
A Multidisciplinary Approach to Coronavirus Disease (COVID-19)

 ,
,
Abstract
:
1. Introduction
2. Understanding the Mechanism and Structure of SARS-CoV-2
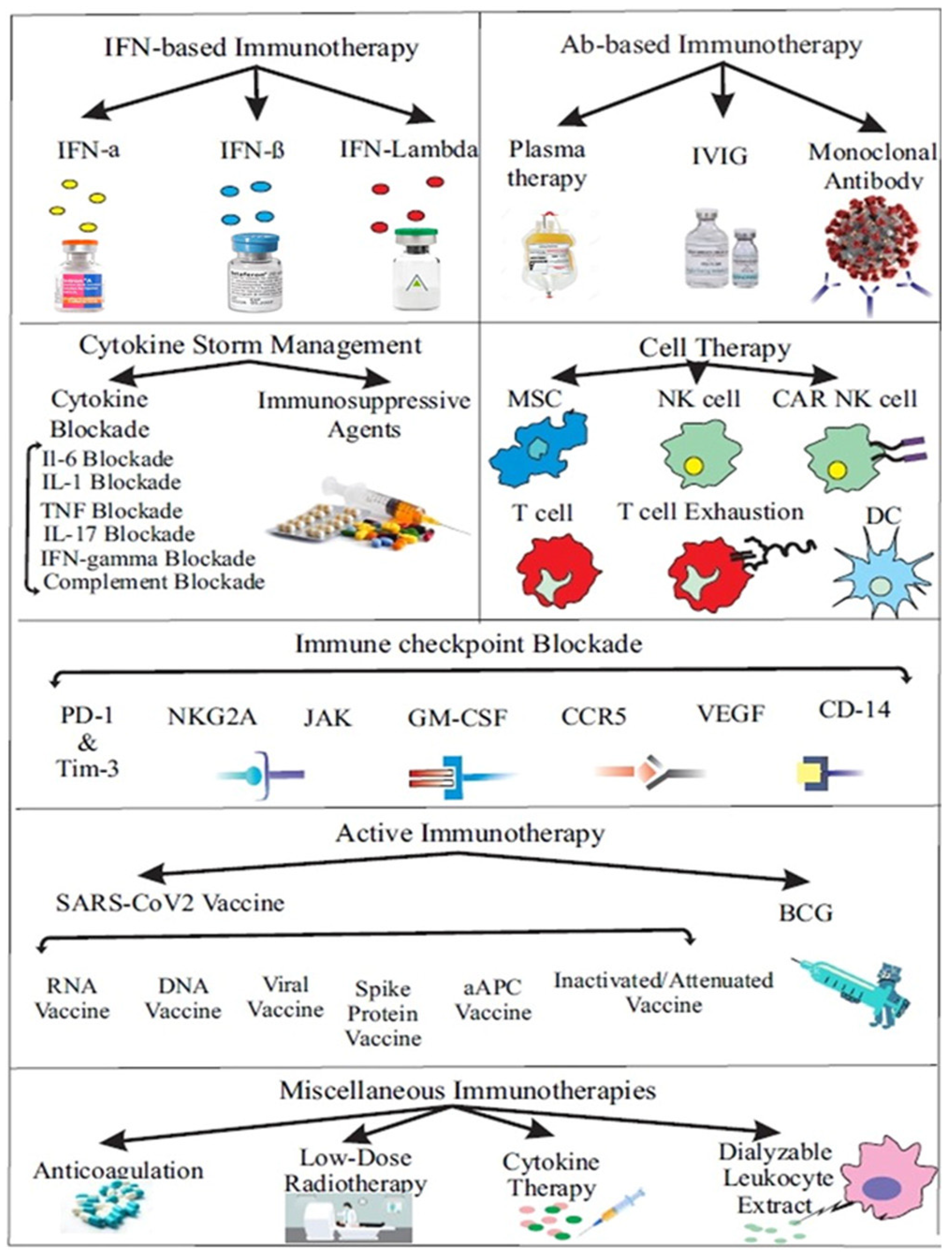
3. Immunotherapy and Vaccine
4. Computer Aided Drug Design
5. Therapeutics for Covid-19 Treatment
5.1. Antivirals
5.2. Anti-Inflammatory Compounds
5.3. Antibacterial Compounds
5.4. Corticosteroid Compounds
5.5. Antiparasitic Compounds
5.6. Anticancer Compounds
5.7. Antipsychotic and Antihistamine Compounds
5.8. Antihypertensive Compounds
5.9. Antidiabetic Compounds
5.10. Immunosuppressive Compounds
5.11. Immunomodulators
6. Synthesis of New Molecular Structures against COVID-19
- (i)
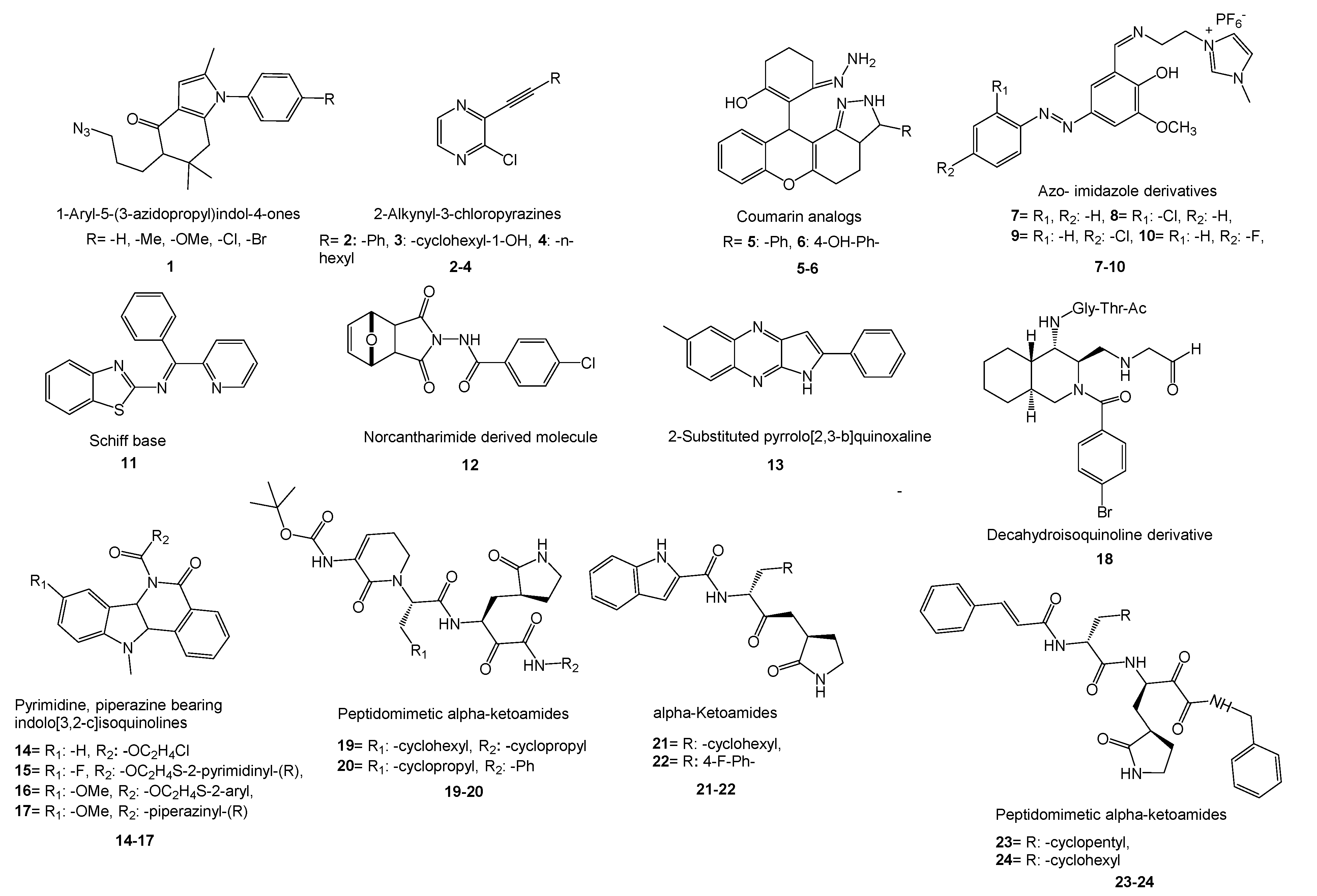
- Domínguez-Villa and co-workers reported the synthesis of five new azidopropylindol-4-ones (1) as a potential inhibitor of SARS-CoV-2 3CLpro [276]. Molecular docking studies, ADME/Tox profile, and drug-likeness works showed favorable properties of the compounds with low toxicity. Therefore, fine bioavailability levels were foreseen. Research on copper-catalyzed sonochemical synthesis of 2-alkynyl-3-chloropyrazines showed binding affinity of the compounds onto the NTD of N-protein of SARS-CoV-2 [277]. The researchers followed SAR, molecular docking, and ADME studies to show the compounds are prospective ligands for SARS-CoV-2. Three new compounds (2–4) were found as potential agents for further studies. In a promising work, novel coumarin analogs and some natural coumarin analogs were investigated to inhibit SARS-CoV-2 Mpro via molecular docking and PK studies of ADME and drug-likeness [278]. Among the synthetic coumarin analogs, two compounds (5,6) revealed good binding energy inhibition potential. A recent work on the inhibitory action of azo- imidazole derivatives against SARS-CoV-2 Mpro presented four new synthesized compounds (7–10) as promising agents by comparing the efficacy of the molecules with FDA-approved and some repurposed antiviral drugs using molecular docking and ADME research [279]. Ahmed et al. synthesized three new Schiff bases as potential SARS-CoV-2 3CLpro inhibitors [280].
- (ii)
- Novel pyrimidine, piperazine-bearing indolo[3,2-c]isoquinolines were synthesized as potent COVID-19 Mpro inhibitors by Verma’s group [283]. Molecular docking studies exhibited good interactions of 4 compounds (14–17) with 6LZE (COVID-19) and 6XFN (SARS-CoV-2) at active sites. In another work on the synthesis of isoquinoline derivatives, molecular docking and in vitro studies revealed that the decahydroisoquinoline scaffold (18) is a good hydrophobic moiety to interact with S2 site of SARS 3CLpro [284].
- (iii)
- Research covering the synthesis of a series of lipopeptides demonstrated that these peptides are potent coronavirus fusion inhibitors [285]. Cytotoxicity studies, in vitro cell–cell fusion assays, and in vivo mouse infection studies demonstrated that one peptide drug is the most potent fusion inhibitor against SARS-CoV-2 and can be used in an inhalation formulation to treat patients.
- (iv)
- Zhang and co-workers reported the synthesis of peptidomimetic α-ketoamides and PK properties of optimized SARS-CoV-2 Mpro inhibitors (19,20). They revealed a pronounced lung tropism showing that it is suitable for use by the inhalative route [286]. In another work, new α-ketoamides were synthesized, and two of them (21,22) showed satisfactory SARS-CoV-2 3CLpro inhibitory activity [287]. The aldehyde groups were covalently linked to cysteine145 of 3CLpro and showed in vivo PK properties. Additionally, in vivo toxicity studies with (21) in SD rats and hunting dogs revealed no significant toxicity in either group. In another study, designed peptidomimetic α-ketoamides were synthesized against Mpro of coronaviruses and 3CLpro of enteroviruses [288]. The researchers found two near-equipotent inhibitors (23,24) by testing the compounds against recombinant proteases, in viral replicons and virus-infected cell cultures. They showed once again that structure-based approaches in the development of broad-spectrum antivirals are a powerful tool.
7. COVID-19 Pandemic and Food: Safety and Functional Food Components
8. Potential Natural Products against COVID-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical | Plant Molecule/ (Natural Source) | Mechanism of Action/(Experimental Results) | Refs |
|---|---|---|---|
| Flavonoids | Baicalin (Citrus) | ACE2 inhibitor of SARS in FRhK-4 cell line (IC50: 2.24 μM; EC50: 12.5 μg/mL) | [322] |
| Binding with TMPRSS2 of SARS-CoV-2 (ΔG: −8.46 kcal/mol) | [324] | ||
| 3CLpro inhibitor of SARS-CoV-2 on Vero E6 cells (IC50: 6.41 µM, EC50: 10.27 µM, SI: 19 µM) | |||
| Quercetin (Vegetables) | Binding with ACE2 of SARS-CoV-2 (ΔG: −8.66 kcal/mol) | ||
| 3CLpro inhibitor of SARS-CoV-2 (ΔG: −6.6 kcal/mol EC50 = 83.4 μM; CC50 = 3.32 μM) | [323] | ||
| Sotetsuflavonen (Dacrydium araucarioides) | RdRp inhibitor of SARS-CoV-2 (IC50: 0.16 μM) | [324] | |
| Hesperetin (Citrus) | 3CLpro inhibitor of SARS-CoV-2 in a cell-based cleavage assay (IC50: 8.3 μM) | [330] | |
| Kaempferol (Sambucus formosana Nakai) | Binding with ACE2 and 3CLpro of SARS-CoV-2 (ΔG: −7.20 kcal/mol) TMPRSS2 inhibitor of SARS-CoV-2 (ΔG: −7.80 kcal/mol) | [326] | |
| Rhoifolin (Hypericum perforatum) | Binding with 3CLpro of SARS-CoV-2 (ΔG: −8.37 kcal/mol) | [323] | |
| Scutellarin (Hypericum perforatum) | Binding with of 3CLpro of SARS-CoV-2 (ΔG: −8.32 kcal/mol) | ||
| Naringenin (Citrus) | Binding with RdRp of SARS-CoV-2 (ΔG: −5.69 kcal/mol) | [331] | |
| Glycosylated flavonoids | Baicalein (Curcuma longa L.) | 3CLpro inhibitor of SARS-CoV-2 on Vero E6 cells (IC50: 0.94 µM, EC50: 1.69 µM, SI: 118 µM) | [324] |
| Hesperidin (Citrus) | Binding with ACE2 protein of SARS-CoV-2 (ΔG: −8.3 kcal/mol) | [325] | |
| Polyphenols | (-)-Epigallocatechin gallate (Hypericum perforatum) | Binding with 3CLpro of SARS-CoV-2 (ΔG: −7.96 kcal/mol) | [323] |
| Caffeic acid (Sambucus formosana Nakai) | ACE2 inhibitor of HCoV-NL63 (IC50: 8.1 μM) | ||
| Ellagic acid (Berry) | Binding with RdRp of SARS-CoV-2 (ΔG: −4.59 kcal/mol) | [331] | |
| Psoralidin (Cullen corylifolium (L.)) | PLpro inhibitor of SARS-CoV (IC50: 4.2 μM) | [327] | |
| Polyphenols (tannins) | Tannic acid (Camellia sinensis) | 3CLpro inhibitor of SARS-CoV-2 (IC50: 3 μM) | [17] |
| 3-Isotheaflavin-3 gallate (Camellia sinensis) | 3CLpro inhibitor of SARS-CoV-2 (IC50: 7 μM) | ||
| Diarylheptanoids | Hirsutenone (Alnus japonica) | PLpro inhibitor of SARS-CoV (IC50: 4.1 µM) | [322] |
| Curcumin (Curcuma longa) | PLpro inhibitor of SARS-CoV (IC50: 5.7 μM) | ||
| 3CLpro inhibitor of SARS-CoV-2 (ΔG: −8.15 kcal/mol) | [323] | ||
| Prenylated phloroglucinol | Hyperforin (Hypericum perforatum) | 3CLpro inhibitor of SARS-CoV-2 (Inhibition of various cytokines at the 1.0 μM level in isolated rats and human pancreatic islets) | [329] |
| Glycosylated seco-iridoid | Oleuropein (Olea europaea) | RdRp inhibitor of SARS-CoV-2 (ΔG: −4.94 kcal/mol) | [331] |
| Alkylated chalcones | Isobavachalcone (Cullen corylifolium (L.)) | PLpro inhibitor of SARS-CoV (IC50: 7.3 μM) | [327] |
| Prenylated chalcones | Xanthoangelol E (Angelica keiskei) | PLpro inhibitor of SARS-CoV (IC50: 1.2 μM) | |
| Xanthoangelol F (Angelica keiskei) | PLpro inhibitor of SARS-CoV (IC50: 5.6 μM) | ||
| Saponins | Glycyrrhizin (Glycyrrhiza glabra L.) | Binding with ACE2 of SARS-CoV (ΔG: −9 kcal/mol) | [323] |
| Binding with 3CLpro of SARS-CoV (ΔG: −8.9 kcal/mol) | [17] | ||
| Terpenoids | Iguesterin (Tripterygium regelii) | 3CLpro inhibitor of SARS-CoV (IC50: 2.6 μM) | [323] |
| Pristimerin (Tripterygium regelii) | 3CLpro inhibitor of SARS-CoV (IC50: 5.5 μM) | ||
| Saikosaponin B2 (Heteromorpha spp.) | ACE2 inhibitor of HCoV-229E (EC50 = 1.7 μM) | ||
| Abietane diterpene | Cryptotanshinone (Salvia miltiorrhiza) | TMPRSS2 inhibitor in the LNCaP cells (IC50: 2.42 μM) | [332] |
| PLpro inhibitor of SARS-CoV (IC50: 0.8 μM) | [323] | ||
| Tanshinone IIA (Salvia miltiorrhiza) | PLpro inhibitor of SARS-CoV (IC50: 1.6 μM) | ||
| Dihydrotanshinone I (Salvia miltiorrhiza) | PLpro inhibitor of SARS-CoV (IC50: 4.9 μM) | ||
| Alkaloids | Emetine (Carapichea ipecacuanha) | 3CLpro inhibitor of HCoV-OC43 (EC50: 0.30 μM) | |
| 3CLpro inhibitor of HCoV-NL63 (EC50: 1.43 μM) | |||
| Caffeine (Cocoa beans) | Binding with RdRp of SARS-CoV-2 (ΔG: −6.10 kcal/mol) | [331] | |
| Tryptanthrin (Strobilanthes cusia) | 3CLpro inhibitor of HCoV-NL63 (IC50: 1.52 μM; ΔG: −8.2 kcal/mol) | [330] | |
| Berberine (Aloe barbadensis) | Binding with 3CLpro of SARS-CoV-2 (ΔG: −8.1 kcal/mol) | [333] | |
| Anthraquinone | Rhein (Aloe barbadensis) | Binding with 3CLpro of SARS-CoV-2 (ΔG: −8.9 kcal/mol) | |
| Quinone | Thymoquinone (Nigella sativa) | Binding with ACE2 of SARS-CoV-2 (ΔG: −5.5 kcal/mol) | [334] |
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez-Morales, A.J.; Bonilla-Aldana, D.K.; Balbin-Ramon, G.J.; Rabaan, A.A.; Sah, R.; Paniz-Mondolfi, A.; Pagliano, P.; Esposito, S. History is repeating itself: Probable zoonotic spillover as the cause of the 2019 novel Coronavirus Epidemic. Infez. Med. 2020, 28, 3–5. [Google Scholar]
- Gralinski, L.E.; Menachery, V.D. Return of the coronavirus: 2019-nCoV. Viruses 2020, 12, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China. N. Engl. J. Med. 2019, 382, 727–733. [Google Scholar] [CrossRef]
- Wei, X.; Li, X.; Cui, J. Evolutionary perspectives on novel coronaviruses identified in pneumonia cases in China. Nat. Sci. Rev. 2020, 7, 239–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munster, V.J.; Koopmans, M.; Van Doremalen, N.; Van Riel, D.; De Wit, E. A novel coronavirus emerging in China—key questions for impact assessment. N. Engl. J. Med. 2020, 382, 692–694. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, K.; Shi, Z.L.; Zhou, P. Bat Coronaviruses in China. Viruses 2019, 11, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Ng, O.W.; Tan, Y.J. Understanding bat SARS-like coronaviruses for the preparation of future coronavirus outbreaks—implications for coronavirus vaccine development. Hum. Vaccines Immunother. 2017, 13, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Aldana, D.K.; Holguin-Rivera, Y.; Cortes-Bonilla, I.; Cardona-Trujillo, M.C.; García-Barco, A.; Bedoya-Arias, H.A.; Rabaan, A.A.; Sah, R.; Rodriguez-Morales, A.J. Coronavirus infections reported by ProMED, february 2000–january 2020. Travel. Med. Infect. Dis. 2020, 35, 101575. [Google Scholar] [CrossRef]
- Zhang, L.; Shen, F.M.; Chen, F.; Lin, Z. Origin and evolution of the 2019 novel coronavirus. Clin. Infect. Dis. 2020, 71, 882–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Pradoet, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar]
- WHO. Vaccines. Guidance Document. Status of COVID-19 Vaccines within WHO EUL/PQ Evaluation Process. 23 March 2021. Available online: https://extranet.who.int/pqweb/sites/default/files/documents/Status_COVID_VAX_23March2021_0.pdf (accessed on 30 March 2021).
- WHO. Draft Landscape and Tracker of COVID-19 Candidate Vaccines. 9 April 2021. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 21 April 2021).
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.; Wu, G.; Xu, M.; Lou, Z.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: A randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect. Dis. 2021, 21, 39–51. [Google Scholar] [CrossRef]
- Smith, T.R.; Patel, A.; Ramos, S.; Elwood, D.; Zhu, X.; Yan, J.; Broderick, K.E. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, Y.; Tariq, A.; Jiang, X.; Ahmed, Z.; Zhihao, Z.; Idrees, M.; Azizullah, A.; Adnan, M.; Bussmann, R.W. Food as medicine: A possible preventive measure against coronavirus disease (COVID-19). Phytother. Res. 2020, 34, 3124–3136. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirüs: Implications for virüs origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar]
- Brian, D.A.; Baric, R.S. Coronavirus genome structure and replication. Curr. Topics. Microbiol. Immunology 2005, 287, 1–30. [Google Scholar]
- Chang, C.K.; Sue, S.C.; Yu, T.H.; Hsieh, C.-M.; Tsai, C.-K.; Chiang, Y.-C.; Lee, S.-J.; Hsiao, H.-H.; Wu, W.-J.; Chang, W.-L.; et al. Modular organization of SARS coronavirus nucleocapsid protein. J. Biomed. Sci. 2006, 13, 59–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef]
- Kuldeep, D.; Khan, S.; Tiwari, R.; Sircar, S.; Bhat, S.; Malik, Y.S.; Singh, K.P.; Chaicumpa, W. Coronavirus disease 2019–COVID-19. Clinic. Microbiol. Rev. 2020, 33, 20–28. [Google Scholar]
- Schoeman, D.; Fielding, B.C. Coronavirus envelope protein: Current knowledge. Virol. J. 2019, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Nicholls, J.M.; Chen, Y.G. Severe acute respiratory syndrome-associated coronavirus nucleocapsid protein interacts with Smad3 and modulates transforming growth factor-β signaling. J. Biol. Chem. 2008, 283, 3272–3280. [Google Scholar] [CrossRef] [PubMed]
- Beniac, D.R.; Andonov, A.; Grudeski, E.; Booth, T.F. Architecture of the SARS coronavirus prefusion spike. Nat. Struct. Mol. Biol. 2006, 13, 751–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benvenuto, D.; Giovanetti, M.; Ciccozzi, A.; Spoto, S.; Angeletti, S.; Ciccozzi, M. The 2019-new coronavirus epidemic: Evidence for virus evolution. J. Med. Virol. 2020, 92, 455–459. [Google Scholar] [CrossRef] [Green Version]
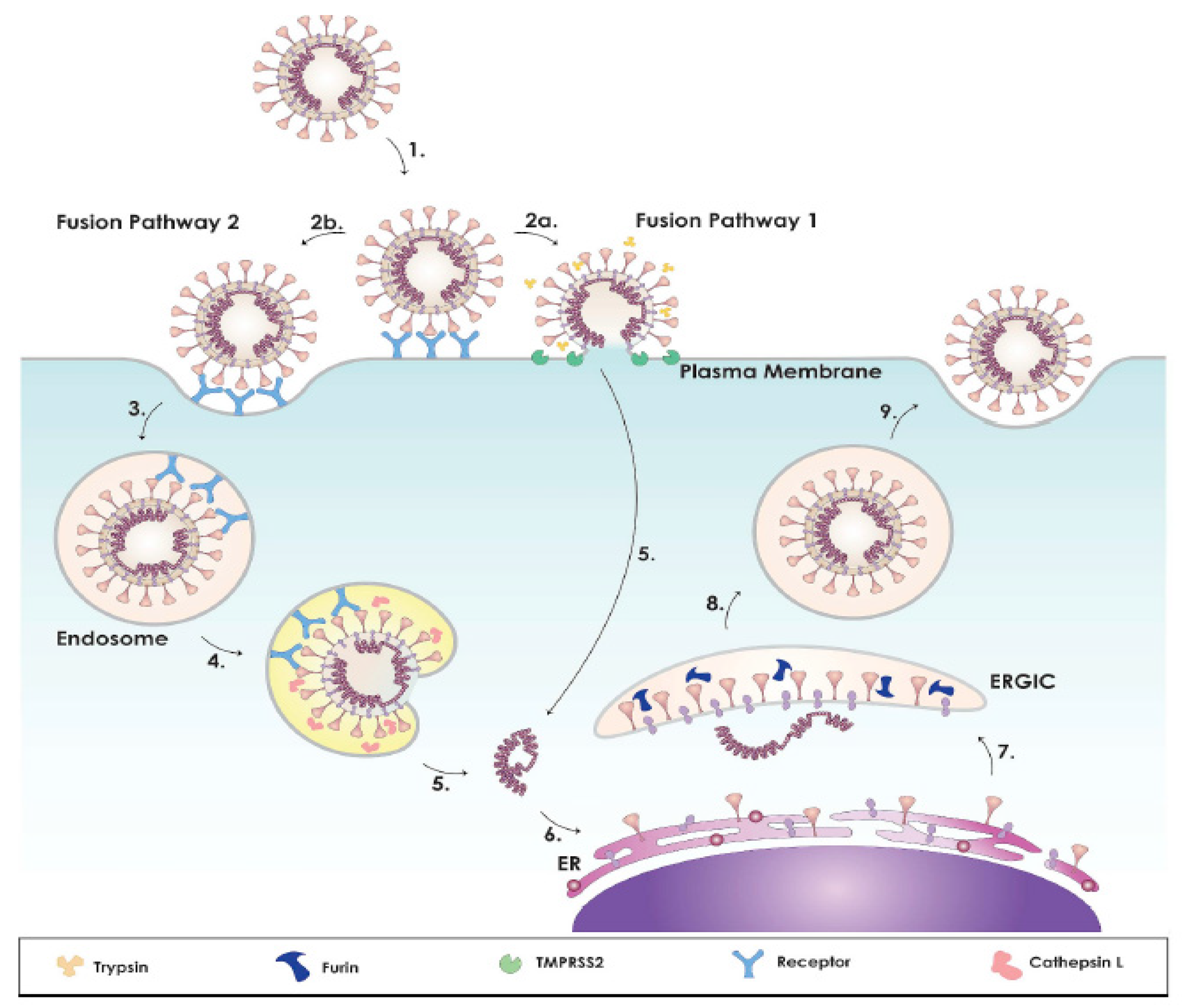
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittakerb, G.R.; Daniel, S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antivir. Res. 2020, 178, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nal, B.; Chan, C.; Kien, F.; Siu, L.; Tse, J.; Chu, K.; Kam, J.; Staropoli, I.; Crescenzo-Chaigne, B.; Escriou, N.; et al. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S., M and E. J. Gen. Virol. 2005, 86, 1423–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.Y.; Li, J.L.; Yang, X.L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Receptor recognition mechanisms of coronaviruses: A decade of structural studies. J. Virol. 2015, 89, 1954–1964. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Heurich, A.; Hofmann-Winkler, H.; Gierer, S.; Liepold, T.; Jahn, O.; Pöhlmann, S. TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein. J. Virol. 2014, 88, 1293–1307. [Google Scholar] [CrossRef] [Green Version]
- Yost, S.A.; Marcotrigiano, J. Viral precursor polyproteins: Keys of regulation from replication to maturation. Curr. Opin. Virol. 2013, 3, 137–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.; Mukherjee, R.; Grewe, D.; Bojkova, D.; Baek, K.; Bhattacharya, A.; Schulz, L.; Widera, M.; Mehdipour, A.R.; Tascher, G.; et al. Papain-like protease regulates SARS-CoV-2 viral spread and innate immunity. Nature 2020, 587, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Sisay, M. 3CLpro inhibitors as a potential therapeutic option for COVID-19: Available evidence and ongoing clinical trials. Pharmacol. Research 2020, 156, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Qiang, T.; Liang, C.; Ren, X.; Jia, M.; Zhang, J.; Li, J.; Wan, M.; Wen, X.Y.; Li, H.; et al. RNA-dependent RNA polymerase (RdRp) inhibitors: The current landscape and repurposing for the COVID-19 pandemic. Eur. J. Med. Chem. 2021, 213, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Kok, K.H.; Zhu, Z.; Chu, H.; Kai-Wang, T.K.; Yuan, S.; Yuen, K.Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes. Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, N.C.; Smith, M.; Vreede, F.T.; Fodor, E. NS2/NEP protein regulates transcription and replication of the influenza virus RNA genome. J. Gen. Virol. 2009, 90, 1398–1407. [Google Scholar] [CrossRef]
- Huang, J.; Song, W.; Huang, H.; Sun, Q. Pharmacological therapeutics targeting RNA-dependent RNA polymerase, proteinase and spike protein: From mechanistic studies to clinical trials for COVID-19. J. Clin. Med. 2020, 9, 1131. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sui, J.; Huang, I.-C.; Kuhn, J.H.; Radoshitzky, S.R.; Marasco, W.A.; Choe, H.; Farzan, M. The S proteins of human coronavirus NL63 and severe acute respiratory syndrome coronavirus bind overlapping regions of ACE2. Virology 2007, 367, 367–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVID-19 or SARSCoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agent. 2020, 34, 327–331. [Google Scholar]
- Channappanavar, R.; Fehr, A.R.; Vijay, R.; Mack, M.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause Lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe. 2016, 19, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Behrens, E.M.; Koretzky, G.A. Review: Cytokine storm syndrome: Looking toward the precision medicine Era. Arthritis Rheumatol. 2017, 69, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, C.W.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 2020, 181, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.J.; Su, I.J.; Theron, M.; Wu, Y.C.; Lai, S.K.; Liu, C.C.; Le, H.-Y. An interferon-gamma-related cytokine storm in SARS patients. J. Med. Virol. 2005, 75, 185–194. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z.; et al. Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononuclear cells in COVID-19 patients. Emerg. Microbes. Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; et al. Risk factors associated with acute respiratory distress syndrome and death in patients with coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Henry, B.M.; Vikse, J.; Benoit, S.; Favaloro, E.J.; Lippi, G. Hyperinflammation and derangement of renin- angiotensin-aldosterone system in COVID-19: A novel hypothesis for clinically suspected hypercoagulopathy and microvascular immunothrombosis. Clin. Chim. Acta 2020, 507, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.A.; Hossain, N.; Kashem, M.A.; Shahid, M.A.; Alam, A. Immune response in COVID-19: A review. J. Infect. Public Health 2020, 13, 1619–1629. [Google Scholar] [CrossRef]
- Understanding Immunotherapy, Cancer Council. Available online: https://www.cancer.org.au/assets/pdf/understanding-immunotherapy-fact-sheet (accessed on 21 March 2021).
- Puronen, C.E.; Ford, E.S.; Uldrick, T.S. Immunotherapy in people with HIV and cancer. Front. Immunol. 2019, 10, 2060. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.; Liu, J.; Zhang, Z.; Wan, X.; Huang, B.; Chen, Y.; Zhang, Y. COVID-19: Immunopathogenesis and Immunotherapeutics. Sig. Transduct. Target Ther. 2020, 5, 128. [Google Scholar]
- Khosroshahi, L.M.; Rokni, M.; Mokhtari, T.; Noorbakhsh, F. Immunology, immunopathogenesis and immunotherapeutics of COVID-19; an overview. Int. Immunopharmacol. 2021, 93, 107364. [Google Scholar] [CrossRef]
- Esmaeilzadeh, A.; Elahi, R. Immunobiology and immunotherapy of COVID-19: A clinically updated overview. J. Cell. Physiol. 2021, 236, 2519–2543. [Google Scholar] [CrossRef] [PubMed]
- Berraondo, P.; Sanmamed, M.F.; Ochoa, M.C.; Etxeberria, I.; Aznar, M.A.; Pérez-Gracia, J.L.; Rodríguez-Ruiz, M.E.; Ponz-Sarvise, M.; Castañón, E.; Melero, I. Cytokines in clinical cancer immunotherapy. Br. J. Cancer 2019, 120, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, K.C.; Miljkovic, M.D.; Waldmann, T.A. Cytokines in the treatment of cancer. J. Interferon Cytokine Res. 2019, 39, 6–21. [Google Scholar] [CrossRef] [PubMed]
- AminJafari, A.; Ghasemi, S. The possible of immunotherapy for COVID-19: A systematic review. Int. Immunopharmacol. 2020, 83, 106455. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J. Passive antibody therapy in COVID-19. Nat. Rev. Immunol. 2020, 20, 401–403. [Google Scholar] [CrossRef]
- Prévost, Jérémie; Finzi, Andrés. The great escape? SARS-CoV-2 variants evading neutralizing responses. Cell Host Microbe 2021, 29, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Munnink, B.B.O.; Haagmans, B.L. Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef] [Green Version]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 infection: The role of cytokines in COVID-19 disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef] [PubMed]
- García, L.F. Immune response, inflammation, and the clinical spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef]
- Perez, C.R.; De Palma, M. Engineering dendritic cell vaccines to improve cancer immunotherapy. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brewitz, A.; Eickhoff, S.; Dähling, S.; Quast, T.; Bedoui, S.; Kroczek, R.A.; Kastenmüller, W. CD8+ T cells orchestrate pDC-XCR1+ dendritic cell spatial and functional cooperativity to optimize priming. Immunity 2017, 46, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Saadeldin, M.K.; Abdel-Aziz, A.K.; Abdellatif, A. Dendritic Cell Vaccine Immunotherapy; the beginning of the end of Cancer and COVID-19. A Hypothesis. Med. Hypotheses 2021, 146, 110365. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tenchov, R.; Smoot, J.; Liu, C.; Watkins, S.; Zhou, Q. A comprehensive review of the global efforts on COVID-19 vaccine development. ACS Cent. Sci. 2021, 7, 512–533. [Google Scholar] [CrossRef] [PubMed]
- Valérie, N.; Damien, M. COVID-19 Tracker Status of the Top 5 Vaccines Candidates. Available online: https://www.avertim.com/en/news/covid-19-tracker-status-top-5-vaccines-candidates (accessed on 21 April 2021).
- Batty, C.J.; Heise, M.T.; Bachelder, E.M.; Ainslie, K.M. Vaccine formulations in clinical development for the prevention of severe acute respiratory syndrome coronavirus 2 infection. Adv. Drug Deliv. Rev. 2020, 169, 168–189. [Google Scholar] [CrossRef] [PubMed]
- Yap, T.A.; Siu, L.L.; Calvo, E.; Lolkema, M.P.; LoRusso, P.M.; Soria, J.C.; Banerji, U. SARS-CoV-2 vaccination and phase 1 cancer clinical trials. Lancet Oncol. 2021, 22, 298–301. [Google Scholar] [CrossRef]
- Bajaj, V.; Gadi, N.; Spihlman, A.P.; Wu, S.C.; Choi, C.H.; Moulton, V.R. Aging, immunity, and COVID-19: How age influences the host immune response to coronavirus infections? Front. Physiol. 2021, 11, 1793. [Google Scholar] [CrossRef]
- Kaur, S.P.; Gupta, V. COVID-19 Vaccine: A comprehensive status report. Virus Res. 2020, 288, 198114. [Google Scholar] [CrossRef]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in mRNA vaccines for infectious diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef] [Green Version]
- WHO. Update on COVID-19 Vaccine Development, The Latest ön The COVID-19 Global Situation & Vaccine Development. Available online: https://www.who.int/docs/default-source/coronaviruse/risk-comms-updates/update45-vaccinesdevelopement.pdf?sfvrsn=13098bfc_5 (accessed on 30 March 2021).
- WHO. WEvidence Assessment: Sinovac/CoronaVac COVID-19 Vaccine. Available online: https://cdn.who.int/media/docs/default-source/immunization/sage/2021/april/5_sage29apr2021_critical-evidence_sinovac.pdf (accessed on 21 May 2021).
- Felsenstein, S.; Herbert, J.A.; McNamara, P.S.; Hedrich, C.M. COVID-19: Immunology and treatment options. Clin. Immunol. 2020, 215, 108448. [Google Scholar] [CrossRef]
- Florindo, H.F.; Kleiner, R.; Vaskovich-Koubi, D.; Acúrcio, R.C.; Carreira, B.; Yeini, E.; Satchi-Fainaro, R. Immune-mediated approaches against COVID-19. Nat. Nanotech. 2020, 15, 630–645. [Google Scholar] [CrossRef]
- Singh, R.; Kang, A.; Luo, X.; Jeyanathan, M.; Gillgrass, A.; Afkhami, S.; Xing, Z. COVID-19: Current knowledge in clinical features, immunological responses, and vaccine development. FASEB J. 2021, 35, e21409. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Cao, B. 6-Month consequences of COVID-19 in patients discharged from hospital: A cohort study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Astell, C.R.; Holt, R.A.; Jones, S.J.; Marra, M.A. Genome Organization and Structural Aspects of the SARS-Related Virus. In Coronaviruses with Special Emphasis on First Insights Concerning SARS, 1st ed.; Schmidt, A., Weber, O., Wolff, M.H., Eds.; Birkhäuser Advances in Infectious Diseases BAID, Birkhäuser: Basel, Switzerland, 2005; pp. 101–128. [Google Scholar]
- Noureddine, O.; Issaoui, N.; Medimagh, M.; Al-Dossary, O.; Marouani, H. Quantum chemical studies on molecular structure, AIM, ELF, RDG and antiviral activities of hybrid hydroxychloroquine in the treatment of COVID-19: Molecular docking and DFT calculations. J. King Saud. Univ. Sci. 2021, 33, 101334. [Google Scholar] [CrossRef]
- Mizukami, Y. Character of Frontier Orbitals of Antiviral Drugs: Candidate Drugs against COVID-19. Open J. Phys. Chem. 2020, 10, 158–165. [Google Scholar] [CrossRef]
- Chatterjee, S.; Kumar, N.; Sehrawat, H.; Yadav, N.; Mishra, V. Click triazole as a linker for drug repurposing against SARs-CoV-2: A greener approach in race to find COVID-19 therapeutic. Curr. Res. Green Sustain. Chem. 2021, 4, 100064. [Google Scholar] [CrossRef]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Fatimawali, F.; Kepel, B.J.; Idroes, R.; Effendi, Y.; Sakib, S.A.; Emran, T.B. Potential of plant bioactive compounds as SARS-CoV-2 main protease (Mpro) and spike (S) glycoprotein inhibitors: A molecular docking study. Scientifica 2020, 2020, 1–18. [Google Scholar] [CrossRef]
- Sagaama, A.; Brandan, S.A.; Issa, T.B.; Issaoui, N. Searching potential antiviral candidates for the treatment of the 2019 novel coronavirus based on DFT calculations and molecular docking. Heliyon 2020, 6, e04640. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, S.; Dubey, A.; Yadava, U.; Mishra, S.K.; Kang, S.G.; Dwivedi, V.D. Exploration of natural compounds with anti-SARS-CoV-2 activity via inhibition of SARS-CoV-2 Mpro. Brief Bioinform. 2021, 22, 1361–1377. [Google Scholar] [CrossRef]
- Rasool, N.; Yasmin, F.; Sahai, S.; Hussain, W.; Inam, H.; Arshad, A. Biological perspective of thiazolide derivatives against Mpro and MTase of SARS-CoV-2: Molecular docking, DFT and MD simulation investigations. Chem. Phys. Lett. 2021, 771, 138463. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, D.; Kumar, R.; Singh, P.; Chandra, R.; Kumari, K. DFT and docking studies of designed conjugates of noscapines & repurposing drugs: Promising inhibitors of main protease of SARS-CoV-2 and falcipan-2. J. Biomol. Struct. Dyn. 2020, 1–21. [Google Scholar] [CrossRef]
- Jordaan, M.A.; Ebenezer, O.; Damoyi, N.; Shapi, M. Virtual screening, molecular docking studies and DFT calculations of FDA approved compounds similar to the non-nucleoside reverse transcriptase inhibitor (NNRTI) efavirenz. Heliyon 2020, 6, e04642. [Google Scholar] [CrossRef] [PubMed]
- Hagar, M.; Ahmed, H.A.; Aljohani, G.; Alhaddad, O.A. Investigation of some antiviral N-heterocycles as COVID-19 drug: Molecular docking and DFT calculations. Int. J. Mol. Sci. 2020, 21, 3922. [Google Scholar] [CrossRef] [PubMed]
- Gad, E.A.M.; Ahmed, M.E.; Al-Fahemi, J.H. Comparative study on quantum descriptors, molecular docking and dynamic simulation of antiviral drugs with COVID-19. Egypt. J. Pet. 2021, 30, 45–51. [Google Scholar]
- Khelfaoui, H.; Harkati, D.; Saleh, B.A. Molecular docking, molecular dynamics simulations and reactivity, studies on approved drugs library targeting ACE2 and SARS-CoV-2 binding with ACE2. J. Biomol. Struct. Dyn. 2020, 1–17. [Google Scholar] [CrossRef]
- Lokhande, K.B.; Doiphode, S.; Vyas, R.; Swamy, K.V. Molecular docking and simulation studies on SARS-CoV-2 Mpro reveals Mitoxantrone, Leucovorin, Birinapant, and Dynasore as potent drugs against COVID-19. J. Biomol. Struct. Dyn. 2020, 1–12. [Google Scholar] [CrossRef]
- Altalhi, T.A.; Alswat, K.; Alsanie, W.F.; Ibrahim, M.M.; Aldalbahi, A.; El-Sheshtawy, H.S. Chloroquine and hydroxychloroquine inhibitors for COVID-19 sialic acid cellular receptor: Structure, hirshfeld atomic charge analysis and solvent effect. J. Mol. Struct. 2021, 1228, 129459. [Google Scholar] [CrossRef]
- Noor, H.; Ikram, A.; Rathinavel, T.; Kumarasamy, S.; Iqbal, M.N.; Bashir, Z. Immunomodulatory and anti-cytokine therapeutic potential of curcumin and its derivatives for treating COVID-19—A computational modeling. J. Biomol. Struct. Dyn. 2021, 1–16. [Google Scholar] [CrossRef]
- El Kalai, F.; Çınar, E.B.; Lai, C.H.; Daoui, S.; Chelfi, T.; Allali, M.; Dege, N.; Karrouchi, K.; Benchat, N. Synthesis, spectroscopy, crystal structure, TGA/DTA study, DFT and molecular docking investigations of (E)-4-(4-methylbenzyl)-6-styrylpyridazin-3 (2H)-one. J. Mol. Struct. 2021, 1228, 129435. [Google Scholar] [CrossRef] [PubMed]
- Alagumuthu, M.; Rajpoot, S.; Baig, M.S. Structure-based design of novel peptidomimetics targeting the SARS-CoV-2 spike protein. Cell. Mol. Bioeng. 2021, 14, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Fast identification of possible drug treatment of coronavirus disease-19 (COVID-19) through computational drug repurposing study. J. Chem. Inf. Model. 2020, 60, 3277–3286. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Gao, C.; Zhou, Q.; Driel, W.V.; Ye, H.; Zhang, G. The inactivation mechanism of chemical disinfection against SARS-CoV-2: From MD and DFT perspectives. RSC Adv. 2020, 10, 40480–40488. [Google Scholar] [CrossRef]
- Geoffrey, A.S.B.; Madaj, R.; Sanker, A.; Tresanco, M.S.V.; Davidd, H.A.; Roy, G.; Saji, R.S.; Yakubu, A.H.; Malgija, B. Automated in Silico Identification of Drug Candidates for Coronavirus Through a Novel Programmatic Tool and Extensive Computational (MD, DFT) Studies of Select Drug Candidates. Preprint 2020. [Google Scholar] [CrossRef]
- Abdelli, I.; Hassani, F.; Brikci, S.B.; Ghalem, S. In silico study the inhibition of angiotensin converting enzyme 2 receptor of COVID-19 by Ammoides verticillata components harvested from Western Algeria. J. Biomol. Struct. Dyn. 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Adebambo, K.F. Computational investigation of the interaction of anti-influenza drugs with CoVID-19 protein. Comput. Mol. Biosci. 2020, 10, 45–60. [Google Scholar] [CrossRef]
- Mishra, S.S.; Ranjan, S.; Sharma, C.S.; Singh, H.P.; Kalra, S.; Kumar, N. Computational investigation of potential inhibitors of novel coronavirus 2019 through structure-based virtual screening, molecular dynamics and density functional theory studies. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef]
- Gupta, P.S.S.; Biswal, S.; Panda, S.K.; Ray, A.K.; Rana, M.K. Binding mechanism and structural insights into the identified protein target of COVID-19 and importin-α with in-vitro effective drug ivermectin. J. Biomol. Struct. Dyn. 2020, 1–10. [Google Scholar] [CrossRef]
- Secretan, P.H.; Yayé, H.S.; Sogaldi, A.; Antignac, M.; Tortolano, L.; Thirion, O.; Vieillard, V.; Yagoubi, N.; Do, B. Intrinsic stability of the antiviral drug umifenovir by stress testing and DFT studies. J. Pharm. Biomed. Anal. 2021, 196, 113934. [Google Scholar] [CrossRef] [PubMed]
- Vlasiou, M.C.; Pafti, K.S. Screening possible drug molecules for COVID-19. The example of vanadium (III/IV/V) complex molecules with computational chemistry and molecular docking. Comput. Toxicol. 2021, 18, 100157. [Google Scholar] [CrossRef]
- Sliwoski, G.; Kothiwale, S.; Meiler, J.; Lowe, E.W. Computational methods in drug discovery. Pharmacol. Rev. 2014, 66, 334–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, P.K.; Arul, M.N.; Patel, P.; Verma, S.K.; Luo, W.; Rubahn, H.G.; Mishra, Y.K.; Suar, M.; Ahuja, R. Structure-based drug designing and immunoinformatics approach for SARS-CoV-2. Sci. Adv. 2020, 6, eabb8097. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Krumberger, M.; Morris, M.A.; Parrocha, C.M.T.; Kreutzer, A.G.; Nowick, J.S. Structure-based drug design of an inhibitor of the SARS-CoV-2 (COVID-19) main protease using free software: A tutorial for students and scientists. Eur. J. Med. Chem. 2021, 218, 113390. [Google Scholar] [CrossRef]
- Chou, K.C.; Wei, D.Q.; Zhong, W.Z. Binding mechanism of coronavirus main proteinase with ligands and its implication to drug design against SARS. Biochem. Biophys. Res. Commun. 2003, 308, 148–151. [Google Scholar] [CrossRef]
- Joshi, R.S.; Jagdale, S.S.; Bansode, S.B.; Shankar, S.S.; Tellis, M.B.; Pandya, V.K.; Chugh, A.; Giri, A.P.; Kulkarni, M.J. Discovery of potential multi-target-directed ligands by targeting host-specific SARS-CoV-2 structurally conserved main protease. J. Biomol. Struct. Dyn. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.G. Quaternary Structure of the SARS Coronavirus Main Protease. In Molecular Biology of the SARS-Coronavirus; Lal, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 115–128. [Google Scholar]
- Kneller, D.W.; Phillips, G.; O’Neill, H.M.; Jedrzejczak, R.; Stols, L.; Langan, P.; Joachimiak, A.; Coates, L.; Kovalevsky, A. structural plasticity of SARS-CoV-2 3CL Mpro active site cavity revealed by room temperature X-ray crystallography. Nat. Commun. 2020, 11, 3202. [Google Scholar] [CrossRef]
- Jin, Z.; Zhao, Y.; Sun, Y.; Zhang, B.; Wang, H.; Wu, Y.; Zhu, Y.; Zhu, C.; Hu, T.; Du, X.; et al. Structural basis for the inhibition of SARS-CoV-2 main protease by antineoplastic drug carmofur. Nat. Struct. Mol. Biol. 2020, 27, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Sarkar, A.; Maulik, U. Molecular docking study of potential phytochemicals and their effects on the complex of SARS-CoV2 spike protein and human ACE2. Sci. Rep. 2020, 10, 1–15. [Google Scholar]
- Gupta, S.; Singh, A.K.; Kushwaha, P.P.; Prajapati, K.S.; Shuaib, M.; Senapati, S.; Kumar, S. Identification of potential natural inhibitors of SARS-CoV2 main protease by molecular docking and simulation studies. J. Biomol. Struct. Dyn. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Quimque, M.T.J.; Notarte, K.I.R.; Fernandez, R.A.T.; Mendoza, M.A.O.; Liman, R.A.D.; Lim, J.A.K.; Pilapil, L.A.E.; Ong, J.K.H.; Pastrana, A.M.; Khan, A.; et al. Virtual screening-driven drug discovery of SARS-CoV2 enzyme inhibitors targeting viral attachment, replication, post-translational modification and host immunity evasion infection mechanisms. J. Biomol. Struct. Dyn. 2020, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Maurya, S.K.; Maurya, A.K.; Mishra, N.; Siddique, H.R. Virtual screening, ADME/T, and binding free energy analysis of anti-viral, anti-protease, and anti-infectious compounds against NSP10/NSP16 methyltransferase and main protease of SARS CoV-2. J. Recept. Signal Transduct. Res. 2020, 40, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, C.; Panwar, U.; Dinesh, D.C.; Boura, E.; Singh, P.; Dubey, V.K.; Singh, S.K.; Microsecond, M.D. Simulation and multiple-confirmation virtual screening to identify potential anti-COVID-19 inhibitors against SARS-CoV-2 main protease. Front. Chem. 2020, 8, 1179. [Google Scholar]
- Huang, X.; Pearce, R.; Zhang, Y. De Novo Design of Protein Peptides to Block Association of the SARS-CoV-2 Spike Protein with Human ACE2. Aging 2020, 12, 11263. [Google Scholar] [CrossRef]
- Elmezayen, A.D.; Al-Obaidi, A.; Şahin, A.T.; Yelekçi, K. Drug repurposing for coronavirus (COVID-19): In silico screening of known drugs against coronavirus 3CL hydrolase and protease enzymes. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef]
- De Oliveira, O.V.; Rocha, G.B.; Paluch, A.S.; Costa, L.T. Repurposing approved drugs as inhibitors of SARS-CoV-2 S-protein from molecular modeling and virtual screening. J. Biomol. Struct. Dyn. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mortier, J.; Rakers, C.; Bermudez, M.; Murgueitio, M.S.; Riniker, S.; Wolber, G. The impact of molecular dynamics on drug design: Applications for the characterization of ligand–macromolecule complexes. Drug Discov. Today 2015, 20, 686–702. [Google Scholar] [CrossRef]
- Weglarz-Tomczak, E.; Tomczak, J.M.; Talma, M.; Burda-Grabowska, M.; Giurg, M.; Brul, S. Identification of ebselen and its analogues as potent covalent inhibitors of papain-like protease from SARS-CoV-2. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Scala, C.D.; Chahinian, H.; Yahi, N. Structural and molecular modelling studies reveal a new mechanism of action of chloroquine and hydroxychloroquine against SARS-CoV-2 infection. Int. J. Antimicrob. Agents 2020, 55, 105960. [Google Scholar] [CrossRef]
- Patel, D.; Athar, M.; Jha, P.C. Computational investigation of binding of chloroquinone and hydroxychloroquinone against PLPro of SARS-CoV-2. J. Biomol. Struct. Dyn. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, N.; Sakthivel, R.; Velmurugan, D.; Gromiha, M.M. Computational studies of drug repurposing and synergism of lopinavir, oseltamivir and ritonavir binding with SARS-CoV-2 protease against COVID-19. J. Biomol. Struct. Dyn. 2020, 39, 2673–2678. [Google Scholar] [CrossRef]
- Tiwari, V. De-Novo design, retrosynthetic analysis and combinatorial synthesis of a hybrid antiviral (VTAR-01) to inhibit the interaction of SARS-CoV2 spike glycoprotein with human angiotensin-converting enzyme 2. Biol. Open 2020, 9, bio054056. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Huang, H.J.; Zhuang, D.L.; Nasser, M.I.; Yang, M.H.; Zhu, P.; Zhao, M.Y. Biological, clinical and epidemiological features of COVID-19, SARS and MERS and AutoDock simulation of ACE2. Infect. Dis. Poverty 2020, 9, 1–11. [Google Scholar] [CrossRef]
- Bailly, C.; Vergoten, G. Glycyrrhizin: An alternative drug for the treatment of COVID-19 infection and the associated respiratory syndrome? Pharmacol. Ther. 2020, 107618. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Chen, W.; Xiao, D.; Wang, C. Computational molecular docking and virtual screening revealed promising SARS-CoV-2 drugs. Precis. Clin. Med. 2021, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Maffioli, P.; D’Angelo, A.; Di Pierro, F. A role for quercetin in coronavirus disease 2019 (COVID-19). Phytother. Res. 2021, 35, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.L.; Lee, N.Y.; Cia, C.T.; Ko, W.C.; Hsueh, P.R. A review of treatment of coronavirus disease 2019 (COVID-19): Therapeutic repurposing and unmet clinical needs. Front. Pharmacol 2020, 11, 1782. [Google Scholar] [CrossRef] [PubMed]
- Cusinato, J.; Cau, Y.; Calvani, A.M.; Mori, M. Repurposing drugs for the management of COVID-19. Expert Opin. Ther. Pat. 2020, 1–13. [Google Scholar] [CrossRef]
- Frediansyah, A.; Tiwari, R.; Sharun, K.; Dhama, K.; Harapan, H. Antivirals for COVID-19: A critical review. Clin. Epidemiol. Glob. Health 2021, 9, 90–98. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E.; Li, G. Approved antiviral drugs over the past 50 years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, J.M.; Monogue, M.L.; Jodlowski, T.Z.; Cutrell, J.B. Pharmacologic treatments for coronavirus disease 2019 (COVID-19): A Review. JAMA 2020, 323, 1824–1836. [Google Scholar] [CrossRef] [PubMed]
- Biot, C.; Daher, W.; Chavain, N.; Fandeur, T.; Khalife, J.; Dive, D.; De Clercq, E. Design and synthesis of hydroxyferroquine derivatives with antimalarial and antiviral activities. J. Med. Chem. 2006, 49, 2845–2849. [Google Scholar] [CrossRef] [Green Version]
- Devaux, C.A.; Rolain, J.M.; Colson, P.; Raoult, D. New insights on the antiviral effects of chloroquine against coronavirus: What to expect for COVID-19? Int. J. Antimicrob. Agents 2020, 55, 105938. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Trivedi, N. Disease-drug and drug-drug interaction in COVID-19: Risk and assessment. Authorea 2020, 1–2. [Google Scholar] [CrossRef]
- Saghir, S.A.; AlGabri, N.A.; Alagawany, M.M.; Attia, Y.A.; Alyileili, S.R.; Elnesr, S.S.; Shafi, M.E.; Al-shargi, O.Y.; Al-balagi, N.; Alwajeeh, A.S.; et al. Chloroquine and Hydroxychloroquine for the Prevention and Treatment of COVID-19: A Fiction, Hope or Hype? An Updated Review. Ther. Clin. Risk Manag. 2021, 17, 371–387. [Google Scholar] [CrossRef]
- Painter, W.P.; Holman, W.; Bush, J.A.; Almazedi, F.; Malik, H.; Eraut, N.C.J.E.; Morin, M.J.; Szewczyk, L.J.; Painter, G.R. Human safety, tolerability, and pharmacokinetics of Molnupiravir, a novel broad-spectrum oral antiviral agent with activity against SARS-CoV-2. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Paymode, D.; Vasudevan, N.; Kadam, A.; Cardoso, F.; Burns, J.; Cook, D.; Stringham, R.; Snead, D. Toward a Practical, Two-Step Process for Molnupiravir from Cytidine. Preprint 2021. [Google Scholar] [CrossRef]
- Merck and Ridgeback Biotherapeutics Provide Update on Progress of Clinical Development Program for Molnupiravir, an Investigational Oral Therapeutic for the Treatment of Mild-to-Moderate COVID-19. Available online: https://www.merck.com/news/merck-and-ridgeback-biotherapeutics-provide-update-on-progress-of-clinical-development-program-for-molnupiravir-an-investigational-oral-therapeutic-for-the-treatment-of-mild-to-moderate-covid-19/ (accessed on 15 April 2021).
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef]
- Jean, S.S.; Lee, P.I.; Hsueh, P.R. Treatment options for COVID-19: The reality and challenges. J. Microbiol. Immunol. Infect. 2020, 53, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Deng, Q.; Dai, S. Remdesivir for severe acute respiratory syndrome coronavirus 2 causing COVID-19: An evaluation of the evidence. Travel. Med. Infect. Dis. 2020, 35, 101647. [Google Scholar] [CrossRef]
- Harrison, C. Coronavirus puts drug repurposing on the fast track. Nat. Biotechnol. 2020, 38, 379–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Cho, S.; Meyyur Aravamudan, V.; Sanda, H.Y.; Palraj, R.; Molton, J.S.; Venkatachalam, I. Remdesivir in Coronavirus Disease 2019 (COVID-19) treatment: A review of evidence. Infection 2021, 49, 401–410. [Google Scholar] [CrossRef]
- Gordon, C.J.; Tchesnokov, E.P.; Feng, J.Y.; Porter, D.P.; Götte, M. The antiviral compound remdesivir potently inhibits RNA-dependent RNA polymerase from Middle East respiratory syndrome coronavirus. J. Biol. Chem. 2020, 295, 4773–4779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ita, K. Coronavirus Disease (COVID-19): Current status and prospects for drug and vaccine development. Arch. Med. Res. 2021, 52, 15–24. [Google Scholar] [CrossRef]
- FDA U.S. Food and Drug. FDA Approves First Treatment for COVID-19. FDA Approval of Remdesivir. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-treatment-covid-19 (accessed on 21 April 2021).
- Eurropean Medicine Agency. Veklury. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/veklury (accessed on 21 April 2021).
- McKee, D.L.; Sternberg, A.; Stange, U.; Laufer, S.; Naujokat, C. Candidate drugs against SARS-CoV-2 and COVID-19. Pharmacol. Res. 2020, 157, 104859. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Hu, S.; Gao, J. Discovering drugs to treat coronavirus disease 2019 (COVID-19). Drug Discov. Ther. 2020, 14, 58–60. [Google Scholar] [CrossRef] [Green Version]
- Delang, L.; Abdelnabi, R.; Neyts, J. Favipiravir as a potential countermeasure against neglected and emerging RNA viruses. Antiviral. Res. 2018, 153, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Vicenti, I.; Zazzi, M.; Saladini, F. SARS-CoV-2 RNA-dependent RNA polymerase as a therapeutic target for COVID-19. Expert Opin. Ther. Pat. 2021, 1–13. [Google Scholar] [CrossRef]
- Bai, C.Q.; Mu, J.S.; Kargbo, D.; Song, Y.B.; Niu, W.K.; Nie, W.M.; Kanu, A.; Liu, W.W.; Wang, Y.P.; Dafae, F.; et al. Clinical and virological characteristics of ebola virus disease patients treated with Favipiravir (T-705)-Sierra Leone 2014. Clin. Infect. Dis. 2016, 63, 1288–1294. [Google Scholar] [CrossRef]
- Shiraki, K.; Daikoku, T. Favipiravir, an anti-influenza drug against life-threatening RNA virus infections. Pharmacol. Ther. 2020, 209, 107512. [Google Scholar] [CrossRef]
- Cai, Q.; Yang, M.; Liu, D.; Chen, J.; Shu, D.; Xia, J.; Liao, X.; Gu, Y.; Cai, Q.; Yang, Y.; et al. Experimental treatment with Favipiravir for COVID-19: An open-label control study. Engineering 2020, 6, 1192–1198. [Google Scholar] [CrossRef]
- Udwadia, Z.F.; Singh, P.; Barkate, H.; Patil, S.; Rangwala, S.; Pendse, A.; Kadam, J.; Wu, W.; Caracta, C.F.; Tandon, M. Efficacy and safety of favipiravir, an oral RNA-dependent RNA polymerase inhibitor, in mild-to-moderate COVID-19: A randomized, comparative, open-label, multicenter, phase 3 clinical trial. Int. J. Infect. Dis. 2021, 103, 62–71. [Google Scholar] [CrossRef]
- Walmsley, S.; Bernstein, B.; King, M.; Arribas, J.; Beall, G.; Ruane, P.; Johnson, M.; Johnson, D.; Lalonde, R.; Japour, A.; et al. M98-863 Study Team. Lopinavir-ritonavir versus nelfinavir for the initial treatment of HIV infection. N. Engl. J. Med. 2002, 346, 2039–2046. [Google Scholar] [CrossRef]
- Teimury, A.; Mahmoodi Khaledi, E. Current options in the treatment of COVID-19: A Review. Risk Manag. Healthc. Policy 2020, 13, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.M.; Cheng, V.C.; Hung, I.F.; Wong, M.M.; Chan, K.H.; Chani, K.S.; Kao, R.Y.; Poon, L.L.; Wong, C.L.; Guan, Y.; et al. HKU/UCH SARS study group. Rrole of iopinavir/ritonavir in the treatment of SARS: Initial virological and clinical findings. Thorax 2004, 59, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Jeon, S.; Shin, H.Y.; Kim, M.J.; Seong, Y.M.; Lee, W.J.; Choe, K.W.; Kang, Y.M.; Lee, B.; Park, S.J. Case of the index patient who caused tertiary transmission of COVID-19 infection in Korea: The Application of lopinavir/ritonavir for the treatment of COVID-19 infected pneumonia monitored by quantitative RT-PCR. J. Korean Med Sci. 2020, 35, 79. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. Trial of lopinavir-ritonavir in adults hospitalized with severe COVID-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Zhu, Z.; Lu, Z.; Xu, T.; Chen, C.; Yang, G.; Zha, T.; Lu, J.; Xue, Y. Arbidol monotherapy is superior to lopinavir/ritonavir in treating COVID-19. J. Infect. 2020, 81, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Coronavirus: COVID-19 has killed more people than SARS and MERS combined, despite lower case fatality rate. BMJ 2020, 18, 368. [Google Scholar] [CrossRef] [Green Version]
- Ahern, M.J.; Reid, C.; Gordon, T.P.; McCredie, M.; Brooks, P.M.; Jones, M. Does colchicine work? The results of the first controlled study in acute gout. Aust. N. Z. J. Med. 1987, 17, 301–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terkeltaub, R.A.; Furst, D.E.; Bennett, K.; Kook, K.A.; Crockett, R.S.; Davis, M.W. High versus low dosing of oral colchicine for early acute gout flare: Twenty-four-hour outcome of the first multicenter, randomized, double-blind, placebo-controlled, parallel-group, dose-comparison colchicine study. Arthritis Rheum. 2010, 62, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, N.; Mizushima, Y. Leucocyte movement and colchicine treatment in Behcet’s disease. Lancet 1975, 2, 813. [Google Scholar] [CrossRef]
- Davatchi, F.; Abdollahi, B.S.; Banihashemi, A.T.; Shahram, F.; Nadji, A.; Shams, H.; Davachi, C.C. Colchicine versus placebo in Behçet’s disease: Randomized, double-blind, controlled crossover trial. Mod. Rheumatol. 2009, 19, 542–549. [Google Scholar] [CrossRef]
- Richter, M.; Boldescu, V.; Graf, D.; Streicher, F.; Dimoglo, A.; Bartenschlager, R.; Klein, C.D. Synthesis, biological evaluation and molecular docking of combretastatin and colchicine derivaties and their hCE1-activated prodrugs as antiviral agents. ChemMedChem. 2019, 14, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Worachartcheewan, A.; Songtawee, N.; Siriwong, S.; Prachayasittikul, S.; Nantasenamat, C.; Prachayasittiku, V. Rational design of colchicine derivatives as anti-HIV agents via QSAR and molecular docking. Med. Chem. 2019, 15, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, N.; Firestein, B.L.; Brunetti, L. Colchicine in COVID-19: An old drug, new use. Curr. Pharmacol. Rep. 2020, 6, 137–145. [Google Scholar] [CrossRef]
- Reyes, A.Z.; Hu, K.A.; Teperman, J.; Muskardin, T.L.W.; Tardif, J.C.; Shah, B.; Pillinger, M.H. Anti-inflammatory therapy for COVID-19 infection: The case for colchicine. Ann. Rheum. Dis. 2020, 80, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Salah, H.M.; Mehta, J.L. Meta-analysis of the Effect of Colchicine on Mortality and Mechanical Ventilation in COVID-19. Am. J. Cardiol. 2021, 145, 170–172. [Google Scholar] [CrossRef]
- Lejal, N.; Tarus, B.; Bouguyon, E.; Chenavas, S.; Bertho, N.; Delmas, B.; Ruigrok, R.W.H.; Di Primo, C.; Slama-Schwoka, A. Structure-based discovery of the novel antiviral properties of Naproxen against the nucleoprotein of influenza A virus. Antimicrob. Agents Chemother. 2013, 57, 2231–2242. [Google Scholar] [CrossRef] [Green Version]
- Crofford, L.J. Use of NSAIDs in treating patients with arthritis. Arthritis Res. Ther. 2013, 15, S2. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Tai, Y.L.; Huang, L.M.; Chi, H.; Huang, F.Y.; Chiu, N.C.; Huang, C.Y.; Tu, Y.H.; Wang, J.Y.; Huang, D.T.N. Efficacy of clarithromycin-naproxen-oseltamivir combination therapy versus oseltamivir alone in hospitalized pediatric influenza patients. J. Microbiol. Immunol. Infect. 2020. [Google Scholar] [CrossRef] [PubMed]
- Vahedi, E.; Ghanei, M.; Ghazvini, A.; Azadi, H.; Izadi, M.; Panahi, Y.; Fathi, S.; Salesi, M.; Saadat, S.H.; Ghazale, A.H.; et al. The clinical value of two combination regimens in the Management of Patients Suffering from Covid-19 pneumonia: A single centered, retrospective, observational study. DARU J. Pharm. Sci. 2020, 28, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Parnham, M.J.; Haber, V.E.; Giamarellos-Bourboulis, E.J.; Perletti, G.; Verleden, G.M.; Vos, R. Azithromycin: Mechanisms of action and their relevance for clinical applications. Pharmacol. Ther. 2014, 143, 225–245. [Google Scholar] [CrossRef]
- Porter, J.D.; Watson, J.; Roberts, L.R.; Gill, S.K.; Groves, H.; Dhariwal, J.; Almond, M.H.; Wong, E.; Walton, R.P.; Jones, L.H.; et al. Identification of novel macrolides with antibacterial, anti-inflammatory and type I and III IFN-augmenting activity in airway epithelium. J. Antimicrob. Chemother. 2016, 71, 2767–2781. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.E.; Hinks, T.S.C. Azithromycin in viral infections. Rev. Med. Virol. 2021, 31, e2163. [Google Scholar] [CrossRef] [PubMed]
- Chilamakuri, R.; Agarwal, S. COVID-19: Characteristics and therapeutics. Cells 2021, 10, 206. [Google Scholar] [CrossRef]
- Esnal, D.E.; Ontiyuelo, C.M.; Rouco, M.E.N.; Cuscó, M.D.A.; Ferrández, O.; Horcajada, J.P.; Grau, S. Azithromycin in the treatment of COVID-19: A Review. Exp. Rev. Anti-Infect. Ther. 2021, 19, 147–163. [Google Scholar] [CrossRef] [PubMed]
- RECOVERY Collaborative Group. Azithromycin in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2021, 397, 605–612. [Google Scholar] [CrossRef]
- Kelleni, M.T. Nitazoxanide/azithromycin combination for COVID-19: A suggested new protocol for early management. Pharmacol. Res. 2020, 157, 104874. [Google Scholar] [CrossRef] [PubMed]
- Al Kuraishy, H.M.; Hussien, N.R.; Al Naimi, M.S.; Al Buhadily, A.K.; Al Gareeb, A.I.; Lungnier, C. Is Ivermectin–Azithromycin combination the next step for COVID-19? Biomed. Biotechnol. Res. J. 2020, 4, 101–103. [Google Scholar]
- ClinicalTrials.gov. Ivermectin-Azithromycin-Cholecalciferol (IvAzCol) Combination Therapy for COVID-19 (IvAzCol). Available online: https://clinicaltrials.gov/ct2/show/NCT04399746 (accessed on 21 April 2021).
- Khamesipour, F.; Hashemian, S.M.; Tabarsi, P.; Velayati, A.A. Review of Teicoplanin used in the prevention and treatment of serious infections caused by gram-positive bacteria and compared its effects with some other antibiotics. Biomed. Pharmacol. J. 2015, 8, 513–521. [Google Scholar]
- Wang, Y.; Cui, R.; Li, G.; Gao, Q.; Yuan, S.; Altmeyer, R.; Zou, G. Teicoplanin inhibits Ebola pseudovirus infection in cell culture. Antivir. Res. 2016, 215, 1–7. [Google Scholar] [CrossRef]
- Baron, S.A.; Devaux, C.; Colson, P.; Raoult, D.; Rolain, J.M. Teicoplanin: An alternative drug for the treatment of COVID-19? Inter. J. Antimic. Agents 2020, 55, 105944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, X.; Yu, F.; Liu, J.; Zou, F.; Pan, T.; Zhang, H. Teicoplanin potently blocks the cell entry of 2019-nCoV. Preprint 2020. [Google Scholar] [CrossRef] [Green Version]
- Kroot, E.J.A.; Huisman, A.M.; Zeben, J.V.; Wounters, J.M.G.W.; Paassen, H.C.V. Oral Pulsed Dexamethasone Therapy in Early Rheumatoid Arthritis. Ann. N. Y. Acad. Sci. 2006, 1069, 300–306. [Google Scholar] [CrossRef]
- Roila, F.; Ballatori, E.; Ruggeri, B.; De Angelis, V. Dexamethasone alone or in combination with ondansetron for the prevention of delayed nausea and vomiting induced by chemotherapy. N. Engl. J. Med. 2000, 342, 1554–1559. [Google Scholar]
- Gogali, A.; Kyriakopoulos, C.; Kostikas, K. Corticosteroids in Covid-19: One size does not fit all. Eur. Respir. J. 2021, 57, 2100224. [Google Scholar] [CrossRef] [PubMed]
- Águas, R.; Mahdi, A.; Shretta, R.; Horby, P.; Landray, M.; White, L.; The CoMo Consortium. Potential health and economic impacts of dexamethasone treatment for patients with COVID-19. Nat. Commun. 2021, 12, 915. [Google Scholar] [CrossRef] [PubMed]
- The RECOVERY Collaborative Group. Dexamethasone in hospitalized patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Halpin, D.M.G.; Singh, D.; Harfield, R.M. Inhaled corticosteroids and COVID-19: A systematic review and clinical perspective. Eur. Respir. J. 2020, 57, 2001009. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.M.; Head, H.; Neurology, H.O. Corticosteroids in autoimmune diseases. Aust Prescr. 1999, 22, 9–11. [Google Scholar]
- Yamano, Y.; Taniguchi, H.; Kondoh, Y.; Ando, M.; Kataoka, K.; Furukawa, T.; Johkoh, T.; Fukuoka, J.; Sakamoto, K.; Hasegawa, Y. Multidimensional improvement in connective tissue disease-associated interstitial lung disease: Two courses of pulse dose methylprednisolone followed by low-dose prednisone and tacrolimus. Respirology 2018, 23, 1041–1048. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, X.; Huang, Y.; Shan, H.; Huang, J. Successful use of methylprednisolone for treating severe COVID-19. J. Allergy Clin. Immunol. 2020, 146, 325–327. [Google Scholar] [CrossRef]
- Gopalaswamy, R.; Subbian, S. Corticosteroids for COVID-19 Therapy: Potential Implications on Tuberculosis. Int. J. Mol. Sci. 2021, 22, 3773. [Google Scholar] [CrossRef]
- Rnajbar, K.; Moghadami, M.; Mirahmadizadeh, A.; Fallahi, M.J.; Khaloo, V.; Shahriarirad, R.; Erfani, A.; Khodamoradi, Z.; Saadi, M.H.G. Methylprednisolone or dexamethasone, which one is superior corticosteroid in the treatment of hospitalized COVID-19 patients: A triple-blinded randomized controlled trial. BMC Infect. Dis. 2021, 21, 337. [Google Scholar]
- Rossignol, J.R. Nitazoxanide: A first-in-class broad-spectrum antiviral agent. Antivir. Res. 2014, 110, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Geller, M.; Blum, V.F.; Oliveira, L.; Mezitis, S.G.E.; Nigri, R.; Neto, M.S.; Nunes, C.P.; da Fonseca, A.S.; Cunha, K.S. Nitazoxanide: Antiviral properties relevant to current global situation. Int. J. Curr. Res. 2020, 12, 11733–11737. [Google Scholar]
- Rossignol, J.F. Nitazoxanide, a new drug candidate for the treatment of Middle East respiratory syndrome coronavirus. J. Infect. Public Health 2016, 9, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Lokhande, A.S.; Devarajan, P.V. A review on possible mechanistic insights of Nitazoxanide for repurposing in COVID-19. Eur. J. Pharmacol. 2021, 891, 173748. [Google Scholar] [CrossRef]
- Campbell, W.C. Ivermectin as an antiparasitic agent for use in humans. Annu. Rev. Microbiol. 1991, 45, 445–474. [Google Scholar] [CrossRef] [PubMed]
- Heidary, F.; Gharebaghi, R. Ivermectin: A systematic review from antiviral effects to COVID-19 complementary regimen. J. Antibiot. 2020, 73, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in-vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef]
- Jans, D.A.; Wagstaff, K.M. The broad-spectrum host-directed agent ivermectin as an antiviral for SARS-CoV-2? Biochem. Biophyl. Res. Commun. 2021, 538, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Morales, R.L.; Hernandez, P.M.; Flores, Y.N.; Romero, P.O.; Rugerio, E.C.; Zamora, A.N.; Galdamez, D.R.H.; Duenas, D.K.R.; Salmeron, J. Effectiveness of a multidrug therapy consisting of ivermectin, azithromycin, montelukast and acetylsalicylic acid to prevent hospitalization and death among ambulatory COVID-19 cases in Tlaxcala, Mexico. Int. J. Infect. Dis. 2021, 105, 598–605. [Google Scholar] [CrossRef]
- Ramsey, M.L.; Nuttall, J.; Hart, P.A.; TACTIC Investigate Team. A phase 1/2 trial to evaluate the pharmacokinetics, safety, and efficacy of NI-03 in patients with chronic pancreatitis: Study protocol for a randomized controlled trial on the assessment of camostat treatment in chronic pancreatitis (TACTIC). Trials 2019, 20, 501. [Google Scholar] [CrossRef] [Green Version]
- Ota, S.; Hara, Y.; Kanoh, S.; Shinoda, M.; Kawano, S.; Fujikura, Y.; Kawana, A.; Shinkai, M. Acute eosinophilic pneumonia caused by camostat mesilate: The first case report. Respir. Med. Case. Rep. 2016, 19, 21–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breining, P.; Frølund, A.L.; Højen, J.F.; Gunst, J.D.; Staerke, N.B.; Saedder, E.; Thomas, M.C.; Little, P.; Nielsen, L.P.; Søgaard, O.S.; et al. Camostat Mesylate against SARS-CoV-2 and COVID-19-rationale, dosing and safety. Basic Clin. Pharmacol. Toxicol. 2021, 128, 204–212. [Google Scholar] [CrossRef]
- Hoffmann, M.; Winkler, H.H.; Smith, J.C.; Krüger, N.; Sørensen, L.K.; Søgaard, O.S.; Hasselstrøm, J.B.; Winkler, M.; Hempel, T.; Raich, L.; et al. Camostat mesylate inhibits SARS-CoV-2 activation by TMPRSS2-related proteases and its metabolite GBPA exerts antiviral activity. EBioMedicine. 2021, 65, 103255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Ma, Y.X.; Sun, Y.; Cao, Y.B.; Li, Q.; Xu, C.A. Gemcitabine in combination with a second cytotoxic agent in the first-line treatment of locally advanced or metastatic pancreatic cancer: A systematic review and meta-analysis. Target. Oncol. 2017, 12, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Clouser, C.L.; Holtz, C.M.; Mullett, M.; Crankshaw, D.L.; Briggs, J.E.; O’Sullivan, M.G.; Patterson, S.E.; Mansky, L.M. Activity of a novel combined antiretroviral therapy of gemcitabine and decitabine in a mouse model for HIV-1. Antimicrob. Agents Chemother. 2012, 56, 1942–1948. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Middle East Respiratory Syndrome-Coronavirus (MERS-CoV): An updated overview and pharmacotherapeutics. Med. Chem. 2015, 5, 361–372. [Google Scholar] [CrossRef]
- Bagheri, A.; Moezzi, S.M.I.; Mosaddeghi, P.; Parashkouhi, S.M.; Hoseini, S.M.F.; Badakhshan, F.; Negahdaripour, M. Interferon-inducer antivirals: Potential candidates to combat COVID-19. Int. Immunopharmacol. 2021, 91, 107245. [Google Scholar] [CrossRef] [PubMed]
- Passerini, C.G.; Antolini, L.; Mahon, F.X.; Guilhot, F.; Deininger, M.; Fava, C.; Nagler, A.; Casa, C.M.D.; Morra, E.; Abruzzese, E.; et al. Multicenter independent assessment of outcomes in chronic myeloid leukemia patients treated with imatinib. J. Natl. Cancer Inst. 2011, 103, 553–561. [Google Scholar] [CrossRef]
- Ortega, A.M.; Bello, D.B.; Barroso, C.L.; Pérez, B.F.; Millán, M.A.D.M.; García, V.G.V.; Sedano, A.I.F.; Castrillero, E.C.; Giardín, J.M.R.; Ruiz, J.R.; et al. Imatinib for COVID-19: A case report. Clin. Immunol. 2020, 218, 1–3. [Google Scholar]
- Ortega, A.M.; Prado, L.R.; Pérez, B.F.; Barrios, B.J.; Sedano, A.S.I.F.; Parra, C.J.G.; Muniesa, B.H.; Millán, M.A.D.; Cerezo, E.M.; Nasarre, A.O.; et al. Early clinical experience with imatinib in COVID-19: Searching for a dual effect. J. Infect. 2021, in press. [Google Scholar]
- Jordan, V.C. Fourteenth Gaddum Memorial Lecture. A current view of tamoxifen for the treatment and prevention of breast cancer. Br. J. Pharmacol. 1993, 110, 507–517. [Google Scholar] [CrossRef]
- Montoya, M.C.; Krysanb, D.J. Repurposing estrogen receptor antagonists for the treatment of infectious disease. mBio 2018, 9, e02272-18. [Google Scholar] [CrossRef] [Green Version]
- Almosawey, H.A.S.; Al Khikani, F.H.O.; Hameed, R.M.; Abdullah, Y.J.; Al Ibraheemi, M.K.M.; Al Asadi, A.A. Tamoxifen from chemotherapy to antiviral drug: Possible activity against COVID-19. Biomed. Biotechnol. Res. J. 2020, 4, 108–116. [Google Scholar]
- Karacin, C.; Karacin, P.; Ergun, Y. Tamoxifen in breast cancer survivors with COVID 19: Stop or go? Med. Oncol. 2021, 38, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Varga, B.; Csonka, A.; Csonka, A.; Molnar, J.; Amaral, L.R.D.; Spengler, G. Possible biological and clinical applications of phenothiazines. Anticancer Res. 2017, 37, 5983–5993. [Google Scholar] [PubMed] [Green Version]
- Muric, N.N.; Arsenijevic, N.N.; Borovcanin, M.M. Chlorpromazine as a potential Antipsychotic Choice in COVID-19 Treatment. Front. Psychiatry 2020, 11, 612347. [Google Scholar] [CrossRef]
- Hoertel, N.; Rico, M.S.; Vernet, R.; Jannot, A.S.; Neuraz, A.; Blanco, C.; Lemogne, C.; Airagnes, G.; Paris, N.; Daniel, C.; et al. Observational study of chlorpromazine in hospitalized patients with COVID-19. Clin. Drug Investig. 2021, 41, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Sukhatme, V.P.; Reiersen, A.M.; Vayttaden, S.J.; Sukhatme, V.V. Fluvoxamine: A Rewiew of its mechanism of action and its role in Covid-19. Front. Pharmacol. 2021, 12, 652688. [Google Scholar] [CrossRef] [PubMed]
- Lenze, E.J.; Mattar, C.; Zorumski, C.F.; Stevens, A.; Schweiger, J.; Nicol, G.E.; Miller, J.P.; Yang, L.; Yingling, M.; Avidan, M.S.; et al. Fluvoxamine vs placebo and clinical deterioration in outpatients with symptomatic COVID-19 A randomized clinical trial. JAMA 2020, 324, 2292–2300. [Google Scholar] [CrossRef] [PubMed]
- Seftel, D.; Boulware, D.R. Prospective cohort of fluvoxamine for early treatment of COVID-19. Open Forum Infect. Dis. 2021, 8, ofab050. [Google Scholar] [CrossRef]
- Deppe, S.; Boger, R.H.; Weiss, J.; Benndorf, R.A. Telmisartan: A review of its pharmacodynamic and pharmacokinetic properties. Expert Opin. Drug Metab. Toxicol. 2010, 6, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Huang, F.; Xu, J.; Yang, P.; Qin, Y.; Lv, J.; Zhang, S.; Ye, L.; Gong, M.; Liu, Z.; et al. Antihypertensive drugs are associated with reduced fatal outcomes and improved clinical characteristics in elderly COVID-19 patients. Cell Discov. 2020, 6, 77. [Google Scholar] [CrossRef]
- Bengtson, C.D.; Montgomery, R.N.; Nazir, U.; Satterwhite, L.; Kim, M.D.; Bahr, N.C.; Castro, M.; Baumlin, N.; Salathe, M. An open label trial to assess safety of losartan for treating worsening respiratory illness in COVID-19. Front. Med. 2021, 8, 630209. [Google Scholar] [CrossRef]
- Nejat, R.; Sadr, A.S. Are losartan and imatinib efective against SARS CoV2 pathogenesis? A pathophysiologic based in silico study. Silico Pharmacol. 2021, 9, 1. [Google Scholar] [PubMed]
- UK Prospective Diabetes Study Group. Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). Lancet 1998, 352, 854–865. [Google Scholar] [CrossRef]
- Bramante, C.T.; Ingraham, N.E.; Murray, T.A.; Marmor, S.; Hovertsen, S.; Gronski, J.; McNeil, C.; Feng, R.; Guzman, G.; Abdelwahab, N.; et al. Metformin and risk of mortality in patients hospitalised with COVID-19: A retrospective cohort analysis. Lancet Healthy Longev. 2021, 2, e34–e41. [Google Scholar] [CrossRef]
- Wang, J.; Cooper, J.M.; Gokhale, K.; Acosta-Mena, D.; Dhalla, S.; Byne, N.; Chandan, J.S.; Anand, A.; Okoth, K.; Subramanian, A.; et al. Association of Metformin with Susceptibility to COVID-19 in people with type 2 diabetes. J. Clin. Endocrinol. Metab. 2021, 106, 1255–1268. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, S.N. Sirolimus: Its discovery, biological properties, and mechanism of action. Transplant. Proc. 2003, 35, 7–14. [Google Scholar] [CrossRef]
- Bischof, E.; Siow, R.C.; Zhavoronkov, A.; Kaeberlein, M. The potential of rapalogs to enhance resilience against SARS-CoV-2 infection and reduce the severity of COVID-19. Lancet Healthy Longev. 2021, 2, e105–e111. [Google Scholar] [CrossRef]
- Ramaiah, M.J. mTOR inhibition and p53 activation, microRNAs: The possible therapy against pandemic COVID-19. Gene Rep. 2020, 20, 100765. [Google Scholar] [CrossRef]
- Choudhary, S.; Sharma, K.; Silakari, O. The interplay between inflammatory pathways and COVID-19: A critical review on pathogenesis and therapeutic options. Microb. Pathog. 2021, 150, 104673. [Google Scholar] [CrossRef] [PubMed]
- Cure, E.; Kucuk, A.; Cure, M.C. Cyclosporine therapy in cytokine storm due to coronavirus disease 2019 (COVID-19). Rheumatol. Int. 2020, 40, 1177–1179. [Google Scholar] [CrossRef]
- Sanchez-Pernaute, O.; Romero-Bueno, F. Why choose cyclosporin A as first-line therapy in COVID-19 pneumonia. Reumatol. Clínica 2020. [Google Scholar] [CrossRef]
- Bozzi, G.; Mangioni, D.; Minoia, F.; Aliberti, S.; Grasselli, G.; Barbetta, L.; Castelli, V.; Palomba, E.; Alagna, L.; Lombardi, A.; et al. Anakinra combined with methylprednisolone in patients with severe COVID-19 pneumonia and hyperinflammation: An observational cohort study. J. Allergy Clin. Immunol. 2021, 147, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Kooistra, E.J.; Waalders, N.J.B.; Grondman, I.; Janssen, N.A.F.; De Nooijer, A.H.; Netea, M.G.; Van de Veerdonk, F.L.; Ewalds, E.; Van Der Hoeven, J.G.; Kox, M.; et al. Anakinra treatment in critically ill COVID-19 patients: A prospective cohort study. Crit. Care 2020, 24, 688. [Google Scholar] [CrossRef]
- Lilly Announces Agreement with U.S. Government to Supply 300,000 Vials of Investigational Neutralizing Antibody Bam-lanivimab (LY-CoV555) in An Effort to Fight COVID-19 (Press release). Eli Lilly Co. 2020. Available online: https://investor.lilly.com/news-releases/news-release-details/lilly-announces-agreement-us-government-supply-300000-vials (accessed on 28 October 2020).
- Gottlieb, R.L.; Nirula, A.; Chen, P.; Boscia, J.; Heller, B.; Morris, J.; Huhn, G.; Cardona, J.; Mocherla, B.; Stosor, V.; et al. Effect of bamlanivimab as monotherapy or in combination with etesevimab on viral load in patients with mild to moderate COVID-19. A randomized clinical trial. JAMA 2021, 325, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, S.C.J.; Tse, C.L.Y.; Burry, L.; Dresser, L.D. Baricitinib: A Review of pharmacology, safety, and emerging clinical experience in COVID-19. Pharmacotherapy 2020, 40, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Qiao, W.; Zhang, J.; Qi, Z. Baricitinib, a drug with potential effect to prevent SARS-COV-2 from entering target cells and control cytokine storm induced by COVID-19. Int. Immunopharmacol. 2020, 86, 106749. [Google Scholar] [CrossRef] [PubMed]
- Goletti, D.; Cantini, F. Baricitinib therapy in Covid-19 pneumonia-An unmet need fulfilled. N. Engl. J. Med. 2021, 384, 867–869. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Hurwitz, H.I.; Sandler, A.B.; Miles, D.; Coleman, R.L.; Deurloo, R.; Chinot, O.L. Bevacizumab (Avastin®) in cancer treatment: A review of 15 years of clinical experience and future outlook. Cancer Treat Rev. 2020, 86, 102017. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Xu, F.; Aondio, G.; Li, Y.; Fumagalli, A.; Lu, M.; Valmadre, G.; Wei, J.; Bian, Y.; Canesi, M.; et al. Efficacy and tolerability of bevacizumab in patients with severe Covid-19. Nat. Commun. 2021, 12, 814. [Google Scholar] [CrossRef]
- Lamb, Y.N.; Deeks, D. Sarilumab: A review in moderate to severe rheumatoid arthritis. Drugs 2018, 78, 929–940. [Google Scholar] [CrossRef]
- Tu, Y.F.; Chien, C.S.; Yarmishyn, A.A.; Lin, Y.Y.; Luo, Y.H.; Lin, Y.T.; Lai, W.Y.; Yang, D.M.; Chou, S.J.; Yang, Y.P.; et al. A review of SARS-CoV-2 and the ongoing clinical trials. Int. J. Mol. Sci. 2020, 21, 2657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benucci, M.; Giannas, G.; Cecchini, P.; Li Gobbi, F.; Damiani, A.; Grossi, V.; Infantino, M.; Manfredi, M. COVID-19 pneumonia treated with Sarilumab: A clinical series of eight patients. J. Med. Virol. 2020, 92, 2368–2370. [Google Scholar] [CrossRef]
- Masia, M.; Fernandez-Gonzalez, M.; Padilla, S.; Ortega, P.; Garcia, J.A.; Agullo, V.; Garcia-Abellan, J.; Telenti, G.; Guillen, L.; Gutierrez, F. Impact of interleukin-6 blockade with tocilizumab on SARS-CoV-2 viral kinetics and antibody responses in patients with COVID-19: A prospective cohort study. EBioMedicine 2020, 60, 102999. [Google Scholar] [CrossRef]
- Stone, J.H.; Frigault, M.J.; Serling Boyd, N.J.; Fernandes, A.D.; Harvey, L.; Foulkes, A.S.; Horick, N.K.; Healy, B.C.; Shah, R.; Bensaci, A.M.; et al. Efficacy of tocilizumab in patients hospitalized with Covid-19. N. Engl. J. Med. 2020, 383, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Padron, E.; Painter, J.S.; Kunigal, S.; Mailloux, A.W.; McGraw, K.; McDaniel, J.M.; Kim, E.; Bebbington, C.; Baer, M.; Yarraton, G.; et al. GM-CSF-dependent pSTAT5 sensitivity is a feature with therapeutic potential in chronic myelomonocytic leukemia. Blood 2013, 121, 5068–5077. [Google Scholar] [CrossRef] [PubMed]
- Temesgen, Z.; Burger, C.D.; Baker, J.; Polk, C.; Libertin, C.; Kelley, C.; Marconi, V.C.; Orenstein, R.; Durrant, C.; Chappell, D.; et al. Lenzilumab efficacy and safety in newly hospitalized Covid-19 subjects: Results from the live-air phase 3 randomized double-blind placebocontrolled trial. medRxiv 2021. [Google Scholar] [CrossRef]
- Talukdar, D.; Jain, V.; Balaramnavar, V.; Srivastava, S.P.; Sivanandy, P.; Gupta, M.M. Potential Drugs for COVID -19 Treatment Management With Their Contraindications and Drug- Drug Interaction. Preprints 2021. [Google Scholar] [CrossRef]
- Erlanson, D.A. Many small steps towards a COVID-19 drug. Nat. Commun. 2020, 11, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pant, S.; Singh, M.; Ravichandiran, V.; Murty, U.S.N.; Srivastava, H.K. Peptide-like and small-molecule inhibitors against Covid-19. J. Biomol. Struct. Dyn. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Villa, F.X.; Duran-Iturbide, N.A.; Avila-Zarraga, J.G. Synthesis, molecular docking, and in-silico ADME/Tox profiling studies of new 1-aryl-5-(3-azidopropyl)indol-4-ones: Potential inhibitors of SARS CoV-2 main protease. Bioorg. Chem. 2021, 106, 104497. [Google Scholar] [CrossRef]
- Satyanarayana, M.V.; Reddy, A.G.; Yedukondalu, M.; Tej, M.B.; Hossain, K.A.; Rao, M.V.B.; Pal, M. In silico assessment and sonochemical synthesis of 2-alkynyl 3-chloropyrazines as prospective ligands for SARS-CoV-2. J. Mol. Struct. 2021, 1231, 129981. [Google Scholar] [CrossRef] [PubMed]
- Chidambaram, S.; El-Sheikh, M.A.; Alfarhan, A.H.; Radhakrishnan, S.; Akbar, I. Synthesis of novel coumarin analogues: Investigation of molecular docking interaction of SARS-CoV-2 proteins with natural and synthetic coumarin analogues and their pharmacokinetics studies. Saudi J. Biol. Sci. 2021, 28, 1100–1108. [Google Scholar] [CrossRef]
- Chhetri, A.; Chettri, S.; Rai, P.; Sinha, B.; Brahman, D. Exploration of inhibitory action of azo imidazole derivatives against COVID-19 main protease (Mpro): A computational study. J. Mol. Struct. 2021, 1224, 129178. [Google Scholar] [CrossRef]
- Al-Janabi, A.S.M.; Elzupir, A.O.; Yousef, T.A. Synthesis, anti-bacterial evaluation, DFT study and molecular docking as a potential 3-chymotrypsin-like protease (3CLpro) of SARS-CoV-2 inhibitors of a novel Schiff bases. J. Mol. Struct. 2021, 1228, 129454. [Google Scholar] [CrossRef] [PubMed]
- Özkan, H.; Adem, Ş. Synthesis, spectroscopic characterizations of novel norcantharimides, their ADME properties and docking studies against COVID-19 Mpro. ChemistrySelect 2020, 5, 5422–5428. [Google Scholar] [CrossRef] [PubMed]
- Chemboli, R.; Kapavarapu, R.; Deepti, K.; Prasad, K.R.S.; Reddy, A.G.; Kumar, A.V.D.N.; Rao, M.V.B.; Pal, M. Pyrrolo[2,3-b]quinoxalines in attenuating cytokine storm in COVID-19: Their sonochemical synthesis and in silico / in-vitro assessment. J. Mol. Struct. 2021, 1230, 129868. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.A.; Saundane, A.R.; Meti, R.S.; Vennapu, D.R. Synthesis of novel indolo[3,2-c] isoquinoline derivatives bearing pyrimidine, piperazine rings and their biological evaluation and docking studies against COVID-19 virus main protease. J. Mol. Struct. 2021, 1229, 129829. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Hattori, Y.; Kobayashi, K.; Akaji, K. Evaluation of a non-prime site substituent and warheads combined with a decahydroisoquinolin scaffold as a SARS 3CL protease inhibitor. Bioorg. Med. Chem. 2019, 27, 425–435. [Google Scholar] [CrossRef]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Zhang, B.; Jiang, X.M.; Su, H.; Li, J.; Zhao, Y.; Xie, X.; Jin, Z.; Peng, J.; Liu, F.; et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Lin, D.; Kusov, Y.; Nian, Y.; Ma, Q.; Wang, J.; Von Brunn, A.; Leyssen, P.; Lanko, K.; Neyts, J.; et al. α-Ketoamides as broad-spectrum inhibitors of coronavirus and enterovirus replication: Structure-based design, synthesis, and activity assessment. J. Med. Chem. 2020, 63, 4562–4578. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Beer, J.C.; Sankaranarayanan, N.V.; Swanson-Mungerson, M.; Desai, U.R. Discovering small-molecule therapeutics against SARS-CoV-2. Drug Discov. Today 2020, 25, 1535–1544. [Google Scholar] [CrossRef]
- Rizou, M.; Galanakis, I.M.; Aldawoud, T.M.; Galanakis, C.M. Safety of foods, food supply chain and environment within the COVID-19 pandemic. Trends Food Sci. Technol. 2020, 102, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Laborde, D.; Martin, W.; Swinnen, J.; Vos, R. COVID-19 risks to global food security. Science 2020, 369, 500–502. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, S.; Toussaint, E.C. Food access in crisis: Food security and COVID-19. Ecol. Econ. 2021, 180, 1–11. [Google Scholar] [CrossRef]
- Swinnen, J.; McDermott, J. COVID-19 and global food security. EuroChoices 2020, 19, 26–33. [Google Scholar] [CrossRef]
- Chitrakar, B.; Zhang, M.; Bhandari, B. Improvement strategies of food supply chain through novel food processing technologies during COVID-19 pandemic. Food Control 2021, 125, 1–10. [Google Scholar] [CrossRef]
- Torero, M. COVID-19 and Global Food Security-Prepare Food Systems for a Long-Haul Fight against COVID-19; International Food Policy Research Institute: Washington, DC, USA, 2020; pp. 118–121. [Google Scholar]
- FDA (U.S. Food and Drup Administration). Coronavirus Disease 2019 (COVID-19) and the Food Supply Chain. Available online: https://www.fda.gov/food/food-safety-during-emergencies/food-safety-and-coronavirus-disease-2019-covid-19 (accessed on 15 April 2020).
- WHO (World Health Organization). Coronavirus Disease 2019 (COVID-19) Situation Report–32. Available online: https://www.who.int/docs/default-source/coronaviruse/situationreports/20200221-sitrep-32-covid-19.pdf?sfvrsn=4802d 089_2 (accessed on 15 April 2020).
- Arens, M. Tönnies Meat-Processing Plant in Germany Restarted Despite Ongoing Danger from COVID-19. Available online: https://www.wsws.org/en/articles/2020/07/20/tonn-j20.html (accessed on 23 April 2020).
- Xinhua Headlines: China’s Dalian Going All Out to Stem New COVID-19 Infections. Available online: http://www.xinhuanet.com/english/2020-07/30/c_139249565.htm (accessed on 15 April 2020).
- China Recalls Ecuadorian Shrimp from Three Companies after COVID-19 Risks Found. Available online: https://www.globaltimes.cn/content/1194334.shtml (accessed on 15 April 2020).
- Pei, S.; Kandula, S.; Shaman, J. Differential effects of intervention timing on COVID-19 spread in the U.S. Sci. Adv. 2020, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Roy, P.K.; Hossain, I.; Byun, K.H.; Choi, C.; Ha, S.D. COVID-19 pandemic crisis and food safety: Implications and inactivation strategies. Trends Food Sci. Technol. 2021, 25–36. [Google Scholar] [CrossRef]
- Sportelli, M.C.; Izzi, M.; Kukushkina, E.A.; Hossain, S.I.; Picca, R.A.; Ditaranto, N.; Cioffi, N. Can nanotechnology and materials science help the fight against SARS-CoV 2? Nanomaterials 2020, 10, 802. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.-C.; Zhang, M.; Mujumdar, A.S.; Adhikari, B. Recent developments in smart freezing technology applied to fresh foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Mohd Ali, M.; Hashim, N.; Abd Aziz, S.; Lasekan, O. Principles and recent advances in electronic nose for quality inspection of agricultural and food products. Trends Food Sci. Technol. 2020, 99, 1–10. [Google Scholar] [CrossRef]
- Khan, A.; Munir, M.T.; Yu, W.; Young, B.R. A review towards hyperspectral imaging for real-time quality control of food products with an illustrative case study of milk powder production. Food Bioproc. Tech. 2020, 13, 739–752. [Google Scholar] [CrossRef]
- Han, B.; Hoang, B.X. Opinions on the current pandemic of COVID-19: Use functional food to boost our immune functions. J. Infect. Public Health 2020, 13, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Tripathi, M.K.; Yasir, M.; Khare, R.; Tripathi, M.K.; Shrivastava, R. Potential inhibitors for SARS-CoV-2 and functional food components as nutritional supplement for COVID-19: A review. Plant Food Hum. Nutr. 2020, 75, 458–466. [Google Scholar] [CrossRef]
- Park, Y.; Subar, A.F.; Hollenbeck, A.; Schatzkin, A. Dietary fiber intake and mortality in the NIH-AARP diet and health study. Int. Arch. Intern. Med. 2011, 171, 1061–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripari, N.; Sartori, A.A.; Da Silva Honorio, M.; Conte, F.L.; Tasca, K.I.; Santiago, K.B.; Sforcin, J.M. Propolis antiviral and immunomodulatory activity: A review and perspectives for COVID-19 treatment. J. Pharm. Pharmacol. 2021, 73, 281–299. [Google Scholar] [CrossRef]
- Maruta, H.; He, H. PAK1-blockers: Potential therapeutics against COVID-19. Med. Drug Discov. 2020, 6, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Sanduzzi Zamparelli, M.; Santoro, C.; Ciampaglia, R.; Stornaiuolo, M.; Tenore, G.C.; Novellino, E. May polyphenols have a role against coronavirus infection? An overview of in-vitro evidence. Front. Med. 2020, 7, 1–7. [Google Scholar] [CrossRef]
- Suravajhala, R.; Parashar, A.; Malik, B.; Nagaraj, A.V.; Padmanaban, G.; Kishor, P.K. Comparative docking studies on curcumin with COVID-19 proteins. Preprints 2020, 1–15. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Reiter, R.J. COVID-19: Melatonin as a potential adjuvant treatment. Life Sci. 2020, 250, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, J.; Hekmat, S.; Reid, G. The development of micronutrient supplemented probiotic yogurt for people living with HIV: Laboratory testing and sensory evaluation. Innov. Food Sci. Emerg. Technol. 2021, 12, 79–84. [Google Scholar] [CrossRef]
- Lips, P.; Cashman, K.D.; Lamberg-Allardt, C.; Bischoff-Ferrari, H.A.; Obermayer-Pietsch, B.; Bianchi, M.L.; Bouillon, R. Current vitamin D status in European and Middle East countries and strategies to prevent vitamin D deficiency: A position statement of the European Calcified Tissue Society. Eur. J. Endocrinol. 2019, 180, 23–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, L.; Zhao, W.; Wang, J.; Wang, H.; Zhao, Y.; Tseng, Y. Extra dose of vitamin C based on a daily supplementation shortens the common cold: A meta-analysis of 9 randomized controlled trials. BioMed Res. Int. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Calder, S.D.; Claessen, M.; Ebbels, S.; Leitão, S. The efficacy of an explicit intervention approach to improve past tense marking for early school-age children with developmental language disorder. J. Speech Lang. Heart Res. 2021, 64, 91–104. [Google Scholar] [CrossRef]
- Mossad, S.B.; Macknin, M.L.; Medendorp, S.V.; Mason, P. Zinc gluconate lozenges for treating the common cold. A randomized, doulbe-blind placebo-controlled study. ACP J. Club 1996, 125, 81–88. [Google Scholar]
- Paraiso, I.L.; Revel, J.S.; Stevens, J.F. Potential use of polyphenols in the battle against COVID-19. Curr. Opin. Food Sci. 2020, 32, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Omrani, M.; Keshavarz, M.; Ebrahimi, S.N.; Mehrabi, M.; McGaw, L.J.; Abdalla, M.A.; Mehrbod, P. Potential natural products against respiratory viruses: A perspective to develop anti-COVID-19. Front. Pharmacol. 2021, 11, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Boozari, M.; Hosseinzadeh, H. Natural Products for COVID-19 prevention and treatment regarding to previous coronavirus infections and novel studies. Phytother. Res. 2021, 35, 864–876. [Google Scholar] [CrossRef]
- Wijayasinghe, Y.S.; Bhansali, P.; Viola, R.E.; Kamal, M.A.; Poddar, N.K. Natural products: A rich source of antiviral drug lead candidates for the management of COVID-19. Curr. Pharm. Des. 2021, 27, 1–26. [Google Scholar]
- Da Silva, A.A.; Wiedemann, L.S.M.; Veiga-Junior, V.F. Natural products’ role against COVID-19. RSC Adv. 2020, 10, 23379–23393. [Google Scholar]
- Yonekawa, M.; Shimizu, M.; Kaneko, A.; Matsumura, J.; Takahashi, H. Suppression of R5-type of HIV-1 in CD4 + NKT cells by Vδ1 + T cells activated by flavonoid glycosides, hesperidin and linarin. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da, J.; Xu, M.; Wang, Y.; Li, W.; Lu, M.; Wang, Z. Kaempferol promotes apoptosis while inhibiting cell proliferation via androgen-dependent pathway and suppressing vasculogenic mimicry and invasion in prostate cancer. Anal. Cell. Pathol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benarba, B.; Pandiella, A. Medicinal pants as sources of active molecules against COVID-19. Front. Pharmacol. 2020, 11, 1189. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F.; Khan, A.; Bertuccioli, A.; Maffioli, P.; Derosa, G.; Khan, S.; Khan, B.A.; Nigar, R.; Ujjan, I.; Devrajani, B.R. Quercetin phytosome® as a potential candidate for managing COVID-19. Minerva Gastroenterol. Dietol. 2020, 66, 1–6. [Google Scholar]
- Masiello, P.; Novelli, M.; Beffy, P.; Menegazzi, M. Can hypericum perforatum (SJW) prevent cytokine storm in COVID-19 Patients? Phytother. Res. 2020, 34, 1471–1473. [Google Scholar] [CrossRef] [PubMed]
- Meneguzzo, F.; Ciriminna, R.; Zabini, F.; Pagliaro, M. Hydrodynamic Cavitation-based rapid expansion of hesperidin-rich products from waste citrus peel as a potential tool against COVID-19. Processes 2020, 8, 549. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.M.; Shehata, M.G.; Olfat, M.; Awad, E.; El-Sohaimy, S.A. Inhibition of COVID-19 RNA-dependent RNA polymerase by natural bioactive compounds: Molecular docking analysis. Egypt. J. Chem. 2012, 64, 1989–2001. [Google Scholar]
- Xu, D.; Lin, T.H.; Li, S.; Da, J.; Wen, X.Q.; Ding, J.; Chang, C.; Yeh, S. Cryptotanshinone suppresses androgen receptor-mediated growth in androgen dependent and castration resistant prostate cancer cells. Cancer Lett. 2012, 316, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of natural products as potential inhibitors of COVID-19 main protease (Mpro): In-silico evidences. Nat. Prod. Bioprospect. 2020, 10, 297–306. [Google Scholar] [CrossRef]
- Ahmad, A.; Rehman, M.U.; Alkharfy, K.M. An alternative approach to minimize the risk of coronavirus (COVID-19) and similar infections. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4030–4034. [Google Scholar] [PubMed]
- Sekiou, O.; Bouziane, I.; Bouslama, Z.; Djemel, A. In-silico identification of potent inhibitors of COVID-19 main protease (Mpro) and Angiotensin converting enzyme 2 (ACE2) from natural products: Quercetin, hispidulin, and Ccrsimaritin exhibited better potential inhibition than hydroxy-chloroquine against COVID-19 main protease active site and ACE2. Preprint 2020. [Google Scholar] [CrossRef]
- Huang, J.; Tao, G.; Liu, J.; Cai, J.; Huang, Z.; Chen, J. Current prevention of COVID-19: Natural products and herbal medicine. Front. Pharmacol. 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Thota, S.M.; Balan, V.; Sivaramakrishnan, V. Natural products as home-based prophylactic and symptom management agents in the setting of COVID-19. Phytother. Res. 2020, 34, 3148–3167. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Huang, J.; Yeung, A.W.K.; Tzvetkov, N.T.; Horba’nczuk, J.O.; Willschke, H.; Gai, Z.; Atanasov, A.G. The Significance of Natural Product Derivatives and Traditional Medicine for COVID-19. Processes 2020, 8, 937. [Google Scholar] [CrossRef]






| Agent | Chemical Structure | Classification | Approved for |
|---|---|---|---|
| Molnupiravir |  | Antiviral | Phase 3 trial for COVID-19 treatment |
| Remdesivir |  | Antiviral | Treatment of COVID-19 and Ebola |
| Favipiravir |  | Antiviral | Treatment of Influenza |
| Chloroquine |  | Antimalarial | Treatment of Malaria |
| Hydroxychloro-quine |  | Antimalarial | Treatment of Malaria and some auto-immune diseases |
| Lopinavir |  | Antiviral | Treatment of HIV |
| Ritonavir |  | Antiviral | Treatment of HIV |
| Colchicine |  | Anti-inflammatory | Treatment of familial Mediterranean fever (FMF) and acute gout flares |
| Naproxen |  | Anti-inflammatory | Treatment of acute gout, ankylosing spondylitis, bursitis, polyarticular juvenile idiopathic arthritis, osteoarthritis, tendonitis, rheumatoid arthritis, pain, and primary dysmenorrhea |
| Azithromycin |  | Antibacterial | Treatment of a number of bacterial infections. |
| Teicoplanin |  | Antibacterial | Treatment of a number of Gram-positive bacterial infections and Ebola |
| Dexamethasone |  | Corticosteroid | Treatment of a number of inflammatory conditions and for reducing the body’s immune response in the treatment of allergies and autoimmune diseases |
| Methylpredniso-lone |  | Corticosteroid | Treatment of allergic conditions, arthritis, asthma exacerbations, long-term asthma maintenance, acute exacerbation of multiple sclerosis |
| Nitazoxanide |  | Antiparasitic | Treatment of diarrhea caused by Giardia lamblia |
| Ivermectin |  | Antiparasitic | Treatment of parasitic infections such as intestinal strongyloidiasis and onchocerciasis |
| Camostat mesylate |  | Anticancer | Treatment of chronic pancreatitis in Japan |
| Gemcitabine |  | Anticancer | Treatment of a number of types of cancer |
| Imatinib |  | Anticancer | Treatment of a number of types of leukemia |
| Tamoxifen |  | Anticancer | Treatment of breast cancer |
| Chlorpromazine |  | Antipsychotic and antihistamine | The management of Schizophrenia and other psychoses, mania and hypomania.In anxiety psychomotor agitation excitementNausea and vomiting |
| Fluphenazine |  | Antipsychotic and antihistamine | The management of Schizophrenia |
| Promethazine |  | Antipsychotic and antihistamine | Treatment of allergic rhinitis, Vasomotor rhinitis. In addition to its antihistaminic action, it provides clinically useful sedative and antiemetic effects |
| Fluvoxamine |  | Antipsychotic | Treatment of depression, anxiety and other mood disorders |
| Losartan |  | Antihypertensive | Treatment of high blood pressure |
| Metmorfin |  | Antidiabetic | Treatment of type 2 diabetes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gediz Erturk, A.; Sahin, A.; Bati Ay, E.; Pelit, E.; Bagdatli, E.; Kulu, I.; Gul, M.; Mesci, S.; Eryilmaz, S.; Oba Ilter, S.; et al. A Multidisciplinary Approach to Coronavirus Disease (COVID-19). Molecules 2021, 26, 3526. https://doi.org/10.3390/molecules26123526
Gediz Erturk A, Sahin A, Bati Ay E, Pelit E, Bagdatli E, Kulu I, Gul M, Mesci S, Eryilmaz S, Oba Ilter S, et al. A Multidisciplinary Approach to Coronavirus Disease (COVID-19). Molecules. 2021; 26(12):3526. https://doi.org/10.3390/molecules26123526
Chicago/Turabian StyleGediz Erturk, Aliye, Arzu Sahin, Ebru Bati Ay, Emel Pelit, Emine Bagdatli, Irem Kulu, Melek Gul, Seda Mesci, Serpil Eryilmaz, Sirin Oba Ilter, and et al. 2021. "A Multidisciplinary Approach to Coronavirus Disease (COVID-19)" Molecules 26, no. 12: 3526. https://doi.org/10.3390/molecules26123526