 Danushka S. Tennakoon1,2
Danushka S. Tennakoon1,2 Kasun M. Thambugala3
Kasun M. Thambugala3 Nimali I. de Silva1,2
Nimali I. de Silva1,2 Nakarin Suwannarach1,2
Nakarin Suwannarach1,2 Saisamorn Lumyong1,2,4*
Saisamorn Lumyong1,2,4*- 1Research Center of Microbial Diversity and Sustainable Utilization, Faculty of Science, Chiang Mai University, Chiang Mai, Thailand
- 2Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai, Thailand
- 3Genetics and Molecular Biology Unit, Faculty of Applied Sciences, University of Sri Jayewardenepura, Gangodawila, Sri Lanka
- 4Academy of Science, The Royal Society of Thailand, Bangkok, Thailand
Fungal taxonomy has a long history and changed significantly in the last few decades. Most recent studies have witnessed morphology combined with DNA-based molecular analyses as the main research tool for fungal species identification. During field surveys, some interesting Didymosphaeriaceae species were found from plant litter in China and Thailand. Morphology combined with phylogenetic analyses (Bayesian and maximum likelihood) of ITS, LSU, SSU, tef1-α, and tub2 loci was used to identify fungal taxa. In this article, three new species and six new host records are described. The new species, Montagnula acaciae, Paraconiothyrium zingiberacearum, and Paraphaeosphaeria brachiariae, can be distinguished from other species of the respective genera based on their distinct size differences (ascomata, asci, and ascospores) and DNA sequence data. The new host records, Montagnula jonesii, Paraconiothyrium fuckelii, Spegazzinia deightonii, and S. tessarthra are reported from Ficus benjamina, Dimocarpus longan, Hedychium coronarium, and Acacia auriculiformis respectively, for the first time. Also, Paraconiothyrium archidendri and P. brasiliense are reported for the first time from Magnolia sp. in China. Moreover, Paraconiothyrium rosae is synonymized under P. fuckelii based on close phylogeny affinities and morphological characteristics. In-depth morphological descriptions, micrographs, and phylogenetic trees are provided to show the placement of new taxa.
Introduction
Fungi are an essential and indispensable component of the ecosystem (Hawksworth, 2001; Gafforov, 2017; Hernandez-Restrepo et al., 2017; Purahong et al., 2017; Field and Pressel, 2018; Hyde et al., 2018a,b). They perform irreplaceable functions in the ecosystem such as nutrient cycling, responding to plant growth, maintaining plant diversity, and organic matter decomposition (Osono, 2017; Purahong et al., 2017; Li et al., 2018). Fungi are regarded as “key players” in the decomposition of litter due to their capability of secreting different kinds of enzymes (Promputtha et al., 2017; Tennakoon et al., 2021). The lignocellulose matrix of litter, which cannot be broken down by most species, is degraded by these enzymes (Romaní et al., 2006; Osono, 2007; Zhang et al., 2018; Tennakoon et al., 2021). Fungi have different lifestyles, such as biotrophs, endophytes, hemi-biotrophs, necrotrophs, and saprotrophs (Barelli et al., 2016; Hyde et al., 2018a; Tang et al., 2018; Tennakoon et al., 2021). They also differ greatly in terms of morphological characteristics, phylogeny characteristics, reproduction modes, and life cycles (Drinkwater et al., 2008; Purahong et al., 2017; Willis, 2018; Phookamsak et al., 2019). Therefore, identification of fungi is crucial to understanding their diversity and roles in the ecosystem. However, proper identification of fungal species is challenging. Despite challenges, mycologists have put considerable effort to identify fungal species worldwide (Lumbsch et al., 2011; Błaszkowski et al., 2013; Csata et al., 2013; Hyde et al., 2013; Giraldo et al., 2019).
Didymosphaeriaceae, one of the Dothideomycetes families, is an interesting taxonomic group of fungi in Pleosporales (Hyde et al., 2013; Hongsanan et al., 2020). Munk (1953) reported this family to contain Didymosphaeria. Didymosphaeriaceae members are morphologically diverse, and most of the sexual morphs have uni-septate, brown ascospores (Hongsanan et al., 2020). Asexual morph can be either coelomycetes (e.g., Alloconiothyrium, Letendraea, Paraconiothyrium, and Paraphaeosphaeria) or hyphomycetes (e.g., Spegazzinia) (Ariyawansa et al., 2014a,b; Thambugala et al., 2017; Hongsanan et al., 2020). The members of this family are cosmopolitan, and most of them have been recognized as saprobes. Meanwhile, some have been found as pathogens or endophytes in diverse plant substrates (e.g., leaves and twigs) in different ecosystems (e.g., marine, terrestrial, and mangroves) (Ariyawansa et al., 2013; Hyde et al., 2013; Liu et al., 2015; Wanasinghe et al., 2016; Hongsanan et al., 2020). Recently, numerous genera have been introduced in Didymosphaeriaceae. For instance, during the past 2 years period, four genera have been introduced, namely, Cylindroaseptospora (Jayasiri et al., 2019), Neptunomyces (Gonçalves et al., 2019), Vicosamyces (Phookamsak et al., 2019), and Chromolaenicola (Mapook et al., 2020). Hongsanan et al. (2020) added 32 genera in this family. Of them, some are highly diverse (e.g., Didymosphaeria, Kalmusia, Paraphaeosphaeria, Pseudocamarosporium, Pseudopithomyces, and Spegazzinia), and some genera have a few number of species (e.g., Alloconiothyrium, Austropleospora, Barria, Bimuria, Cylindroaseptospora, Deniquelata, Didymocrea, Kalmusibambusa, Lineostroma, Vicosamyces, and Xenocamarosporium) (Hongsanan et al., 2020).
As a part of the fungal diversity study, we have identified nine taxa from plant litter substrates (China and Thailand), which belong to the family Didymosphaeriaceae. Their taxonomic positions were established through morphology combined with phylogenetic analyses.
Materials and methods
Sample collection, morphological studies, and isolation
Plant litter (dead leaves and stems) samples were collected from China and Thailand. All the collected samples were examined under a stereo-microscope (AXIOSKOP 2 PLUS Series, Göttingen, Germany). Squash mount preparations were prepared to determine fungal microscopic features (e.g., asci, ascospores, conidia, and pseudoparaphyses). All the images were captured under an Axioskop 2 Plus (Göttingen, Germany) compound microscope equipped using a Canon Axiocam 506 color digital camera (Hanover, Germany). Lactoglycerol and nail polish were used to prepare the permanent slides. All the photo plates were prepared by Adobe Photoshop CS3 Extended version 10.0 software (Adobe Systems, USA), and measurements were taken by ZEN2 (blue edition).
DNA extraction and polymerase chain reaction (PCR) amplification
The isolation process was used for the single spore isolation, as described by Senanayake et al. (2020). Potato dextrose agar (PDA) was used to transfer germinated spores, which were then incubated at 25°C. Following that, careful sub-culturing was carried out to obtain pure cultures. After 3 weeks, culture characteristics (on PDA) were observed. All the type specimens were deposited in the herbariums of Mae Fah Luang University (MFLU) and National Chiayi University (NCYU). The culture collections of Mae Fah Luang University (MFLUCC) and National Chiayi University (NCYUCC) were used to deposit the living cultures. Numbers for Faces of Fungi and Index Fungorum were used as mentioned in Jayasiri et al. (2015) and Index Fungorum (2022).
The fungal colonies growing on PDA (3 weeks) were used for genomic DNA extraction. Liquid nitrogen was used to grind the mycelium into a fine powder, and the DNA was extracted using the DNA extraction kit (E.Z.N.A Fungal DNA Mini Kit, D3390-02, Omega Bio-Tek) following the manufacturer's instructions. Montagnula jonesii (MFLU 18-0084) was subjected to direct DNA extraction using a DNA extraction kit from BioFlux®, Hangzhou, P.R. China, in accordance with the manufacturer's instructions. The DNA products were stored for a long period of time at −20 °C and retained at 4°C for DNA amplification. DNA was amplified by polymerase chain reaction (PCR) for obtaining the five genes: the large subunit (28S, LSU), small subunit (18S, SSU), internal transcribed spacers (ITS1-5.8S-ITS2), translation elongation factor 1-alpha gene (tef1-α), and β-tubulin (tub2). The LSU gene was amplified using the primers LR0R (5′-TCCTGAGGGAAACTTCG-3′) and LR5 (5′-ACCCGCTGAACTTAAGC-3′) (Vilgalys and Hester, 1990; Rehner and Samuels, 1994); the SSU gene was amplified using the primers NS1 (5′-GTAGTCATATGCTTGTCTC-3′) and NS4 (5′-CTTCCGTCAATTCCTTTAAG-3′) (White et al., 1990); nuclear ITS was amplified using the primers ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) (White et al., 1990); tef1-α gene was amplified using the primers EF1-983F (5′-GCYCCYGGHCAYCGTGAYTTYAT-3′) and EF1-2218R (5′-ATGACACCRACRGCRACRGTYTG-3′) (Rehner, 2001); and beta-tubulin (tub2) gene was amplified using the primers BT2a (5′-GGTAACCAAATCGGTGCTGCTTTC-3′) and BT2b (5′-ACCCTCAGTGTAGTGACCCTTGGC-3′) (Glass and Donaldson, 1995). Sterilized water (9.5 μl), 2 × Power Taq PCR MasterMix (Bioteke Co., China) (12.5 μl), each forward and reverse primers (1 μl), and DNA template (1 μl) were used for amplification reactions. The PCR thermal cycle program for ITS, LSU, SSU, tef1-α, and tub2 was performed, as described by Conforto et al. (2019) and Tennakoon et al. (2020). PCR products were sent to Shanghai Sangon Biological Engineering Technology and Services Co., Ltd, China, for the purification and sequencing. All the obtained sequences were deposited in GenBank (Tables 1–4).
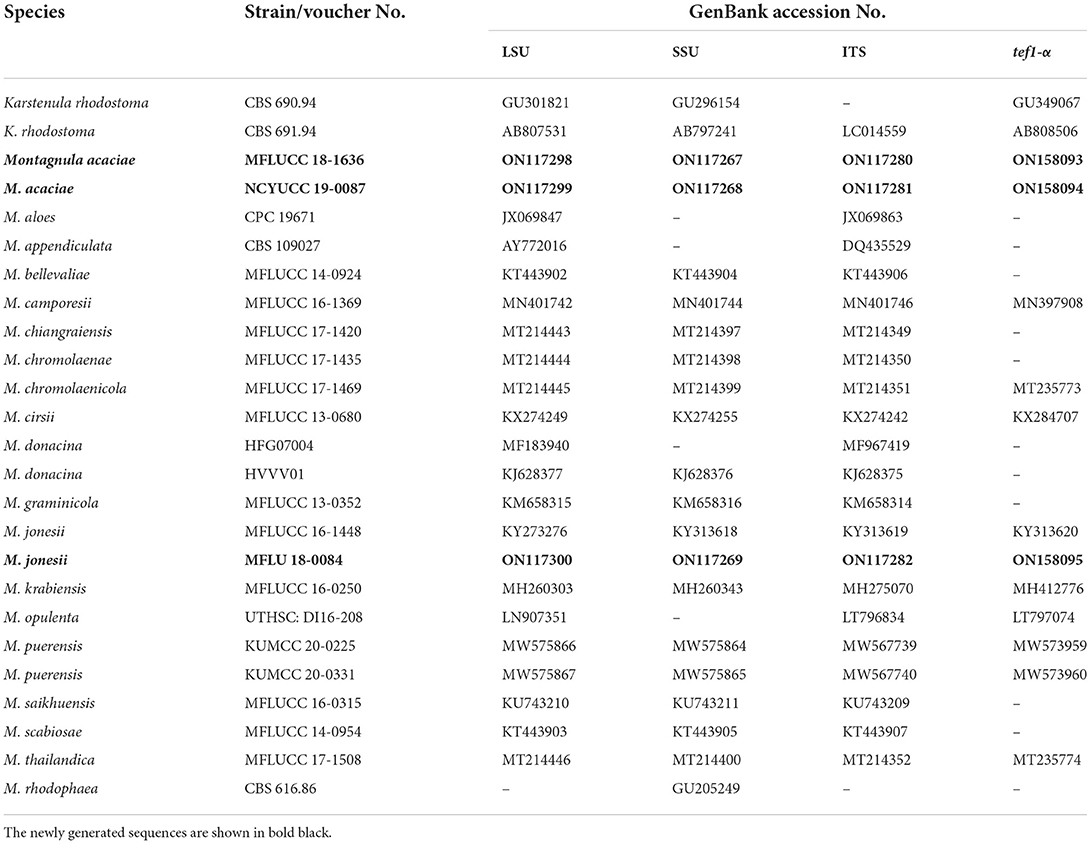
Table 1. GenBank and culture collection accession numbers of species included in this phylogenetic study (Montagnula tree).
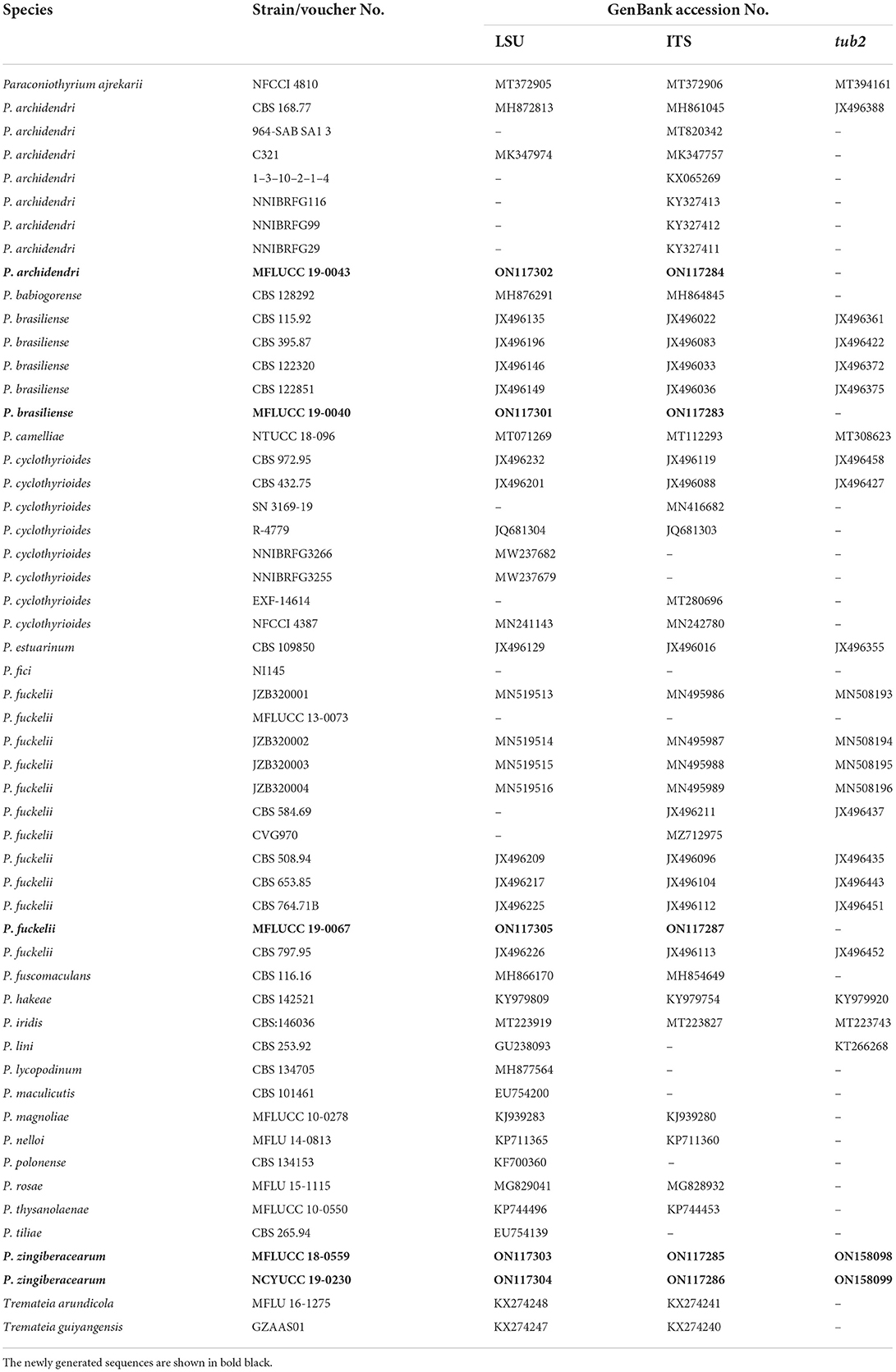
Table 2. GenBank and culture collection accession numbers of species included in this phylogenetic study (Paraconiothyrium tree).
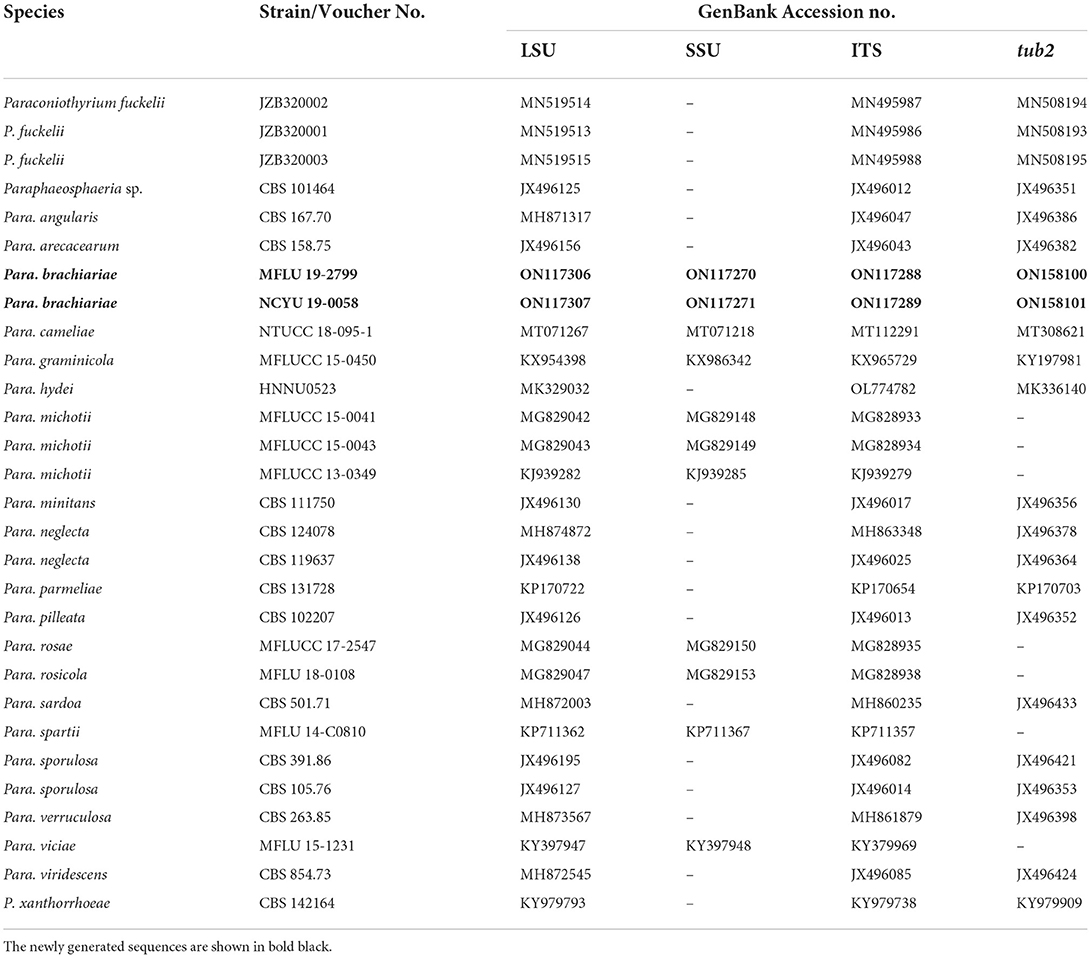
Table 3. GenBank and culture collection accession numbers of species included in this phylogenetic study (Paraphaeosphaeria tree).
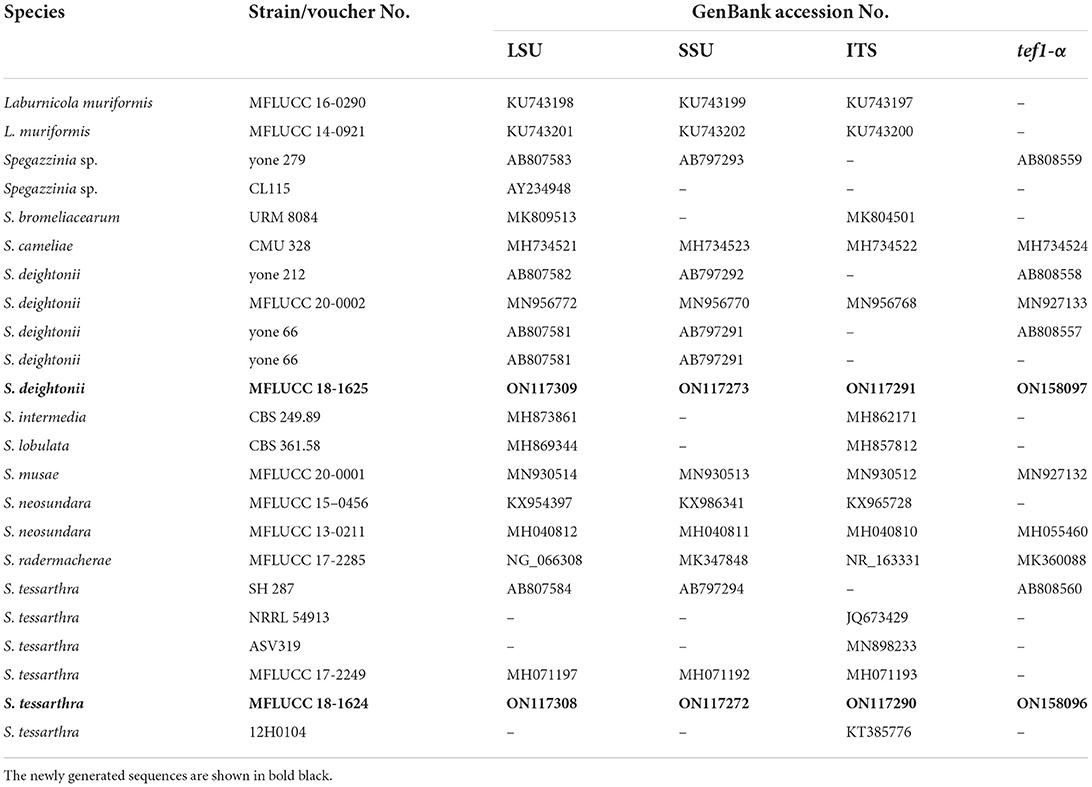
Table 4. GenBank and culture collection accession numbers of species included in this phylogenetic study (Spegazzinia tree).
Phylogenetic analyses
A combined gene dataset of ITS, LSU, SSU, tef1-α, and tub2 was used for the phylogenetic analyses. The newly attained sequences were initially subjected to BLASTn searches in GenBank (http://www.ncbi.nlm.nih.gov/) to identify the taxa that shared the most similarities with our strains. The additional sequences included in the analysis were from previous publications (Phookamsak et al., 2019; Samarakoon et al., 2020; Boonmee et al., 2021; Tennakoon et al., 2021). MAFFT v.7 web server (http://mafft.cbrc.jp/alignment/server) was used to make multiple alignments (Katoh and Standley, 2013). The alignment was corrected manually using BioEdit v.7.0.5.2 (Hall, 1999), where necessary.
The CIPRES Science Gateway platform (Miller et al., 2010) was used to create maximum likelihood trees using the GTR + I + G model of evolution and RAxML-HPC2 on XSEDE (8.2.8) (Stamatakis et al., 2008; Stamatakis, 2014). MrModeltest v.3.7 (Posada and Crandall, 1998) was carried out to check the evolutionary models for phylogenetic analyses under the Akaike Information Criterion (AIC). Model test results revealed “GTR + I + G” as the best-fit model for each locus for Bayesian and maximum likelihood analyses. MrBayes v. 3.1.2 (Huelsenbeck and Ronquist, 2001) was used for Bayesian analysis (Rannala and Yang, 1996; Zhaxybayeva and Gogarten, 2002). The number of generations used in Bayesian analysis were 1,000,000 to 5,000,000, and trees were sampled every 100th or 1,000th generations. The FigTree v1.4.0 (Rambaut, 2012) program was used to view the phylogenetic trees and was reorganized by using Microsoft PowerPoint (2010). All the final alignments were deposited in TreeBASE (http://www.treebase.org/) (Submission ID: 29615).
Results
Phylogenetic and taxonomic results of Montagnula
Montagnula Berl
Notes: Berlese (1896) established this genus to include M. infernalis as the generic type. Montagnula members mostly have globose, spherical, immersed, or semi-immersed ascomata, clavate to cylindrical asci, and multi-septate, fusoid, or ellipsoid ascospores (Ariyawansa et al., 2014a; Pitt et al., 2014). Montagnula species serves a crucial role in the environment as saprobes, which generally grow on the wood and bark of dead plants but also rarely on dead leaves (Hongsanan et al., 2020; Mapook et al., 2020). This genus has a vast range of species all around the world. In addition, given the number of new species revealed through recent investigations, Montagnula appears to be phylogenetically diverse as well (Mapook et al., 2020). Up to date, 39 accepted Montagnula species are listed in Index Fungorum (2022). In this study, we introduce Montagnula acaciae as a new species and new host record of M. jonesii from Ficus benjamina.
Montagnula acaciae Tennakoon and S. Lumyong, sp. nov. (Figure 1).

Figure 1. Montagnula acaciae (MFLU 18-2569, holotype). (a) Ascomata on a dead stem of Acacia auriculiformis. (b) Vertical section through an ascoma. (c) Peridium. (d) Pseudoparaphyses. (e–h) Immature and mature asci. (i–o) Ascospores. (p) Germinating ascospore. (q) Upper side of the culture. (r) Lower side of the culture. Scale bars: (b) = 100 μm, (c) = 8 μm, (d–h) = 30 μm, (i–p) = 5 μm.
Etymology: Name reflects the host genus Acacia.
Index Fungorum Number: IF559599.
Facesoffungi number: FoF 10801.
Holotype: MFLU 18-2569.
Saprobic on dead stem of Acacia auriculiformis A. Cunn. (Fabaceae). Sexual morph: Ascomata 140–180 μm high × 150–200 μm diam. ( = 160 × 175 μm, n = 10), solitary or scattered, coriaceous, immersed to semi-immersed, partly erumpent, unilocular, globose to obpyriform, brown to dark brown, ostiolate, with minute papilla, filled with hyaline cells. Peridium 10–20 μm wide, outer layer comprising light brown to dark brown, thick-walled, cells of textura angularis, inner layer comprising hyaline, flattened, thin-walled cells of textura angularis. Hamathecium comprising 1.3–2.4 μm wide, cylindrical to filiform, branched, septate pseudoparaphyses. Asci 50–90 × 8–10 μm ( = 72 × 9 μm, n = 20), 8-spored, bitunicate, fissitunicate, clavate to cylindrical, with a long pedicel, slightly curved, rounded at the apex and with a shallow ocular chamber. Ascospores 10–15 × 4–7 μm ( = 12 × 6 μm, n = 30), overlapping, 1–2-seriate, fusiform to ellipsoid, initially hyaline or pale brown, becoming light brown to dark brown, 1-septate, constricted at the septum, upper cell slightly wider, tapering toward ends, slightly curved. Asexual morph: Undetermined.
Culture characteristics: Colonies growing on PDA, 15–20 mm diam. after 3 weeks at 25°C, colonies circular, dense, flattened, surface slightly rough, entire edge, velvety; from above: dark brown at margin, gray to slightly greenish at center; reverse: dark brown to black at the margin and center, mycelium greenish to greenish gray.
Material examined: Thailand, Chiang Mai, on dead stem of Acacia auriculiformis (Fabaceae), 25 March 2016, D.S. Tennakoon, TD001A (MFLU 18-2569, holotype); ex-type living culture, MFLUCC 18-1636. ibid. 28 March 2016, TD001B (NCYU 19-0255, Paratype); ex-paratype living culture, NCYUCC 19-0087.
Notes: The morphological characteristics of Montagnula acaciae agree well with those described for Montagnula species (immersed to semi-immersed or erumpent ascomata; clavate to cylindrical, long pedicellate asci; and light brown to dark brown, 1-septate ascospores) (Hyde et al., 2013; Hongsanan et al., 2020; Mapook et al., 2020). The generated phylogeny (LSU, SSU, ITS, and tef1-α) shows that M. acaciae forms a distinct sister lineage to the clade formed by M. chromolaenicola, M. donacina, M. graminicola, M. puerensis, M. saikhuensis, and M. thailandica with strong statistical support (96% ML, 1.00 BYPP, Figure 2). Morphologically, M. acaciae differs from M. saikhuensis and M. puerensis in having smaller ascomata (140–180 × 150–200 μm), whereas M. saikhuensis and M. puerensis have larger ascomata (400–450 × 400–500 and 300–600 × 230–380 μm). Montagnula donacina differs from M. acaciae in having carbonaceous ascostromata with a flat bottom (Pitt et al., 2014). In addition, M. graminicola can be differentiated from M. acaciae in having verrucous ascospores, not constricted in the middle, and surrounded by obvious sheaths (Liu et al., 2015). Interestingly, we found that the morphological characteristics of M. acaciae share similarities with M. thailandica, but they are phylogenetically distinct (Figure 2). We compared the base pair differences of the ITS (+5.8S) gene region of M. acaciae and M. thailandica, and there were 12 base pair differences (2.18%) across 550 nucleotides. In addition, there were 22 base pair differences (2.51%) across 880 nucleotides of tef1-α gene region as well. The main morphology differences of Montagnula species are provided in Table 5.

Figure 2. Phylogram generated from maximum likelihood analysis is based on combined ITS, LSU, and SSU sequence data (Final likelihood value of −9216.328354). The tree is rooted with Karstenula rhodostoma (CBS 690.94 and CBS 691.94). The ex-type strains are indicated in bold, and the new isolates are indicated in red. Bootstrap support values ≥65% from the maximum likelihood (ML) and Bayesian posterior probabilities (PP) values ≥0.90 are given above the nodes.
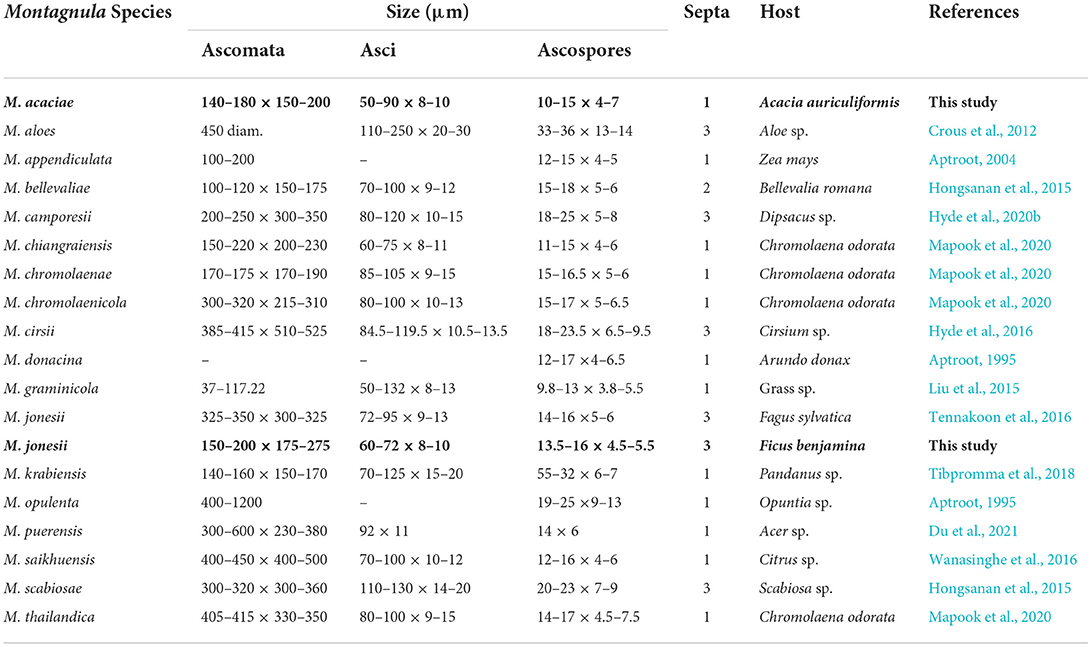
Table 5. Synopsis of Montagnula species treated in this study.
Montagnula jonesii Tennakoon, Wanas., Phook. and K.D. Hyde, Mycosphere 7: 1350 (2016) (Figure 3).

Figure 3. Montagnula jonesii (MFLU 18-0084, new host record). (a,b) Ascomata on a dead stem of Ficus benjamina. (c) Vertical section through an ascoma. (d) Peridium. (e) Pseudoparaphyses. (f–h) Asci. (i–l) Ascospores. Scale bars: (c) = 50 μm, (d) = 10 μm, (e–h) = 30 μm, (i–l) = 8 μm.
Index Fungorum Number: IF552577.
Facesoffungi number: FoF 02719.
Saprobic on dead stem of Ficus benjamina L. (Moraceae). Sexual morph: Ascomata 150–200 μm high × 175–275 μm diam. ( = 175 × 210 μm, n = 10), scattered to clustered, solitary, immersed to semi-immersed, erumpent, globose to sub-globose, glabrous, dark brown to black, uniloculate, ostiolate. Peridium 10–20 μm wide, thin-walled with equal thickness, composed of several layers of hyaline, pseudoparenchymatous cells, arranged in a textura angularis. Hamathecium comprising 1.5–2.5 μm wide, cylindrical to filiform, cellular pseudoparaphyses, septate, not constricted at the septum, anastomosing at the apex, embedded in a gelatinous matrix. Asci 60–72 × 8–10 μm ( = 65 × 9 μm, n = 20), bitunicate, 8-spored, clavate, apically rounded, with a long pedicel, with an ocular chamber. Ascospores 13.5–16 × 4.5–5.5 μm ( = 15 × 5 μm), overlapping 1–2-seriate, ellipsoidal to fusiform with rounded ends, hyaline or pale brown when immature, brown to reddish-brown when mature, mostly 1-septate at immature, 3-septate at mature, constricted at the septa, slightly curved, second cell enlarge from apex, guttulate, smooth-walled. Asexual morph: Undetermined.
Known hosts: Fagus sylvatica and Ficus benjamina (Tennakoon et al., 2016; This study).
Known distribution: Italy and China (Tennakoon et al., 2016; This study).
Material examined: China, Taiwan region, Chiayi, Dahu Forest, on a dead stem of Ficus benjamina (Moraceae), 26 August 2018, D.S. Tennakoon, D008 (MFLU 18-0084).
Notes: Tennakoon et al. (2016) introduced Montagnula jonesii from Fagus sylvatica in Italy. The morphological characteristics of our collection (MFLU 18-0084) resemble M. jonesii (MFLU 16-1363) in having globose to sub-globose, immersed to semi-immersed, erumpent ascomata; clavate, apically rounded, long pedicellate asci; and brown to reddish-brown, ellipsoidal to fusiform, 3-septate ascospores with enlarge second cell from apex (Tennakoon et al., 2016). Our phylogeny also shows that our collection cluster with M. jonesii (MFLU 16-1363), and it is supported by phylogenetic data (100% ML, 1.00 BYPP, Figure 4). We therefore consider MFLU 18-0084 as a new host record of M. jonesii from Ficus benjamina.
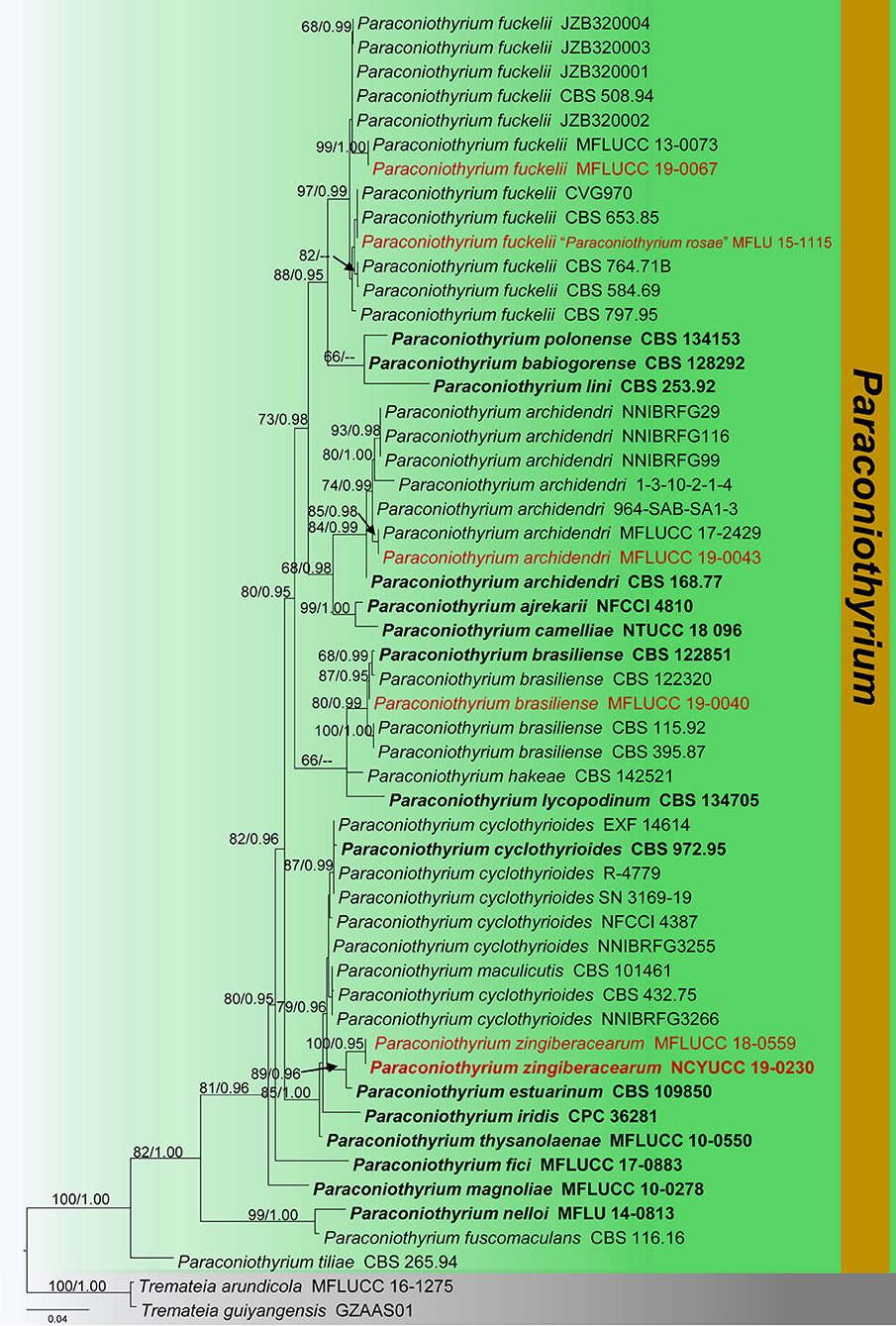
Figure 4. Phylogram generated from maximum likelihood analysis is based on combined ITS, LSU, and tub2 sequence data (final likelihood value of −8278.609544). The tree is rooted with Tremateia arundicola and T. guiyangensis (MFLUCC 16-1275 and GZAAS01). The ex-type strains are indicated in bold, and the new isolates are indicated in red. Bootstrap support values ≥65% of maximum likelihood (ML) and Bayesian posterior probabilities (PP) values ≥0.90 are given above the nodes.
Phylogenetic and taxonomic results of Paraconiothyrium
Paraconiothyrium Verkley
Notes: Paraconiothyrium is a diverse genus, which was introduced by Verkley et al. (2004). Members of this genus play a crucial function as saprobes on dead plants, particularly dead wood, and occasionally on dead leaves (Verkley et al., 2004; Hyde et al., 2013; Ariyawansa et al., 2014b; Hongsanan et al., 2020; Boonmee et al., 2021). In addition, since the species has been found in both temperate and tropical countries (e.g., Australia, China, India, Iran, Laos, Myanmar, Poland, Thailand, the United States, and Uzbekistan), Paraconiothyrium appears to have a global distribution (Verkley et al., 2004, 2014; Budziszewska et al., 2011; Ariyawansa et al., 2014b, 2020; Crous et al., 2017; Gafforov, 2017; Hongsanan et al., 2020; Boonmee et al., 2021). Paraconiothyrium has diverse morphology characteristics, such as eustromatic to pycnidial conidiomata, phialidic or annelidic conidiogenous cells, and hyaline to brown conidia (Verkley et al., 2004, 2014). Currently, 19 Paraconiothyrium species are accepted in Index Fungorum (2022). In this study, we introduce Paraconiothyrium zingiberacearum as a new species, and P. archidendri, P. brasiliense and P. fuckelii as new host records. In addition, P. rosae was synonymized under P. fuckelii based on strong morphological similarities and phylogeny data.
Paraconiothyrium archidendri Verkley, Göker and Stielow, Persoonia 32: 37 (2014) (Figure 5).

Figure 5. Paraconiothyrium archidendri (MFLU 18-2653, new host record): (a) Conidiomata on a dead twig of Magnolia sp. (b,c) Close-up of conidiomata on the host. (d,e) Sections through conidiomata. (f) Conidiomatal wall. (g) Conidiogenous cells with developing conidia. (h–j) Conidia. Scale bars: (a) = 500 μm, (b,c) = 200 μm, (d,e) = 50 μm, (f,g) = 10 μm, (h–j) = 2 μm.
Index Fungorum Number: IF800761.
Facesoffungi number: FoF 05243.
Saprobic on dead twigs of Magnolia sp. (Magnoliaceae). Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 90–120 μm high, 100–140 μm diam. ( = 100 × 125 μm, n = 10), pycnidial, superficial or semi-immersed, dark brown to black, solitary to aggregated, globose to sub-globose. Conidiomatal wall 19–23 μm wide, 2–3 cell layers, composed of brown, cells of textura angularis with relatively thick-walled. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 5–7 × 3–4 μm ( = 6 × 3.5 μm, n = 20), discrete, globose to doliiform, holoblastic, hyaline. Conidia 6–7 × 3–4 ( = 6.5 × 3.5 μm, n = 40) μm, variable in shape, ellipsoid, rarely obovoid, ends rounded, sub-globose or sometimes one end more or less blunt, aseptate, initially hyaline, becoming light brown at maturity, smooth-walled.
Culture characteristics: Colonies growing on PDA, 23 mm diam. after 1 week at 25°C, colonies circular, dense, surface smooth, undulate edge, cottony; from above: pale brown; reverse: brown.
Known hosts: Leucaena sp., Pithecellobium bigeminum, and Magnolia sp. (Verkley et al., 2014; Jayasiri et al., 2015; Goh et al., 2016; this study).
Known distribution: China, Korea, and Myanmar (Verkley et al., 2014; Jayasiri et al., 2015; Goh et al., 2016; this study).
Material examined: China, Xishuangbanna, on dead twigs of Magnolia sp. (Magnoliaceae), 27 March 2017, N.I. de Silva, NI275 (MFLU 18-2653), living culture, MFLUCC 19-0043.
Notes: Verkley et al. (2014) described Paraconiothyrium archidendri, which was associated with leaf spots of Pithecellobium bigeminum in Myanmar. Phylogenetic analyses show that a new strain (MFLUCC 19-0043) clustered with other strains of P. archidendri, in particular close to the isolate MFLUCC 17-2429 with 85% ML and 0.98 BYPP statistical support (Figure 4). The new collection (MFLU 18-2653) morphologically fits with the description of P. archidendri (CBS 168.77 and MFLUCC 17-2429) in having globose to sub-globose pycnidia, globose to doliiform conidiogenous cells and olivaceous-brown, sub-globose or ellipsoid, aseptate conidia (Verkley et al., 2014; Jayasiri et al., 2019). Thus, we report this new collection as a new host record of P. archidendri from dead twigs of Magnolia sp. in China.
Paraconiothyrium brasiliense Verkley, Stud. Mycol. 50: 329 (2004) (Figure 6).

Figure 6. Paraconiothyrium brasiliense (MFLU 18-2649, new host record): (a,b) Conidiomata on a dead twig of Magnolia sp. (c) Close-up of conidiomata. (d,e) Sections through conidiomata. (f) Conidiomatal wall. (g) Sections through conidiomata showing neck region. (h) Conidiogenous cells and developing conidia. (i,j) Conidia. Scale bars: (a) = 500 μm, (b,c) = 200 μm, (d,e) = 50 μm, (f) = 10 μm, (g) = 20 μm, (h–j) = 2 μm.
Index Fungorum Number: IF500082.
Facesoffungi number: FoF 00053.
Saprobic on dead twigs of Magnolia sp. (Magnoliaceae). Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 170–230 μm high, 180–250 μm diam. ( = 200 × 210 μm, n = 10), pycnidial, superficial or semi-immersed, dark brown to black, solitary to aggregated, globose to sub-globose. Conidiomatal wall 15–20 μm wide, outer layer comprising cells of textura angularis with somewhat thickened, brown walls, inner layer comprising flattened, hyaline, thin-walled cells of textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 4–5 × 1–2 μm ( = 4.6 × 1.5 μm, n = 20), discrete or integrated in short, formed from the inner cells all over the conidiomatal wall, simple, broadly ampulliform to globose, holoblastic, phialidic, hyaline to pale yellow. Conidia 2–4 × 1–2 ( = 3 × 1.5 μm, n = 40) μm, variable in shape, ellipsoid to short-cylindrical or obovoid, hyaline at immature, light brown at maturity, aseptate, smooth, rounded at both ends.
Culture characteristics: Colonies growing on PDA, 26 mm diam. after 3 weeks at 25°C, colonies circular, dense, smooth surface, undulate edge; from above: pale brown; reverse: brown.
Known hosts: Acer pentaphyllum, Alliaria petiolate, Cinnamomum camphora, Coffea arabica, Ginkgo biloba, Magnolia sp., Pinus tabulaeformis, Picea glauca and Prunus salicina (Damm et al., 2008; Paul and Lee, 2014; Verkley et al., 2014; Nakashima et al., 2019; this study).
Known distribution: Brazil, Canada, China, Japan, Korea, South Africa and the USA (Damm et al., 2008; Paul and Lee, 2014; Verkley et al., 2014; Nakashima et al., 2019; this study).
Material examined: China, Yunnan Province, Xishuangbanna, on dead twigs of Magnolia sp. (Magnoliaceae), 27 March 2017, N.I. de Silva, NI270 (MFLU 18-2649), living culture, MFLUCC 19-0040.
Notes: Phylogenetic analyses of a concatenated LSU, ITS, and tub2 sequence data show that our strain (MFLUCC 19-0040) forms a well-supported clade, grouping with several strains of Paraconiothyrium brasiliense (80% ML, 0.99 BYPP, Figure 4). In particular, our strain provides a close phylogenetic relationship (87% ML, 0.95 BYPP) with the type of P. brasiliense (CBS 122851) and another strain of P. brasiliense (CBS 1223200). Paraconiothyrium brasiliense was described from fruits of Coffea arabica in Brazil (Verkley et al., 2014). This species shows widespread distribution and found on a wide range of host plants (Damm et al., 2008). Paraconiothyrium brasiliense was isolated as endophytes from Ginkgo biloba (Damm et al., 2008), Acer pentaphyllum in Korea (Paul and Lee, 2014), and healthy twig of Cinnamomum camphora in Japan (Nakashima et al., 2019). P. brasiliense was found associated with necrotic symptoms on Prunus salicina in South Africa (Damm et al., 2008). In this study, we identify our saprobic strain (MFLUCC 19-0040) as P. brasiliense and report its occurrence on Magnolia sp. in China for the first time.
Paraconiothyrium fuckelii (Sacc.) Verkley and Gruyter, Stud. Mycol. 75: 25 (2012) (Figure 7).
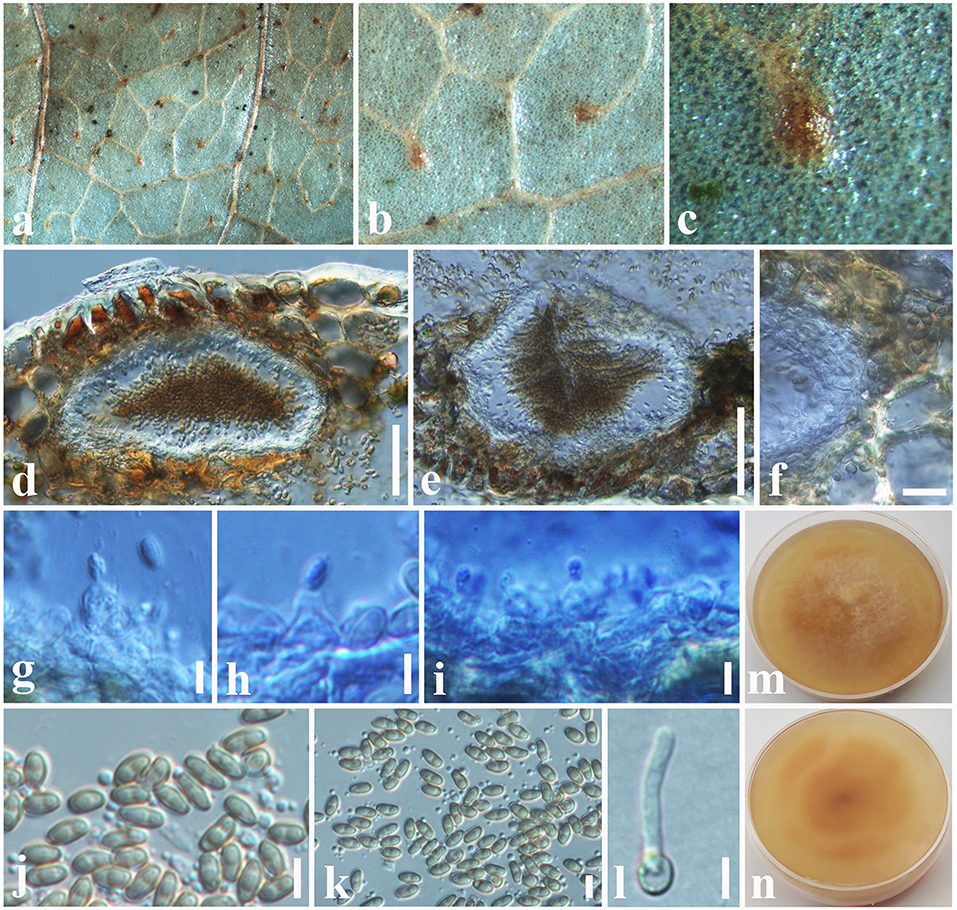
Figure 7. Paraconiothyrium fuckelii (MFLU 18-2605, new host record): (a,b) Conidiomata on a dead leaf of Dimocarpus longan. (c) Close-up of conidioma. (d,e) Sections through conidiomata. (f) Conidiomatal wall. (g–i) Conidiogenous cells and developing conidia. (j,k) Conidia. (l) Germinating conidium. (m) Upper side of the culture. (n) Lower side of the culture. Scale bars: (d,e) = 100 μm, (f) = 10 μm, (g–l) = 3 μm.
Index Fungorum Number: IF564787.
Facesoffungi number: FoF 00055.
≡ Coniothyrium fuckelii Sacc., Fungi venet. nov. vel. Crit., Sér. 5: 200 (1878).
= Paraconiothyrium rosae Senan., Wanas., Camporesi and K.D. Hyde, Fungal Divers. 31 (2018).
Saprobic on dead leaves of Dimocarpus longan Lour. (Sapindaceae). Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 150–300 μm high, 250–500 μm diam. ( = 240 × 320 μm, n = 10), pycnidial, immersed, light brown to dark brown, solitary, globose to sub-globose. Conidiomatal wall 15–20 μm wide, outer layer composed of somewhat thickened, brown-walled cells of textura angularis, inner layer composed hyaline, flattened, thin-walled cells of textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 4–8 × 3–5 μm ( = 5.2 × 4 μm, n = 20), discrete or integrated in short, simple, broadly ampulliform to globose, holoblastic, often annellidic, hyaline. Conidia 3–4 × 1–2.6 ( = 3.5 × 1.5 μm, n = 40) μm, variable in shape, sub-globose to ellipsoid or obovoid, rarely cylindrical, hyaline at immature, light brown at maturity, aseptate, smooth-walled.
Culture characteristics: Colonies growing on PDA, 15–20 mm diam. after 3 weeks at 25°C, colonies circular, slightly dense, smooth surface, crenate edge, velvety; from above: yellowish brown at the margin, brown in the center; reverse: yellowish brown at the margin, light brown in the center, mycelium white to whitish cream.
Known hosts: Dimocarpus longan, Picea abies, Prunus sp., Punica granatum, Rosa hybrid, and Rubus sp. (De Gruyter et al., 2006; Verkley et al., 2014; Gafforov, 2017; Jamali, 2020; this study).
Known distribution: China, Denmark, Germany, Iran, Netherlands, Thailand and Uzbekistan (De Gruyter et al., 2006; Ariyawansa et al., 2014a,b; Verkley et al., 2014; Gafforov, 2017; this study).
Material examined: China, Taiwan region, Chiayi, Ali Shan Mountain, on dead leaves of Dimocarpus longan (Sapindaceae), 05 September 2018, D.S. Tennakoon, TAP020 (MFLU 18-2605); living culture, MFLUCC 19-0067.
Notes: Paraconiothyrium fuckelii was first introduced by Saccardo (1876) from cane lesions on raspberry, similar to Coniothyrium fuckelii. Subsequently, C. fuckelii was synonymized under Paraconiothyrium by De Gruyter et al. (2006), based on morphology combined with phylogeny data. The morphological characteristics of our collection (MFLU 18-2605) resemble previously introduced authentic P. fuckelii isolates in having pycnidial, immersed, light brown to dark brown, globose to sub-globose conidiomata, ampulliform to globose, holoblastic conidiogenous cells and sub-globose to ellipsoid or obovoid, hyaline to light brown, aseptate conidia (Ariyawansa et al., 2014b; Verkley et al., 2014). Multi-gene phylogeny also directs that our collection (MFLU 18-2605) groups with other P. fuckelii isolates in a highly supported clade (97% ML, 0.99 BYPP, Figure 4). In particular, our isolate shows a close phylogeny relationship (99% ML, 1.00 BYPP) with the isolate MFLUCC 13-0073, which was introduced by Ariyawansa et al. (2014b) from Thailand (Figure 4). Hence, we consider MFLU 18-2605 as a new host record of P. fuckelii from Dimocarpus longan (Sapindaceae). Paraconiothyrium fuckelii seems to have a cosmopolitan distribution since it has been reported from different plant families, such as Fabaceae, Rosaceae, Pinaceae, and Sapindaceae worldwide (Kuter, 1986; De Gruyter et al., 2006; Mułenko et al., 2008; Kowalski and Andruch, 2012; Ariyawansa et al., 2014b; Verkley et al., 2014). Many researchers have identified the asexual morph of this species and a sexual morph introduced by Ariyawansa et al. (2014b), which has immersed to erumpent, globose or sub-globose, coriaceous, ostiolate ascomata; clavate asci; and pale brown, 3-septate, narrowly ovoid to clavate ascospores.
In addition, P. rosae was introduced by Wanasinghe et al. (2018) from dead aerial spines of Rosa canina (Rosaceae) without statistical support. In the present phylogenetic analyses, P. rosae groups well with P. fuckelii isolates in a highly supported clade (97% ML, 0.99 BYPP, Figure 4). The morphological characteristics of P. rosae also agree well with P. fuckelii in having pycnidial, immersed or semi-immersed, globose to sub-globose conidiomata and sub-globose to ellipsoid or obovoid, hyaline to light brown, aseptate conidia (Ariyawansa et al., 2014b; Verkley et al., 2014; Wanasinghe et al., 2018). Therefore, we treat P. rosae as a synonym of P. fuckelii in this study.
Paraconiothyrium zingiberacearum Tennakoon and S. Lumyong, sp. nov. (Figure 8).
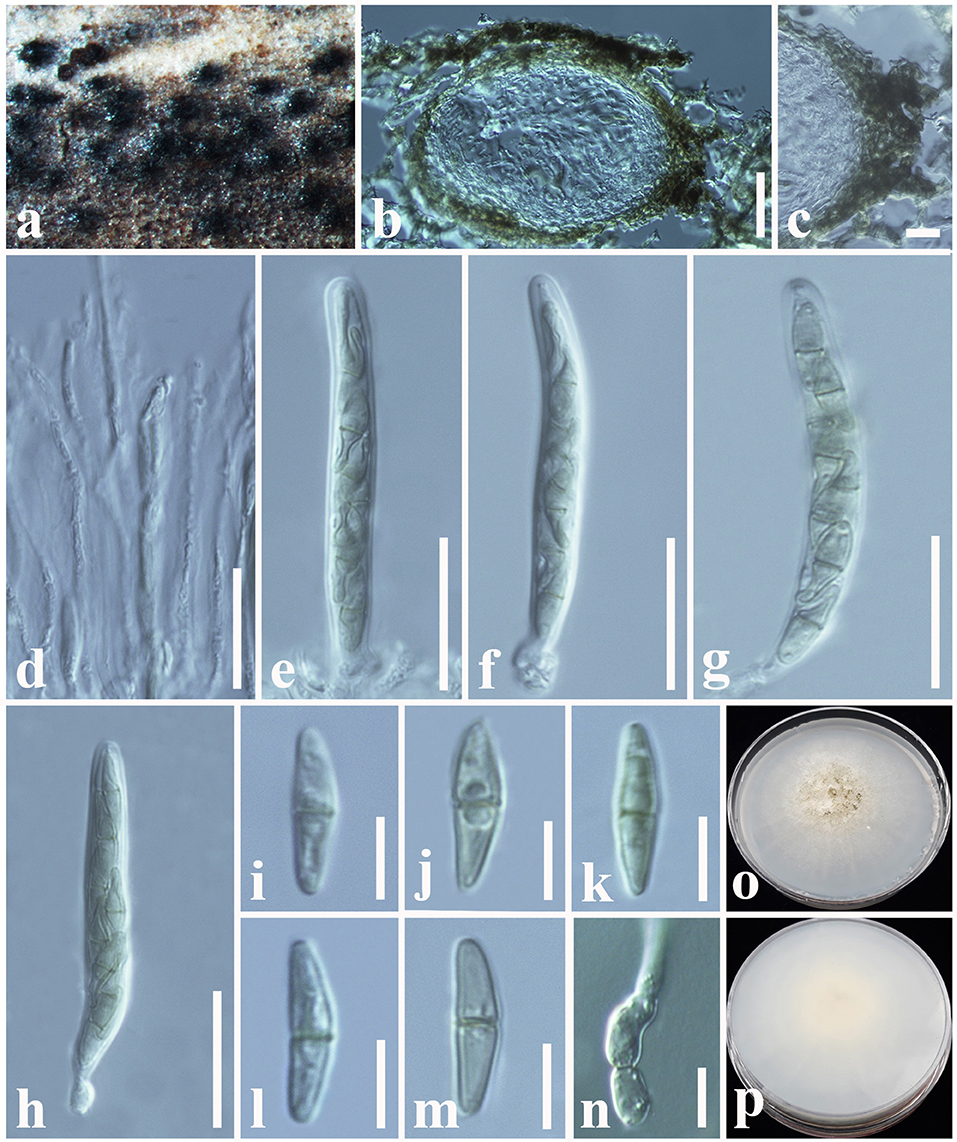
Figure 8. Paraconiothyrium zingiberacearum (NCYU 19-0320, holotype). (a) Ascomata on a dead stem of Hedychium coronarium. (b) Vertical section through an ascoma. (c) Peridium. (d) Pseudoparaphyses. (e–h) Asci. (i–m) Ascospores. (n) Germinating ascospore. (o) Upper side of the culture. (p) Lower side of the culture. Scale bars: (b) = 50 μm, (c) = 10 μm, (d–h) = 20 μm, (i–n) = 6 μm.
Index Fungorum Number: IF559600.
Facesoffungi number: FoF 10802.
Etymology: The species name reflects the host family Zingiberaceae, from which the holotype was collected.
Holotype: NCYU 19-0320.
Saprobic on dead stem of Hedychium coronarium J. Koenig (Zingiberaceae). Sexual morph: Ascomata 100–170 μm high × 120–220 μm diam. ( = 150 × 180 μm, n = 10), solitary or clustered, coriaceous, immersed to semi-immersed, erumpent, globose to sub-globose, unilocular, brown to dark brown. Peridium 20–30 μm wide, outer layer containing light brown to dark brown, thick-walled, tightly packed cells of textura angularis, inner layer containing flattened, hyaline, thin-walled, loosely packed cells of textura angularis. Hamathecium comprising 1.5–3 μm wide, cylindrical to filiform, septate, cellular pseudoparaphyses. Asci 50–60 × 5.5–7 μm ( = 53 × 6.5 μm, n = 20), bitunicate, 8-spored, fissitunicate, clavate to cylindrical, apex rounded, short pedicel, with a shallow ocular chamber. Ascospores 11–14 × 2.5–4 μm ( = 12.5 × 3 μm, n = 30), overlapping, 1–2-seriate, light brown, broadly fusiform with acute ends, 1-septate, slightly curved, rough-walled, guttulate, without a mucilaginous sheath. Asexual morph: Undetermined.
Culture characteristics: Colonies growing on PDA, 10–12 mm diam. after 3 weeks at 25°C, colonies circular, slightly dense, surface smooth, crenate edge, cottony; from above: white to cream at the margin, cream to gray in the center; reverse: white at the margin, light brown to yellowish in the center.
Material examined: China, Taiwan region, Chiayi, on a dead stem of Hedychium coronarium (Zingiberaceae), 05 August 2017, D.S. Tennakoon, NSP002a (NCYU 19-0320, holotype); ex-type living culture, NCYUCC 19-0230. ibid. 08 August 2017, NSP002b (MFLU 18-0091, Paratype); ex-paratype living culture, MFLUCC 18-0559.
Notes: Phylogenetic analyses indicate that our collection (MFLU 18-0091 and NCYU 19-0320) is grouped with Paraconiothyrium estuarinum with high statistical support (89% ML, 0.96 BYPP, Figure 4). The asexual morph of P. estuarinum was introduced by Verkley et al. (2004) from an estuarine sediment polluted with industrial discharges in Brazil. However, we were unable to compare our collection with the asexual morph of P. estuarinum (CBS 109850) because it failed to sporulate even after 2 months of the incubation period (Verkley et al., 2004). Therefore, we compared the base pair differences between our collection and P. estuarinum. There are 10 base pair differences (2.21%) across 451 nucleotides across the tub2 gene region and 11 base pair differences (2.17%) across 506 nucleotides across the ITS (+5.8S) gene region. Phylogeny also indicates that the clade containing our collection and P. estuarinum provides sister lineage to P. cyclothyrioides (Figure 4). The sexual morph of P. cyclothyrioides differs from our collection in having olive-brown, 3-septate, cylindrical to ellipsoidal ascospores with obtusely rounded ends, whereas our collection has light brown, 1-septeate, broadly fusiform ascospores with acute ends (Hyde et al., 2020a). Hence, based on significant differences, we present our collection as a novel species, P. zingiberacearum from dead stems of Hedychium coronarium (Zingiberaceae).
Phylogenetic and taxonomic results of Paraphaeosphaeria
Paraphaeosphaeria O. E. Erikss
Notes: Paraphaeosphaeria was established by Eriksson (1967) to include four species, which have oblong-cylindric ascospores. The sexual morphs of Paraphaeosphaeria have immersed or semi-immersed ascomata, short pedicellate asci, and broadly elliptical, yellowish brown, multi-septate ascospores (Ariyawansa et al., 2014b; Hongsanan et al., 2020). Asexual morphs have pycnidial conidiomata and aseptate or 1-septate conidia (Wanasinghe et al., 2018; Hongsanan et al., 2020). Ariyawansa et al. (2014b) verified the phylogenetic placement of this genus in Didymosphaeriaceae. Currently, 30 species epithets are listed under Paraphaeosphaeria in Index Fungorum (2022). In this study, we introduce a new species (P. brachiariae) from dead leaves of Brachiaria mutica.
Paraphaeosphaeria brachiariae Tennakoon and S. Lumyong, sp. nov. (Figure 9).
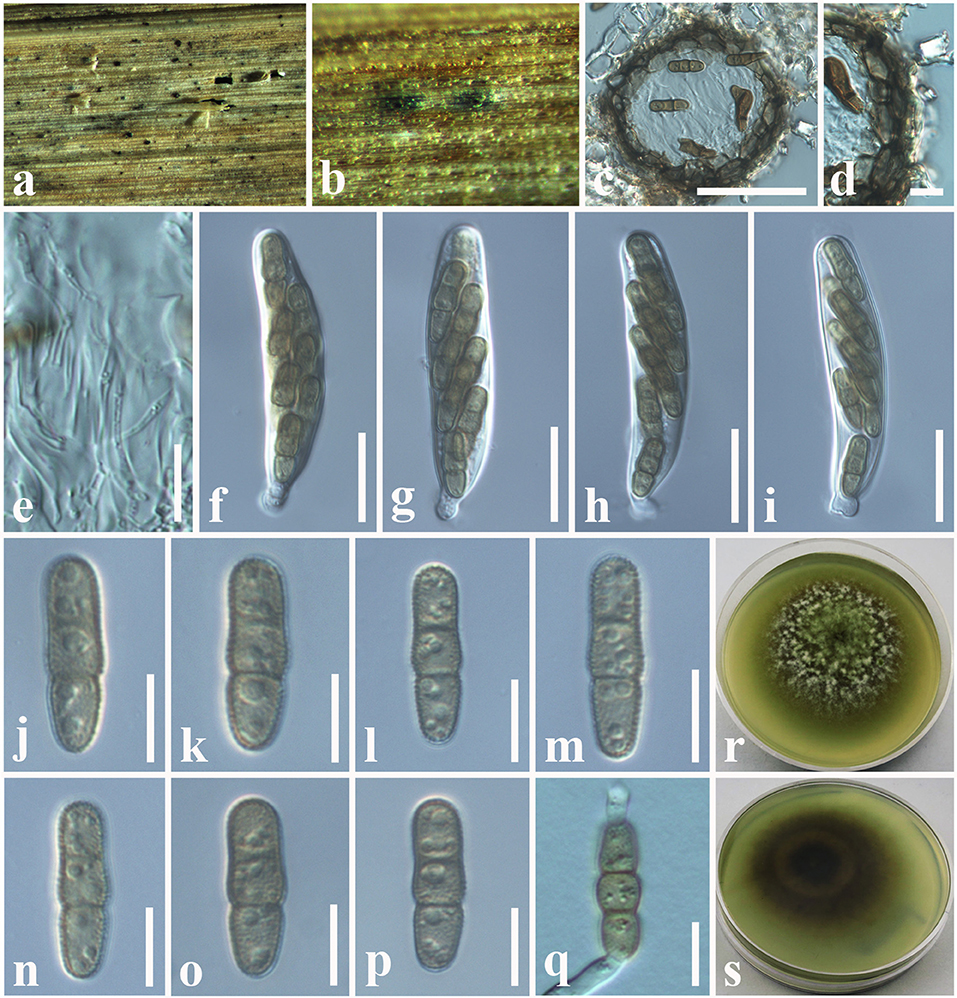
Figure 9. Paraphaeosphaeria brachiariae (NCYU 19-0058, holotype). (a,b) Ascomata on a dead leaf of Brachiaria mutica. (c) Vertical section through an ascoma. (d) Peridium. (e) Pseudoparaphyses. (f–i) Asci. (j–p) Ascospores. (q) Germinating ascospore. (r) Upper side of the culture. (s) Lower side of the culture. Scale bars: (c) = 50 μm, (d) = 10 μm, (e–i) = 20 μm, (j–q) = 7 μm.
Index Fungorum Number: IF559601
Facesoffungi number: FoF 10803
Etymology: Name reflects the host genus, Brachiaria.
Holotype: NCYU 19-0058
Saprobic on dead leaves of Brachiaria mutica (Forssk.) Stapf. (Poaceae). Sexual morph: Ascomata 60–120 μm high × 50–100 μm diam. ( = 75 × 80 μm, n = 10), solitary, scattered, coriaceous, immersed to semi-immersed, partly erumpent, unilocular, globose to sub-globose, brown to dark brown, ostiolate, with minute papilla, filled with hyaline cells. Peridium 10–20 μm wide, comprising light brown to dark brown, thick-walled, loosely packed cells of textura angularis. Hamathecium comprising 1.5–2.5 μm wide, cylindrical to filiform, branched, septate, pseudoparaphyses. Asci 58–65 × 11–13 μm ( = 62 × 12.5 μm, n = 20), bitunicate, 8-spored, fissitunicate, clavate to cylindrical, apex rounded, short pedicellate, with a shallow ocular chamber. Ascospores 14–20 × 4–6 μm ( = 16 × 5 μm, n = 30), overlapping, 1–2-seriate, light brown, fusiform with rounded ends, straight or slightly curved, with 1–2-septate, guttulate, rough-walled, without a mucilaginous sheath. Asexual morph: Undetermined.
Culture characteristics: Colonies growing on PDA, 12–15 mm diam. after 3 weeks at 25°C, colonies circular, smooth surface, crenate edge, cottony; from above: light brown to greenish at the margin, gray in the center; reverse: light brown to yellowish at the margin, brown in the center, mycelium greenish to whitish gray.
Material examined: China, Taiwan region, Chiayi, on dead leaves of Brachiaria mutica (Poaceae), 10 July 2019, D.S. Tennakoon, PC058A (NCYU 19-0058, holotype); ex-type living culture, NCYUCC 19-0342. ibid. 18 July 2019, PC058A (MFLU 19-2799, Paratype); paratype living culture, NCYUCC 19-0343.
Notes: The morphological characteristics of our collection (MFLU 19-2799 and NCYU 19-0058) agree well within the species concept of Paraphaeosphaeria in having immersed or semi-immersed ascomata, short pedicellate asci, and broadly elliptical, multi-septate, yellowish brown ascospores (Ariyawansa et al., 2014b; Hongsanan et al., 2020). Phylogeny (LSU, SSU, ITS, and tub2) shows that our collection (MFLU 19-2799 and NCYU 19-0058) grouped with P. angularis with high statistical support (99% ML, 1.00 BYPP). The asexual morph of P. angularis was introduced by Verkley et al. (2014) from Saccharum officinarum in Brazil. A comparison of 539 nucleotides across the ITS (+5.8S) gene region of P. angularis (CBS 167.70) and our collection (MFLU 19-2799) shows 11 base pair differences (2.04%). We compared 452 nucleotides across the tub2 gene as well, and there were 12 base pair differences (2.65%). Thus, we present our collection as a new species, P. brachiariae from Brachiaria mutica.
Phylogenetic and taxonomic results of Spegazzinia
Spegazzinia Sacc
Notes: Saccardo (1880) established Spegazzinia to include S. ornata as the type species. This genus was previously placed in Apiosporaceae (Sordariomycetes) by Hyde et al. (1998), based on its morphological characteristics. Subsequently, its phylogenetic placement was revealed by Tanaka et al. (2015) and accommodated in Didymosphaeriaceae (Dothideomycetes). Spegazzinia has a diverse distribution worldwide as saprobes on dead materials of vascular plants (Leão-Ferreira and Gusmão, 2010; Manoharachary and Kunwar, 2010; Thambugala et al., 2017; Samarakoon et al., 2020; Tennakoon et al., 2021), endophytes from lichens, and plant leaves (Crous et al., 2019), and even they have been recorded from soil (Ellis, 1971). The conidial morphology and basauxic conidiogenesis of Spegazzinia differentiate it apart from other dematiaceous hyphomycetes. The majority has distinct morphological characteristics in the shapes of α and β conidia (Hongsanan et al., 2020). A total of 14 Spegazzinia species are currently listed in Index Fungorum (2022). In this study, we introduce two new host records of Spegazzinia deightonii and S. tessarthra from Hedychium coronarium and Acacia auriculiformis, respectively.
Spegazzinia deightonii (S. Hughes) Subram., J. Indian Bot. Soc. 35: 78 (1956) (Figure 10).

Figure 10. Spegazzinia deightonii (MFLU 18-2564, new host record). (a–c) Fungal colonies on a dead stem of Hedychium coronarium. (d,e) α Conidia with conidiophores. (f) α Conidium with conidiogenous cell. (g–m) β Conidia. (n) A germinating β conidium. (o) Upper side of the culture. (p) Lower side of the culture. Scale bars: (b,c) = 20 mm, (d–n) = 10 μm.
Index Fungorum number: IF306062.
Facesoffungi number: FoF 07238.
Saprobic on dead stem of Hedychium coronarium J. Koenig (Zingiberaceae). Sexual morph: Undetermined. Asexual morph: Hyphomycetous. Sporodochia 1–2 mm diam., dense, dark, velvety, powdery. Conidiophores long and micronematous. Conidiophores of α conidia up to 50–100 × 1–2 μm ( = 78 × 1.5 μm, n = 20) long, light brown to dark brown, verruculose, narrow, erect or flexuous, unbranched. Conidiogenous cells basauxic, verrucose, pale brown. Conidia holoblastic, two types, α conidia 17–27 × 18–26 μm ( = 24 × 22 μm, n = 25), stellate, solitary, globose to variously shaped, with spines 4–5 μm long, 4–8-celled, frequently 4–6-celled, deeply constricted at the septa. β conidia 15–20 × 10–15 μm ( = 18 × 13 μm, n = 25), hyaline at immature, light brown to dark brown at mature, 8-celled, disc-shaped, both sides slightly flat with short and blunt spines.
Culture characteristics: Colonies growing on PDA, 15–20 mm diam. after 3 weeks at 25°C, colonies circular, slightly dense, surface smooth, velvety; from above: gray to light brown at the margin, dark gray to brown in the center; reverse: yellowish brown at the margin, dark brown to black in the center, mycelium greenish to light brown.
Known hosts: Areca catechu, Cocos nucifera, Hedychium coronarium, Musa sp. and Panicum maximum (Matsushima, 1980; Lu et al., 2000; Tianyu, 2009; Samarakoon et al., 2020; Farr and Rossman, 2022; this study).
Known distribution: China, and Thailand (Matsushima, 1980; Lu et al., 2000; Tianyu, 2009; Samarakoon et al., 2020; Farr and Rossman, 2022; this study).
Material examined: Thailand, Chiang Rai, a dead stem of Hedychium coronarium (Zingiberaceae), 11 July 2018, D.S. Tennakoon, DP017 (MFLU 18-2564); living culture, MFLUCC 18-1625.
Notes: Due to the high similarities of morphological characteristics and phylogenetic affinities, we introduce our collection (MFLU18-2564) as a new host record of S. deightonii from Hedychium coronarium (Zingiberaceae) in Thailand. Phylogenetic analyses of this study indicate that our collection is nested with other S. deightonii isolates in a well-supported clade (92% ML, 0.95 BYPP) (Figure 11). As well as, our collection (MFLU 18-2564) shows a close phylogenetic relationship with the isolate yone 212, which was introduced by Tanaka et al. (2015) from Japan. Both isolates share similar morphological characteristics, such as 8-celled, disked-shaped, dark brown, spiny conidia (Tanaka et al. 2015). Spegazzinia deightonii has previously been recorded from many host species (e.g., Areca catechu, Cocos nucifera, Musa sp., and Panicum maximum), and this is the first host record from Hedychium coronarium (Zingiberaceae) in Thailand.

Figure 11. Phylogram generated from maximum likelihood analysis is based on combined ITS, LSU, SSU, and tef1-α sequence data (Final likelihood value of −8278.609544). The tree is rooted with Laburnicola muriformis (MFLUCC 14-0921, MFLUCC 16-0290). The ex-type strains are indicated in bold, and the new strains are indicated in red. Bootstrap support values ≥65% of maximum likelihood (ML) and Bayesian posterior probabilities (PP) values ≥0.90 are given above the nodes.
Spegazzinia tessarthra (Berk. and M.A. Curtis) Sacc., Syll. Fung. (Abellini) 4: 758 (1886) (Figure 12).

Figure 12. Spegazzinia tessarthra (MFLU 18-2557, new host record). (a–c) Fungal colonies on a dead stem of Acacia auriculiformis. (d–f) α Conidia with conidiophores. (g–o) β Conidia. (p) A germinating β conidium. (q) Upper side of the culture. (r) Lower side of the culture. Scale bars: (b,c) = 2 mm, (d–f) = 30 μm, (g–p) = 6 μm.
Index Fungorum Number: IF219777.
Facesoffungi number: FoF 08241.
= Sporidesmium tessarthrum Berk. and M.A. Curtis,. Soc., Bot. 10(no. 46): 355 (1868).
Saprobic on dead stem of Acacia auriculiformis A. Cunn. (Fabaceae). Sexual morph: Undetermined. Asexual morph: Hyphomycetous. Sporodochia 1–3 mm diam., dense, dark, powdery, velvety. Conidiophores of α conidia up to 30–60 × 1.5–2.5 μm ( = 48 × 1.8 μm, n = 15), erect or flexuous, pale brown to dark brown, rough-walled, narrow and long, generally unbranched. Conidiogenous cells basauxic, verrucose, pale brown. Conidia solitary, two types, α conidia stellate, 15–20 × 14–18 μm ( = 17 × 16 μm, n = 20), globose to sub-globose, 4–6-celled, deeply constricted at the septa, conspicuously spinulate, 4–6 spines, each spine 2–8 μm long. β conidia 10–15 × 8–12 μm ( = 12 × 11 μm, n = 20), disc-shaped, initially hyaline, becoming light brown to dark brown at maturity, 4-celled, rough-walled, crossed septate, smooth-walled, somewhat obovoid, deeply constricted at the septa, dark brown in constricted areas, flat from side view.
Culture characteristics: Colonies growing on PDA, 10–12 mm diam. after 3 weeks at 25°C, colonies circular, dense, smooth surface, entire edge, cottony; from above: light brown to yellowish at the margin, dark brown to black in the center; reverse: yellowish brown at the margin, dark brown to black in the center, mycelium dark brown to dark gray.
Known hosts: Acacia auriculiformis, Ochroma pyramidale and Zea mays (Berkeley and Curtis, 1869; Tanaka et al., 2015; Farr and Rossman, 2022; this study).
Known distribution: Cuba, India, Japan, and Thailand (Berkeley and Curtis, 1869; Tanaka et al., 2015; Farr and Rossman, 2022; this study).
Material examined: Thailand, Ching Rai, a dead stem of Acacia auriculiformis (Fabaceae), 15 July 2018, D.S. Tennakoon, DP003 (MFLU 18-2557); living culture, MFLUCC 18-1624.
Notes: New collection (MFLU 18-2557) resembles Spegazzinia tessarthra (SH 287), which was introduced by Tanaka et al. (2015) from balsa wood in Japan. Both isolates have two types of conidia (α conidia: 4–6-celled, globose to sub-globose, 4–6 spines; β conidia: disc-shaped, 4-celled, light brown to dark brown) (Tanaka et al., 2015). Multi-gene (LSU, SSU, ITS, and tef1-α) phylogenetic analyses also indicate that our collection (MFLU 18-2557) groups with S. tessarthra isolates (NRRL 54913, SH 287) in a well-supported clade (93% ML, 0.99 BYPP, Figure 11). Hence, we consider MFLU 18-2557 as a new host record of S. tessarthra from Acacia auriculiformis (Fabaceae) in Thailand. This species has previously been recorded from Acacia auriculiformis, Ochroma pyramidale, and Zea mays (Berkeley and Curtis, 1869; Tanaka et al., 2015; Farr and Rossman, 2022).
Discussion
Didymosphaeriaceae species has a great variation in morphologies, phylogenies, ecologies, and nutritional modes (Hyde et al., 2013; Ariyawansa et al., 2014b; Liu et al., 2015; Gonçalves et al., 2019; Htet et al., 2021; Suwannarach et al., 2021). It has a diverse range of nutritional modes, such as endophytes, pathogens, and saprobes in plant substrates (Zhang et al., 2012; Liu et al., 2015; Gonçalves et al., 2019; Suwannarach et al., 2021). Researchers have referred this family into several higher taxa due to the uncertainty of the taxonomic placement (von Arx and Müller, 1975; Lumbsch and Huhndorf, 2007; Crous et al., 2012; Zhang et al., 2012). For instance, this family was a synonym of Pleosporaceae (von Arx and Müller, 1975), and Barr (1990) referred Didymosphaeriaceae in Melanommatales. Aptroot (1995) assigned this as a separate family within Pleosporales, and Lumbsch and Huhndorf (2007) treated Didymosphaeriaceae members to the Montagnulaceae in their outline of Ascomycota. However, the confusion surrounding the genera of Didymosphaeriaceae and Montagnulaceae was debated by Ariyawansa et al. (2014a), and they pointed out Didymosphaeriaceae as a distinct family in Pleosporales upon morphology, but phylogenetically, it fits well with Montagnulaceae. Thus, Ariyawansa et al. (2014b) synonymized Montagnulaceae under Didymosphaeriaceae and accepted 16 genera in this family. Over time, numerous genera have been introduced, and currently, 32 genera are accepted in Didymosphaeriaceae (Hongsanan et al., 2020).
Various approaches have been used to identify Didymosphaeriaceae species over the years. Earlier, morphology-based species recognition was the key method for identifying Didymosphaeriaceae species, and most studies have been used drawings with descriptions (von Arx and Müller, 1975; Sutton, 1980; Barr, 1990). However, morphology-based species identification suffered various issues, including phenotypic plasticity, which may lead to countless misinterpretations. However, by using molecular techniques for species delineation, identification, and taxonomic classifications, this fungal taxonomy undergone a revolution (Ariyawansa et al., 2014a; Das et al., 2014; Chethana et al., 2020). Therefore, most of recent studies have integrated morphology and phylogeny data for Didymosphaeriaceae species identification, taxonomic classification, and phylogenetic inferences (Ariyawansa et al., 2014a,b; Wanasinghe et al., 2016; Mapook et al., 2020; Htet et al., 2021; Suwannarach et al., 2021). Apart from morphology and phylogeny classifications, some researchers have been carried out to estimate the divergence time of this family using molecular clock analyses (Khodaei et al., 2019), and some have investigated the secondary metabolites or biological functions of Didymosphaeriaceae species (Wang et al., 2021).
The phylogeny recovered in this study agrees well with previous Didymosphaeriaceae studies within Pleosporales (Ariyawansa et al., 2014b; Wanasinghe et al., 2018; Phookamsak et al., 2019; Mapook et al., 2020; Phukhamsakda et al., 2020). The morphological characteristics of the novel species (Montagnula acaciae, Paraconiothyrium zingiberacearum, and Paraphaeosphaeria brachiariae) fit well with the respective genera, are phylogenetically distinct from other species, and group with high statistical evidence (Figures 2, 4, 11, 13). Also, the morphology of the new host records (Montagnula jonesii, Paraconiothyrium archidendri, P. brasiliense, P. fuckelii, Spegazzinia deightonii, and S. tessarthra) also agrees with their respective type species (Saccardo, 1876; Ariyawansa et al., 2014a,b; Tanaka et al., 2015; Tennakoon et al., 2016). Overall, this work offers fascinating taxonomic insights into Didymosphaeriaceae species. In particular, the findings provide evidence to show the vast range of fungal diversity in litter substrates (e.g., dead leaves and dead stems), even within a single family, Didymosphaeriaceae. But, to determine whether these fungal species are generalists or specialists, more ecological studies are required. Furthermore, it is worthy to note that several Didymosphaeriaceae genera still lack molecular data to determine the phylogenetic placement (e.g., Barria and Lineostroma). Therefore, it is necessary to collect, isolate, and retrieve sequence data for more Didymosphaeriaceae species in future studies.

Figure 13. Phylogram generated from maximum likelihood analysis is based on combined ITS, LSU, SSU, and tub2 sequence data (final likelihood value of −9171.108559). The tree is rooted with Paraconiothyrium fuckelii (JZB320001, JZB320002, and JZB320003). The ex-type strains are indicated in bold, and the new isolates are indicated in red. Bootstrap support values ≥65% of maximum likelihood (ML) and Bayesian posterior probabilities (PP) values ≥0.90 are given above the nodes.
Conclusion
Studies on fungal species in plant litter are particularly pertinent in the current climate change scenario since climate plays a crucial role in decomposition and, ultimately, nutrient cycling, soil fertility, and the global carbon cycle. This study revealed plant litter inhabiting three new fungal species (Montagnula acaciae, Paraconiothyrium zingiberacearum, and Paraphaeosphaeria brachiariae) and six new host records (Montagnula jonesii, Paraconiothyrium archidendri, P. brasiliense, P. fuckelii, Spegazzinia deightonii, and S. tessarthra) from China and Thailand. In addition, Paraconiothyrium rosae was synonymized under P. fuckelii based on morphology and phylogeny data. We believe that this study sheds light on the plant litter-dwelling fungal communities, which are diverse and astonishing.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Author contributions
DT, KT, and NIS: conceptualization, methodology, formal analysis, and writing—original draft preparation. DT and NIS: software and data curation. KT and NS: validation. KT and SL: investigation. DT: resources. KT, NIS, NS, and SL: writing—review and editing. NS and SL: supervision and funding acquisition. SL: project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Post-Doctoral Fellowship 2022 for Reinventing Chiang Mai University (R000030885). This project was also funded by the National Research Council of Thailand (NRCT) N42A650198.
Acknowledgments
DT would like to thank Chang-Hsin Kuo and department of plant medicine, National Chiayi University (NCYU) for lab facilities. The authors thank Shaun Pennycook for checking species names. DT, NS, and SL thank Chiang Mai University, Thailand, for partial support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer SCK is currently organizing a Research Topic with the author NS.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aptroot, A. (2004). Two new Ascomycetes with long gelatinous appendages collected from monocots in the tropics. Stud. Mycol. 50, 307–311.
Ariyawansa, H. A., Camporesi, E., Thambugala, K. M., Mapook, A., Kang, J. C., Alias, S. A., et al. (2014a). Confusion surrounding Didymosphaeria—phylogenetic and morphological evidence suggest Didymosphaeriaceae is not a distinct family. Phytotaxa 176, 102–119. doi: 10.11646/phytotaxa.176.1.12
Ariyawansa, H. A., Maharachchikumbura, S. S., Karunarathna, S. C., Chukeatirote, E., Bahkali, A. H., Kang, J. K., et al. (2013). Deniquelata barringtoniae gen. et sp. nov., associated with leaf spots of Barringtonia asiatica. Phytotaxa 105, 11–20. doi: 10.11646/phytotaxa.105.1.2
Ariyawansa, H. A., Tanaka, K., Thambugala, K. M., Phookamsak, R., Tian, Q., Camporesi, E., et al. (2014b). A molecular phylogenetic reappraisal of the Didymosphaeriaceae (= Montagnulaceae). Fungal Divers. 68, 69–104. doi: 10.1007/s13225-014-0305-6
Ariyawansa, H. A., Tsai, I., Thambugala, K. M., Chuang, W. Y., Lin, S. R., Hozzein, W. N., et al. (2020). Species diversity of Pleosporalean taxa associated with Camellia sinensis (L.) Kuntze in Taiwan. Sci. Rep. 10, 1–20. doi: 10.1038/s41598-020-69718-0
Barelli, L., Moonjely, S., Behie, S. W., and Bidochka, M. J. (2016). Fungi with multifunctional lifestyles: endophytic insect pathogenic fungi. Plant Mol. Biol. 90, 657–664. doi: 10.1007/s11103-015-0413-z
Berkeley, M. J., and Curtis, M. A. (1869). On a collection of Fungi from Cuba. Part. II., including those belonging to the families gasteromycetes, coniomycetes, hyphomycetes, physomycetes, and ascomycetes. J. Linn. Soc.10, 341–392. doi: 10.1111/j.1095-8339.1868.tb00648a.x
Błaszkowski, J., Chwat, G., Kovács, G. M., Gáspár, B. K., Ryszka, P., Orłowska, E., et al. (2013). Septoglomus fuscum and S. furcatum, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycologia 105, 670–680. 10.3852/12-127
Boonmee, S., Wanasinghe, D. N., Calabon, M. S., Huanraluek, N., Chandrasiri, S. K., Jones, G. E., et al. (2021). Fungal diversity notes 1387–1511: taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers 111, 1–335. doi: 10.1007/s13225-021-00489-3
Budziszewska, J., Szypuła, W., Wilk, M., and Wrzosek, M. (2011). Paraconiothyrium babiogorense sp. nov., a new endophyte from fir club moss Huperzia selago (Huperziaceae). Mycotaxon 115, 457–468. doi: 10.5248/115.457
Chethana, K. W. T., Jayawardena, R. S., and Hyde, K. D. (2020). Hurdles in fungal taxonomy: effectiveness of recent methods in discriminating taxa. Megataxa 1, 114–122. doi: 10.11646/megataxa.1.2.2
Conforto, C., Lima, N. B., Silva, F. J. A., Câmara, M. P. S., Maharachchikumbura, S., and Michereff, S. J. (2019). Characterization of fungal species associated with cladode brown spot on Nopalea cochenillifera in Brazil. Eur. J. Plant Pathol. 155, 1179–1194. doi: 10.1007/s10658-019-01847-3
Crous, P. W., Carnegie, A. J., Wingfield, M. J., Sharma, R., Mughini, G., Noordeloos, M. E., et al. (2019). Fungal Planet description sheets: 868–950. Persoonia 42, 291–473. doi: 10.3767/persoonia.2019.42.11
Crous, P. W., Summerell, B. A., Shivas, R. G., Burgess, T. I., Decock, C. A., Dreyer, L. L., et al. (2012). Fungal planet description sheets: 107–127. Persoonia 28, 138–182. doi: 10.3767/003158512X652633
Crous, P. W., Wingfield, M. J., Burgess, T. I., Hardy, G. S. J., Barber, P. A., Alvarado, P., et al. (2017). Fungal planet description sheets: 558–624. Persoonia 38, 240–384. doi: 10.3767/003158517X698941
Csata, E., Czekes, Z., Eros, K., Német, E., Hughes, M., Csosz, S., et al. (2013). Comprehensive survey of Romanian myrmecoparasitic fungi: new species, biology and distribution. North-West J. Zool. 9, 23–29.
Damm, U., Verkley, G. J. M., Crous, P. W., Fourie, P. H., Haegi, A., and Riccioni, L. (2008). Novel Paraconiothyrium species on stone fruit trees and other woody hosts. Persoonia 20, 9–17. doi: 10.3767/003158508X286842
Das, S., Dash, H. R., Mangwani, N., Chakraborty, J., and Kumari, S. (2014). Understanding molecular identification and polyphasic taxonomic approaches for genetic relatedness and phylogenetic relationships of microorganisms. J. Microbiol. Methods 103, 80–100. doi: 10.1016/j.mimet.2014.05.013
De Gruyter, J., Woudenberg, J. H. C., Aveskamp, M. M., Verkley, G. J. M., Groenewald, J. Z., and Crous, P. W. (2006). Redisposition of Phoma-like anamorphs in Pleosporales. Stud. Mycol. 75, 1–36. doi: 10.3114/sim0004
Drinkwater, L. E., Schipanski, M., Snapp, S. S., and Jackson, L. E. (2008). “Ecologically based nutrient management,” in Agricultural Systems: Agroecology and Rural Innovation for Development, eds Snapp S and Pound B (Boston: Elsevier/Academic Press), 159–208.
Du, T., Hyde, K. D., Mapook, A., Mortimer, P. E., Xu, J., Karunarathna, S. C., et al. (2021). Morphology and phylogenetic analyses reveal Montagnula puerensis sp. nov. (Didymosphaeriaceae, Pleosporales) from southwest China. Phytotaxa 514, 1–25. doi: 10.11646/phytotaxa.514.1.1
Eriksson, O. E. (1967). On graminicolous Pyrenomycetes from Fennoscandia. I, II, III. Ark Bot. Ser. 26, 339–466.
Farr, D. F., and Rossman, A. Y. (2022). Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. Available from: http://nt.ars-grin.gov/fungaldatabases (accessed March 1, 2022).
Field, K. J., and Pressel, S. (2018). Unity in diversity: structural and functional insights into the ancient partnerships between plants and fungi. New Phytol. 220, 996–1011. doi: 10.1111/nph.15158
Gafforov, Y. S. (2017). A preliminary checklist of Ascomycetous microfungi from Southern Uzbekistan. Mycosphere 8, 660–696. doi: 10.5943/mycosphere/8/4/12
Giraldo, A., Hernández-Restrepo, M., and Crous, P. W. (2019). New plectosphaerellaceous species from Dutch garden soil. Mycol. Prog. 18, 1135–1154. doi: 10.1007/s11557-019-01511-4
Glass, N. L., and Donaldson, G. C. (1995). Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 61, 1323–1330. doi: 10.1128/aem.61.4.1323-1330.1995
Goh, J., Mun, H. Y., Oh, Y., and Chung, N. (2016). Four species of Montagnulaceae unrecorded in Korea and isolated from plant litter in freshwater. Kor. J. Mycol. 44, 263–270. doi: 10.4489/KJM.2016.44.4.263
Gonçalves, M. F., Vicente, T. F., Esteves, A. C., and Alves, A. (2019). Neptunomyces aureus gen. et sp. nov. (Didymosphaeriaceae, Pleosporales) isolated from algae in Ria de Aveiro, Portugal. MycoKeys 60, 31–44. doi: 10.3897/mycokeys.60.37931
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95–98.
Hawksworth, D. L. (2001). The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol. Res. 105, 1422–1432. doi: 10.1017/S0953756201004725
Hernandez-Restrepo, M., Gené, J., Castañeda-Ruiz, R. F., Mena-Portales, J., Crous, P. W., and Guarro, J. (2017). Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 86, 53–97. doi: 10.1016/j.simyco.2017.05.002
Hongsanan, S., Hyde, K. D., Bahkali, A. H., Camporesi, E., Chomnunti, P., Ekanayaka, H., et al. (2015). Fungal biodiversity profiles 11–20. Cryptogam. Mycol. 36, 355–380. doi: 10.7872/crym/v36.iss3.2015.355
Hongsanan, S., Hyde, K. D., Phookamsak, R., Wanasinghe, D. N., McKenzie, E. H. C., Sarma, V. V., et al. (2020). Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 11, 1553–2107. doi: 10.5943/mycosphere/11/1/13
Htet, Z. H., Mapook, A., Gafforov, Y., Chethana, K. T., Lumyoung, S., and Hyde, K. D. (2021). Molecular phylogeny and diversity of Laburnicola (Didymosphaeriaceae): a new species from Uzbekistan. Phytotaxa 527, 177–190. doi: 10.11646/phytotaxa.527.3.2
Huelsenbeck, J. P., and Ronquist, F. (2001). MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17, 754–755. doi: 10.1093/bioinformatics/17.8.754
Hyde, K. D., Chaiwan, N., Norphanphoun, C., Boonmee, S., Camporesi, E., Chethana, K. W., et al. (2018a). Mycosphere notes 169–224. Mycosphere 9, 271–430. doi: 10.5943/mycosphere/9/2/8
Hyde, K. D., de Silva, N. I., Jeewon, R., Bhat, D. J., Phookamsak, R., Doilom, M., et al. (2020a). AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 3, 22–294. doi: 10.5943/ajom/3/1/3
Hyde, K. D., Dong, Y., Phookamsak, R., Jeewon, R., Bhat, D. J., Jones, E. B., et al. (2020b). Fungal diversity notes 1151–1276: taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 100, 1–273. doi: 10.1007/s13225-020-00439-5
Hyde, K. D., Fröhlich, J., and Taylor, J. E. (1998). Fungi from palms. XXXVI. Reflections on unitunicate Ascomycetes with apiospores. Sydowia 50, 21–80.
Hyde, K. D., Hongsanan, S., Jeewon, R., Bhat, D. J., McKenzie, E. H. C., Jones, E. B. G., et al. (2016). Fungal diversity notes 367–490: taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 80, 1–270. doi: 10.1007/s13225-016-0373-x
Hyde, K. D., Jones, E. B., Liu, J. K., Ariyawansa, H., Boehm, E., Boonmee, S., et al. (2013). Families of Dothideomycetes. Fungal Divers. 63, 1–313. doi: 10.1007/s13225-013-0263-4
Hyde, K. D., Norphanphoun, C., Chen, J., Dissanayake, A. J., Doilom, M., Hongsanan, S., et al. (2018b). Thailand's amazing diversity: up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 93, 215–239. doi: 10.1007/s13225-018-0415-7
Index Fungorum (2022). Index Fungorum. Available from: http://www.indexfungorum.org/names/Names.asp (accessed May 6, 2022).
Jamali, S. A. M. A. D. (2020). First report of Paraconiothyrium fuckelii (Didymosphaeriaceae, Pleosporales), causing stem canker in Rosa hybrida, from Iran. Czech Mycol. 72, 71–82. doi: 10.33585/cmy.72105
Jayasiri, S. C., Hyde, K. D., Ariyawansa, H. A., Bhat, J., Buyck, B., Cai, L., et al. (2015). The Faces of Fungi database: fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 74, 3–18. doi: 10.1007/s13225-015-0351-8
Jayasiri, S. C., Hyde, K. D., Jones, E. B. G., McKenzie, E. H. C., Jeewon, R., Phillips, A. J. L., et al. (2019). Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 10, 1–186. doi: 10.5943/mycosphere/10/1/1
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Khodaei, S., Arzanlou, M., Babai-Ahari, A., Rota-Stabelli, O., and Pertot, I. (2019). Phylogeny and evolution of Didymosphaeriaceae (Pleosporales): new Iranian samples and hosts, first divergence estimates, and multiple evidences of species mis-identifications. Phytotaxa 424, 131–146. doi: 10.11646/phytotaxa.424.3.1
Kowalski, T., and Andruch, K. (2012). Mycobiota in needles of Abies alba with and without symptoms of Herpotrichia needle browning. For. Pathol. 42, 183–190. doi: 10.1111/j.1439-0329.2011.00738.x
Kuter, G. A. (1986). Microfungal populations associated with the decomposition of sugar maple leaf litter. Mycologia 78, 114–126. doi: 10.1080/00275514.1986.12025213
Leão-Ferreira, S. M., and Gusmão, L. F. P. (2010). Conidial fungi from the semi-arid Caatinga biome of Brazil. New species of Endophragmiella and Spegazzinia with new records for Brazil, South America, and Neotropica. Mycotaxon 111, 1–10. doi: 10.5248/111.1
Li, P., Li, Y., Zheng, X., Ding, L., Ming, F., Pan, A., et al. (2018). Rice straw decomposition affects diversity and dynamics of soil fungal community, but not bacteria. J. Soils Sedim. 18, 248–258. doi: 10.1007/s11368-017-1749-6
Liu, J. K., Hyde, K. D., Jones, E. B., Ariyawansa, H. A., Bhat, D. J., Boonmee, S., et al. (2015). Fungal diversity notes 1–110: taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 72, 1–197. doi: 10.1007/s13225-015-0324-y
Lu, B., Hyde, K. D., Ho, W. H., Tsui, K. M., Taylor, J. E., Wong, K. M., et al. (2000). Checklist of Hong Kong Fungi. Hong Kong: Fungal Diversity Research Series.
Lumbsch, H. T., Ahti, T., Altermann, S., De Paz, G. A., Aptroot, A., Arup, U., et al. (2011). One hundred new species of lichenized fungi: a signature of undiscovered global diversity. Phytotaxa 18, 1–127. doi: 10.11646/phytotaxa.18.1.1
Manoharachary, C., and Kunwar, I. K. (2010). Spegazzinia Species from India. Taxonomy and Ecology of Indian Fungi. New Delhi: IK Internat Pvt Ltd, 13–18.
Mapook, A., Hyde, K. D., McKenzie, E. H., Jones, E. B., Bhat, D. J., Jeewon, R., et al. (2020). Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 101, 1–175. doi: 10.1007/s13225-020-00444-8
Matsushima, T. (1980). Matsushima Mycological Memoirs No. 1. Saprophytic Microfungi from Taiwan, Part 1. Hyphomycetes. Kobe: Matsushima Fungus Collection, 82.
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES science gateway for inference of large phylogenetic trees,” in Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 1–8. doi: 10.1109/GCE.2010.5676129
Mułenko, W., Majewski, T., and Ruszkiewicz-Michalska, M. (2008). A Preliminary Checklist of Micromycetes in Poland. Krakow: W. Szafer Institute of Botany, Polish Academy of Sciences.
Munk, A. (1953). The System of the Pyrenomycetes: A Contribution to a Natural Classification of the Group Sphaeriales Sensu Lindau: Repr. for Dansk Botanisk Arkiv as Bd. 15, Nr. 2 (Doctoral dissertation).
Nakashima, K. I., Tomida, J., Hirai, T., Kawamura, Y., and Inoue, M. (2019). Paraconiothins A–J: sesquiterpenoids from the endophytic fungus Paraconiothyrium brasiliense ECN258. J. Nat. Prod. 82, 3347–3356. doi: 10.1021/acs.jnatprod.9b00638
Osono, T. (2007). Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 22, 955–974. doi: 10.1007/s11284-007-0390-z
Osono, T. (2017). Leaf litter decomposition of 12 tree species in a subtropical forest in Japan. Ecol. Res. 32, 413–422. doi: 10.1007/s11284-017-1449-0
Paul, N. C., and Lee, H. B. (2014). First record of endophytic Paraconiothyrium brasiliense isolated from Chinese maple leaves in Korea. Kor. J. Mycol. 42, 349–352. doi: 10.4489/KJM.2014.42.4.349
Phookamsak, R., Hyde, K. D., Jeewon, R., Bhat, D. J., Jones, E. B., Maharachchikumbura, S. S., et al. (2019). Fungal diversity notes 929–1035: taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 95, 1–273. doi: 10.1007/s13225-019-00421-w
Phukhamsakda, C., McKenzie, E. H., Phillips, A. J., Jones, E. B. G., Bhat, D. J., Stadler, M., et al. (2020). Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 102, 1–203. doi: 10.1007/s13225-020-00448-4
Pitt, W. M., Úrbez-Torres, J. R., and Trouillas, F. P. (2014). Munkovalsaria donacina from grapevines and Desert Ash in Australia. Mycosphere 5, 656–661. doi: 10.5943/mycosphere/5/5/6
Posada, D., and Crandall, K. A. (1998). MODELTEST: testing the model of DNA substitution. Bioinformatics 14, 817–818. doi: 10.1093/bioinformatics/14.9.817
Promputtha, I., McKenzie, E. H., Tennakoon, D. S., Lumyong, S., and Hyde, K. D. (2017). Succession and natural occurrence of saprobic fungi on leaves of Magnolia liliifera in a tropical forest. Cryptogam. Mycol. 38, 213–225. doi: 10.7872/crym/v38.iss2.2017.213
Purahong, W., Pietsch, K. A., Lentendu, G., Schöps, R., Bruelheide, H., Wirth, C., et al. (2017). Characterization of unexplored deadwood mycobiome in highly diverse subtropical forests using culture-independent molecular technique. Front. Microbiol. 8, 574. doi: 10.3389/fmicb.2017.00574
Rambaut, A. (2012). FigTree Version 1.4.0. Edinburgh: Institute of Evolutionary Biology; University of Edinburgh. Available online at: http://tree.bio.ed.ac.uk/software/figtree/ (accessed May 10, 2022).
Rannala, B., and Yang, Z. (1996). Probability distribution of molecular evolutionary trees: a new method of phylogenetic inference. J. Mol. Evol. 43, 304–311. doi: 10.1007/BF02338839
Rehner, S. (2001). Primers for Elongation Factor 1-α (EF1-α). Available online at: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf (accessed June 16, 2022).
Rehner, S. A., and Samuels, G. J. (1994). Taxonomy and phylogeny of Gliocladium analyzed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 98, 625–634. doi: 10.1016/S0953-7562(09)80409-7
Romaní, A. M., Fischer, H., Mille-Lindblom, C., and Tranvik, L. J. (2006). Interactions of bacteria and fungi on decomposing litter: differential extracellular enzyme activities. Ecology 87, 2559–2569. doi: 10.1890/0012-9658(2006)872559:IOBAFO2.0.CO
Saccardo, P. A. (1876). Fungi Veneti novi vel critici. Series V. Nuovo Giornale Botanico Italiano 8,161–211.
Samarakoon, B. C., Wanasinghe, D. N., Samarakoon, M. C., Phookamsak, R., McKenzie, E. H., Chomnunti, P., et al. (2020). Multi-gene phylogenetic evidence suggests Dictyoarthrinium belongs in Didymosphaeriaceae (Pleosporales, Dothideomycetes) and Dictyoarthrinium musae sp. nov. on Musa from Thailand. MycoKeys 71, 101–118. doi: 10.3897/mycokeys.71.55493
Senanayake, I. C., Rathnayaka, A. R., Marasinghe, D. S., Calabon, M. S., Gentekaki, E., Lee, H. B., et al. (2020). Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 11, 2678–2754. doi: 10.5943/mycosphere/11/1/20
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Stamatakis, A., Hoover, P., and Rougemont, J. (2008). A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 57, 758–771. doi: 10.1080/10635150802429642
Sutton, B. C. (1980). The Coelomycetes. Fungi Imperfecti with Pycnidia, Acervuli and Stromata. Kew: Commonwealth Mycological Institute, 1–696.
Suwannarach, N., Kumla, J., and Lumyong, S. (2021). Spegazzinia camelliae sp. nov. (Didymosphaeriaceae, Pleosprales), a new endophytic fungus from northern Thailand. Phytotaxa 483, 117–128. doi: 10.11646/phytotaxa.483.2.4
Tanaka, K., Hirayama, K., Yonezawa, H., Sato, G., Toriyabe, A., Kudo, H., et al. (2015). Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 82, 75–136. doi: 10.1016/j.simyco.2015.10.002
Tang, C., Xu, Q., Zhao, M., Wang, X., and Kang, Z. (2018). Understanding the lifestyles and pathogenicity mechanisms of obligate biotrophic fungi in wheat: the emerging genomics era. Crop J. 6, 60–67. doi: 10.1016/j.cj.2017.11.003
Tennakoon, D. S., Hyde, K. D., Wanasinghe, D. N., Bahkali, A. H., Camporesi, E., Khan, S., et al. (2016). Taxonomy and phylogenetic appraisal of Montagnula jonesii sp. nov. (Didymosphaeriaceae, Pleosporales). Mycosphere 7, 1346–1356. doi: 10.5943/mycosphere/7/9/8
Tennakoon, D. S., Kuo, C. H., Maharachchikumbura, S. S., Thambugala, K. M., Gentekaki, E., Phillips, A. J., et al. (2021). Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 108, 1–215. doi: 10.1007/s13225-021-00474-w
Tennakoon, D. S., Thambugala, K. M., Wanasinghe, D. N., Gentekaki, E., Promputtha, I., Kuo, C. H., et al. (2020). Additions to Phaeosphaeriaceae (Pleosporales): Elongaticollum gen. nov., Ophiosphaerella taiwanensis sp. nov., Phaeosphaeriopsis beaucarneae sp. nov. and a new host record of Neosetophoma poaceicola from Musaceae. MycoKeys 70, 59–88. doi: 10.3897/mycokeys.70.53674
Thambugala, K. M., Wanasinghe, D. N., Phillips, A. J. L., Camporesi, E., Bulgakov, T. S., Phukhamsakda, C., et al. (2017). Mycosphere notes 1–50: grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 8, 697–796. doi: 10.5943/mycosphere/8/4/13
Tianyu, Z. (2009). Flora Fungorum Sincorum (Vol 31): 26 Genera of Dematiaceous Dictyosporous Hyphomycetes Excluding Alternaria. Beijing: Science Press.
Tibpromma, S., Hyde, K. D., McKenzie, E. H., Bhat, D. J., Phillips, A. J., Wanasinghe, D. N., et al. (2018). Fungal diversity notes 840–928: micro-fungi associated with Pandanaceae. Fungal Divers. 93, 1–160. doi: 10.1007/s13225-018-0408-6
Verkley, G. J., da Silva, M., Wicklow, D. T., and Crous, P. W. (2004). Paraconiothyrium, a new genus to accommodate the mycoparasite Coniothyrium minitans, anamorphs of Paraphaeosphaeria, and four new species. Stud. Mycol. 50, 323–335.
Verkley, G. J. M., Dukik, K., Renfurm, R., Göker, M., and Stielow, J. B. (2014). Novel genera and species of coniothyrium-like fungi in Montagnulaceae (Ascomycota). Persoonia 32, 25–51. doi: 10.3767/003158514X679191
Vilgalys, R., and Hester, M. (1990). Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 172, 4238–4246. doi: 10.1128/jb.172.8.4238-4246.1990
von Arx, J. A., and Müller, E. (1975). A re-evaluation of the bitunicate Ascomycetes with keys to families and genera. Stud. Mycol. 9, 1–159.
Wanasinghe, D. N., Jones, E. B. G., Camporesi, E., Dissanayake, A. J., Kamolhan, S., Mortimer, P. E., et al. (2016). Taxonomy and phylogeny of Laburnicola gen. nov. and Paramassariosphaeria gen. nov. (Didymosphaeriaceae, Massarineae, Pleosporales). Fungal Biol. 120, 1354–1373. doi: 10.1016/j.funbio.2016.06.006
Wanasinghe, D. N., Phukhamsakda, C., Hyde, K. D., Jeewon, R., Lee, H. B., Jones, E. B. G., et al. (2018). Fungal diversity notes 709–839: taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 89, 1–236. doi: 10.1007/s13225-018-0395-7
Wang, J., Shao, S., Liu, C., Song, Z., Liu, S., and Wu, S. (2021). The genus Paraconiothyrium: Species concepts, biological functions, and secondary metabolites. Crit. Rev. Microbiol. 47, 781–810. doi: 10.1080/1040841X.2021.1933898
White, T. J., Bruns, T., Lee, S. J. W. T., and Taylor, J. (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 18, 315–322.
Zhang, N., Li, Y., Wubet, T., Bruelheide, H., Liang, Y., Purahong, W., et al. (2018). Tree species richness and fungi in freshly fallen leaf litter: unique patterns of fungal species composition and their implications for enzymatic decomposition. Soil Biol. Biochem. 127, 120–126. doi: 10.1016/j.soilbio.2018.09.023
Zhang, Y., Crous, P. W., Schoch, C. L., and Hyde, K. D. (2012). Pleosporales. Fungal Divers. 52, 1–225. doi: 10.1007/s13225-011-0117-x
Keywords: 3 new species, China, molecular phylogeny, morphology, new host records, saprobic fungi, taxonomy, Thailand
Citation: Tennakoon DS, Thambugala KM, Silva NId, Suwannarach N and Lumyong S (2022) A taxonomic assessment of novel and remarkable fungal species in Didymosphaeriaceae (Pleosporales, Dothideomycetes) from plant litter. Front. Microbiol. 13:1016285. doi: 10.3389/fmicb.2022.1016285
Received: 11 August 2022; Accepted: 11 October 2022;
Published: 22 November 2022.
Edited by:
Jesús Navas-Castillo, La Mayora Experimental Station (CSIC), SpainReviewed by:
ZongLong Luo, Dali University, ChinaSamantha Chandranath Karunarathna, Qujing Normal University, China
Yusufjon Gafforov, Academy of Science of the Republic of Uzbekistan, Uzbekistan
Sinang Hongsanan, Shenzhen University, China
Copyright © 2022 Tennakoon, Thambugala, Silva, Suwannarach and Lumyong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Saisamorn Lumyong, scboi009@gmail.com