
Abstract
Endophytes were investigated for their potential to protect Cinnamomum cassia (cinnamon) from Phytophthora root rot. From healthy cinnamon trees in diseased plantations, 134 bacterial and fungal endophytes were isolated and screened in vitro for their ability to antagonise the root rot pathogens Phytophthora cinnamomi, P. multibullata and P. × vanyenensis. Seventeen endophytes exhibiting high levels of antagonism in vitro (more than 45%) were then tested in a glasshouse study for their ability to reduce the impact of Phytophthora infection in cinnamon seedlings. Trials using cinnamon seeds or seedlings inoculated with an endophyte and then infested with Phytophthora identified three endophytes expressing high levels of disease suppression (based on root damage reduction) and plant growth promotion. These were Penicillium citrinum, Xylaria curta and Clonostachys rosea. These endophytes can potentially be used in the biological control of root rot in cinnamon, but this must be explored further under field conditions.
Similar content being viewed by others


Introduction
Selected endophytic microorganisms may improve plant growth and increase resistance to pathogens. Besides these positive effects, they have the advantage of providing a nonchemical method of disease management with no adverse effects on the environment or human health. Endophytic fungi, bacteria or actinomycetes are associated with many plants. They survive inside a host without causing disease symptoms and can be isolated from surface-disinfected plant tissues (Rosenblueth and Martínez Romero 2006; Hallmann et al. 2011). Their beneficial effect on crop yield (Azevedo et al. 2000) may be through their production of secondary metabolites that promote growth, such as auxin (Sudha et al. 2016), or through the production of fungicidal or bactericidal compounds (Kumar and Kaushik 2012; Soliman et al. 2013; Chatterjee et al. 2019). Their effects may also be mediated through changes in plant anatomy/histology that improve nutrition and increase plant vigour (Eid et al. 2019). Endophytes strongly inhibiting plant pathogens have the potential as biological control agents. They have been used successfully with many plant species, including herbaceous crop species and forest trees (Gunatilaka 2006; Eid et al. 2019). This success has resulted in a recent increase in research efforts to identify endophytes with the ability to reduce the effects of pathogens (Zheng et al. 2017; Abro et al. 2019).
Of particular interest here are that endophytes have been successfully utilised to control Phytophthora diseases in several commercially important species: black pepper, rubber, avocado, cocoa and bell pepper, and have also been found to promote growth in some of these species (Table S1). In avocado trees, two species of fungi (Trichoderma harzianum and Trichoderma hamatum) and two bacteria (Bacillus cereus and Paenibacillus polymyxa) reduced the incidence of root rot disease to ratings of 2–40% in comparison to the positive controls in which disease ratings were 94–100% (Hakizimana et al. 2011). In the woody vine, black pepper, several endophytic Trichoderma species (including Trichoderma harzianum and T. viride) and an Alcaligenes sp. were shown to protect plants in the nursery and the field from infection by P. capsici (Anandaraj and Sarma 1995; Anith and Manomohandas 2001). Additional examples are shown in Table S1.
Products derived from the bark of Cinnamomum cassia are one of the most important exports from Vietnam. However, since early 2017, cinnamon plantations in Vietnam have been affected by the ‘root rot dieback’ of trees (Dang et al. 2017). The roots of the trees rot, and the bark peels off, result in wilt and eventual tree death. Root rot disease is most severe during the rainy season in Vietnam and causes significant economic losses. This disease results from infection by Phytophthora cinnamomi, P. × vanyenensis or P. multibullata (Dang et al. 2021, 2022). Using chemical control measures is undesirable due to the cost of the fungicides and their application, possible negative impacts on the environment, and consumer suspicion of an effect on the quality of the cinnamon bark, which is used as a spice and for medical purposes (Madan and Kannan 2004; Parthasarathy et al. 2008). Alternative control methods need to be considered. Endophytes are an attractive option as there are no toxic residues for the environment or in the harvested plant products. However, there has been no research on managing root rot in cinnamon in Vietnam using endophytes. Although there is no knowledge of the endophytes naturally present in C. cassia, Trichoderma hamatum has been recorded from C. vera (Andrade-Hoyos et al. 2020), Pestalotiopsis spp. from C. camphora (Kharwar et al. 2012) and Muscodor albus from C. zeylanicum (Worapong et al. 2001).
Consequently, the present study examined the range of endophytic species isolated from healthy cinnamon trees. These were tested for their antagonistic activity against Phytophthora cinnamomi, P. × vanyenensis and P. multibullata in vitro and using seedlings in the glasshouse. The goal is to use endophytes as biocontrol agents to treat Phytophthora root rot disease in cinnamon trees in Vietnam.
Materials and methods
Endophyte isolations
Endophytic fungi and bacteria were isolated from healthy tissues (branches and leaves) of cinnamon trees collected from four major cinnamon-producing provinces in Vietnam including the Van Yen district of Yen Bai province, the Tien Yen district of Quang Ninh province, the Thuong Xuan district of Thanh Hoa, and the Tra Bong, Bac Tra My and Nam Tra My districts of Quang Nam provinces. All samples were immediately transferred to the laboratory, and the tissues were screened for endophytes following the modified methods of Weber et al. (2004) and Taylor et al. (2009). Briefly, stems with bark free of visible injuries were surface sterilised by passage through a sequence of full-strength bleach (37% sodium hypochlorite) for 1 min, 70% ethanol for 1 min and six washes for 1 min each in sterile distilled water. The effectiveness of surface sterilisation was tested using the method of Cao et al. (2004). After this treatment, surface sterilisation was checked by immersing samples in 5 mL of sterile water and swirling for 1 min. An aliquot of 0.3 mL was plated on potato dextrose agar (PDA). No growth of microbes was observed after incubation. Surface-sterilised samples were soaked in sterile water and stirred for 1 min. Samples were then dried on sterilised filter paper then cut into 0.5 × 0.5-cm pieces and placed in Petri dishes on half-strength PDA: (BBL, Becton, Dickinson & Co, Sparks MD 21152 USA); (19.5 g PDA (BBL, Becton, Dickinson & Co, USA), 7.5 g agar, 1 L distilled water) with the addition of tetracycline (3 μg mL/L) and streptomycin (10 μg mL/L) for fungi and NB medium (Nutrient Broth -NutriSelect® Basic, Merck) for bacteria. Cultures were kept in an incubator at 25 °C for fungi and 28 °C for bacteria in the dark. Isolations were made on day 2 for bacteria and on day 10 or earlier for fungi (Sayed et al. 2020). Colonies of distinctive bacteria were chosen based on their colour and morphology. After streak plating, single bacterial colonies were picked and stored in 15% glycerol at −20 °C for further experiments. Hyphal tips of actively growing fungi were subcultured, purified and transferred to PDA or oatmeal agar for further characterisation. Single pure cultures were identified morphologically. The remaining mycelium was divided into small agar blocks for future investigations and stored in 10 mL sterile water at 4 °C.
Impact of endophytes against Phytophthora species in dual cultures
An in vitro screening trial was conducted to assess the effect of the endophytes on the growth of the isolates P. cinnamomi (TB5.2, collected from a dead tree in a plantation in Quang Nam), P. × vanyenensis (QD13, from a stream in a plantation with infected trees) and P. multibullata (QD5, from a dead tree in a plantation in Yen Bai). All isolates were virulent in cinnamon (Dang et al. 2022). Each isolate was subcultured on PDA for 7 days. A 5-mm diam mycelial plug of each pathogen was cut out and placed on a fresh PDA plate, 1 cm from the edge. A 5-mm diam mycelium plug of each endophytic fungus or a streak of each endophytic bacterial isolate was placed on the opposite side (~7-cm distance). There were five replicate plates of each endophyte with each Phytophthora species, and five replicate control plates of each Phytophthora species with no endophyte. The plates were incubated in the dark at 25 °C, and radial growth of the Phytophthora was measured after 10 days. Inhibition of Phytophthora growth due to the presence of the endophyte was calculated as the reduction in hyphal radial growth on plates with endophytes compared to the controls with no endophyte, expressed as a percentage of the control growth.
The endophyte species were identified using morphology and analysis of ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′)/ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) (White et al. 1990). The bacterial endophytes were identified from the 16S ribosomal DNA sequence analysis. The polymerase chain reaction was carried out using the universal forward 16SF: 5′-GAGTTTGATCCTGGCTCAG-3′ and reverse 16SR: 5′-GAAAGGAGGTGATCCAGCC-3′ primers (Yuwa-Amornpita 2012). The PCR products were cleaned and sequenced by the First BASE Laboratory Sdn. Bhd., Selangor, Malaysia, using both forward primer ITS1 and reverse primer ITS4. The sequences obtained were edited and blasted using Geneious Prime 2019 (Biomatters Ltd.).
Establishing endophytes in cinnamon by inoculation of seeds or seedlings and assessing plant responses to Phytophthora species
Experimental design
Two methods of inoculating cinnamon with endophytes were tested. For the first method, seeds and the soil in which they germinated were treated with a suspension of the endophyte, and additional irrigation and spraying leaves with endophytes were applied to 1-month-old plants. The second method involved spraying the leaves and irrigating the soil of 6-month-old seedlings with the endophyte suspension.
Based on the in vitro results, five endophytic bacteria and twelve endophytic fungi were tested for their ability to protect cinnamon plants from Phytophthora damage using seedlings in a glasshouse (Table 1). The same experimental design was used to assess the response to exposure to Phytophthora of plants from inoculated seeds or seedlings. The same Phytophthora isolates used in the in vitro trials were used to infect the plants. In each test of each endophyte, there were 40 plants; 10 controls and 10 plants infested with each of the three Phytophthora species. The trial using seed inoculation of endophytes was conducted in February and repeated in October 2021. The trial using endophyte inoculation of seedlings was conducted only in February.
Cinnamon seed inoculation with endophytes and infestation with Phytophthora
Seeds were purchased in February 2021 from the Van Yen district in Yen Bai province. They were planted immediately in the glasshouse at the Vietnamese Academy of Forest Sciences in Ha Noi, Vietnam, as they have a short period of viability. Seeds were surface sterilised by immersing them for 2 min in 0.5% sodium hypochlorite and 2 min in 70% ethanol, followed by rinsing three times in sterile distilled water (Parsa et al. 2013). After the last rinse, 50 μL of the rinse water was dispensed onto PDA plates and incubated at 25 °C for 10 days to assess the success of the sterilisation (Parsa et al. 2013). Seeds were incubated in moist sand that had been irrigated with a solution of a selected endophyte at a concentration of 106 spores/mL of sterile water for bacteria and around 500 μg cells/mL for fungi with 0.05% Tween 80. Once the seeds germinated (usually after 5–7 days), they were transferred to 50 cell trays (each cell 5 × 5 cm), watered with clean potable tap water and fertilised with NPK (16–16–13) (Binh Dien Fertilizer Joint Stock Company). After 1 month, they were inoculated with the endophyte again by irrigating the soil around each seedling with ~5 mL of a spore suspension (106 spores/mL) in sterile water and by spraying the leaves with 5 mL of a spore suspension (concentration 106/mL) in sterile water containing 1% Tween 80. Care was taken to avoid water splashes between treatments. At the time of each inoculation treatment, the spore viability in the suspension was evaluated by placing three, 30 μL droplets onto PDA and incubating in the dark at 28 °C, to check for germination. After 2 months, the inoculated seedlings were transferred to free-draining 10-cm diameter polyurethane pots with a 1:1:1 mix of humus soil:sand:coconut coir, each of which had been dried in the sun for 3 days before use.
Plants were inoculated with the Phytophthora species 75 days after germination. Three grammes of vermiculite/millet inoculum, prepared as described in Dang et al. (2022), was buried to a depth of 5 cm next to the seedling roots. Plant height was measured at this time. Plants were watered daily with deionised water; plant deaths were recorded, and all plants were harvested 60 days after inoculation with Phytophthora (i.e. when they were 135 days old). At harvest, plant height was measured, and root health was assessed using a 1–5 scale (Fig. 1): 1 = healthy roots, 2 = 2–25% roots brown, 3 = 26–50% roots dark brown, 4 = 51–75% roots dark brown and 5 = 76–100% roots dark brown and black, or dead (Belhaj et al. 2018). At harvest, attempts were made to reisolate the endophytes from representative plants and to isolate the Phytophthora species from lesioned roots (see below).

Cinnamon roots showing the range of root damage and the scale used to quantify the observations 1 = healthy roots, 2 = 1–25% damage, 3 = 26–50% damage, 4 = 51–75% damage and 5 = 76–100% damage
Cinnamon seedling inoculation with endophytes and infestation with Phytophthora
Six-month-old cinnamon seedlings in small plastic bags were obtained from a nursery in Van Yen district, Yen Bai Province, and transferred to 10-cm diameter free-draining polyurethane pots in the potting mix described above. After 2 weeks, they were inoculated with the endophytes by spraying ~10 mL of a spore suspension (106 spores/mL in sterile water containing 1% Tween 80 (Hicks et al. 2001) onto the leaves and by pouring 10 mL into a hole near the base of the plant. These inoculation treatments were repeated after two weeks. Spore viability was checked (as above) at the time of each inoculation. One month after inoculating with the endophytes, 3 g of inoculum (as described above) of the appropriate Phytophthora species was placed in a hole near the base of each plant. Plant height was measured at the time of inoculation with Phytophthora.
Plants were watered daily to container capacity with deionised water, and deaths were recorded. Plants were harvested one hundred days after soil infestation with the Phytophthora species. Heights were measured, roots rated for disease damage and reisolation attempts were made for both the endophytes and the Phytophthora species (see below).
Reisolation of endophytes and Phytophthora species from inoculated plants
The presence of endophytes in the seedling roots was examined using four plants for each endophyte from each inoculation method from plants with no Phytophthora, and three plants of each treatment with the Phytophthora species. The method of Rat et al. (2021) (with modifications) was used to detect the endophytes. Briefly, soil particles were removed from the roots with tap water. The roots were then submerged for 5 min in a 70% ethanol solution, before surface sterilisation in a solution of 1.4% NaOCl for 2 min, followed by a rinse with sterile water. The roots were then submerged in 2% sodium thiosulfate for 2 min to counteract the effects of bleach and washed again with sterile water. To evaluate the effectiveness of the sterilisation procedure, the surface of the roots was imprinted onto ½ PDA Petri dishes, which were then incubated in the dark at 25 °C, and roots for which the impression showed microbial development were discarded. Surface-sterilised roots were crushed in 10 mL of sterile phosphate buffer saline (PBS), and a tenfold dilution series was prepared. Finally, the dilutions 10−2–10−7 were triple plated on NB medium, then incubated at 28 °C in the dark for endophytic bacteria.
Similarly, attempts were made to isolate the Phytophthora species from lesioned roots. Lesioned roots were sampled from three plants from each treatment with an endophyte and a Phytophthora species. Roots were sterilised for 1 min in 70% ethanol, followed by three washes in sterile water, blotted dry on sterile filter paper, cut into 1-cm segments and placed onto a Phytophthora-selective medium (NARH).
Data analysis
The data on percentage inhibition in vitro of P. cinnamomi, P. multibullata and P. × vanyenensis were assessed by analysis of variance (ANOVA) and Duncan’s multiple-range test. The package used for analysis was SPSS version 20 for Windows. Shoot growth (plant height at harvest minus plant height at time of Phytophthora inoculation) was assessed for plants inoculated with Phytophthora with or without endophyte application. The data were analysed using the ‘metafor’ package of the R software (Team R C 2017; Viechtbauer 2010).
To check the effect treatments had on controlling root damage caused by Phytophthora, the qualitative data for root damage for each treatment was assessed separately using chi-square tests. All the data analyses were conducted, and bar graphs were created in RStudio 2021.09.1 Build 372 (© 2009-2021 RStudio, PBC).
The results from the two repeat inoculation experiments using seeds were statistically similar and were combined before the effects of the endophytes and Phytophthora were analysed.
Results
Isolation of endophytic fungal and bacterial isolates and testing for inhibition of growth of Phytophthora cinnamomi, P. multibullata and P. × vanyenensis in a dual culture test
The healthy cinnamon trees yielded 110 isolates of endophytic fungi and 24 endophytic bacteria. The data for all isolates screened are given in Table S2. The degree of inhibition was variable for each Phytophthora species. No endophyte gave a very high level of inhibition against all three Phytophthora species, but several were very effective against two species. Endophyte E4, an undescribed species of Penicillium, showed the highest inhibition of P. cinnamomi and P. × vanyenensis but was less effective against P. multibullata. Overall, higher levels of inhibition were observed using the fungal endophytes, and no bacterial endophyte gave more than 57% inhibition against P. multibullata (Table 1, Table S2). The most effective fungal isolates were predominantly Penicillium species, while Bacillus and Burkholderia were the best bacterial species. Seventeen isolates were selected as being highly suppressive of one or more species of Phytophthora (Table 1 and Fig. 2) and were tested for disease inhibition and promotion of plant growth in cinnamon plants.

Inhibition of Phytophthora spp. by fungal and bacteria endophyte isolates after 10 days at 25 °C on ½ PDA. 1. Penicillium citrinum – P. cinnamomi; 2. Penicillium citrinum – P. multibullata; 3. Penicillium fusisporum – P. cinnamomi; 4. Penicillium fusisporum – P. multibullata; 5. Penicillium sp. nov – P. cinnamomi; 6. Penicillium sp. nov – P. multibullata; 7. Penicillium sp. nov – P. × vanyenensis; 8. Penicillium austrosinicum – P. cinnamomi; 9. Penicillium austrosinicum – P. × vanyenensis; 10. Xylaria curta – P. cinnamomi; 11. Xylaria curta – P. multibullata; 12. Clonostachys rosea – P. cinnamomi; 13. Clonostachys rosea – P. × vanyenensis; 14. Achromobacter sp. – P. cinnamomi; 15. Achromobacter sp. – P. × vanyenensis; 16. Bacillus sp. 2 – P. cinnamomi; 17. Bacillus sp. 2 – P. multibullata and 18. Burkholderia gladioli – P. cinnamomi; 19. Burkholderia gladioli – P. × vanyenensis, 20. control – P. × vanyenensis
Effect of inoculation of cinnamon plants with endophytes on plant growth and resistance to Phytophthora
Effect of using seed inoculation of endophytes
Several endophytes significantly increased (P < 0.05) the shoot height of the plants not infected with Phytophthora (Fig. 3). The three fungal isolates showing the most stimulation of cinnamon growth was Penicillium citrinum, Xylaria curta and Clonostachys rosea. Four bacterial isolates also promoted growth: Achromobacter sp., Bacillus sp. 1, Bacillus sp. 2 and Burkholderia gladioli. Although the average growth was higher than the controls with the remaining endophytes, the increase was not significantly different.

Shoot growth following seed inoculation with endophytes. Control: plants with no endophyte. Plants were not inoculated with Phytophthora. Vertical lines indicate SE, and letters indicate a significant difference (α < 0.05)
Forest plots of the comparative shoot height of plants with or without inoculation with a Phytophthora species showed that several endophytes significantly decreased the damage caused by Phytophthora to shoot growth. More of the endophytes effectively antagonised P. cinnamomi (12 endophytes) than P. multibullata (8 endophytes) or P. × vanyenensis (7 endophytes). Seven endophytes (Penicillium citrinum, Penicillium exsudans, Penicillium austrosinicum, Achromobacter sp., Bacillus sp. 1, Bacillus sp. 2 and Burkholderia gladioli) were effective against two Phytophthora species and three (Penicillium aff. multicolor, Xylaria curta and Clonostachys rosea) against all three Phytophthora species (Fig. 4).

Forest plots comparing the shoot growth of plants with or without endophyte inoculation of seeds when plants were inoculated with P. cinnamomi, P. × vanyenensis or P. multibullata. The mean difference and confidence intervals (95%) of the height increment for each endophyte treatment are shown. When the confidence interval of the value reaches or exceeds zero, plants with that endophyte have shown significantly more shoot growth than the control plants. When the confidence intervals do not overlap between endophytes, there is a significant (α < 0.05) difference in plant shoot growth between the different endophytes
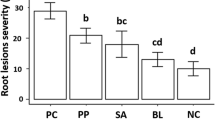
Seed inoculation with endophytes did not result in any differences in root damage of plants not inoculated with a Phytophthora species (Fig. 5). There was significantly (P < 0.05) more root damage after inoculation with the different Phytophthora species than in the non-inoculated plants regardless of which endophyte had been applied (according to the chi-square test) (Fig. 5). However, 12 of the endophytes significantly reduced root damage after inoculation with P. cinnamomi (Fig. 5a), 11 against P. multibullata (Fig. 5b) and 4 against P. × vanyenensis (Fig. 5c). Four of the endophytes (Penicillium citrinum, Penicillium sp. nov, Xylaria curta and Clonostachys rosea) reduced root damage from all three Phytophthora species.

Means of root damage scores of plants inoculated with endophytes as seeds, and the control with no endophyte, when plants were inoculated with a P. cinnamomi, b P. × vanyenensis or c P. multibullata (black bars) compared with control plants with no Phytophthora (grey bars). Vertical lines indicate SE, and letters indicate significant (α < 0.05) differences between means for inoculated plants based on Duncan’s multiple-range tests
Effect of using seedling inoculation of endophytes
Inoculation of seedlings with endophytes identified four (Penicillium citrinum, Penicillium exsudans, Xylaria curta and Clonostachys rosea) that significantly increased shoot height compared to the control (in the absence of Phytophthora) (Fig. 6). However, the magnitude of the increase was less than that those observed using seed inoculation. The variability in plant growth rate within treatments was higher than when the endophytes were applied to seeds.

The effect on shoot growth of seedling inoculation with endophytes. Control: plants with no endophyte. Plants were not inoculated with Phytophthora. Vertical lines indicate SE, and letters indicate a significant (α < 0.05) difference
Forest plots of the comparative shoot height of plants with or without an endophyte and inoculated with a Phytophthora showed that 12 endophytes reduced seedling growth by P. cinnamomi, 11 were effective against P. × vanyenensis and 13 against P. multibullata. Eight endophytes (Penicillium citrinum, Penicillium aff. multicolour, Penicillium austrosinicum, Penicillium paxillin, Xylaria curta, Clonostachys rosea, Bacillus sp. 1 and Bacillus sp. 2) were effective against all three Phytophthora species (Fig. 7).

Forest plots comparing the shoot growth of plants with or without endophyte inoculation of seedlings when plants were infected with either P. cinnamomi, P. × vanyenensis or P. multibullata. The mean difference and confidence intervals (95%) of the height increment for each endophyte treatment are shown. When the confidence interval of the value reaches or exceeds the zero, plants with that endophyte have shown significantly more shoot growth than the control plants. When the confidence intervals do not overlap between endophytes, there is a significant (P < 0.05) difference in plant shoot growth with the different endophytes
Seedling inoculation with endophytes did not result in any differences in root damage of plants not inoculated with a Phytophthora (Fig. 8). The presence of four endophytes (Penicillium citrinum, Penicillium paxillin, Xylaria curta and Clonostachys rosea) resulted in reduced root damage from P. cinnamomi, five (Penicillium citrinum, Penicillium paxillin, Penicillium sclerotiorum, Xylaria curta and Clonostachys rosea) reduced damage from P. multibullata and three (Penicillium citrinum, Xylaria curta and Clonostachys rosea) reduced damage from P. × vanyenensis (Fig. 8).

Means (and SEs shown by vertical lines above bars) of root damage scores of plants inoculated with endophytes as 6-month-old seedlings and the control with no endophyte when plants were inoculated with a P. cinnamomi, b P. × vanyenensis or c P. multibullata (black bars) compared with control plants with no Phytophthora (grey bars). Significant (α < 0.05) differences between means for controls and plants inoculated
A comparison of the results of the two inoculation methods shows that the endophytes that gave the best shoot height results and reduced root damage in both seed and seedling inoculations were Penicillium citrinum, Xylaria curta and Clonostachys rosea.
Reisolation of endophytes and Phytophthora species from inoculated plants
All endophyte isolates were reisolated from at least one plant of the control seedlings not inoculated with Phytophthora. In the plants inoculated with P. cinnamomi or P. multibullata, isolate E14 a Bacillus species was not recovered, but it was present in plants inoculated with P. × vanyenensis (Table 2). Eighty-four percent of control plants were successfully inoculated, and the percentage was slightly lower (70.6–78.4%) in plants inoculated with Phytophthora. The percentage of endophytes recovered from roots was 70.6% for P cinnamomi, 74.5% for P. multibullata and 78.4% for P. × vanyenensis (Table 2). The percentage of Phytophthora recovered from lesioned roots was 82.4% for P. cinnamomi, 72.5% for P. multibullata and 76.5% for P. × vanyenensis (Table 3).
Correlations between the ability of endophytes to inhibit Phytophthora in vitro and in planta
There was a negative correlation between the assessment of endophyte inhibition of P. cinnamomi, P. multibullata and P. × vanyenensis through dual in vitro cultures and the assessment of damage root ratings of seedlings inoculated either as seeds or seedlings (Fig. 9). There was a positive correlation between the root damage assessments for the different endophytes using the seed or seedling inoculation techniques (data from Table 1 and Fig. 9a–i).

Correlation between the percentage root damage caused by P. cinnamomi (a–c), P. multibullata (d–f) and P. × vanyenensis (g–i) on plants inoculated with endophytes as seeds (a, d, g) or seedlings (b, e, h) with percentage inhibition in vitro
Discussion
Healthy cinnamon trees were found to have a wide array of fungal and bacterial endophytes, including species that significantly inhibited in vitro growth of one or more of the Phytophthora species tested. Seventeen endophytes with high levels of inhibition to the Phytophthora species in vitro were tested in glasshouse trials using either seed or seedling inoculation with the endophytes. Inoculation of either seeds or seedlings was effective for disease control. Three fungal endophytes, Penicillium citrinum, Xylaria curta and Clonostachys rosea, reduced root and shoot damage after inoculation with P. cinnamomi, P. multibullata or P. × vanyenensis.
This is the first study to consider endophytes of C. cassia and their ability to suppress Phytophthora species. However, several bacterial and fungal endophytes are effective against P. cinnamomi in a range of woody and herbaceous species, including one study of endophytes of C. camphora against three Phytophthora species (Kharwar et al. 2012) (Table S1). More fungal species than bacteria were found effective against Phytophthora in vitro, and the most effective fungi were Penicillium (Pe) species (Tables 2 and S2). This is the first record of Penicillium austrosinicum, P. fusisporum, P. citrinum, P. aff. multicolour, P. paxillin and P. sclerotiorum inhibiting a Phytophthora species. However, in other studies, P. striatisporum, P. funiculosum and P. oxalicum are effective inhibitors of several plant pathogens, including Phytophthora spp. (Fang and Tsao 1995; Ma et al. 2008; Dam et al. 2018). Penicillium citrinum resulted in one of the highest levels of disease suppression in the seedling trial. As a rhizosphere organism, it is effective against several fungal pathogens in various crops (Sharma et al. 2021). Penicillium species are particularly effective against oomycete pathogens in the soil as they produce surfactants that lyse the zoospores (Stanghellini and Miller, 1997; Bruisson et al. 2019). In the present study, this would be an effective mode of action against Phytophthora in the rhizosphere, but it is difficult to see how an impact on zoospores would make it an effective endophyte. Further studies are required to determine the mechanisms of action.
The other suppressive endophytic fungi from C. cassia included Cladosporium cladosporioides, Xylaria curta and Clonostachys rosea. Clonostachys rosea has also been isolated from Theobroma cacao and as an endophyte shown to inhibit P. palmivora in that host (Mejía et al. 2008). Endophytic Xylaria spp. are known to inhibit some fungal species and, as endophytes, promote plant growth (Adnan et al. 2018; Elias et al. 2018; Bills and Polishook 1992; Rodrigues 1993). Its effect in cinnamon observed here is the first record of it being effective against Phytophthora. Cladosporium cladosporioides also has not been previously identified as effective against Phytophthora. No endophytic Trichoderma species was particularly effective in vitro against the Phytophthora species used here (Table S2), but several species are known to suppress Phytophthora species (Table S1).
The most effective bacterial endophytes isolated included an Achromobacter sp., two unknown Bacillus spp. and Burkholderia gladioli. Several endophytic Bacillus spp. suppress Phytophthora (Table S1) and promote plant growth (Broadbent et al. 1971; Merriman et al. 1974; Caesar and Burr 1987; Backman et al. 1994; Yobo 2000). Bacillus spp. have also been shown to be antagonistic to P. parasitica (Abraham 2005), P. capsici and a Phytophthora sp. (Chung et al. 2008), P. cactorum (Utkhede 1984) and P. infestans (Orfila and Knox 2000). Burkholderia gladioli is known to be effective against Pythium ultimum (Bae et al. 2007), and P. infestans (An et al. 2010), as is Burkholderia cepacia (Yan et al. 2002). The ability of the Achromobacter sp. from cinnamon to inhibit Phytophthora mycelium growth appears to be the first record.
Although a high percentage of the inoculated plants contained the introduced endophyte in their roots, relatively few plants (30%) were assessed after inoculation. More extensive tests are required to accurately determine the success rate of the two inoculation techniques for each isolate and whether a high level of success could be obtained using fewer steps in the inoculation protocol. The correlation between the inhibition of Phytophthora mycelial growth in vitro and the ability of an endophyte to reduce root damage in infected plants indicates that a preliminary screening of isolates in vitro is an effective way of selecting those most likely to protect the plant from the pathogen. The positive correlation between the root damage observed in plants inoculated with endophytes as seeds or seedlings suggests that the techniques are equally effective.
As this is the first study of endophytic diversity in C. cassia, there are reasonable prospects of finding further unique and interesting endophytes in this and other Cinnamomum species. Under greenhouse conditions, the endophytes isolated from the cinnamon successfully inhibited P. cinnamomi, P. multibullata and P. × vanyenensis. Further experiments are needed to determine whether inoculation of seedlings results in beneficial effects in older trees in the field and if inoculation with multiple endophytes will suppress more species of Phytophthora or result in a higher level of control. Another aspect worthy of further investigation is whether endophyte inoculation is cost-effective, even in the absence of Phytophthora, because of their promotion of plant growth.
This research has shown that endophytes from healthy cinnamon trees in diseased areas have significant potential as biocontrol agents against root rot. More endophytes showed suppression of Phytophthora species in vitro than in planta. While several bacterial endophytes showed promise in vitro, none was as effective as the fungal isolates in planta. In the glasshouse trials, no endophyte gave a very high level of inhibition against all three Phytophthora species. However, several were very effective against two, the best being an undescribed Penicillium species (endophyte Penicillium sp. nov.) which showed the highest inhibition of P. cinnamomi and P. × vanyenensis but was less effective against P. multibullata. The results reported here are an encouraging step towards using endophytes as biocontrol agents to treat Phytophthora root rot disease, reduce the use of chemical fungicides and enable farmers to move towards the production of organically grown cinnamon.
Data availability
All sequences have been uploaded to GenBank.
Code availability
Not applicable.
References
Abraham A, Narayanan S, Philip S, Nair D, Chandrasekharan A, Kochupurackal J (2013a) In silico characterization of a novel β-1,3-glucanase gene from Bacillus amyloliquefaciens–a bacterial endophyte of Hevea brasiliensis antagonistic to Phytophthora meadii. J Mol Model 19:999–1007. https://doi.org/10.1007/s00894-012-1645-3
Abraham A, Philip S, Kuruvilla Jacob C, Jayachandran K (2013b) Novel bacterial endophytes from Hevea brasiliensis as biocontrol agent against Phytophthora leaf fall disease. J Int Org Biol Cont 58:675–684. https://doi.org/10.1007/s10526-013-9516-0
Abraham AO (2005) Biological control of Phytophthora root rot of citrus seedlings and cuttings. Dissertation University of KwaZulu-Natal - Pietermaritzburg South Africa
Abro M, Sun X, Li X, Jatoi G, Guo L (2019) Biocontrol potential of fungal endophytes against Fusarium oxysporum f. sp. cucumerinum causing wilt in cucumber. Plant Pathol J 35:598–608. https://doi.org/10.5423/PPJ.OA.05.2019.0129
Adnan M, Alshammari E, Ashraf SA, Patel K, Lad K, Patel M (2018) Physiological and molecular characterization of biosurfactant producing endophytic fungi Xylaria regalis from the cones of Thuja plicata as a potent plant growth promoter with its potential application. Biomed Res Int 2018:11. https://doi.org/10.1155/2018/7362148
Anandaraj M, Sarma Y (1995) Diseases of black pepper (Piper nigrum L.) and their management. J Spices Arom Crops 4:17–23
Andrade-Hoyos P, Silva-Rojas HV, Romero-Arenas O (2020) Endophytic Trichoderma species isolated from Persea americana and Cinnamomum verum roots reduce symptoms caused by Phytophthora cinnamomi in avocado. Plants 9:1220. https://doi.org/10.3390/plants9091220
Anith K, Manomohandas T (2001) Combined application of Trichoderma harzianum and Alcaligenes sp. strain AMB 8 for controlling nursery rot disease of black pepper. Indian Phytopath 54:335–339
Aravind R, Kumar A, Eapen S (2012) Pre-plant bacterisation: a strategy for delivery of beneficial endophytic bacteria and production of disease-free plantlets of black pepper (Piper nigrum L.). Arch Phytopath Plant Protect 45: 1115–1126. https://doi.org/10.1080/03235408.2012.657894
Azevedo J, Maccheroni W, Pereira J, Araújo W (2000) Endophytic microorganisms: a review on insect control and recent advances on tropical plants. Electronic J Biotech 3:41–65
Backman P, Brannen PM, Mahaffee WF (1994) Plant response and disease control following seed inoculation with Bacillus subtilis. Improving plant productivity with rhizosphere bacteria. CSIRO Division of Soil, Adelaide, Australia, pp 3–9
Belhaj R, McComb JA, Burgess T, Hardy GSJ (2018) Pathogenicity of 21 newly described Phytophthora species against seven Western Australian native plant species. Plant Pathol 67:1140–1149. https://doi.org/10.1111/ppa.12827
Bills GF, Polishook J (1992) Recovery of endophytic fungi from Chamaecyparis thyroides. Sydowia 44:1–12
Broadbent P, Baker K, Waterworth Y (1971) Bacteria and actinomycetes antogonistic to fungal root pathogens in Australian soils. Aust J Biol Sci 24:925–944
Caesar A, Burr T (1987) Growth promotion of apple seedlings and rootstocks by specific strains of bacteria. Phytopathology 77:1583–1588
Cao L, Qiu Z, You J, Tan H, Zhou S (2004) Isolation and characterization of endophytic Streptomyces strains from surface-sterilized tomato (Lycopersicon esculentum) roots. Lett Appl Microbiol 39:425–430. https://doi.org/10.1111/j.1472-765X.2004.01606.x
Chatterjee S, Ghosh R, Mandal N (2019) Production of bioactive compounds with bactericidal and antioxidant potential by endophytic fungus Alternaria alternata AE1 isolated from Azadirachta indica A. Juss. PLoS ONE 14: e0214744 https://doi.org/10.1371/journal.pone.0214744
Chung S, Kong H, Buyer JS, Lakshman DK, Lydon J, Kim SD, Roberts DP (2008) Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soilborne pathogens of cucumber and pepper. Appl Microbiol Biot 80:115–123. https://doi.org/10.1007/s00253-008-1520-4
Dam TV, Pham TQ, Dang QN, Nguyen CM (2018) Endophytes and using for management of Phytophthora leaf fall disease of rubber tree plantations in south east Vietnam. Sci Tech J Agric Rural Dev 22:25–32
Dang QN, Pham NQ, Pham TQ, Nguyen HM (2017) First report of Phytophthora cinnamomi on Cinnamomum cassia in Vietnam. In: Phytophthora in Forests and Natural Ecosystems, Section 8: Pathogenicity, IUFRO Working Party S07-02-09 Sapa, Vietnam, pp 40
Dang QN, Pham TQ, Arentz F, Hardy GESJ, Burgess TI (2021) New Phytophthora species in clade 2a from the Asia-Pacific region including a re-examination of P. colocasiae and P. meadii. Mycol Prog 20:111–129. https://doi.org/10.1007/s11557-020-01656-7
Dang QN, Burgess TI, McComb J, Pham TQ, Le BV, Nguyen TH, Le XT, Hardy GESJ (2022) Root rot pathogens of Cinnamomum cassia in Vietnam. Plant Pathol 71:1969–1979. https://doi.org/10.1111/ppa.13617
de Vries S, von Dahlen J, Schnake A, Ginschel S, Schulz B, Rose L (2018) Broad-spectrum inhibition of Phytophthora infestans by fungal endophytes. FEMS Microl Ecol 94(4) fiy037. https://doi.org/10.1093/femsec/fiy037
Eid A, Salim S, Hassan S, Ismail M, Fouda A (2019) Role of endophytes in plant health and abiotic stress management. In Microbiome in plant health and disease, pp 119–144. Springer. https://doi.org/10.1007/978-981-13-8495-0_6
Elias LM, Fortkamp D, Sartori SB, Ferreira MC, Gomes LH, Azevedo JL et al (2018) The potential of compounds isolated from Xylaria spp. as antifungal agents against anthracnose. Braz J Microbiol 49:840–847. https://doi.org/10.1016/j.bjm.2018.03.003
Fang J, Tsao P (1995) Efficacy of Penicillium funiculosum as a biological control agent against Phytophthora root rots olf azalea and citrus. Phytopathology 85:871–878
García-Latorre C, Rodrigo S, Santamaria O (2022) Protective effects of filtrates and extracts from fungal endophytes on Phytophthora cinnamomi in Lupinus luteus. Plants 11:1455. https://doi.org/10.3390/plants11111455
Gunatilaka A (2006) Natural products from plant associated microorganisms: distribution, structural diversity, bioactivity, and implications of their occurrence. J Nat Prod 69:509–526. https://doi.org/10.1021/np058128n
Hakizimana J, Gryzenhout M, Coutinho T, Van den Berg N (2011) Endophytic diversity in Persea americana (avocado) trees and their ability to display biocontrol activity against Phytophthora cinnamomi. Proceedings VII World Avocado Congress, Cairns, Australia
Hallmann J, Von Quadt A, Mahaffee W, Kloepper J (2011) Endophytic bacteria in agricultural crops. Can J Microbiol 43:895–914. https://doi.org/10.1139/m97-13
Hanada RE, de Jorge ST, Pomella AW, Hebbar KP, Pereira JO, Ismaiel A, Samuels GJ (2008) Trichoderma martiale sp. nov., a new endophyte from sapwood of Theobroma cacao with a potential for biological control. Mycol Res 112:1335–1343. https://doi.org/10.1016/j.mycres.2008.06.022
Hicks BJ, Watt AD, Cosens D (2001) The potential of Beauveria bassiana (Hyphomycetes: Moniliales) as a biological control agent against the pine beauty moth, Panolis flammea (Lepidoptera: Noctuidae). For Ecol Manag 149:275–281. https://doi.org/10.1016/S0378-1127(00)00561-2
Hong CE, Kwon SY, Park JM (2016) Biocontrol activity of Paenibacillus polymyxa AC-1 against Pseudomonas syringae and its interaction with Arabidopsis thaliana. Microbiol Res 185:13–21. https://doi.org/10.1016/j.micres.2016.01.004
Irabor A, Mmbaga M (2017) Evaluation of selected bacterial endophytes for biocontrol potential against Phytophthora blight of bell pepper (Capsicum annuum L.). J Plant Pathol Microbiol 8:31–34. https://doi.org/10.4172/2157-7471.1000424
Kharwar R, Maurya A, Verma V, Kumar A, Gond S, Mishra A (2012) Diversity and antimicrobial activity of endophytic fungal community isolated from medicinal plant Cinnamomum camphora. Proc Nat Acad Sci India Section B Biol Sci 82:557–565. https://doi.org/10.1007/s40011-012-0063-8
Kollakkodan N, Anith K, Nysanth N (2020) Endophytic bacteria from Piper colubrinum suppress Phytophthora capsici infection in black pepper (Piper nigrum L.) and improve plant growth in the nursery. Arch Phytopathol Pl Protection 54: 86–108. https://doi.org/10.1080/03235408.2020.1818493
Kumar S, Kaushik N (2012) Metabolites of endophytic fungi as novel source of biofungicide: a review. Phytochem Rev 11:507–522. https://doi.org/10.1007/s11101-013-9271-y
Ma Y, Chang ZZ, Zhao JT, Zhou MG (2008) Antifungal activity of Penicillium striatisporum Pst10 and its biocontrol effect on Phytophthora root rot of chilli pepper. Biol Control 44:24–31. https://doi.org/10.1016/j.biocontrol.2007.10.005
Madan M, Kannan S (2004) Economics and marketing of Cinnamon and Cassia–a global view. In: Ravindran PN, Babu KN (eds) Cinnamon and cassia: the genus Cinnamomum. CRC Press, pp 301–326
Masanto Arif W, Tri J, Siti S, Koji K (2020) Antagonistic potential of endophytic bacteria against Phytophthora palmivora causing black pod rot disease on cacao (Theobroma cacao L.) in Indonesia. Plant Pathol 19:22–41. https://doi.org/10.3923/ppj.2020.22.41
Mejía L, Rojas E, Maynard Z, Bael S, Arnold A, Hebbar P, Samuels GJ, Robbins N, Herre E (2008) Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol Control 46:4–14. https://doi.org/10.1016/j.biocontrol.2008.01.012
Merriman P, Price R, Kollmorgen J, Piggott T, Ridge E (1974) Effect of seed inoculation with Bacillus subtilis and Streptomyces griseus on the growth of cereals and carrots. Aust J Agric Res 25:219–226. https://doi.org/10.1071/AR9740219
Mitchell AM, Strobel GA, Moore E, Robison R, Sears J (2009) Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 156:270–277. https://doi.org/10.1099/mic.0.032540-0
Mmbaga M, Gurung S, Maheshwari A (2018) Screening of plant endophytes as biological control agents against root rot pathogens of pepper (Capsicum annum l.). J Pl Pathol Microbiol 9(3). https://doi.org/10.4172/2157-7471.1000435
Ngo V A, Wang SL, Nguyen VB, Doan CT, Tran TN, Tran DM, Tran TD Nguyen AD (2020) Phytophthora antagonism of endophytic bacteria isolated from roots of black pepper (Piper nigrum L.). Agronomy 10:286. https://doi.org/10.3390/agronomy10020286
Orfila CC, Knox JP (2000) Spatial regulation of pectic polysaccharides in relation to pit fields in cell walls of tomato fruit pericarp. Plant Physiol 122:775–782. https://doi.org/10.1104/pp.122.3.775
Parsa S, Ortiz V, Vega F (2013) Establishing fungal entomopathogens as endophytes: towards endophytic biological control. J Visual Exp74:e50360
Parthasarathy VA, Chempakam B, Zachariah TJ (2008) Chemistry of spices. CABI, Oxfordshire, pp 405–407
Rat A, Naranjo HD, Krigas N, Grigoriadou K, Maloupa E, Alonso AV et al (2021) Endophytic bacteria from the roots of the medicinal plant Alkanna tinctoria Tausch (Boraginaceae): exploration of plant growth promoting properties and potential role in the production of plant secondary metabolites. Front Microbiol 12:1–14. https://doi.org/10.3389/fmicb.2021.633488
Rodrigues K (1993) Endophytic species of Xylaria: cultural and isozymic studies. Sydowia 45:116–138
Rosenblueth M, Martínez Romero E (2006) Bacterial endophytes and their interactions with hosts. Mol Plant Microbe În 19:827–837. https://doi.org/10.1094/MPMI-19-0827
Sharma H, Rai AK Chettri R, Nigam PS (2021) Bioactivites of Penicillium citrinum isolated from a medicinal plant Swertia chirayita. Arch Microbiol 203:5173–5182. https://doi.org/10.1007/s00203-021-02498-x
Simamora AV, Hahuly MV, Henuk JB (2021) Endophytic fungi as potential biocontrol agents of Phytophthora palmivora in the cocoa plant. Biodiversitas J Biol Div 22(5). https://doi.org/10.13057/biodiv/d220519
Soliman S, Trobacher C, Tsao R, Greenwood J, Raizada M (2013) A fungal endophyte induces transcription of genes encoding a redundant fungicide pathway in its host plant. BMC Plant Biol 13:1–10. https://doi.org/10.1186/1471-2229-13-93
Sreeja K, Anandaraj M, Bhai R (2016) In vitro evaluation of fungal endophytes of black pepper against Phytophthora capsici and Radopholus similis. J Spices Arom Crops 25:113–122
Sudha V, Govindaraj R, Baskar K, Al Dhabi NA, Duraipandiyan V (2016) Biological properties of endophytic fungi. Brazil Arch Biol Techn 59. https://doi.org/10.13057/biodiv/d220519
Taylor K, Barber PA, Hardy G, Burgess T (2009) Botryosphaeriaceae from tuart (Eucalyptus gomphocephala) woodland, including descriptions of four new species. Mycol Res 113:337–353. https://doi.org/10.1016/j.mycres.2008.11.010
Team R C (2017) R: a language and environment for statistical computing. R Foundation for statistical computing. www.R-project.org. Accessed 31 May 2022
Utkhede R (1984) Antagonism of isolates of Bacillus subtilis to Phytophthora cactorum. Can J Bot 62:1032–1035
Viechtbauer W (2010) Conducting meta-analyses in R with the metafor package. J Stat Software 36:1–48
Weber RW, Stenger E, Meffert A, Matthias H (2004) Brefeldin a production by Phoma medicaginis in dead pre-colonized plant tissue: a strategy for habitat conquest? Mycol Res 108:662–671. https://doi.org/10.1017/S0953756204000243P
Worapong J, Strobel EJ, Ford JY, Li JY, Baird G Hess WM (2001) Muscador albus gen. et sp. nov. an endophyte from Cinnamomum zelandicum. Mycotaxon 79: 67–79
White T, Bruns T, Lee S, Taylor J, Innis M, Gelfand D, Sninsky J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innes MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic Press, San Diego, pp 315–322
Yan Z, Reddy M, Ryu CM, McInroy JA, Wilson M, Kloepper JW (2002) Induced systemic protection against tomato late blight elicited by plant growth-promoting rhizobacteria. Phytopathology 92:1329–1333
Yobo KS (2000) Utilization of Bacillus spp. as plant probiotics. Dissertation University of Natal. Pietermaritzburg, Republic of South Africa
Yuwa-Amornpita T (2012) Bio-hydrogen production from biodiesel glycerol waste from used oil by bacterium isolated from waste water sludge. J Environ Sci Technol 5:373–380
Zheng K, Miao CP, Chen HH, Huang FF, Xia YM, Chen YW, Zhao LX (2017) Endophytic fungi harbored in Panax notoginseng: diversity and potential as biological control agents against host plant pathogens of root-rot disease. J Ginseng Res 41:353–360. https://doi.org/10.1016/j.jgr.2016.07.005
Zouaoui M, Essghaier B, Weslati M, Smiri M, Hajlaou M, Sadfi-Zouaoui N (2019) Biological control of clementine branch canker, caused by Phytophthora citrophthora. Phytopathol Mediterr 58:547–558. https://doi.org/10.14601/Phyto-10754
Acknowledgements
The authors would like to thank the staff from the Forest Protection Research Centre of Vietnam Academy of Forest Sciences including Hoai Thu, Nhat Tan, A Tong and Minh Hang who helped collect data. We express appreciation for advice on statistical analyses from Hoa Duong and Ha Do.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions Resources and funding were provided by Murdoch University and the Ministry of Agriculture and Rural Development of Vietnam (MARD) of Vietnam. The senior author received a PhD scholarship jointly funded by the Vietnam Government and Murdoch University.
Author information
Authors and Affiliations
Contributions
Design: Q. D., T. B., T. P, G. H. and J. M.; field, glasshouse and laboratory work: Q. D., B. L., T. T. and L. N.; data analysis: Q.D, B. L.; preparation of manuscript: Q.D.; review and editing: Q. D., T. B., J. M and G. H. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Section Editor: Tanay Bose
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the “Topical collection - Since de Bary: Progress in Phytophthora research”.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dang, Q.N., Burgess, T.I., McComb, J. et al. Fungal and bacterial endophytes antagonistic to Phytophthora species causing root rot in Cinnamomum cassia. Mycol Progress 22, 28 (2023). https://doi.org/10.1007/s11557-023-01878-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11557-023-01878-5