
【Evolution of Bambusoideae】Bamboos

Subfamily Bambusoideae
Poaceae, the family of grasses. One of the most successful and ubiquitous lineages of the monocot class (move way, Orchidaceae).[1] Members of this family are known for dominating dry treeless plains, hence the name grassland. Bamboos, however, are an anomaly. Contrasting with many of their iconic relatives, Bambusoideae presents the only major grass lineage to diversify primarily in association with mesic-to-wet forests.[2][3] Their superficial tree-like appearance, built by its strongly lignified stem, also stands them out in their kindred of midgets.[3]
Not all bamboos are tall and mighty, though. As bamboos evolved from grassy poacean ancestors (bamboos’ closest subfamily is Pooideae, containing wheat), naturally, groups that diverged early will have retained the herbaceous plesiomorphy. Bambusoideae has three tribes: the herbaceous Olyreae, the woody Arundinarieae (Ah-run-dee-neh-ree-ee-ee?), and the woody “Bambuseae.” [2]
(Tribe is apparently a taxonomic level between family and genus, but like why do taxonomists add this if those categories aren’t enough for the matryoshka clades they establish all day anyways…)
Even though the existence of those three tribes is fairly certain (except Bambuseae, which I’ll elaborate later), the intertribal relationships between those clades remain disputed. Molecular analyses of plastid DNA tend to support the sister relationship between Olyreae and Bambuseae, indicating that the woody bamboo syndrome either is paraphyletic or evolved twice. On the other hand, molecular analyses on nuclear DNA end up with results that support the monophyly of the woody bamboos (note that here doesn’t use “syndrome”), though with complex hybridization within, and Olyreae being sister with the woody bamboos.[4]



This is apparently known as plastid-nuclear incongruence. Since plastid analyses test only exclusively maternal inheritance in bamboos,[5] the resulting phylogeny can be inaccurate when noised by evolutionary biases like hybridization and incomplete lineage sorting (see below).[6] Anyways, the author decides to adopt the second hypothesis using nuclear DNA because it is more holistic in genetic information and explains the polyploidy present in many bamboos better. Plus, according to [6], when dealing with polyploid organisms, choose nuclear phylogeny.


Caption: Diagram for hybridization and incomplete lineage sorting. Different colors represent homologous genes in each species; at the right is the actual phylogeny. Presumably, incomplete lineage sorting means the separation of genes before the separation of species, and hybridization just means gene exchange between species. Then how to explain the illusion of Olyreae being sister with Bambuseae in the plastid DNA analysis? Since Olyreae couldn’t possibly have hybridized in a way that affected their ploidy levels (because olyreans are normal diploids), and it’s implausible for incomplete lineage sorting to occur because olyrean divergence happened way before bambusean divergence, therefore, the hypothetical explanation the author proposes is that olyreans hybridized with one of the ancestral lineages of Bambuseae but then somehow lost the bambusean genome, thus leaving olyrean ploidies the same. But the olyreans passed on the plastid DNA from Bambuseae, creating the supposed illusion that Olyreae is sister with Bambuseae.[7] Figure from [8].
[9], [10] mention a phenomenon of biased diploidization, where ploidy from one parent is biasedly eliminated in an attempt to return to a more stable diploidy, which can potentially be applied to the author’s aforementioned hypothesis.
Sorry to have babbled so much, and sorry more to let me babble more about polyploidy. The question is, why are plants more resistant against polyploidy than animals do? Frankly, this subject is still of academic debate.[11] Possible explanations include polyploidy has less of an impact on plants’ “simpler” structures and that plants are more prone to asexual reproduction and gene duplication, procedures that can facilitate polyploidization. Of course, those proposals have all met scientific rejections.[12] What is true is that plants are probably more easily hybridized due to passive fertilization;[13] polyploid plants also tend to be larger and more resilient,[12] and since plants can’t just run away from unfavorable environments like animals, they kind of have the evolutionary motivation to complicate their genome and diversify their adaptational strategies.[14] Still, this seems somewhat indirect to the author, but we’ll leave it here.


Ok, back to topic. Bambusoideae can be said to be born in a house of crops. Of the two subfamilies Bambusoideae is closest to, Ehrhartoideae includes rice while Pooideae is literally providing half of the staple crops we consume today, containing wheat, barley, and oats, let alone being the most diverse group within Poaceae.[16][17][18] Bambusoideae belonging to this clade no doubt contributes to the miracle of this lineage.


Late Cretaceous, 70 million years ago. The group we know as Poaceae was still in its infant stage. Most likely in what is now South Asia, the two descendants of an insignificant little grass parted ways, henceforth giving rise to two of the most notable subfamilies of grass, Pooideae and Bambusoideae.[18][19]
The first major event to have occurred in bambusoid evolution was during the Eocene 40 million years ago. The stem bamboo group diverged into two lineages, marking the establishment of the crown clade of Bambusoideae. One of the two is our familiar Olyreae, who have retained a herbaceous stature to this day. Due to its early divergence, many botanists consider members of Olyreae to represent the early ancestor of modern woody bamboos.



Fossil records of olyreans are basically none. However, from the genetic evidence and distribution of modern olyreans, we can piece together a vague evolutionary history. Today, the vast majority of olyreans inhabit the tropics of Central and South America, except for one species, Buergersiochloa bambusoides, which is native to the island of New Guinea. Previously, botanists thought that just like the non-native olyrean populations in Africa, Buergersiochloa must have been recently dispersed to New Guinea via sea birds or introduced by humans, until genetic analyses showed that Buergersiochloa is actually the earliest diverging lineage of Olyreae and must have been endemic to New Guinea.[4] This means that Olyreae must have had a much wider distribution than they do now, but throughout the years, they have declined, finally retreating into the Americas at around 14 million years ago, the common ancestor of all modern New World olyreans,[21] leaving the lonely buergersiochloa guarding the last footholds of Olyreae on the Old World…


Caption: Molecular phylogeny of Olyreae; as can be seen, Buergersiochloa is the earliest diverging lineage of Olyreae and is the sister to the rest of its members. Who knows how many stories and history is behind the representative of this lonely family the strata decided not to record? It is indeed a relict of the past. From [5].
(Uh, of course, if you want the atmosphere to be broken, the newest study suggests that Buergersiochloa actually has close relatives in the Americas,[18] which could mean all the theory up there is just plain false… But since this proposal is so recent (2021), it deserves some time to face revisions and contrasting opinions, so we’ll leave it for now.)
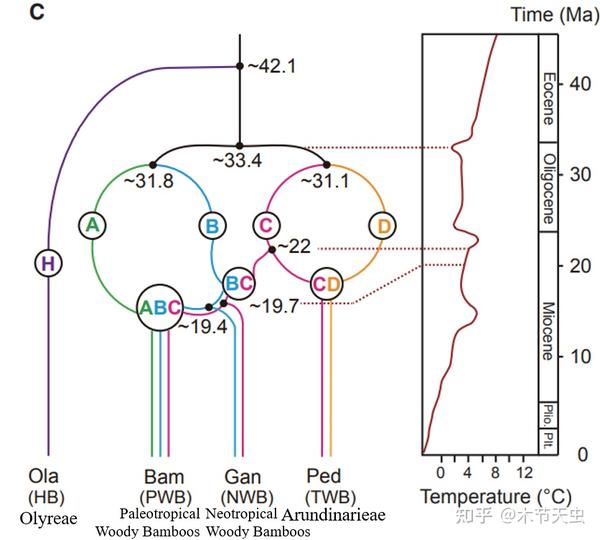
Now let us turn our attention to that other lineage which diverged alongside Olyreae. Prepare to get confused. So apparently, at Oligocene around 31 million years ago, it radiated into four different lineages, which we’ll now call A, B, C, and D. Of them, Lineage C and D hybridized in Miocene to breed the temperate bamboo lineage, known to us as Arundinarieae. This is why arundinarieans have a tetraploid genome rather than the diploid genome of their olyrean relatives.


Arundinarieae, most of the archetypal bamboos we imagine pertaining their evergreen even when covered by snow are members of this tribe. Throughout their millennia of evolution, they have adapted to live in temperate conditions, that is habitats at high latitudes and altitudes, that their tropical ancestors and most of their bambusoid counterparts today are unable to tolerate. (There are still temperate bamboos living in tropical areas though, but just not vice versa) Many innovations are of their patent. For example tessellated, frost-hardy leaves,[22] and a famous one, the evolution of running bamboos.
Woody bamboos can be informally classified into two groups: clumping bamboos, or professionally called pachymorphs, and running bamboos, called leptomorphs.[23] Pachymorphy is supposed to be the plesiomorphy of Bambusoideae, and the ability to “run” allows one bamboo to spread rapidly and invasively, and in theory permits indeterminate extension. Not only do leptomorphs possess an extraordinary ability to occupy available living space, their interconnected rhizomes also form an underground nutrition network, and the distancing between bamboos mean less competition than clumping bamboos, presenting them a significant advantage in competing with pachymorphs.[24][25] There are evidence of bamboos benefiting through deforestation by rapidly occupying those empty lands where they were previously obstructed.[26] (Of course, there can’t be no advantages for pachymorphs, otherwise they would go extinct. For example, their clumplike rhizomes store more water that aid pachymorph growth in drier tropical conditions; they are also better at consolidation, enhancing their survival in sloped soil, and their forcefulness allow them to live in compact soil runners aren’t able to penetrate.[27][28] However, overall, they are at a disadvantage.)


Arundinarieae experienced a rapid diversification around the mid-Miocene. This is probably a result of the intensification of the East Asian Monsoon, bringing a moist climate facilitating expansion and speciation.[24] Another significant event in arundinariean history was during the Pliocene, when a warm period provided suitable conditions for ancestors of Genus Arundinaria to cross the Bering Land Bridge and enter North America, thus extending the territory of the temperate bamboos into the New World and bringing a seemingly alien figure to this faraway continent.[21]


Caption: Distribution map for different groups of Bambusoideae; red represents those of Arundinarieae. As can be seen, in addition to the disparate distribution in North America, there are also patches of arundinariean distribution in Africa. In fact, it is common to find one bamboo group concentrated in one area and turning to find an anomalous species or two making their habitat half a globe away from their kin.[30] It is an actually strange phenomenon since bamboos are poor pioneering species and bamboo seeds are not adapted at all to dispersal by bird or wind. A once-wide coverage until a shrinking of distribution into fragmented habitats is also unreasonable since the disparate species in Arundinarieae aren’t basal.[22] So the explanation should be, as in the above hypothesis of Arundinaria’s expansion to North America, that the species in disparate areas were spread naturally from a heartland, but the intermediate species eventually became extinct, leaving the façade of random distributions all over the world. Map from [31].
On the other hand, Lineage B and C hybridized to form the hybrid lineage BC. The tetraploid Neotropical Woody Bamboos are descended from this line. But before the birth of the ancestor for the Neotropical Woody Bamboos, Lineage BC hybridized with A once again to form hybrid lineage ABC, whose progeny forms the hexaploid Paleotropical Woody Bamboos. Both occurred during the Miocene.
Caption: From [3]. The evolution of Bambusoideae has strong correlation with temperature fluctuations, as indicated at the right of the figure. The temperature variations at the boundary of Eocene and Oligocene led to the major divergences of A, B, C, and D while the temperature variations at the Oligocene-Miocene boundary directly caused the myriad of hybridizations that made the Bambusoideae we know today. This is because temperature fluctuations result in overlapping of habitats, making it more probable for species originally separated in different ecosystems to hybridize. In addition to those, the intensification of the monsoon during the Miocene was also a momentous event for bamboos living in Asia, with the divergence of the paleotropical and neotropical woody bamboos; it also led to significant diversification and expansion for the paleotropical and temperate bamboos, leaving many new groups that persisted to this day.[21][32]
As able to judge from the names, the distribution of Paleotropical Woody Bamboos (abbreviated PWB) is in Africa and Asia in the “Old World” while Neotropical Woody Bamboos (NWB) are distributed in the Americas of the “New World.”
Speaking of here, keen-eyed readers might notice this, aren’t the three tribes of Bambusoideae supposed to be Olyreae, Arundinarieae, and Bambuseae? Where did Bambuseae go? Well, as supposed to the orthodox classification, Bambuseae is a valid tribe, and PWB and NWB are two monophyletic subclades under Bambuseae.[4] However, as elaborated before, those are based on plastid analyses (most likely plastid DNA retained from Lineage B, which is shared exclusively by PWB and NWB), and the author seems unable to find any place for Bambuseae under the hypothesis from nuclear analyses that is adopted here. In fact, there seems to be no taxonomic rules for hybrids whatsoever that justify PWB and NWB should be united by any way, and ancient-hybridized lineages are treated as monophylies. So, does that mean, if we are accepting this hypothesis for bambusoid evolution that PWB and NWB are equal in taxonomic status as Olyreae and Arundinarieae, they too should be treated as tribes? No author has addressed this issue in any way yet, but I have found traces of authors being discontent with the establishment of Bambuseae in works citing the article of this hypothesis. First of all, [3], the original article, doesn’t mention Bambuseae once in the entire paper. Furthermore, articles citing and accepting [3]’s theory, the most iconic of which being [32], also avoid using the clade Bambuseae. Since [32] is a study particularly dealing with the evolution of PWB, it would be implausible for the author to unintentionally not mention Bambuseae. Even studies mentioning Bambuseae, like the study of [10] that is trying to propose a new theory on bamboo evolution, are de facto treating PWB and NWB as separate clades. Therefore, the conclusion that the author has come to is that the academic community has already rejected the validity of Bambuseae, and it is just a matter of time before PWB and NWB are elevated as tribes, and not doing it now is because revolutionary theories like this need time to be digested and accepted? (keep in mind that the author is very unprofessional, and is just trying to come up with a self-consistent line of reasoning)
Anyways, PWB and NWB (crown group at least) most likely originated respectively in Southeast Asia and the Neotropics.[18] Flourishing by the opportunity of the monsoons, PWB diversified with all their might and secured their status in the family of bamboos. By at least Late Miocene, paleotropical bamboos are already inhabiting Eastern and Southern China.



Neotropics, or tropical areas in the New World, that is Central America and South America, the area where the ancestor of neotropical woody bamboos most likely originate and now concentrated in. Sorry to be so brief, but there really isn’t much drama in PWB’s and NWB’s evolutionary processes nor much studies dedicated to them. So I’ll share some pictures.






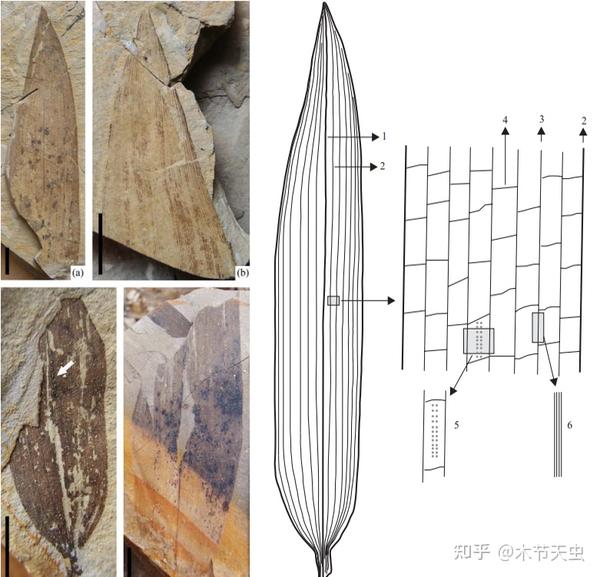
Caption: Incomplete compilation of every bamboo fossil unearthed. Site of excavation is marked on the map, with color of dot representing time period. Note that a lot of bamboo fossils are concentrated in Europe. What’s interesting is that Europe is today one of the only two continents in which bamboos aren’t distributed in, the other being Antarctica. Curious what happened. Don’t believe in that Early Cretaceous one tho, I checked the original paper, and it doesn’t mention anything about bambusoids. From [33].
The process of hybridization does not merely affect the bamboos in the size of their genomes. One of the most characteristic publicities bamboos have is the widespread trivia of “bamboos being a kind of grass.” The fact that creates this surprise is the highly derived lignification taking place in bamboo culm (stem) cells, which is linked to their hybrid history. Today, every grass has genes that code the deposit of cellulose and lignin for basic support, though at a moderate level. However, since bamboos experienced polyploidizations that allowed their genes to double or more, some of their functions are magnified as well, for example the deposit of lignin. With tetraploid to even hexaploid genomes, the deposit of lignin is multiplied to a quantity fair to that of trees. [3][35] And this of course explains why just the polyploid bamboos happen to all be woody while the herbaceous ones are normal diploids. The incredible heights and speeds (growing up to 1m per day) we are familiar today are all but mere sequels of this amazing event.[36]
(Remember my saying before of how I avoided referring to the woody bamboo syndrome as monophyly? Well, this is because according to this theory, woodiness is not a synapomorphy but a result of the polyploidizations, and it is likely that Lineages A, B, C, and D are all herbaceous)


Caption: From [37]. At phase two, large concentrations of hormones like auxin accumulate in shoot apexes to induce fast growth. The extensive and sturdy rhizomes underground secure bamboo’s high and potentially disbalanced culm, and the rigid culm sheaths enveloping young culms can ensure stable growth and penetration. Speaking of culm sheaths, many sources have claimed that they evolve from leaves; however, there are studies concluding that those sheaths are modified from branches in purpose of protecting the inner stem.[38] The second hypothesis might not necessarily be right though, but do keep in mind that the matter is not set in stone.
Another interesting feature of bamboos is their extremely long lifecycle, the most extreme ones flowering once every 120 years.[37] After bamboos flower, they immediately die, the phenomenon of which is known as “monocarpy” (flowering once; as with biology, not all bamboos are like this). The result of these is what are thought to be the most advanced woody bambusoids actually have had much infrequent of a chance to evolve and hone their primitive inflorescence while their olyrean relatives, most of them flowering annually, possess much more specialized reproductive systems than woody bamboos.[39]


Caption: A visual reflection of just how long the generation time of woody bamboos is compared to their relatives. As we know, using molecular mutations to estimate divergences and phylogenies requires knowing the mutation rate, aka generation time of the object organism. Those are usually indicated by phylogenetic branch length. As can be seen here, amidst the long branches of other grasses, the brevity of woody bamboo lengths is incredible. It is also worth noting that the lengths of Arundinarieae are shorter than those of tropical bamboos, probably due to the lower metabolisms and thus lower rates of evolution from living in temperate conditions.[31]


Bamboos are extremely shade-intolerant and have poor dispersal. Their seeds are also vulnerable against the myriad of predators coexisting with them at their habitat. Therefore, evolution has driven them to adopt a very long lifecycle, and invest all their accumulated nutrients at the end of their lives to the production of an enormous number of seeds so that some seeds are bound to escape predation and germinate the next generation. This strategy is known as predator swamping, overwhelming the predators. Monocarpy also maximizes the success rate by dying off and providing the best shade-less lands for their progeny to quickly colonize.[26]
One might have also heard about the gregariousness of bamboo flowering, that is individuals of every bamboo species flowering and dying at the same time. This phenomenon definitely exists for certain species, but it is much less frequent than some might superficially perceive because the “gregariousness” identified by some might just be clusters of running bamboo clones connected by rhizomes that appear to be different individuals but is strictly one organism. They flowering together does not mean that species has gregarious flowering. (of course, even clones might flower at different times for some species. Keep in mind that there are almost always exceptions in biology) There must be the following incentives in order for a species to pursue gregarious flowering: the species distribution cannot be scattered, otherwise predators are only likely to prey on the seeds of one individual and hence no reason for different individuals to collaborate; and whether there are constant fluctuations of climate and conditions, where if so sporadic flowering would be the best strategy to ensure not all seeds die in one catastrophic event.[26]
(As can be seen, gregarious bamboos are extremely vulnerable at the transition to the next generation, and any disturbance during seeding will potentially lead to the extinction of that species. Many bamboos are threatened in this way.[26])
Naturally, the long and unpredictable flowering is a pain in the arse for taxonomists trying to classify bamboos by their inflorescence, geneticists trying to study their hybridization, and botanists trying to understand the fricking mechanisms under their unique flowering cycle.[3][37][41] Yet, despite all those difficulties, respectably they have come up with many results. Today, the most accepted theory explaining gregarious flowering is that it is corely decided by a biological clock, but slight variations in time are synchronized with the modification of shared environmental triggers.[42] The underlying genetic mechanisms for this behavior is still linked with polyploidy, as the genes previously controlling the flowering of grasses are duplicated in bamboos and lead to differentiation of function--- certain transposons found their way into paralogues that cause a delay in the flowering time.[3][37] The actual evolutionary processes are probably something like this: At first, ancestors of one species have the same synchronized flowering time. But then, some old six decides to double or triple his flowering interval, and that results in him having more time to accumulate resources for the production of seeds while still able to share in the benefits of gregarious flowering. As time goes on, the shorter-interval or mis-synced bamboos are outcompeted and eliminated while the process goes all over again for the survivors. Of course, those having the same interval but different synchronizations than the rest of its species’ members technically have equal advantage, so one will either get eliminated by chance or diverge into a different species.[43] On the other hand, all of those are simply in theory extensions from the annual monocarpic lifecycles of their grassy ancestors, so bamboos still die immediately after flowering.[40]



So this is the story of the bamboos. Since their establishment in the Late Cretaceous, they have persisted to this day with species numbering in the thousands and a truly global distribution. They no doubt innovated an unprecedented way of living and thrived in their own niche. And about how their future will turn out? I have great hope for them, but we’ll have to wait and see. Since a group of valiant grasses decided to stand tall amid this world of danger, it is already destined that yet another miracle will be added to this teaming Earth.


Works Cited
[1] COL contributors. “Liliopsida.” Catalogue of Life, http://www.catalogueoflife.org/data/taxon/L2L. Accessed 9 Jan 2023.
[2] Clark, Lynn and Patricia de Oliveira, R. “Diversity and Evolution of the New World Bamboos (Poaceae: Bambusoideae: Bambuseae, Olyreae).” Iowa State University and Universidade Estadual de Feira de Santana, 2018, pp. 1-13. Accessed 9 Jan 2023.
[3] Guo, Zhen-Hua, et al. “Genome Sequences Provide Insights into the Reticulate Origin and Unique Traits of Woody Bamboos.” Molecular Plant, vol. 12, no. 10, 9 May 2019, pp. 1353-1365. Accessed 9 Jan 2023.
[4] Clark, Lynn et al. Bamboo- The Plant and its Uses, Springer, 2015. Accessed 9 Jan 2023.
[5] Wysocki, William et al. “Evolution of the Bamboos (Bambusoideae; Poaceae): A Full Plastome Phylogenomic Analysis.” BMC Evolutionary Biology, vol. 15, no. 15, 18 Mar 2015, pp. 1-12. Accessed 9 Jan 2023.
[6] Dong, Shanshan et al. “Plastid and Nuclear Phylogenomic Incongruences and Biogeographic Implications of Magnolia s.l. (Magnoliceae).” Journal of Systematics and Evolution, vol. 60, no. 1, 15 Jan 2021, pp. 1-15. Accessed 9 Jan 2023.
[7] Wu, Miaoli et al. “The Complete Chloroplast Genome of Guadua augustifolia and Comparative Analyses of Neotropical-Paleotropical Bamboos.” Plos One, vol. 10, no. 12, 2 Dec 2015, pp. 1-19. Accessed 9 Jan 2023.
[8] Twyford, A. and Ennos R. “Next Generation Hybridization and Introgression.” Heredity, vol. 108, no. 3, 7 Sep 2011, pp. 179-189. Accessed 9 Jan 2023.
[9] Xie, Ting et al. "Biased Gene Retention during Diploidization in Brassica Linked to Three-dimensional Genome Organization." Nature Plants, vol. 5, no. 8, 2019, pp. 822-832. Accessed 21 Jan 2023.
[10] Chalopin, Domitille et al. “Integrated Genomic Analyses from Low-depth Sequencing Help Resolve Phylogenetic Incongruence in Bamboos (Poaceae: Bambusoideae).” Frontiers in Plant Science, vol. 12, 3 Sep 2021, pp. 1-18. Accessed 9 Jan 2023.
[11] Miller, Kenneth and Levine, Joseph. Miller & Levine Biology. Pearson Education, Inc, 2014. Accessed 16 Oct 2022.
[12] Mable, Barbara. “‘Why Polyploidy is Rarer in Animals than in Plants:’ Myths and Mechanisms.” Biological Journal of the Linnean Society, vol. 82, no. 4, 9 Aug 2004, pp. 453-466. Accessed 9 Jan 2023.
[13] LeoZSJ. “为什么植物比动物容易杂交?” Zhihu, 7 Oct 2022, http://www.zhihu.com/question/558073342. Accessed 9 Jan 2023.
[14] 生物演化菌. “竹子是如何从草本演化为木本的?” 163, 31 Jul 2019, http://www.163.com/dy/article/ELENGGC305329I9M.html. Accessed 9 Jan 2023.
[15] Hegarty, Matthew and Hiscock, Simon. “Genomic Clues to the Evolutionary Success of Polyploid Plants.” Current Biology, vol. 18, no. 10, 20 May 2008, pp. R435-R444. Accessed 16 Oct 2022.
[16] Prasad, V. et al. “Late Cretaceous Origin of the Rice Tribe Provides Evidence for Early Diversification in Poaceae.” Nature Communications, vol. 2, no. 480, 20 Sep 2011, pp. 1-9. Accessed 9 Jan 2023.
[17] Davis, J. “Systematics of the Grass Family (Poaceae).” Cornell University, 1 Sep 2003. Accessed 9 Jan 2023.
[18] Gallaher, Timothy et al. “Grasses through Space and Time: An Overview of the Biogeographical and Macroevolutionary History of Poaceae.” Journal of Systematics and Evolution, vol. 60, no. 3, 27 Apr 2022, pp. 522-569. Accessed 9 Jan 2023.
[19] Goh, W. et al. “Multi-gene Region Phylogenetic Analyses Suggest Reticulate Evolution and a Clade of Australian Origin among Paleotropical Woody Bamboos (Poaceae: Bambusoideae: Bambuseae).” Plant Systematics and Evolution, vol. 299, no. 1, 6 Nov 2012, pp. 239-257. Accessed 9 Jan 2023.
[20] Silvestrini, Steve. “Lithachne pauciflora.” iNaturalist, 17 Sep 2021, http://www.inaturalist.org/observations/95093668. Accessed 9 Jan 2023.
[21] Burke, Sean et al. “Biogeography and Phylogenomics of New World Bambusoideae (Poaceae), Revisited.” American Journal of Botany, vol. 101, no. 5, 1 May 2014, pp.1-6. Accessed 9 Jan 2023.
[22] Hodkinson, Trevor et al. “Phylogenetic Analyses of Plastid and Nuclear DNA Sequences Indicate a Rapid Late Miocene Radiation of the Temperate Bamboo Tribe Arundinarieae (Poaceae, Bambusoideae).” Plant Ecology and Diversity, vol. 3, no. 2, 13 Sep 2010, pp. 109-120. Accessed 9 Jan 2023.
[23] Hornaday, Fred. “Woody vs Herbaceous Bamboo: Introducing Olyreae.” Bambu Batu, 16 Sep 2020, http://bambubatu.com/woody-vs-herbaceous-bamboo-olyreae/. Accessed 9 Jan 2023.
[24] Cen, Guo et al. “Parallel ddRAD and Genome Skimming Analyses Reveal a Radiative and Reticulate History of the Temperate Bamboos.” Systematic Biology, vol. 70, no. 4, 15 Oct 2020, pp. 756-773. Accessed 9 Jan 2023.
[25] Tachiki, Yuuya et al. “A Spatially Explicit Model for Flowering Time in Bamboos: Long Rhizomes Drive the Evolution of Delayed Flowering.” Journal of Ecology, vol. 103, no. 3, 25 Feb 2015, pp. 585-593. Accessed 9 Jan 2023.
[26] Gadgil, Madhav and Sunkara, Narendra. “Ecological Determinants of Life History Evolution of Two Indian Bamboo Species.” Biotropica, vol. 16, no. 3, 1 Sep 1984, pp. 161-172. Accessed 9 Jan 2023.
[27] Stapleton, Chris. “Form and Function in the Bamboo Rhizome.” Journal of American Bamboo Society, vol. 12, no.1, 28 Nov 1994, pp. 21-29. Accessed 10 Jan 2023.
[28] Palmco editor. “Running Bamboo and Clumping Bamboo.” Palmco, http://www.palmco.com/resources/running-bamboo-and-clumping-bamboo.html. Accessed 10 Jan 2023.
[29] http://staging.joepato.org editor. “New Bylaw Approved Running Bamboo Control.” staging.joepato.org, http://staging.joepato.org/2021/06/new-bylaw-approved-running-bamboo-control/. Accessed 9 Jan 2023. (Error 404-Not found)
[30] Hornaday, Fred. “Where does Bamboo Come from?” Bambu Batu, 18 Nov 2020, http://bambubatu.com/where-does-bamboo-come-from/. Accessed 9 Jan 2023.
[31] Wang, Wencai et al. “Tropical Plants Evolve Faster than their Temperate Relatives: A Case from the Bamboos (Poaceae: Bambusoideae) based on Chloroplast Genome Data.” Biotechnology and Biotechnological Equipment, vol. 34, no. 1, 16 Jun 2020, pp. 482-493. Accessed 9 Jun 2023.
[32] Zhou, Meng-Yuan et al. “Plastid Phylogenomics Shed Light on Intergeneric Relationships and Spatiotemporal Evolutionary History of Melocanninae (Poaceae: Bambusoideae).” Journal of Systematics and Evolution, vol. 60, no. 3, 15 Mar 2020, pp. 640-652. Accessed 9 Jun 2023.
[33] Wang, Qiujun et al. “Bamboo Leaf and Pollen Fossils from the Late Miocene of Eastern Zhejiang, China and their Phytogeological Significance.” Acta Geologica Sinica, vol. 88, no. 4, 28 Aug 2014, pp. 1066-1083. Accessed 9 Jun 2023.
[34] Brea, Mariana et al. “A New Bambusoideae (Poaceae: Bambusoideae: Bambuseae: Guaduinae) from the Ituzaingó Formation (Pliocene-Pleistocene) Entre Ríos, Argentina.” Review of Palaeobotany and Palynology, vol. 192, 23 Jan 2013, pp. 1-9. Accessed 9 Jan 2023.
[35] Que, Feng et al. “Genome-Wide Identification, Expansion, and Evolution Analysis of Homeobox Gene Family Reveals TALE Genes Important for Secondary Cell Wall Biosynthesis in Moso Bamboo (Phyllostachys edulis).” International Journal of Molecular Sciences, vol. 23, no. 8, pp. 1-20. Accessed 9 Jan 2023.
[36] Jin, Guihua et al. “New Genes Interacted with Recent Whole-Genome Duplicates in the Fast Stem Growth of Bamboos.” Molecular Biology and Evolution, vol. 38, no. 12, 28 Sep 2021, pp. 5752-5768. Accessed 9 Jan 2023.
[37] Basak, Mridushree et al. “Genomic Insights into the Growth and Development of Bamboos: What have We Learnt and what more to Discover?” Trees, vol. 35, 21 Aug 2021, pp. 1771-1791. Accessed 9 Jan 2023.
[38] Wang, Shuguang. “Bamboo Sheath--- A Modified Branch Based on the Anatomical Observations.” Scientific Reports, vol. 7, no. 1, 23 Nov 2017, pp. 1-8. Accessed 9 Jan 2023.
[39] Soderstrom, Thomas. “Some Evolutionary Trends in the Bambusoideae (Poaceae).” Annals of the Missouri Botanical Garden, vol. 68, no. 1, 1 Jan 1981, pp. 15-47. Accessed 9 Jan 2023.
[40] 零壹贰012. “幸运的竹子隐藏在最深的山林里——不幸的也是.” 360doc, 3 Jun 2021, http://www.360doc.com/content/21/0603/13/15604356_980270163.shtml. Accessed 9 Jan 2023.
[41] Ganie, Irfan et al. Biotechnological Advances in Bamboo. Springer, 2021. Accessed 9 Jan 2023.
[42] Zheng, Xiao et al. “The Bamboo Flowering Cycle Sheds Light on Flowering Diversity.” Frontiers in Plant Science, vol. 11, 17 Apr 2020, pp. 1-14. Accessed 9 Jan 2023.
[43] Veller, Carl et al. “Extended Flowering Intervals of Bamboos Evolved by Discrete Multiplication.” Ecology Letters, vol. 18, no. 7, 11 May 2015, pp. 653-659. Accessed 9 Jan 2023.
[44] Dreamstime contributors. “424,679 Bamboo Stock Photos, Images & Pictures.” Dreamstime, http://www.dreamstime.com/photos-images/bamboo.html. Accessed 9 Jan 2023.
Cover source:
WallpaperAccess contributors. “Bamboo Wallpapers.” WallpaperAccess, http://wallpaperaccess.com/bamboo. Accessed 9 Jan 2023.
Here is another hypothesis on the evolution of Bambusoideae that is in some ways more holistic and logical than the one adopted here. But since the evolutionary processes are pretty cumbersome and sources citing it are scarce, people who are interested are welcome to explore this hypothesis by themselves.


Basal: indicating a group has diverged from the rest of its clade early on and is a subjective comparison when that group is much less biodiverse than the rest of its relatives. Used for convenience, no species is truly more primitive than another extant species.