
Endophytic Fungal Community of Huperzia serrata: Diversity and Relevance to the Production of Huperzine A by the Plant Host


 , ,
, ,
Abstract
:1. Introduction
2. Results
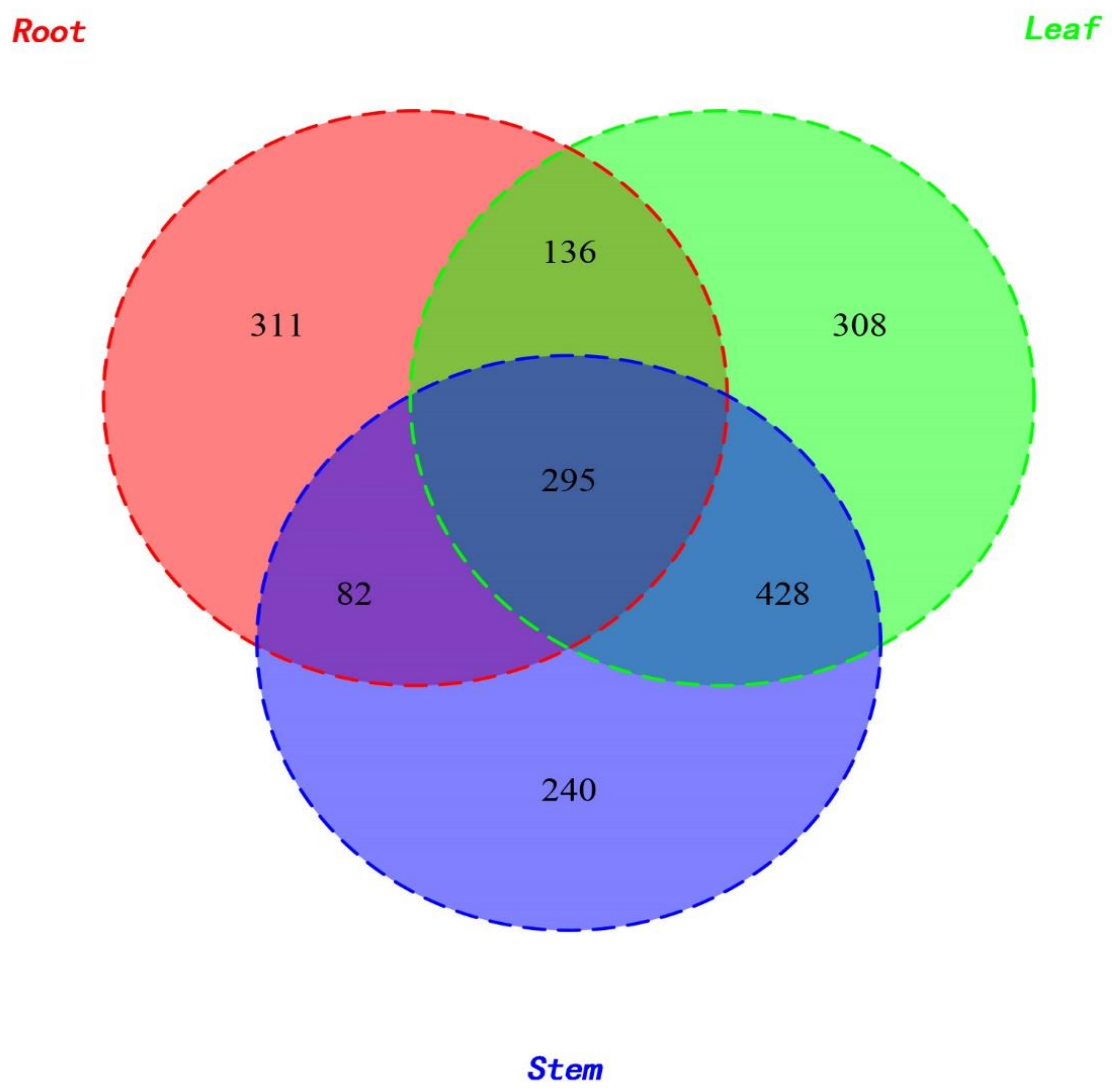
2.1. Taxonomic Distribution of Endophytic Fungi
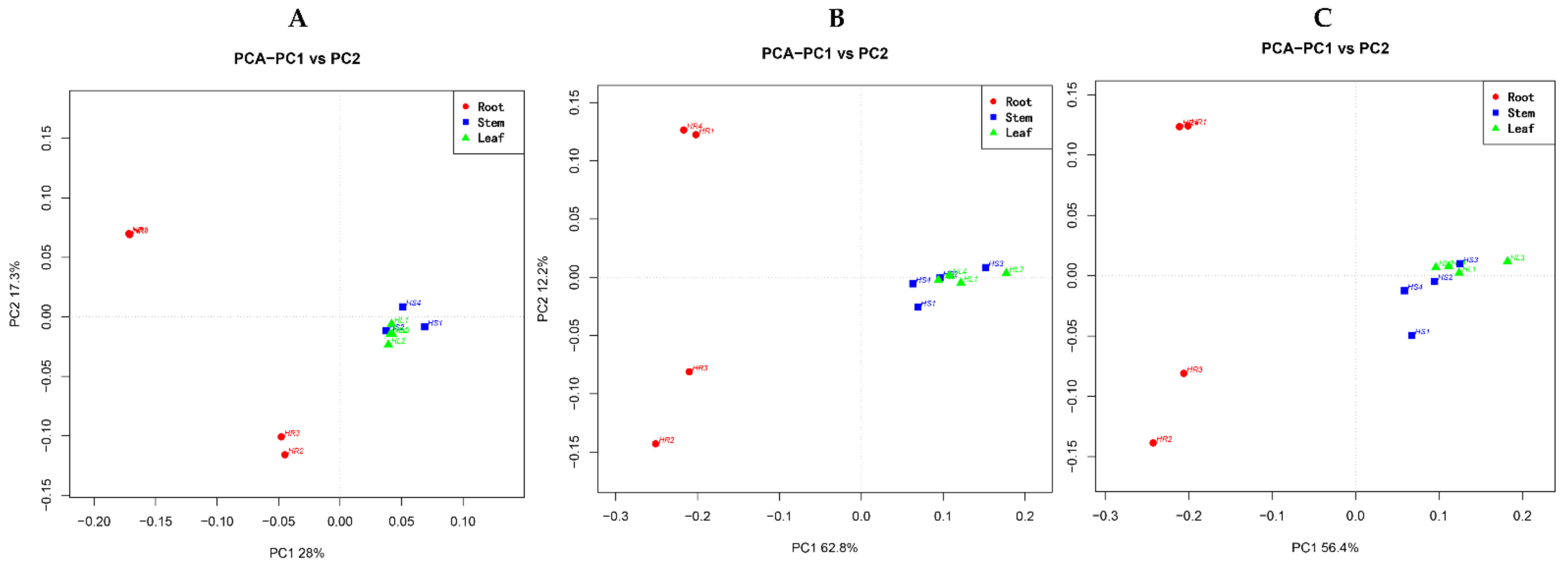
2.2. Diversity of Fungal Endophytes within Plant Organs
2.3. Correlation between Fungal Endophytes and HupA
2.4. Isolation and Identification of Endophytic Fungi
2.5. Screening of Endophytic Fungi Producing HupA
2.6. Detection of Metabolites of the HS7-1 Strain by Electrospray Ionization Mass Spectrometry (ESI-MS)
3. Discussion
4. Materials and Methods
4.1. Collection and Surface Sterilization of H. serrata Plants
4.2. Endophytic Fungal Isolation
4.3. Identification of Fungal Endophytes
4.4. Quantification of HupA in H. serrata
4.5. Endophytic Fungal Diversity Analysis
4.6. Morphological Identification Method
4.7. Fermentation of Endophytic Fungi and Extraction of Huperzine A from Huperzia serrate
4.8. Determination of Alkaloids by Electrospray Ionization Mass Spectrometry (ESI-MS)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability: Samples of the compounds are not available from the authors. |
References
- Ashani, Y.; Peggins, J.O.; Doctor, B.P. Mechanism of inhibition of cholinesterases by huperzine A. Biochem. Biophys. Res. Commun. 1992, 184, 719–726. [Google Scholar] [CrossRef]
- Wang, R.; Yan, H.; Tang, X.C. Progress in studies of huperzine A, a natural cholinesterase inhibitor from Chinese herbal medicine. Acta Pharmacol. Sin. 2006, 27, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Szypuła, W.J.; Mistrzak, P.; Olszowska, O. A new and fast method to obtain in vitro cultures of Huperzia selago (Huperziaceae) sporophytes, a club moss which is a source of huperzine A. Acta Soc. Bot. Pol. 2013, 82, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Li, Y.; Sun, C.; Wu, Q.; Song, J.; Sun, Y.; Steinmetz, A.; Chen, S. Comparison of 454-ESTs from Huperzia serrata and Phlegmariurus carinatus reveals putative genes involved in lycopodium alkaloid biosynthesis and developmental regulation. BMC Plant Biol. 2010, 10, 209. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Tan, C.; Zhu, D.; Gang, D.R. A survey of potential huperzine A natural resources in China: The Huperziaceae. J. Ethnopharmacol. 2006, 104, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; He, C. Population structure and genetic diversity of Huperzia serrata (Huperziaceae) based on amplified fragment length polymorphism (AFLP) markers. Biochem. Syst. Ecol. 2010, 38, 1137–1147. [Google Scholar] [CrossRef]
- Damar, U.; Gersner, R.; Johnstone, J.T.; Schachter, S.; Rotenberg, A. Huperzine A as a neuroprotective and antiepileptic drug: A review of preclinical research. Expert Rev. Neurother. 2016, 16, 671–680. [Google Scholar] [CrossRef]
- Zangara, A. The psychopharmacology of huperzine A: An alkaloid with cognitive enhancing and neuroprotective properties of interest in the treatment of Alzheimer’s disease. Pharmacol. Biochem. Behav. 2003, 75, 675–686. [Google Scholar] [CrossRef]
- Yang, M.; You, W.; Wu, S.; Fan, Z.; Xu, B.; Zhu, M.; Li, X.; Xiao, Y. Global transcriptome analysis of Huperzia serrata and identification of critical genes involved in the biosynthesis of huperzine A. BMC Genom. 2017, 18, 245. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.X.; Li, Z.; Chen, Y.; Yang, Y.H.; Li, G.H.; Zhao, P.J. Four new steroids from the endophytic fungus Chaetomium sp. M453 derived of Chinese herbal medicine Huperzia serrata. Fitoterapia 2017, 117, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Rodrigues, M.; Fortuna, A.; Falcão, A.; Alves, G. Huperzine A from Huperzia serrata: A review of its sources, chemistry, pharmacology and toxicology. Phytochem. Rev. 2016, 15, 51–85. [Google Scholar] [CrossRef]
- Ma, X.; Tan, C.; Zhu, D.; Gang, D.R.; Xiao, P. Huperzine A from Huperzia species—An ethnopharmacolgical review. J. Ethnopharmacol. 2007, 113, 15–34. [Google Scholar] [CrossRef]
- Gao, W.Y.; Li, Y.M.; Jiang, S.H.; Zhu, D.Y. Two new nitrone alkaloids from Huperzia serrata. Helv. Chim. Acta 2008, 91, 1031–1035. [Google Scholar] [CrossRef]
- Tan, C.H.; Zhu, D.Y. Lycopodine-type Lycopodium alkaloids from Huperzia serrata. Helv. Chim. Acta 2004, 87, 1963–1967. [Google Scholar] [CrossRef]
- Sui, X.; Gao, C. Huperzine A ameliorates damage induced by acute myocardial infarction in rats through antioxidant, anti-apoptotic and anti-inflammatory mechanisms. Int. J. Mol. Med. 2014, 33, 227–233. [Google Scholar] [CrossRef]
- Liu, J.S.; Zhu, Y.L.; Yu, C.M.; Zhou, Y.Z.; Han, Y.Y.; Wu, F.W.; Qi, B.F. The structures of huperzine A and B, two new alkaloids exhibiting marked anticholinesterase activity. Can. J. Chem. 1986, 64, 837–839. [Google Scholar] [CrossRef]
- Mishima, K.; Egashira, N.; Hirosawa, N.; Fujii, M.; Matsumoto, Y.; Iwasaki, K.; Fujiwara, M. Characteristics of learning and memory impairment induced by Δ9-tetrahydrocannabinol in rats. Jpn. J. Pharmacol. 2001, 87, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.-C.; De Sarno, P.; Sugaya, K.; Giacobini, E. Effect of huperzine A, a new cholinesterase inhibitor, on the central cholinergic system of the rat. J. Neurosci. Res. 1989, 24, 276–285. [Google Scholar] [CrossRef]
- Qian, Z.M.; Ke, Y. Huperzine A: Is it an effective disease-modifying drug for Alzheimer’s disease? Front. Aging Neurosci. 2014, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Xu, Y.; Chen, J.; Liu, Q.; Gu, J.; Wang, X.; Ma, J.; Li, H.; Onuchic, J.N.; Jiang, H. Free energy landscape for the binding process of Huperzine A to acetylcholinesterase. Proc. Natl. Acad. Sci. USA 2013, 110, 4273–4278. [Google Scholar] [CrossRef] [Green Version]
- Modrego, P.J. Depression in Alzheimer’s disease. Pathophysiology, diagnosis, and treatment. J. Alzheimers Dis. 2010, 21, 1077–1087. [Google Scholar] [CrossRef]
- Dong, L.; May, B.H.; Feng, M.; Hyde, A.J.; Tan, H.Y.; Guo, X.; Zhang, A.L.; Lu, C.; Xue, C.C. Chinese Herbal Medicine for Mild Cognitive Impairment: A Systematic Review and Meta-Analysis of Cognitive Outcomes. Phyther. Res. 2016, 30, 1592–1604. [Google Scholar] [CrossRef]
- Yang, G.; Wang, Y.; Tian, J.; Liu, J.P. Huperzine A for Alzheimer’s Disease: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wu, H.M.; Zhou, R.L.; Liu, G.J.; Dong, B.R. Huperzine A for Alzheimer’s disease. Cochrane Database Syst. Rev. 2008, CD005592. [Google Scholar] [CrossRef]
- Fu, L.M.; Li, J.T. A systematic review of single Chinese herbs for Alzheimer’s disease treatment. Evid. Based Complement. Altern. Med. 2011, 2011, 640284. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Hamayun, M.; Kang, S.M.; Kim, Y.H.; Jung, H.Y.; Lee, J.H.; Lee, I.J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, Z.A.; Mirza, D.N.; Arora, P.; Riyaz-Ul-Hassan, S. Molecular phylogeny, diversity, community structure, and plant growth promoting properties of fungal endophytes associated with the corms of saffron plant: An insight into the microbiome of Crocus sativus Linn. Fungal Biol. 2016, 120, 1509–1524. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic fungi: A source of potential antifungal compounds. J. Fungi 2018, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.-H.; Fan, S.-W.; Ling, Q.-Z.; Huang, B.-B.; Wei, Z.-J. Taxomyces andreanae, a proposed new taxon for a bulbilliferous hyphomycete associated with Pacific yew (Taxus brevifolia). World J. Microbiol. Biotechnol. 2014, 47, 71–80. [Google Scholar] [CrossRef]
- Su, J.; Yang, M. Huperzine A production by Paecilomyces tenuis YS-13, an endophytic fungus isolated from Huperzia serrata. Nat. Prod. Res. 2015, 29, 1035–1041. [Google Scholar] [CrossRef]
- Shu, S.; Zhao, X.; Wang, W.; Zhang, G.; Cosoveanu, A.; Ahn, Y.; Wang, M. Identification of a novel endophytic fungus from Huperzia serrata which produces huperzine A. World J. Microbiol. Biotechnol. 2014, 30, 3101–3109. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.H.; Fan, S.W.; Ling, Q.Z.; Huang, B.B.; Wei, Z.J. Indentification of huperzine A-producing endophytic fungi isolated from Huperzia serrata. World J. Microbiol. Biotechnol. 2014, 30, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, Q.G.; Zhang, Z.B.; Yan, R.M.; Wang, L.Y.; Zhu, D. Isolation and characterization of endophytic huperzine A-producing fungi from Huperzia serrata. J. Ind. Microbiol. Biotechnol. 2011, 38, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Zeng, Q.G.; Yan, R.M.; Wang, Y.; Zou, Z.R.; Zhu, D. Endophytic fungus Cladosporium cladosporioides LF70 from Huperzia serrata produces Huperzine A. World J. Microbiol. Biotechnol. 2011, 27, 479–486. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, J.; Zeng, Q.; Zhang, Z.; Yan, R. A novel endophytic Huperzine A-producing fungus, Shiraia sp. Slf14, isolated from Huperzia serrata. J. Appl. Microbiol. 2010, 109, 1469–1478. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles: Tansley review. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Saikkonen, K.; Faeth, S.H.; Helander, M.; Sullivan, T.J. Fungal endophytes: A continuum of interactions with host plants. Annu. Rev. Ecol. Syst. 1998, 29, 319–343. [Google Scholar] [CrossRef]
- Kogel, K.H.; Franken, P.; Hückelhoven, R. Endophyte or parasite-what decides? Curr. Opin. Plant Biol. 2006, 9, 358–363. [Google Scholar] [CrossRef]
- Unterseher, M.; Schnittler, M. Dilution-to-extinction cultivation of leaf-inhabiting endophytic fungi in beech (Fagus sylvatica L.)—Different cultivation techniques influence fungal biodiversity assessment. Mycol. Res. 2009, 113, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Moricca, S.; Ragazzi, A. Fungal endophytes in mediterranean oak forests: A lesson from Discula quercina. Phytopathology 2008, 98, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef]
- Tudzynski, B.; Sharon, A. Biosynthesis, biological role and application of fungal phytohormones. In The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Osiewacz, H.D., Ed.; Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2002; Volume 10, pp. 183–211. [Google Scholar]
- Deckert, R.J.; Melville, L.H.; Peterson, R.L. Structural features of a Lophodermium endophyte during the cryptic life-cycle phase in the foliage of Pinus strobus. Mycol. Res. 2001, 105, 991–997. [Google Scholar] [CrossRef]
- Menéndez, A.; Bertoni, M.D.; Cabrai, D. Fungal endophytes in Juncus imbricatus var chamissonis: Identification of colonization patterns. Rev. Iberoam. Micol. 1997, 14, 125–128. [Google Scholar]
- Ngwene, B.; Boukail, S.; Söllner, L.; Franken, P.; Andrade-Linares, D.R. Phosphate utilization by the fungal root endophyte Piriformospora indica. Plant. Soil 2016, 405, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Bialer, M.; Johannessen, S.I.; Kupferberg, H.J.; Levy, R.H.; Perucca, E.; Tomson, T. Progress report on new antiepileptic drugs: A summary of the Eighth Eilat Conference (EILAT VIII). Epilepsy Res. 2007, 73, 1–52. [Google Scholar] [CrossRef]
- Bialer, M.; Johannessen, S.I.; Levy, R.H.; Perucca, E.; Tomson, T.; White, H.S. Progress report on new antiepileptic drugs: A summary of the Tenth Eilat Conference (EILAT X). Epilepsy Res. 2010, 92, 89–124. [Google Scholar] [CrossRef]
- Bai, D. Development of huperzine A and B for treatment of Alzheimer’s disease. Pure Appl. Chem. 2007, 79, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Ha, G.T.; Wong, R.K.; Zhang, Y. Huperzine A as potential treatment of Alzheimer’s disease: An assessment on chemistry, pharmacology, and clinical studies. Chem. Biodivers. 2011, 8, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhou, Z.; Liu, Y.; Hu, X.; Guo, Y.; Li, J. Application of high-throughput internal transcribed spacer rRNA metagenomics analysis in deciphering endophytic fungi diversity of dendrobium officinale. J. Biobased Mater. Bioenergy 2017, 11, 106–118. [Google Scholar] [CrossRef]
- Pohanka, M.; Zemek, F.; Bandouchova, H.; Pikula, J. Toxicological scoring of Alzheimer’s disease drug huperzine in a guinea pig model. Toxicol. Mech. Methods 2012, 22, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Gasong, B.T.; Tjandrawinata, R.R. Production of secondary metabolite E2.2 from Phaleria macrocarpa endophytic fungus. Asian Pac. J. Trop. Biomed. 2016, 6, 881–885. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D. Endophyte: The Evolution of a Term, and Clarification of Its Use and Definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Petrini, O. Fungal endophytes of tree leaves. In Microbial Ecology of Leaves; Andrews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Gloer, J.B. Applications of fungal ecology in the search for new bioactive natural products. In Environmental and Microbial Relationships; Springer: New York, NY, USA, 2007; pp. 257–283. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 16, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Lodge, D.J.; Fisher, P.J.; Sutton, B.C. Endophytic fungi of Manilkara bidentata leaves in Puerto Rico. Mycologia 1996, 88, 733–738. [Google Scholar] [CrossRef]
- Wilson, D.; Carrol, G.C. Infection studies of Discula quercina, an endophyte of Quercus garryana. Mycologia 1994, 86, 635–647. [Google Scholar] [CrossRef]
- Fisher, P.J.; Petrini, O.; Petrini, L.E.; Sutton, B.C. Fungal endophytes from the leaves and twigs of Quercus ilex L. from England, Majorca and Switzerland. New Phytol. 1994, 127, 133–137. [Google Scholar] [CrossRef]
- Verma, V.C.; Gond, S.K.; Kumar, A.; Kharwar, R.N.; Strobel, G. The endophytic mycoflora of bark, leaf, and stem tissues of Azadirachta indica A. Juss (Neem) from Varanasi (India). Microb. Ecol. 2007, 54, 119–125. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich, J.; Hyde, K.D.; Petrini, O. Endophytic fungi associated with palms. Mycol. Res. 2000, 104, 1202–1212. [Google Scholar] [CrossRef]
- Raviraja, N.S. Fungal endophytes in five medicinal plant species from Kudremukh Range, Western Ghats of India. J. Basic Microbiol. 2005, 45, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Fisher, P.J.; Graf, F.; Petrini, L.E.; Sutton, B.C.; Wookey, P.A. Fungal Endophytes of Dryas octopetala from a High Arctic Polar Semidesert and from the Swiss Alps. Mycologia 1995, 87, 319–323. [Google Scholar] [CrossRef]
- Fernandes, E.G.; Pereira, O.L.; da Silva, C.C.; Bento, C.B.P.; de Queiroz, M.V. Diversity of endophytic fungi in Glycine max. Microbiol. Res. 2015, 181, 84–92. [Google Scholar] [CrossRef]
- Carroll, G. Fungal endophytes in stems and leaves: From latent pathogen to mutualistic symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Boyle, C.; Götz, M.; Dammann-Tugend, U.; Schulz, B. Endophyte-host interactions III. Local vs. Systemic colonization. Symbiosis 2001, 31, 259–281. [Google Scholar]
- Sun, H.; Wei, Y.; Fang, X.; Yu, L.N.; Zhang, Y. Diversity of endophytic bacteria isolated from Huperzia serrata. Wei Sheng Wu Xue Bao 2016, 56, 614–628. [Google Scholar]
- Zhou, W.; Verma, V.C.; Wheeler, T.A.; Woodward, J.E.; Starr, J.L.; Sword, G.A. Tapping into the cotton fungal phytobiome for novel nematode biological control tools. Phytobiomes J. 2020, 4, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Miao, L.; Li, H.; Lin, A.; Song, F.; Zhang, P. Illumina-based analysis yields new insights into the diversity and composition of endophytic fungi in cultivated Huperzia serrata. PLoS ONE 2020, 15, e0242258. [Google Scholar] [CrossRef]
- Peng, W.; Guo, L.; Zheng, C.J.; Zhang, Q.Y.; Jia, M.; Jiang, Y.P.; Han, T.; Qin, L.P. Two new azaphilone alkaloids dimers from endophyte Chaetomium fusiforme of the liverwort Scapania verrucosa Heeg. Biochem. Syst. Ecol. 2012, 45, 124–126. [Google Scholar] [CrossRef]
- Sang, X.; Yang, M.; Su, J. Research on endophytic fungi for producing huperzine A on a large-scale. Crit. Rev. Microbiol. 2020, 46, 654–664. [Google Scholar] [CrossRef]
- Ohba, T.; Yoshino, Y.; Ishisaka, M.; Abe, N.; Tsuruma, K.; Shimazawa, M.; Oyama, M.; Tabira, T.; Hara, H. Japanese Huperzia serrata extract and the constituent, huperzine A, ameliorate the scopolamine-induced cognitive impairment in mice. Biosci. Biotechnol. Biochem. 2015, 79, 1838–1844. [Google Scholar] [CrossRef]
- Ma, X.; Tan, C.; Zhu, D.; Gang, D.R. Is there a better source of huperzine A than Huperzia serrata? Huperzine A content of huperziaceae species in China. J. Agric. Food Chem. 2005, 53, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Wang, J.; Pan, S.L. Isolation and preliminary identification of the endophytic fungi which produce Hupzine A from four species in Hupziaceae and determination of Huperzine A by HPLC. Fudan Univ. J. Med. Sci. 2009, 36, 445–449. [Google Scholar]
- Zhang, D.; Yang, Y.; Castlebury, L.A.; Cerniglia, C.E. A method for the large scale isolation of high transformation efficiency fungal genomic DNA. FEMS Microbiol. Lett. 1996, 145, 261–265. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Retention Time (Min) | Concentration (μg/g) | Area (mAu·sec) |
|---|---|---|---|
| Control | 10.247 | 4.4 | 459.66324 |
| HS7-1 | 10.255 | 3.7 | 388.57199 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; Noushahi, H.A.; Zhang, Y.; Liu, J.; Cosoveanu, A.; Liu, Y.; Yan, L.; Zhang, J.; Shu, S. Endophytic Fungal Community of Huperzia serrata: Diversity and Relevance to the Production of Huperzine A by the Plant Host. Molecules 2021, 26, 892. https://doi.org/10.3390/molecules26040892
Cui L, Noushahi HA, Zhang Y, Liu J, Cosoveanu A, Liu Y, Yan L, Zhang J, Shu S. Endophytic Fungal Community of Huperzia serrata: Diversity and Relevance to the Production of Huperzine A by the Plant Host. Molecules. 2021; 26(4):892. https://doi.org/10.3390/molecules26040892
Chicago/Turabian StyleCui, Lingli, Hamza Armghan Noushahi, Yipeng Zhang, Jinxin Liu, Andreea Cosoveanu, Ying Liu, Ling Yan, Jing Zhang, and Shaohua Shu. 2021. "Endophytic Fungal Community of Huperzia serrata: Diversity and Relevance to the Production of Huperzine A by the Plant Host" Molecules 26, no. 4: 892. https://doi.org/10.3390/molecules26040892