ja 13 lajia tropiikin metsissä.[4]
Kaksi sukua ja 11 lajia trooppisessa Afrikassa.[4]
Sukuja 212 ja lajeja 3316 tropiikissa ja lauhkealla vyöhykkeellä. Runsaslajisimmat suvut: helmihirssit (Paspalum, 330 lajia), harjasheinät (Cenchrus, 105 lajia; sisältää entisen sulkahirssien suvun, Pennisetum), varrasheinät (Andropogon, 100 lajia), hirssit (Panicum, 100 lajia), nukkahirssit (Dichanthelium, 55 lajia) ja Eriachne (40 lajia).[4]
Kolme sukua ja 365 lajia lämpimänlauhkeilla alueilla; Euroopassa muutama laji. Suurimmat suvut ovat vihniöt (Aristida, 250–290 lajia) ja untuvaheinät (Stipagrostis, 50 lajia). [4]
Sukuja 130 ja lajeja 1721 tropiikissa ja lämpimillä alueilla enmmän tai vähemmän kuivissa kasvuympäristöissä erityisesti Afrikassa ja Australiassa. Runsaslajisimmat suvut: röllinurmikat (Eragrostis, 300 lajia), vasuheinät (Muhlenbergia, 155 lajia), karisteheinät (Sporobolus, 180 lajia) ja viuhkaheinät (Chloris, 55 lajia). [4]
Sukuja yhdeksän ja lajeja 188 tropiikissa. Suurimmat suvut: Isachne (100 lajia) ja Eriachne (35 lajia). [4]
Sukuja 19 ja lajeja 46 lauhkealla ja trooppisella vyöhykkeellä. Kasvuympäristö vaihtelee vedestä ja kosteikoista kuiviin kasvupaikkoihin.[4]
Sukuja 17 ja lajeja 281 laajalle levinneinä, eniten eteläisellä pallonpuoliskolla; Kaakkois-Aasiassa ja Malaijien saaristossa vähän lajeja. Suurimmat suvut: hinat (Danthonia, 100 lajia) ja Rytidosperma (90 lajia).[4]
Sukuja 116 ja lajeja 2123 tropiikissa ja lauhkeilla alueilla, usein metsäkasveja. Bambujen alaheimossa on kolme tribusta.[4]
Tribuksessa Olyreae on 21 sukua ja 120 lajia Keski- ja Etelä-Amerikassa sekä Afrikassa; Uudessa-Guineassa kasvaa suku Buergersiochloa. Eniten lajeja on suvussa Pariana (35 lajia). Tribuksen kasvit ovat ruohovartisia, muissa tribuksissa on puuvartisia bambuja.[4]
Arundinarieae-tribus käsittää 26 sukua ja 533 lajia hajanaisesti levinneinä Itä-Yhdysvaltoihin, Itä-Aasiaan ja Afrikkaan; enemmän tai vähemmän vuoristoissa. Runsaslajisimmat suvut: ruokobambut (Fargesia, 60 lajia), sasabambut (Sasa, 40–60 lajia), jättibambut (Phyllostachys) (55 lajia), kaislabambut (Arundinaria, 50 lajia).[4]
Bambuseae-tribus koostuu 84–101 suvusta, joissa on 1470 juurakollista lajia tropiikissa, lämpimillä ja lauhkeilla alueilla. Suurimmat suvut: Chusquea (200 lajia), kimppubambut (Bambusa, 120 lajia), Merostachys (50 lajia) ja Schizostachyum (50 lajia).[4]
Sukuja 17 ja lajeja 111 laajalti levinneinä, mutta erityisesti eteläisellä pallonpuoliskolla. Suurimmat suvut: riisit (Oryza, 20 lajia) ja hukkariisit (Leersia, 20 lajia).[4]
Sukuja 177 ja lajeja 3850 enimmäkseen pohjoisessa lauhkealla vyöhykkeellä. Alaheimo jakaantuu neljäksi kehityslinjaksi eli kladiksi.[4]
Brachyelytreae käsittää vain yhden kolmilajisen suvun, joka kasvaa Itä-Aasiassa ja Pohjois-Amerikan itäosassa. Toinen kladi on nimeltään Nardeae, ja siinä on kaksi yksilajista sukua Euroopassa. Kolmas kladi (Phaenospermateae) koostuu seitsemästä suvusta ja niiden 11 lajista, joiden levinneisyys on hajanainen: niitä on Keski-Aasiasta Itä-Aasiaan, Australiassa, Meksikossa, Balkanin niemimaalla ja Kaukasuksella. Neljäs kladi on nimetön ja suurin, siinä on 169 sukua ja 3833 lajia, jotka kasvavat enimmäkseen pohjoisella pallonpuoliskolla. Sen runsaslajisimmat suvut ovat nurmikat (Poa, 500 lajia), nadat (Festuca, 470; sisältää entisen raiheinien suvun, Lolium), höyhenheinät (Stipa, <300 lajia), kastikat (Calamagrostis, 230 lajia), röllit (Agrostis, 220 lajia), villivehnät (Elymus, 150 lajia), Nassella (122 lajia), töpökattarat (Bromus, 100 lajia) ja simakkeet (Anthoxanthum, 50 lajia).[4]
Heinäkasveihin kuuluvat monet kaikkein tärkeimmistä viljelykasveista. Heinäkasveja, joiden jyviä käytetään ihmisten ravinnoksi, kutsutaan viljoiksi. Viljoista tärkeimpiä ovat vehnä, riisi, maissi, ohra, ruis ja kaura. Ne muodostavat tärkeän energian- ja proteiininlähteen ihmisille. Sokeriruoko on sokerintuotannon tärkeä lähde.
Lisäksi heimoon kuuluu joukko kotieläinten, etenkin nautojen ja lampaiden rehuksi viljeltäviä kasveja, joita tavallisesti tarkoitetaankin puhuttaessa heinistä tai heinästä. Sellaisia ovat esimerkiksi nurmipuntarpää ja timotei. Niitä voidaan käyttää karjan rehuksi useilla tavoilla: karja voi laiduntaa niitä suoraan pellolta tai erilaisilta luonnonbiotoopeilta, ne voidaan niittää karjalle tuoreena nurmirehuksi tai valmistaa säilörehuksi yhdessä muiden nurmikasvien kanssa tai niistä voidaan valmistaa karjan talvirehuksi kuivaa heinää. Näistä kuiva heinä oli yhdessä esimerkiksi lehdesten kanssa aiemmin pääasiallinen talvirehun tuotantotapa pohjoisilla alueilla, mutta säilörehun yleistyttyä kuivan heinän tuotanto talvirehuksi on vastaavasti vähentynyt.
Heinäkasvit (Poaceae, aikaisemmin myös Graminaceae) on yksisirkkaisten luokkaan kuuluva kasviheimo. Heimoon kuuluu noin 600 sukua ja 10 000 lajia. Useimmat lajit elävät luonnostaan vain yhdellä mantereella, mutta ihmisen mukana ne ovat levinneet laajalle. Arviolta 20 % maapallon kasvillisuuspinnasta on heinien peittämää. Heinäkasvit on taloudellisesti tärkein kasviheimo.
Les Poaceae, ou graminées, sont une famille de plantes monocotylédones de l'ordre des Poales, qui comprend environ 12 000 espèces groupées en 780 genres, à répartition cosmopolite. C'est, par le nombre d'espèces, la cinquième famille de plantes à fleurs, après les Asteraceae, Orchidaceae, Fabaceae et Rubiaceae. On y trouve la plupart des espèces appelées communément herbes et les céréales.
Ce sont généralement des plantes herbacées, plus rarement ligneuses (bambous), qui partagent des caractéristiques morphologiques qui les distinguent nettement des autres familles végétales : tiges (chaumes) cylindriques aux entrenœuds creux, feuilles alternes à disposition distique, au limbe linéaire à nervation parallèle, et dont la gaine enveloppe la tige, inflorescence élémentaire en épillets, fleurs réduites aux organes sexuels (étamines et ovaire), fruits dont le péricarpe est soudé à la graine (caryopses).
Les formations graminéennes, telles que les savanes et les prairies, dans lesquelles les Poaceae sont l'élément dominant, couvrent plus de 40 % de la surface terrestre (Groenland et Antarctique exclus)[3]. Les graminées forment également une part importante d'autres habitats, notamment des zones humides, des forêts et de la toundra.
C'est la famille de plantes la plus importante sur le plan économique, qui fournit une part essentielle de l'alimentation de base directement grâce aux espèces domestiquées comme les céréales (blé, riz, maïs, orge et millet), la canne à sucre, et indirectement grâce aux plantes fourragères, sans compter les matières utiles à l'industrie ou l'artisanat comme les bambous, la paille, le chaume et la biomasse (éthanol). Les graminées sont aussi cultivées pour l'agrément, notamment pour constituer des pelouses et des terrains de sport (golf), et pour lutter contre l'érosion des sols (oyat, vétiver). La famille compte aussi de nombreuses espèces adventices qui affectent les cultures, cinq espèces de Poaceae (Cynodon dactylon, Echinochloa colona, Eleusine indica, Sorghum halepense, Imperata cylindrica) figurant parmi les dix « pires » mauvaises herbes sur le plan mondial[4].
L'étude des phytolithes a montré que les Poaceae sont probablement apparues au cours du Crétacé, il y a plus de 85 millions d'années, faisant déjà partie de l'alimentation des dinosaures[5].
Le nom de « Poaceae » a été donné à cette famille par John Hendley Barnhart en 1895[6], en référence au nom de la tribu des Poeae (taxon décrit en 1814 par Robert Brown), et au nom du genre-type, Poa (décrit en 1753 par Linné). Ce terme dérive du grec ancien, πόα (poa), désignant l'herbe.
« Poaceae » est un nomen alternativum (nom de famille alternatif), correctement formé à partir du nom d'un genre, mais dont l'usage est autorisé en application de l'article 18.6 du Code international de nomenclature pour les plantes (CIN), aux côtés de « Graminae », considéré comme un nom valide en raison d'un « long usage » (art. 18.5 du CIN)[7].
Les graminées sont en général des plantes herbacées, annuelles ou vivaces à tige cylindrique creuse portant des nœuds, le chaume, généralement non ramifiée sauf au niveau du sol où se produit souvent le phénomène du tallage, qui conduit à la formation de touffes caractéristiques. Certaines espèces produisent des rhizomes et des stolons qui permettent l'occupation du terrain en surface et la formation de pelouses.
À maturité, le système racinaire des graminées est fibreux et fasciculé, constitué exclusivement de nombreuses racines adventives qui forment un chevelu important. Ces racines adventives remplacent les racines séminales et les racines nodales primaires qui ont une durée de vie éphémère. Elles prennent naissance au niveau des nœuds de la base des tiges, des rhizomes et des stolons, en fait de tous les nœuds en contact avec le sol. Dans certains cas, chez le maïs et le sorgho par exemple, on peut voir apparaître, nettement au-dessus du niveau du sol, des verticilles de racines, appelées racines coronaires ou racines d'ancrage[8].
La tige des graminées, ou chaume, a une structure caractéristique. C'est une tige cylindrique, à section circulaire à elliptique, articulée par une succession de nœuds pleins, séparés par des entrenœuds généralement creux par résorption de la moelle. Quelques espèces ont toutefois une tige pleine, par exemple la canne à sucre dont la moelle est exploitée pour la production de sucre.
Les nœuds sont un peu plus épais que les entrenœuds et c'est à leur niveau que naissent les feuilles et les bourgeons. Les entrenœuds sont parfois quelque peu aplatis dans la zone où se développent des ramifications. Juste au-dessus du nœud, on trouve un méristème intercalaire en forme d'anneau qui détermine l'allongement de la tige.
Au niveau des nœuds, se trouvent des éléments de renfort fibreux qui donnent à la tige plus de stabilité et de résistance à la traction. Ces tiges restent ainsi souples et flexibles, et sont capables de se redresser à nouveau après une exposition au vent ou à la pluie.
Chez certains genres, il existe deux à six nœuds plus rapprochés les uns des autres (qui sont appelés « nœuds composés »), chacun d'entre eux portant la feuille correspondante. Chez Cynodon dactylon, par exemple, les nœuds sont groupés deux à deux de sorte que les feuilles paraissent opposées. En général les entrenœuds sont plus courts à la base des tiges que ceux de la partie supérieure ; lorsque de nombreux nœuds à la base de la plante sont très rapprochés, les feuilles sont disposées de telle manière qu'elles simulent une rosette.
On peut distinguer différents types de tiges chez les graminées :
Les feuilles des graminées, à disposition alterne distique, se caractérisent par un limbe allongé, étroit, généralement linéaire à nervation parallèle. Elles sont généralement sessiles, s'attachent à la tige par une gaine. Le limbe est simple et peut être parfois aplati ou roulé en tube. La gaine, généralement fendue, entoure étroitement la tige, ses marges se chevauchant mais sans se souder (il y a des exceptions dans lesquelles la gaine forme un tube). À la jonction du limbe avec la gaine, se trouve un petit appendice membraneux situé dans la partie adaxiale, la ligule, plus rarement réduite à un groupe de poils (trichomes).
Ces différents éléments présentent une certaine variabilité :
Les graminées sont des plantes anémophiles qui possèdent des caractéristiques communes que les botanistes regroupent sous le terme du « syndrome de l'anémophilie » : fleurs discrètes souvent vert-pâle au périanthe réduit (à des écailles appelées lodicules[11], turgescentes à la floraison, ce qui permet la sortie des organes reproducteurs par écartement des glumelles) ou absent (pas d'investissement dans ces pièces florales, dans le parfum ou le nectar qui font partie des stratégies d'attraction des pollinisateurs par les fleurs entomophiles). La fleur est enveloppée par deux bractées appelées glumelles. La glumelle inférieure, appelée lemme, est une bractée insérée sur le rachillet, la glumelle supérieure, ou paléole, est une préfeuille insérée sur l'axe floral. La fleur entourée de ses glumelles constitue le fleuron[12]. L'appareil reproducteur fleur comprend en général un verticille de trois étamines (mais six chez les Bambusoideae), parfois seulement deux (ex. : Imperata sp.), très rarement une seule (Vulpia ciliata). Ces étamines sont composées d'anthères médifixes (d'où la forme typique d'X) insérées sur de longs filets grêles qui pendent hors de l'épillet (anthères dites versatiles ou oscillantes car pouvant s'incliner sous différents angles selon la direction du vent). L'ovaire uniloculaire (résultant de la fusion des trois carpelles des monocotylédones) est généralement surmonté de deux longs stigmates ramifiés (aspect « plumeux » typique des Poacées) dont la surface d'échange augmente la probabilité de capturer les grains de pollen des Poaceae. Le caractère aléatoire de l'anémogamie est le risque d'autofécondation. L'autopollinisation existe chez les espèces dont les organes reproducteurs arrivent à maturité en même temps, mais ne conduit pas forcément à l'autofécondation et la majorité des espèces pratiquent la séparation des sexes dans l'espace (diécie) ou dans le temps (protandrie), ce qui favorise la pollinisation croisée[13].
L'inflorescence élémentaire est l'épillet, qui comprend un nombre variable de fleurons, disposés de façon distique le long d'une axe, le rachillet. L'épillet est sous-tendu par deux bractées stériles, les glumes, en général au nombre de deux. La glume inférieure, toujours plus petite que la glumelle supérieure, est parfois absente. Le nombre de fleurs se réduit souvent à un, comme chez le riz (oryza sativa), mais de tels exemples se trouvent dans presque toutes les tribus[12].
Dans certains cas, par exemple chez Poa bulbosa, les épillets se transforment en bulbilles, permettant une reproduction végétative[14].
On distingue, selon le mode de regroupement des épillets, trois types principaux d'inflorescences secondaires chez les graminées : panicule (avoine, agrostis, phragmites), racème et épi (blé, chiendent). Il existe cependant de nombreux types intermédiaires[8].
À maturité, l'ovaire se transforme en un fruit particulier, le caryopse, fruit caractéristique des Poaceae, souvent appelé « grain ». Il s'agit d'un type particulier d'akène, fruit sec indéhiscent renfermant une seule graine, qui se différencie par le fait que le péricarpe est fusionné avec le tégument de la graine[15]. Toutefois, on rencontre chez certains genres des caryopses particuliers, par exemple à péricarpe épais et charnu en forme de baies globuleuses (caryopses baccoïdes) chez certains genres de Bambuseae tels que Dinochloa, Melocanna, Melocalamus et Ochlandra[16],[17]. Chez Coix lacryma-jobi, le caryopse est inclus dans un involucre en forme d'utricule présentant un trou à son sommet. Ce « pseudo-fruit » particulier, dont la forme évoque une perle ou une larme, est gris bleuté, dur et d'apparence vernissée dans les formes sauvages[18],[19].
Dans le caryopse, l'embryon est en position latérale, séparé de l'albumen par le scutellum, considéré comme un cotylédon modifié. Il présente un axe embryonnaire hautement différencié, avec vers le haut le méristème de la tige (gemmule ou plumule), enveloppée par le coléoptile, et vers le bas celui de la racine (radicule), enveloppée par la coléorrhize. Chez certaines espèces, on trouve à l'opposé du scutellum, l'épiblaste, considéré parfois comme le rudiment d'un second cotylédon. L'épiblaste est absent dans les sous-familles des Arundinoideae et des Panicoideae. Chez les Arundinoideae, Chloridoideae, Centothecoideae et Panicoideae, l'embryon présente un premier entrenœud, le mésocotyle, situé entre l'insertion du scutellum et le coléoptile[20].
Le grain de pollen présente une remarquable uniformité au niveau de la famille : de forme sphéroïdale, il est protégé par une paroi externe, l'exine, mince, lisse et à ornementation réduite ou nulle (absence de plages visqueuses qui permettent aux pollens entomophiles d'adhérer entre eux). Il est monoporé ou monoaperturé (une seule aperture ou pore germinatif, entourée par un anneau épaissi). Il est parfois muni de ballonnets aérifères (sacs résultant d'une expansion de l'exine) qui diminuent leur densité et facilite la pollinisation anémophile). Pulvérulents, secs et de petite taille, généralement entre 10 et 40 μm de diamètre (un diamètre supérieur suggère une domestication de ces plantes)[21], ils sont libérés par les anthères par temps sec, ces propriétés lui confèrent une légèreté favorisant aussi l'anémophilie. La pollinisation entomophile existe cependant : dans les forêts ombrophiles tropicales caractérisées par une canopée agitée par le vent mais un étage inférieur où la vitesse du vent, considérablement freinée, est pratiquement nulle, de nombreux coléoptères consomment divers pollens de Graminées[22].
Conséquence du caractère aléatoire de la pollinisation anémophile, les grains de pollen des Poacées sont produits en grande quantité : le hasard qui préside à leur pollinisation est responsable d'un « gaspillage » phénoménal : « un épillet de seigle libère en un jour 50 000 grains de pollen[23] », et « d'une manière générale, la fécondation d'un seul ovule exige, si la surface stigmatique est de 1 mm2, l'émission d'un million de grains de pollen[24] ».
S'il existe quelques curiosités végétales avec des distance de transport de pollen sur des centaines de kilomètres, la distance efficace moyenne est généralement de quelques mètres, la probabilité qu'un pollen atterrisse sur un stigmate au-delà de cette distance devenant très aléatoire[25].
Pour identifier des taxons à un niveau infra-familial (notamment en paléopalynologie), les chercheurs doivent recourir à des caractères tels que l'ornementation et la texture de l'exine en surface, observables en microscopie électronique à balayage (MEB)[26].
Le génome de graminées se caractérise par la variation importante du nombre chromosomique de base selon les espèces, ce nombre pouvant prendre les valeurs x = 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 et 18, l'incidence élevée de la polyploïdie, estimée à 80 %, l'hybridation fréquente et l'ampleur de la variation de la taille du génome, la valeur de l'ADN 2C variant par exemple de 0,7 chez Chloris gayana à 27,6 pg chez Lygeum spartum[27]. Le nombre total de chromosomes varie de 2n = 4 chez Zingeria biebersteiniana à 2n = 266 chez Poa litorosa (espèce polyploïde)[28].
La taille et le nombre des chromosomes ont une grande importance dans la systématique des graminées. Il existe deux types chromosomiques extrêmes : le type « festucoïde » caractérisé par des chromosomes grands et un nombre chromosomique de base principalement égal à x = 7, et le type « panicoïde » avec des chromosomes petits et les nombres de base prédominants x = 9 et x = 10. Le type festucoïde se rencontre chez presque toutes les tribus de la sous-famille des Pooideae, à quelques exceptions près. Par exemple, la tribu des Stipeae a des chromosomes petits et les nombres de base x = 9, 10, 11, 12, 14, 16 et 17. Les autres sous-familles de graminées présentent le type chromosomique panicoïde avec de chromosomes petits et la prédominance des nombres de base x = 9 et 10. Chez les Bambusoideae, Ehrhartoideae et Arundinoideae, les chromosomes sont petits et le nombre de base est x = 12. Chez la sous-famille des Danthonioideae, les chromosomes sont de taille intermédiaire et le nombre de base est x = 6 et 7. La sous-famille des Chloridoideae a des chromosomes petits et plusieurs nombres de base : x = 7, 8, 9, 10, 11, 12 et 14. Les Panicoideae ont toujours des chromosomes petits avec les nombres de base x = 9 et x = 10, bien que certaines espèces présentent d'autres nombres de base, variant de x = 4 à x = 19[10].
Liste des espèces de Poaceae dont le génome a été séquencé :
La plupart des espèces de la famille des Poaceae sont des plantes hermaphrodites qui présentent une fertilisation croisée (plantes allogames) et qui sont pollinisées par le vent (pollinisation anémophile). Ce schéma de reproduction présente cependant de nombreuses exceptions : autogamie, apomixie, cléistogamie, monoécie, diécie...
L'apomixie se définit comme la reproduction asexuelle par l'intermédiaire de graines. Dans ce système reproductif, les embryons se développent par mitose à partir d'une oosphère non réduite, sans qu'ait lieu la fécondation. En d'autres termes, chaque embryon est génétiquement identique à la plante-mère. L'apomixie a été décrite pour la première fois chez les graminées en 1933 chez une espèce de Poa. On a depuis identifié ce mécanisme chez des centaines d'espèces de Poaceae, en particulier chez les Paniceae et chez les Andropogoneae. Parmi les genres comprenant des espèces apomictiques figurent Apluda, Capillipedium, Heteropogon, Themeda, Sorghum, Bothriochloa, Dichanthium, Cenchrus, Setaria et Paspalum[41],[42],[43].
L'autopollinisation et l'autofécondation sont très répandues chez les graminées. En général, c'est un mécanisme plus commun chez les espèces annuelles que chez les espèces vivaces, et beaucoup plus fréquent chez les espèces colonisatrices. Ce mécanisme a été déterminé chez environ 45 genres de graminées, et notamment chez des genres très importants sur le plan économique, tels que Triticum, Oryza, Secale, Avena, Agropyron et Lolium. Un type d'autogamie extrême est la « cléistogamie », dans laquelle la pollinisation et la fécondation se réalisent à l'intérieur même du fleuron, sans que se produise l'anthèse. La cléistogamie est présente chez plus de 70 genres appartenant à 20 tribus de graminées[41],[44].
Bien que majoritairement hermaphrodites de nombreuses espèces de graminées sont incapables de produire des graines quand le pollen d'une plant pollinise ses propres stigmates. Cela est dû au fait qu'un grand nombre de ces espèces présentent une auto-incompatibilité, de type gamétophytique, due à l'action de deux gènes indépendants (appelés S et Z), ayant chacun divers allèles. Ce système d'autoincompatibilité a été observé chez différents genres de la famille (Festuca, Secale, Lolium, Hordeum, Dactylis entre autres) et n'est pas parfaitement efficace. En effet, chez la majorité des espèces auto-incompatibles, on peut obtenir une certaine proportion de graines (même si elle est réduite) par autofécondation d'une plante[41],[45].
La monoécie est un système reproductif dans lequel les sexes sont séparés (sur des fleurs différentes) mais chaque individu présente des inflorescences mâles et femelles séparées ou parfois des fleurs mâles et femelles distinctes dans des inflorescences mixtes. Zea, Humbertochloa, Luziola, Ekmanochloa et Mniochloa sont des exemples de genres comprenant des espèces monoïques.
Beaucoup plus communes chez les graminées sont les espèces « andromonoïques », en particulier chez les Andropogoneae et les Paniceae. Chez les premières, les deux sexes se présentent sur des épillets différents en paires hétérogames. Une paire hétérogame d'épillets consiste généralement en un épillet sessile, avec une fleur neutre et une fleur hermaphrodite, et un épillet pédicellé, avec une fleur neutre et une fleur mâle. Chez les épillets biflores des Paniceae, en revanche, la fleur inférieure est généralement mâle ou neutre, et la fleur supérieure est hermaphrodite. Parmi les genres qui comprennent des espèces andromonoïques figurent notamment : Alloteropsis, Brachiaria, Cenchrus, Echinochloa, Melinis, Oplismenus, Panicum, Setaria, Whiteochloa et Xyochlaena. Certaines espèces de ces genres peuvent avoir seulement des fleurs hermaphrodites puisque la fleur inférieure est toujours neutre. Il est rare que les deux fleurs soient hermaphrodites. Outre les Paníceae et les Andropogoneae, les Arundinelleae sont une autre tribu comprenant des espèces andromonoïques. Dans le reste de la famille, les espèces andromonoïques se rencontrent de façon très sporadique, comme chez les genres Arrhenatherum, Hierochloe et Holcus[41].
La diécie est un type de système reproductif, dans lequel des fleurs mâles et femelles sont portées par des individus différents est assez rare chez les graminées. Seuls 18 genres sont dioïques (ou comprennent des espèces dioïques), parmi lesquels le genre Poa est le plus connu. En fait, les espèces dioïques de Poa ont été classées dans un sous-genre particulier, Dioicopoa[41].
La gynodiécie est un système reproductif dans lequel coexistent des individus femelles et des individus hermaphrodites. Ce système est rare chez les graminées. Bouteloua chondrosioides et quelques espèces du sous-genre Poa subgen. Andinae sont gynodioïques, même si Cortaderia est l'exemple le plus remarquable[41].
Les graines (caryopses) sont riches en amidon, qui peut se présenter sous forme de grains d'amidon individuels (seigle, blé, orge) ou de plusieurs grains d'amidon composites (avoine).
Les graminées accumulent aussi dans les rhizomes et autres organes végétatifs de l'amidon, du saccharose ou des fructanes. Les fructanes se présentent, outre le type inuline non ramifié, sous la forme de type phléine ramifié. Le type de fructane qui dépend du degré de polymérisation est souvent caractéristique de l'espèce[46].
Les grains de céréales (caryopses) contiennent de 10 à 15 % de protéines (en pourcentage de matière sèche), dont la moitié environ sont des protéines de réserve, présentes principalement dans la couche externe de l'albumen (couche d'aleurone) et dans l'embryon. Ces parties de la graine sont souvent ôtées lors de la mouture du blé ou du polissage du riz. Les protéines de réserve sont principalement de l'albumine, des globulines, des glutélines (solubles uniquement dans des acides et bases dilués) et des prolamines (solubles dans l'éthanol à 70-80 %). Les prolamines, qui forment la principale fraction des protéines de réserves des principales céréales (à l'exception de l'avoine et du riz), sont déficientes en certains acides aminés essentiels : lysine, thréonine et tryptophane (en particulier chez le maïs)[47].
De la silice (sous forme de particules opalines de dioxyde de silicium), ou phytolithes, est souvent stockée dans l'épiderme foliaire chez de nombreuses espèces de graminées, notamment le riz. Cet élément peut représenter de 2 à 5 % de la matière sèche totale chez certaines espèces, soit 10 à 20 fois plus que chez les dicotylédones. La présence de silice dans les feuilles serait un mécanisme de défense contre les herbivores, vertébrés ou invertébrés, d'une part en augmentant leur caractère abrasif et d'autre part en réduisant leur digestibilité[48].
Certaines espèces de graminées produisent des glycosides cyanogènes, notamment la dhurrine chez le sorgho, qui sont susceptibles d'être hydrolysés dans certaines conditions et de produire de l'acide cyanhydrique. La synthèse de ces métabolites secondaires est vraisemblablement liée à une stratégie de défense de ces plantes contre les herbivores[49].
Des espèces de graminées, telles que la fétuque, le brome, le dactyle pelotonné, le raygrass, le maïs, les sorghos fourragers et le mil à chandelle (Pennisetum glaucum) sont susceptibles dans certaines conditions d'accumuler des nitrates, ce qui peut les rendre toxiques pour les animaux herbivores. Ce phénomène peut se produire lorsque les plantes poussent sur des sols riches en nitrates, mais que les conditions ne sont pas réunies pour une métabolisation normale et la transformation de ces nitrates en protéines, par exemple en cas de sécheresse, de temps trop froid, ou quand les plantes sont flétries, comme après l'application d'herbicides[50],[51].
Les Poacées sont une des familles les plus cosmopolites. On les trouve sur tous les continents, y compris en Antarctique[52], depuis les zones équatoriales jusqu'aux cercles polaires et depuis les bords de mer jusqu'au sommet des montagnes.
Cette famille de plantes très « sociables » représente environ 24 % de la couverture végétale du globe terrestre. Elles sont l'élément dominant de plusieurs formations végétales très étendues comme les steppes eurasiennes, la savane et le veld en Afrique, la pampa en Amérique du Sud, la Prairie américaine et les pelouses alpines. Elles ont permis le développement de toute une faune herbivore. En dehors des prairies herbacées, les bambous ligneux jouent un rôle central dans les forêts en Asie tropicale et tempérée.
On rencontre les graminées aussi bien dans les déserts que dans des habitats aquatiques d'eau douce ou marine, et à pratiquement toutes les altitudes, sauf les plus élevées de la planète.
De vastes biomes naturels dominés par les graminées se sont développés dans le monde, caractérisés aussi bien par des sècheresses périodiques, que par des reliefs plane ou accidentés, par l'occurrence d'incendies fréquente, par certaines conditions particulières de sols ou par la pression du broutage et du pâturage.
De nombreuses chenilles, comme celles des papillons de nuit suivants, se nourrissent de graminées :
Les graminées sont parasitées par diverses espèces de champignons pathogènes provoquant d'importantes maladies telles que les rouilles, les charbons et les ergots, bien connues chez le céréales, les graminées fourragères et les graminées à gazon[53]. Parmi les genres de champignons pathogènes les plus importants figurent les suivants : Alternaria, Aspergillus, Botrytis, Claviceps, Fusarium, Helminthosporium, Penicillium, Puccinia, Rhizopus, Septoria, Stachybotrys, Ustilago.
Les champignons endophytes des graminées appartiennent à la famille des Clavicipitaceae (Ascomycètes) et se développent dans les tissus végétaux dans l'espace intercellulaire. Ils se rencontrent dans toutes les sous-familles des Poaceae et sont parfois pathogènes, mais les plus fréquents, appartenant en particulier au genre anamorphe Neotyphodium, forment des symbioses mutualistes avec les plantes, principalement chez les Pooideae à photosynthèse en C3. La plupart des champignons endophytes synthétisent des alcaloïdes qui jouent un rôle dans la défense des plantes-hôtes contre les prédateurs phytophages et herbivores[53].
Les mycorhizes, qui sont des associations symbiotiques au niveau des racines, sont très répandues chez les graminées, et sont pour la plupart des « endomycorhizes »[53].
Diverses espèces de bactéries fixatrices de l'azote (ou diazotrophes) sont associées à des graminées, notamment des céréales, la canne à sucre et diverses espèces fourragères. Pratiquement toutes ces bactéries sont des organismes microaérophiles, fixant le diazote (N2) uniquement en présence de faibles pressions partielles d'oxygène. Ces bactéries, appartenant notamment aux genres Azospirillum, Acetobacter, et Herbaspirillum, sont associées (endophytes) aux graminées au niveau des racines principalement, mais aussi des tiges ou des feuilles, et peuvent apporter une contribution agronomique significative pour la fixation de l'azote atmosphérique[54].
D'autres bactéries sont des organismes phytopathogènes. Différents pathovars de Xanthomonas translucens provoquent des flétrissements bactériens.
Les graminées comprennent certaines des formes de vie végétale parmi les plus polyvalentes. Elles se sont répandues vers la fin du Crétacé, et on a retrouvé des déjections fossilisées (coprolithes) de dinosaures contenant des phytolithes provenant notamment de plantes apparentées aux espèces modernes de riz et de bambous[55]. Les graminées se sont adaptées à des conditions de milieu très variées, des forêts humides luxuriantes comme des déserts arides, des montagnes au climat froid et même à l'écologie particulière de l'estran, et sont actuellement le type de plantes le plus répandu dans le monde. Elles sont une source précieuse de nourriture et d'énergie pour toutes sortes d'organismes de la faune et la flore sauvage. Les fossiles découverts jusqu'en 2005 indiquaient que l'évolution des graminées avait commencé il y a environ 55 millions d'années. La découverte récente de phytolithes graminoïdes dans des coprolithes de dinosaures du Crétacé a permis de repousser cette date jusqu'à 66 millions d'années[56],[57]. En 2011, une révision de la datation des origines de la tribu des Oryzeae (les riz) a reporté celle-ci entre 107 et 129 millions d'années[58]. La découverte, en 2014, du squelette fossilisé d'un mammifère herbivore multituberculé, présentant des adaptations à la consommation des graminées, semble suggérer que ces plantes existaient déjà il y a environ 120 millions d'années[59].
Les relations entre les trois sous-familles, Bambusoideae, Oryzoideae et Pooideae du clade BEP ont été résolues : les Bambusoideae et les Pooideae sont plus étroitement liées entre elles qu'avec les Oryzoideae[60]. Cette séparation est intervenue dans un laps de temps relativement court d'environ 4 millions d'années.
Les analyses phylogéniques à l'aide de séquences exprimées de l'ADN et la structure générale des génomes suggèrent que les graminées diffèrent beaucoup plus des autres monocotylédones que ces dernières des dicotylédones[61]. De telles conclusions sur les relations des Poaceae avec les autres familles de monocotylédones et avec les dicotylédones ne sont pas surprenantes. En fait, les graminées sont facilement à reconnaitre et à distinguer de toute autre famille, et leur monophylie est soutenue tant par la morphologie que par les analyses moléculaires de l'ADN. Les caractères phénotypiques qui soutiennent la monophylie de la famille sont l'inflorescence avec des bractées, le périanthe réduit, le type de fruit et les caractères de l'embryon et de la paroi du grain de pollen. Les similitudes avec les carex (Cyperaceae) dans le port général des plantes et dans le type d'épillets résultent d'une évolution convergente, et pas d'une synapomorphie. En fait, les Cypéracées sont plus apparentées aux joncs (Juncaceae) qu'aux graminées, qui appartiennent au noyau des Poales.
L'importance économique et écologique de la famille a conduit à la réalisation d'un nombre important d'études de systématique. Au début du XIXe siècle, les différences entre les épillets des Pooideae et des Panicoideae ont conduit le botaniste anglais Robert Brown à subdiviser la famille dans ces deux groupes de base. Au début du XXe siècle, les caractères de l'épiderme des feuilles et le nombre de chromosomes ont conduit à séparer les Chloridoideae des Pooideae. Au milieu du XXe siècle, l'anatomie interne de la feuille (en particulier la présence ou l'absence de l'anatomie de type kranz et les caractères de l'embryon (présence ou absence de l'épiblaste et certains caractéristiques du nœud cotylédonaire), ont conduit à la reconnaissance de cinq à huit sous-familles. Depuis la fin du XXe siècle, les études phylogénétiques à partir de plusieurs séquences de gènes ont conduit à des résultats compatibles avec la plupart des relations phylogénétiques précédemment inférées à partir de caractères structurels et physiologiques. Les études moléculaires les plus avancées soutiennent la reconnaissance de 12 ou 13 sous-familles[62],[63],[64],[65],[66].
Cladogramme des Poaceae PoaceaeAnomochlooideae (4)
Pharoideae (13)
Puelioideae (11)
Oryzoideae (110)
Bambusoideae - bambous (1450)
Pooideae (3850)
Aristidoideae (350)
Panicoideae (3250)
Arundinoideae (200)
Micrairoideae (50)
Danthonioideae (300)
Chloridoideae (1600)
Les trois premières lignées qui ont divergé sont les Anomochlooideae (originaire du Brésil), les Pharoideae (originaire des régions tropicales de l'Ancien et du Nouveau Monde) et les Puelioideae (originaires d'Afrique de l'Ouest). Les membres de ces trois groupes ne représentent qu'environ 30 des quelque 12 000 espèces de la famille. Le reste des espèces est réparti en deux grands groupes ou clades : le clade BEP (ou BOP) regroupe les Bambusoideae, Oryzoideae et Pooideae ; le second, appelé clade PACMAD (ou PACCMAD), regroupe les Panicoideae, Arundinoideae, Chloridoideae, Micrairoideae, Centothecoideae, Aristidoideae et Danthonioideae[67]. Le clade PACMAD est soutenu par un caractère de l'embryon : un entre-nœud long dans le mésocotyle. L'anatomie C3 est l'état plésiomorphe ou ancestral de la famille. Toutes les espèces à photosynthèse en C4 se trouvent dans le clade PACMAD.
Toutes les sous-familles mentionnées sont monophylétiques, bien que seules quelques-unes aient des synapomorphies morphologiques qui caractérisent tous leurs membres. De plus, leur monophylie est soutenue par des groupes de caractères morphologiques qui doivent être observés dans leur ensemble. Les relations au sein des grands clades PACMAD et BEP sont pour la plupart encore peu claires, en effet la position des Pooideae n'est pas claire dans certaines analyses[68],[69]. Le cladogramme ci-contre montre les relations entre les 12 clades mentionnés[66] :
Bambou géant (Dendrocalamus giganteus, Bambusoideae).
Folle-avoine (Avena fatua, Pooideae).
Canne à sucre (Saccharum officinarum, Panicoideae).
Canne de provence (Arundo donax, Arundinoideae).
Herbe de la pampa (Cortaderia selloana, Danthonoideae).
La famille des Poaceae, ou Graminae, est classée parmi les plantes monocotylédones, dans l'ordre des Poales. Les graminées sont des Angiospermes (Magnoliopsida). À la différence des Gymnospermes, chez les Angiospermes, l'ovule est enfermé dans l'ovaire. Elle comprend environ 12 000 espèces regroupées en 780 genres[70]. C'est la cinquième famille botanique, par le nombre d'espèces, après les Asteraceae, Orchidaceae, Fabaceae et Rubiaceae[71].
Les Poaceae sont subdivisées en douze sous-familles de tailles très inégales, elles-mêmes subdivisées en 51 tribus. Le genre type de la famille des Poaceae est Poa (les pâturins). Les genres les plus représentatifs, en nombre d'espèces, sont les suivants : Panicum (500 espèces), Poa (500), Festuca (450), Eragrostis (350), Paspalum (330) et Aristida (300)[72].
La famille des graminées occupe une place essentielle dans l'économie mondiale et certaines espèces sont vitales pour l'alimentation de l'homme et de son bétail. Au niveau mondial, environ 70 % de la superficie cultivée est emblavée avec des graminées et 50 % des calories consommées par l'humanité proviennent des nombreuses espèces de graminées utilisées directement dans notre alimentation ou bien, indirectement dans l'alimentation des animaux d'élevage, en fourrage ou en grains. Les quatre espèces de plantes cultivées les plus importantes en volume de production sont des graminées : canne à sucre (Saccharum officinarum), maïs, blé, riz. l'orge et le sorgho figurent parmi les douze premières. D'autre part, diverses espèces de graminées sont utilisées dans l'industrie et pour la production d'énergie.
Ce sont principalement les céréales dont on consomme les grains (caryopses) soit entiers, soit moulus sous forme de farine, de semoule ou de gruau. Ces plantes sont cultivées depuis au moins 10 000 ans. Depuis le début de leur domestication, le blé tendre (Triticum aestivum), l'orge (Hordeum vulgare) et l'avoine (Avena sativa) dans le croissant fertile au Proche-Orient, le sorgho (Sorghum bicolor) et le mil ([Pennisetum glaucum) en Afrique, le riz (Oryza sativa) dans le sud-est de l'Asie et le maïs (Zea mays) en Méso-Amérique, ont rendu possible l'établissement de communautés humaines et le développement de civilisations. Les céréales cultivées occupent la moitié du total des terres arables et ont produit près de 3 milliards de tonnes en 2017[75]. Le blé (sous-famille des Pooideae, en particulier le blé tendre Triticum aestivum), qui a commencé à être domestiqué il y a plus de 11 000 ans, fournit le cinquième des calories consommées par l'homme. La plupart des formes domestiquées sont polyploïdes, et la plasticité du génome en connexion avec la polyploïdie explique le succès de la culture de cette céréale[76]. Il est énormément utilisé en boulangerie, viennoiserie, pâtisserie et en cuisine (sauces, desserts). Le blé dur (Triticum durum) est utilisé pour produire la semoule, le boulghour et les pâtes alimentaires. Le maïs (Zea mays, sous-famille des Panicoideae) est une céréale aux multiples applications, de la consommation directe comme « maïs doux », à la production de farine pour la préparation de nombreux plats et même de boissons alcoolisées. Le riz (Oryza sativa, Oryzoideae) est l'espèce la plus importante en alimentation humaine. L'avoine (Avena sativa, Pooideae), l'orge (Hordeum vulgare) et le seigle (Secale cereale) sont trois autres céréales couramment utilisées comme aliments.
Des céréales anciennement cultivées comme l'engrain et l'épeautre reprennent une place en alimentation diététique et biologique.
Outre les céréales, les graminées alimentaires comptent certaines espèces de bambous, comme Phyllostachys edulis et Sinocalamus beecheyanus dont on consomme les jeunes pousses comme légumes en Asie[9].
La canne à sucre, première culture mondiale par l'importance des tonnages récoltés (1,3 milliard de tonnes en 2008[77]), est la première source de sucre.
De nombreuses espèces de graminées permettent de produire du fourrage pour le bétail, tant dans les prairies naturelles que dans les prairies cultivées permanentes ou temporaires, pâturées, fanées ou ensilées. On cultive à cet effet de nombreuses espèces de graminées vivaces, aussi bien dans les climats tempérés que dans les climats tropicaux ou subtropicaux. Les espèces fourragères des régions tempérées produisent de l'herbe principalement au printemps et en automne. Les espèces les plus courantes[78] sont le ray-grass anglais (Lolium perenne), le ray-grass d'Italie (Lolium multiflorum), le ray-grass hybride (Lolium ×hybridum), le dactyle (Dactylis glomerata), la fétuque élevée (Festuca arundinacea), la fétuque des prés (Festuca pratensis), la fléole des prés (Phleum pratense), plusieurs espèces de bromes dont Bromus unioloides (espèce endémique d'Amérique du Sud), Thinopyrum ponticum, et l'alpiste tubéreux (Phalaris aquatica). Les graminées cultivées comme plantes fourragères vivaces provenant des climats tropicaux ou subtropicaux ont une production d'été et parmi elles figurent Paspalum dilatatum, Panicum elephantypes, l'Herbe de Rhodes (Chloris gayana), le mil à chandelle (Pennisetum glaucum), l'herbe de Bahia (Paspalum notatum) et Eragrostis curvula.
Certaines espèces annuelles sont utilisées pour produire de grandes quantités de fourrage en une seule récolte. Parmi les espèces d'hiver figurent les avoines (Avena fatua, Avena sativa), le seigle (Secale cereale), le triticale et certaines variétés de ray-grass d'Italie. Pour les espèces d'été, on utilise le maïs, le sorgho fourrager (Sorghum sudanense) et le moha (setaria italica).
En outre, les céréales sont utilisées en grains pour nourrir les animaux : chevaux, ruminants, porcs, volailles; c'est le cas du millet commun (Panicum miliaceum) et de l'alpiste (Phalaris arundinacea) pour nourrir des oiseaux[9],[79]. Le maïs grain est largement utilisé pour l'alimentation des porcs. Le maïs et le sorgho sont aussi cultivés comme plantes fourragères, récoltées en plantes entières, elles sont souvent conservées en ensilage.
Les utilisations industrielles des graminées sont aussi variées que la famille en elle-même.
Les espèces à parfum, dont des extraits sont utilisés dans la préparation de nombreux parfums, tels que la citronnelle (Cymbopogon citratus), dont on extrait une essence appelé huile de citronnelle et le vétiver (Chrysopogon zizanioides).
D'autres graminées, principalement l'orge brassicole (Hordeum vulgare), sont utilisées pour la préparation de malt utilisés par les industries alimentaires, surtout dans la fabrication de boissons alcoolisées par fermentation, telles que la bière, le whisky, le gin, la vodka, le genièvre, etc[80], les succédanés de café (malt torréfié) et des préparations diverses (extraits de malt). Le saké (ou nihonshu) est élaboré à partir du riz au Japon. Le Bourbon est élaboré à partir d'un malt à 51 % de maïs au moins, selon la règlementation fédérale américaine[81], généralement 70 %.
Selon les pays, on fabrique bières, et alcools après distillation d'un moût fermenté analogue à la bière, à partir des grains disponibles localement. Le malt d'orge constitue généralement la base de ces préparations mais l'orge peut être remplacée ou mélangée à n'importe quelle autre céréale. On rajoute parfois des grains non maltés cuits ou crus avant brassage pour des goûts particuliers.
L'industrie huilière utilise également des grains pour produire des huiles alimentaires ou non, comme l'huile de maïs, l'huile de germe de blé.
Voir Amidon#Utilisations industrielles
Les industries (alimentaires ou non) utilisent les amidons, les sucres et sirops issus des céréales comme matières premières ou adjuvants.
Le biogazole peut être produit à partir de céréales comme le maïs. Le bioéthanol peut être fabriqué à partir de canne à sucre.
Diverses espèces de graminées sont exploitées pour leurs fibres. L'alfa et certaines espèces de bambous permettent de produire de la pâte à papier. On extrait de certains bambous une matière première textile, la fibre de bambou. Certains genres ont un grand intérêt en sparterie, vannerie et pour la fabrication de chaussures traditionnelles, comme les espadrilles. Tel est le cas du sparte (Lygeum spartum), et surtout de l'alfa (Stipa tenacissima), qui sont tous deux utilisés en Espagne et dans le nord de l'Afrique pour produire le spart, matière première pour la fabrication de tous ces éléments, et de la canne flèche (Gynerium) en Amérique du Sud.
En outre, on utilise des espèces telles que Sorghum bicolor pour faire des balais, Epicampes microura et Aristida pallens pour faire des brosses et Stipa tenacissima pour produire des éponges[9].
Les bambous fournissent avec leur tiges épaisses un matériau apprécié dans le secteur de la construction (échafaudages, lamellé-collé) et une matière première pour la production de meubles et de divers objets artisanaux.
Les graminées utilisées pour leur grain (céréales mais aussi les cultures destinées à la production de semences) sont battues pour récupérer le grain, il reste alors la paille. Celle-ci est traditionnellement utilisée pour la confection des litières et comme fourrage pour les ruminants en cas de pénurie. Cependant, ses usages sont très variés : composts, toîtures en chaumes, combustible, vannerie, etc.
De nombreuses espèces des genres Poa, Lolium, Festuca, Axonopus, Stenotaphrum et Paspalum, sont utilisées pour créer des pelouses dans les parcs et jardins, terrains de sport, espaces verts, et pour la végétalisation des talus de routes et voies ferrées. Agrostis capillaris et Agrostis stolonifera, en particulier, sont utilisés pour les « greens » des parcours de golf[9].
Outre les graminées à gazon, de nombreuses graminées vivaces de grande taille sont devenues de plus en plus populaires pour constituer des éléments centraux dans la conception des parcs et jardins. Cela s'explique non seulement par leurs caractéristiques de longévité, d'adaptation, de rusticité et de facilité d'entretien, mais aussi par leur valeur ornementale ou par des qualités esthétiques particulières liées aux mouvements, à la transparence, à l'éclairage et aux changements saisonniers spectaculaires. Parmi les espèces utilisées comme plantes ornementales figurent notamment Alopecurus pratensis, Cortaderia selloana (Herbe de la pampa), Festuca glauca, Imperata cylindrica, Leymus condensatus, Miscanthus sinensis, Pennisetum setaceum et Phyllostachys aurea[82].
Différentes espèces de Poaceae sont exploitées pour la production d'énergie. On cultive certaines graminées vivaces, telles que Panicum virgatum, Spartina pectinata, Phalaris arundinacea[83], Miscanthus ×giganteus[84], spécifiquement pour la production de biomasse. On utilise comme combustible des sous-produits de céréales comme la paille et la balle, de la canne à sucre (bagasse) ou de l'industrie de transformation des bambous. On cultive aussi des céréales (maïs, sorgho notamment) et la canne à sucre pour la production de bio-éthanol, notamment aux États-Unis et au Brésil. Certaines espèces de bambou sont aussi cultivées pour la production d'énergie principalement en Asie, soit par combustion directe comme c'est le cas traditionnellement, soit par conversion en d'autres formes d'énergie (cogénération d'électricité, gaséification, pyrolyse, etc)[85].
Différentes espèces de graminées sont utilisées pour lutter contre l'érosion et comme fixatrices des dunes. Les espèces suivantes, par exemple, sont utilisées à cet effet : Sporobolus arundinaceus, Panicum urvilleanum, Spartine ciliata, Poa lanuginosa, Ammophila arenaria, Elymus arenarius et Arundo donax (canne de Provence).
D'autres espèces comme le ray-grass d'Italie (Lolium italicum), le moha (Setaria italica), le seigle (Secale cereale) sont utilisées comme engrais verts ou comme cultures intermédiaires destinées à piéger les nitrates.
Certains instruments de musique sont fabriqués avec des cannes de graminées, notamment des bambous, c'est le cas de certaines flûtes en utilisant Rhipidocladum harmonicum[86].
La majorité des Poaceae a des feuilles et tiges comestibles. Les enfants aiment à mâchonner la partie inférieure, tendre et juteuse, à la base de leur tige[87].
D'autres espèces, telles le chiendent officinal (Elymus repens) et le chiendent pied-de-poule (Cynodon dactylon), ont été utilisées pendant des siècles comme plantes médicinales en particulier comme diurétiques.
Les Poacées peuvent être utilisées pour pratiquer de la musique verte[88] : sifflet d'herbe en tirant des sons stridents des feuilles[89], hautbois à une anche avec la tige[90].
Ils sont également utilisés pour des jeux buissonniers entre des garçons et des filles : des espèces comme l'orge des rats ont les arêtes des épillets qui empêchent l'épi de glisser en arrière. En plaçant cet épi entre leurs bras ou dans leur manche, à chaque mouvement du bras, il remonte seul, jusqu'à venir piquer l'aisselle[91].
De nombreuses espèces de graminées sont des mauvaises herbes des cultures. Certaines d'entre elles sont très difficiles à éradiquer ou à maîtriser, et causent d'importantes pertes de rendement en concurrençant les espèces végétales cultivées. C'est le cas par exemple du sorgho d'Alep (Sorghum halepense), du chiendent pied-de-poule (Cynodon dactylon) , du panic pied-de-coq (Echinochloa crus-galli), de la digitaire sanguine (Digitaria sanguinalis) et de Brachiaria extensa[92]. Cinq espèces de graminées adventices (Cynodon dactylon, Echinochloa colona, Eleusine indica, Sorghum halepense, Imperata cylindrica) figurent parmi les dix « pires » mauvaises herbes sur le plan mondial[4].
Un cas particulier est celui des riz nuisibles qui sont des espèces voisines du riz cultivé, appartenant au même genre Oryza, voire des variétés de la même espèce, Oryza sativa, retournées à l'état sauvage et difficiles à extirper des rizières du fait de leur proximité biologique avec la plante cultivée. Ces riz adventices sont connus sous le nom de « riz crodo » en Camargue[93].
Diverses populations de graminées adventices ont été signalées comme résistantes à des herbicides. C'est le cas des espèces suivantes : Agrostis stolonifera, Alopecurus aequalis, Alopecurus japonicus, Alopecurus myosuroides, Apera spica-venti, Alopecurus aequalis, Avena fatua, Avena sterilis, Brachiaria eruciformis, Brachypodium distachyon, Bromus diandrus, Bromus japonicus, Bromus rubens, Bromus secalinus, Bromus sterilis, Bromus tectorum, Chloris barbata, Chloris elata, Chloris truncata, Chloris virgata, Cynodon hirsutus, Cynosurus echinatus, Digitaria ciliaris, Digitaria ciliaris, Digitaria insularis, Digitaria ischaemum, Digitaria sanguinalis, Echinochloa colona, Echinochloa colona, Echinochloa crus-galli, Echinochloa crus-pavonis, Echinochloa erecta, Echinochloa oryzoides, Echinochloa phyllopogon, Ehrarta longiflora, Eleusine indica, Hordeum murinum, Ischaemum rugosum, Ixophorus unisetus, Leptochloa chinensis, Leptochloa panicoides, Leptochloa scabra, Leptochloa virgata, Lolium perenne, Lolium persicum, Lolium rigidum, Oryza sativa var. sylvatica, Panicum capillare, Panicum dichotomiflorum, Paspalum paniculatum, Phalaris brachystachys, Phalaris minor, Phalaris paradoxa, Polypogon fugax, Polypogon monspeliensis, Rostraria smyrnacea, Polypogon fugax, Rottboellia cocinchinensis, Sclerochloa dura, Sclerochloa kengiana, Setaria faberi, Setaria pumila, Setaria verticillata, Setaria viridis, Sorghum bicolor, Sorghum halepense, Sporobolus fertilis, Urochloa panicoides, Urochloa plantaginea, Vulpia bromoides[94].
L'ivraie enivrante (Lolium temulentum), adventice des champs de blé, est citée dans le Nouveau Testament (Matthieu 13, 24-30 et 36-38) dans la « parabole de l'ivraie et du bon grain ». Elle est à l'origine de l'expression « semer la zizanie » (de zizania, nom latin de la plante ou Zizanion en grec ancien langue originale des évangiles)[95],[96].
Les graminées peuvent se révéler toxiques surtout pour les animaux herbivores dont elles constituent l'essentiel de l'alimentation. Cette toxicité peut être d'origine intrinsèque du fait de substances toxiques synthétisées ou accumulées par les plantes, soit extrinsèque par suite d'infections fongiques essentiellement.
Les substances toxiques accumulées par la plante peuvent être par exemple des oxalates, des alcaloïdes, des glycosides cyanogènes, des saponosides, etc[97].
L'infection par des champignons endophytes ou parasites peut provoquer chez différentes espèces de graminées des toxicités dangereuses pour l'alimentation humaine ou animale. C'est le cas par exemple de mycotoxines telles que celle produites par l'ergot du seigle ou certaines fusarioses.
Les graminées, du fait qu'elles sont très répandues sur toute la planète et qu'elles sont pollinisées par le vent, sont la famille de plantes qui produit le plus de pollen allergisant, provoquant chez l'homme diverses affections (pollinose, rhinite allergique). Les espèces les plus allergisantes appartiennent principalement à cinq sous-familles : Pooideae (Festucoideae), Chloridoideae (Eragrostoideae), Panicoideae, Arundinoideae et Oryzoideae. La morphologie très uniforme des grains de pollen de graminées ne permet pas de les distinguer facilement à un niveau infra-familial, sauf pour les variétés cultivées qui ont des grains de pollen plus gros que les types sauvages[98].
Les Poaceae, ou graminées, sont une famille de plantes monocotylédones de l'ordre des Poales, qui comprend environ 12 000 espèces groupées en 780 genres, à répartition cosmopolite. C'est, par le nombre d'espèces, la cinquième famille de plantes à fleurs, après les Asteraceae, Orchidaceae, Fabaceae et Rubiaceae. On y trouve la plupart des espèces appelées communément herbes et les céréales.
Ce sont généralement des plantes herbacées, plus rarement ligneuses (bambous), qui partagent des caractéristiques morphologiques qui les distinguent nettement des autres familles végétales : tiges (chaumes) cylindriques aux entrenœuds creux, feuilles alternes à disposition distique, au limbe linéaire à nervation parallèle, et dont la gaine enveloppe la tige, inflorescence élémentaire en épillets, fleurs réduites aux organes sexuels (étamines et ovaire), fruits dont le péricarpe est soudé à la graine (caryopses).
Les formations graminéennes, telles que les savanes et les prairies, dans lesquelles les Poaceae sont l'élément dominant, couvrent plus de 40 % de la surface terrestre (Groenland et Antarctique exclus). Les graminées forment également une part importante d'autres habitats, notamment des zones humides, des forêts et de la toundra.
C'est la famille de plantes la plus importante sur le plan économique, qui fournit une part essentielle de l'alimentation de base directement grâce aux espèces domestiquées comme les céréales (blé, riz, maïs, orge et millet), la canne à sucre, et indirectement grâce aux plantes fourragères, sans compter les matières utiles à l'industrie ou l'artisanat comme les bambous, la paille, le chaume et la biomasse (éthanol). Les graminées sont aussi cultivées pour l'agrément, notamment pour constituer des pelouses et des terrains de sport (golf), et pour lutter contre l'érosion des sols (oyat, vétiver). La famille compte aussi de nombreuses espèces adventices qui affectent les cultures, cinq espèces de Poaceae (Cynodon dactylon, Echinochloa colona, Eleusine indica, Sorghum halepense, Imperata cylindrica) figurant parmi les dix « pires » mauvaises herbes sur le plan mondial.
L'étude des phytolithes a montré que les Poaceae sont probablement apparues au cours du Crétacé, il y a plus de 85 millions d'années, faisant déjà partie de l'alimentation des dinosaures.
As gramíneas ou poáceas (Poaceae Barnhart) son unha familia de plantas herbáceas, ou moi raramente leñosas, pertencente á orde Poales das monocotiledóneas (Liliopsida). Con máis de 670 xéneros e preto de 10.000 especies descritas, as gramíneas son a cuarta familia con maior riqueza de especies logo das compostas (Asteraceae), as orquídeas (Orchidaceae) e as leguminosas (Fabaceae); mais, definitivamente, é a primeira en importancia económica global.[1] De feito, a maior parte da dieta dos seres humanos provén das gramíneas, tanto de xeito directo (grans de cereais e os seus derivados, como fariñas e aceites) ou indirecto (carne, leite e ovos que proveñen do gando e das aves de curral que se alimentan de pastos ou grans). É unha familia cosmopolita, que conquistou a maioría dos nichos ecolóxicos do planeta, desde as zonas desérticas até os ecosistemas de auga salgada, e desde as zonas deprimidas até os sistemas montañosos máis altos. Esta incomparable capacidade de adaptación está sustentada nunha enorme diversidade morfolóxica, fisiolóxica e reprodutiva e en varias asociacións mutualísticas con outros organismos, que converten ás gramíneas nunha fascinante familia, non só pola súa importancia económica, senón tamén pola súa relevancia biolóxica.
En xeral son herbas, aínda que poden ser leñosas como os bambús tropicais, cespitosas, rizomatosas ou estoloníferas. Pola duración do seu ciclo de vida poden ser anuais, bienais ou perennes. As gramíneas anuais, como é lóxico supor, reprodúcense unha soa vez durante o seu ciclo vital (o caso do trigo ou da avea, por exemplo). As especies perennes, en cambio, poden reproducirse varias veces (unha vez ao ano, normalmente) ou unha soa vez. No primeiro caso as especies denomínanse iteróparas (a maioría das especies de pastos. por exemplo) e no segundo caso, semélparas (as diferentes especies de bambús).
Teñen talos cilíndricos a elípticos na súa sección transversal, articulados, chamados ordinariamente canas, en xeral con nós macizos e entrenós ocos (mais poden ser totalmente macizos como no caso do millo e algúns bambús). Os nós son algo máis grosos que os entrenós e neles nacen as follas e as xemas. Os entrenós son ás veces algo achatados na zona onde se desenvolven as ramificacións. Un pouco máis arriba do nó existe un meristema intercalar en forma de anel que determina o alongamento do talo. Nalgúns xéneros existen de 2 a 6 nós moi próximos entre si (os cales se denominan nós compostos), cada un dos cales leva a súa correspondente folla. En Cynodon dactylon, por exemplo, os nós están en grupos de dous en dous polo que as follas parecen opostas. En xeral os entrenós basais son máis curtos que os superiores; cando hai varios nós basais moi próximos, as follas semellan arrosetadas, é dicir, disponse de xeito tal que simulan formar unha roseta basal de follas. Os principais tipos de talos nas gramíneas son os seguintes: Talos aéreos: Talos ascendentes, xeralmente con entrenós curtos na base e gradualmente máis longos cara ao ápice. Poden ser simples ou ramificados. Talos rastreiros: que medran deitados sobre o chan, arraigando nos nós; frecuentemente presentan nós compostos, como en Stenotaphrum e Cynodon. Cando os talos rastreiros orixinan novas plantas en cada nó denomínanse estolóns. Talos flotantes, os cales flotan na auga grazas aos entrenós ocos ou á presenza de aerénquima. Talos subterráneos:
Posúen follas de disposición alterna, dísticas, compostas tipicamente de vaíña, lígula e limbo. A vaíña rodea ao talo, as súas marxes superpóñense mais non se fusionan entre si (só ocasionalmente poden ser atopadas formando un tubo). A lígula é un pequeno apéndice membranoso, ou raramente un grupo de pelos, situado na zona de unión do limbo coa vaíña, na parte adaxial. O limbo (ou lámina) é simple, xeralmente lineal, con nervación paralela. Pode ser aplanado ou ás veces enrolado nun tubo, pode ser continuo coa vaíña ou posuír pecíolo. Ademais desta descrición, é necesario abordar a variabilidade que se pode achar para cada un destes órganos:
Os macollos ou macollas son a unidade estrutural da maioría das especies de gramíneas. Fórmanse a partir das xemas axilares ou secundarias do meristema basal do eixo principal. Cada un destes brotes secundarios ou macollos comenzan a súa aparición cando as plantas presentan entre dúas e tres follas. Cada un deles, logo de producir as súas primeiras follas, xera o seu propio sistema radicular. A suma ou adición de macollos é o que conforma a estrutura e a forma dunha planta de gramínea. Cando as gramíneas áchanse en estado vexetativo producen continuamente novos macollos e follas. Cada macollo, á súa vez, comezará no seu momento a producir novos macollos.
A inflorescencia elemental das gramíneas é unha pequena espiga formada por unha ou máis flores sentadas sobre un raque articulado, e protexido por brácteas estériles chamadas glumas. Este tipo de inflorescencia recibe o nome de espigueta. As flores poden ser hermafroditas ou unisexuais e presentan un perianto rudimentario de 2 ó 3 pezas, as lodículas ou glumélulas. Estas lodículas son os órganos que, determinan a apertura do antecio ou casa floral durante a floración, permitindo que se expoñan os estigmas e os estames. Os antecios están formados pola lemma, adherida á raquilla, e a pálea inserida sobre o eixo floral que nace sobre a raquilla e soporta aos órganos florais propiamente devanditos. A lemma (ou glumela inferior) ten forma de quilla, e abraza á pálea cos seus bordos. A aresta nace na extremidade da lemma ou no seu dorso. A pálea ou glumela superior é lanceolada, binervada e é como unha tapa que pecha á flor. As glumas están inseridas sobre a raquilla, unha máis abaixo que a outra. Todos estes elementos son moi variables polo que é conveniente analizalos por separado.
O froito ou gran das gramíneas é unha cariopse, froito seco, cunha semente cuxa testa está ligada co pericarpo formando unha envoltura moi delgada. Esta envoltura encerra ao embrión e ao endosperma. Este froito é basicamente unha variante do aquenio, aínda que se pode atopar unha variedade de froitos na familia (ver por exemplo Werker 1997). Nalgúns xéneros como Zizianopsis ou Eleusine, o pericarpio non está soldado coa semente, de modo que o froito é un aquenio (ou un utrículo segundo outros autores). Nalgunhas bambúseas o froito é unha noz ou unha baga, mentres no xénero Sporobolus o pericarpio deixa saír á semente cando se embebe en auga. Moitos xéneros, como Aristida, Stipa, Piptochaetium, Oryza e case todas as Paníceas posúen cariopses que se desprenden da planta envolvidos pola lemma e pola pálea. Nas Andropogóneas son as glumas as que persisten pechando á cariopse. En Pennisetum e Cenchrus despréndese toda a espiguilla rodeada dun invólucro de porcas ou de espiñas. A forma da cariopse varía moito segundo os xéneros, podendo ser case circular como en Briza, oblonga como en Hordeum, lanceolada como en Poa até case lineal, como en Vulpia. Na parte inferior da cariopse vista dorsalmente, vese o embrión máis ou menos elíptico cuberto polo pericarpio transparente. Do outro lado, correspondente ao asuco ou sutura carpelar, distínguese, tamén por transparencia, a mácula fiar ou fío (ou zona de unión da semente co carpelo), que pode ser punctiforme, como en Poa e nas Paníceas, ovada, como en Briza subaristata, ou lineal, como en Hordeum, Vulpia ou Festuca.
O tamaño e o número de cromosomas teñen grande importancia na sistemática das Gramíneas. Hai dous tipos cromosómicos extremos: o tipo festucoide caracterizado por presentar cromosomas grandes e número básico predominantemente x=7 e o tipo panicoide con cromosomas pequenos e números básicos predominantes x=9 e x=10. O tipo festucoide atópase en case todas as tribos da subfamilia Festucoideae (sen.: Pooideae), con algunhas excepcións. Por exemplo, a tribo Stipeae desta subfamilia posúe cromosomas pequenos e números básicos x=9, 10, 11, 12, 14, 16 e 17. Os restantes subfamilias das gramíneas presentan o tipo cromosómico panicoide, con cromosomas pequenos e predominio do número básico x=9 e 10. Nas Bambusóideas, ehrhartóideas e arundinóideas os cromosomas son pequenos e o número básico é x=12. Na subfamilia Danthonioideae preséntanse cromosomas de tamaño intermedio e número básico x=6 e 7. A subfamilia Chloridoideae presenta cromosomas pequenos e varios números básicos, x=7, 8, 9, 10, 11, 12 e 14. As Panicóideas sempre teñen cromosomas pequenos, con números básicos x=9 ou x=10, aínda que existen especies con outros números básicos, que varían desde x=4 a x=19.[3]
As gramíneas son morfoloxicamente distintas de calquera outra familia de plantas e, ademais, son moi diversas en canto a morfoloxía e hábito de crecemento. As diferentes especies de gramíneas - como se describiu na sección previa - difiren nos seus tamaños e números cromosómicos. Así mesmo, difiren no tamaño (ou contido de ADN) dos seus xenomas. O xenoma do arroz, por exemplo, é máis de 11 veces máis pequeno que o xenoma da cebada, a pesar de que ambas as especies son diploides e aparentan ter a mesma complexidade morfolóxica e fisiolóxica.[5] O contido de xenes das diferentes especies de gramíneas, no entanto, non varia tan amplamente como o contido de ADN total. O arroz e a cebada, novamente, non difiren máis que en 2 veces no número media de fragmentos de restrición que hibridan coas mesmas sondas.[6] A maior parte das diferenzas no tamaño do xenoma entre especies de gramíneas débese a diferenzas no ADN repetitivo. Os xenomas máis grandes, como os de cebada ou trigo, están compostos nun 75% de ADN repetitivo, mentres que os xenomas máis pequenos, como o do arroz, só conteñen menos do 50% de ADN altamente repetitivo. Máis aínda, determinouse que boa parte dese ADN repetitivo está composto de retrotransposóns insertos entre os xenes.[7][8] A maior parte das diferenzas no tamaño do xenoma entre especies de gramíneas débese a diferenzas no ADN repetitivo. Os xenomas máis grandes, como os de cebada ou trigo, están compostos nun 75% de ADN repetitivo, mentres que os xenomas máis pequenos, como o do arroz, só conteñen menos do 50% de ADN altamente repetitivo. Máis aínda, determinouse que boa parte dese ADN repetitivo está composto de retrotransposóns insertos entre os xenes.[7][8] Os estudos de mapeo xenómico en moitas especies de gramíneas empregando as mesmas sondas de ADN amosaron que non só o contido de xenes está moi conservado, senón tamén a orde dos xenes dentro dos cromosomas. A extensa conservación no contido de xenes e na orde dos mesmos entre o millo e o sorgo non é inesperada xa que ambas as especies "só" contan con 15 a 20 millóns de anos de evolución independente. No entanto, similares observacións para o arroz e o millo, as cales afástaronse hai de 60 a 80 millóns de anos, indican que todas as especies da familia proveñen dun mesmo antepasado común e que todas elas conservan un mesmo repertorio de xenes no mesmo orde aproximado.[9][10][11] Os grandes arranxos xenómicos que diferencian entre si a todas as gramíneas son o resultado de investimentos ou duplicacións cromosómicas que involucran a maior parte dos brazos cromosómicos.[12] A maioría, senón todas, as gramíneas son poliploides. Baseados no suposto que todos os xéneros e familias que presentan un número cromosómico básico x=12 son derivados de devanceiros que sufriron duplicacións cromosómicas durante a súa evolución e que as subfamilias de gramíneas máis primitivas (Anomochlooideae, Pharoideae, e Puelioideae) teñen un número cromosómico básico x=12, pódese deducir que o devanceiro das gramíneas xa era un poliploide. Séguese, ademais, que todas as gramíneas que se clasifican como diploides son, en realidade, paleopoliploides (ou sexa, poliploides antigos que presentan herdanza disómica.[13] e cuxos proxenitores non poden ser identificados mediante ferramentas citogenéticas ou marcadores moleculares).[14][15] A familia contén máis do 60% de especies, distribuídas en todos os clados, que se clasifican como neopoliploides, ou sexa que sufriron un ciclo adicional de duplicación xenómica. Nestas especies, os xenomas duplicados non han divergido moito do xenoma dos seus devanceiros e o seu número de cromosomas e comportamento citológico durante a meiose son indicativos da duplicación cromosómica que os orixinou. A maioría destes neopoliploides (máis do 65%) derivaron de cruzamientos interespecíficos ou intergenéricos polo que se lles clasifica como alopoliploides.[14]
As hemicelulosas e os polisacáridos de pectinas da parede celular primaria dos pastos son moi diferentes dos das demais espermatófitas, tanto en estrutura como nas particularidades da composición dos xiloglucanos. Os polisacáridos son menos ramificados que en todas as demais familias de plantas, aínda que esta afirmación está baseada nunha mostraxe aínda escasa de especies. As poáceas poden ser cianoxenéticas ou non. Cando son cianoxenéticas, os compostos cianoxenéticos son derivados da tirosina. Poden presentar alcaloides (ás veces isoquinolina, pirrolizidina e indol). Raramente pode haber proantocianidinas e cianidinas, en cantidades traza, e só en representantes das subfamilias Panicoideae e Chloridoideae. Os flavonoides acháronse só nalgúns xéneros, Bouteloua Glyceria e Melica,[16] cando están presentes son quercetina, ou kaempferol xunto con quercetina.
Unha xeneralización acerca do modo de reprodución das gramíneas é que os membros desta familia son plantas hermafroditas, que presentan fertilización cruzada (son alógamos) e se polinizan polo vento. Obviamente, unha familia con preto de 10.000 especies conta con moitas excepcións a esta regra, as cales se describen a continuación.
Este tipo de sistema reprodutivo, no cal existen plantas femininas e plantas masculinas, non é moi frecuente nas gramíneas. Só 18 xéneros son dioicos ou presentan especies dioicas, sendo Poa o máis coñecido deles. De feito, as especies dioicas de Poa inclúense nun subxénero afastado, Dioicopoa.[17]
Este sistema reprodutivo describe o feito de que nas poboacións naturais dunha especie coexisten individuos femininos e individuos hermafroditas. Esta condición é bastante rara nas gramíneas. Bouteloua chondrosioides e algunhas especies do subxénero Andinae de Poa son xinodioicas, aínda que Cortadeira é o exemplo máis conspicuo.[17]
Neste sistema os sexos están separados espacialmente pero no mesmo individuo, ou sexa, cada planta presenta inflorescencias femininas e masculinas. Zea, Humbertochloa, Luziola, Ekmanochloa e Mniochloa son exemplos de xéneros con especies monoicas. Moito máis común entre as gramíneas son as especies andromonoicas, unha condición moi común nas andropogóneas e paníceas. Nas primeiras, os dous sexos preséntanse en espiguillas diferentes de pares heterógamos. Un par heterógamo de espiguillas consiste xeralmente nunha espiguilla sésil, cunha flor neutra e outra hermafrodita, e unha espiguilla pedicelada, cunha flor neutra e outra masculina. Nas espiguillas bifloras das Paniceas, en troques, a flor inferior é xeralmente masculina ou neutra, e o superior é hermafrodita. Algúns dos xéneros que exemplifican este tipo de sistema son Alloteropsis, Brachiaria, Cenchrus, Echinochloa, Melinis, Oplismenus, Panicum, Setaria, Whiteochloa, e Xyochlaena. Algunhas especies dentro destes xéneros poden ter só flores hermafroditas xa que a flor inferior é sempre neutra, raramente ambas as flores son hermafroditas. á parte das paníceas e as andropogóneas, Arundinelleae é outra tribo con especies andromonoicas. No resto da familia, as especies andromonoicas atópanse moi esporádicamente, por exemplo en Arrhenatherum, Hierochloe e Holcus.[17]
A gran maioría das especies de gramíneas son hermafroditas, no entanto, frecuentemente son incapaces de producir sementes cando o pole dunha planta poliniza os seus propios estigmas. Isto débese a que unha gran parte das especies da familia presentan autoincompatibilidade, de tipo gametofítica e debido á acción de dous xenes independentes (chamados S e Z) con varios alelos cada un. Este sistema de autoincompatibilidade foi observado en varios xéneros da familia (Festuca, Secale, Lolium, Hordeum, Dactylis, entre moitos outros) e non é perfectamente eficiente. De feito, da maioría das especies autoincompatibles pode obterse unha proporción - aínda que reducida - de sementes ao autofecundar unha planta.[17][18]
A autopolinización e a autofecundación están moi distribuídas entre as gramíneas. En xeral, é un mecanismo máis común entre as especies anuais que entre as perennes e, decididamente, moito máis frecuente entre as especies colonizadoras. Este mecanismo determinouse en aproximadamente 45 xéneros de gramíneas, entre os cales se achan xéneros economicamente moi importantes como Triticum, Oryza, Secale, Avea, Agropyron e Lolium. Unha condición de autogamia extrema é a cleistogamia. Este último sistema está distribuído en máis de 70 xéneros pertencentes a 20 tribos de gramíneas.[17][19]
A apomixia defínese como a reprodución asexual a través de sementes. Neste sistema reprodutivo os embrións desenvólvense por mitose a partir dunha oosfera non reducida sen que teña lugar a fecundación. Noutras palabras, cada embrión producido é xeneticamente idéntico a plánta nai. Nas gramíneas, a apomixia foi descrita por primeira vez en 1933 nunha especie de Poa. Desde aquel momento identificouse este mecanismo en centos de especies de poáceas, particularmente nas Paníceas e nas Andropogóneas. Algúns dos xéneros que presentan especies apomícticas son Apluda, Capillipedium, Heteropogon, Themeda, Sorghum, Bothriochloa, Dichanthium, Cenchrus, Setaria e Paspalum.[17][20][21]
As gramíneas ou poáceas (Poaceae Barnhart) son unha familia de plantas herbáceas, ou moi raramente leñosas, pertencente á orde Poales das monocotiledóneas (Liliopsida). Con máis de 670 xéneros e preto de 10.000 especies descritas, as gramíneas son a cuarta familia con maior riqueza de especies logo das compostas (Asteraceae), as orquídeas (Orchidaceae) e as leguminosas (Fabaceae); mais, definitivamente, é a primeira en importancia económica global. De feito, a maior parte da dieta dos seres humanos provén das gramíneas, tanto de xeito directo (grans de cereais e os seus derivados, como fariñas e aceites) ou indirecto (carne, leite e ovos que proveñen do gando e das aves de curral que se alimentan de pastos ou grans). É unha familia cosmopolita, que conquistou a maioría dos nichos ecolóxicos do planeta, desde as zonas desérticas até os ecosistemas de auga salgada, e desde as zonas deprimidas até os sistemas montañosos máis altos. Esta incomparable capacidade de adaptación está sustentada nunha enorme diversidade morfolóxica, fisiolóxica e reprodutiva e en varias asociacións mutualísticas con outros organismos, que converten ás gramíneas nunha fascinante familia, non só pola súa importancia económica, senón tamén pola súa relevancia biolóxica.
Trave (travovke, prave trave, lat. Poaceae) je naziv za porodicu biljaka jednosupnica koje obilježava neprimjetni cvijet, dugo i usko lišće. Botanički pripadaju redu travolike (Poales).
Trave služe kao hrana brojnih životinja, pogotovo preživača i konja, a neke vrste i u prehrani ljudi, kao što je kukuruz i druga žita, a raširene su po svim naseljenim kontinentima. Obuhvaća 759 poznatih rodova sa ukupno 11.554 priznate vrste među kojima pšenica, riža, zob, etc:
Trave (travovke, prave trave, lat. Poaceae) je naziv za porodicu biljaka jednosupnica koje obilježava neprimjetni cvijet, dugo i usko lišće. Botanički pripadaju redu travolike (Poales).
Trave služe kao hrana brojnih životinja, pogotovo preživača i konja, a neke vrste i u prehrani ljudi, kao što je kukuruz i druga žita, a raširene su po svim naseljenim kontinentima. Obuhvaća 759 poznatih rodova sa ukupno 11.554 priznate vrste među kojima pšenica, riža, zob, etc:
Słódke trawy (Poaceae) su swójba symjencowych rostlinow (Spermatophyta).
Wobsahuje sćěhowace rody:
Słódke trawy (Poaceae) su swójba symjencowych rostlinow (Spermatophyta).
Wobsahuje sćěhowace rody:
arundo (Arundo) hoberska sćina (Arundo donax) bambus (Bambusa) arundobodobna bambusnica (Bambusa arundinaceae) błysknica (Phalaris) hłójčkata błysknica (Phalaris canariensis) wódna błysknica (Phalaris arundinacea) brodačk (Danthonia) wšědny brodačk (Danthonia decumbens) brodawa (Bothriochloa) citronowa brodawa (Bothriochloa nardus) wšědna brodawa (Bothriochloa ischaemum) cokorina (Saccharum) prawa cokorina (Saccharum officinarum) česačk (Cynosurus) jěžowy česačk (Cynosurus echinatus) pastwišćowy česačk (Cynosurus cristatus) dornica (Festuca) alpska dornica (Festuca alpina) ametystowa dornica (Festuca amethystina) blěda dornica (Festuca pallens) ćeńkołopjenata dornica (Festuca trichophylla) ćěmna dornica (Festuca nigrescens) čerwjena dornica (Festuca rubra) čorna dornica (Festuca puccinellii) dołhołopjenata dornica (Festuca longifolia) duvalowa dornica (Festuca duvalii) fijałkomódra dornica (Festuca violacea) hładka dornica (Festuca laevigata) hruba dornica (Festuca brevipila) hruba wowča dornica (Festuca heteropachys) kamzykowa dornica (Festuca rupicaprina) kosmata dornica (Festuca rupicola) łučna dornica (Festuca pratensis) lěsna dornica (Festuca altissima) mała dornica (Festuca airoides) mnohokwětkata čerwjena dornica (Festuca heteromalla) módrozelena dornica (Festuca glaucina) nawěwowa dornica (Festuca polesica) něžna dornica (Festuca pulchella) niska dornica (Festuca quadriflora) njeprawa wowča dornica (Festuca pulchra) noriska dornica (Festuca norica) patzkowa dornica (Festuca patzkei) pisana dornica (Festuca varia) sćinojta dornica (Festuca arundinacea) šwižna dornica (Festuca heterophylla) waliziska dornica (Festuca valesiaca) włosata dornica (Festuca filiformis) wowča dornica (Festuca ovina) wysoka dornica (Festuca gigantea) dornica (Micropyrum) jednora dornica (Micropyrum tenellum) dornicowa sćina (Scolochloa) prawa dornicowa sćina (Scolochloa festucacea) dornička (Vulpia) mikawčkata dornička (Vulpia ciliata) myšaca dornička (Vulpia myuros) přibrjóžna dornička (Vulpia unilateralis) wjewjerča dornička (Vulpia bromoides) dubrowka (Hierochloë) awstralska dubrowka (Hierochloë australis) kosmata dubrowka (Hierochloë hirta) wonjata dubrowka (Hierochloë odorata) gawdina (Gaudinia) rozłamliwa gawdina (Gaudinia fragilis) chrósćica (Oreochloa) dwojorjadkata chrósćica (Oreochloa disticha) chrósćica (Psilathera) jejkojta chrósćica (Psilathera ovata) chrósćica (Sesleria) módra chrósćica (Sesleria albicans) jahły (Panicum) nahe jahły (Panicum dichotomiflorum) prawe jahły (Panicum miliaceum) ćeńkohałžkate jahły (Panicum capillare) jahlička (Digitaria) čerwjena jahlička (Digitaria sanguinalis) ćeńka jahlička (Digitaria ischaemum) jahlička (Echinochloa) kałačkojta jahlička (Echinochloa muricata) kurjaca jahlička (Echinochloa crus-galli) ječmjeń (Hordeum) hriwaty ječmjeń (Hordeum jubatum) myšacy ječmjeń (Hordeum murinum) přimórski ječmjeń (Hordeum maritinum) rožkowy ječmjeń (Hordeum secalinum) sywny ječmjeń (Hordeum vulgare) jiłowka (Spartina) jendźelska jiłowka (Spartina anglica) prerijowa jiłowka (Spartina pectinata) wysoka jiłowka (Spartina x townsendii) koleantus (Coleanthus) wšědny koleantus (Coleanthus subtilis) korakan (Eleusine) dźiwi korakan (Eleusine indica) kostrjawa (Bromus) andska kostrjawa (Bromus catharticus) hłucha kostrjawa (Bromus sterilis) japanska kostrjawa (Bromus japonicus) kaliforniska kostrjawa (Bromus carinatus) kitkojta kostrjawa (Bromus racemosus) krótka kostrjawa (Bromus brachystachys) lěsna kostrjawa (Bromus ramosus) mjechka kostrjawa (Bromus hordeaceus) něžna kostrjawa (Bromus lepidus) niska kostrjawa (Bromus tectorum) přeměnita kostrjawa (Bromus commutatus) rólna kostrjawa (Bromus arvensis) runa kostrjawa (Bromus erectus) srjedźomórska kostrjawa (Bromus madritensis) šepjerjaca kostrjawa (Bromus squarrosus) tołsta kostrjawa (Bromus grossus) tupa kostrjawa (Bromus inermis) wisata kostrjawa (Bromus patulus) wulka kostrjawa (Bromus diandrus) zažna kostrjawa (Bromus benekenii) žitowa kostrjawa (Bromus secalinus) kostrjenka (Brachypodium) lěsna kostrjenka (Brachypodium sylvaticum) pjerkojta kostrjenka (Brachypodium pinnatum) skalna kostrjenka (Brachypodium rupestre) kowjel (Stipa) kosmičkata kowjel (Stipa dasyphylla) něžna kowjel (Stipa eriocaulis) pěskowa kowjel (Stipa borysthenica) pjerkojta kowjel (Stipa pennata) włosojta kowjel (Stipa capillata) wopuškojta kowjel (Stipa tirsa) wulka kowjel (Stipa pulcherrima) kowjel (Achnatherum) smjelzynojta kowjel (Achnatherum calamagrostis) kukurica (Zea) prawa kukurica (Zea mays) lipnica (Poa) alpska lipnica (Poa alpina) badenska lipnica (Poa badensis) cybličkata lipnica (Poa bulbosa) ćeńka lipnica (Poa nemoralis) drapata lipnica (Poa trivialis) dwojorjadkata lipnica (Poa cenisia) hórska lipnica (Poa chaixii) łučna lipnica (Poa pratensis) langowa lipnica (Poa langiana) mała lipnica (Poa minor) mólička lipnica (Poa annua) niska lipnica (Poa supina) přeměnita lipnica (Poa hybrida) róznokłosata lipnica (Poa remota) sucha lipnica (Poa compressa) šěra lipnica (Poa caesia) wisata lipnica (Poa laxa) wódna lipnica (Poa palustris) wuska lipnica (Poa angustifolia) liščak (Alopecurus) łučny liščak (Alopecurus pratensis) pucherkojty liščak (Alopecurus rendlei) rólny liščak (Alopecurus myosuroides) ryzy liščak (Alopecurus aequalis) stawacy liščak (Alopecurus geniculatus) sćinojty liščak (Alopecurus arundinaceus) sučkaty liščak (Alopecurus bulbosus) lubočinka (Eragrostis) etiopiska lubočinka (Eragrostis tef) japanska lubočinka (Eragrostis multicaulis) łobjowa lubočinka (Eragrostis albensis) mała lubočinka (Eragrostis minor) włosata lubočinka (Eragrostis pilosa) wulka lubočinka (Eragrostis cilianensis) mědlina (Holcus) łučna mědlina (Holcus lanatus) lěsna mědlina (Holcus mollis) motawa (Lolium) hładka motawa (Lolium perenne) jědojta motawa (Lolium temulentum) lenowa motawa (Lolium remotum) wulka motawa (Lolium multiflorum) pasnik (Phleum) alpski pasnik (Phleum alpinum) bayerski pasnik (Phleum hirsutum) drapaty pasnik (Phleum paniculatum) dulojty pasnik (Phleum bertolonii) hórski pasnik (Phleum rhaeticum) łučny pasnik (Phleum pratense) pěskowy pasnik (Phleum arenarium) suchi pasnik (Phleum phleoides) paćernik (Melica) drobny paćernik (Melica uniflora) mikawčkaty paćernik (Melica ciliata) pisany paćernik (Melica picta) transsilvaniski paćernik (Melica transsilvanica) wisaty paćernik (Melica nutans) wysoki paćernik (Melica altissima) pěskač (Tragus) kićowy pěskač (Tragus racemosus) pjerchowanka (Milium) lěsna pjerchowanka (Milium effusum) porčizna (Glyceria) hybridiska porčizna (Glyceria x pedicellata) lěsna porčizna (Glyceria nemoralis) módrozelena porčizna (Glyceria declinata) ropata porčizna (Glyceria notata) słódka porčizna (Glyceria fluitan) smuhata porčizna (Glyceria striata) wulka porčizna (Glyceria maxima) prostak (Molinia) sćinowy prostak (Molinia arundinacea) wšědny prostak (Molinia caerulea) psyči zubik (Cynodon) wšědny psyči zubik (Cynodon dactylon) pšeńca (Triticum) jednozorno (Triticum monococcum) krupnik (Triticum spelta) pólska pšeńca (Triticum polonicum) prawa pšeńca (Triticum aestivum) přimórski wows (Ammophila) wšědny přimórski wows (Ammophila arenaria) pyr (Elytrigia) drobnokłóskaty pyr (Elytrigia x oliveri) kałačkojty pyr (Elytrigia x mucronata) módrozeleny pyr (Elytrigia intermedium) nachileny pyr (Elytrigia x laxa) pěskowy pyr (Elytrigia arenosa) pontiski pyr (Elytrigia obtusiflora) přibrjóžny pyr (Elytrigia atherica) syćowy pyr (Elytrigia junceiformis) runy pyr (Elytrigia x obtusiuscula) wšědny pyr (Elytrigia repens) pyr (Roegneria) rěčny pyr (Roegneria canina) rajs (Oryza) prawy rajs (Oryza sativa) rjapuška (Sclerochloa) wšědna rjapuška (Sclerochloa dura) rožka (Secale) jerica (Secale aestivum) sywna rožka (Secale cereale) sćina (Phragmites) Schilf wšědna sćina (Phragmites australis) seršćik (Nardus) wšědny seršćik (Nardus stricta) smjelzyna (Calamagrostis) alpska smjelzyna (Calamagrostis varia) bahnowa smjelzyna (Calamagrostis canescens) hatna smjelzyna (Calamagrostis lanceolata) kosmata smjelzyna (Calamagrostis villosa) łuhojta smjelzyna (Calamagrostis stricta) lěsna smjelzyna (Calamagrostis arundinacea) něžna smjelzyna (Calamagrostis tenella) pěskowa smjelzyna (Calamagrostis epigejos) přirěčna smjelzyna (Calamagrostis pseudophragmites) purpurowa smjelzyna (Calamagrostis phragmitoides) sakska smjelzyna (Calamagrostis pseudopurpurea) sólnica (Puccinellia) bahnowa sólnica (Puccinellia limosa) ćeńka sólnica (Puccinellia capillaris) přimórska sólnica (Puccinellia maritima) wotstejaca sólnica (Puccinellia distans) sukač (Dactylis) łučny sukač (Dactylis glomerata) lěsny sukač (Dactylis polygama) šěrawka (Koeleria) běłojta šěrawka (Koeleria arenaria) jehłancojta šěrawka (Koeleria pyramidata) pólska šěrawka (Koeleria grandis) šěrozelena šěrawka (Koeleria glauca) šwižna šěrawka (Koeleria macrantha) waliziska šěrawka (Koeleria vallesiana) šmica (Deschampsia) bahnowa šmica (Deschampsia wibeliana) dwojobarbna šmica (Deschampsia discolor) łučna šmica (Deschampsia cespitosa) lěsna šmica (Deschampsia flexuosa) pobrjóžna šmica (Deschampsia littoralis) seršćowa šmica (Deschampsia setacea) syćinowa šmica (Deschampsia media) šmička (Aira) chuda šmička (Aira praecox) něžna šmička (Aira elegantissima) pyšna šmička (Aira caryophyllea) šćětkowy lišćak (Polypogon) wšědny šćětkowy lišćak (Polypogon monspeliensis) šćótka (Corynephorus) wšědna šćótka (Corynephorus canescens) tajawka (Leersia) wšědna tajawka (Leersia oryzoides) tradwička (Parapholis) zhibnjena tradwička (Parapholis strigosa) trawička (Mibora) pěskowa trawička (Mibora minima) třasawka (Apera) přetorhnjena třasawka (Apera interrupta) wšědna třasawka (Apera spica-venti) třepjetawka (Briza) wšědna třepjetawka (Briza media) twjerdačk (Catapodium) wšědny twjerdačk (Catapodium rigidum) wjechelca (Agrostis) alpska wjechelca (Agrostis alpina) běła wjechelca (Agrostis stolonifera) čerwjena wjechelca (Agrostis capillaris) hoberska wjechelca (Agrostis gigantea) hruba wjechelca (Agrostis scabra) kastiliska wjechelca (Agrostis castellana) něžna wjechelca (Agrostis agrostiflora) pochowa wjechelca (Agrostis canina) schleicherowa wjechelca (Agrostis schleicheri) skalna wjechelca (Agrostis rupestris) wuskokitkata wjechelca (Agrostis vinealis) włosanc (Setaria) čerwjeny włosanc (Setaria pumila) faberowy włosanc (Setaria faberi) kosmaty włosanc (Setaria verticilliformis) seršćojty włosanc (Setaria vertillata) zeleny włosanc (Setaria viridis) zornjaty włosanc (Setaria italica) wonjanka (Anthoxanthum) alpska wonjanka (Anthoxanthum alpinum) kóchtata wonjanka (Anthoxanthum aristatum) łučna wonjanka (Anthoxanthum odoratum) wows (Avena) krótki wows (Avena nuda subsp. brevis) nahi wows (Avena nuda subsp. nuda) njerodny wows (Avena fatua) rólny wows (Avena sativa) smuhaty wows (Avena nuda subsp. strigosa) zmjećeny wows (Avena planiculmis) wows (Helictotrichon) łučny wows (Helictotrichon pratensis) mjechki wows (Helictotrichon pubescens) pisany wows (Helictotrichon versicolor) wutrajny wows (Helictotrichon parlatoreï) wowsnik (Arrhenatherum) wšědny wowsnik (Arrhenatherum elatius) wowsyčk (Trisetum) dwojorjadkaty wowsyčk (Trisetum distichophyllum) kłóskaty wowsyčk (Trisetum spicatum) židźany wowsyčk (Trisetum flavescens) wowsyčk (Ventenata) šmicojty wowsyčk (Ventenata dubia) zbožowna trawa (Beckmannia) ameriska zbožowna trawa (Beckmannia syzigachne) sibirska zbožowna trawa (Beckmannia eruciformis) žórłowka (Catabrosa) wšědna žórłowka (Catabrosa aquatica)Suku padi-padian, Poaceae, Glumiflorae, atau Graminae adalah salah satu suku anggota tumbuhan berbunga. Menurut sistem klasifikasi APG II suku ini termasuk ke dalam bangsa Poales, klad commelinids (euMonokotil).
Anggota suku ini adalah yang paling tinggi populasinya di dunia karena banyak tanaman budidaya yang menjadi anggotanya dan ditanam luas sebagai bahan pangan utama. Di dalamnya termasuk tumbuhan seperti padi, gandum, jagung, jelai, jewawut, serta sorgum (cantel). Selain itu, bambu dan tebu juga termasuk di dalamnya. Bahan pakan ternak juga banyak memanfaatkan anggota suku ini, seperti rumput gajah dan rumput raja. Anggota suku ini beberapa di antaranya merupakan tumbuhan pengganggu (gulma) yang penting, seperti alang-alang dan rumput bandotan. Ada anggotanya yang merupakan sumber wangi-wangian, yaitu rumput akar wangi dan serai (termasuk sitronela).
Pada umumnya, ciri tumbuhan Poaceae berbatang beruas-ruas, bunga tak bermahkota, serta daun berbentuk pita.
Secara taksonomi, suku rumput-rumputan dibagi menjadi tujuh anaksuku yang masing-masing memiliki kekhasan yang dapat dengan mudah dilihat:
Suku padi-padian, Poaceae, Glumiflorae, atau Graminae adalah salah satu suku anggota tumbuhan berbunga. Menurut sistem klasifikasi APG II suku ini termasuk ke dalam bangsa Poales, klad commelinids (euMonokotil).
Grasaætt (fræðiheiti: Poaceae, áður Gramineae) er ætt einkímblöðunga. Um 600 ættkvíslir eru innan grasaættarinnar og á milli 9 og 10 þúsund tegundir grasa. Grasaættin er ein mikilvægasta fjölskylda plantna í heiminum, enda gefur hún fóður dýra og næringu manna auk bambusreyrsins sem notaður er til bygginga í Asíu. Áætlað er að 20% af yfirborði jarðar séu þakin tegundum úr grasaættinni. Tegundir af grasaætt hafa aðlagað sig fjölbreytilegum aðstæðum sem ríkja á mismunandi stöðum í heiminum og finnast frá köldustu svæðum jarðar til regnskóga hitabeltisins og eyðimarka.
Grös hefja lífsferil sinn með geldvexti. Hlutverk grasplöntu í geldvexti er aðallega að mynda rætur og blöð. Sjálfur stöngullinn (einnig kallaður vaxtarbroddur) er aðeins örfáa millimetrar á lengd og sést ekki með berum augum. Hann er liðskiptur og vex ávallt rót eða blað út úr hverjum lið. Rætur myndast eðlilega út frá neðstu liðum stönguls en blöð út frá efri liðum. Fyrsta blaðið vex út frá neðsta lið sem ekki myndar rót. Blaðið myndar hólk utan um stöngulinn og vex uppávið.
Blað grasa skiptist í tvennt: blaðslíður og blöðku. Blaðslíðrið myndar hólk en blaðkan er efri hluti blaðsins og breiðir úr sér. Næsta blaðið myndast út frá næstneðsta lið stöngulsins og hefur vöxt sinn innan við eldra blaðið. Eftir því sem blaðið stækkar vex það upp úr blaðslíðri elsta blaðsins og hækkar upp fyrir það. Svona heldur vöxtur blaða áfram koll af kolli, þar sem yngsta blaðið kemur ávallt innan úr næstyngsta blaðinu.
Mörg blaðslíður myndan þá hólk sem kallast gervistrá, þar sem ekki er um raunverulegt strá er að ræða. Stöngullinn sjálfur er enn örfáa millimetrar á lengd og leynist neðst við jörðina innan í blaðslíðrunum. Á mótum blaðslíðurs og blöðkunnar finnst oftast lítil himna, slíðurhimna. Hún er gott greiningareinkenni grasa.
Flestar plöntur fara í kynvöxt þegar réttar aðstæður ríkja. Ýmsar breytingar verða á vaxtarhætti grassprota við það. Stöngullinn hættir að mynda nýja blaðvísa en myndar þess í stað blómvísa á enda þess. Stöngulliðirnir fara nú að lengjast í þeim tilgangi að ýta blóminu upp úr yngsta blaðslíðrinu. Við það hækkar plantan umtalsvert. Stöngulliðirnir bólgna upp og mynda svokölluð hné. Til þess að halda uppi blómskipuninni sem verður sífellt þyngri eftir því sem hún stækkar, myndar stöngullinn tormelt efni, svo sem lignín og tréni.
Blóm grasa raða sér saman í smáöx. Grösum er skipt í þrjá flokka eftir blómskipun þeirra, það er á hvaða hátt smáöxin raða sér á stöngulinn; axgrös, axpuntgrös og puntgrös.
Grös hafa þann hæfileika að mynda dóttursprota. Þeir myndast inn á milli blaða og stönguls á móðursprotanum og getur einn dóttursproti myndast við hvert blað á móðursprotanum. Fjöldi dóttursprota er því háður fjölda blaða á móðursprotanum. Það eru þó ekki allir dóttursprotar sem ná að dafna og stækka. Það er háð næringarástandi móðursprotans, hversu há gróðurinn er (skuggamyndun) eða hversu snöggklipptur hann er eftir beit eða slátt. Eftir að móðursprotinn er farinn í kynvöxt hefur hann forgang og dóttursprotar þurfa að bíða afgangs. Þeir sem hafa náð að mynda sjálfir rætur hafa hugsanlega tök á að dafna og stækka, aðrir bíða betri tíma. Ef stöngulendi sprota hefur verið fjarlægður með beit og/eða slátt eru það dóttursprotarnir sem taka við og viðhalda grassverðinum. Dóttursprotum er skipt í tvo flokka, eftir því hvort þeir vaxa innanslíðurs eða utanslíðurs. Plöntur er mynda innanslíðurs dótturspota eru oftast með þúfukenndan vöxt, eins og til dæmi snarrótarpuntur.
Hjá plöntum með utanslíðurs dóttursprotum lengjast neðstu stöngulliðir dóttursprotans mikið og brjóta sér leið í gegnum blaðslíðrin móðursprotans. Er þá talað um renglur og geta þær ýmist vaxið ofanjarðar eða neðanjarðar. Sem dæmi má nefna vallarsveifgras sem myndar öflugar neðanjarðarrenglur og skriðlíngresi sem myndar ofanjarðarrenglur.
Helstu tegundir grasaættar flokkaðar eftir nýtingu.
Grasaætt (fræðiheiti: Poaceae, áður Gramineae) er ætt einkímblöðunga. Um 600 ættkvíslir eru innan grasaættarinnar og á milli 9 og 10 þúsund tegundir grasa. Grasaættin er ein mikilvægasta fjölskylda plantna í heiminum, enda gefur hún fóður dýra og næringu manna auk bambusreyrsins sem notaður er til bygginga í Asíu. Áætlað er að 20% af yfirborði jarðar séu þakin tegundum úr grasaættinni. Tegundir af grasaætt hafa aðlagað sig fjölbreytilegum aðstæðum sem ríkja á mismunandi stöðum í heiminum og finnast frá köldustu svæðum jarðar til regnskóga hitabeltisins og eyðimarka.
Le Poacee, comunemente conosciute come Graminacee[1] (Poaceae [R.Br.] Barnhart, 1895 [2] o Gramineae [nomen cons.] Juss.), sono una famiglia di piante angiosperme monocotiledoni appartenente all'ordine Cyperales.
Nella regione mediterranea, allo stato spontaneo, pur localizzandosi soprattutto in ambienti aperti, occupano praticamente tutti i tipi di habitat, dai boschi ai luoghi umidi, dalle dune sabbiose agli ambienti rurali.
Si spingono a tutte le latitudini, con notevole escursione altimetrica. Sono piante perfettamente adattate ai climi aridi, anche se non mancano specie mesofile e altre persino acquatiche. Notevole l'importanza nel quadro del rivestimento vegetale della terra: costituiscono fitte formazioni vegetali molto estese alle quali conferiscono una particolare fisionomia come praterie, savane, steppe.
Dominano i biomi delle savane, delle praterie e delle steppe e interi gruppi animali (es. ungulati) si sono evoluti contestualmente. Particolare importanza assumono anche nella storia e nell'economia umana. La coltura dei cereali è alla base dello sviluppo delle prime società civilizzate.
Si tratta di piante erbacee, perenni o annuali. L'habitus dominante è rizomatoso erbaceo.
Il fusto (denominato culmo) è articolato in nodi e internodi. Nella maggior parte delle specie è cavo negli internodi e pieno ai nodi. Poche specie (es. mais) presentano fusti pieni anche agli internodi. A livello dei nodi si originano le foglie e le ramificazioni laterali. Il culmo non si ramifica salvo che in basso; una porzione meristematica in corrispondenza del nodo conferisce al culmo la capacità di raddrizzarsi nel caso venga flesso. Molte Graminacee mostrano il fenomeno dell'accestimento: da un rizoma perenne si formano, anche nella stessa annata, più culmi vicini tra loro.
Le foglie presentano una guaina, che avvolge il culmo e un lembo, che si stacca nettamente dalla guaina in corrispondenza di una piccola struttura membranosa detta ligula. La guaina presenta una incisione per tutta la sua lunghezza, i margini possono coprirsi per un breve tratto. Il lembo si distanzia dal fusto ad angolo ottuso e termina a punta, sovente presenta delle striature. La ligula è una membrana sottile posizionata tra il lembo e la guaina: può essere più o meno sviluppata o sostituita da un ciuffo di peli ed è importante come carattere diagnostico. In alcuni generi (es. Hordeum, Lolium) possono essere presenti delle auricole alla base della foglia.
I fiori sono sempre raccolti in particolari infiorescenze, distintive della famiglia, denominate spighette riunite a loro volta in spighe o pannocchie. L'impollinazione è anemofila (con conseguente perdita della funzione vessillare del fiore). Le Poaceae, sulla base della struttura della spighetta, vengono distinte in vari gruppi (tribù).
Il fiore possiede un asse centrale, rappresentato dal gineceo, supero, costituito da 2 (3) carpelli che delimitano una sola loggia (gineceo uniloculare) con un solo ovulo, uno stilo e 2(3) stimmi piumosi. Alla base dell'ovario si inseriscono gli stami (che formano l'androceo), quasi sempre in numero di tre, ognuno costituito da un filamento e da un'antera pendula. Talora i fiori sono unisessuali per mancanza del pistillo o sterili per aborto degli organi sessuali. L'ovario e gli stami sono avvolti da due glumette col valore di brattee fertili, di cui l'inferiore prende il nome di lemma, e la superiore, più piccola e generalmente racchiusa dalla precedente, viene detta palea. Il lemma può presentare, nella parte mediana, un angolo (carena) dalla quale può dipartirsi un'arista (talvolta in luogo dell'arista si trova un aculeo, oppure la glumetta si presenta mutica, ovvero priva di aculeo). Nel fiore si osservano i resti del perianzio, rappresentati da 2 (3) lodicule, di consistenza membranosa.
I fiori sono sempre raccolti in infiorescenze denominate spighette, distintive della famiglia. La spighetta, che può essere uniflora o, più frequentemente, pluriflora, è provvista di un asse (rachilla) portante alla base le glume, opposte, con il significato di brattee sterili. Le glume sono generalmente più piccole delle glumette e si distinguono in superiore e inferiore (lemma), generalmente di grandezza diversa. Le glume possono essere carenate e, più raramente delle glumette, portano aculei e ariste (dette anche reste) consistenti in prolungamenti filiformi. Le forme non aristate sono dette mutiche.
I fiori sono sempre raccolti in infiorescenze denominate spighette, distintive della famiglia, e riunite a loro volta in infiorescenze denominate:
Il frutto è una cariosside in cui l'albume è a diretto contatto con il frutto per digestione dei tegumenti seminali. L'albume è amilaceo e presenta la porzione più esterna formata da uno strato di cellule ricche di proteine (strato aleuronico). Il seme ha endosperma ricco di amido.
La sistematica di questa numerosa famiglia è complessa.
Comprende 733 generi riconosciuti e circa 11.600 specie di cui circa 350 italiane.
Secondo il sistema Cronquist la famiglia va collocata, insieme alle sole Cyperaceae, nell'ordine delle Cyperales.
Secondo la classificazione APG le Poaceae vengono incluse nell'ordine delle Poales, insieme a 15 altre famiglie di piante monocotiledoni.
La classificazione APG elenca 12 sottofamiglie:[3]
La classificazione è complicata dal fatto che, oltre a numerose sottofamiglie, vengono riconosciuti vari gruppi di livello tassonomico intermedio (tribù), soprattutto in base alla struttura della spighetta.
Le tribù principali sono:
Alcuni studiosi introducono anche supertribù o sottotribù. In totale, i vari gruppi di posizione intermedia tra la famiglia e il genere (sottofamiglie, supertribù, tribù e sottotribù), compresi i sinonimi, raggiungono il totale oltre 200 entità[4]
Della famiglia fanno parte i seguenti generi (lista non esaustiva):[5]
Le Poaceae possono causare allergie. Di seguito le specie che causano pollinosi:
Le Poaceae hanno un'enorme importanza nell'economia della biosfera: comprendono infatti la maggior parte dei cereali prodotti alla base dell'alimentazione umana.
Tipicamente, vengono consumati solo i semi (chicchi), previa cottura. In alcuni casi, i semi vengono consumati interi (procedimento comune in particolare per il riso, ma applicato anche ad altri cereali), mentre in altri casi essi vengono prima macinati, producendo una farina; alla farina vengono aggiunti acqua e in molti casi lievito, con risultati molto diversi (pane, pasta, focacce, pizza, polenta, porridge, cracker ecc.).
Diversi cereali sono usati anche per la produzione di bevande alcoliche. Soprattutto l'orzo, ma anche il grano, vengono usati per la produzione di birre. Cereali diversi (grano, orzo, segale) sono usati tradizionalmente per produrre la vodka, il whisky, il gin. Il riso è invece usato per il sakè. Una specie Hierochloe odorata (Erba del bisonte), ovvero la pianta di cui si cibano i bisonti europei, viene utilizzata in Polonia per la produzione di una vodka, la Żubrówka, nome locale della pianta in questione, derivante dal termine polacco Żubr (bisonte). Dalle lacrime di Giobbe vengono ricavati alcuni liquori cinesi e coreani.
Inoltre diversi cereali, insieme a piante erbacee di altre famiglie, sono usati per l'alimentazione del bestiame domestico.
I cereali usati come alimenti per l'uomo (prescindendo quindi dai cereali usati solo per il bestiame) sono:
Fuori dal campo alimentare, va segnalato soprattutto il bambù, nome collettivo che comprende diverse specie affini, usate come materiale da costruzione.
Alcune graminacee appartenenti al genere Bromus o la specie Avena sativa sono anche detti forasacchi e le loro piccole spighe possono infilarsi nelle orecchie dei cani, compromettendone a volte definitivamente l'udito; causano ascessi anche semplicemente conficcandosi tra le "dita" delle zampe dei cani e dei gatti.[6]
Le Poacee, comunemente conosciute come Graminacee (Poaceae [R.Br.] Barnhart, 1895 o Gramineae [nomen cons.] Juss.), sono una famiglia di piante angiosperme monocotiledoni appartenente all'ordine Cyperales.
Poaceae, olim Gramineae, sunt familia plantarum florentium classis Liliopsidorum (monocotyledonum). Sunt in familia circa 600 genera et fere 9000–10 000 vel plus species (Kew Index of World Grass Species).
Monocotyledones
Commelinidae
Ordo : Poales
Familia : Poaceae
Poaceae, olim Gramineae, sunt familia plantarum florentium classis Liliopsidorum (monocotyledonum). Sunt in familia circa 600 genera et fere 9000–10 000 vel plus species (Kew Index of World Grass Species).
Migliniai, Varpiniai (lot. Poaceae, Gramineae, vok. Süßgräser) – magnolijūnų (Magnoliophyta) skyriui priklausanti labai didelė augalų grupė, kurią sudaro apie 500 genčių, apimančių daugiau kaip 6000 rūšių. Lietuvoje – 122 rūšys. Yra daug kultūrinių bei introdukuotų rūšių. Vienmečiai, dvimečiai bei daugiamečiai žoliniai augalai (tik bambukinių stiebai sumedėję). Šaknų sistema kuokštinė. Stiebas stačias, bambliuotas, tarpubambliuose dažniausiai tuščiaviduris. Lapai išsidėstę dvieiliškai; juos sudaro lakštas, makštis ir liežuvėlis. Žiedai susitelkę į varputes, o jos – į sudėtinius žiedynus (varpa, kekė, šluotelė).
Migliniai yra vėjo apdulkinami (anemofiliniai). Paplitę visame Žemės rutulyje. Daugiau rūšių yra atogrąžų kraštuose, čia jie vyrauja savanų augalų bendrijose. Šių augalų reikšmė žmogui labai didelė. Čia priklauso daug žemės ūkio kultūrų – javų, kurie yra vieni pagrindinių maisto produktų visame pasaulyje: kviečiai, miežiai, ryžiai, kukurūzai, cukranendrės ir kt.
Migliniai, Varpiniai (lot. Poaceae, Gramineae, vok. Süßgräser) – magnolijūnų (Magnoliophyta) skyriui priklausanti labai didelė augalų grupė, kurią sudaro apie 500 genčių, apimančių daugiau kaip 6000 rūšių. Lietuvoje – 122 rūšys. Yra daug kultūrinių bei introdukuotų rūšių. Vienmečiai, dvimečiai bei daugiamečiai žoliniai augalai (tik bambukinių stiebai sumedėję). Šaknų sistema kuokštinė. Stiebas stačias, bambliuotas, tarpubambliuose dažniausiai tuščiaviduris. Lapai išsidėstę dvieiliškai; juos sudaro lakštas, makštis ir liežuvėlis. Žiedai susitelkę į varputes, o jos – į sudėtinius žiedynus (varpa, kekė, šluotelė).
Migliniai yra vėjo apdulkinami (anemofiliniai). Paplitę visame Žemės rutulyje. Daugiau rūšių yra atogrąžų kraštuose, čia jie vyrauja savanų augalų bendrijose. Šių augalų reikšmė žmogui labai didelė. Čia priklauso daug žemės ūkio kultūrų – javų, kurie yra vieni pagrindinių maisto produktų visame pasaulyje: kviečiai, miežiai, ryžiai, kukurūzai, cukranendrės ir kt.
Graudzāļu dzimta (Poaceae vai Gramineae) ir liela viendīgļlapju klases augu dzimta, sastopama gandrīz visā pasaulē. Dzimtā ietilpst apmēram 10 tūkstoši sugu, tādējādi tā ir piektā izplatītākā augu dzimta. Graudzāļu dzimtas augi tiek uzskatīti par svarīgākajiem augiem cilvēku ekonomikā — dzimtā ietilpst liela patēriņa pārtikas graudi un graudaugu kultūras, kas tiek audzētas visā pasaulē. Graudzāļu dzimta ir viena no lielākajām dzimtām Latvijas florā. Parasti graudzāļu dzimtas augi ir daudzgadīgi, retāk viengadīgi vai divgadīgi augi. Latvijā ir sastopamas aptuveni 175 sugas, visbiežāk tie ir pļavu un mežu augi.[1] Tiem ir trīs putekšņlapas un divas augļlapas. Starp populārākajiem dzimtas kultūraugiem ir auzas, rudzi, kvieši un mieži.[2]
Graudzāļu dzimta (Poaceae vai Gramineae) ir liela viendīgļlapju klases augu dzimta, sastopama gandrīz visā pasaulē. Dzimtā ietilpst apmēram 10 tūkstoši sugu, tādējādi tā ir piektā izplatītākā augu dzimta. Graudzāļu dzimtas augi tiek uzskatīti par svarīgākajiem augiem cilvēku ekonomikā — dzimtā ietilpst liela patēriņa pārtikas graudi un graudaugu kultūras, kas tiek audzētas visā pasaulē. Graudzāļu dzimta ir viena no lielākajām dzimtām Latvijas florā. Parasti graudzāļu dzimtas augi ir daudzgadīgi, retāk viengadīgi vai divgadīgi augi. Latvijā ir sastopamas aptuveni 175 sugas, visbiežāk tie ir pļavu un mežu augi. Tiem ir trīs putekšņlapas un divas augļlapas. Starp populārākajiem dzimtas kultūraugiem ir auzas, rudzi, kvieši un mieži.
De grassenfamilie (botanische naam: Gramineae of Poaceae, beide namen zijn toegestaan) is een van de soortenrijkste plantenfamilies van de bedektzadigen: de familie omvat ongeveer 8000 soorten. Leden van deze familie komen op alle werelddelen voor. Op Antarctica groeit een smele-soort. Ook granen, zoals tarwe en rijst, zijn vertegenwoordigers van de grassenfamilie.
Tot recente datum gaven fossiele vondsten aan dat gras circa 55 Ma (55 miljoen jaar) geleden evolueerde. Recente vondsten van de grasachtigen in coprolieten (fossiele fecaliën) van dinosauriërs uit het Krijt verwezen naar 65 Ma geleden.[1][2] Dateringen van de herkomst van rijst suggereren 107 tot 129 Ma.[3]
De relaties tussen de subfamilies Bambusoideae (bamboe-achtigen), Ehrhartoideae (waaronder rijst) en Pooideae (onder andere graan) worden als volgt verklaard: de Bambusoideae en de Pooideae zijn fylogenetisch meer met elkaar verwant dan met de Ehrhartoideae.[4] De scheiding voltrok zich in een relatief korte periode van 4 Ma.
Grassen zijn goed herkenbaar: ze hebben een vrij karakteristiek bouwplan. Sommige kenmerken bevinden zich aan de stengel en het blad, andere aan de bloem en aan de bloeiwijze. Ook hebben ze een kenmerkende geur.
De stengel is tussen de knopen (verdikkingen van de stengel) meestal hol. Onder andere mais en de Europese hanenpoot hebben echter een gevulde stengel.
De bladen staan in twee tegenoverstaande rijen ingeplant op de knopen. De bladen zijn gewoonlijk lijnvormig (zeer lang, smal) en parallelnervig.
De bladbasis, het eerste deel van het blad, vormt een kokervormige bladschede rond de stengel. Naar boven toe wordt de omsluiting van de stengel door de schede minder volledig, hier kan dan aan de randen van de schede een vlies zijn gevormd.
Het tweede deel is de bladschijf en staat af van de stengel. Op de overgang tussen bladschede en bladschijf is vaak een tongetje (ligula) te vinden, dit is een vliesje of een rij haartjes. De voet van de bladschijf omsluit de stengel vaak gedeeltelijk. Aan weerszijden van deze gedeeltelijke omsluiting zijn ook vaak de oortjes, die uit korte vliesjes bestaan, te vinden. In de lengte van de bladschijf lopen vaak ribben. Bij het topblad van de stengel heet de bladschijf het vlagblad.
Grassen zijn windbestuivers. De bloemblaadjes zijn niet felgekleurd: de bloeiwijze is vaak groenig, bruinig of doorschijnend en vormt veel stuifmeel. Vrij veel mensen vertonen een allergische reactie tegen stuifmeel: hooikoorts.
De aartjes vormen een centraal en kenmerkend onderdeel van de bloeiwijze van grassen. Een aartje kan één of meer bloemen hebben.
De onderste twee blaadjes van een aartje heten de kelkkafjes. Die kunnen een aartje helemaal omsluiten. Wanneer de kelkkafjes het aartje niet omsluiten, staan de achtereenvolgende blaadjes van het aartje dakpansgewijs.
Dan volgen twee schubjes, de kroonkafjes. Het eerstvolgende schubje (kroonkafje) aan de as van het aartje is het onderste kroonkafje: lemma. Het lemma bezit enige stevigheid, hoewel het heel dun is. Het volgende schubje is het bovenste kroonkafje: de palea, die vaak vliezig en slap is. Aan de top of op de rug van het lemma treedt bij veel grassen een meer of minder lange naald uit, de kafnaald; het is de verlenging van de middennerf.
Daarna komen de centrale bloemonderdelen: lodiculae, meeldraden en de stamper. De lodiculae zijn kleine schubjes die bij het begin van de bloei gaan zwellen en zo lemma en palea uit elkaar duwen. Hierdoor krijgen de meeldraden en de stamper ruimte om zich uiteen te gaan vouwen.
De aartjes kunnen op verschillende wijzen de bloeiwijze vormen. Die bloeiwijze bevindt zich altijd boven in de plant en vormt het meest kenmerkende onderdeel van de grashalm. Die bloeiwijze bestaat vrijwel altijd uit meer aartjes, maar het kan soms voorkomen dat die maar uit één enkele aartje bestaat.
De aartjes kunnen samen een aar vormen. Een aar wil zeggen dat de aartjes om het verlengde van de grasstengel, de aarspil, staan. Die aarspil is verder niet vertakt. De aartjes zitten dan op zogenaamde tanden aan die aarspil vast. Per tand kan er één aartje zijn, maar het kunnen er ook drie zijn. Vaak zitten de aartjes in twee rijen langs de aarspil, waarbij na een aartje van de ene rij, een aartje uit de andere rij volgt. Soms is de aar vlak, bij andere soorten juist vierkant.
Maar ze kunnen ook in een pluim gegroepeerd zijn. Dan ontspringt aan het verlengde van de grasstengel op verschillende punten zijtakjes. Aan deze zijtakjes kunnen de aartjes zitten, maar soms ontspringen er weer andere zijtakjes aan. Hieraan kunnen de aartjes zich bevinden, soms alleen, soms weer in groepen van drie. Soms is het eerste zijtak lang en zijn de andere die daaraan ontspringen kort, dan bestaat de pluim uit een aantal kluwens.
De typische grasgeur wordt veroorzaakt door cis-3-hexenol (bladalcohol).
Het blad, met al of niet een stukje stengel, wordt door grazende dieren gegeten. Een aanzienlijk deel van het voedsel van mensen wordt gevormd door grazende dieren. Vegetaties waar grassen in domineren worden graslanden genoemd.
Het begraasde gras is zo succesvol omdat het zijn investering voornamelijk onder de grond houdt. Het slaat zijn voedselvoorraad in zijn wortels op. De groei ontstaat in een verdikking net boven of net onder de grond. Dit maakt dat het helemaal niet erg is wanneer een grazend dier alle sprietvormige bladeren boven de grond opeet, want het kan binnen korte tijd door uitstoeling weer nieuwe bladeren vormen vanuit de energiereserves in de wortels. Ook een steppebrand deert het gras niet.
Het gras vormt een soort symbiose met de grazende dieren, door een wederzijdse afhankelijkheid: de dieren zijn afhankelijk van het gras, maar het gras ook van de dieren, want die dieren eten niet alleen al het gras op, maar ook de andere planten, die anders hoger zouden groeien dan het gras en het zonlicht zouden wegnemen. Daarbij bemesten ze het gras.
De mens kan de bladeren en de stengels van grassen niet verteren. Hij bezit namelijk geen verteringssappen die de wand van een plantaardige cel kunnen afbreken. Deze bestaat voornamelijk uit cellulose. Planteneters kunnen dit wel, dankzij de aanwezigheid van micro-organismen in hun maag-darmkanaal. Bij herkauwers, zoals de koe, het schaap, de wisent, de bizon en de gnoe vindt deze microbiële vertering (fermentatie) plaats in de voormagen (netmaag, pens en boekmaag). Bij andere planteneters zoals paard en olifant, vindt deze fermentatie plaats in de dikke darm.
De mens eet alleen de zaden van grassen. Die bevatten geen cellulose, maar hoofdzakelijk zetmeel. Soms kunnen de zaden tot wel 20% suiker bevatten, zoals bij suikermais, waarvan de onrijpe zaden als groente gegeten worden. Door selectie en veredeling zijn grassoorten ontstaan, die zo veel mogelijk energie in zaadproductie stoppen in plaats van in celwanden en nerven.
Voor de cultuurgewassen uit de familie van de grassen die door mensen worden gegeten, gebruikt men de verzamelnaam graan. Graan is wereldwijd de belangrijkste voedingsbron voor de mens. Granen zijn:
In de vorm van grasvelden wordt gras al eeuwen door de mens gebruikt. Grasvelden dienen uiteenlopende doelen: van decoratie van tuin (wordt dan gazon genoemd) of park tot het gebruik als sportveld, zoals een voetbalveld.
De grassenfamilie is onder te verdelen in onderfamilies en geslachtengroepen. Zie bijvoorbeeld de uitgebreide tabel op de pagina onderverdeling van de familie Poaceae, waar alle Nederlandse geslachten te vinden zijn.
De volgende (onder)soorten worden behandeld in een eigen artikel:
Alsook:
In het Cronquist-systeem (1981) was de plaatsing van de familie in diens orde Cyperales.
VoetnotenLiteratuur
Externe links
De grassenfamilie (botanische naam: Gramineae of Poaceae, beide namen zijn toegestaan) is een van de soortenrijkste plantenfamilies van de bedektzadigen: de familie omvat ongeveer 8000 soorten. Leden van deze familie komen op alle werelddelen voor. Op Antarctica groeit een smele-soort. Ook granen, zoals tarwe en rijst, zijn vertegenwoordigers van de grassenfamilie.
Grasfamilien er ei stor gruppe plantar einfrøblada dekkfrøplantar som veks over nesten heile verda. Ho omfattar over 10 000 ville og dyrka artar, og er den femte største plantefamilien[a][1] I Noreg finst kring 160 grarartar, fordelt på kring 60 slekter.
Grassletter utgjer rundt 20 % av vegetasjonsdekket på jorda. Grasplantar veks også i andre habitat, som skog, tundra og våtmark. Grasfamilien er økonomisk svært viktig, i og med at alle kornsortane høyrer til her. Korn vert skild frå dei andre artane som gruppa åkervekster. Dei viktigaste grasplantene er mellom anna havre, rug, bygg, kveite, ris, mais, durra, sukkerrøyr, bambus, timotei og raps.
Grasliknande vekstar som siv, sjøgras og halvgras høyrer ikkje til grasfamilien.
Grasplantene har vore nytta av menneske frå førhistorisk tid og har vorte nytta som mat til menneske i form av korn og som fôr til husdyr.
Frå gras til mat er det det mogne frøet (kornet) den viktigaste delen av planta. Kornet blei sådd om våren, i våronna, og hausta om hausten, i skuronna.
Frå gras til fôr er det blada og stenglane som utgjer den viktige delen av avlinga. Gras til fôr beitast direkte eller vert hausta fleire gongar i sesongen (slåttonn) og vert konservert anten ved tørking til høy eller vert ensilert som surfôr i tårnsilo, plansilo eller rundballar.
Grasfamilien er ei stor gruppe plantar einfrøblada dekkfrøplantar som veks over nesten heile verda. Ho omfattar over 10 000 ville og dyrka artar, og er den femte største plantefamilien I Noreg finst kring 160 grarartar, fordelt på kring 60 slekter.
Grassletter utgjer rundt 20 % av vegetasjonsdekket på jorda. Grasplantar veks også i andre habitat, som skog, tundra og våtmark. Grasfamilien er økonomisk svært viktig, i og med at alle kornsortane høyrer til her. Korn vert skild frå dei andre artane som gruppa åkervekster. Dei viktigaste grasplantene er mellom anna havre, rug, bygg, kveite, ris, mais, durra, sukkerrøyr, bambus, timotei og raps.
Grasliknande vekstar som siv, sjøgras og halvgras høyrer ikkje til grasfamilien.
Grasfamilien eller gressfamilien (Poaceae) omfatter cirka 700 planteslekter. I Norge finnes ca. 160 arter, fordelt på ca. 60 slekter. Familien er økonomisk svært viktig, i og med at alle kornsortene hører til her. Totalt artstall anslås til rundt 12 000.
De viktigste grasplantene i verden er blant annet havre, rug, bygg, hvete, ris, mais, durra, sukkerrør, bambus, timotei og rapp.
Grasplantene har vært benyttet fra førhistorisk tid og har blitt brukt som mat til mennesker i form av korn, og som fôr til husdyr. Fra gras til mat er det det modne frøet (kornet) som er den viktige delen av planta. Kornet ble sådd om våren, i våronna, og høstet om høsten, i skuronna.
Fra gras til fôr er det bladene og stenglene som utgjør den viktige delen av avlinga. Gras til fôr beites direkte eller høstes flere ganger i sesongen (slåttonn) og konserveres enten ved tørking til høy eller ensileres som silofôr i tårnsilo, plansilo eller rundballer.
Grasfamilien eller gressfamilien (Poaceae) omfatter cirka 700 planteslekter. I Norge finnes ca. 160 arter, fordelt på ca. 60 slekter. Familien er økonomisk svært viktig, i og med at alle kornsortene hører til her. Totalt artstall anslås til rundt 12 000.
De viktigste grasplantene i verden er blant annet havre, rug, bygg, hvete, ris, mais, durra, sukkerrør, bambus, timotei og rapp.
Wiechlinowate, trawy (Poaceae (R. Br.) Barnh., Gramineae Juss.[2]) – rodzina roślin należąca do rzędu wiechlinowców. Liczy ok. 11 tys. gatunków. Stanowi ona główny komponent roślinności stepowej, łąkowej i pastwiskowej. Należą do niej również ważne rośliny uprawne, w tym zboża. W Polsce występuje ponad 150 gatunków traw.
Są to rośliny zielne, (częściej byliny niż rośliny jednoroczne). Ich charakterystyczną cechą jest zdolność krzewienia się, czyli wypuszczania licznych źdźbeł z tzw. punktów krzewienia występujących w dolnych częściach pędu. Są przeważnie wiatropylne, czasami samopylne. U niektórych gatunków (np. u pszenicy zwyczajnej) występuje klejstogamia.
Klad okrytonasienne, klad jednoliścienne (monocots), rząd wiechlinowce (Poales), rodzina wiechlinowate (Poaceae). Rodzina stanowi grupę siostrzaną dla rodzin Ecdeiocoleaceae i Joinvilleaceae[1].
W różnych miejscach kuli ziemskiej znajdują się rozległe tereny porośnięte głównie trawami, a pozbawione drzew i krzewów. Noszą one nazwy wywodzące się najczęściej z miejscowego języka:
Dawna nazwa rodziny pojawia się w piosence „Opowieść o trawie” autorstwa Leszka Wójtowicza. Określana jest tam jako: ciekawa, dzielna, zwykła, bezczelna, uparta, mocna[5].
Wiechlinowate, trawy (Poaceae (R. Br.) Barnh., Gramineae Juss.) – rodzina roślin należąca do rzędu wiechlinowców. Liczy ok. 11 tys. gatunków. Stanowi ona główny komponent roślinności stepowej, łąkowej i pastwiskowej. Należą do niej również ważne rośliny uprawne, w tym zboża. W Polsce występuje ponad 150 gatunków traw.
Poaceae é uma família de plantas das angiospermas da classe Liliopsida (Monocotiledôneas), subclasse Commelinidae, também conhecidas como capins, gramas ou relvas. São plantas floríferas, frequentemente rizomáticas (ou não), anuais ou perenes, sublenhosas até lenhosas, com forma de vida em arbusto, árvore, bambu, erva, liana/volúvel/trepadeira ou subarbusto, vive sob substrato aquático, rupícula ou terrícula.[1][2]
O sistema de classificação APG II, de 2003, reconhece esta família incluindo-a na ordem Poales. A família é constituida por 668 gêneros com 10 035 espécies. O sistema de classificação APG, de 1998, reconhecia a família na ordem Poales com o nome de Gramineae (Juss.), com aproximadamente 700 gêneros e 12 000 espécies.
O Sistema de Cronquist, de 1981, também reconhecia esta família, porém colocando-a na ordem Cyperales (Wettst., 1911), que não existe mais.
No Sistema de Jussieu, de 1789, Gramineae é o nome de uma ordem botânica da classe Monocotyledones com estames hipogínicos, com 58 gêneros.
Estima-se que ambientes onde predominam espécies de Poacae correspondem a 20% da vegetação que cobre a terra, como os pastos e as savanas/cerrados. No Brasil, que possui cerrados e campos (naturais e antrópicos), ocorrem cerca de 180 gêneros e 1 500 espécies.
Nesta família encontram-se algumas plantas conhecidas como o trigo (Triticum aestivum), centeio, cevada (Hordeum vulgare), aveia (Avena sativa), arroz (Oryza sativa), sorgo (Sorghum bicolor), milheto (Pennisetum americanum), milho (Zea mays), cana-de-açúcar (Saccharum officinale), Bambu a Brachiaria entre outros.[3]
"Capim" é proveniente do termo tupi caá pi'i, que significa "folha delgada".[4] "Grama" vem do latim gramen, que significa "erva, relva".[5] "Cereal" vem do latim cereale, que significa "referente a Ceres" (a deusa romana da agricultura).[6] "Relva" vem do latim relevare, "levantar".[7]
Por serem monocotiledôneas tem a raiz fasciculada, abrangendo bastante área ao seu redor
As folhas das gramíneas são alternas dísticas, alongadas, paralelinérvias, invaginantes, com bainha fendida. Por serem sésseis não possuem pecíolo (o que liga a folha ao caule) exceto o bambu que possui um pseudo pecíolo. Há duas pequenas expansões na base da lâmina foliar denominada aurículas. Na junção da lâmina foliar com a bainha há a formação da lígula|lígula que pode ser membranosa, pilosa ou mista, característica muito importante para a taxonomia, a lígula fica entre a bainha e o limbo da folha na parte interna da mesma. A epiderme é rica em silício.
Possui caules de secção circular, em sua maioria o colmo, exceto o bambu que possui o caule lignificado, em seus entrenós são encontrados meristemas intercalados que os protegem do fogo e de herbivorias, já que nesses meristas são constituídos de gemas que fazem a planta crescer novamente.
Apresenta um perigônio formado por 3 verticilos de 2 peças que se reúnem em 1º grau para formar a inflorescência denominada “espigueta". Esta espigueta pode ser formada por 1 a 50 flores, onde algumas podem não ser funcionais.
Constituição da espigueta: ráquila, 2 glumas e flor em número variado conforme a espécie, que pode ser unissexual, hermafrodita ou neutra. A flor está formada de "lodículas", "androceu" e "gineceu". Apresentam brácteas: a “lema” e a “pálea”, que protegem o androceu e o gineceu, podem ser férteis ou estéreis. A lema é a mais externa, monoquilhada, pode apresentar "arista" (espigueta aristada), e quando não a tem denomina-se espigueta "mútica", passando este termo a integrar ou classificar a cultivar ou variedade; a pálea é biquilhada e cobre total ou parcialmente a pálea; o conjunto formado por lema e pálea denomina-se de "antécio" que pode estar ou não protegido pelas 2 brácteas da espigueta, denominadas “glumas I e II”; o androceu é formado por 3 estames na maioria das espécies, com filetes longos e tênues, o que facilita a anemofilia; excepcionalmente podem ocorrer 1,2,6 ou mais estames.[1]
Na sua grande maioria, os frutos das gramíneas são cariópses, que são frutos secos, indeiscentes (que não abrem) e possuem em sua maioria a semente presa ao pericarpo (endocarpo, mesocarpo e epicarpo) do fruto.
Plantas cosmopolitas, presentes tanto em ambientes desérticos como marinhos e de água doce. Registro em todos os tipos de ecossistemas, exceto em grandes altitudes. Segundo Judd, (2009), os campos nativos se desenvolvem em ecossistemas que apresentam estiagem sazonal, relevo ondulado e fogo periódico. Locais de presença e dominância das gramineas são as pradarias da América do Norte, os Pampas e os Cerrados Sul-americanos, o Veld africano (savana) e as estepes euro-asiáticas. Ocupam cerca de 24% da vegetação terrestre. Não é uma família extritamente endêmica do Brasil.[2][3]
Área antrópica, caatinga (stricto sensu), campinarana, campo de altitude, campo de várzea, campo limpo, campo rupestre, carrasco, cerrado (lato sensu), floresta ciliar ou galeria, floresta de igapó, floresta de terra firme, floresta de várzea, floresta estacional decidual, floresta estacional perenifólia, floresta estacional semidecidual, floresta ombrófila (= floresta pluvial), floresta ombrófila mista, manguezal, palmeiral, restinga, savana amazônica, vegetação aquática, vegetação sobre afloramentos rochosos.[2]
Dados da biogeografia da família no Brasil:[2]
Ocorrências confirmadas
Possíveis ocorrências
Amazônia, Caatinga, Cerrado, Mata Atlântica, Pampa, Pantanal
![Relações filogenéticas em Poaceae segundo Angiosperm Phylogeny Group (APG) Mapeamento de Vester, (1940) e Hultén, (1961) ([8]](http://pt.wikipedia.org/wiki/Ficheiro:Captura_de_tela_de_2017-01-29_14-38-19.png)
Anomochlooideae: Grupo irmão de todas as outras dentro do clado Poaceae. Inclui ervas perenes que crescem no chão sombreado de florestas nos Neotrópicos. Existem dois gêneros, Anomochloa e Streptochaeta, cada um em sua própria tribo.
Esta subfamília é a linhagem mais precoce divergente das gramíneas.
Pharoideae: Contém uma única tribo Phareae, com dois gêneros: Leptaspis e Pharus. Seus membros crescem nos pisos sombreados de florestas temperadas tropicais e quentes.
Esta subfamília é uma das linhagens de gramíneas mais antigas divergentes.
Puelioideae: Possui dois gêneros: Guaduella e Puelia, cada um em sua própria tribo. Seus membros crescem no sub-bosque de florestas tropicais.
Esta subfamília é uma das linhagens de gramíneas mais antigas divergentes.
Aristideae: é a única tribo de gramíneas na subfamília monotípica Aristidoideae. Seus membros são herbáceos anuais ou perenes encontrados em florestas subtropicais e zonas temperadas. A tribo tem mais de 300 espécies em três gêneros, onde a subfamília é um membro do clado de gramíneas, o grupo evolutivo em que a fotossíntese C4 evoluiu muitas vezes.
Panicoideae: Segunda maior subfamília das gramíneas com mais de 3.500 espécies, distribuídas principalmente em regiões temperadas e tropicais quentes. Compreende algumas culturas agrícolas importantes, incluindo a cana-de-açúcar, o milho, o sorgo e o capim. A fotossíntese C4 evoluiu independentemente um número de vezes na subfamília, onde tinha um antepassado C3.
Arundinoideae: Possui cerca de 40 espécies, incluindo o junco gigante e junco comum. Ao contrário de muitos outros membros do clado nas gramíneas, os Arundinoideae todos usam C3 fotossíntese. Seu grupo irmão é a subfamília Micrairoideae. Arundinoideae costumava ser bastante grande em sistemas taxonômicos mais antigos, com mais de 700 espécies, mas a maioria deles foram movidos para outras subfamílias após análises filogenéticas. Atualmente, as espécies são colocadas em 16 gêneros e duas tribos.
Micrairoideae: Distribuída em regiões tropicais e subtropicais. Dentro do clado é irmão da subfamília Arundinoideae. Inclui cerca de 190 espécies em nove gêneros. A classificação filogenética das gramíneas reconhece quatro linhagens principais, classificadas como tribos. Apenas as espécies da tribo Eriachneae (gêneros Eriachne e Pheidochloa) desenvolveram a via fotossintetizante C4.
Danthonioideae: Distribuida principalmente do hemisfério sul, contendo a única tribo Danthonieae e três gêneros, com 300 espécies. Inclui erva herbácea e parcialmente lenhosa ou anual (menos comum) gramas que crescem em pastagens abertas, arvoredos e florestas.
Danthonioideae exclusivamente utiliza a fotossíntese C3. Seu grupo irmão é a subfamília Chloridoideae, onde existem 20 gêneros, 17 dos quais são colocados na tribo Danthonieae, enquanto três ainda não foram classificados. As relações dentro do grupo são complicadas; Evidências filogenéticas conflitantes do DNA nuclear e do cloroplasto sugerem que os eventos de hibridização desempenharam um papel importante no Danthonioideae.
Chloridoideae é uma das maiores subfamílias de gramíneas, com aproximadamente 130 gêneros e 1 600 espécies, encontradas principalmente em pastagens áridas tropicais ou subtropicais. Dentro do clado, seu grupo irmão são os Danthonioideae. A subfamília inclui ervas daninhas e as espécies do milho crescidas em algumas regiões tropicais.
Com exceção de algumas espécies em Elliscochloa, a maioria das espécies da subfamília utiliza a via fotossintetizante C4. A primeira transição evolutiva da fotossíntese C3 para C4 nas gramíneas provavelmente ocorreu nesta subfamília, cerca de 32 a 25 milhões de anos.
Oryzoideae: Inclui cerca de 120 espécies em 20 gêneros, principalmente o arroz. Dentro das gramíneas, esta subfamília é uma das três pertencentes doclado rico em espécies que utilizam a fotossíntese C3; É a linhagem basal da subfamília. Contém quatro tribos e um gênero de posição incerta, Suddia (provavelmente Phyllorachideae). As análises filogenéticas resolveram a ordem de ramificação destes clados dentro da subfamília.
Bambus são perenes florescendo plantas na subfamília Bambusoideae. No bambu, tal como noutras gramíneas, as regiões internodais do caule são geralmente ocos e os feixes vasculares na secção transversal estão espalhados por todo o caule em vez de numa disposição cilíndrica. O xilema lenhoso dicotiledóneo também está ausente. A ausência de madeira de crescimento secundário faz com que as hastes de monocotiledóneas, incluindo as palmas e os grandes bambus, sejam colunares em vez de afiladas.
Pooideae é a maior subfamília de Poaceae com 4.200 espécies em 14 tribos e aproximadamente e 200 géneros. Incluem alguns cereais principais como o trigo, a cevada, a aveia, o centeio e muitas gramas. Eles são muitas vezes referidos como gramíneas de estação fria porque elas são distribuídas em clima temperado. Todos usam a via fotossintética C3.
Índice com os géneros da família Poaceae segundo o sistema: APG II
Índice com os gêneros da família Poaceae segudo o sistema: Flora do Brasil
Há 458 espécies de gramíneas catalogadas no Estado de São Paulo, sendo 403 plantas nativas e 55 originárias do exterior.[9]
De fundamental importância para alimentação humana, esta família botânica é a mais importante de todas as famílias de plantas para economia, cerca de 70% das terras cultivadas e responsável por mais de 50% das calorias consumidas pela humanidade, sendo cultivada pelo menos há 10 mil anos. Graças a domesticação de plantas como trigo, cevada, arroz e aveia foi possível o surgimento das primeiras formas de civilização. Os grãos são o principal alimento cultivado em torno do mundo e o bambu, usado extensamente para a construção e os brotos jovens na alimentação. As gramas agriculturais cultivadas para a produção de alimento são chamadas cereais. Os cereais constituem a fonte principal de calorias para seres humanos. Incluem o arroz na Ásia, o milho no México e trigo e cevada na Europa e na América do Norte. O milho e outros cereais são também cultivados em muitos países para a produção de alimentos para animais, na pecuária, as fibras para fabricar papel, a polpa para a produção de raiom, os açucares para fermentação de bebidas alcoólicas, tais como cerveja e uísque e os caules resistentes utilizados como material de construção.[3]
Elas têm uma grande importância também na agronomia. O trigo é um exemplo de gramínea, que se adaptou largamente entre várias regiões do planeta para a produção de cervejas. O arroz é outro exemplo de gramínea que se adaptou largamente ao paladar, não só do brasileiro assim como outros países que tem ele como um dos seus principais alimentos.
As gramíneas estão largamente relacionadas com a agronomia e agroecologia. O ramo da botânica que as estuda é a agrostologia.
Poaceae é uma família de plantas das angiospermas da classe Liliopsida (Monocotiledôneas), subclasse Commelinidae, também conhecidas como capins, gramas ou relvas. São plantas floríferas, frequentemente rizomáticas (ou não), anuais ou perenes, sublenhosas até lenhosas, com forma de vida em arbusto, árvore, bambu, erva, liana/volúvel/trepadeira ou subarbusto, vive sob substrato aquático, rupícula ou terrícula.
O sistema de classificação APG II, de 2003, reconhece esta família incluindo-a na ordem Poales. A família é constituida por 668 gêneros com 10 035 espécies. O sistema de classificação APG, de 1998, reconhecia a família na ordem Poales com o nome de Gramineae (Juss.), com aproximadamente 700 gêneros e 12 000 espécies.
O Sistema de Cronquist, de 1981, também reconhecia esta família, porém colocando-a na ordem Cyperales (Wettst., 1911), que não existe mais.
No Sistema de Jussieu, de 1789, Gramineae é o nome de uma ordem botânica da classe Monocotyledones com estames hipogínicos, com 58 gêneros.
Estima-se que ambientes onde predominam espécies de Poacae correspondem a 20% da vegetação que cobre a terra, como os pastos e as savanas/cerrados. No Brasil, que possui cerrados e campos (naturais e antrópicos), ocorrem cerca de 180 gêneros e 1 500 espécies.
Nesta família encontram-se algumas plantas conhecidas como o trigo (Triticum aestivum), centeio, cevada (Hordeum vulgare), aveia (Avena sativa), arroz (Oryza sativa), sorgo (Sorghum bicolor), milheto (Pennisetum americanum), milho (Zea mays), cana-de-açúcar (Saccharum officinale), Bambu a Brachiaria entre outros.
Poaceae sau gramineae (graminee) este o familie de plante erbacee (rar lemnoase) monocotiledonate, cu rădăcina fasciculată, cu tulpina formată din noduri și internoduri, cu inflorescența în formă de spic; (și la sg.) plantă care face parte din această familie. Familia conține între 9.000-10.000 de specii de plante.
Poaceae sau gramineae (graminee) este o familie de plante erbacee (rar lemnoase) monocotiledonate, cu rădăcina fasciculată, cu tulpina formată din noduri și internoduri, cu inflorescența în formă de spic; (și la sg.) plantă care face parte din această familie. Familia conține între 9.000-10.000 de specii de plante.
Lipnicovité (Poaceae; starší názov trávy – Graminae sa stále používa v USA) je čeľaď rastlín z radu lipnicotvarých.
Je to morfologicky aj biologicky vyhranená čeľaď s vyše 400 rodmi a takmer 10 000 druhmi (iné zdroje 600 rodmi a vyše 6 000 druhmi). Sú to jednoročné až trvalé rastliny. Sú rozšírené po celom svete a majú pre človeka rozhodujúci význam ako základ pestovania obilnín a krmovín. S výnimkou bambusu ide o byliny s valcovitými stonkami (steblami), ktoré sú článkované, v kolienkach delené priečnymi prepážkami, duté (výnimky: cukrová trstina, kukurica); listy objímajú steblo a sú dlhé a úzke, väčšinou sú striedavé so súbežnou žilnatinou a bývajú rozlíšené na čepeľ a pošvu. Zatváravé bunky prieduchov majú netypický činkovitý tvar.
Nevýrazné kvety sú usporiadané v klasoch, metlinách alebo stiahnutých metlinách, majú tri tyčinky, jeden piestik s dvomi perovitými bliznami a zvonka sú kryté plievočkou, plevicou a vonkajšími (čiže spodnými) plevami. Po opelení vetrom stena jediného semenníka (osemenie) zrastá s oplodím a vytvára plod tráv, obilku (caryopsis). Z jednej obilky môže vyrásť odnožovaním niekoľko stebiel.
Väčšina druhov obilnín bola vypestovaná z planých tráv juhozápadnej Ázie. Oproti rozpadavému vretenu klasov planých druhov zostáva pri kultúrnych obilninách klas v celku a obilky sa uvoľňujú z pliev, plevíc a plievočiek až mlátením.
Lipnicovité súvisia morfologicky s Commelinales.
Podľa APG (2006), Grass Phylogeny Working Group (2005) a [1] sa delí takto:
Staršie delenie podľa [2]:
Tradičné delenie na Slovensku:
Lipnicovité (Poaceae; starší názov trávy – Graminae sa stále používa v USA) je čeľaď rastlín z radu lipnicotvarých.
Je to morfologicky aj biologicky vyhranená čeľaď s vyše 400 rodmi a takmer 10 000 druhmi (iné zdroje 600 rodmi a vyše 6 000 druhmi). Sú to jednoročné až trvalé rastliny. Sú rozšírené po celom svete a majú pre človeka rozhodujúci význam ako základ pestovania obilnín a krmovín. S výnimkou bambusu ide o byliny s valcovitými stonkami (steblami), ktoré sú článkované, v kolienkach delené priečnymi prepážkami, duté (výnimky: cukrová trstina, kukurica); listy objímajú steblo a sú dlhé a úzke, väčšinou sú striedavé so súbežnou žilnatinou a bývajú rozlíšené na čepeľ a pošvu. Zatváravé bunky prieduchov majú netypický činkovitý tvar.
Nevýrazné kvety sú usporiadané v klasoch, metlinách alebo stiahnutých metlinách, majú tri tyčinky, jeden piestik s dvomi perovitými bliznami a zvonka sú kryté plievočkou, plevicou a vonkajšími (čiže spodnými) plevami. Po opelení vetrom stena jediného semenníka (osemenie) zrastá s oplodím a vytvára plod tráv, obilku (caryopsis). Z jednej obilky môže vyrásť odnožovaním niekoľko stebiel.
Väčšina druhov obilnín bola vypestovaná z planých tráv juhozápadnej Ázie. Oproti rozpadavému vretenu klasov planých druhov zostáva pri kultúrnych obilninách klas v celku a obilky sa uvoľňujú z pliev, plevíc a plievočiek až mlátením.
Lipnicovité súvisia morfologicky s Commelinales.
Arundinoideae
Bambusoideae
Centothecoideae
Chloridoideae
Panicoideae
Pooideae
Stipoideae
Gramineae
Trave (znanstveno ime Poaceae) so večinoma zelnate rastline, redkeje olesenele. Obsegajo okoli 9000 vrst, združenih v okoli 650 rodov. So vir vseh vrst žit (vključno z rižem), večine svetovnega pridelka sladkorja in hrane za domače in divje živali ter bambusa in trstike. Družina je razširjena po vsem svetu, od polarnih krogov do ekvatorja, od gora do morja.
Trave so kozmopoliti. Pokrivajo približno 20% kopnega. Dominirajo v pasu med gozdovi in puščavami, to je v pasu step, prerij, savan in pamp. Pod vplivom človeka so nastale obsežne kulturne stepe na nekoč gozdnatih območjih.
Med travami prevladujejo zelišča, zelnate trajnice in enoletnice. Pravih lesnatih rastlin ni, čeprav pri nekaterih stebla olesenijo (bambus).
Pri tipičnih travah je koreninski sistem šopast in pogosto dopolnjen z nadomestnimi koreninami, ki poganjajo iz spodnjih stebelnih kolenc.
Steblo trav (bil ali bilka) je pokončno, izrazito kolenčasto, večinoma nerazraslo in največkrat votlo. Včasih je steblo tudi polno ali celo lesnato, vendar ne zaradi sekundarne debelitve, ampak zaradi lignifikacije primarnih tkiv (bambus).
Trave imajo pogosto podzemna stebla, iz katerih poganjajo nadzemni deli. Stebla se razraščajo v glavnem tik pod ali nad površjem.
Listi rastejo v dveh vrstah v določenih presledkih vzdolž stebla. Mesto kjer so pritrjeni na steblo se imenuje kolence (nodij). Sestavljeni so iz nožnice in ploskve. Nožnica, ki je morfološka značilnost trav, tesno obdaja steblo in daje mehansko oporo mehkemu delu stebla, ki je tik nad kolencem. Kjer prehaja listna nožnica v listno ploskev, je pri mnogih travah nežna prosojna listna kožica ali ligula. Kolence ni votlo, tu je tkivo, ki omogoča travam rast v dolžino. Odebelina na kolencih nastane zato, ker na tem mestu močno odebeli listna nožnica; s tem je nežno tkivo v notranjosti kolenca dobro zavarovano.
Socvetja trav so sestavljena socvetja. Glavna os socvetja ne nosi posameznih cvetov, temveč manjša socvetja, ki jim pravimo klaski. Urejeni so na različne načine, od posamičnega klasa ali grozda do vmesnih stopenj z nekaj klaski in večvejnatega lata.
V klasu pšenice, rži, ječmena in mnogih drugih trav so klaski sedeči, to je nepecljati; v latu pa so klaski pecljati, tako na primer pri ovsu.
Trave so vetrocvetke in imajo dvospolne cvetove - ena izmed redkih izjem je koruza. Opraševanje traja zelo kratek čas.
Velik pomen trav, ki ga imajo v vegetaciji, je predvsem v njihovi sposobnosti, da rastejo v silnih množinah in da tvorijo masovne sestoje, kot so pašniki, senožeti, travniki, stepe, pampe, prerije in savane.
Trave so najožje povezane z evolucijo človeka, saj so se vse človeške civilizacije razvile vsaj z eno od vrst trav kot glavnim virom prehrane:
Trave dajejo poleg škroba tudi sladkor (Saccharum officinarium), krmo živalim, lignificirana stebla bambusov se uporabljajo za pohištvo. Pomembne so tudi za krajinsko oblikovanje (trate, zelenice, igrišča).
Tipski rod trav je latovka (Poa), odtod tudi poimenovanje družine Poaceae. Ime Poa izvira iz grščine in pomeni zelišče, trava, rastlina. Zastarel izraz za družino je Gramineae. Uporaba starejšega izraza je še vedno dovoljena: Poaceae = Gramineae Jussieu nom. cons. (nomina conservanda) et nom. alt. (nomen alternativum)[1].
Trave so kritosemenke (Magnoliophyta). V nasprotju z golosemenkami (Gymnospermae) je pri njih semenska zasnova v plodnici. Družina trav pripada redu travovci (Poales) in razredu enokaličnice (Liliopsida). Družina vsebuje približno 10.000 vrst, ki so razdeljene v 600 do 700 rodov. Trave so razdeljene v 13 poddružin, ki so po velikosti neenakomerno porazdeljene, potem so razčlenjene še v skupno 64 tribusov. Poddružine lahko glede na filogenetiko združimo v dve glavni skupini: „BEP-klad“ in „PACC-klad“.
BEB-klad
PACC-klad
Trave (znanstveno ime Poaceae) so večinoma zelnate rastline, redkeje olesenele. Obsegajo okoli 9000 vrst, združenih v okoli 650 rodov. So vir vseh vrst žit (vključno z rižem), večine svetovnega pridelka sladkorja in hrane za domače in divje živali ter bambusa in trstike. Družina je razširjena po vsem svetu, od polarnih krogov do ekvatorja, od gora do morja.
Gräs (Poaceae, tidigare Gramineae) är en familj enhjärtbladiga växter med omkring 9 000 arter i hela världen.
Gräs är ett- eller fleråriga örter med kraftigt varierande storlek, och rötterna är samlade i en jämn knippa. Stjälken kallas strå och är ofta ihålig, men arter som majs och sockerrör har märg, medan bambu har en förvedad stam. Strået är ledat och lederna skiljs åt av förtjockade noder. Bladen utgår från noderna och omsluter strået med en bladslida. Mellan denna och bladskivan sitter en hinna, snärpet. Blommorna skyddas av skärmfjäll och blomfjäll som kallas agnar, och är samlade i småax som i sin tur bildar ax eller vippor.
Gräs kan växa enstaka eller i täta tuvor och förökar sig ofta via underjordiska revor. De är vanligen korsbefruktande och pollineras av vinden. Det stärkelserika, mjöliga fröet är inneslutet i ett slags nöt.
Arter i familjen halvgräs betraktas ofta felaktigt som gräs. Halvgräs kan särskiljas genom att de saknar en tydlig led på strået och ofta genom att halvgräsens strån inte har rund (utan till exempel trekantig) genomskärningsyta.
Grupp 1: Panicoideae
Grupp 2: Andropogonodae
Gräs (Poaceae, tidigare Gramineae) är en familj enhjärtbladiga växter med omkring 9 000 arter i hela världen.
Gräs är ett- eller fleråriga örter med kraftigt varierande storlek, och rötterna är samlade i en jämn knippa. Stjälken kallas strå och är ofta ihålig, men arter som majs och sockerrör har märg, medan bambu har en förvedad stam. Strået är ledat och lederna skiljs åt av förtjockade noder. Bladen utgår från noderna och omsluter strået med en bladslida. Mellan denna och bladskivan sitter en hinna, snärpet. Blommorna skyddas av skärmfjäll och blomfjäll som kallas agnar, och är samlade i småax som i sin tur bildar ax eller vippor.
Gräs kan växa enstaka eller i täta tuvor och förökar sig ofta via underjordiska revor. De är vanligen korsbefruktande och pollineras av vinden. Det stärkelserika, mjöliga fröet är inneslutet i ett slags nöt.
Arter i familjen halvgräs betraktas ofta felaktigt som gräs. Halvgräs kan särskiljas genom att de saknar en tydlig led på strået och ofta genom att halvgräsens strån inte har rund (utan till exempel trekantig) genomskärningsyta.
Buğdaygiller (Poaceae), tohum kabuğu, meyve kabuğu ile bitişik, sapları boğumlu ve ekseriya içi boş, başakçıklar, başak vaziyetinde toplanmış halde, 4000 kadar türe sahip, Poales takımına bağlı bitki familyası.
Buğday, pirinç, çavdar, mısır, darı, yulaf, arpa, bambu gibi bitkiler buğdaygillerdendir.
Anadolu'da, bütün buğdaygillere ait çeşitler yetişmekte ve yetiştirilmektedir. Buğdaygillerin, bütün dünyada ve Türkiye'de yetişme alanı geniştir. İnsan ve hayvan beslenmesinde büyük öneme sahiptir. Memleketimizde, buğday ve pirinç yemeklik olarak, çavdar, mısır ve darı hem yemeklik, hem de hayvan yemi, arpa ve yulaf ise hayvan yemi olarak sarf edilmektedir.
Buğday Türkiye'nin hemen hemen her tarafında, daha çok İç Anadolu bölgesinde yetiştirilir. Pirinç en fazla Trakya, Kastamonu, Kahramanmaraş ve Hatay illerimizde; mısır Akdeniz bölgesinde; darı Hatay'da yetiştirilmektedir.
Türkiye'nin Yıllık Buğdaygiller Üretimi (1991) Tahıl Miktar Buğday 20.400.000 ton Arpa 7.800.000 ton Mısır 2.100.000 ton Çavdar 280.000 ton Yulaf 276.000 ton Pirinç 158.000 ton Darı 9.000 tonКолись більш уживана латинська назва родини Graminea частіше перекладалась як зл́аки Лунтика
Коренева система, як і у всіх однодольних, — мичкуватого типу.
Мають стебло — соломину. Пагін часто є кореневищем. Злаковим властивий інтеркалярний ріст (пагін складається з вузлів та міжвузль). Тонконогові мають порожнисте стебло — соломину, на якому чітко видно вузли та міжвузля. Листки лінійні, з паралельним жилкуванням, сидячі; у місці переходу листкової пластинки в піхву є язичок (плівчастий виріз).
Квіти дрібні, анемогамні, в основному двостатеві. Суцвіття тонконогових, як правило, — складний колос (у культурної кукурудзи жіноча квітка — складний початок, а чоловіча квітка — волоть).
Квітка злакових складається з двох лусок, має трипроменеву симетрію. Маточка одна з двома приймочками. Тичинок 3 або 6.
Плід — зернівка, горішок або ягода (у бамбуку) серед злаків відомі як самозапильні.
Розвиток родини злакових пов'язаний із пристосуванням до запилення вітром. Злакові є основою рослинного покриву Землі.
До останнього часу вважалось, що трави як форма рослинності з'явились близько 55 мільйонів років тому; але, згідно з останніми палеонтологічними знахідками скам'янілих залишків трав (попередників рису та бамбуку), вони існували вже в Крейдовому періоді, приблизно 65 мільйонів років тому. Вважають, що еволюційно трави з'явились в результаті утворення неотенічних форм багаторічних рослин, що надало їм ряд екологічних переваг, насамперед пов'язаних із вкороченим життєвим циклом і, як результат, більшою плодючістю на довгостроковому проміжку часу.
Порівняно з іншими покритонасінними, еволюція трав проходила досить повільно. Їхнє широке розповсюдження та швидке збільшення кількості видів почалося лише в Міоцені (20-10 мільйонів років тому), коли трав'яні екосистеми стали домінувати на великих територіях. Це звичайно пов'язують із кліматичними змінами того періоду (періодичні зледеніння), завдяки яким різко збільшилась площа суходолу із сухим та холодним кліматом; а саме до такого типу клімату трави екологічно більше пристосовані, ніж інші форми рослин. Окрім того, трави швидше відновлюються після пошкодження при випасі тварин та пожежах; почастішання цих явищ по мірі розвитку людства також мало сприяти заміні трав'янистими екосистемами всіх інших.
Злакові мають технічне та декоративне значення.
Тонконогові використовуються як агрокультура. Це, в основному, — пшениця, жито, ячмінь і овес, але до них також належать рис, просо і кукурудза. Тонконогові містять близько 75% складних вуглеводів і 10% білків, а також жири і клітковину. Добре зберігаються.
Значення злакових у житті людини надзвичайно велике та різноманітне. Перше місце посідають хлібні та круп'яні культури, серед яких пшениця, рис посівний і кукурудза, вони є основними харчовими рослинами людства. Не менш важливим є використання кормових злакових культур. Найкращими з них є тимофіївка лучна, лисохвіст лучний, тонконіг лучний, костриця лучна тощо. Є серед злакових і лікарські рослини (кукурудза, пирій), рослини що використовуються у косметиці (з рисового борошна виробляють найвишуканіші види пудри). Чимало серед злакових і бур'янів (пирій, плоскуха, мишій).
Див. Список родів родини Тонконогових.
Họ Hòa thảo hay họ Lúa hoặc họ Cỏ ("cỏ" thực thụ) là một họ thực vật một lá mầm (lớp Liliopsida), với danh pháp khoa học là Poaceae, còn được biết dưới danh pháp khác là Gramineae. Trong họ này có khoảng 668 chi và khoảng 10.035 loài cỏ. Người ta ước tính rằng các đồng cỏ chiếm khoảng 20% toàn bộ thảm thực vật trên Trái Đất. Họ này là họ thực vật quan trọng nhất đối với toàn bộ nền kinh tế của loài người, bao gồm cả các bãi cỏ và cỏ cho gia súc cũng như là nguồn lương thực chủ yếu (ngũ cốc) cho toàn thế giới, hay các loại tre, trúc được sử dụng rộng rãi ở châu Á trong xây dựng.
Các loài thuộc họ Hòa thảo có các đặc điểm sau:
Cho đến thời gian gần đây, các loài cỏ được cho là đã tiến hóa vào khoảng 55 triệu năm trước, khi người ta căn cứ vào các mẫu hóa thạch đã có. Tuy nhiên, các nghiên cứu gần đây về các loại thực vật hóa đá 65 triệu năm tuổi, bao gồm các tổ tiên của lúa và tre trong phân hóa đá của khủng long thời kỷ Phấn trắng ([1], [2]), đã đặt sự đa dạng của các loài cỏ về thời kỳ sớm hơn.
Cây phát sinh chủng loài của họ Hòa thảo so với các họ khác trong bộ Hòa thảo liệt kê dưới đây là lấy theo APG III.
Poales
Typhaceae s. l.
Typhaceae s. s.
Sparganiaceae (Sparganium)
Poaceae
Cây phát sinh chủng loài trong nội bộ họ Hòa thảo như liệt kê dưới đây là lấy theo APG III.
Poaceae
Nhánh BEP
Các loài cỏ được trồng để cung cấp lương thực cho loài người được gọi chung là ngũ cốc. Các loại ngũ cốc cung cấp phần lớn năng lượng để nuôi sống con người và có lẽ cũng là nguồn cung cấp chính về protein. Các loại ngũ cốc bao gồm lúa ở Nam và Đông Nam Á, ngô ở Trung và Nam Mỹ, lúa mì và lúa mạch ở châu Mỹ và khu vực miền bắc đại lục Á-Âu. Nhiều loại cỏ khác cũng được trồng để làm cỏ tươi và cỏ khô cho gia súc, cụ thể là cho trâu, bò và cừu.
Một số loại "cỏ" được biết đến rộng rãi là:
Họ Hòa thảo hay họ Lúa hoặc họ Cỏ ("cỏ" thực thụ) là một họ thực vật một lá mầm (lớp Liliopsida), với danh pháp khoa học là Poaceae, còn được biết dưới danh pháp khác là Gramineae. Trong họ này có khoảng 668 chi và khoảng 10.035 loài cỏ. Người ta ước tính rằng các đồng cỏ chiếm khoảng 20% toàn bộ thảm thực vật trên Trái Đất. Họ này là họ thực vật quan trọng nhất đối với toàn bộ nền kinh tế của loài người, bao gồm cả các bãi cỏ và cỏ cho gia súc cũng như là nguồn lương thực chủ yếu (ngũ cốc) cho toàn thế giới, hay các loại tre, trúc được sử dụng rộng rãi ở châu Á trong xây dựng.
Các loài thuộc họ Hòa thảo có các đặc điểm sau:
Cây thân cỏ, sống lâu năm, ít khi 1 hay 2 năm, một số có dạng thân gỗ thứ sinh (tre, nứa v.v). thân khí sinh chia gióng (cọng) và mấu (đốt): gióng thường rỗng (trừ một số loài như nứa, kê, ngô có thân đặc), không phân nhánh (trừ tre) mà chỉ phân nhánh từ thân rễ hoặc từ gốc. Lá mọc cách (so le), xếp hai dãy theo thân (trên cùng một mặt phẳng), ít khi có dạng xoắn ốc, gân lá song song. Bẹ lá to, dài, hai mép của bẹ không dính liền nhau. Lá không có cuống (trừ tre), phiến lá hình dải hẹp. Giữa bẹ và phiến lá có lưỡi bẹ nhỏ hình bản mỏng hay hình dãy lông mi. nguồn gốc của lưỡi không rõ ràng, một số tác giả cho là do hai lá kèm dính nhau biến đổi thành. Vai trò sinh học của nó là cản bớt nước chảy vào thân non ở đốt. Gốc bẹ lá hơi phồng lên, mép ôm chặt lấy thân và che chở cho mô phân sinh đốt, nhờ đó mà mô này duy trì hoạt động được khá lâu. Hoa nhỏ, thụ phấn nhờ gió, tập trung thành cụm bông, cơ sở là các hoa nhỏ. Các hoa nhỏ này lại hợp thành những cụm bông phức tạp hơn như bông kép, chùm, chùy v.v nhưng không có các cánh hoa. Mỗi bông mang từ 1 - 10 hoa nhỏ. Ở gốc bông nhỏ thường có 2 mày (lá bắc) bông xếp đối nhau: còn ở gốc mỗi hoa có 2 mày hoa, mày hoa dưới ôm lấy mày hoa trên, nhỏ và mềm hơn, mày hoa dưới chỉ có 1 gân ở chính giữa, còn mày hoa trên có 2 gân bên. Ở nhiều loài, mày hoa dưới kéo dài ra thành chỉ ngón. Phía trong 2 mày hoa còn 2 mày cực nhỏ rất bé và mềm. Như vậy, thông thường mỗi hoa có 4 mày, nhưng trong thực tế số lượng này có khi không đầy đủ. Nhị thường là 3 (đôi khi 6), chỉ nhị dài bao phấn đính lưng, hai bao phấn khi chín thường xòe ra thành hình chữ X. Bầu trên có một ô, 1 noãn, 2 vòi nhụy ngắn và 2 đầu nhụy dài mang chùm lông quét, thường màu nâu hoặc tím. Quả là loại quả thóc (caryopsis).Cho đến thời gian gần đây, các loài cỏ được cho là đã tiến hóa vào khoảng 55 triệu năm trước, khi người ta căn cứ vào các mẫu hóa thạch đã có. Tuy nhiên, các nghiên cứu gần đây về các loại thực vật hóa đá 65 triệu năm tuổi, bao gồm các tổ tiên của lúa và tre trong phân hóa đá của khủng long thời kỷ Phấn trắng ([1], [2]), đã đặt sự đa dạng của các loài cỏ về thời kỳ sớm hơn.
Poaceae Barnhart (1895), nom. cons.
СинонимыЗла́ки, или Злаковые[2] (лат. Gramíneae), или Мя́тликовые[2] (лат. Poáceae) — семейство однодольных растений, к которому относятся такие известные и давно используемые в хозяйстве растения, как пшеница, рожь, овёс, рис, кукуруза, ячмень, просо, бамбук, сахарный тростник.
Злаки распространены на всех континентах. Даже в Антарктиде встречаются 2 вида злаков — щучка антарктическая (луговик антарктический) и обнаруженный недавно заносный вид — мятлик однолетний.
Злаки составляют значительную часть фитомассы во многих биоценозах, а в степях и саваннах — подавляющую часть.
Жизненные формы злаков разнообразны — деревья, кустарники, многолетние и однолетние травы. Среди злаковых нет эпифитов, сапрофитов и паразитов. Группа многолетних злаков наиболее разнообразна. Среди них есть растения дерновинные, длиннокорневищные, столонообразующие. В каждой из указанных групп можно выделить несколько подгрупп, различающихся деталями строения надземных и подземных побегов растения. Например, дерновинные злаки в зависимости от типа возобновления побегов — вневлагалищного или внутривлагалищного — подразделяются на рыхлодерновинные и плотнодерновинные, соответственно.
Побеги злаков подразделяются на вегетативные и генеративные. Каждый из них может быть как однолетним, так и многолетним. Механизма вторичного утолщения стебля нет. Ветвление происходит в зоне кущения или в области соцветия. Стебли цилиндрические, членистые, в междоузлиях полые — соломины. Исключением являются несколько видов подсемейства бамбуковых, кукуруза и сахарный тростник[3].
Листья очерёдные, двурядные, узкие, с незамкнутым влагалищем. На границе влагалища и пластинки находится плёнчатый язычок или ряд волосков.
Общие соцветия злаков могут быть представлены колосьями, метёлками, колосовидными метелками (султан), кистями, початками. Общие соцветия состоят из многих элементарных соцветий — колосков. В колосках может находиться от одного до нескольких цветков. Цветки мелкие, ветро- или самоопыляемые. Цветок в типичном случае имеет нижнюю и верхнюю цветковые чешуи. Верхняя цветковая чешуя имеет 2 киля и, вероятно, возникла от срастания 2 чешуй. Внутри цветковых чешуй находятся 3 очень маленькие цветковые чешуйки — лодикулы, которые считаются элементами внутреннего круга околоцветника. Такая структура околоцветника даёт основание считать его трёхчленным (трёхмерным). В цветке обычно 3 тычинки с качающимися пыльниками. Завязь цветка имеет 2 перистых рыльца. У некоторых злаков строение цветка отличается от этой типичной схемы. Например, виды бамбуков имеют 6 или более тычинок, а виды рода цинна — одну тычинку.
Формула цветка: ↑ P ( 2 ) + 2 A 3 G ( 2 _ ) o r ( 3 _ ) {displaystyle uparrow P_{{(2)}+2};A_{3};G_{({underline {2}})or({underline {3}})}} 
Плод — псевдомонокарпий: зерновка, у которой плёнчатый околоплодник плотно прилегает к семени и иногда слипается со спермодермой.
Большую часть единственного семени составляет эндосперм. Зародыш сравнительно небольшой, боковой.
Корни злаков мочковатые[5].
Плоды злаков — зерновки — содержат большое количество крахмала и протеина. Некоторые злаки содержат кумарины, значительно реже эфирные масла[6].
Злаки — важные кормовые и древнейшие пищевые культурные растения. Из них получают хлебопекарную муку, крупы, сахар (сахарный тростник), строительные материалы и волокна (бамбук, тростник). Дикорастущие злаки составляют основной корм домашнего скота[6].
Семейство злаков включает 759 родов и 11 554 вида[7].
Современная классификация APG II выделяет следующие подсемейства:
В классических системах цветковых растений, распространённых в XX веке, деление семейства злаков на подсемейства было другим. Так, А. Л. Тахтаджян в 1982 году выделял 6 подсемейств: Бамбуковые (Bambusoideae), Рисовые[en] (Oryzoideae), Мятликовые (Pooideae), Просовые (Panicoideae), Тростниковые[en] (Arundinoideae) и Полевичковые (Eragrostideae)[9]; в 1987 году он же делил злаки всего на 2 подсемейства: Бамбуковые (Bambusoideae) и Мятликовые (Pooideae)[10].
По данным кладистического анализа, проведённого в 2012 году методами молекулярной филогенетики группой исследователей (Grass Phylogeny Working Group II), в составе семейства Poaceae удаётся выделить 12 подсемейств, филогенетические связи между которыми можно отобразить следующей кладограммой (в скобках через дробную черту приведены количества включённых в каждое подсемейство видов)[11]:
Pharoideae[en] (13)
Puelioideae[en] (11)
Oryzoideae[en] (111)
Bambusoideae (1462)
Pooideae (3850)
Aristidoideae[en] (349)
Panicoideae (3236)
Arundinoideae[en] (45)
Micrairoideae[en] (188)
Danthonioideae[en] (281)
Chloridoideae[en] (1607)
Зла́ки, или Злаковые (лат. Gramíneae), или Мя́тликовые (лат. Poáceae) — семейство однодольных растений, к которому относятся такие известные и давно используемые в хозяйстве растения, как пшеница, рожь, овёс, рис, кукуруза, ячмень, просо, бамбук, сахарный тростник.
禾本科(学名:Poaceae),禾本科主要包括稻亚科、竹亚科、早熟禾亚科等12个亚科和少数不确定类群。最近研究有668属,10000多种。中国有190余属约1200多种。除了荞麦以外,几乎所有的人類主食都是禾本科植物。大自然中的野草不只是动物的食物,还能制造大量氧气,防止水土流失。
地球陆地大约有20%覆盖着草。多种俗称作“某某草”的植物是该科物种,但是必须指出,不是所有的“草”都是禾本科植物。同样,并不是所有禾本科植物都是低矮的“草”,就如竹子,也可以高达十数米,连片成林。
茎(特称为秆)有明显的节,节间常中空,少数为实心。
叶子互生,两列,由叶片和叶鞘组成,叶片狭长,叶脉平行,叶鞘抱茎,一边常开裂,叶鞘同叶片连接处通常有一叶舌。
花很小不明显,两性有时单性,由雄蕊和雌蕊及2-3枚内质的浆片(亦称鳞被)组成,生于2苞片(外面的苞片称外稃,里面的名内稃)内,这些部分合称一小花,一至多朵,两列于一纤细的短轴上,基部通常有2枚无花的苞片,名为颖片,诸小花与颖合成一小穗,小穗无柄或具柄,排成顶生或侧生的圆锥、总状或穗状花序。
果实通常为一颖果,含有大量淀粉质胚乳,稀有浆果、囊果或坚果。
本科与禾本目各科的亲疏关系如下(节点有“†”标志的分支表示其自助法支持率介于60-80%之间,无标志者为80%以上)[2]:
禾本目 Poales二柱草科 Ecdeiocoleaceae
拟苇科 Joinvilleaceae
禾本科 Poaceae
须叶藤科 Flagellariaceae
帚灯草科 Restionaceae
刺鳞草科 Centrolepidaceae
苞穗草科 Anarthriaceae
† †莎草科 Cyperaceae
灯心草科 Juncaceae
梭子草科 Thurniaceae
苔草科 Mayacaceae
黄眼草科 Xyridaceae
谷精草科 Eriocaulaceae
偏穗草科 Rapateaceae
香蒲科 Typhaceae香蒲属 Typha
黑三棱属 Sparganium
凤梨科 Bromeliaceae
本科可分成12个亚科,包含两大演化支——BOP类群与PACMAD类群,关系如下[3][4][5]:
禾本科 Poaceae PACMAD类群虎尾草亚科 Chloridoideae
扁芒草亚科 Danthonioideae
百生草亚科 Micrairoideae
芦竹亚科 Arundinoideae
黍亚科 Panicoideae
三芒草亚科 Aristidoideae
BOP类群早熟禾亚科 Pooideae
竹亚科 Bambusoideae
稻亚科 Oryzoideae
姜叶竺亚科 Puelioideae
服叶竺亚科 Pharoideae
柊叶竺亚科 Anomochlooideae
禾本科植物具有极其重要的意义。
除了荞麦以外,几乎所有的粮食都是禾本科植物,如小麦、稻米、玉米、大麦、高粱等。猪、牛、马、羊等各类家畜也都吃草。竹子在日常生活处处可见,在东南亚还有竹子造的房屋。大自然中的野草不止是动物的食物,还能制造大量氧气,防止水土流失。
此外,许多种类可作造纸和纺织原料,或做牧草、防沙固堤和庭园绿化作用。
禾本科(学名:Poaceae),禾本科主要包括稻亚科、竹亚科、早熟禾亚科等12个亚科和少数不确定类群。最近研究有668属,10000多种。中国有190余属约1200多种。除了荞麦以外,几乎所有的人類主食都是禾本科植物。大自然中的野草不只是动物的食物,还能制造大量氧气,防止水土流失。
地球陆地大约有20%覆盖着草。多种俗称作“某某草”的植物是该科物种,但是必须指出,不是所有的“草”都是禾本科植物。同样,并不是所有禾本科植物都是低矮的“草”,就如竹子,也可以高达十数米,连片成林。
 分類(APG III) 界 : 植物界 Plantae 階級なし : 被子植物 Angiosperms 階級なし : 単子葉類 Monocots 階級なし : ツユクサ類 Commelinids 目 : イネ目 Poales 科 : イネ科 Poaceae (Gramineae) 学名 Poaceae (R.Br.) Barnhart クレード
分類(APG III) 界 : 植物界 Plantae 階級なし : 被子植物 Angiosperms 階級なし : 単子葉類 Monocots 階級なし : ツユクサ類 Commelinids 目 : イネ目 Poales 科 : イネ科 Poaceae (Gramineae) 学名 Poaceae (R.Br.) Barnhart クレード イネ科(イネか、Poaceae)は、おおよそ700属と8000種が属する被子植物単子葉類の大きな科である。世界中で広く分布する[1]。古くは禾本科(かほんか)又はホモノ科とも呼んだ。
草本か、あるいは木質化した中空の茎をもつ木本的な植物である[1]。
葉は平行脈をもち、細長く、薄いものが多い。一部で基部でねじれて裏表逆転するものがある。葉は根元から生える根出葉と、茎の途中に生える茎葉がある。ススキのように根出葉が多いものもあれば、ヨシのように茎葉が多いものもある。一部のものは、匍匐枝や地下茎があって横に這う。
葉の基部が茎に巻き付いて葉鞘を形成するが、カヤツリグサ科のように、葉鞘の両端が融合して筒になることは少ない。また、葉鞘の先端にヒレのような出っ張りが生じ、これを葉舌(ようぜつ)という。葉身が葉鞘に至るところで膨らむことがあるがここを葉耳と呼ぶ。葉舌、葉耳を種の同定の目安にするグループも少なくない。
茎は節があり、節ごとに葉がつき、根が円周上に出る。茎は往々にして中空で、節の所がしきりになる。茎は木質化して堅くなる場合があり、特にタケ亜科のものは材木に使えるほど丈夫になる。ただし、形成層はないので、成長して太くなることはない。タケ亜科は分類体系によってはイネ科から分離し、タケ科として扱うこともある。
イネ科植物は、花粉媒介を風によっておこなう風媒花へと進化したものである[1]。そのため、その花は花弁などを失い、雌しべは長くて毛が生えていることが多い。また、花序が変形した小穂と呼ばれる偽花を単位とし、これが集合して穂を形成する。小穂内の子房が成熟した果実を穎果と呼び、これがさらに外穎と内穎、時には苞穎も加わった籾殻にくるまれる。風媒ではあるが、花粉を食物とするハエの仲間やハナバチなどの昆虫が訪れるのを見ることも多く、イネ科の花粉を専門に集めるハナバチもある。ただし、麦角菌に感染したイネ科植物の穂に昆虫が集まるのは、麦角菌が分生子を含んだ蜜を分泌するためである。
比較的下等なものでは小穂は多数の同じような花を並べた形であり、より進んだものでは、部分的な退化や様々な特殊化が見られる。雄花と雌花の分化が見られるものもある。なお、このような特徴はイネ科の属の分類には欠かせない重要な特徴である。そのため、不明のイネ科の同定をおこなう場合、花が咲く直前のものを調べるのがよいとされている。
非常に種類が多く、実に様々な環境に生育するものがある。森林や高山に生育するものもあるが、草原はイネ科の植物を中心に構成されることが多い。
双子葉植物が生育する草地では、一番上の層に同化組織が集中し、その内部は暗くなる傾向があるが、イネ科植物の形成する草原は、茎も葉も立っているので、根元まで光が入る。
水辺にもカヤツリグサ科やイグサ科とともに大きな群落をなす種がある。
イネ(米)、コムギ(小麦)、トウモロコシ、オオムギ、ライムギなど、狭義の穀物はイネ科に属する。その他サトウキビやタケなど馴染深い資源植物が多く含まれる。ススキやパンパスグラスもイネ科に属する。
代表的な使用例
また肉食動物のネコも燕麦などのイネ科の植物を時折食べる習性を持つため、飼い猫用に猫草としてペットショップなどで売られている事もある。
イネ科は、初期に枝分かれしたいくつかの原始的で互いには疎遠な亜科と、BEP clade(イネ、ムギ、タケなど)、PACAD clade(トウモロコシ、キビ、ヒエ、ススキなど)という2つの大きな単系統に分かれる。BEP clade と PACAD clade は互いに姉妹群である。[2]
亜科・連への分け方は諸説あるが、系統が明らかになるにつれまとまりつつある。ここに示すのはGrass Phylogeny Working Group[3] による12亜科約40連である。さらに Micrairoideae 亜科を認める説もある[4]が、最近の研究によれば Micrairoideae は単系統ではなく[2]位置づけは不確実である。
亜連は省略し、属・種は重要なもののみリストアップしている。
タケ類。かつてはイネ科のうち最初に分岐したという説もあり、タケ科 Bambusaceae とする場合もあった。
イネ亜科 Oryzoideae とも。
小穂は多数花、二花の場合、先端から退化傾向。
Micrairoideae亜科を認める場合は PACCMAD clade とすることもある。
小穂は二花、下側は実をつけない傾向。
벼과(-科, 학명: Poaceae 포아케아이[*])는 벼목의 과이다.[1] 화본(禾本, 영어: grass 그래스[*])으로도 알려져 있다. 고등 식물 중 가장 큰 과의 하나이다.
세계 각지에서 흔히 볼 수 있는데, 700여 속의 12,000여 종 가량이 알려져 있다. 벼나 밀, 옥수수를 비롯하여 식량이나 사료로 이용되는 매우 중요한 종류가 많다.
한해살이 또는 여러해살이 초본이 대부분이지만 대나무류와 같이 목질화되는 것도 있다. 줄기는 속이 빈 원통형으로, 잎의 밑부분은 잎집이 되어 줄기를 둘러싸고 있는데 그 위 끝에는잎혀라고 부르는 돌기가 있다. 꽃차례는 1개 또는 몇 개의 꽃과 몇 종류의 포엽이 모여서 작은이삭이라는 특별한 단위를 이루고 있다. 작은이삭의 밑부분에는 2개의 포영이 있고, 그것에 이어 1개 또는 몇 개의 호영이 2열로 어긋나는데, 그 겨드랑이에 꽃이 달린다. 호영은 까락이 생기는 일도 있다.
한편, 호영의 안쪽에는 2개의 주맥을 가진 내영이 있는데, 이것은 2개의 바깥 꽃덮이조각이 합쳐진 것이다. 또, 그 안쪽에 2개의 작은 비늘껍질이 있다. 이것은 내꽃덮이조각에 해당되는데, 꽃이 필 때는 팽윤하여 영류(穎類)를 밀고 벌어지게 된다. 수술은 3-6개가 있고, 씨방은 원래 3개의 심피로 이루어져 있다고 생각되었으나, 실제로는 격벽이 없어져 1개의 방으로 되어 있으며, 그 속에 1개의 밑씨가 생길 뿐이다. 암술머리는 2-3개인데 깃볼로 털이 나 있다. 대부분 풍매화이며 씨는 성숙하면 과피의 안쪽과 합쳐져 영과라고 불리는 폐과가 된다. 꽃차례는 다수의 작은이삭으로 이루어지는데, 작은이삭에 복자루가 있을 경우에는 원추꽃차례가 되는 데 비해, 꽃자루가 없을 경우에는 이삭꽃차례가 된다.
계통 분류는 다음과 같다. 괄호 안의 숫자는 추정되는 종의 수이다.[2]
벼과아노모클로아아과 C3 (4)
파루스아과 C3 (13)
푸엘리아아과 C3 (11)
BOP 분지군벼아과 C3 (110)
대나무아과 C3 (1450)
포아풀아과 C3 (3850)
PACMAD 분지군아리스티다아과 C3 & C4 (350)
기장아과 C3 & C4 (3250)
물대아과 C3 (50)
미크라이라아과 C3 & C4 (200)
꼬인새풀아과 C3 (300)
나도바랭이아과 C3 & C4 (1600)
벼과(-科, 학명: Poaceae 포아케아이[*])는 벼목의 과이다. 화본(禾本, 영어: grass 그래스[*])으로도 알려져 있다. 고등 식물 중 가장 큰 과의 하나이다.
세계 각지에서 흔히 볼 수 있는데, 700여 속의 12,000여 종 가량이 알려져 있다. 벼나 밀, 옥수수를 비롯하여 식량이나 사료로 이용되는 매우 중요한 종류가 많다.
한해살이 또는 여러해살이 초본이 대부분이지만 대나무류와 같이 목질화되는 것도 있다. 줄기는 속이 빈 원통형으로, 잎의 밑부분은 잎집이 되어 줄기를 둘러싸고 있는데 그 위 끝에는잎혀라고 부르는 돌기가 있다. 꽃차례는 1개 또는 몇 개의 꽃과 몇 종류의 포엽이 모여서 작은이삭이라는 특별한 단위를 이루고 있다. 작은이삭의 밑부분에는 2개의 포영이 있고, 그것에 이어 1개 또는 몇 개의 호영이 2열로 어긋나는데, 그 겨드랑이에 꽃이 달린다. 호영은 까락이 생기는 일도 있다.
한편, 호영의 안쪽에는 2개의 주맥을 가진 내영이 있는데, 이것은 2개의 바깥 꽃덮이조각이 합쳐진 것이다. 또, 그 안쪽에 2개의 작은 비늘껍질이 있다. 이것은 내꽃덮이조각에 해당되는데, 꽃이 필 때는 팽윤하여 영류(穎類)를 밀고 벌어지게 된다. 수술은 3-6개가 있고, 씨방은 원래 3개의 심피로 이루어져 있다고 생각되었으나, 실제로는 격벽이 없어져 1개의 방으로 되어 있으며, 그 속에 1개의 밑씨가 생길 뿐이다. 암술머리는 2-3개인데 깃볼로 털이 나 있다. 대부분 풍매화이며 씨는 성숙하면 과피의 안쪽과 합쳐져 영과라고 불리는 폐과가 된다. 꽃차례는 다수의 작은이삭으로 이루어지는데, 작은이삭에 복자루가 있을 경우에는 원추꽃차례가 되는 데 비해, 꽃자루가 없을 경우에는 이삭꽃차례가 된다.
_flower.JPG)
_plant.JPG)

_3.jpg)














































.jpg)






.jpg)












































































































-fr.jpg)




.jpg)













.jpg)










.jpg)
.jpg)






































.jpg)






.jpg)

.jpg)

.jpg)




{kind=link}
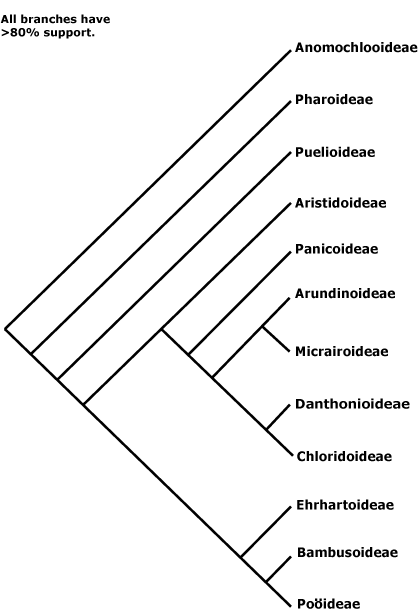{kind=link}