
Huperzine A and Huperzine B Production by Prothallus Cultures of Huperzia selago (L.) Bernh. ex Schrank et Mart



Abstract
:1. Introduction
2. Results
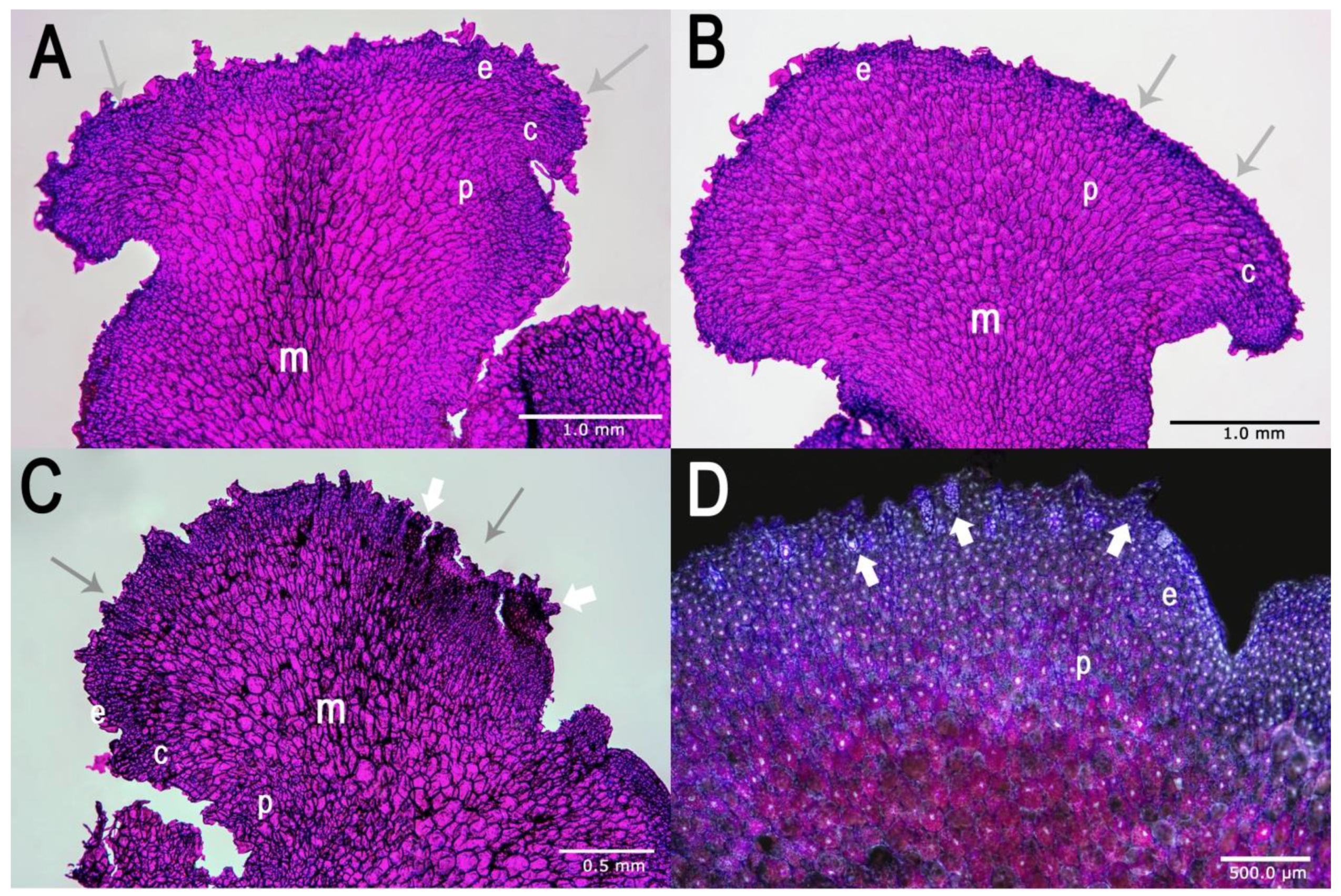
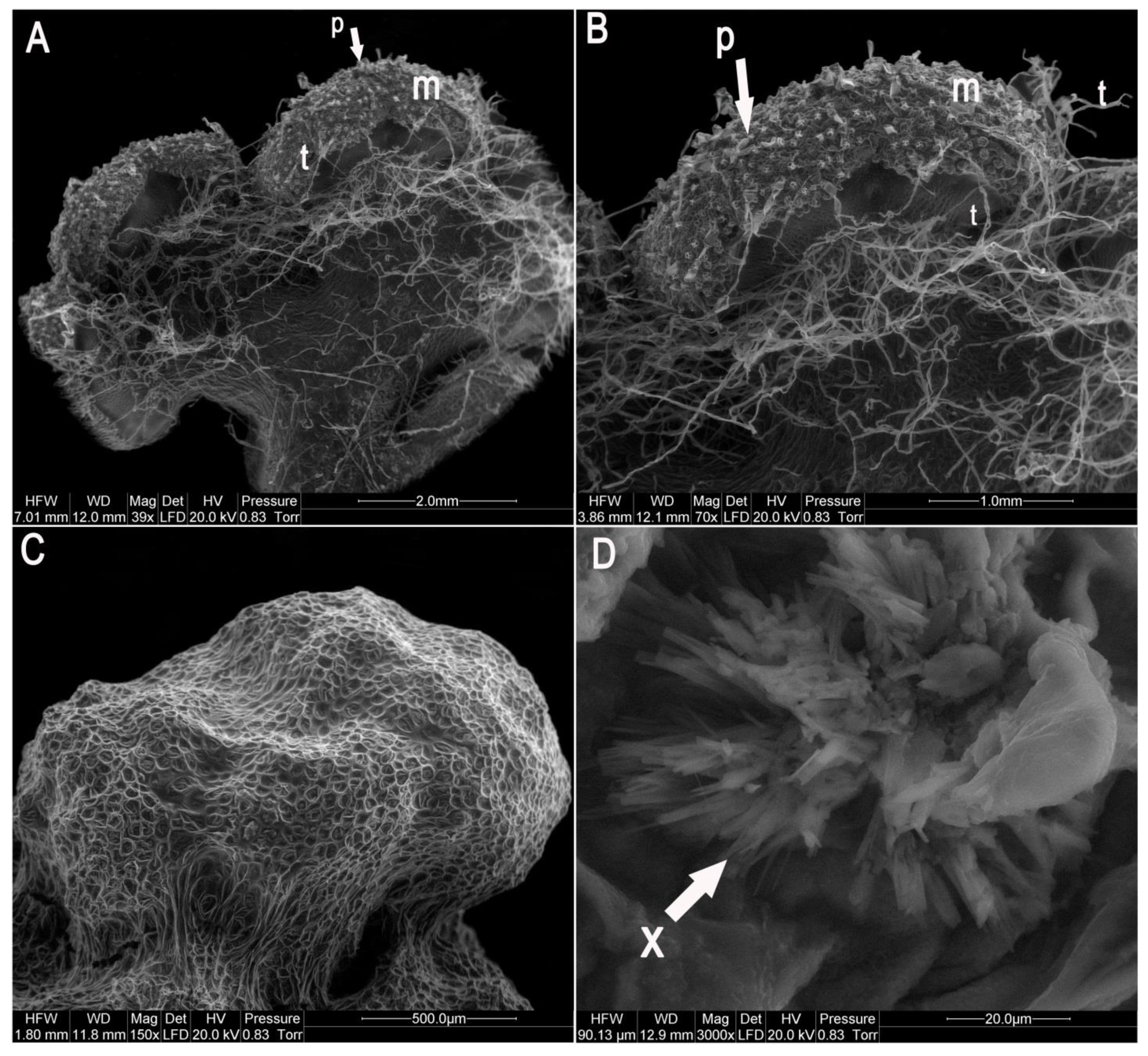
2.1. Establishment of In Vitro Culture and Morphological Characteristics of Gametophytes
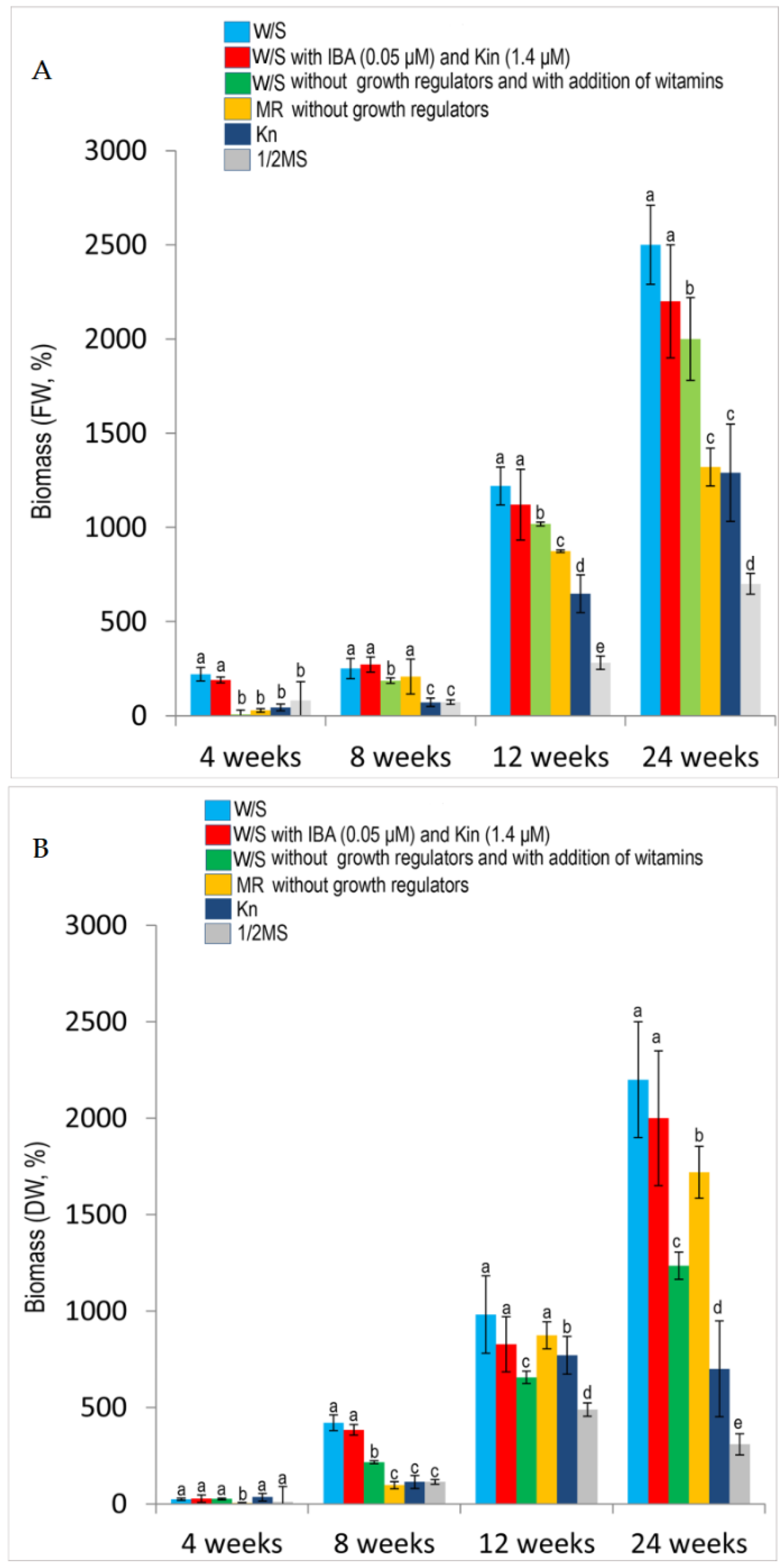
2.2. Effects of Media and Growth Regulators on Cell and Biomass Growth
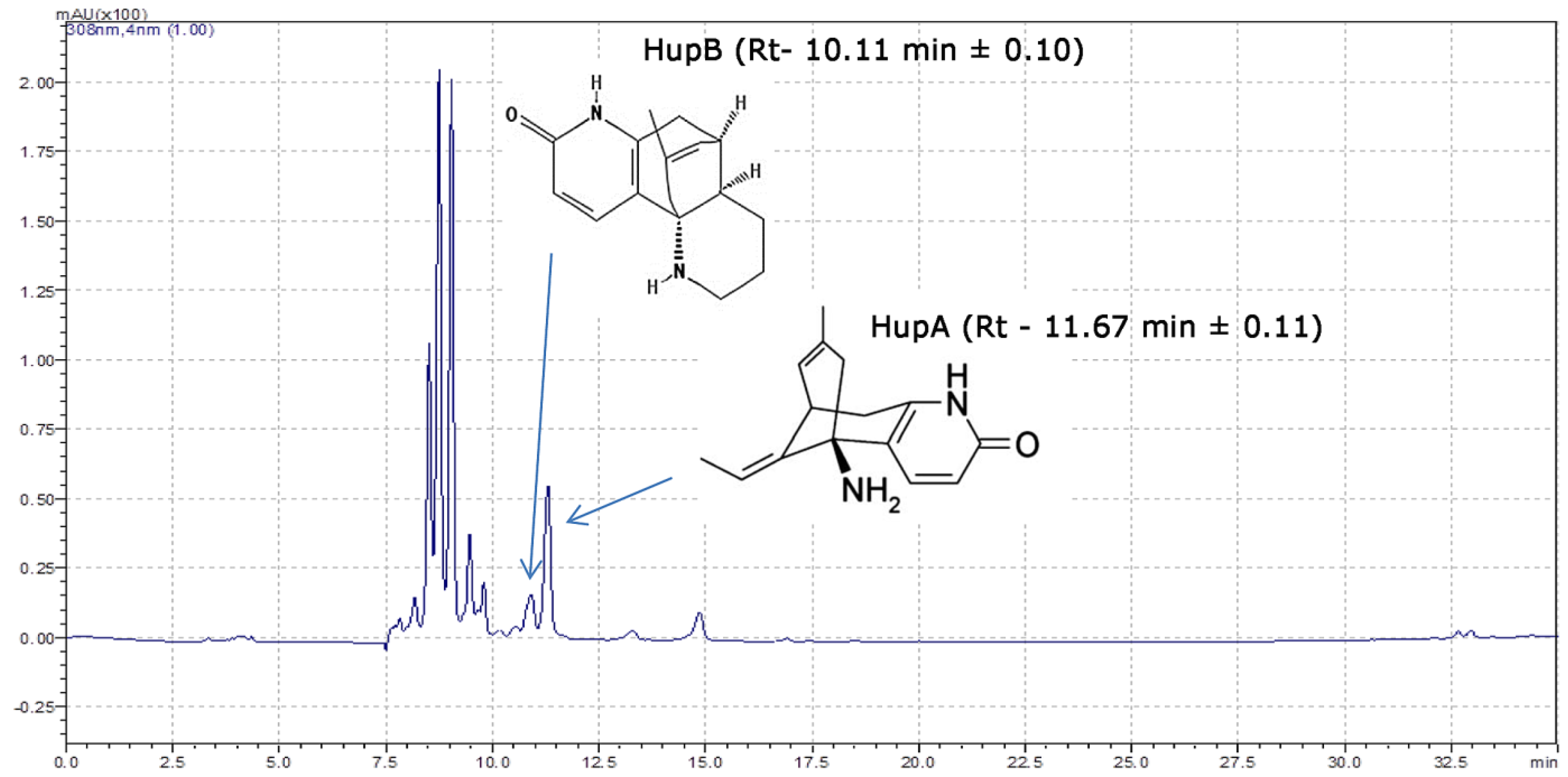
2.3. Characterization of Huperzia Selago Alkaloids from Gametophytes Using HPLC-DAD and Mass Spectrometry
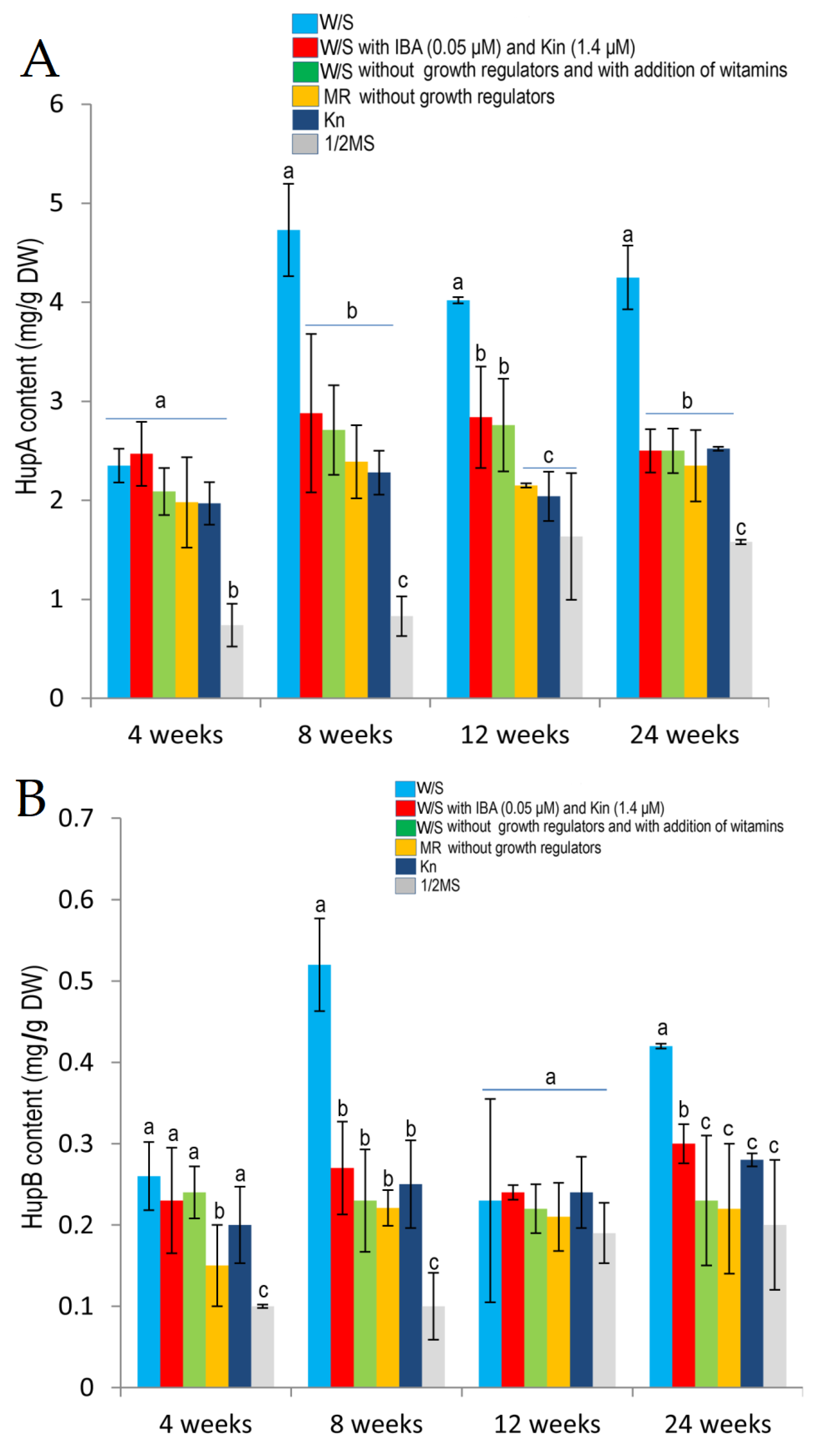
2.4. Effects of Different Media and Their Composition on Huperzine A and Huperzine B Production
3. Discussion and Conclusions
4. Materials and Methods
4.1. Reagents and Standards
4.2. Plant Material and Establishment of In Vitro Culture
4.3. Morphological Analysis of Huperzia Selago Prothallus Cultures
4.4. Histological Analysis
4.5. Alkaloid Extraction and Determination
4.5.1. Preparation of Extracts and Their Purification
4.5.2. Qualitative and Quantitative HPLC-DAD Analyses of HupA and HupB
4.5.3. LC-MS Analysis of HupA and HupB
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Valenta, Z.; Yoshimura, H.; Rogers, E.; Ternbah, M.; Wiesner, K. The structure of selagine. Tetrahedron Lett. 1960, 1, 26–33. [Google Scholar] [CrossRef]
- Yoshimura, H.; Valenta, Z.; Wiesner, K. A rigorous proof of the selagine structure. Tetrahedron Lett. 1960, 1, 14–17. [Google Scholar] [CrossRef]
- Ayer, W.A.; Browne, L.M.; Orszanska, H.; Valenta, Z.; Liu, J.-S. Alkaloids of Lycopodiumselago. On the identity of selagine with huperzine A and the structure of a related alkaloid. Can. J. Chem. 1989, 67, 1538–1540. [Google Scholar] [CrossRef]
- Liu, J.-S.; Zhu, Y.-L.; Yu, C.-M.; Zhou, Y.-Z.; Han, Y.-Y.; Wu, F.-W.; Qi, B.-F. The structures of huperzine A and B, two new alkaloids exhibiting marked anticholinesterase activity. Can. J. Chem. 1986, 64, 837–839. [Google Scholar] [CrossRef]
- Ma, X.; Gang, D.R. The Lycopodium alkaloids. Nat. Prod. Rep. 2004, 21, 752. [Google Scholar] [CrossRef]
- Ferreira, A.; Rodrigues, M.; Fortuna, A.; Falcão, A.; Alves, G. Huperzine A from Huperzia serrata: A review of its sources, chemistry, pharmacology and toxicology. Phytochem. Rev. 2014, 15, 51–85. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, C.Y.; Yan, H.; Wang, Z.F.; Tang, L.L.; Gao, X.; Tang, X.C. Potential therapeutic targets of huperzine A for Alzheimer’s disease and vascular dementia. Chem. Interactions 2008, 175, 396–402. [Google Scholar] [CrossRef]
- Zhang, H.Y. New insights into huperzine A for the treatment of Alzheimer’s disease. Acta Pharm. Sinic. 2012, 33, 1170–1175. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Yan, H.; Tang, X.C. Progress in studies of huperzine A, a natural cholinesterase inhibitor from Chinese herbal medicine1. Acta Pharmacol. Sin. 2006, 27, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhang, H.; Tang, X.C. Huperzine A attenuates cognitive dysfunction and neuronal degeneration caused by β-amyloid protein-(1–40) in rat. Eur. J. Pharmacol. 2001, 421, 149–156. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, X.C. Neuroprotective effects of huperzine A: New therapeutic targets for neurodegenerative disease. Trends Pharmacol. Sci. 2006, 27, 619–625. [Google Scholar] [CrossRef]
- Feng, S.; Xia, Y.; Han, D.; Zheng, C.; He, X.; Tang, X.; Bai, D. Synthesis and acetylcholinesterase inhibition of derivatives of huperzine B. Bioorg. Med. Chem. Lett. 2005, 15, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Gong, K.; Yan, Y.; Zhang, L.; Tang, P.-F.; Zhang, X.; Gong, Y. Huperzine A promotes hippocampal neurogenesis in vitro and in vivo. Brain Res. 2013, 1506, 35–43. [Google Scholar] [CrossRef]
- Xu, H.; Tang, X.C. [Cholinesterase inhibition by huperzine B]. Zhongguo yao li xue bao = Acta Pharmacol. Sin. 1987, 8, 18–22. [Google Scholar]
- Ma, R.; Tan, C.-H.; Zhu, D.; Gang, D.R. A survey of potential huperzine A natural resources in China: The Huperziaceae. J. Ethnopharmacol. 2006, 104, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Tan, C.-H.; Zhu, D.; Gang, D.R.; Xiao, P. Huperzine A from Huperzia species—An ethnopharmacolgical review. J. Ethnopharmacol. 2007, 113, 15–34. [Google Scholar] [CrossRef]
- Szypuła, W.; Pietrosiuk, A.; Suchocki, P.; Olszowska, O.; Furmanowa, M.; Kazimierska, O. Somatic embryogenesis and in vitro culture of Huperzia selago shoots as a potential source of huperzine A. Plant Sci. 2005, 168, 1443–1452. [Google Scholar] [CrossRef]
- Wu, Q.; Gu, Y. Quantification of huperzine A in Huperzia serrata by HPLC-UV and identification of the major constituents in its alkaloid extracts by HPLC-DAD-MS-MS. J. Pharm. Biomed. Anal. 2006, 40, 993–998. [Google Scholar] [CrossRef]
- Ishiuchi, K.; Park, J.-J.; Long, R.M.; Gang, D.R. Production of huperzine A and other Lycopodium alkaloids in Huperzia species grown under controlled conditions and in vitro. Phytochemistry 2013, 91, 208–219. [Google Scholar] [CrossRef]
- Szypuła, W.J.; Kiss, A.K.; Pietrosiuk, A.; Świst, M.; Danikiewicz, W.; Olszowska, O. Determination of Huperzine A in Huperzia selago Plants from Wild Population and Obtained in In Vitro Culture by High-Performance Liquid Chromatography Using a Chaotropic Mobile Phase. Acta Chrom. 2011, 23, 339–352. [Google Scholar] [CrossRef]
- Borloz, A.; Marston, A.; Hostettmann, K. The determination of huperzine A in European Lycopodiaceae species by HPLC-UV-MS. Phytochem. Anal. 2006, 17, 332–336. [Google Scholar] [CrossRef]
- Lim, W.H.; Goodger, J.Q.D.; Field, A.R.; Holtum, J.A.M.; Woodrow, I.E. Huperzine alkaloids from Australasian and southeast Asian Huperzia. Pharm. Biol. 2010, 48, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szypuła, W.J.; Pietrosiuk, A. Production of Cholinesterase-Inhibiting Compounds in In Vitro Cultures of Club Mosses. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer International Publishing: Cham, Swizerland, 2019; pp. 1–40. [Google Scholar]
- Ma, X.; Gang, D.R. In vitro production of huperzine A, a promising drug candidate for Alzheimer’s disease. Phytochemistry 2008, 69, 2022–2028. [Google Scholar] [CrossRef]
- Szypuła, W.; Mistrzak, P.; Olszowska, O. A new and fast method to obtain in vitro cultures of Huperzia selago (Huperziaceae) sporophytes, a club moss which is a source of huperzine A. Acta Soc. Bot. Pol. 2013, 82, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Whittier, D.P.; Štorchová, H. The Gametophyte of Huperzia selago in Culture. Am. Fern J. 2007, 97, 149–154. [Google Scholar] [CrossRef]
- Freeberg, J.A.; Wetmore, R.H. Gametophytes of Lycopodium as grown in vitro. Phytomorphology 1957, 7, 204–217. [Google Scholar]
- Knudson, L. Nonsymbiotic Germination of Orchid Seeds. Int. J. Plant Sci. 1922, 73, 1–25. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Plant Physiol. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jerkowich, A.D.; Vivilecchia, R.V. Developmental of fast HPLC Methods. In HPLC for Pharmaceutical Scientists; Kazakevich, Y., Lobrutto, R., Eds.; Wiley-Interscience John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 783–810. [Google Scholar]
- Czapski, G.A.; Szypuła, W.; Kudlik, M.; Wilenska, B.; Kania, M.; Danikiewicz, W.; Adamczyk, A. Original article Assessment of antioxidative activity of alkaloids from Huperzia selago and Diphasiastrum complanatum using in vitro systems. Folia Neuropathol. 2014, 4, 394–406. [Google Scholar] [CrossRef] [Green Version]
- Lenkiewicz, A.; Czapski, G.A.; Jęśko, H.; Wilkaniec, A.; Szypuła, W.; Pietrosiuk, A.; Uszyńska, A.M.; Adamczyk, A. Potent effects of alkaloid-rich extract from Huperzia selago against sodium nitroprusside-evoked PC12 cells damage via attenuation of oxidative stress and apoptosis. Folia Neuropathol. 2016, 2, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Eames, A.J. Illustrations of Some Lycopodium Gametophytes. Am. Fern J. 1942, 32, 1. [Google Scholar] [CrossRef]
- Bruchmann, H. Das Prothallium von Lycopodium. Bot. Zentralblatt 1885, 21, 23–28. [Google Scholar]
- Bruchmann, H. Über die Prothallien und die Keimpfl anzen Mehrerer europäischer Lycopodien, und Zwar über die von Lycopodium clavatum, L. annotinum, L. complanatum und L. selago; Friedrich & Andreas Perthes: Gotha, Germany, 1898. [Google Scholar]
- Freeberg, J.A. The apogamous development of sporelings of Lycopodium cernuum L., L. complanatum var. flabelliforme Fernald and L. selago L. in vitro. Phytomorphology 1957, 7, 217–229. [Google Scholar]
- Freeberg, J.A. lycopodium prothalli and their endophytic fungi as studied in vitro. Am. J. Bot. 1962, 49, 530–535. [Google Scholar] [CrossRef]
- Whittier, D.P. Gametophytes of Lycopodium obscurum as grown in axenic culture. Can. J. Bot. 1977, 55, 563–567. [Google Scholar] [CrossRef]
- Whittier, D.P. Gametophytes of Lycopodium digitatum (Formerly L. Complanatum var. flabelliforme) as Grown in Axenic Culture. Int. J. Plant Sci. 1981, 142, 519–524. [Google Scholar] [CrossRef]
- Whittier, P.; Webster, T.R. Gametophytes of Lycopodium lucidulum from Axenic Culture. Am. Fern J. 1986, 76, 48. [Google Scholar] [CrossRef]
- Atmane, N.; Blervacq, A.; Michaux-Ferriere, N.; Vasseur, J. Histological analysis of indirect somatic embryogenesis in the Marsh clubmoss Lycopodiella inundata (L.) Holub (Pteridophytes). Plant Sci. 2000, 156, 159–167. [Google Scholar] [CrossRef]
- Meijer, E.G.M.; Brown, D.C.W. Role of exogenous reduced nitrogen and sucrose in rapid high frequency somatic embryogenesis in Medicago sativa. Plant Cell Tissue Organ Cult. 1987, 10, 11–19. [Google Scholar] [CrossRef]
- Trigiano, R.N.; May, R.A.; Conger, B.V. Reduced nitrogen influences somatic embryo quality and plant regeneration from suspension cultures of orchardgrass. Vitr. Cell. Dev. Boil.-Anim. 1992, 28, 187–191. [Google Scholar] [CrossRef]
- Debeaujon, I.; Branchard, M. Somatic embryogenesis in Cucurbitaceae. Plant Cell Tissue Organ Cult. 1993, 34, 91–100. [Google Scholar] [CrossRef]
- Gaj, M.D. Factors Influencing Somatic Embryogenesis Induction and Plant Regeneration with Particular Reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Xu, M.; Heidmarsson, S.; Thorsteinsdottir, M.; Wąsowicz, P.; Sun, H.; Deng, T.; Omarsdottir, S.; Olafsdottir, E.S. Infraspecific Variation of Huperzine A and B in Icelandic Huperzia selago Complex. Planta Med. 2018, 85, 160–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achmatowicz, O.; Rodewald, W. The alkaloids of Lycopodium selago. Bull. Acad. Polon. Sci. Classe III 1955, 3, 553–555. [Google Scholar]
- Achmatowicz, O.; Rodewald, W. Alkaloidy rodzaju Lycopodium. III. Alkaloidy Lycopodium selago L. Rocz. Chem. 1956, 30, 232–242. [Google Scholar]
- Rodewald, W.J.; Grynkiewicz, G. Alkaloidy rodzaju Lycopodium. VI. Alkaloidy Lycopodium selago L. Rocz. Chem. 1968, 42, 465–475. [Google Scholar]
- Ayer, W.A.; Kasitu, G.C. Some new Lycopodium alkaloids. Can. J. Chem. 1989, 67, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.-Q.; Jiang, S.-H.; Zhu, D.-Y. Alkaloid patterns in Huperzia and some related genera of Lyopodiaceae sensu lato occurring in China and their contribution to classification. Biochem. Syst. Ecol. 1998, 26, 723–728. [Google Scholar] [CrossRef]
- Stærk, D.; Larsen, J.; Larsen, L.A.; Olafsdottir, E.S.; Witt, M.; Jaroszewski, J.W.; Staerk, D. Selagoline, A new Alkaloid fromHuperzia Selago. Nat. Prod. Res. 2004, 18, 197–203. [Google Scholar] [CrossRef]
- Valentine, D.H.; Moore, D.M. Lycopodiaceae L. In Flora Europea, 2nd ed.; Tutin, T.G., Burges, N.A., Chater, A.O., Edmondson, J.R., Heywood, V.H., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; Volume I, pp. 1–4. [Google Scholar]
- Street, H.; Henshaw, G. Introduction and Methods Employed in Plant Tissue Culture. In Cells and Tissues in Culture Methods, Biology and Physiology; Elsevier BV: New York, NY, USA, 1966; Volume 3, pp. 459–532. [Google Scholar]
Sample Availability: Samples are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| An alkaloid Name | Formula | Theoretical Mass | Measured Mass | m/z Error (ppm) |
|---|---|---|---|---|
| Fawcettimine | C16H25NO2 | 264.1958 | 264.1962 | 1.51 |
| Deacetylfawcettine | C16H27NO2 | 266.2115 | 266.2109 | −2.25 |
| 6β-hydroxyhuperzine A | C15H18N2O2 | 259.1441 | 259.1431 | −3.86 |
| 16-hydroxyhuperzine B | C16H21N2O2 | 273.1598 | 273.1599 | 0.37 |
| Deacetyllycoclavine | C16H27NO2 | 266.2115 | 266.2109 | −2.25 |
| Huperzine B | C16H20N2O | 257.1648 | 257.1642 | −2.33 |
| Huperzine A | C15H18N2O | 243.1492 | 243.1481 | −4.52 |
| Serratinidine | C18H28N2O2 | 305.2224 | 305.2217 | −2.29 |
| Annopodine | C17H25NO3 | 292.1907 | 292.1901 | −2.05 |
| Lycopodine | C16H25NO | 248.2009 | 248.2002 | 2.82 |
| Selagoline | C16H25NO | 248.2009 | 248.1998 | −4.48 |
| Lycopecurine | C16H27NO | 250.2165 | 250.2156 | −3.60 |
| Des-N-methylfastigiatine | C18H26N2O | 287.2118 | 287.2113 | −1.74 |
| Flabelline | C18H29N2O | 289.2274 | 289.2275 | 0.35 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szypuła, W.J.; Wileńska, B.; Misicka, A.; Pietrosiuk, A. Huperzine A and Huperzine B Production by Prothallus Cultures of Huperzia selago (L.) Bernh. ex Schrank et Mart. Molecules 2020, 25, 3262. https://doi.org/10.3390/molecules25143262
Szypuła WJ, Wileńska B, Misicka A, Pietrosiuk A. Huperzine A and Huperzine B Production by Prothallus Cultures of Huperzia selago (L.) Bernh. ex Schrank et Mart. Molecules. 2020; 25(14):3262. https://doi.org/10.3390/molecules25143262
Chicago/Turabian StyleSzypuła, Wojciech J., Beata Wileńska, Aleksandra Misicka, and Agnieszka Pietrosiuk. 2020. "Huperzine A and Huperzine B Production by Prothallus Cultures of Huperzia selago (L.) Bernh. ex Schrank et Mart" Molecules 25, no. 14: 3262. https://doi.org/10.3390/molecules25143262