
Is the lamprey,
Pteromyzon (Fig. 1), the basalmost fish? Or is it a large, derived lancelet (Fig. 2) with eyes, external gill openings and parasagittal fins?

Figure 1. Lamprey adult (Pteromyzon) and larvae in vivo and sagittal section.Pteromyzon marinus (Linneaus 1758) is the extant lamprey, a large lancelet in which some metamorphiosized adults attach themselves to fish to suck their blood. Others do not feed as adults, but live off reserves obtained by filter feeding while young. The gill openings open to the outside, not into an atrium.
The answer might come down to the question,
is a skull present in lampreys? Fish have a skull (Fig. 4). Lancelets (Fig. 2) do not.

Figure 2. Extant lancelet (genus: Amphioxus) in cross section and lateral view. The gill basket nearly fills an atrium, which intakes water + food, sends the food into the intestine and expels the rest of the water.
Cambrian lancelets
like Metaspriggina (Fig. 3) have eyes, but no skull. So do flatworms, lampreys and conodont animals. Transitional taxa between flatworms and lampreys, like nematodes and extant lancelets, lack eyes. So do all the closest relatives of basal chordates: echinoderms, crinoids, tunicates. The loss of eyes appears to be a derived trait in hagfish (Fig. 3) and, by convergence, chordate relatives derived from lancelets. Lamprey eyes are complex structures that took a long time to evolve. Apparently blindness is a derived trait on generally sessile chordate cousins, starting with lancelets.
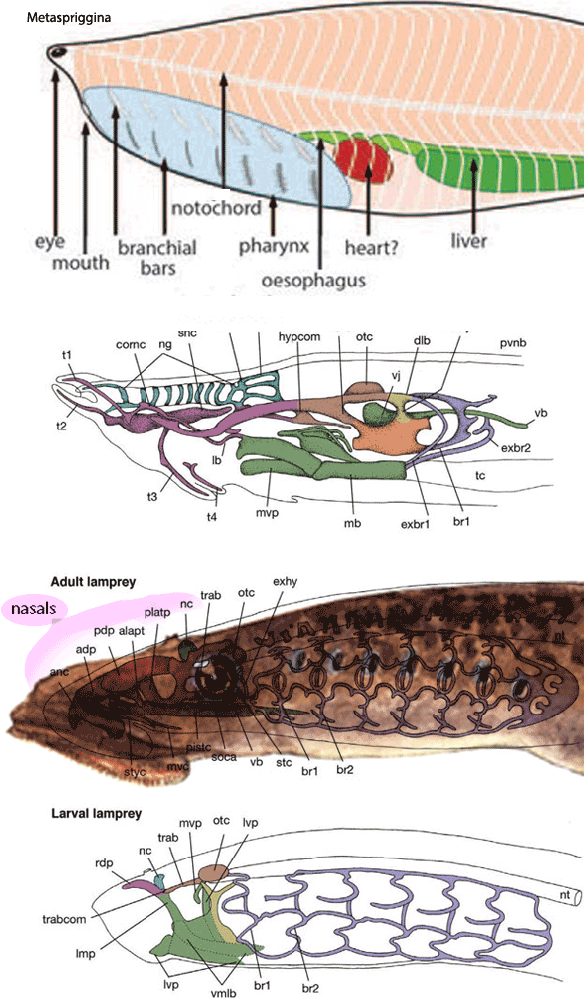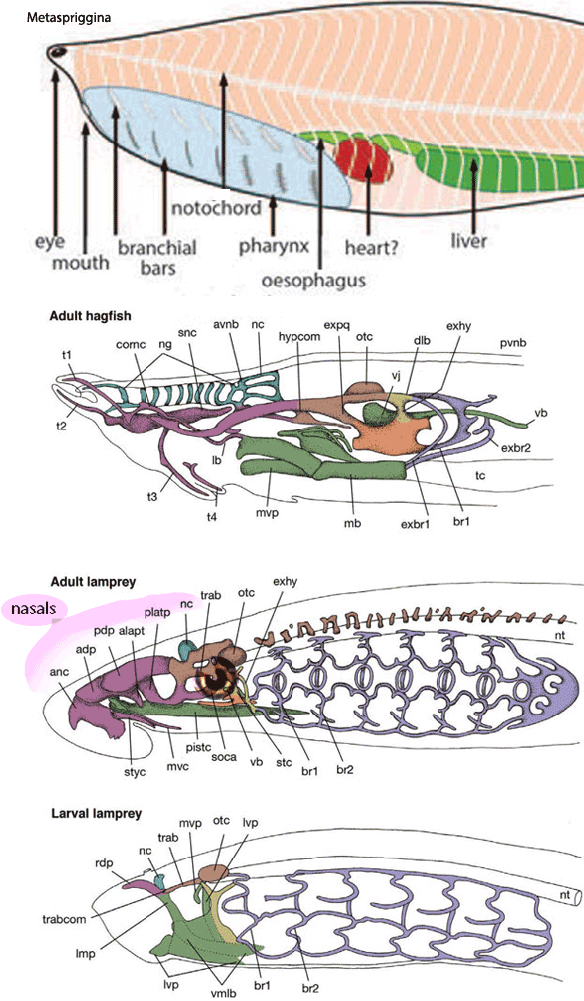
Figure 3. Hagfish and lamprey cranial organs compared to the lancelet, Metaspriggina. No dermal skull material, other than the nasal, is present in the lamprey.
Perhaps now is a good time to consider definitions
to help decide the lamprey ‘fish or lancelet’ question.
According to Wikipedia (Lamprey),
“Lampreys are an ancient extant lineage of jawless fish of the order Petromyzontiformes, placed in the superclass Cyclostomata.
According to Wikipedia (Fish),
“Fish are gill-bearing aquatic craniate animals that lack limbs with digits.”
Note that both definitions rely on traits,
not phylogenetic placement.
Even so,
does the lamprey skull (= cranial and annular cartilage) have homologs in the dermal skull of craniates?
Not really, except perhaps, the nasal,
which is the largest bone/cartilage in many basal fish. The appearance of more dermal cranial bones would have been phylogenetically gradual. We wait until Drepanaspis, Drepanolepis and Birkenia for cranial homologs of many bones to appear in these valid jawless craniates.
You might think the loss of eyes in lancelets
resulted in the loss of primitive cartilage structures that held the eyes, as in large-eyed conodonts, but even in lampreys, such dermal skull structures are not present (Fig. 1).
It’s always phylogeny, not a short list of defining traits.
In the large reptile tree (LRT, 1729+ taxa; subset Fig. 5) the last common ancestor of lampreys (genus: Pteromyzon; Fig. 1), is the lancelet Metaspriggina (Fig. 3) The last common ancestor of all extant fish is Euphanerops, another jawless fish from the Late Devonian with an earlier radiation in the Silurian. Middle Silurian Birkenia (Fig. 4) is the most primitive fish with skull bones homologous with those of extant fish. A less primitive, Early Silurian Jamoytius (Fig. 5) has been considered a sister to lampreys, but in the LRT nests between Birkenia and Thelodus.

Figure 4. Birkenia skull has bones still found in derived craniates, including fish, birds and mammals. Here the nasal is still the largest skull bone.
Armored lancelets
Arandaspis, and Porapsis have an armored gill chamber with few to no bone homologs with those of living craniates. Yes, that armor forms a sort of skull, but it is not the same skull as in Birkenia and its craniate descendants. Arandaspis, and Porapsis have been called primitive fish, but phylogenetically precede valid craniates, which means they cannot be ‘fish’, by definition. For the same reason, neither can the lamprey, Pteromyzon, be called a fish.

Figure 5. Subset of the LRT focusing on basal chordates and Jamoytius (at right).
Lamprey origins
go back to the early Ordovician. 38 extant and 5 extinct species are known. Like other lancelets and basal fish, primitive and young lampreys are filter feeders. The closest living relatives of lampreys are hagfish (Fig. 2) and lancelets.
As hypothesized earlier
here, the lack of a lens, iris and eyeball in lancelets was retained by derived taxa including tunicates and echinoderms, distinct from the order proposed by Romer and others.
Another point-of-view, according to Gregory 1933:
“The profound researches of Stensio (1927) and Kiser (1924) have left no reasonable doubt however, that one or another of the ostracoderms gave rise to the modern class of cyclostomes, including the lampreys and hags, thus confirming the earlier views of Cope and others.”
Gregory 1933 continues:
“Both Gaskell and Stensio have shown the striking similarities of the larval lamprey head to that of the cephalaspid ostracoderms and in the light of much evidence it seems highly probable that the lamprey skull type has been derived from a cephalaspid-like type in the following way:
- “the bony exoskeleton has lost its bone cells and become membranous;
- thorny epidermal teeth have developed around the sucker-like mouth;
- a rasping apparatus has developed out of the so-called tongue, which is a specialized part of the branchial apparatus;
- the rest of the branchial arches have been displaced backward;
- the cartilages that support the sucker and its teeth have also been enlarged;
- the originally continuous cartilaginous septa between the gill-pouches have become fenestrated, giving rise to the branchial basket;
- a special hydraulic organ, described by T. E. Reynolds (1931) has been developed in the oral chamber to assist in the sucking action of the mouth.”
LRT arguments to Gregory’s 1933
ostracoderm ancestry for lampreys based on the LRT:
- Gregory has it backwards: the bony exoskeleton had not developed yet;
- Agreed. Lamprey ‘teeth’ are not homologous with those of gnathostomes;
- Or the tongue is a new medial organ not associated with paired lateral branchials;
- Not displaced relative to LRT sisters;
- Agreed;
- Or the fenestrations are derived from sisters to Metaspriggina, unknown in 1933;
- Agreed, but only a few lampreys are blood suckers and no juveniles are bloodsuckers.
This is not the first time the LRT has been in the minority.
In the LRT(subset Fig. 5) the lamprey is several nodes more primitive than the ostracoderm, Hemicyclaspis, a sturgeon ancestor with pectoral fins, armor and a heterocercal tail. Again, the best way to find out what a taxon is, is to nest it in a wide gamut cladogram like the LRT.
References
Gregory WK 1933. Fish skulls. A study of the evolution of natural mechanisms. American Philosophical Society 23(2) 1–481.
Linnaeus C 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata.
wiki/Branchiostoma
wiki/Lamprey_Pteromyzon
wiki/Birkenia
wiki/Jamoytius

There’s a lot to address here, so I’ll only address a couple of them:
– Of course there’s nothing in the lamprey chondrocranium that corresponds to anything in the gnathostome dermatocranium. Dermal bones don’t have a cartilaginous precursor. They form in tissue just beneath the dermis, hence the name “dermal bone.” It’s a waste of time to compare the lamprey braincase to any vertebrate dermatocranium; a better comparison would be between a lamprey chondrocranium and a shark chondrocranium. Neither structure has any homolog in anyone’s dermatocranium.
This is why I was puzzled by your claim that the nasal is “the largest bone/cartilage in many basal fish.” Except that it’s never a cartilage. It has no cartilaginous precursor. It doesn’t form in the nasal capsule, either, which is an entirely different anatomical structure. Lampreys have a nasal capsule (which makes sense, as they’re vertebrates and lancelets are not), but no nasal.
I would urge you to read up on skeletal development before pursuing this line further; I’m not trying to be rude, but there’s quite a lot out there on bone and cartilage development that would inform your approach.
– you seem to equate an origin of lampreys among early vertebrates with an “ostracoderm” origin. Except the current consensus doesn’t derive lampreys from “ostracoderms.” It has cyclostomes (lampreys + hagfish) branching off fairly early on the vertebrate tree, with most bony traits arising in other lineages.
I’m honestly trying to help you here. You seem to have some misconceptions about skeletal development that are leading you toward absurd conclusions.
Thanks, Chris. If you look at the tetrapod skull homologs I identify in fish, I think you’ll see what I mean about the nasal. On this blog alone take a look at Birkenia, then Chondrosteus, Hemicyclaspis and Pseudoscaphorhynchus. Importantly, the nasal in tetrapods begins as several homologous bones that fuse or disappear in basal vertebrates, like sturgeons. In the LRT topology cartilage in sharks appears after bone appears in sturgeons and paddlefish, not as a precursor.
You wrote: “you seem to equate an origin of lampreys among early vertebrates with an “ostracoderm” origin. Except the current consensus doesn’t derive lampreys from “ostracoderms.”
Please re-read the text. I quoted from Gregory 1933. The subset of the LRT on Fig. 5 shows lampreys and ostestracans are separated by several taxa, with lampreys branching off immediately after Cambrian headless chordates. You are trying to correct me with an identical tree topology to the one you misread in the LRT and Fig. 5.
You’re misunderstanding what I mean by “precursor.” I’m talking about development, not phylogeny. The dermal bones in a skull don’t ossify in cartilage. They are neither endochondral nor perichondral. They ossify in or beneath the dermis. That’s why it’s meaningless to look for a homolog to the nasal in a lamprey or any other vertebrate with no dermatocranium. No dermatocranium, no homolog to the nasal. You might as well look for something homologous to the radius in a snake – no limb, no radius.
Claiming that homologs of the nasal occur in other early vertebrates is irrelevant to the argument. In those cases, you’re trying to find something homologous to the nasal among dermal bones of the head. I don’t buy the claims for homology, but at least you’re looking at the dermatocranium, where a homolog could, in principle, exist.
Thank you, Chris. Understood. What you report are reflected in the matrix scores.
Then what, exactly, did you mean when you described the nasal as “the largest bone/cartilage in many basal fish?”
Look at http://reptileevolution.com/pseudoscaphirhynchus.htm
http://reptileevolution.com/birkenia.htm
http://reptileevolution.com/polyodon.htm
the pink bone.