 Elīna Brauna-Morževska1*
Elīna Brauna-Morževska1* Frederick L. Stoddard2
Frederick L. Stoddard2 Biruta Bankina1Jānis Kaņeps1Gunita Bimšteine1Irina Petrova1Ingrīda Neusa-Luca1
Biruta Bankina1Jānis Kaņeps1Gunita Bimšteine1Irina Petrova1Ingrīda Neusa-Luca1 Ance Roga3
Ance Roga3 Dāvids Fridmanis3
Dāvids Fridmanis3- 1Institute of Soil and Plant Sciences, Latvia University of Life Sciences and Technologies, Jelgava, Latvia
- 2Viikki Plant Science Centre, and Helsinki Sustainability Science Centre, Department of Agricultural Sciences, University of Helsinki, Helsinki, Finland
- 3Human Genetics and Disease Mechanisms, Latvian Biomedical Research and Study Centre, Riga, Latvia
Fungi of genus Botrytis are important pathogens of legumes, causing gray mold and chocolate spot diseases. The use of molecular methods to identify pathogens has resulted in the discovery of several new Botrytis species and new associations of pathogens with diseases. Thus, chocolate spot of faba bean is now associated with at least four species: B. fabae, B. cinerea, B. pseudocinerea and B. fabiopsis. Species of Botrytis differ in host plant, pathogenicity, fungicide resistance and other relevant properties that affect disease control. The aim of this study was to identify the species of Botrytis isolated from different legume crops and to evaluate their in vitro pathogenicity. Between 2014 and 2019, 278 isolates of Botrytis were obtained from infected legumes in Latvia. A phylogenetic analysis was carried out by sequencing three nuclear genes, RPB2, HSP60, and G3PDH, considered to be diagnostic for species in this genus. A set of 21 representative isolates was selected for pathogenicity tests on detached leaves of faba bean, field pea, lupin and soybean using 5-mm mycelium-agar plugs. The diameter of the formed lesions under the inoculated plug was measured crosswise each day. The datasets were subjected to analysis of variance with the split-plot design of the experiment and repeated-measures model. Six species were identified: B. cinerea, B. fabae, B. pseudocinerea, B. fabiopsis, B. euroamericana and B. medusae. In addition to the expected combinations of host and pathogen, naturally occurring infections of B. fabiopsis were found on chickpea, B. euroamericana on faba bean and B. medusae in lupin seeds. Species and isolate had significant effects on pathogenicity on all crops tested. Several isolates were pathogenic on two or more host species: two of B. pseudocinerea, two of B. cinerea, two of B. fabiopsis and the one of B. medusae. One isolate of B. pseudocinerea and two of B. fabiopsis caused primary lesions on all five host species. The results show that these Botrytis species have a broad host range that should be borne in mind when planning crop sequences and rotations.
1 Introduction
Legume crops provide various benefits for cropping systems and are regaining an important role in European agriculture. Between 2009-2014 and 2015-2020 the area for production of dry pulses and protein crops increased by 64.5% in the European Union. In the same period in the Baltic countries, areas sown to grain legumes increased an average of five times (from 5820 to 42250 ha in Latvia, 51850 to 181090 ha in Lithuania and 10730 to 48600 ha in Estonia) (Eurostat, 2021).
In Latvia, the important grain legume crops are faba bean (Vicia faba L.) and field pea (Pisum sativum L.). Lupins (Lupinus angustifolius L. and L. luteus L.), common vetch (Vicia sativa L.), soybean (Glycine max (L.) Merr.) and common bean (Phaseolus vulgaris L.) are grown on small areas (Sterna et al., 2020; Central Statistical Bureau of Latvia, 2022). Lucerne (Medicago sativa L.), clovers (Trifolium spp.), fodder galega (Galega orientalis Lam.), and white sweet clover (Melilotus albus Medik.) are legumes grown for forage (Līpenīte and Kārkliņš, 2015). Given this diversity of legumes and their uses in agriculture, it is important to know about their shared diseases.
Chocolate spot disease of faba bean has usually been attributed to Botrytis fabae and to a lesser extent to B. cinerea (Davidson et al., 2007; Elad et al., 2016). Distinguishing species by the symptoms they cause is usually not possible, considering that species can occur in complexes. In 2010 in China, a new Botrytis sp., named B. fabiopsis, was found and described in complex with B. cinerea and B. fabae in faba bean where it occurred with higher frequency than B. fabae (Zhang et al., 2010). B. pseudocinerea was first discovered in French vineyards, then isolated from faba bean in Germany, and increasing numbers of studies have revealed its presence on different host-plants in Europe, North America, China and Australia (Walker et al., 2011; Saito et al., 2014; Harper et al., 2019; Xue et al., 2019; Plesken et al., 2021). These four Botrytis species have all been associated with the chocolate spot disease of faba bean, most recently in Latvia (Bankina et al., 2021).
Recent studies have shown the ability of the lesser known Botrytis species to infect different legumes in vitro. B. euroamericana was able to infect chickpea, field pea, lentil and faba bean leaves during pathogenicity tests in vitro at the same rate as B. cinerea (Moparthi et al., 2023). B. caroliniana, B. pyriformis, B. aclada and B. paeoniae formed lesions on faba bean leaves in vitro, but have not been isolated from field-grown faba bean (Li et al., 2012; Zhang et al., 2016; Garfinkel et al., 2017).
The species of Botrytis mainly have narrow host-plant specificity, but there are several species that infect a wide range of host plants. B. cinerea as a generalist can infect over 1400 plant species, including many legume crops (Ma & Michailides, 2005; Elad et al., 2016; Garfinkel, 2021). B. pseudocinerea has been found on several host plants, including grapevine, strawberry and oilseed rape, and was the predominant pathogen isolated from a faba bean field in Germany (Walker et al., 2011; Plesken et al., 2015; Weber and Hahn, 2019; Xue et al., 2019). The role of B. pseudocinerea in other legumes is unclear yet, but the species has a worldwide distribution and it seems to have a wide host range.
The minimum recommended interval for legumes in a crop rotation is 3-4 years (Pande et al., 2009). According to the ability of several Botrytis species to infect different legumes, we may suggest that Botrytis spp. can transfer between host-plants, volunteer plants and alternate hosts. Thus, for the successful inclusion of legume crops in agriculture, it is necessary to identify the Botrytis species occurring in legumes and to evaluate their pathogenicity for each legume crop. Exploring the interaction between Botrytis species and legume hosts will provide important information for understanding disease control methods in legumes and possible sources of inoculum.
Hence, we set out to identify the range of Botrytis species that could be isolated from grain legume crops in Latvia and to test their pathogenicity on other legume crop species. In this way, we could determine the potential for cross-infection of legume crops from other legumes or their post-harvest residues.
2 Materials and methods
2.1 Sample collection and fungal isolation
Infected plant parts of different legumes were collected between 2014 and 2019 (Figure 1; Table 1). Small tissue parts of infected leaves, pods and flowers or whole seeds were surface sterilized for 1 min with 1% sodium hypochlorite, rinsed three times in sterile distilled water and placed onto potato-dextrose agar (PDA) with streptomycin (100 mg L-1) and penicillin (100 mg L-1). Cultures were incubated at 20°C under 12 h light and 12 h darkness for one week. Single-hyphal isolates were obtained from each colony and grown on PDA (Leyronas et al., 2012).

Figure 1 Legume crops with visible fungal infection symptoms: common bean pod (A), chickpea pod (B) and red clover flowers (C).
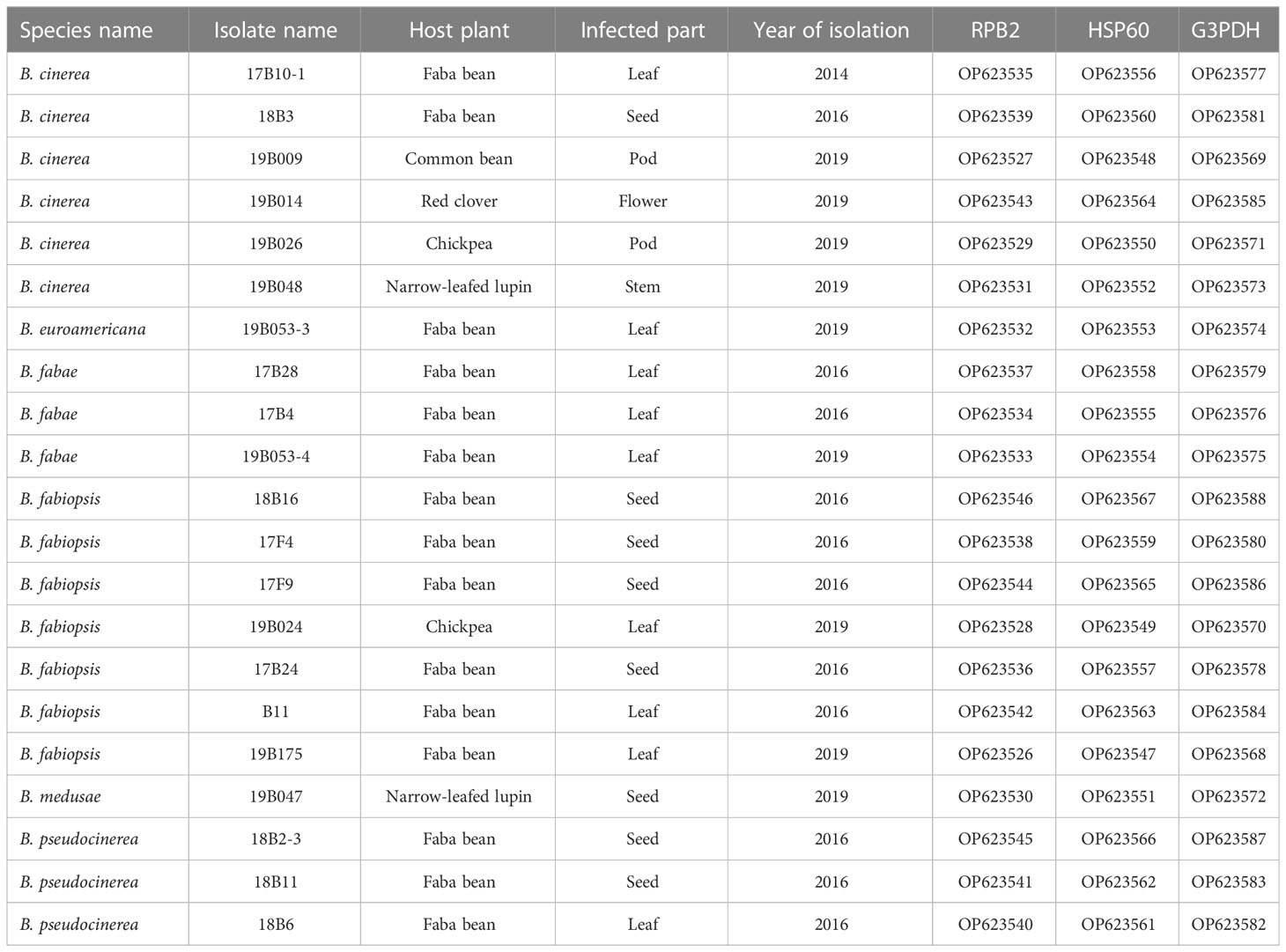
Table 1 Identities, origins and GenBank accession numbers of the sequences for the selected Botrytis spp. isolates.
Pure cultures were evaluated for morphological characteristics and subjected to preliminary molecular genetic analyses of the ITS region, using ITS1-F, ITS4 (reverse) primers. The final collection comprised 278 Botrytis putative isolates showing traits typical of the genus. After the identification to species level by sequencing three nuclear genes RPB2, HSP60 and G3PDH (described in the following section), a set of 21 isolate was selected for further investigations. Isolates of all determined species originated from different hosts were choosen.
For each selected isolate, 10 working cultures were established by transferring a mycelial plug (5 mm ø) from the margins of the one-week-old culture: nine for pathogenicity tests and one for molecular genetic analyses.
2.2 Molecular genetic analyses
About 10 mg of fungal material was collected from petri dishes using a sterile cell scraper with a 20 mm lifter blade (Bellco Glass, USA) (in our experience this amount covers ~ 1/3 of scraper blade) and suspended in 500 µl NucleoMag® 96 Plant kit (Macherey-Nagel, Germany) Lysis buffer. This was followed by homogenization for 2×60 sec using FastPrep®-24 instrument and Lysing Matrix D (MP Biomedicals, USA), phenol (Alfa Aesar, Germany) - chloroform (Merck, USA) treatment and subsequent purification with the NucleoMag® 96 Plant kit according to the manufacturer’s instructions.
The identification to species level was carried out by sequencing three nuclear genes: RNA polymerase II (RPB2), Heat shock protein 60 (HSP60) and Glyceraldehyde 3-phosphate dehydrogenase (G3PDH). Primer combinations (Table 2) were used according to Staats et al. (2005), and Raja et al. (2017) who showed that these three gene sequences were diagnostic for discriminating Botrytis species from each other.
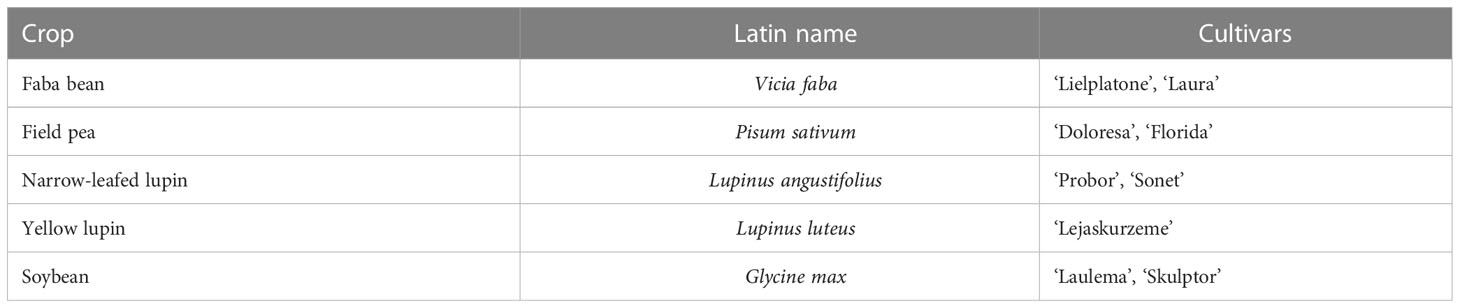
Table 2 Legume crops used in the pathogenicity tests.
The amplification of RPB2, HSP60 and G3PDH by PCR was carried out for 40 cycles (98°C-5 sec, 59°C-5 sec, 72°C-20 sec, GeneAmp PCR System 9700 (Applied Biosystems, USA)) in a total volume of 20 µl. The reaction mixture was comprised of following components: 10 µl of H2X Phire Plant Direct PCR Master Mix (Thermo Fisher Scientific, USA), 0.3 µM forward primer and 0.3 µM reverse primer (Table 3) (Staats et al., 2005) and eluted 1 µl of fungal DNA solution. The success of the amplification was verified through the inspection of PCR products by agarose gel electrophoresis. Samples were considered positive when they contained a >700bp PCR fragment.

Table 3 Primers for PCR amplification and sequencing.
Positive reaction mixtures were cleaned up from excess of dNTPs and primers through employment of Exonuclease I (0.5 µl) (Thermo Fisher Scientific, USA) and Shrimp Alkaline Phosphatase (2 µl) (Thermo Fisher Scientific, USA) (incubated for 40 min at 37°C and inactivated at 95°C for 20 min). Further, 1 µl of cleaned fragment solution was transferred to BigDye® Terminator v3.1 Cycle Sequencing reaction mixture which was prepared according to the manufacturer’s instructions (Applied Biosystems, USA). Both DNA strands of every PCR product were sequenced using primers ITS1-F, ITS4 (reverse), G3PDHfor1, G3PDHrev1, HSP60for1, HSP60rev1, RPB2for1 and RPB2rev1 and sequencing products were analyzed on 3130xl Genetic Analyzer (Applied Biosystems, USA). The ITS region sequences from each isolate were aligned using MEGA11 software (v. 11.0.13) and the created contig was BLAST® searched against the NCBI nucleotide database to identify taxonomic sources of records with the highest sequence identity (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
G3PDH, HSP60, and RPB2 gene sequences were aligned with other Botrytis sequences, downloaded from NCBI nucleotide database, including the sequences of Sclerotinia sclerotiorum and Monilinia fructigena as outgroup taxa (Staats et al., 2005). The alignment of sequences was carried out by Multiple Alignment using Fast Fourier Transform (MAFFT) (Madeira et al., 2022), alignments were trimmed by trimAl software employing the strict method (Capella-Gutiérrez et al., 2009) and clustered by MEGA5 software employing the ‘Neighbor Joining’ algorithm to construct the phylogenetic tree (Tamura et al., 2011).
2.3 Pathogenicity test
The test of pathogenicity was performed in vitro on leaves of two cultivars of each of faba bean, field pea, narrow-leafed lupin and soybean, and one of yellow lupin (Table 2).
Two layers of sterile filter paper sheets were used to line metal trays (40 × 30 × 5 cm). Four 28 cm long glass sticks were arranged into each tray to support the leaves. An equal amount of water was poured into each tray, so that it was moist but water did not accumulate in puddles. Leaves were obtained from the middle level of plants at BBCH growth stage 35 (5 visibly extended internodes) and rinsed twice with sterile water. Whole leaves of faba bean and field pea were put onto the glass sticks in the metal trays so that only the petiole touched the wet filter paper. Soybean leaves were divided into leaflets and placed in metal trays so that only the cut surface touched the water. For lupin leaves, three glass sticks were tied together and three middle leaflets were detached and placed on them so as not to touch the filter paper.
Each tray contained 30 plant leaflets, one for each of the isolates and some controls. Agar plugs with Botrytis isolates (5 mm ø) were placed on leaflets with the mycelium side down. Two non-inoculated agar plugs were used in each tray as a control. After inoculation, the trays were covered with plastic film to maintain high humidity and incubated at 20°C in the dark for 72 h. The diameter of the formed lesions under the inoculated plug was measured crosswise once a day until some of lesions reached the edge of the leaf. Four replicates were prepared and the experiment was repeated, giving 8 replicates. A further repeat was conducted for narrow-leafed lupin ‘Probor’, giving 12 replicates. Mean lesion diameter was considered as the measure of pathogenicity.
2.4 Data analysis
Leaflet datasets were subjected to analysis of variance using SPSS v. 27 (IBM Inc., Chicago, IL, USA) within each host genus (yellow and narrow-leafed lupins were analysed together). A simple split-plot model was applied to the data from each day, where the culture box was the main plot and the leaflet was the subplot. The repeated-measures model was then applied to 4 days of data from faba bean and soybean as well as to 3 days of data from lupins and pea. The output was corrected to take into account the split-plot design of the experiment.
3 Results
3.1 Molecular-genetic analysis
The 26 isolates collected from legumes in this study with the Botrytis spp. isolates from GenBank formed a clade with 99% bootstrap support, distinct from the closely related M. fructigena and S. sclerotiorum outgroups. Branches with under 50% bootstrap support were not included in the phylogenetic tree (Figure 2). Phylogenetic analysis of the combined gene datasets confirmed that all isolates belonged to the genus Botrytis. Six isolates clustered with B. cinerea references, three with B. fabae, seven with B. fabiopsis and three with B. pseudocinerea. One isolate was grouped with B. medusae and one isolate with B. euroamericana (Table 1). Five isolates did not clearly group with identifiable clades, so they were not considered further in this study.
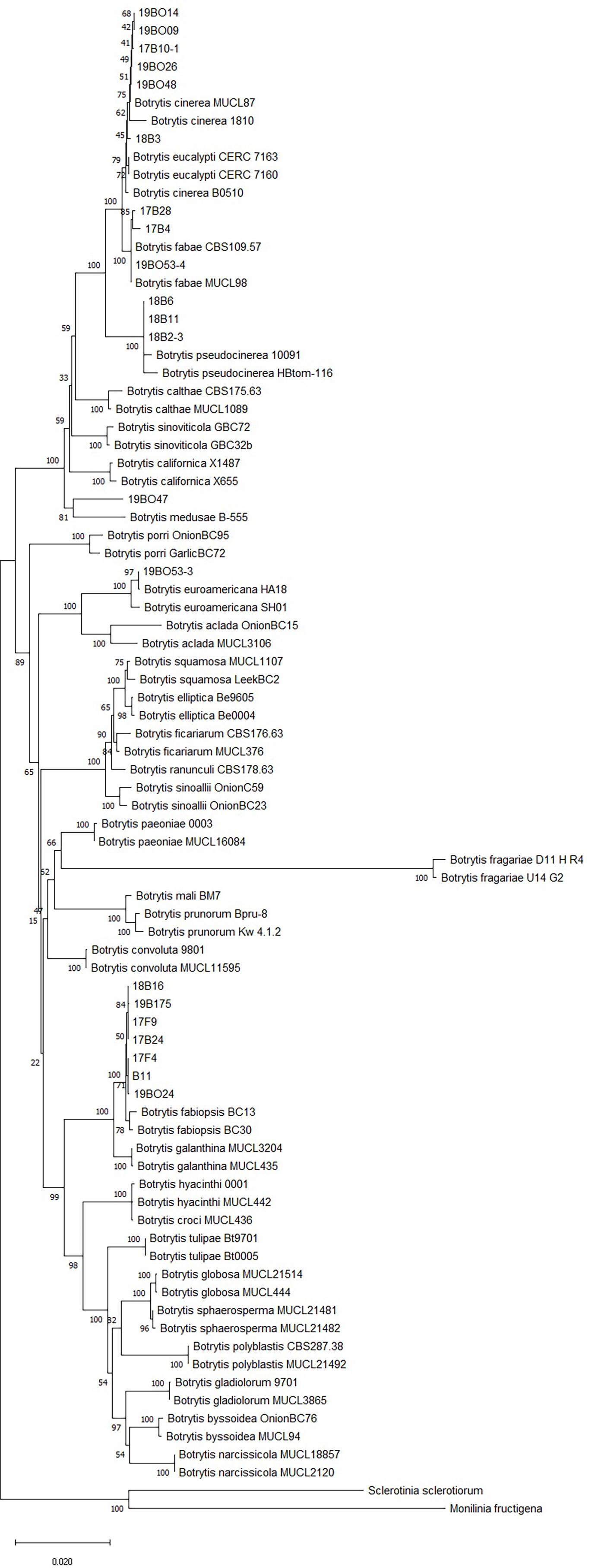
Figure 2 Neighbor joining tree of RPB2, HSP60 and G3PDH gene sequences. The tree describes the relationship of Botrytis species isolates collected from legumes in Latvia to named Botrytis species including the sequences of Sclerotinia sclerotiorum and Monilinia fructigena as outgroup taxa. Branches with <50% bootstrap support are not shown.
3.2 Pathogenicity tests
The first symptoms of infection were observed 24 h after inoculation of all host cultivars. The main effects of Botrytis species and isolates were significant for all measures on all host species (Table 4). The effects of replicate and of the day × replicate interaction were significant for pea, because lesion diameters were 1 mm greater in replicates 1 to 4 than those in replicates 5 to 8 on day 1, which continued until day 3, when it was no longer significant.
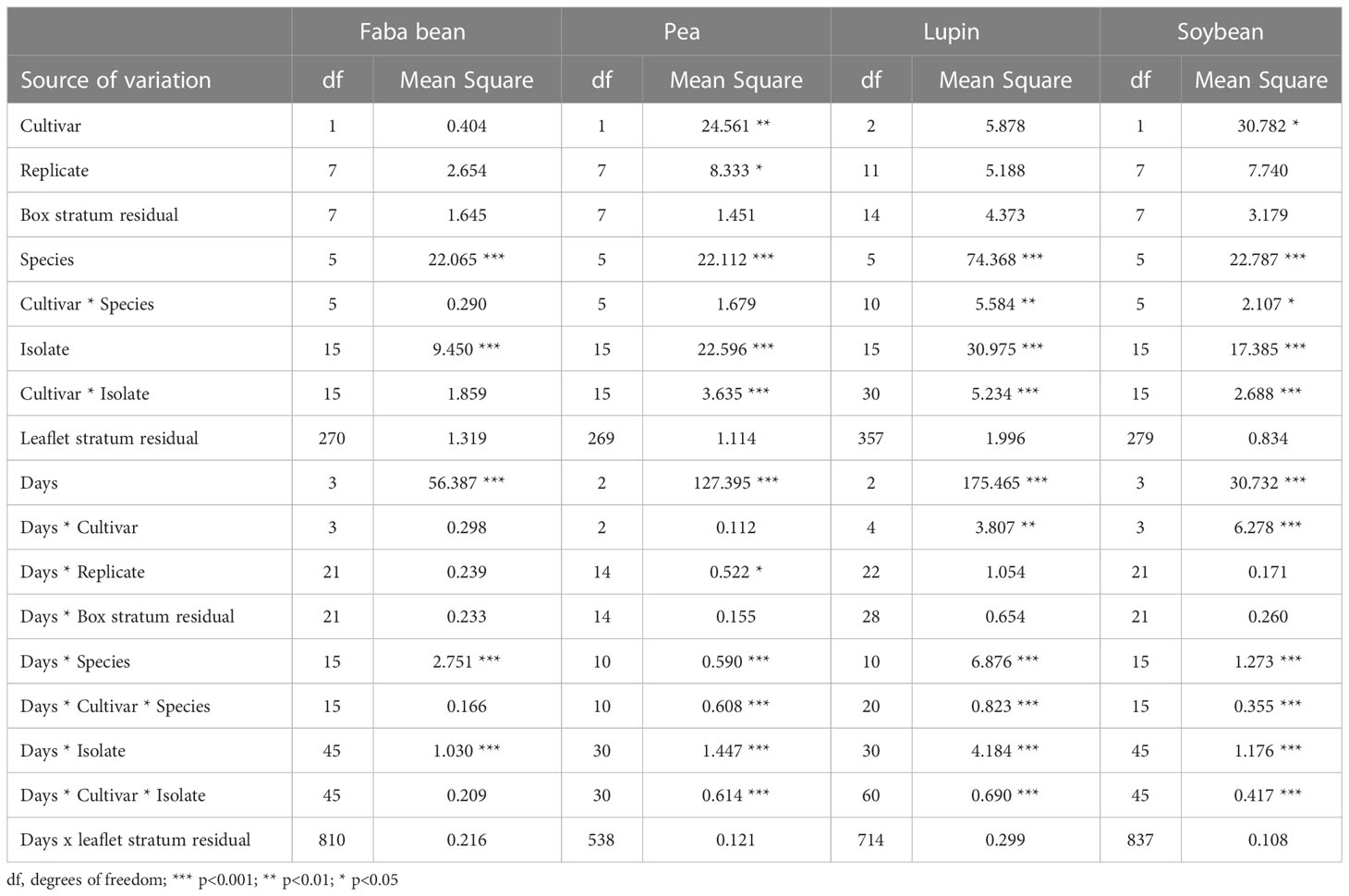
Table 4 Summary analysis of variance of Botrytis spp. pathogenicity on faba bean, pea, lupin and soybean leaves.
Differences between cultivars were significant in soybean, where lesions were larger on ‘Skulptor’ than those on ‘Laulema’ from day 2 onwards, and in pea, where the lesions on ‘Doloresa’ were larger than those on ‘Florida’ on all dates. The cultivar × species interaction was significant in soybean and lupin, while the cultivar × isolate interaction was significant for all species except faba bean. On days 1 and 2, B. cinerea and B. pseudocinerea caused much larger lesions on the yellow lupin ‘Lejaskurzeme’ than on the narrow-leafed cultivars. On day 1, lesions of B. medusae on ‘Probor’ were much larger than those on the other two lupin cultivars and on days 2 and 3, ‘Sonet’ had smaller lesions than the other two cultivars. On days 2 and 3, both B. euroamericana and B. fabae produced larger lesions on ‘Sonet’ than on the other two lupins.
To compare the pathogenicity of Botrytis spp., the optimal assessment day was examined for each host species, when the leaves were still fresh and the lesion diameter was determined. For pea, the optimal assessment day was the third, for lupins the second, for soybean the fourth, and for faba bean the fifth day after infection. Based on the size of the lesions and the characteristics of spot formation in each host species, we divided the pathogenicity of the isolates into three classes: – low (primary lesion only), moderate, and high (Table 5).
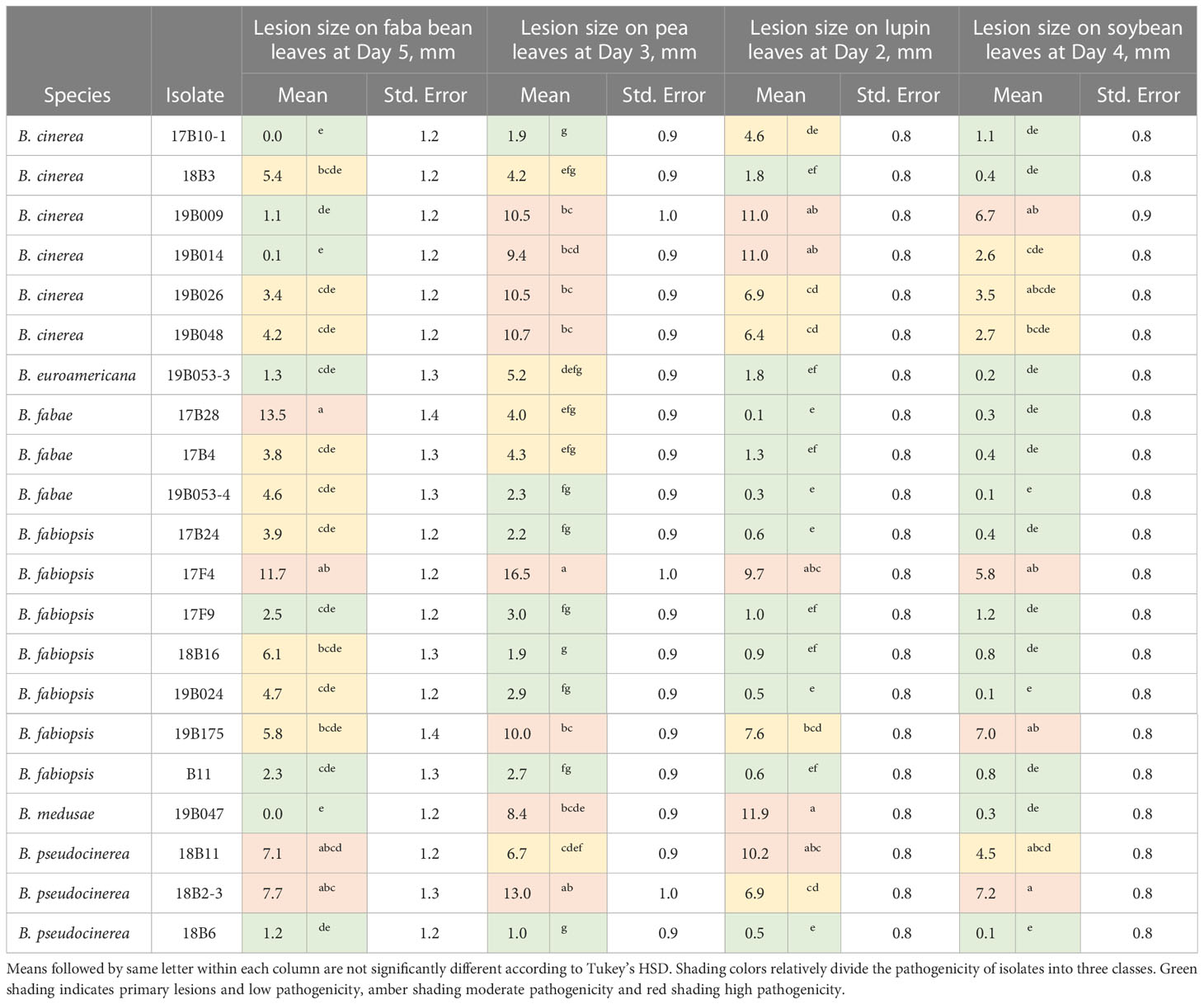
Table 5 Pathogenicity of Botrytis spp. on faba bean, lupin, pea and soybean leaves.
In faba bean, B. fabae caused the largest lesions on day 5 (Figure 3), with isolate 17B28 being the strongest (Table 5). The lesions produced by B. pseudocinerea and B. fabiopsis were somewhat smaller than those caused by B. fabae and were similar to each other. B. fabiopsis isolate 17F4 produced lesions second to those of B. fabae 17B28 in size. Isolates 18B11 and 18B2-3 of B. cinerea exhibited high pathogenicity on faba bean leaves; however, as a species it was less pathogenic than B. pseudocinerea. Lesions caused by B. euroamericana were observed only in cv. ‘Lielplatone’. B. medusae did not cause any injury in either faba bean cultivar.
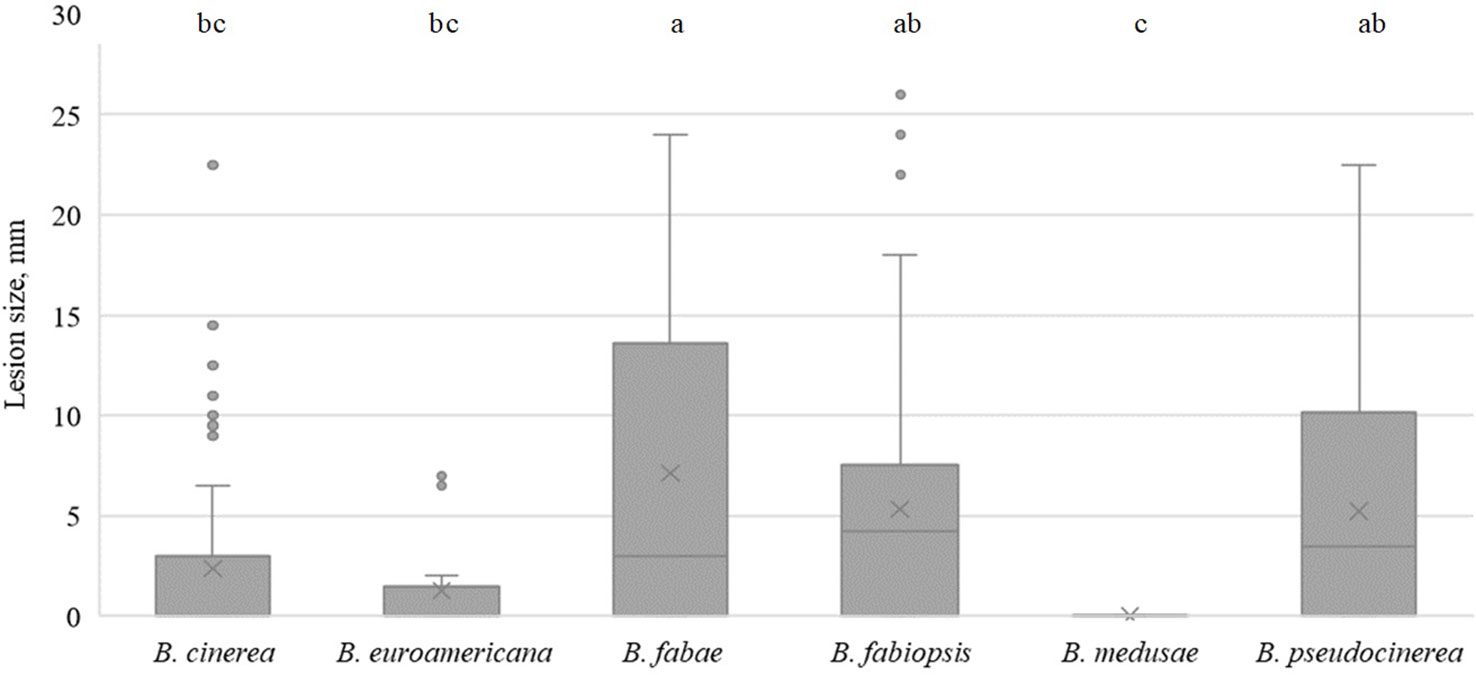
Figure 3 Box-plot diagram of the lesion diameters on faba bean leaves five days after inoculation with isolates of 6 Botrytis species. Boxes with the same letter are not significantly different according to Tukey’s HSD.
On day 3, all species caused lesions on pea leaves, and the most pathogenic were B. medusae and B. cinerea (Figure 4). B. fabae was significantly less pathogenic. In general, B. fabiopsis caused primary or moderate lesions, except 17F4 and 19B175 (Table 4), which formed large lesions on both the tested pea cultivars. Isolate 17F4 caused lesions that were even larger than those of the most pathogenic B. cinerea isolates. B. pseudocinerea isolate 18B2-3 was also responsible for large lesions.
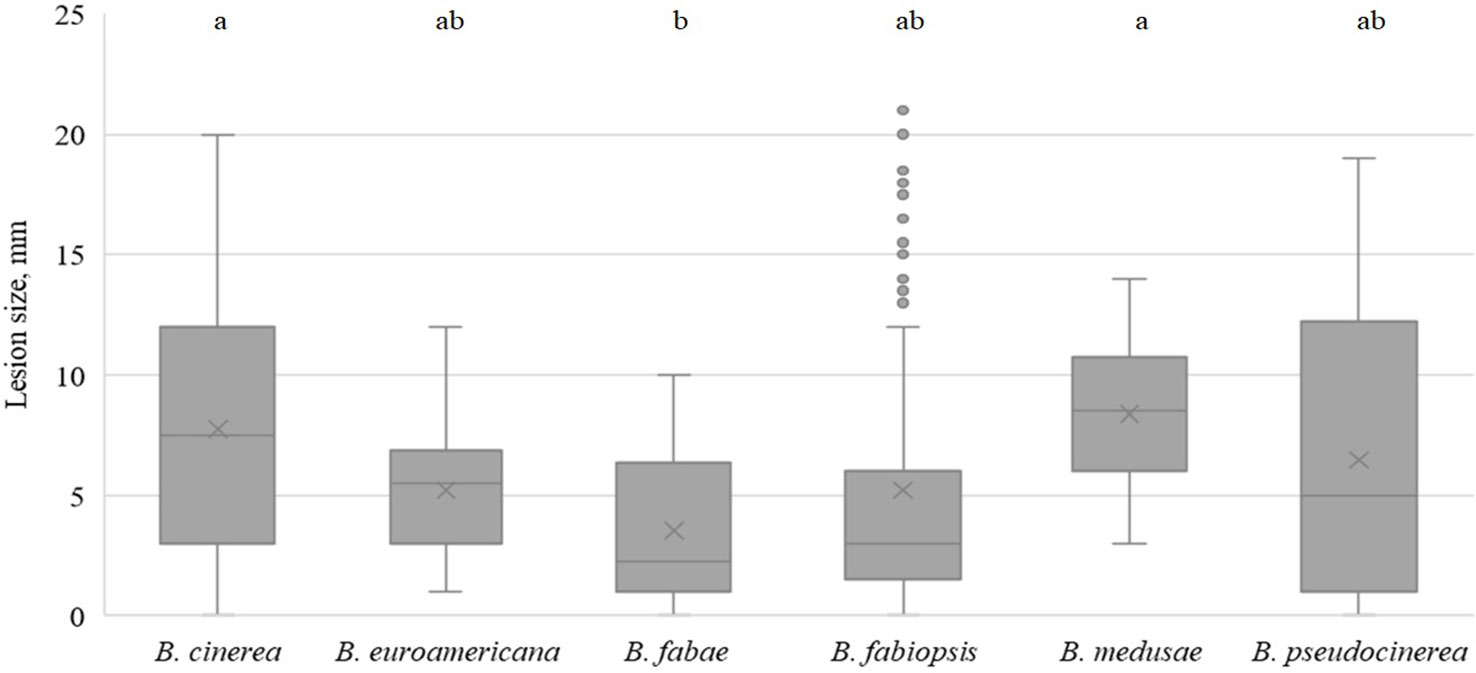
Figure 4 Box-plot diagram of the lesion diameters on field pea leaves three days after inoculation with isolates of 6 Botrytis species. Boxes with the same letter are not significantly different according to Tukey’s HSD.
On lupin leaves, B. medusae was the most pathogenic (Table 5) and caused significantly larger lesions than the other Botrytis spp. (Figure 5). B. pseudocinerea and B. cinerea caused moderate to large lesions, depending on the isolate, and their pathogenicity was significantly higher than that of B. fabae, B. fabiopsis and B. euroamericana, although B. fabiopsis isolate 17F4 produced lesions that were nearly as large as those of the most pathogenic B. cinerea isolates (Table 5).
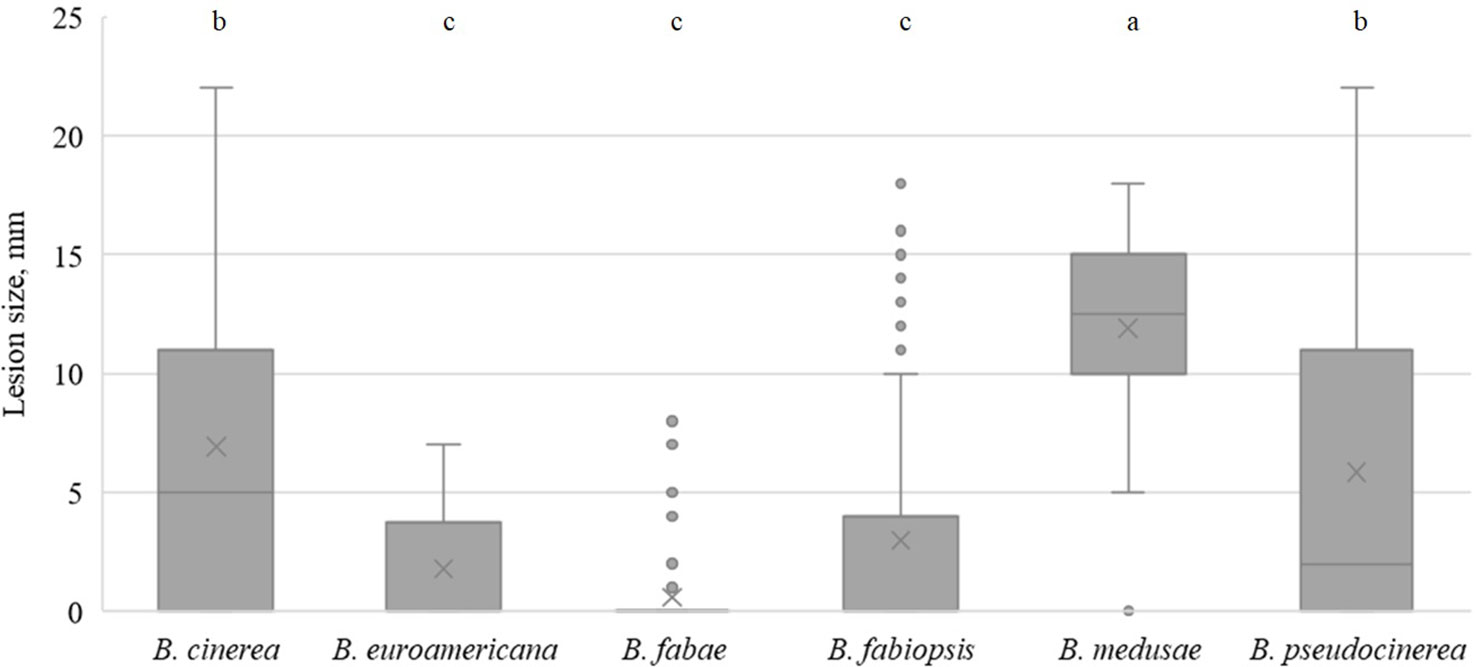
Figure 5 Box-plot diagram of the lesion diameters on lupin leaves two days after inoculation with isolates of 6 Botrytis species. Boxes with the same letter are not significantly different according to Tukey’s HSD.
Lesion sizes were smaller in soybean than on the other host species (Figure 6). B. fabae, B. euroamericana and B. medusae caused only primary lesions on both soybean cultivars. One B. cinerea, two B. fabiopsis and one B. pseudocinerea isolate caused lesions larger than 5.5 mm in diameter (Table 5), that larger than those caused by the agar plug of inoculum. Cultivar ‘Skulptor’ was notably more susceptible to B. pseudocinerea than cultivar ‘Laulema’.
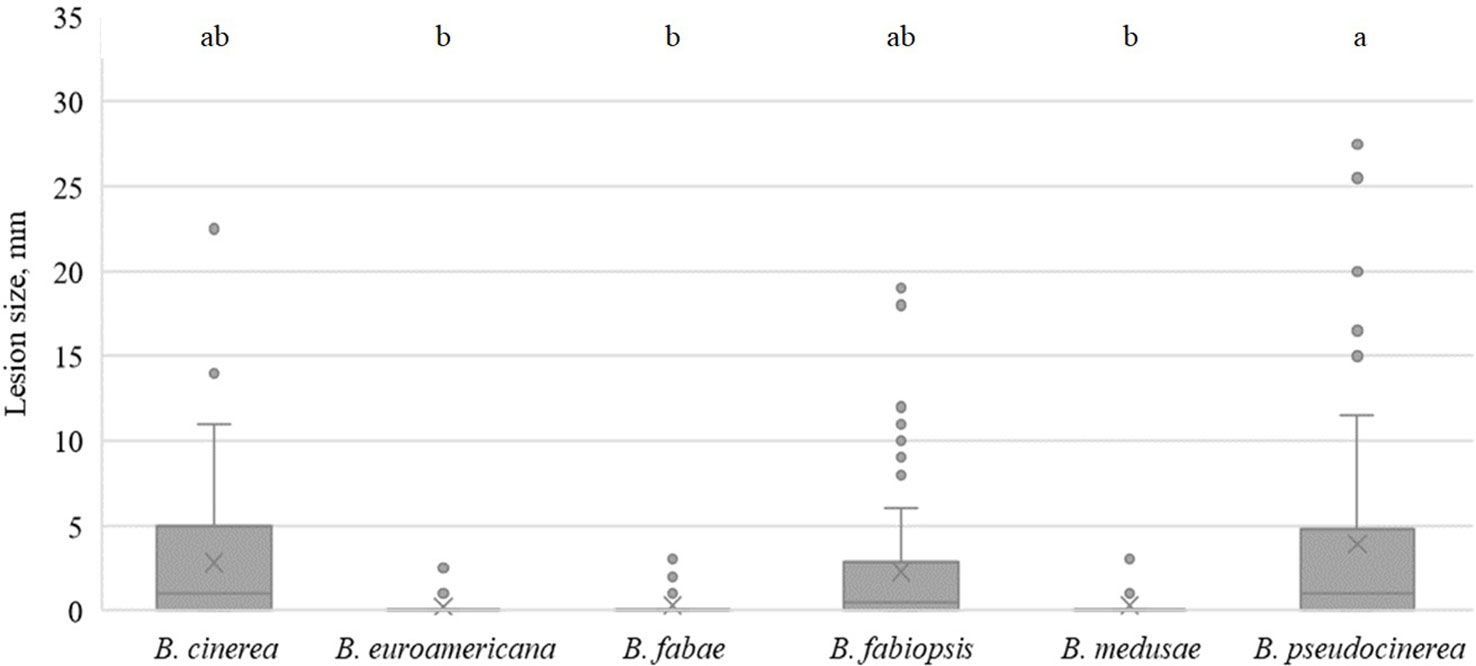
Figure 6 Box-plot diagram of the lesion diameter on soybean leaves four days after inoculation with isolates of 6 Botrytis species. Boxes with the same letter are not significantly different according to Tukey’s HSD.
Certain isolates were highly pathogenic to all host species; notably B. fabiopsis 17F4 (highly pathogenic on all), B. cinerea 19B009 and B. pseudocinerea 18B2-3 (highly pathogenic on three and moderately pathogenic on one), and B. fabiopsis 19B175 and B. pseudocinerea 18B11 (highly pathogenic on two and moderately pathogenic on two host species (Table 5)). At the other extreme, B. fabiopsis 17F9 and B11, along with B. pseudocinerea 18B6 had, low pathogenicity in all hosts.
4 Discussion
A high diversity of Botrytis spp. was recovered from these legumes. In the main legume crop of Latvia, faba bean, five Botrytis spp. were isolated: B. cinerea, B. fabae, B. pseudocinerea, B. fabiopsis and B. euroamericana.
B. fabae is considered the most important pathogen and primary causal agent of chocolate spot disease in faba bean because of its narrow specialization (Davidson et al., 2007). In the present study, B. fabae caused lesions on faba bean and moderate lesions on pea, but only primary lesions on soybean and lupin, thus acting as a narrowly specialized faba bean pathogen. B. fabae has occasionally been isolated from other genera in the Fabaceae, including field pea (Elad et al., 2016).
B. cinerea can be observed as a pathogen of faba bean, either separately or in a complex with B. fabae (Harrison, 1988; Zhang et al., 2010). As expected, B. cinerea was isolated from a wide variety of legumes including faba bean, common bean, red clover, chickpea, and narrow-leafed lupin. Pathogenicity tests did not exhibit clear regularity between the natural host-plants and pathogenicity of these isolates. Isolate 17B10-1 from faba bean did not produce any lesions on faba bean during these tests, while isolate 19B048 from narrow-leafed lupin caused more damage to pea than to lupin. These results suggest that these isolates were saprophytic on plant tissues that were already suffering from the attack of other pathogens. The Botrytis genus is known to have high intraspecific variation, and unstable pathogenicity has been observed within individual isolates (Garfinkel, 2021). In general B. cinerea exhibited a high variability in pathogenicity.
In this study, some isolates of B. pseudocinerea and B. fabiopsis produced large lesions on the detached leaves of all host species and some caused mild disease. These species have been much less studied than the previous two and found in relatively few countries. Both have been implicated in the chocolate spot disease of faba bean in several countries, including Latvia (Bankina et al., 2021). B. pseudocinerea has been found in various unrelated host plants (Plesken et al., 2015; Garfinkel, 2021).
B. fabiopsis was isolated from faba bean leaves and seeds as well as from chickpea leaves. Most isolates acted as a narrow specialist pathogen; however, but one isolate was highly pathogenic in all hosts. The current study is the first report of B. fabiopsis naturally occurring in chickpea and the first demonstration that it can cause disease in pea, narrow-leafed lupin, yellow lupin and soybean. To date, no information is available on the occurrence of B. fabiopsis elsewhere than in faba bean in China (Zhang et al., 2010) and Latvia (Bankina et al., 2021).
The isolate of B. euroamericana was obtained from faba bean leaves but caused only primary lesions on this species, while it was moderately pathogenic on pea leaves. The present isolate was purified from two B. cinerea isolates obtained from a single leaf lesion, so it may have been an endophytic or saprophytic follower on the lesion caused by B. cinerea. This is the first report of B. euroamericana occurring naturally on faba bean leaves. Originally, B. euroamericana was isolated from peony and grapes in North America and Europe (Garfinkel et al., 2017). Later B. euroamericana was reported as a causative agent of grey mold on chickpea leaves in the USA (Moparthi et al., 2020) and was observed on lentil and dry pea seeds. The ability to cause lesions in several hosts from different plant families during pathogenicity tests confirms the wide host range of this pathogen.
The single B. medusae isolate was obtained from lupin seeds and of all the Botrytis species tested here, it showed the highest pathogenicity on lupin leaves, along with convincing pathogenicity on field pea leaves while it formed primary lesions on faba bean leaves. To the best of our knowledge, this study is the first report to B. medusae occurring naturally in lupin. Originally, B. medusae was collected in 2015 from wine grapes (Harper et al., 2019) and has otherwise been little reported on.
Garfinkel's (2021) review suggested that many of the newly discovered Botrytis species may be polyphagous. This study confirmed the occurrence of four new species, discovered since 2010, on crops different from the original host-plant families: B. fabiopsis on chickpea, B. pseudocinerea, B. euroamericana on faba bean, and B. medusae on narrow-leafed lupin.
Pathogenicity tests on detached leaves do not always reflect the true pathogenicity of a pathogen under field conditions; however, if there is a possibility that isolates in nature may behave similar to the pathogenicity tests, the risk of transmission between host plants may be higher. Further research of lesser-known Botrytis sp. in different legumes is needed to clarify their occurrence and significance in disease initiation in all hosts.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author contributions
EB-M and FS performed the data analysis and wrote the manuscript. EB-M and IP performed pathogenicity studies. EB-M and JK did the sampling and genetic characterization of the isolates. BB and GB did the conception and design of the study. IN-L did fungal isolation and creation of fungal collection. AR and DF performed sequencing. All authors contributed to the article and approved the submitted version.
Funding
The research was supported by the project of the Latvian Council of Sciences No. LZP-2019/1-0034 ‘Pathogenicity and diversity of Botrytis spp. – important causal agents of legume diseases’.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bankina, B., Stoddard, F. L., Kaņeps, J., Brauna-Morževska, E., Bimšteine, G., Neusa-Luca, I., et al. (2021). Botrytis four species are associated with chocolate spot disease of faba bean in Latvia. Zemdirbyste-Agriculture 108 (4), 297–302. doi: 10.13080/z-a.2021.108.038
Capella-Gutiérrez, S., Silla-Martínez, J. M., Gabaldón, T. (2009). trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25 (15), 1972–1973. doi: 10.1093/bioinformatics/btp348
Central Statistical Bureau of Latvia (2022) Central statistical bureau of Latvia database. Available at: https://data.stat.gov.lv/pxweb/lv/OSP_PUB/START:NOZ:LA:LAG/LAG020/table/tableViewLayout1/.
Davidson, J. A., Pande, S., Bretag, T. W., Lindbeck, K. D., Krishna-Kishore, G. (2007). ‘Biology and management of Botrytis spp. in legume crops,’ in Botrytis: biology, pathology and control (Dordrecht: Springer), 295–318. doi: 10.1007/978-1-4020-2626-3_16
Elad, Y., Pertot, I., Prado, A. M. C., Stewart, A. (2016). ‘Plant hosts of Botrytis spp,’ in Botrytis–the fungus, the pathogen and its management in agricultural systems (Champagne, IL, USA: Springer), 413–486. doi: 10.1007/978-3-319-23371-0_20
Eurostat (2021) Dry pulses and protein crops for the production of grain (including seed and mixtures of cereals and pulses) by area, production and humidity. Available at: https://ec.europa.eu/eurostat/databrowser/view/APRO_CPSH1:custom_1774212/default/table?lang=en.
Garfinkel, A. R. (2021). The history of Botrytis taxonomy, the rise of phylogenetics, and implications for species recognition. Phytopathology 111 (3), 437–454. doi: 10.1094/PHYTO-06-20-0211-IA
Garfinkel, A. R., Lorenzini, M., Zapparoli, G., Chastagner, G. A. (2017). Botrytis euroamericana, a new species from peony and grape in north America and Europe. Mycologia 109 (3), 495–507. doi: 10.1080/00275514.2017.1354169
Harper, L. A., Derbyshire, M. C., Lopez-Ruiz, F. J. (2019). Identification and characterization of Botrytis medusae, a novel cryptic species causing grey mould on wine grapes in Australia. Plant Pathol. 68 (5), 939–953. doi: 10.1111/ppa.13005
Harrison, J. G. (1988). The biology of Botrytis spp. on Vicia beans and chocolate spot disease–a review. Plant Pathol. 37, 168–201. doi: 10.1111/j.1365-3059.1988.tb02064.x
Leyronas, C., Duffaud, M., Nicot, P. C. (2012). Compared efficiency of the isolation methods for Botrytis cinerea. Mycology 3 (4), 221–225. doi: 10.1080/21501203.2012.727484
Li, X., Kerrigan, J., Chai, W., Schnabel, G. (2012). Botrytis caroliniana, a new species isolated from blackberry in south Carolina. Mycologia 104 (3), 650–658. doi: 10.3852/11-218
Līpenīte, I., Kārkliņš, A. (2015). ‘Tauriņziežu audzēšanas vides riski (Cultivation of leguminous crops and environmental risks),’ in Proceedings of the Scientific and Practical Conference ‘Harmonious Agriculture’, 19-20 February 2015, Jelgava, Latvian. 24–37.
Ma, Z., Michailides, T. J. (2005). Genetic structure of Botrytis cinerea populations from different host-plants in California. Plant Dis. 89 (10), 1083–1089. doi: 10.1094/pd-89-1083
Madeira, F., Pearce, M., Tivey, A. R. N., Basutkar, P., Lee, J., Edbali, O., et al. (2022). Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 50 (W1), W276–W279. doi: 10.1093/nar/gkac240
Moparthi, S., Parikh, L. P., Gunnink Troth, E., Burrows, M. E. (2023). Identification and prevalence of seedborne botrytis spp. in dry pea, lentil, and chickpea in Montana. Plant Dis. 107 (2), 382–392. doi: 10.1094/PDIS-05-22-1236-RE
Moparthi, S., Peluola, C., Agindotan, B., McPhee, K., Burrows, M. (2020). First report of gray mold of chickpea caused by Botrytis euroamericana in the USA. Crop Prot. 137, 105297. doi: 10.1016/j.cropro.2020.105297
Pande, S., Sharma, M., Kumari, S., Gaur, P. M., Chen, W., Kaur, L., et al. (2009). ‘Integrated foliar diseases management of legumes,’ in Proceedings of international conference on grain legumes: Quality improvement, value addition and trade, 14–16 February 2009 (Kanpur, Uttar Pradesh, India: Indian Institute of Pulses Research), 143–167.
Plesken, C., Pattar, P., Reiss, B., Noor, Z. N., Zhang, L., Klug, K., et al. (2021). Genetic diversity of Botrytis cinerea revealed by multilocus sequencing, and identification of b. cinerea populations showing genetic isolation and distinct host adaptation. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.663027
Plesken, C., Weber, R. W., Rupp, S., Leroch, M., Hahn, M. (2015). Botrytis pseudocinerea is a significant pathogen of several crop plants but susceptible to displacement by fungicide-resistant b. cinerea strains. Appl. Environ. Microbiol. 81 (20), 7048–7056. doi: 10.1128/AEM.01719-15
Raja, H. A., Miller, A. N., Pearce, C. J., Oberlies, N. H. (2017). Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 80 (3), 756–770. doi: 10.1021/acs.jnatprod.6b01085
Saito, S., Michailides, T. J., Xiao, C. L. (2014). First report of Botrytis pseudocinerea causing gray mold on blueberry in north America. Plant Dis. 98 (12), 1743. doi: 10.1094/PDIS-06-14-0573-PDN
Staats, M., van Baarlen, P., van Kan, J. A. (2005). Molecular phylogeny of the plant pathogenic genus Botrytis and the evolution of host specificity. Mol. Biol. Evol. 22 (2), 333–346. doi: 10.1093/molbev/msi020
Sterna, V., Zute, S., Jansone, I., Ence, E., Strausa, E. (2020). Evaluation of various legume species and varieties grown in Latvia as a raw material of plant-based protein products. Agron. Res. 18 (4), 2602–2612. doi: 10.15159/ar.20.215
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28 (10), 2731–2739. doi: 10.1093/molbev/msr121
Walker, A. S., Gautier, A., Confais, J., Martinho, D., Viaud, M., Le Pêcheur, P., et al. (2011). Botrytis pseudocinerea, a new cryptic species causing gray mold in French vineyards in sympatry with Botrytis cinerea. Phytopathology 101 (12), 1433–1445. doi: 10.1094/PHYTO-04-11-0104
Weber, R. W., Hahn, M. (2019). Grey mould disease of strawberry in northern Germany: causal agents, fungicide resistance and management strategies. Appl. Microbiol. Biotechnol. 103 (4), 1589–1597. doi: 10.1007/s00253-018-09590-1
Xue, L. H., Liu, Y., Zhang, L., Huang, X. Q., Zhou, X. Q., Yang, X. X., et al. (2019). Botrytis pseudocinerea, a new pathogen causing gray mold on Brassica napus in China. Plant Dis. 103 (2), 367. doi: 10.1094/PDIS-04-18-0688-PDN
Zhang, J., Wu, M. D., Li, G. Q., Yang, L., Yu, L., Jiang, D. H., et al. (2010). Botrytis fabiopsis, a new species causing chocolate spot of broad bean in central China. Mycologia 102 (5), 1114–1126. doi: 10.3852/09-217
Keywords: Botrytis, legumes, chocolate spot, gray mold, pathogenicity tests, B. euroamericana, B. medusae
Citation: Brauna-Morževska E, Stoddard FL, Bankina B, Kaņeps J, Bimšteine G, Petrova I, Neusa-Luca I, Roga A and Fridmanis D (2023) Evaluation of pathogenicity of Botrytis species isolated from different legumes. Front. Plant Sci. 14:1069126. doi: 10.3389/fpls.2023.1069126
Received: 13 October 2022; Accepted: 13 March 2023;
Published: 27 March 2023.
Edited by:
Islam Hamim, Bangladesh Agricultural University, BangladeshReviewed by:
Mora-Romero Guadalupe Arlene, Universidad Autónoma de Occidente, MexicoZhexin Li, Chongqing University of Arts and Sciences, China
Dhruv Aditya Srivastava, Hebrew University of Jerusalem, Israel
Malkhan Singh Gurjar, Indian Agricultural Research Institute (ICAR), India
Delong Li, Qingdao Agricultural University, China
Copyright © 2023 Brauna-Morževska, Stoddard, Bankina, Kaņeps, Bimšteine, Petrova, Neusa-Luca, Roga and Fridmanis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elīna Brauna-Morževska, Biruta.Bankina@lbtu.lv