
This species has a filamentous gametophyte that alternates with a club-shaped zygote (i.e. haplontic life history). Both of these phases contribute to a dark green band in the upper to mid intertidal on rock, the gametophyte typically in winter to early summer (Image A) the zygote obvious later in summer and into autumn (Image B). The gametophytic filaments reach 8 cm in length, and are anchored by a modified basal rhizoidal cell (Images C & D), which can be augmented by 1 to ~4 descending intramatrical (i.e. produced within the “matrix” or cell walls and surrounding cuticle) rhizoids from cells near the base (Image E). The vegetative cells near the base are squat to square (occasionally rectangular; Image D) in outline and 28-34 µm wide by 18-25(33) µm long and contain a parietal chloroplast with 2-3 pyrenoids (Image F). Cells mid filament are also typically squat to disc-shaped, 42-46 µm wide by 14-34 µm long, with a parietal chloroplast containing multiple pyrenoids (Image G). Upper filament cells are typically squat, 52-56 µm wide by 23-35 µm long, again with a parietal chloroplast and multiple pyrenoids (Image H). During presumptive gametangial production, upper cells take on a barrel shape, 47-50 µm wide by 34-40 µm long, with bulging walls (Image I). The zygotes (“Codiolum pusillum” stage) can be 2-2.5 mm in height with a long narrow stalk and a terminal cytoplasmic region that broadens toward the tip, 95-105 µm wide by 450-730 µm tall (Image J), and anchored by a small attachment pad (Image K).
We have genetically verified records from upper to mid intertidal on open rock, as well as in areas of seepage and damp depressions, where this species forms a dark green band along with other filamentous green algae. Records extend from ME to the Maritime Provinces plus the Gulf of St. Lawrence coasts of NL and QC, but this species is reportedly much more widely distributed in the NW Atlantic flora. In addition, we have verified records for this species from Kamchatka and Nome AK, which is consistent with reports of this species in BC but genetically verified records are wanting (likely due to low sampling). However, the possibility that the BC records represent a different species needs to be explored with molecular data (e.g., Urospora sp. 1BC and Urospora sp. 2Nome).
This species, as well as most others in our flora assigned to Urospora, do not join the type species Urospora penicilliformis (Roth) Areschoug in preliminary phylogenetic analyses and taxonomic study is needed. Filaments of Uroposra neglecta are stiffer than those of Ulothrix flacca (Dillwyn)Thuret, and lack the band-shaped plastid with 1-2 pyrenoids as seen in the latter. The filaments of Urospora penicilliformis (Roth) Areschoug are stiffer with the overall dimensions greater.
It is common for what otherwise looks like a unialgal carpet on rock, or mat of green filaments in an estuary, to be a chaotic mix of green species including, among others, various Blidingia spp., Percursaria percursa (C.Agardh) Rosenvinge, Rhizoclonium spp., Rosenvingiella polyrhiza (Rosenvinge) P.C.Silva, Urospora spp. and Ulothrix spp. Identification should be done carefully, as should attempting to match DNA sequences to individual species. Owing to the wonders of PCR, a clean sequence could be from any one of the species present in a collection and not necessarily the dominant one on which the microscopical identification was based. I have some confidence that all of the images here represent this genetic group, but remain uncertain if the correct binomial is applied. This is my best guess after consulting the pertinent literature (mostly Scagel (1966), Burrows (1991), Sears (2002), Brodie et al. (2007), Mathieson & Dawes (2017) and Gabrielson & Lindstrom (2018)). Considerable work remains for this genus in our flora. This name does appear to be applied to the same morphospecies concept as that for Urospora neglecta in Hanic (2005).

Image A. Filamentous gametophyte stage forming a thick covering on a rock in the upper intertidal during spring at our Letete exposed biodiversity site, Bay of Fundy, NB (GWS032356).

Image B. The club-shaped zygote forms a low turf on rocks in the upper intertidal (late summer collection from Herring Cove Head (small bay to east), Campobello Island, NB; GWS013884).
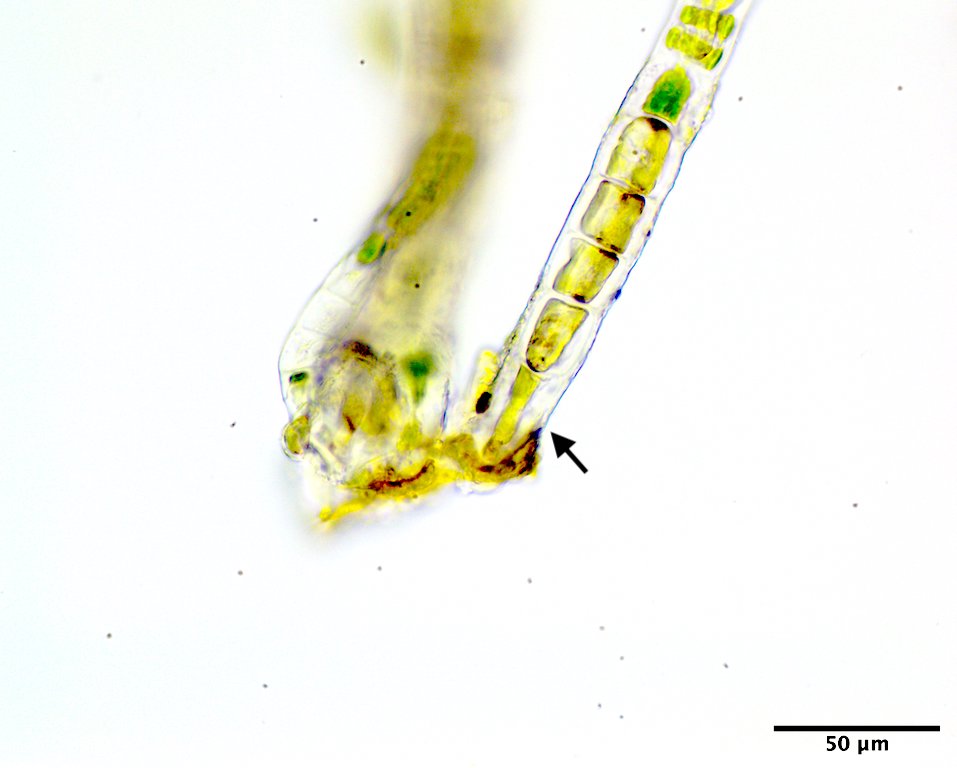
Image C. Basal cell (arrow) rhizoidal in habit (low mid intertidal on rock, Samoset Resort, ME; GWS002687; rehydrated from press).
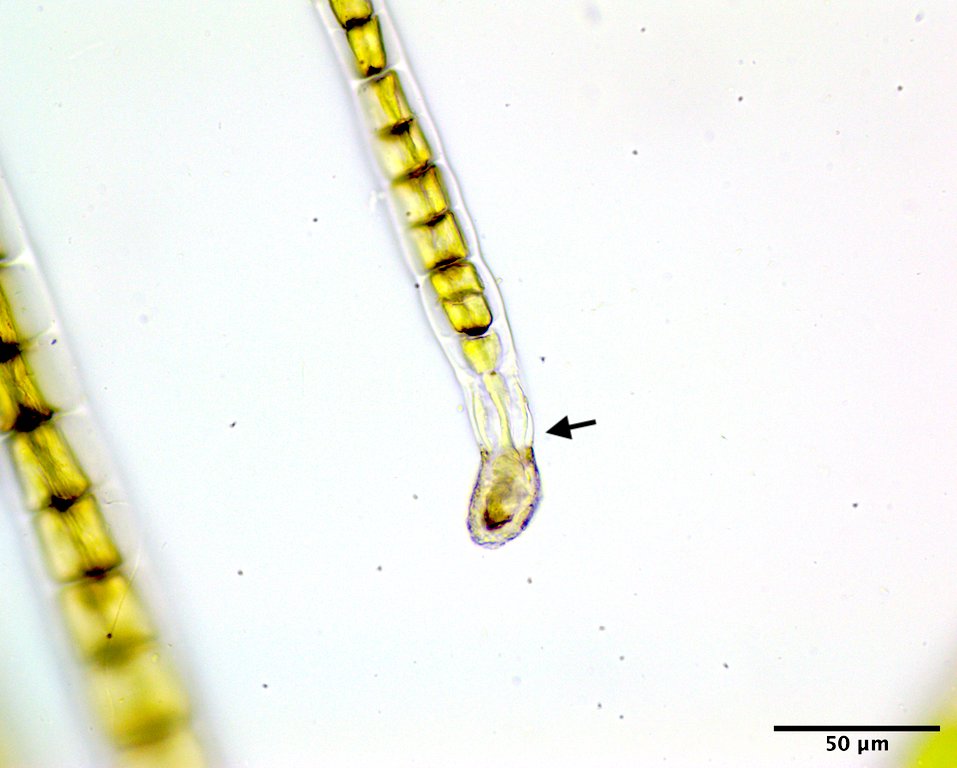
Image D. Basal cell (arrow) rhizoidal in habit (GWS002687; rehydrated from press).
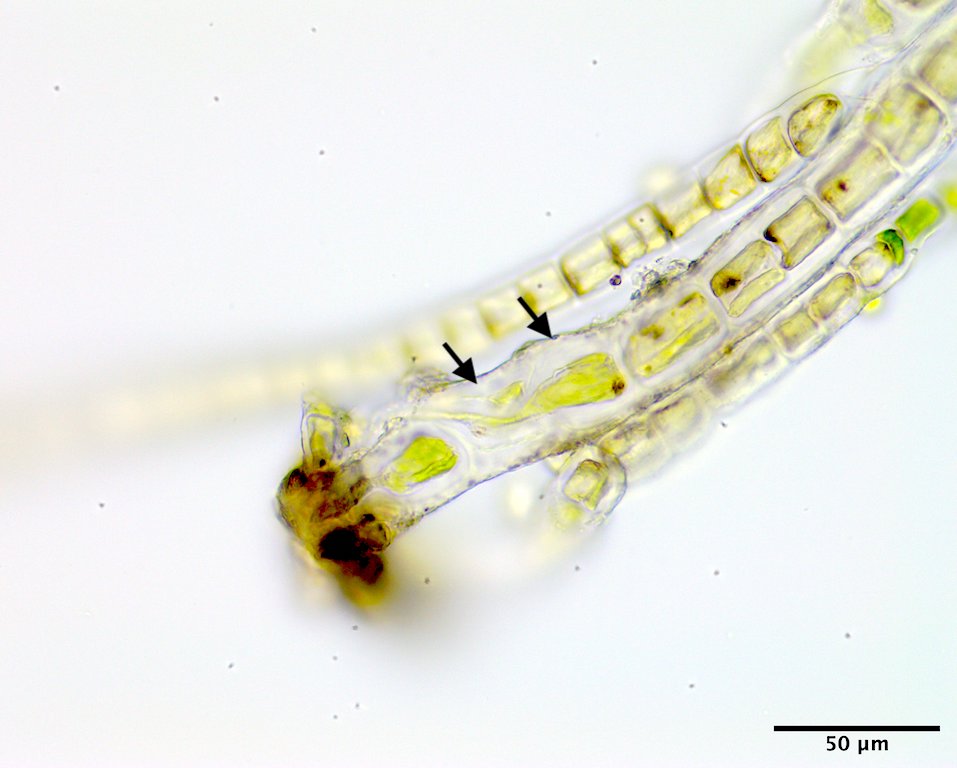
Image E. Two cells (arrows) above the basal rhizoidal cell each producing a descending intramatrical rhizoid (GWS002687; rehydrated from press).
Image F. Vegetative cells low thallus (upper intertidal on rock, Wallace Cove Lighthouse, Bay of Fundy, NB; GWS044492).
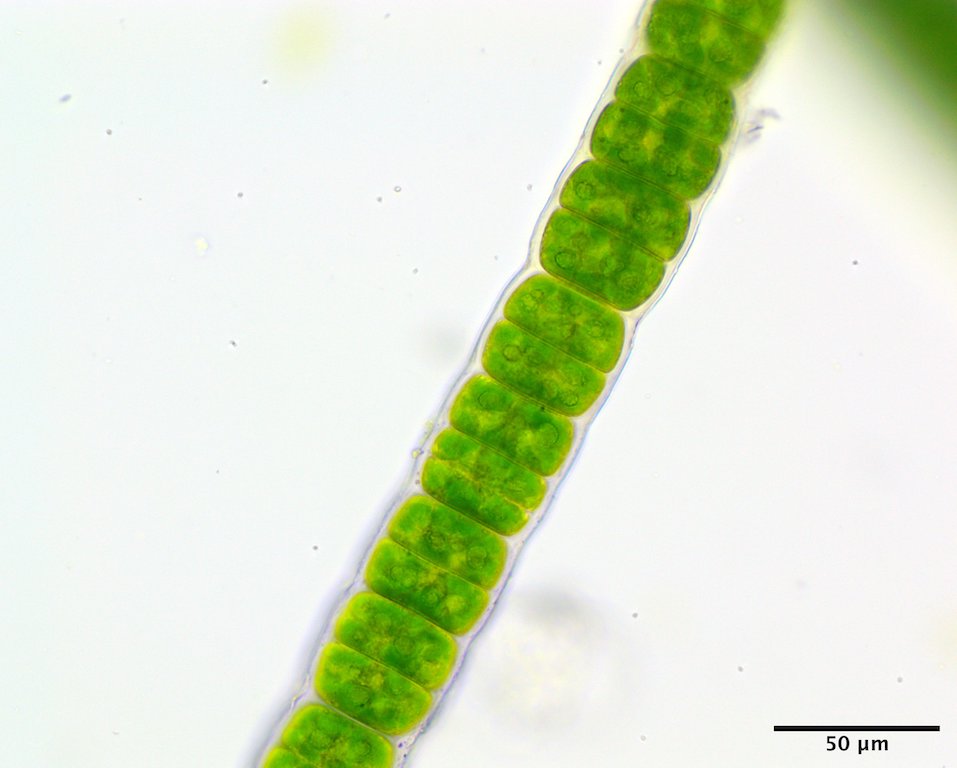
Image G. Vegetative cells mid thallus (mid intertidal on rock, Wallace Cove Lighthouse, Bay of Fundy, NB; GWS044495).
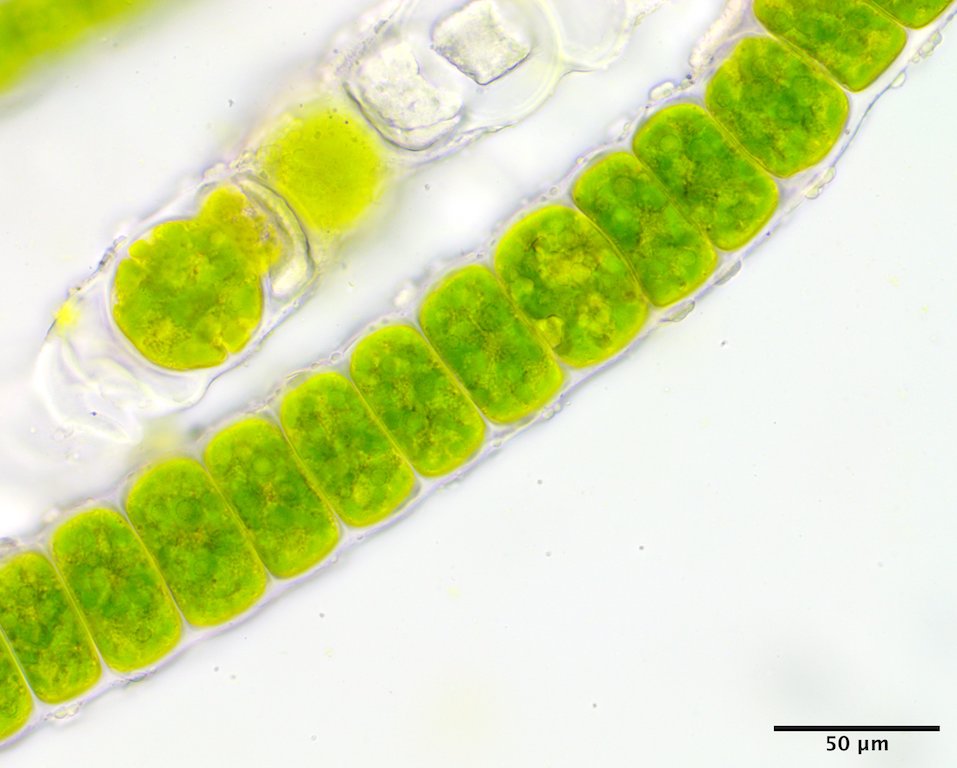
Image H. Vegetative cells upper thallus (GWS044492).
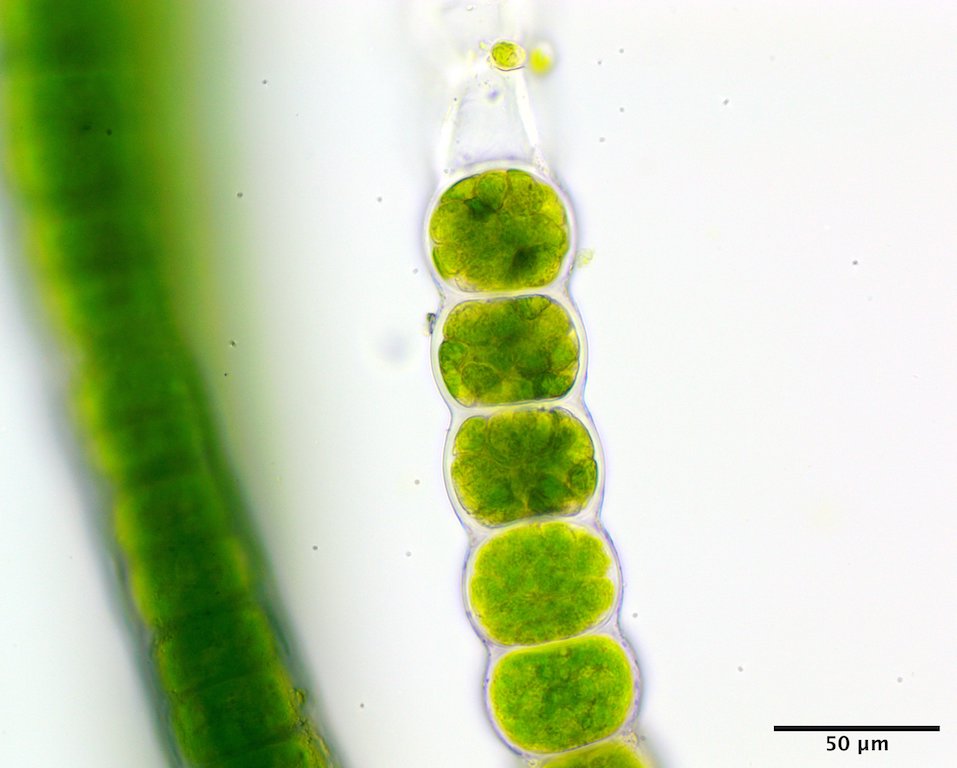
Image I. Reproductive cells, presumably gametangia (GWS044495).

Image J. The zygote (“Codiolum pusillum“) stage in this haplontic life history with the long basal stalk and terminal club-shaped cytoplasm (upper intertidal on rock, Riviere du Loup, QC; GWS008049; rehydrated from press).
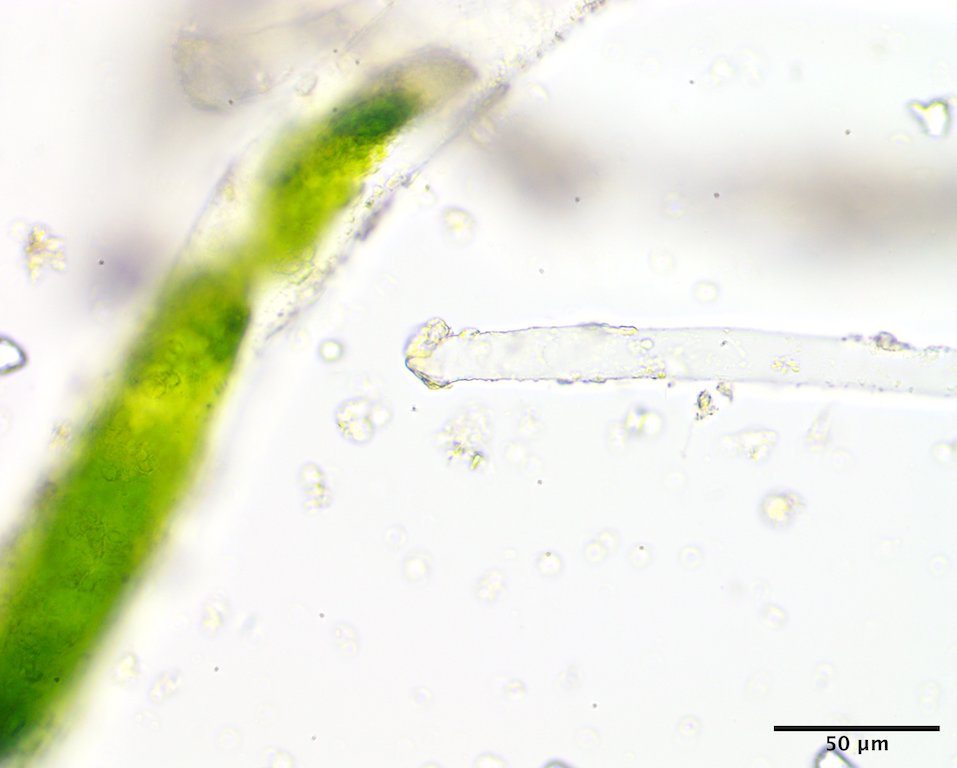
Image K. Basal attachment pad at the base of the long stalk (GWS008049; rehydrated from press).
