Secondary Metabolites and the Risks of Isaria fumosorosea and Isaria farinosa


Key Laboratory of Bio-Pesticide Innovation and Application of Guangdong Province, College of Agriculture, South China Agricultural University, Guangzhou 510642, China
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(4), 664; https://doi.org/10.3390/molecules24040664
Submission received: 21 January 2019
/
Revised: 8 February 2019
/
Accepted: 11 February 2019
/
Published: 13 February 2019
Abstract
:Isaria fumosorosea and Isaria farinosa are important entomopathogenic fungi with a worldwide distribution and multiple host insects. However, the concerns about the safety risks of myco-pesticides have been attracting the attention of researchers and consumers. Secondary metabolites (SMs), especially the mycotoxins, closely affect the biosafety of Isaria myco-insecticides. In the last forty years, more than seventy SMs were identified and isolated from I. fumosorosea and I. farinose. The SMs of I. fumosorosea include the mycotoxins of non-ribosomal peptides (NRPs) (beauvericin and beauverolides), terpenes (trichocaranes and fumosorinone), lactone compounds (cepharosporolides), acids (dipicolinic acid and oxalic acid), etc. Meanwhile, the NRP mycotoxins (cycloaspeptides) and the terpene compounds (farinosones and militarinones) are the main SMs in I. farinosa. Although several researches reported the two Isaria have promised biosafety, the bioactivities and the safety risks of their SMs have not been studied in detail so far. However, based on existing knowledge, most SMs (i.e., mycotoxins) do not come from Isaria myco-insecticide itself, but are from the host insects infected by Isaria fungi, because only the hosts can provide the conditions for fungal proliferation. Furthermore, the SMs from Isaria fungi have a very limited possibility of entering into environments because many SMs are decomposed in insect cadavers. The biosafety of Isaria myco-insecticides and their SMs/mycotoxins are being monitored. Of course, SMs safety risks of Isaria myco-insecticides need further research.
1. Introduction
Isaria fumosorosea and Isaria farinosa—formerly known as Paecilomyces fumosoroseus and Paecilomyces farinosus, respectively—are important entomopathogenic fungi with a worldwide distribution and multiple host insects [1,2]. Although differing from the popular Beauveria bassiana and Metarhizium anisopliae species thoroughly researched in various areas, both I. fumosorosea and I. farinosa attract more attention. They have multiple hosts, do not show harmful effects linked to the use of chemical pesticides, and are considered to be environmentally friendly [3]. Besides their application as pest biocontrol agents, there were some experiments indicating the both fungi have potential uses in the biotransformation of flavonoids glycosides, steroids, etc. [4,5,6].
I. fumosorosea is a species complex and mainly infects hemipteran and lepidopteron insects, such as aphids, leafhoppers, whiteflies, and the Asian citrus psyllid, etc. [7,8]. Other recently reported host insects besides hemipteran and lepidopteron insects include the subterranean termites, Coptotermes curvignathus and Coptotermes gestroi [9], rice weevils, Sitophilus oryzae [10], yellowmargined leaf beetles, and Microtheca ochroloma [11]. I. fumosorosea has been used as a pest biocontrol agent in many countries. In the USA, it was registered under NOFLY™ Technical for use as biocontrol for whiteflies, aphids, thrips, psyllids, mealybugs, and fungus gnats in greenhouses (https://www.epa.gov/pesticides/). The Apopka 97 strain in the European Union (http://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/) and the Challenger or Puma myco-pesticide in Brazil (http://agrofit.agricultura.gov.br/agrofit_cons/) were registered as well. In China, although this fungus has not been registered as a myco-pesticide, it is researched and used widely to control whiteflies and aphids [3,12,13]. Compared to I. fumosorosea, I. farinosa is less researched and used. However, in the former Soviet Union, this species (Paecilomyces farinosus) was used as myco-insecticide named “Paecilomin” to control the apple moth, Siberian pine caterpillar, and larch caterpillar [14,15,16]. It was also reported that, in the laboratory or greenhouse, this fungus is effective against the rice weevil Sitophilus oryzae [10], termite Nasutitermes corniger [2], horn fly Haematobia irritans [17], two spotted spider mite Tetranychus urticae [18], vine mealybug Planococcus ficus [19], sunn pests Eurygaster integriceps and Eurygaster austriaca [20], emerald ash borer Agrilus planipennis [21], Aelia rostrata [22], pine bark-weevil (Pissodes punctatus) [23], etc. However, in China, I. farinosa is also considered as a pathogen of Hepialus sp. and seriously affects the production of the traditional medicinal mushroom, Ophiocordyceps sinensis. This is because Hepialus sp. is the host of O. sinensis [24,25,26].
The biology, ecology, and application for biocontrol agents of both fungi were carefully reviewed 10 years ago [7]. However, during the past decade, numerous research reports about the two fungal species covering various areas were published. In the area of secondary metabolites (SMs), a lot of new compounds have been isolated and identified from the fungi. Some of the SMs are mycotoxins, which have risks contaminating foods and impacting human health. There were several reports on the risk evaluations of myco-pesticides and their mycotoxins, mainly involving B. bassiana, M. anisopliae, and their NRP (non-ribosomal peptide) and PK (polyketide) metabolites [27,28,29,30]. However, there have been few documents about the risks of Isaria fungi and their mycotoxins. In the current review, we will focus on the mycotoxins of both Isaria fungi, including their structures, bioactivities, and toxicities. We will also focus on the risk evaluation of these fungi entering food chains.
2. Secondary Metabolites (SMs) from Isaria fumosorosea
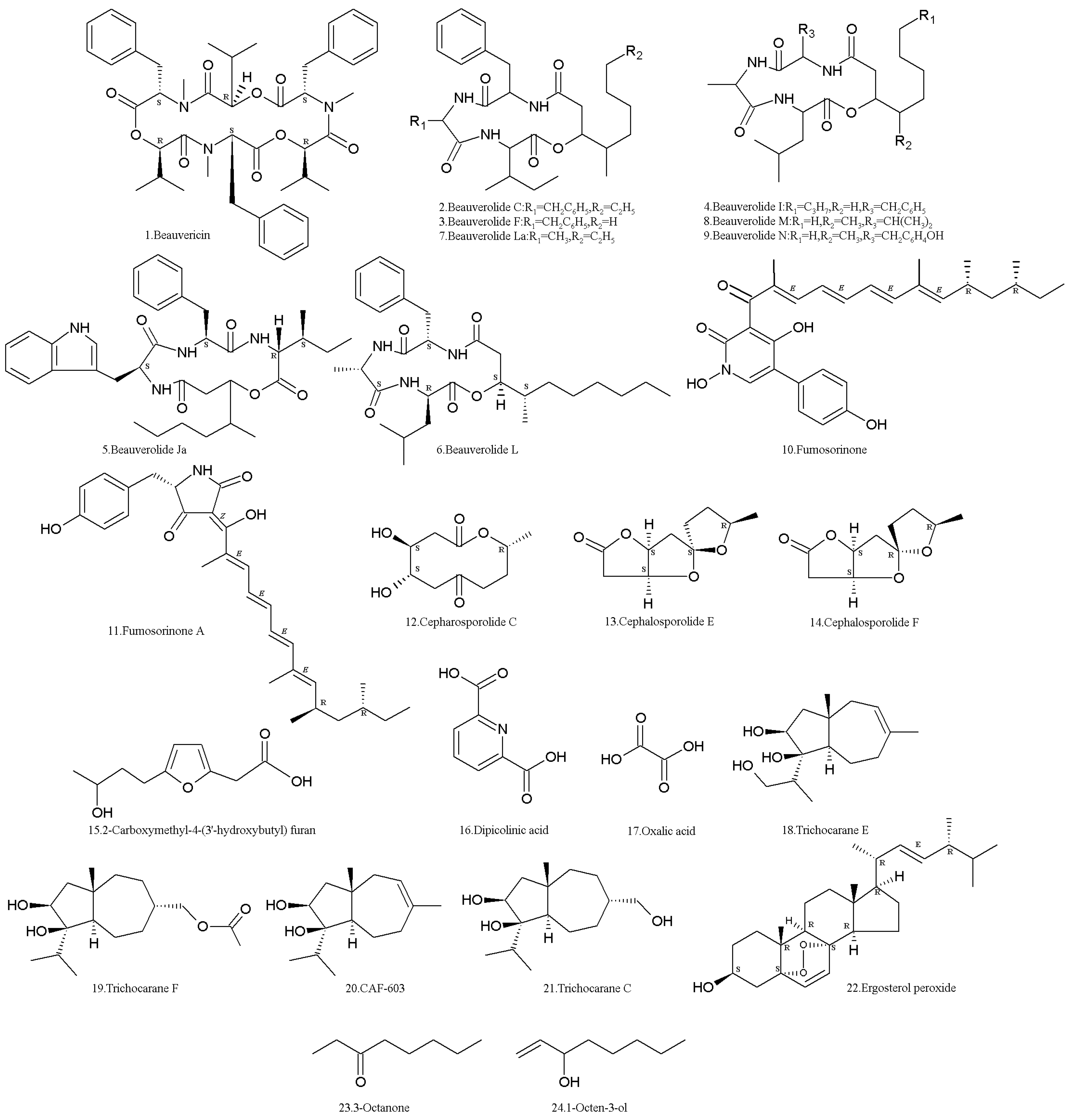
The NRP metabolite, beauvericin (1) (Table 1, Figure 1), was isolated from the strain ACCC37775 of I. fumosorosea (Hebei University, Baoding, China). It showed apparent inhibitory activity to protein tyrosine phosphatase 1B (PTP1B) with an IC50 value of 0.59 M [31]. It is a cyclic hexadepsipeptide mycotoxin with antibacterial, insecticidal, antiviral, and cytotoxic activities, and has potential value in the development of new pesticides [32,33,34,35]. However, the risks of beauvericin contamination are attracting the attention of researchers [36,37,38].
The other NRPs, beauverolides C (2), F (3), I (4), Ja (5), L (6), M (8), and N (9) (Table 1, Figure 1), were isolated from the strain BMFM-UNAM 834 of I. fumosorosea (Universidad Nacional Autonoma de Mexico, Mexico City, Mexico). These cyclotetradepsipeptides displayed a high affinity to calmodulin (CaM), with dissociation constants (Kd) ranging from 0.078–3.440 μM. Beauverolide Ja (5) which is the only one containing a tryptophan residue in its structure showed the highest affinity to CaM [39]. The beauverolides L and La (7) (Table 1, Figure 1) were isolated and identified from the PFR97-Apopka (ATCC 20874) strain (WR Grace & Co, Conn, Columbia, MD, USA) of I. fumosorosea [40]. Beauverolide L has anti-immunity activity against the greater wax moth, Galleria mellonella [41].
The terpene compound, fumosorinone (10) (Table 1, Figure 1) was isolated from the ACCC37775 strain of I. fumosorosea (Hebei University, Baoding, China). Fumosorinone (10) is structurally similar to tenellin and desmethylbassianin, but has different chain length and degree of methylation. Fumosorinone (10) acts as a classic non-competitive inhibitor of protein tyrosine phosphatase 1B (PTP1B) with an IC50 of 14.04 µM, which suggests that it is a potential medicine for the treatment of type II diabetes and other associated metabolic syndromes. The gene cluster of fumosorinone biosynthesis includes a hybrid polyketide synthase–nonribosomal peptide synthetase gene, two cytochrome P450 enzyme genes, a trans-enoyl reductase gene, and other two transcription regulatory genes [42]. Fumosorinone (10) also showed cytotoxic against human cancer lines, including HeLa, MDA-MB-231, and MDA-MB-453 cell lines [43]. A compound similar to fumosorinone, fumosorinone A (11) (Table 1, Figure 1), was identified as well. It is also a PTP1B inhibitor [31].
The lactone compounds, cepharosporolides C (12), E (13), and F (14), and an organic acid, 2-carboxymethyl-4-(3′-hydroxybutyl) furan (15) (Table 1, Figure 1) were isolated [31]. They had no antimalaria activity to Plasmodium falciparum K1 [44], and no inhibition to PTP1B [31].
The other acids, dipicolinic acid (DPA) (16) and oxalic acid (OXA) (17) (Table 1, Figure 1), were found in the I. fumosorosea Pfrd strain (Centro Nacional de Referencia de Control Biológico, Tecomán, Colima, Mexico). DPA (16) was the most abundant metabolite with insecticidal activity against the third-instar nymphs of the whitefly in bioassays involving topical applications. DPA (16) was detected after 24 h when the fungus started growing in submerged cultures. The production of DPA (16) was directly correlated with fungal growth, but the maximal yield was only 0.041 g/L [45]. In submerged fermentation, carbon was significantly directed towards the synthesis of DPA (16) and OXA (17), especially under zinc limitation [46]. OXA (17) has antimicrobial and antioxidant activities [47,48] and can delay the sclerotial formation of Polyporus umbellatus [49], which is called as “Zhuling”, a traditional Chinese medicine used for a wide range of ailments related to the edema, scanty urine, vaginal discharge, urinary dysfunction, jaundice, and diarrhea [50].
Two new carotane-type sesquiterpenes named trichocaranes E (18) and F (19), along with two known ones called CAF-603 (20) and trichocarane C (21) (Table 1, Figure 1), were isolated from the I. fumosorosea ACCC37775 strain (Hebei University, Baoding, China). Compounds 18–20 showed potent cytotoxic activities against six tumor cell lines (i.e., MDA, MCF-7, SKOV-3, Hela, A549, and HepG2), with IC50 values of 0.1–6.0 μg/mL [51].
Peroxy-ergosterol (22) (Table 1, Figure 1) was isolated from the RCEF1253 strain of I. fumosorosea (Anhui Agricultural University, Hefei, China) by high-speed-counter-current chromatography [52]. It has various bioactivities, such as cytotoxicity to cancer cells P-388, KB, A549, and HT-29, with ED50s of 0.4, 2.1, 2.7, and 1.4 μg/mL [53]. It also induces the apoptosis of the human leukemia cell HL-60 [54]. Furthermore, this fungus has a relatively high vitamin A content, which shows that it is a potential producer of vitamin A [55].
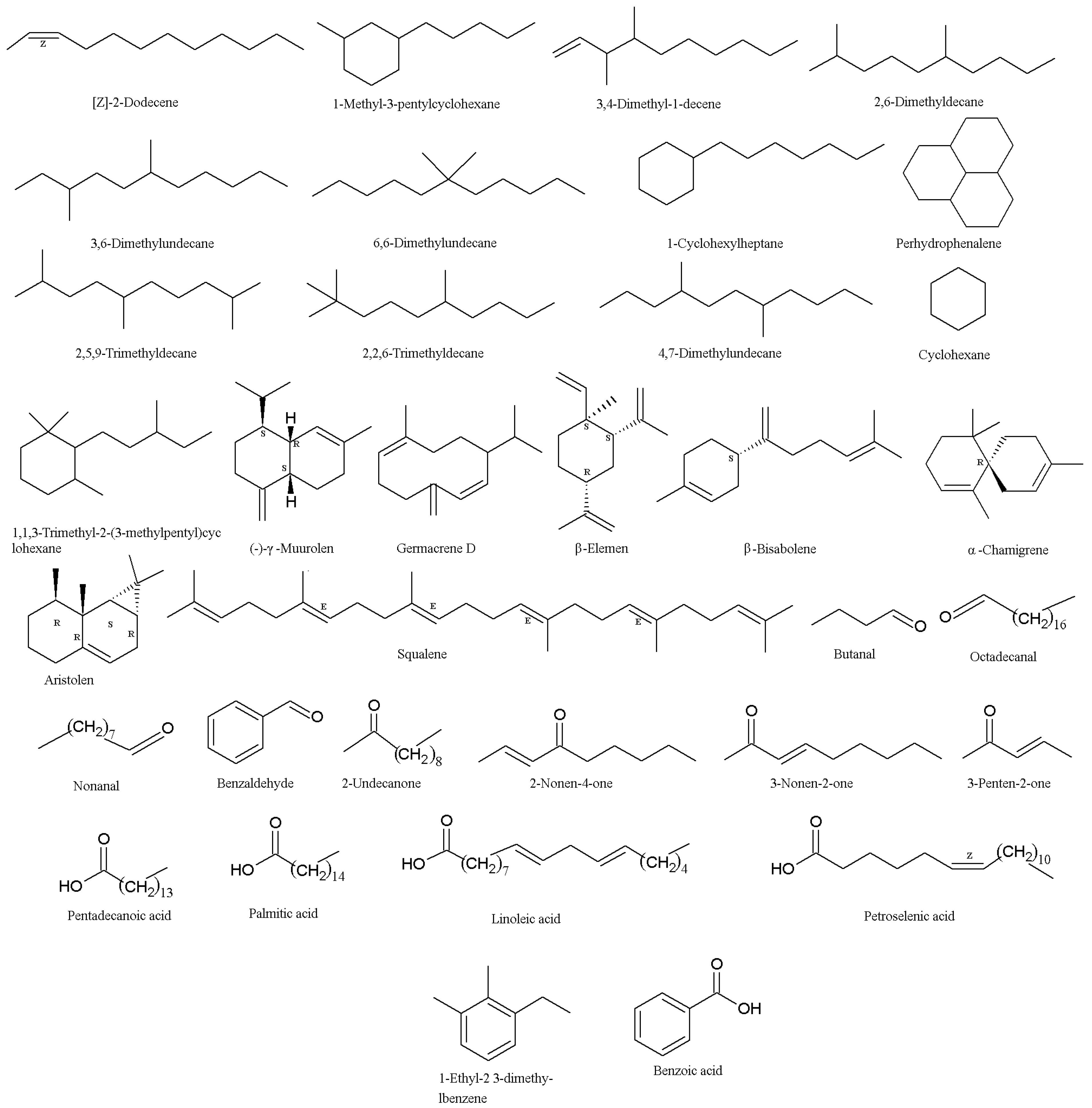
In order to investigate the termite’s response to entomopathogenic fungi, the fungal volatile organic compounds (VOCs) were detected using GC–MS. It was found that 3-octanone (23) and 1-octen-3-ol (24) (Table 1, Figure 1) were the major surface chemical compounds on the conidia of the I. fumosorosea K3 strain (Kyoto University, Kyoto, Japan), and the total quantities of the two chemicals on the surface of fungal conidia were estimated to be approximately 0.01 ng per 107 conidia. The Formosan subterranean termites, Coptotermes formosanus, showed aversion to the two compounds [56]. The results indicated that, in contrast to their reaction to M. anisopliae, I. fumosorosea cultures were not repellent to Formosan subterranean termite workers, which were highly susceptible to infection of I. fumosorosea. The electroantennographic responses of workers to the conidia of I. fumosorosea were approximately 78% less than those to M. anisopliae. The VOC profile of repellent cultures of M. anisopliae mainly consisted of paraffins (60.97%), while the major proportion of the I. fumosorosea profile consisted of branched and cyclic alkanes (84.41%), such as [Z]-2-dodecene, 1-methyl-3-pentyl cyclohexane, 3,4-dimethyl-1-decene, 2,6-dimethyldecane, 3,6-dimethylundecane, 6,6-dimethylundecane, 1-cyclohexylheptane, perhydrophenalene, 2,5,9-trimethyldecane, 2,2,6-trimethyldecane, 4,7-dimethylundecane, cyclohexane, and 1,1,3-trimethyl-2(3-methylpentyl) (Figure 2) [57]. In another report, numerous VOCs were identified in the mycelia of the I. fumosorosea UPH48 strain (Siedlce University of Natural Sciences and Humanities, Siedlce, Poland). The VOCs included terpenes such as γ-muurolen, germacrene D, β-elemen, β-bisabolen, α-chamigren, aristolen, and squalene. The aldehydes included butanal, octadecanal, nonanal, benzaldehyde, fenyloacetaldehyde, and 2-undecanone. The ketones included 2-nonen-4-one, 3-nonen-2-one, and 3-penten-2-one. In addition, five fatty acids (i.e., pentadecanoic, palmitic, g-linolenic, linoleic, and petroselinic) and others compounds (i.e., 1-ethyl-2,3-dimethylbenzene and benzoic acid) (Figure 2) were identified in the mycelia of the I. fumosorosea UPH48 strain (Siedlce University of Natural Sciences and Humanities, Siedlce, Poland) [58].
3. Secondary Metabolites (SMs) from Isaria farinosa
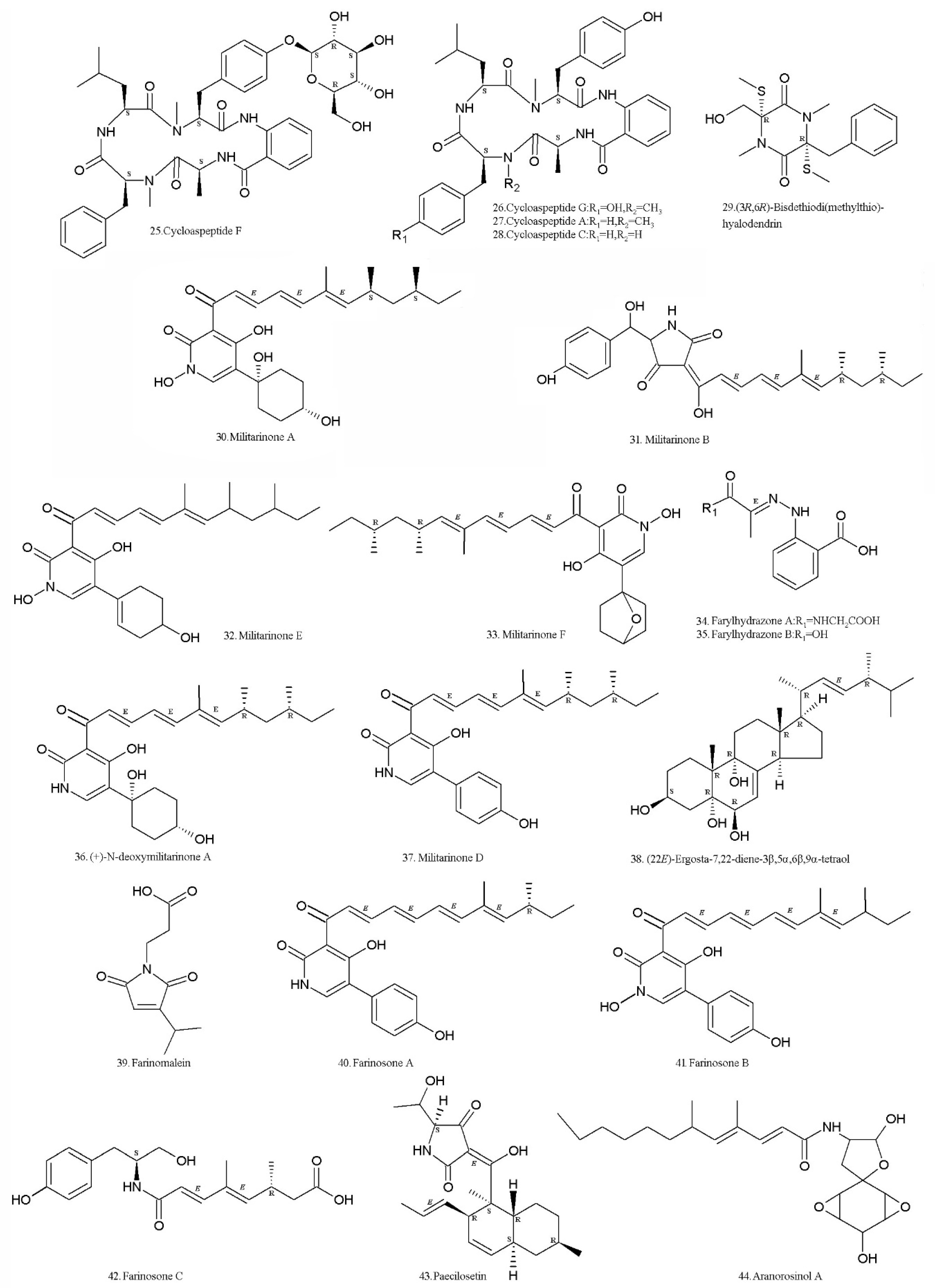
Cycloaspeptides F (25) and G (26) (Table 2, Figure 3), two new cyclic pentapeptides, and the known cycloaspeptides A (27), C (28), and bisdethiodi (methylthio) hyalodendrin (29) (Table 2, Figure 3) were isolated from the fermented rice substrate with the I. farinosa strain XJC04-CT-303 (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) that colonizes Cordyceps sinensis. Cycloaspeptides F (25) and G (26) inhibited the growth of MCF7 cells, which was comparable to the positive control 5-fluorouracil. They also had modest cytotoxic effects on HeLa cells [59]. Cycloaspeptide A (27) has a low cytotoxicity in human lung fibroblasts [60]. Cycloaspeptide C (28) is closely related to cycloaspeptide G (26), but its bioactivity is not reported. The gene cluster responsible for the biosynthesis of the cycloaspeptides were identified in Penicillium soppii and Penicillium jamesonlandense. Heterologous expression in Aspergillus oryzae has demonstrated that the minimal gene set required to produce both cycloaspeptide A and cycloaspeptide E is a 5-module NRPS and a new type of pathway-specific N-methyltransferase (N-MeT). Gene knock-outs and feeding studies have demonstrated that two modules of the NRPS preferentially accept and incorporate N-methylated amino acids, which are provided by the pathway-specific N-MeT. This is a system not previously seen in secondary metabolism [61]. The diketopiperazine derivative, bisdethiodi (methylthio) hyalodendrin (29) (gliovictin), was isolated in 1973. It exhibited weak cytotoxic activity on KB (human epidermoid carcinoma of the mouth) with IC50 of 42 µg/mL, and had IC50 values of >50 µg/mL on HepG2 (human hepatocellular liver carcinoma cell line), A549 (human lung carcinoma cell line), HCC-S102 (hepatocellular carcinoma cell line), HuCCA-1 (human cholangiocarcinoma cancer cells), HeLa (cervical adenocarcinoma cell line), MDA-MB231 (human breast cell line), T47 D (human mammary adenocarcinoma cell line), HL-60 (human promyelocytic leukemia cell line), and P388 (murine leukemia cell line) [62].
The terpene compounds, militarinones A (30), B (31), E (32), and F (33) (Table 2, Figure 3), were isolated from the I. farinosa strain XJC04-CT-303 (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) [63]. Militarinones A (30) and E (32) had significant cytotoxicity on the A549 human carcinoma cell line, whereas militarinone B (31) was active against Staphylococcus aureus, Streptococcus pneumoniae, and Candida albicans [63]. Militarinone A (30) at the concentration of 10 μM, had obvious neurotrophic effects on PC-12 cells [64]. The phenylhydrazones, farylhydrazones A (34) and B (35) (Table 2, Figure 3), were isolated from this strain [63]. No bioactivity report has been found yet. The two compounds can be synthesized with six and five steps respectively, starting from 2-nitrobenzoic acid [65].
A new pyridone alkaloid, (+)-N-deoxymilitarinone A (36), along with the related metabolites, militarinone D (37), militarinone B (31) and the sterol 22E,4R-ergosta-7,22-diene-3β,5α,6β,9α-tetraol (38) (Table 2, Figure 3), were isolated from I. farinosa RCEF 0097 (Entomogenous Research Centre, Anhui Agricultural University, Hefei, China). The (+)-N-deoxymilitarinone A (36) at 33 and 100 μM concentrations induced neurite sprouted in PC 12 cells. A cytotoxic effect was observed in human neurons (IMR-32) at a concentration of 100 μM [66]. A diverted total synthesis approach to the total synthesis of N-deoxymilitarinone A was developed by Ding et al. [67].
A new maleimide-bearing compound, farinomalein (39) (Table 2, Figure 3), was isolated from the strain HF599 (National Institute of Fruit Tree Science, Tsukuba, Japan). It showed potent activity against the plant pathogen Phytophthora sojae [68]. It can be synthesized in two steps from a readily available γ-hydroxybutenolide [69]. The activities against Phytophthora sojae and Aphanomyces cochlioides were confirmed [70].
Two new yellow pigments, farinosones A (40) and B (41), together with farinosone C (42) (Table 2, Figure 3), were isolated from the mycelial extract of the entomogenous fungal strain RCEF 0101 (Entomogenous Research Centre, Anhui Agricultural University, Hefei, China). Farinosone C (42) is a new metabolite derived from an early step of pyridone alkaloid biosynthesis. Farinosones A (40) and C (42) at 50 µM induced neurite outgrowth in the PC-12 cell line, while farinosone B (41) was inactive. The three farinosones had no cytotoxicity against PC-12 cells when tested at 50 µM concentration in the MTT assay [71]. Farinosones are similar to militarinones in structure, belonging to the terpene group of compounds.
A new tetramic acid derivative, paecilosetin (43) (Table 2, Figure 3), was isolated from the strain CANU TE108 (University of Canterbury, Christchurch, New Zealand). It showed activity against the P388 cell line, with IC50 values of 3.1 μg/mL, and was also active against the microorganisms Bacillus subtilis, Trichophyton mentagrophytes, and Cladosporium resinae [72]. Paecilosetin (43) and aranorosinol A (44) (Table 2, Figure 3) were identified in the I. farinosa HF511 strain. They act plant pathogenic oomycetes, Phytophthora sojae and Aphanomyces cochlioides [70]. Aranorosinol A (44) also had weak antibacterial and antifungal properties on other microorganisms [73].
I. farinosa can produce a water soluble anthraquinone-related red pigment with good stability after being exposed to salt solution (96.1% stability after treatment with sodium chloride), acid (72.1% stability with citric acid), heat (86.2% stability at 60 °C), and sunlight (99.4% stability). It shows a potential for pigment production [74,75].
4. Risks of the Secondary Metabolites (SMs) from both Isaria Myco-Insecticides
In recent years, the concerns about the safety risks of myco-pesticides and their SMs have been attracting the attention of researchers and consumers. In fact, the popular myco-insecticides, Beauveria bassiana. and Metarhizium anisopliae, have been proposed as low-risk environmental alternatives to chemical insecticides for controlling agricultural pests and disease vectors [27,28] This is because their safety for humans and the environment were well evaluated [76,77,78,79], while the mycotoxins they produced were considered unlikely to enter food chains [80].
However, there were few experiments on the safety of Isaria myco-insecticides. I. fumosorosea was the subject of two reports involving safety analysis, while I. farinosa has not been paid attention yet. The I. fumosorosea monospore culture EH-506/3 (BMFM-UNAM 834, Universidad Nacional Autonoma de Mexico, Mexico City, Mexico) was subjected to a biosafety test by applying a 2 g/kg of animal body weight dose on the shaved skin of 16 New Zealand rabbits, with an exposure time of 24 h. The results indicated that none of the rabbits showed clinical signs of any disease, and their body weight corresponded to the expected weight for a healthy rabbit. The test data supports the safety of I. fumosorosea EH-506/3 when applied to the skin [29]. Another toxicity test on I. fumosorosea was completed in China. The toxicities of acute oral, dermal, and inhalation to rats were recorded as LD50 > 5000 mg/kg, LD50 (4 h) > 2000 mg/kg, and LC50 (2 h) > 2000 mg/m3, respectively. No irritation action was observed in rabbit eyes, and no dermal sensitization reaction was found on the treated rabbit skin. These results suggested that I. fumosorosea has low toxicities of acute oral, dermal toxicity, and inhalation, and it can be graded as a weak sensitizer [81].
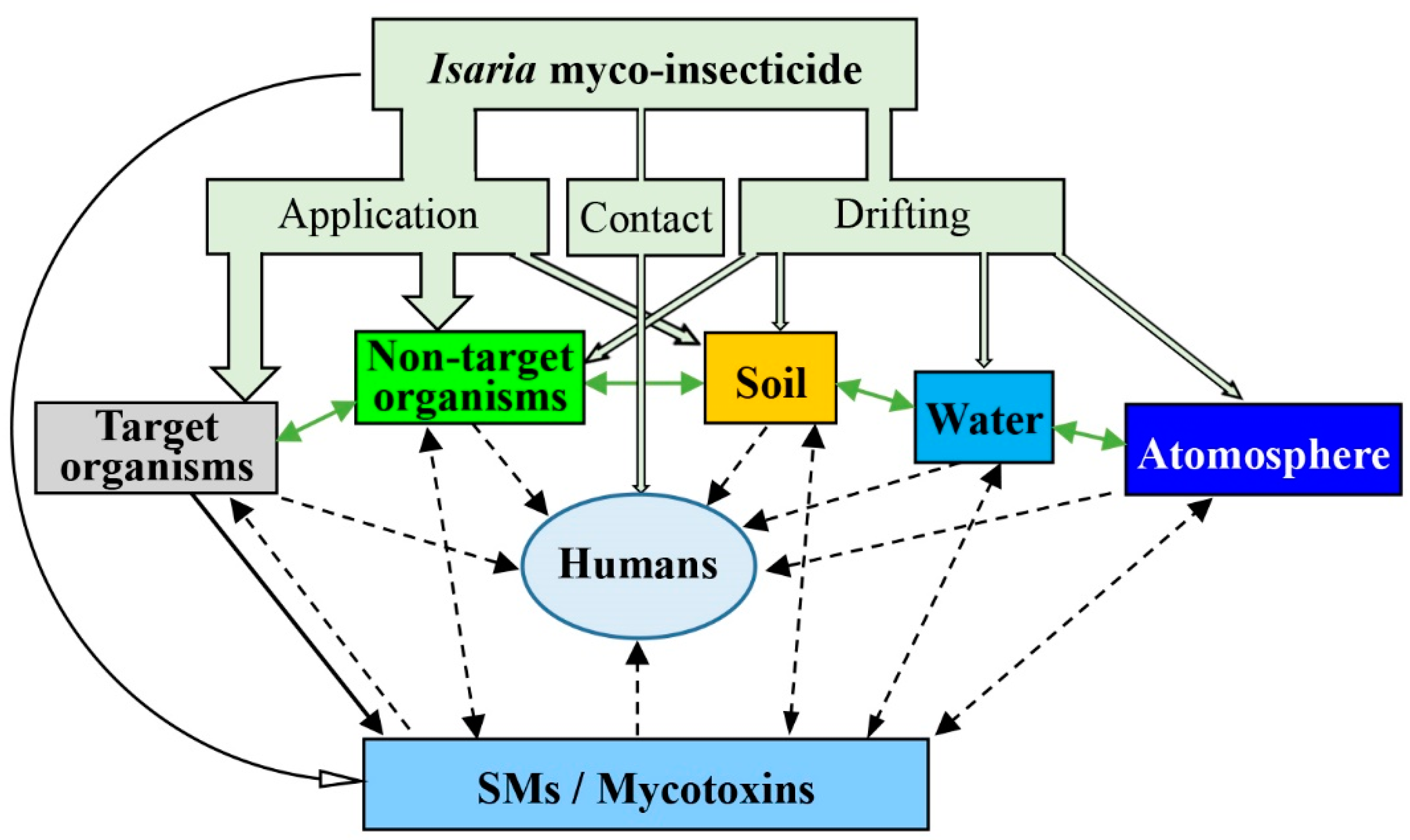
Overall, there are six destinations (i.e., target organisms, non-target organisms, soil, water, atmosphere, and humans) involved in the production and application of Isaria myco-insecticide formulations (Figure 4). The most important destination is target organisms, including the pests and crops, when Isaria myco-insecticide is released in fields. In practice, a few fungal spores of myco-insecticide probably land on insect surfaces, but the fungus will proliferate on the infected insect hosts, which suffer a pathogenic process from spore germination and the formation of the next generation of spores. Many mycotoxins of entomopathogenic fungi are probably biosynthesized in hemocoel to conquer the host’s immunity [82]. The target crops are probably the main destination, especially when the Isaria myco-insecticide is applied by stem-leaf treatment in fields. This is because most of the fungal spores are dropped on the plants with canopy covering the ground. In addition, the endophytic characteristics of entomopathogenic fungi might produce some SMs because Isaria fungi, similar to B. bassiana and M. anisopliae, can colonize plants [83,84,85]. Non-target organisms are an important destination of myco-insecticides as well. They represent a big category, including animals, plants, and microbes, which are not the targets of myco-insecticides but have chances to contact myco-insecticides. Among them, non-target insects might be the most important destination, because some of these insects probably are the hosts of myco-insecticidal fungi. There have been many reports published giving evidence that entomopathogenic fungi infect silkworms [86], bees [87,88], and natural enemy insects [89]. Of course, more studies found that the myco-insecticides are safe to non-target insects if they are used correctly [90,91].
Soil is another important destination, especially when Isaria myco-insecticide is released through soil treatments in fields (Figure 4). Through the drifting pathway from application and dropping pathway from target pest cadavers, fungal phages and mycotoxins can enter the soil system. In fact, entomopathogenic fungi can persist and survive in soil for a long time [92,93,94,95]. Beauveria spp., Metarhizium spp., Paecilomyces spp., and Isaria spp. can be often isolated from the soil [1]. The entomopathogens in soil can be detected after myco-insecticides are used [96], but there are no reports about the growth and proliferation of Isaria fungi and the presence of their SMs in soil. There are also no cases of soil fungi affecting human health.
Water and atmosphere are the destinations of the drifting myco-insecticides (Figure 4). In fact, many entomopathogenic fungi can persist and survive in water. I. farinosa and I. fumosorosea, similarly to M. anisopliae, can infect aquatic insects like mosquitoes [97,98]. However, there are few studies about SMs of entomopathogenic fungi in water. Milner et al. reported that the Metarhizium biopesticide is very unlikely to pose any hazard to aquatic organisms [99]. The atmosphere is another destination of Isaria myco-insecticides, where fungal entomopathogens are obtained from drifting myco-insecticides and the spore dispersal of natural fungi. However, fungi cannot either persist for a long time or proliferate in the air [100]. Also, fungi might be exchanged between soil, water, and atmosphere systems, although there are few studies on this aspect. To the date, there are also no cases of entomopathogenic fungi from water and atmosphere influence on human health.
Humans contact Isaria myco-insecticides through direct and indirect ways (Figure 4). Only people who are involved in the production process of myco-insecticides or use them in farms directly contact the fungi. Several studies have reported that fungal spores lead to allergies in workers who worked in factories of B. bassiana and M. anisopliae for long periods [27,28]. Perhaps, most people indirectly contact the fungi through foods, soil, the atmosphere, and water contaminated by the fungi. However, there have not been any case reports about people’s health affected by indirectly contacting entomopathogenic fungi.
Undoubtedly, the biosafety risks of Isaria myco-insecticides are closely related to the sources and fates of the SMs (especially the mycotoxins) produced by entomopathogenic fungi.
In fact, the SMs of myco-insecticide itself are very limited. Because the active ingredients of myco-insecticide formulations are the spores of the fungal entomopathogen, the fungi cannot proliferate in the formulation and cannot produce new SMs. Most SMs possibly exist in the spore cells rather than outside the spores [80]. Therefore, the main sources of SMs basically include the target pests or host insects infected by fungal entomopathogen of myco-insecticide, because the host insects support the fungi with conditions for proliferation. The source of SMs and mycotoxins is the growing entomopathogenic filamentous fungus. In addition, endophytic entomopathogenic fungi might be an SM source, because some Isaria strains can colonize plants and become endophytic fungi [101,102].
Currently, we do not know the detailed fates of the fungal SMs of myco-insecticides. Obviously, there are certain possibilities that the SMs of entomopathogenic fungi enter environments, however, there have not been any reports that show evidence of the entry of SMs from myco-insecticides into environments. In fact, a few research cases indicate that mycotoxins are scarcely released into environments from insects. For example, destruxin analogues were shortly decomposed by M. anisopliae after the host insects died, which was presumably due to the activity of hydrolytic enzymes in the insects’ cadavers. This appeared to be independent of host or soil type and biota. The study supported that destruxins are essentially restricted to the host and pathogen and are unlikely to contaminate the environment or enter the food chain [103,104].
To date, it has been found that most mycotoxins that contaminate environments and food chains come from the crops and products infected by fungal phytopathogens, such as Fusarium spp., Aspergillus spp., etc., rather than fungal entomopathogens [38,105], despite the fact that both phytopathogenic and entomopathogenic fungi often produce the same mycotoxins [106]. For example, Schenzel et al. reported that beauvericins were detected in drainage water where wheat was inoculated with Fusarium spp., which is a producer of beauvericins [107].
In conclusion, there are more than seventy SMs identified and isolated from I. fumosorosea and I. farinosa. Many of these are mycotoxins attracting people’s concerns about the biosafety. The SMs of I. fumosorosea include the NRP mycotoxins (beauvericin and several beauverolides), terpenes (several trichocaranes and fumosorinone), lactone compounds (several cepharosporolides), and acids (dipicolinic acid and oxalic acid). In I. farinosa, the NRPs (several cycloaspeptides) and terpenes (several farinosones and militarinones) were the main SMs. Currently, the bioactivities and mechanisms of action of the SMs in both Isaria have not been well studied, and neither have the risks of these compounds been carefully assessed. However, it is indicated that most SMs (mycotoxins) come from the host insects infected by Isaria fungi rather than the Isaria myco-insecticide itself, because the hosts provide all the conditions for fungal proliferation. Furthermore, the possibility of SMs from Isaria fungi entering into environments is very limited, because many SMs are decomposed in insect cadavers. Although more careful research in the future is essential, the biosafety of Isaria myco-insecticides and their SMs/mycotoxins in current is under control.
Funding
This research was funded by Guangdong Province Science & Technology Project, 2016B020234005, and the National Natural Science Foundation of China, 31572053.
Acknowledgments
The authors thank the reviewers for their careful and constructive comments.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
References
- Hu, Q.; Liu, S.; Yin, F.; Cai, S.; Zhong, G.; Ren, S. Diversity and virulence of soil-dwelling fungi Isaria spp. and Paecilomyces spp. against Solenopsis invicta (hymenoptera: Formicidae). Biocontrol Sci. Technol. 2011, 21, 225–234. [Google Scholar] [CrossRef]
- Lopes, R.D.; de Lima, G.; Correia, M.T.D.; da Costa, A.F.; Lima, E.A.D.A.; Lima, V.L.D. The potential of Isaria spp. as a bioinsecticide for the biological control of nasutitermes corniger. Biocontrol Sci. Technol. 2017, 27, 1038–1048. [Google Scholar] [CrossRef]
- Gao, T.; Wang, Z.; Huang, Y.; Keyhani, N.O.; Huang, Z. Lack of resistance development in Bemisia tabaci to Isaria fumosorosea after multiple generations of selection. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska, E.; Hoc, N.; Sycz, J.; Urbaniak, M.; Dymarska, M.; Grzeszczuk, J.; Kostrzewa-Suslow, E.; Stepien, L.; Plaskowska, E.; Janeczko, T. Biotransformation of steroids by entomopathogenic strains of Isaria farinosa. Microb. Cell Fact. 2018, 17, 71. [Google Scholar] [CrossRef] [PubMed]
- Dymarska, M.; Janeczko, T.; Kostrzewa-Suslow, E. Glycosylation of 3-hydroxyflavone, 3-methoxyflavone, quercetin and baicalein in fungal cultures of the genus Isaria. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Dymarska, M.; Grzeszczuk, J.; Urbaniak, M.; Janeczko, T.; Plaskowska, E.; Stepien, L.; Kostrzewa-Suslow, E. Glycosylation of 6-methylflavone by the strain Isaria fumosorosea KCH J2. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Majeed, M.Z.; Fiaz, M.; Ma, C.-S.; Afzal, M. Entomopathogenicity of three muscardine fungi, Beauveria bassiana, Isaria fumosorosea and Metarhizium anisopliae, against the asian citrus psyllid, Diaphorina citri (Kuwayama) (Hemiptera: Psyllidae). Egypt. J. Biol. Pest Control 2017, 27, 211–215. [Google Scholar]
- Jessica, J.J.; Peng, T.L.; Sajap, A.S.; Lee, S.H.; Syazwan, S.A. Evaluation of the virulence of entomopathogenic fungus, Isaria fumosorosea isolates against subterranean termites Coptotermes spp. (Isoptera: Rhinotermitidae). J. For. Res. (Harbin, China) 2018. [Google Scholar] [CrossRef]
- Usanmaz-Bozhuyuk, A.; Kordali, S.; Kesdek, M.; Simsek, D.; Altinok, M.A.; Altinok, H.H.; Komaki, A. Mortality effects of six different entomopathogenic fungi strains on rice weevil, Sitophilus oryzae (L.) (Coleoptera: Curculionidae). Fresen. Environ. Bull. 2018, 27, 4373–4380. [Google Scholar]
- Montemayor, C.O.; Avery, P.B.; Cave, R.D. Infection and mortality of Microtheca ochroloma (Coleoptera: Chrysomelidae) by Isaria fumosorosea (Hypocreales: Cordycipitaceae) under laboratory conditions. Biocontrol Sci. Technol. 2016, 26, 605–616. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Shakeel, M.; Li, S.; Wang, S.; Zhou, X.; Yu, J.; Xu, X.; Yu, X.; Jin, F. The entomopathogenic fungi Isaria fumosorosea plays a vital role in suppressing the immune system of Plutella xylostella: Rna-seq and dge analysis of immunity-related genes. Front. Microbiol. 2017, 8, 1421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zou, C.; Hu, Q. Effects of Isaria fumosorosea on TYLCV (tomato yellow leaf curl virus) accumulation and transmitting capacity of Bemisia tabaci. PLoS ONE 2016, 11, e0164356. [Google Scholar] [CrossRef] [PubMed]
- Pu, Z.; Li, Z. Insect Mycology, 1st ed.; Anhui Science and Technology Press: Hefei, China, 1996. [Google Scholar]
- Lappa, N.V.; Goral, V.M.; Drozda, V.P. Effectiveness of boverin and paecilomin in the control of the codling moth. Zashchita Rastenii 1977, 24, 13–18. (in Russian). [Google Scholar]
- Samsinakova, A.; Kalalova, S.; Vlcek, V.; Kybal, J. Mass production of Beauveria bassiana for regulation of Leptinotarsa decemlineata populations. J. Invertebr. Pathol. 1981, 38, 16–174. [Google Scholar] [CrossRef]
- Mochi, D.A.; Monteiro, A.C.; Machado, A.C.; Yoshida, L. Entomopathogenic fungal activity against pupae and adult Haematobia irritans (Diptera: Muscidae). Vet. Parasitol. 2010, 168, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Mustu, M.; Demirci, F.; Koksal, M.; Serbes, C.; Armagan, B. Mortality effects of Isaria farinosa and Purpureocillium lilacinum (Sordariomycetes: Hypocreales) on the two spotted spider mite Tetranychus urticae (Acari: Tetranychidae) and its predator Neoseiulus californicus (Acari: Phytoseiidae) under controlled conditions. Entomol. Gen. 2016, 35, 243–252. [Google Scholar]
- Mustu, M.; Demirci, F.; Kaydan, M.B.; Ulgenturk, S. Laboratory assay of the effectiveness of the entomopathogenic fungus Isaria farinosa (Holmsk.) fries (Sordariomycetes: Hypocreales) against the vine mealybug Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae), even under the use of fungicides. Int. J. Pest Manag. 2015, 61, 264–271. [Google Scholar] [CrossRef]
- Mustu, M.; Demirci, F.; Kocak, E. Mortality of Isaria farinosa and Beauveria bassiana on sunn pests Eurygaster integriceps and Eurygaster austriaca. Phytoparasitica 2014, 42, 93–97. [Google Scholar] [CrossRef]
- Johny, S.; Kyei-Poku, G.; Gauthier, D.; van Frankenhuyzen, K. Isolation and characterisation of Isaria farinosa and Purpureocillium lilacinum associated with emerald ash borer, Agrilus planipennis in canada. Biocontrol Sci. Technol. 2012, 22, 723–732. [Google Scholar] [CrossRef]
- Mustu, M.; Demirci, F.; Kocak, E. Mortality effects of Isaria farinosa (Holm.) and Beauveria bassiana (Balsamo) Vuillemin (Sordariomycetes: Hypocreales) on Aelia rostrata Boh. (Hemiptera: Pentatomidae). Turkiye Entomoloji Dergisi-Turkish 2011, 35, 559–568. [Google Scholar]
- Yang, S.; Zhuang, H.; Li, Y.; Kuang, R. Insecticidal efficacy of Isaria farinosa in different life stages of Pissodes punctatus (Coleoptera: Curculionidae). J. Pest Sci. 2009, 82, 321–325. [Google Scholar] [CrossRef]
- Liu, F.; Wu, X.L.; Liu, Y.; Chen, D.X.; Zhang, D.L.; Yang, D.J. Progress on molecular biology of Isaria farinosa, pathogen of host of ophiocordyceps sinensis during the artificial culture. China J. Chin. Mater. Med. 2016, 41, 403–409. [Google Scholar]
- Lv, Y.; Xia, J.; Zhang, Z.; Li, Q.; Dong, C.; Li, W. Symptoms, infection and histopathology of Hepialus sp. larvae parasitized by Isaria farinosa. Mycosystema 2018, 37, 314–324. [Google Scholar]
- Liu, F.; Xiang, M.; Guo, Y.; Wu, X.; Lu, G.; Yang, Y.; Liu, X.; Chen, S.; Zhang, G.; Shi, W. Culture conditions and nutrition requirements for the mycelial growth of Isaria farinosa (Hypocreales: Cordycipitaceae) and the altitude effect on its growth and metabolome. Sci. Rep. 2018, 8, 15623. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Brunner-Mendoza, C.; Navarro-Barranco, H.; Leon-Mancilla, B.; Perez-Torres, A.; Toriello, C. Biosafety of an entomopathogenic fungus Isaria fumosorosea in an acute dermal test in rabbits. Cutan. Ocul. Toxicol. 2017, 36, 12–18. [Google Scholar] [CrossRef]
- Avery, P.B.; Hunter, W.B.; Hall, D.G.; Jackson, M.A.; Powell, C.A. Efficacy of topical application, leaf residue or soil drench of blastospores of Isaria fumosorosea for citrus root weevil management: Laboratory and greenhouse investigations. Insects 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Meng, L.-L.; Wei, J.-J.; Fan, P.; Liu, S.-S.; Yuan, W.-Y.; Zhao, Y.-X.; Luo, D.-Q. PTP1B inhibitors from the entomogenous fungi Isaria fumosorosea. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Wang, Q.G.; Xu, L.J. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-L.; Lin, H.-I.; Chen, B.-F.; Jow, G.-M. Beauvericin-induced cell apoptosis through the mitogen-activated protein kinase pathway in human nonsmall cell lung cancer A549 cells. J. Toxicol. Sci. 2016, 41, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.W.; Lin, Y.C.; She, Z.G.; Lin, M.T.; Chen, P.X.; Zhang, J.Y. Anticancer activity and mechanism investigation of beauvericin isolated from secondary metabolites of the mangrove endophytic fungi. Anticancer Agents Med. Chem. 2015, 15, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Kuca, K.; Patocka, J.; Wu, Q. Biological activities of beauvericin, a mini review. Mini. Rev. Med. Chem. 2018. [Google Scholar] [CrossRef]
- Panasiuk, L.; Jedziniak, P.; Pietruszka, K.; Piatkowska, M.; Bocian, L. Frequency and levels of regulated and emerging mycotoxins in silage in poland. Mycotoxin Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ojuri, O.T.; Ezekiel, C.N.; Sulyok, M.; Ezeokoli, O.T.; Oyedele, O.A.; Ayeni, K.I.; Eskola, M.K.; Sarkanj, B.; Hajslova, J.; Adeleke, R.A.; et al. Assessing the mycotoxicological risk from consumption of complementary foods by infants and young children in nigeria. Food Chem. Toxicol. 2018, 121, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In vitro mechanisms of beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef]
- Madariaga-Mazon, A.; Gonzalez-Andradeb, M.; Toriello, C.; Navarro-Barranco, H.; Mata, R. Potent anti-calmodulin activity of cyclotetradepsipeptides isolated from Isaria fumosorosea using a newly designed biosensor. Nat. Prod. Commun. 2015, 10, 113–116. [Google Scholar]
- Jegorov, A.; Sedmera, P.; Matha, V.; Simek, P.; Zahradnickova, H.; Landa, Z.; Eyal, J. Beauverolides L and La from Beauveria tenella and Paecilomyces fumosoroseus. Phytochemistry 1994, 37, 1301–1303. [Google Scholar] [CrossRef]
- Vilcinskas, A.; Jegorov, A.; Landa, Z.; Gotz, P.; Matha, V. Effects of beauverolide L and cyclosporin A on humoral and cellular immune response of the greater wax moth, Galleria mellonella. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1999, 122, 83–92. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Chen, C.; Teng, J.; Wang, C.; Luo, D. Structure and biosynthesis of fumosorinone, a new protein tyrosine phosphatase 1B inhibitor firstly isolated from the entomogenous fungus Isaria fumosorosea. Fungal Genet. Biol. 2015, 81, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xue, T.; Fan, P.; Meng, L.; Wei, J.; Luo, D. Cytotoxic activity of SHP2 inhibitor fumosorinone in human cancer cells. Oncol. Lett. 2018, 15, 10055–10062. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Pramjit, S.; Pakawatchai, C.; Isaka, M.; Supothina, S. 10-membered macrolides from the insect pathogenic fungus Cordyceps militaris BCC 2816. J. Nat. Prod. 2004, 67, 1953–1955. [Google Scholar] [CrossRef] [PubMed]
- Asaff, A.; Cerda-Garcia-Rojas, C.; de la Torre, M. Isolation of dipicolinic acid as an insecticidal toxin from Paecilomyces fumosoroseus. Appl. Microbiol. Biotechnol. 2005, 68, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Asaff, A.; Cerda-Garcia-Rojas, C.M.; Viniegra-Gonzalez, G.; de la Torre, M. Carbon distribution and redirection of metabolism in Paecilomyces fumosoroseus during solid-state and liquid fermentations. Process. Biochem. (Amst., The Neth.) 2006, 41, 1303–1310. [Google Scholar] [CrossRef]
- Martinez-Espla, A.; Serrano, M.; Martinez-Romero, D.; Valero, D.; Zapata, P.J. Oxalic acid preharvest treatment increases antioxidant systems and improves plum quality at harvest and during postharvest storage. J. Sci. Food Agric. 2018. [Google Scholar] [CrossRef] [PubMed]
- Valero, D.; Diaz-Mula, H.M.; Zapata, P.J.; Castillo, S.; Guillen, F.; Martinez-Romero, D.; Serrano, M. Postharvest treatments with salicylic acid, acetylsalicylic acid or oxalic acid delayed ripening and enhanced bioactive compounds and antioxidant capacity in sweet cherry. J. Agric. Food Chem. 2011, 59, 5483–5489. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.M.; Yin, W.Q.; Liu, M.M.; Wang, C.L.; Guo, S.X. Oxalic acid and sclerotial differentiation of Polyporus umbellatus. Sci. Rep. 2015, 5, 10759. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y. Traditional uses, phytochemistry, pharmacology, pharmacokinetics and quality control of Polyporus umbellatus (Pers.) fries: A review. J. Ethnopharmacol. 2013, 149, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.S.; Yuan, W.Y.; Wei, J.J.; Zhao, Y.X.; Luo, D.Q. Carotane-type sesquiterpenes from cultures of the insect pathogenic fungus Isaria fumosorosea. J. Asian Nat. Prod. Res. 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, J.; Hu, F.; Wang, X.; Wang, X.; Li, Z.; Fan, M. Isolation and purification of peroxy-ergosterol from Paecilomyces fumosoroseus by high-speed-counter-current chromatography and identification by ESI-MS. Food Ferment. Ind. 2009, 35, 14–17. [Google Scholar]
- Sheu, J.H.; Chang, K.C.; Duh, C.Y. A cytotoxic 5alpha,8alpha-epidioxysterol from a soft coral Sinularia species. J. Nat. Prod. 2000, 63, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Takei, T.; Yoshida, M.; Ohnishi-Kameyama, M.; Kobori, M. Ergosterol peroxide, an apoptosis-inducing component isolated from Sarcodon aspratus (berk.) s. Ito. Biosci. Biotechnol. Biochem. 2005, 69, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, Y.; Liu, X.; Liang, Z. Screening of vitamin a-praducing fungal resources in soil. Guizhou Agric. Sci. 2013, 41, 99–101. [Google Scholar]
- Yanagawa, A.; Imai, T.; Akino, T.; Toh, Y.; Yoshimura, T. Olfactory cues from pathogenic fungus affect the direction of motion of termites, Coptotermes formosanus. J. Chem. Ecol. 2015, 41, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Tian, M.Y.; He, Y.R.; Bland, J.M.; Gu, W.X. Behavioral and electrophysiological responses of Coptotermes formosanus Shiraki towards entomopathogenic fungal volatiles. Biol. Control 2010, 55, 166–173. [Google Scholar] [CrossRef]
- Bojke, A.; Tkaczuk, C.; Stepnowski, P.; Golebiowski, M. Comparison of volatile compounds released by entomopathogenic fungi. Microbiol. Res. 2018, 214, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Liu, H.; Liu, X.; Che, Y. Cycloaspeptides F and G, cyclic pentapeptides from a Cordyceps-colonizing isolate of Isaria farinosa. J. Nat. Prod. 2009, 72, 1364–1367. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Hormazabal, E.; Rodriguez, J.A.; Theoduloz, C. Cycloaspeptide a and pseurotin a from the endophytic fungus penicillium janczewskii. Z Naturforsch C 2008, 63, 383–388. [Google Scholar] [CrossRef]
- de Mattos-Shipley, K.M.J.; Greco, C.; Heard, D.M.; Hough, G.; Mulholland, N.P.; Vincent, J.L.; Micklefield, J.; Simpson, T.J.; Willis, C.L.; Cox, R.J.; et al. The cycloaspeptides: Uncovering a new model for methylated nonribosomal peptide biosynthesis. Chem. Sci. 2018, 9, 4109–4117. [Google Scholar] [CrossRef]
- Prachyawarakorn, V.; Mahidol, C.; Sureram, S.; Sangpetsiripan, S.; Wiyakrutta, S.; Ruchirawat, S.; Kittakoop, P. Diketopiperazines and phthalides from a marine derived fungus of the order pleosporales. Planta Med. 2008, 74, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Li, Y.; Niu, S.; Zhang, H.; Liu, X.; Che, Y. N-hydroxypyridones, phenylhydrazones, and a quinazolinone from Isaria farinosa. J. Nat. Prod. 2011, 74, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Gunther, W.; Stoyanova, S.; Schubert, B.; Li, Z.Z.; Hamburger, M. Militarinone a, a neurotrophic pyridone alkaloid from Paecilomyces militaris. Organ. Lett. 2002, 4, 197–199. [Google Scholar] [CrossRef]
- Tsang, T.H.; Gubler, D.A. Total synthesis of farylhydrazones a and b. Tetrahedron Lett. 2012, 53, 4243–4244. [Google Scholar] [CrossRef]
- Cheng, Y.; Schneider, B.; Riese, U.; Schubert, B.; Li, Z.; Hamburger, M. (+)-n-deoxymilitarinone a, a neuritogenic pyridone alkaloid from the insect pathogenic fungus Paecilomyces farinosus. J. Nat. Prod. 2006, 69, 436–438. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; William, R.; Leow, M.L.; Chai, H.; Fong, J.Z.; Liu, X.W. Directed orthometalation and the asymmetric total synthesis of n-deoxymilitarinone A and torrubiellone B. Organ. Lett. 2014, 16, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Putri, S.P.; Kinoshita, H.; Ihara, F.; Igarashi, Y.; Nihira, T. Farinomalein, a maleimide-bearing compound from the entomopathogenic fungus Paecilomyces farinosus. J. Nat. Prod. 2009, 72, 1544–1546. [Google Scholar] [CrossRef] [PubMed]
- Miles, W.H.; Yan, M. Synthesis of farinomalein. Tetrahedron Lett. 2010, 51, 1710–1712. [Google Scholar] [CrossRef]
- Putri, S.P.; Ishido, K.; Kinoshita, H.; Kitani, S.; Ihara, F.; Sakihama, Y.; Igarashi, Y.; Nihira, T. Production of antioomycete compounds active against the phytopathogens Phytophthora sojae and Aphanomyces cochlioides by clavicipitoid entomopathogenic fungi. J. Biosci. Bioeng. 2014, 117, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.X.; Schneider, B.; Riese, U.; Schubert, B.; Li, Z.Z.; Hamburger, M. Farinosones A-C, neurotrophic alkaloidal metabolites from the entomogenous Deuteromycete Paecilomyces farinosus. J. Nat. Prod. 2004, 67, 1854–1858. [Google Scholar] [CrossRef] [PubMed]
- Lang, G.; Blunt, J.W.; Cummings, N.J.; Cole, A.L.; Munro, M.H. Paecilosetin, a new bioactive fungal metabolite from a new zealand isolate of Paecilomyces farinosus. J. Nat. Prod. 2005, 68, 810–811. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Vijayakumar, E.K.; Mukhopadhyay, T.; Chatterjee, S.; Bhat, R.G.; Blumbach, J.; Ganguli, B.N. Aranorosinol A and aranorosinol B, two new metabolites from pseudoarachniotus roseus: Production, isolation, structure elucidation and biological properties. J. Antibiot. (Tokyo) 1992, 45, 1592–1598. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, P.; Lee, Y.H.; Nanthakumar, K.; Kamala-Kannan, S.; Dufosse, L.; Mapari, S.A.S.; Oh, B.-T. Water-soluble red pigments from Isaria farinosa and structural characterization of the main colored component. J. Basic Microbiol. 2010, 50, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Akilandeswari, P.; Pradeep, B.V. Exploration of industrially important pigments from soil fungi. Appl. Microbiol. Biotechnol. 2016, 100, 1631–1643. [Google Scholar] [CrossRef]
- Genthner, F.J.; Chancy, C.A.; Couch, J.A.; Foss, S.S.; Middaugh, D.P.; George, S.E.; Warren, M.A.; Bantle, J.A. Toxicity and pathogenicity testing of the insect pest control fungus Metarhizium anisopliae. Arch. Environ. Contam. Toxicol. 1998, 35, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Melnikova, E.A.; Murza, V.I. Investigation of the safety of industrial strains of microorganisms and microbial insecticides. J. Hyg. Epidemiol. Microbiol. Immunol. 1980, 24, 425–431. [Google Scholar]
- Smagghe, G.; De Meyer, L.; Meeus, I.; Mommaerts, V. Safety and acquisition potential of Metarhizium anisopliae in entomovectoring with bumble bees, Bombus terrestris. J. Econ. Entomol. 2013, 106, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Haas-Costa, J.; Alves, L.F.A.; Daros, A.A. Safety of Beauveria bassiana (Bals.) Vuill. to Gallus domesticus L. Braz. Arch. Biol. Technol. 2010, 53, 465–471. [Google Scholar] [CrossRef]
- Hu, Q.; Li, F.; Zhang, Y. Risks of mycotoxins from mycoinsecticides to humans. Biomed. Res. Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Liang, L.; Hao, C.; Ma, R. The study on the acute toxicity and dermal sensitization of Isaria fumosorosea. J. Shanxi Agric. Univ. (Nat. Sci. Ed.) 2013, 33, 144–146. [Google Scholar]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [PubMed]
- Mantzoukas, S.; Chondrogiannis, C.; Grammatikopoulos, G. Effects of three endophytic entomopathogens on sweet sorghum and on the larvae of the stalk borer Sesamia nonagrioides. Entomol. Exp. Appl. 2015, 154, 78–87. [Google Scholar] [CrossRef]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Scorsetti, A.C. Endophytic colonisation of tobacco, corn, wheat and soybeans by the fungal entomopathogen Beauveria bassiana (Ascomycota, hypocreales). Biocontrol Sci. Technol. 2015, 25, 475–480. [Google Scholar] [CrossRef]
- Parsa, S.; Ortiz, V.; Vega, F.E. Establishing fungal entomopathogens as endophytes: Towards endophytic biological control. Jove-J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Yang, L.; Qiu, X.; Liu, Y.G.; Zhou, W.; Wan, Y.J. Diversity analysis of Beauveria bassiana isolated from infected silkworm in southwest china based on molecular data and morphological features of colony. World J. Microbiol. Biotechnol. 2013, 29, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Reynaldi, F.J.; Lucia, M.; Genchi Garcia, M.L. Ascosphaera apis, the entomopathogenic fungus affecting larvae of native bees (xylocopa augusti): First report in south america. Rev. Iberoam. Micol. 2015, 32, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Hernandez, R.A.; Ruiz-Toledo, J.; Toledo, J.; Sanchez, D. Effect of three entomopathogenic fungi on three species of stingless bees (Hymenoptera: Apidae) under laboratory conditions. J. Econ. Entomol. 2016. [Google Scholar] [CrossRef]
- Rannback, L.M.; Cotes, B.; Anderson, P.; Ramert, B.; Meyling, N.V. Mortality risk from entomopathogenic fungi affects oviposition behavior in the parasitoid wasp Trybliographa rapae. J. Invertebr. Pathol. 2015, 124, 78–86. [Google Scholar] [CrossRef]
- Yousef, M.; Alba-Ramirez, C.; Garrido Jurado, I.; Mateu, J.; Raya Diaz, S.; Valverde-Garcia, P.; Quesada-Moraga, E. Metarhizium brunneum (Ascomycota; Hypocreales) treatments targeting olive fly in the soil for sustainable crop production. Front. Plant Sci. 2018, 9, 1. [Google Scholar] [CrossRef]
- Herrick, N.J.; Cloyd, R.A. Direct and indirect effects of pesticides on the insidious flower bug (Hemiptera: Anthocoridae) under laboratory conditions. J. Econ. Entomol. 2017, 110, 931–940. [Google Scholar] [CrossRef]
- Fan, M.; Li, J.; Guo, C.; Hashan, E. Persistent forms and survival potential of Metarhizium anisopliae in soil. J. Northwestern Coll. For. 1991, 6, 48–54. [Google Scholar]
- Li, Y.; Hu, Z.; Hu, Z.; Xiao, H.; Li, H. Survival dynamics of Beauveria bassiana BB09 in the muscardine cadaver of Hyphantria cunea and surrounding soil. For. Ecol. Sci. 2018, 33, 172–177. [Google Scholar]
- Zhang, Y.; Wu, X.; Ye, B.; Wang, H.; Shu, J. Inhibition effects of soil bacteria on conidium germination of Metarhizium pingshaense. Chin. J. Biol. Control 2017, 33, 788–795. [Google Scholar]
- Wu, X.; Zhang, Y.; Wu, P.; Ye, B.; Wang, H.; Shu, J. Influence of temperature, moisture, and soil type on conidium germination of Metarhizium pingshaense in soil. Chin. J. Biol. Control 2014, 30, 766–771. [Google Scholar]
- Biswas, C.; Dey, P.; Gotyal, B.S.; Satpathy, S. A method of multiplex pcr for detection of field released Beauveria bassiana, a fungal entomopathogen applied for pest management in jute (Corchorus olitorius). World J. Microbiol. Biotechnol. 2015, 31, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Luz, C.; Tai, M.H.; Santos, A.H.; Rocha, L.F.; Albernaz, D.A.; Silva, H.H. Ovicidal activity of entomopathogenic hyphomycetes on Aedes aegypti (Diptera: Culicidae) under laboratory conditions. J. Med. Entomol. 2007, 44, 799–804. [Google Scholar] [CrossRef]
- Falvo, M.L.; Albornoz Medina, P.; Rodrigues, J.; Lopez Lastra, C.C.; Garcia, J.J.; Fernandes, E.K.K.; Luz, C. Effect of UV-B irradiation on water-suspended Metarhizium anisopliae s.l. (Hypocreales: Clavicipitaceae) conidia and their larvicidal activity in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2018, 55, 1330–1333. [Google Scholar] [CrossRef]
- Milner, R.J.; Lim, R.P.; Hunter, D.M. Risks to the aquatic ecosystem from the application of Metarhizium anisopliae for locust control in australia. Pest Manag. Sci. 2002, 58, 718–723. [Google Scholar] [CrossRef]
- Shah, P.A.; Pell, J.K. Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef]
- Dash, C.K.; Bamisile, B.S.; Keppanan, R.; Qasim, M.; Lin, Y.; Islam, S.U.; Hussain, M.; Wang, L. Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb. Pathog. 2018, 125, 385–392. [Google Scholar] [CrossRef]
- Raya-Diaz, S.; Sanchez-Rodriguez, A.R.; Segura-Fernandez, J.M.; Del Campillo, M.D.C.; Quesada-Moraga, E. Entomopathogenic fungi-based mechanisms for improved fe nutrition in sorghum plants grown on calcareous substrates. PLoS ONE 2017, 12, e0185903. [Google Scholar] [CrossRef] [PubMed]
- Skrobek, A.; Shah, F.A.; Butt, T.M. Destruxin production by the entomogenous fungus Metarhizium anisopliae in insects and factors influencing their degradation. Biocontrol 2008, 53, 361–373. [Google Scholar] [CrossRef]
- Rios-Moreno, A.; Garrido-Jurado, I.; Raya-Ortega, M.C.; Quesada-Moraga, E. Quantification of fungal growth and destruxin a during infection of Galleria mellonella larvae by Metarhizium brunneum. J. Invertebr. Pathol. 2017, 149, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Oyedele, O.A.; Ezekiel, C.N.; Sulyok, M.; Adetunji, M.C.; Warth, B.; Atanda, O.O.; Krska, R. Mycotoxin risk assessment for consumers of groundnut in domestic markets in nigeria. Int. J. Food Microbiol. 2017, 251, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Miller, J.D. A concise history of mycotoxin research. J. Agric. Food Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef] [PubMed]
- Schenzel, J.; Forrer, H.R.; Vogelgsang, S.; Hungerbuhler, K.; Bucheli, T.D. Mycotoxins in the environment: I. Production and emission from an agricultural test field. Environ. Sci. Technol. 2012, 46, 13067–13075. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structures of secondary metabolites (SMs) isolated from Isaria fumosorosea.

Figure 2.
Structures of volatile organic compounds (VOCs) from Isaria fumosorosea.

Figure 3.
Structures of secondary metabolites (SMs) isolated from Isaria farinosa.

Figure 4.
The sources and destinations of the Isaria myco-insecticide and its mycotoxins. ![Molecules 24 00664 i001]() indicates an existing pathway, while
indicates an existing pathway, while ![Molecules 24 00664 i002]() indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).
indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).
 indicates an existing pathway, while
indicates an existing pathway, while  indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).
indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).
Figure 4.
The sources and destinations of the Isaria myco-insecticide and its mycotoxins. ![Molecules 24 00664 i001]() indicates an existing pathway, while
indicates an existing pathway, while ![Molecules 24 00664 i002]() indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).
indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).
indicates an existing pathway, while indicates a pathway that has not been found to date. (SMs = secondary metabolites) (modified based on Hu et al. (2016) [81]).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
SMs from Isaria fumosorosea and their biological activities.
| Metabolite | CAS no. | Strain | Biological activity | References |
|---|---|---|---|---|
| Beauvericin (1) | 26048-05-5 | ACCC37775 (Hebei University, Baoding, China) | Inhibiting HepG2 cells with an IC50 of 2.40 μM. Cytotoxicity to multiple drug-resistant HepG2 cell lines with an IC50 value 25-fold more than that of doxorubicin. Inhibitor of PTP1B with an IC50 of 0.59 μM. | [32,33,34,35,36] |
| Beauverolide C (2) | 75899-64-8 | BMFM-UNAM 834 (Universidad Nacional Autonoma de Mexico, Mexico City, Mexico). | Calmodulin (CaM) inhibitor | [40] |
| Beauverolide F (3) | 75947-00-1 | Same as above | Calmodulin (CaM) inhibitor | [40] |
| Beauverolide I (4) | 62995-91-9 | Same as above | Calmodulin (CaM) inhibitor | [40] |
| Beauverolide Ja (5) | 76265-41-3 | Same as above | Calmodulin (CaM) inhibitor | [40] |
| Beauverolide L (6) | 154491-56-2 | BMFM-UNAM 834 (Universidad Nacional Autonoma de Mexico, Mexico City, Mexico); PFR97-Apopka (ATCC 20874) (WR Grace & Co, Conn, MD, USA) | Calmodulin (CaM) inhibitor Anti-immunity activity against the greater wax moth, Galleria mellonella | [40,41,42] |
| Beauverolide La (7) | 160825-68-3 | PFR97-Apopka (ATCC 20874) (WR Grace & Co, Conn, MD, USA) | [41] | |
| Beauverolide M (8) | BMFM-UNAM 834 (Universidad Nacional Autonoma de Mexico, Mexico City, Mexico). | Calmodulin (CaM) inhibitor | [40] | |
| Beauverolide N (9) | Same as above | Calmodulin (CaM) inhibitor | [40] | |
| Fumosorinone (10) | 1879030-70-2 | ACCC37775 (Hebei University, Baoding, China) | Inhibitor of PTP1B (IC50 of 14.04 μM) | [43,44] |
| Fumosorinone A (11) | 2241028-99-7 | Same as above | Inhibitor of PTP1B (IC50 of 3.24 μM) | [32] |
| Cepharosporolide C (12) | 97344-02-0 | Same as above | No activities to malaria Plasmodium falciparum K1, and PTP1B | [32,45] |
| Cepharosporolide E (13) | 97373-15-4 | Same as above | [32,45] | |
| Cepharosporolide F (14) | 97344-04-2 | Same as above | [32,45] | |
| 2-carboxymethyl-4-(3′-hydroxybutyl)furan (15), | Same as above | [32,45] | ||
| Dipicolinic acid (16) | 499-83-2 | Pfrd (Centro Nacional de Referencia de Control Biológico, Tecomán, Colima, Mexico) | Insecticidal activity against third-instar whitefly nymphs | [47,48,49,50] |
| Oxalic acid (OXA) (17) | 144-62-7 | Same as above | Insecticidal activity against third-instar whitefly nymphs | [47,48,49,50] |
| Trichocarane E (18) | ACCC37775 (Hebei University, Baoding, China) | Cytotoxicity to six tumor cell lines (i.e., MDA, MCF-7, SKOV-3, Hela, A549, and HepG2) with an IC50 of 0.1–6.0 μg/mL. | [52] | |
| Trichocarane F (19) | Same as above | Cytotoxicity to six tumor cell lines (i.e., MDA, MCF-7, SKOV-3, Hela, A549, and HepG2, with an IC50 of 0.1–6.0 μg/mL. | [52] | |
| CAF-603 (20) | Same as above | Cytotoxicity to six tumor cell lines (i.e., MDA, MCF-7, SKOV-3, Hela, A549, and HepG2, with an IC50 of 0.1–6.0 μg/mL. | [52] | |
| Trichocarane C (21) | Same as above | [52] | ||
| Ergosterol peroxide (22) | 2061-64-5 | RCEF1253 (Anhui Agricultural University, Hefei, China) | Cytotoxic to cancer cells P-388, KB, A549, and HT-29 (with ED50 values of 0.4, 2.1, 2.7, and 1.4 μg/mL) and human leukemia cell, HL-60 | [53,54,55,56] |
| 3-octanone (23) | 106-68-3 | Conidia of strain K3 (Kyoto University, Kyoto, Japan). | Repellent to termites | [57] |
| 1-octen-3-ol (24) | 3391-86-4 | Same as above | Repellent to termites | [57] |
Table 2.
Secondary metabolites (SMs) from Isaria farinosa and their biological activities.
| Metabolite | CAS no. | Strain | Biological Activity | References |
|---|---|---|---|---|
| Cycloaspeptide F (25) | 1174132-23-0 | XJC04-CT-303 (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) | Cytotoxic to HeLa and MCF7 cell lines | [60] |
| Cycloaspeptide G (26) | 1174132-24-1 | Same as above | Cytotoxic to HeLa and MCF7 cell lines | [60] |
| Cycloaspeptide A (27) | 109171-13-3 | Same as above | Cytotoxicity to human lung fibroblasts | [60,61] |
| Cycloaspeptide C (28) | 109171-15-5 | Same as above | [60] | |
| (3R,6R)-Bisdethiodi (methylthio) hyalodendrin (29) | 52080-06-5 | Same as above | Weak cytotoxic activity | [60,63] |
| Militarinone A (30) | 400604-05-9 | Same as above | Cytotoxicity to A549 cells. Neurotrophic effects on PC-12 cells | [64,65] |
| Militarinone B (31) | 503584-83-6 | RCEF0097 (Anhui Agricultural University, Hefei, China); XJC04-CT-303 (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) | Anti-microbes to Staphylococcus aureus, Streptococcus pneumoniae, and Candida albicans | [64] |
| Militarinone E (32) | 1261060-55-2 | XJC04-CT-303 (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) | Cytotoxicity to A549 cells | [64] |
| Militarinone F (33) | 1261060-56-3 | Same as above | [64] | |
| Farylhydrazone A (34) | 1261060-57-4 | Same as above | [66] | |
| Farylhydrazone B (35) | 1261060-58-5 | Same as above | [66] | |
| (+)-N-deoxymilitarinone A (36) | 881376-40-5 | RCEF0097 (Anhui Agricultural University, Hefei, China) | Induce neurite sprouting in PC 12 cells when tested at 33 and 100 μM concentrations. Cytotoxic to human neurons (IMR-32) at a concentration of 100 μM. | [67] |
| Militarinone D (37) | 503584-82-5 | RCEF 0097 (Anhui Agricultural University, Hefei, China); XJC04-CT-303 (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) | [67] | |
| (22E)-Ergosta-7,22-diene-3β,5α,6β,9α-tetraol (38) | 88191-06-4 | Same as above | [67] | |
| Farinomalein (39) | 1175521-35-3 | HF599 (National Institute of Fruit Tree Science, Tsukuba, Japan) | Antifungal to phytopathogenic Phytophthora sojae | [69,70,71] |
| Farinosone A (40) | 816431-89-7 | RCEF 0101 (Anhui Agricultural University, Hefei, China) | Neuritogenic in the PC-12 cell model | [72] |
| Farinosone B (41) | 816431-94-4 | Same as above | Inhibitory to Bacillus subtilis and Staphylococcus aureus. Moderate cytotoxicity to brine shrimp larvae (Artemia salina) | [72] |
| Farinosone C (42) | 816431-98-8 | Same as above | Induced neurite outgrowth in the PC-12 cell line at concentrations of 50 μM | [72] |
| Paecilosetin (43) | 856258-89-4 | CANU TE108 (University of Canterbury, Christchurch, New Zealand). HF511 (National Institute of Fruit Tree Science, Tsukuba, Japan.) | Antioomycete activity against both Phytophthora sojae and Aphanomyces cochlioides | [71,73] |
| Aranorosinol A (44) | 145147-04-2 | HF511 (National Institute of Fruit Tree Science, Tsukuba, Japan.) | Antioomycete to both Phytophthora sojae and Aphanomyces cochlioides | [71,74] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Weng, Q.; Zhang, X.; Chen, W.; Hu, Q. Secondary Metabolites and the Risks of Isaria fumosorosea and Isaria farinosa. Molecules 2019, 24, 664. https://doi.org/10.3390/molecules24040664
AMA Style
Weng Q, Zhang X, Chen W, Hu Q. Secondary Metabolites and the Risks of Isaria fumosorosea and Isaria farinosa. Molecules. 2019; 24(4):664. https://doi.org/10.3390/molecules24040664
Chicago/Turabian StyleWeng, Qunfang, Xiaofeng Zhang, Wei Chen, and Qiongbo Hu. 2019. "Secondary Metabolites and the Risks of Isaria fumosorosea and Isaria farinosa" Molecules 24, no. 4: 664. https://doi.org/10.3390/molecules24040664