
1-Iodohexadecane Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in Mice: Possible Involvements of the Skin Barrier and Mast Cell SNARE Proteins


Abstract
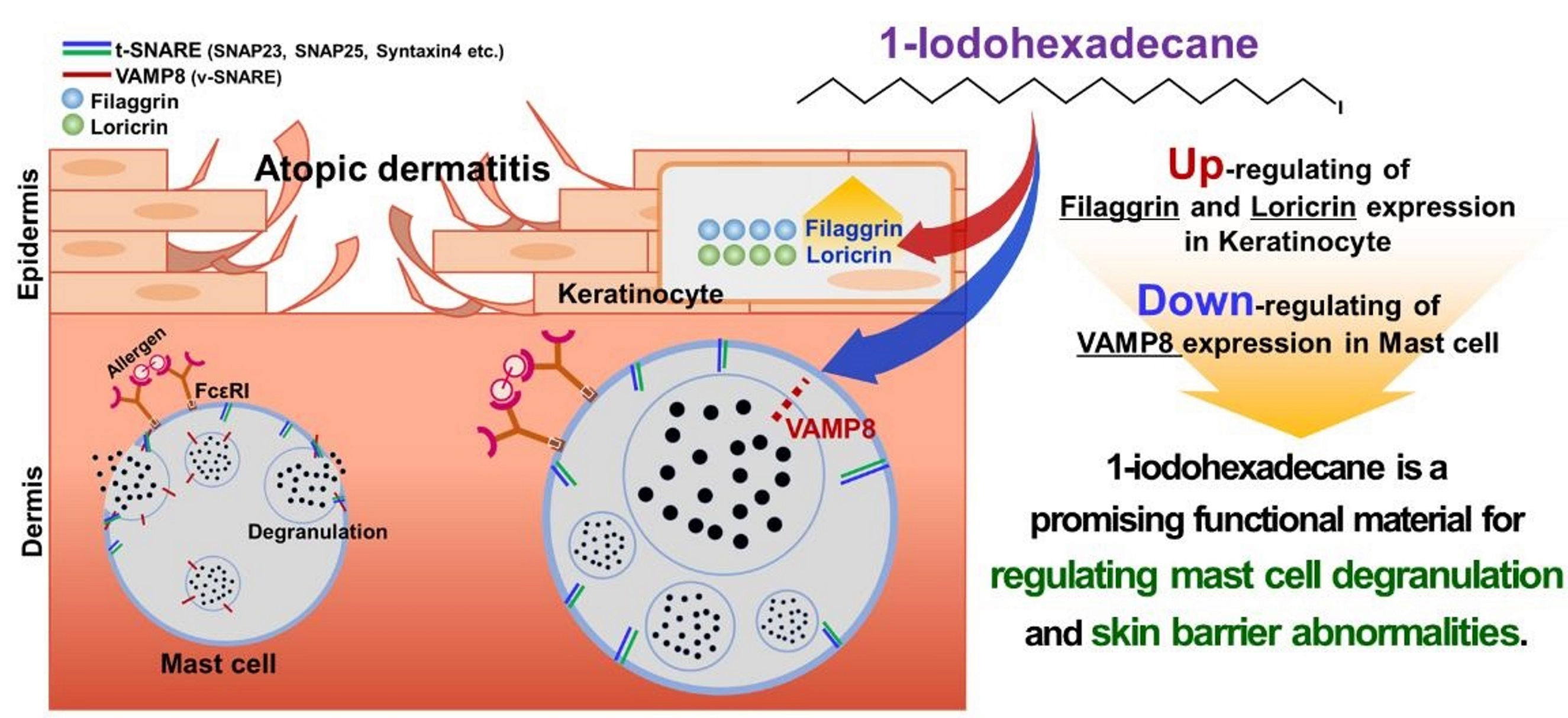
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Effects of 1-Iodohexadecane on the Expressions of SNARE Proteins in Mast Cells
2.2. Effects of 1-Iodohexadecane on Histamine and β-Hexosaminidase Secretion by Mast Cells
2.3. Effects of 1-Iodohexadecane on Skin Barrier Protein Expressions in Human Keratinocytes
2.4. Effect of 1-Iodohexadecane on Skin Lesions in the DNCB-Induced Murine Model
2.5. Histopathological Alterations and Serum Levels of Mast Cell Degranulation Markers in the DNCB-Induced Murine Model Exposed to 1-Iodohexadecane
2.6. Effect of 1-Iodohexadecane on Skin Barrier Proteins in the DNCB-Induced Murine Model
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Viability
4.4. Immunoblotting
4.5. β-Hexosaminidase and Histamine Release Assays
4.6. DNCB-Induced Atopic Dermatitis Animal Model
4.7. Evaluation of Dermatitis Severity
4.8. Histopathological Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Langan, S.M.; Irvine, A.D.; Weidinger, S. Atopic dermatitis. Lancet 2020, 396, 345–360. [Google Scholar] [CrossRef]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.Y.; Lee, H.S.; Lee, J.Y. Skin barrier abnormalities and immune dysfunction in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdis, C.A.; Akdis, M.; Bieber, T.; Bindslev-Jensen, C.; Boguniewicz, M.; Eigenmann, P.; Hamid, Q.; Kapp, A.; Leung, D.Y.; Lipozencic, J.; et al. Diagnosis and treatment of atopic dermatitis in children and adults: European Academy of Allergology and Clinical Immunology/American Academy of Allergy, Asthma and Immunology/PRACTALL Consensus Report. J. Allergy Clin. Immunol. 2006, 118, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Guttman-Yassky, E.; Waldman, A.; Ahluwalia, J.; Ong, P.Y.; Eichenfield, L.F. Atopic dermatitis: Pathogenesis. Semin. Cutan. Med. Surg. 2017, 36, 100–103. [Google Scholar] [CrossRef]
- Sroka-Tomaszewska, J.; Trzeciak, M. Molecular mechanisms of atopic dermatitis pathogenesis. Int. J. Mol. Sci. 2021, 22, 4130. [Google Scholar] [CrossRef]
- Modena, B.D.; Dazy, K.; White, A.A. Emerging concepts: Mast cell involvement in allergic diseases. Transl. Res. 2016, 174, 98–121. [Google Scholar] [CrossRef]
- Kim, H.S.; Kawakami, Y.; Kasakura, K.; Kawakami, T. Recent advances in mast cell activation and regulation. F1000Research 2020, 9, 196. [Google Scholar] [CrossRef] [Green Version]
- Sim, A.T.; Ludowyke, R.I.; Verrills, N.M. Mast cell function: Regulation of degranulation by serine/threonine phosphatases. Pharmacol. Ther. 2006, 112, 425–439. [Google Scholar] [CrossRef]
- Woska, J.R., Jr.; Gillespie, M.E. Small-interfering RNA-mediated identification and regulation of the ternary SNARE complex mediating RBL-2H3 mast cell degranulation. Scand. J. Immunol. 2011, 73, 8–17. [Google Scholar] [CrossRef]
- Puri, N.; Kruhlak, M.J.; Whiteheart, S.W.; Roche, P.A. Mast cell degranulation requires N-ethylmaleimide-sensitive factor-mediated SNARE disassembly. J. Immunol. 2003, 171, 5345–5352. [Google Scholar] [CrossRef] [Green Version]
- Malmersjö, S.; Di Palma, S.; Diao, J.; Lai, Y.; Pfuetzner, R.A.; Wang, A.L.; McMahon, M.A.; Hayer, A.; Porteus, M.; Bodenmiller, B.; et al. Phosphorylation of residues inside the SNARE complex suppresses secretory vesicle fusion. EMBO J. 2016, 35, 1810–1821. [Google Scholar] [CrossRef]
- Woska, J.R., Jr.; Gillespie, M.E. SNARE complex-mediated degranulation in mast cells. J. Cell. Mol. Med. 2012, 16, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kong, B.; Jung, Y.; Park, J.B.; Oh, J.M.; Hwang, J.; Cho, J.Y.; Kweon, D.H. Soluble N-ethylmaleimide-sensitive factor attachment protein receptor-derived peptides for the regulation of mast cell degranulation. Front. Immunol. 2018, 9, 725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proksch, E.; Brandner, J.M.; Jensen, J.M. The skin: An indispensable barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Woodfolk, J.A. Skin barrier defects in atopic dermatitis. Curr. Allergy Asthma Rep. 2014, 14, 433. [Google Scholar] [CrossRef]
- Ahn, K.; Kim, B.E.; Kim, J.; Leung, D.Y. Recent advances in atopic dermatitis. Curr. Opin. Immunol. 2020, 66, 14–21. [Google Scholar] [CrossRef]
- Furue, M. Regulation of filaggrin, loricrin, and involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic implications in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–430. [Google Scholar] [CrossRef]
- Kalinin, A.; Marekov, L.N.; Steinert, P.M. Assembly of the epidermal cornified cell envelope. J. Cell Sci. 2001, 114, 3069–3070. [Google Scholar] [CrossRef]
- Gutowska-Owsiak, D.; Ogg, G.S. Cytokine regulation of the epidermal barrier. Clin. Exp. Allergy 2013, 43, 586–598. [Google Scholar] [CrossRef] [PubMed]
- Hänel, K.H.; Cornelissen, C.; Lüscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.E.; Leung, D.Y. Epidermal barrier in atopic dermatitis. Allergy Asthma Immunol. Res. 2012, 4, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieber, T. Atopic dermatitis: An expanding therapeutic pipeline for a complex disease. Nat. Rev. Drug Discov. 2022, 21, 21–40. [Google Scholar] [CrossRef]
- Yang, G.; Lee, K.; An, D.G.; Lee, M.H.; Ham, I.H.; Choi, H.Y. Effect of Chrysanthemi borealis flos on atopic dermatitis induced by 1-chloro 2,4-dinitrobenzene in NC/Nga mouse. Immunopharmacol. Immunotoxicol. 2012, 34, 413–418. [Google Scholar] [CrossRef]
- Yang, W.S.; Lee, S.R.; Jeong, Y.J.; Park, D.W.; Cho, Y.M.; Joo, H.M.; Kim, I.; Seu, Y.B.; Sohn, E.H.; Kang, S.C. Antiallergic activity of ethanol extracts of Arctium lappa L. undried roots and its active compound, oleamide, in regulating FcεRI-mediated and MAPK signaling in RBL-2H3 cells. J. Agric. Food Chem. 2016, 64, 3564–3573. [Google Scholar] [CrossRef]
- Cho, K.; Parveen, A.; Kang, M.C.; Subedi, L.; Lee, J.H.; Park, S.Y.; Jin, M.R.; Yoon, H.; Son, Y.K.; Kim, S.Y. Pyrus ussuriensis Maxim. leaves extract ameliorates DNCB-induced atopic dermatitis-like symptoms in NC/Nga mice. Phytomedicine 2018, 48, 76–83. [Google Scholar] [CrossRef]
- Kim, D.Y.; Won, K.-J.; Hwang, D.I.; Lee, S.Y.; Choi, I.-H.; Kim, B.; Lee, H.M. Essential oil from Chrysanthemum boreale flowers modulates SNARE protein-linked mast cell response and skin barrier proteins and ameliorates atopic dermatitis-like lesions in mice. Hortic. Environ. Biotechnol. 2022. [Google Scholar] [CrossRef]
- Olubunmi, A.; Gabriel, O.A.; Stephen, A.O.; Scott, F.O. Antioxidant and antimicrobial activity of cuticular wax from Kigelia Africana. FABAD J. Pharm. Sci. 2009, 34, 187–194. [Google Scholar]
- Nadayag, J.; Dapar, M.L.G.; Aranas, A.T.; Mindo, R.A.R.; Cabrido, C.K.; Manting, M.M.E.; Torres, M.A.J.; Demayo, C.G. Qualitative assessment of the antimicrobial, antioxidant, and phytochemical properties of the ethanolic extracts of the inner bark of Atuna racemose. Pharmacophore 2019, 10, 52–59. [Google Scholar]
- Teoh, L.; Gnanasegaran, N.; Adnan, A.F.M.; Taha, R.M. The comparative antimicrobial and anticancer of chemical extract from in vitro and in vivo Peperomia pellucida plantlet. J. Appl. Biol. Biotechnol. 2021, 9, 115–123. [Google Scholar]
- Wu, Z.; Thiyagarajan, S.; O’Shaughnessy, B.; Karatekin, E. Regulation of Exocytotic Fusion Pores by SNARE Protein Transmembrane Domains. Front. Mol. Neurosci. 2017, 10, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, A.J.; Haig, D.M.; Bazin, H.; McGuigan, A.C.; Moqbel, R.; Miller, H.R. IgE-mediated release of rat mast cell protease II, beta-hexosaminidase and leukotriene C4 from cultured bone marrow-derived rat mast cells. Immunology 1989, 67, 414–418. [Google Scholar] [PubMed]
- Andrew, D.; Craig, A.D. Spinothalamic lamina I neurons selectively sensitive to histamine: A central neural pathway for itch. Nat. Neurosci. 2001, 4, 72–77. [Google Scholar] [CrossRef]
- Kim, B.E.; Howell, M.D.; Guttman, E.; Gilleaudeau, P.M.; Cardinale, I.R.; Boguniewicz, M.; Krueger, J.G.; Leung, D.Y. TNF-α Downregulates Filaggrin and Loricrin through c-Jun N-terminal Kinase: Role for TNF-α Antagonists to Improve Skin Barrier. J. Investig. Dermatol. 2011, 131, 1272–1279. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Suárez-Fariñas, M.; Chiricozzi, A.; Nograles, K.E.; Shemer, A.; Fuentes-Duculan, J.; Cardinale, I.; Lin, P.; Bergman, R.; Bowcock, A.; et al. Broad defects in epidermal cornification in atopic dermatitis identified through genomic analysis. J. Allergy Clin. Immunol. 2009, 124, 1235–1244.e58. [Google Scholar] [CrossRef]
- Steinhoff, M.; Steinhoff, A.; Homey, B.; Luger, T.A.; Schneider, S.W. Role of vasculature in atopic dermatitis. J. Allergy Clin. Immunol. 2006, 118, 190–197. [Google Scholar] [CrossRef]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Nutten, S. Atopic Dermatitis: Global Epidemiology and Risk Factors. Ann. Nutr. Metab. 2015, 66, 8–16. [Google Scholar] [CrossRef]
- Avena-Woods, C. Overview of atopic dermatitis. Am. J. Manag. Care 2017, 23, S115–S123. [Google Scholar]
- Udkoff, J.; Waldman, A.; Ahluwalia, J.; Borok, J.; Eichenfield, L.F. Current and emerging topical therapies for atopic dermatitis. Clin. Dermatol. 2017, 35, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Shiohara, T.; Hayakawa, J.; Mizukawa, Y. Animal models for atopic dermatitis: Are they relevant to human disease? J. Dermatol. Sci. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Fan, H.J.; Xie, Z.P.; Lu, Z.W.; Tan, Z.B.; Bi, Y.M.; Xie, L.P.; Wu, Y.T.; Zhang, W.T.; Liu-Kot, K.; Liu, B.; et al. Anti-inflammatory and immune response regulation of Si-Ni-San in 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin dysfunction. J. Ethnopharmacol. 2018, 222, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Castle, J.D.; Guo, Z.; Liu, L. Function of the t-SNARE SNAP-23 and secretory carrier membrane proteins (SCAMPs) in exocytosis in mast cells. Mol. Immunol. 2002, 38, 1337–1340. [Google Scholar] [CrossRef]
- Tiwari, N.; Wang, C.C.; Brochetta, C.; Ke, G.; Vita, F.; Qi, Z.; Rivera, J.; Soranzo, M.R.; Zabucchi, G.; Hong, W.; et al. VAMP-8 segregates mast cell-preformed mediator exocytosis from cytokine trafficking pathways. Blood 2008, 111, 3665–3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, N.; Roche, P.A. Mast cells possess distinct secretory granule subsets whose exocytosis is regulated by different SNARE isoforms. Proc. Natl. Acad. Sci. USA 2008, 105, 2580–2585. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Furuta, K. Roles of IgE and histamine in mast cell maturation. Cells 2021, 10, 2170. [Google Scholar] [CrossRef]
- Nakamura, T.; Yoshida, N.; Yamanoi, Y.; Honryo, A.; Tomita, H.; Kuwabara, H.; Kojima, Y. Eucalyptus oil reduces allergic reactions and suppresses mast cell degranulation by downregulating IgE-FcεRI signalling. Sci. Rep. 2020, 10, 20940. [Google Scholar] [CrossRef]
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef]
- Dębińska, A. New treatments for atopic dermatitis targeting skin barrier repair via the regulation of FLG expression. J. Clin. Med. 2021, 10, 2506. [Google Scholar] [CrossRef]
- Noh, M.; Yeo, H.; Ko, J.; Kim, H.K.; Lee, C.H. MAP17 is associated with the T-helper cell cytokine-induced down-regulation of filaggrin transcription in human keratinocytes. Exp. Dermatol. 2010, 19, 355–362. [Google Scholar] [CrossRef]
- Otsuka, A.; Doi, H.; Egawa, G.; Maekawa, A.; Fujita, T.; Nakamizo, S.; Nakashima, C.; Nakajima, S.; Watanabe, T.; Miyachi, Y.; et al. Possible new therapeutic strategy to regulate atopic dermatitis through upregulating filaggrin expression. J. Allergy Clin. Immunol. 2014, 133, 139–146.e10. [Google Scholar] [CrossRef] [PubMed]
- Gschwandtner, M.; Mildner, M.; Mlitz, V.; Gruber, F.; Eckhart, L.; Werfel, T.; Gutzmer, R.; Elias, P.M.; Tschachler, E. Histamine suppresses epidermal keratinocyte differentiation and impairs skin barrier function in a human skin model. Allergy 2013, 68, 37–47. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.Y.; Won, K.-J.; Hwang, D.I.; Kim, N.Y.; Kim, B.; Lee, H.M. 1-Iodohexadecane Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in Mice: Possible Involvements of the Skin Barrier and Mast Cell SNARE Proteins. Molecules 2022, 27, 1560. https://doi.org/10.3390/molecules27051560
Kim DY, Won K-J, Hwang DI, Kim NY, Kim B, Lee HM. 1-Iodohexadecane Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in Mice: Possible Involvements of the Skin Barrier and Mast Cell SNARE Proteins. Molecules. 2022; 27(5):1560. https://doi.org/10.3390/molecules27051560
Chicago/Turabian StyleKim, Do Yoon, Kyung-Jong Won, Dae Il Hwang, Nan Young Kim, Bokyung Kim, and Hwan Myung Lee. 2022. "1-Iodohexadecane Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in Mice: Possible Involvements of the Skin Barrier and Mast Cell SNARE Proteins" Molecules 27, no. 5: 1560. https://doi.org/10.3390/molecules27051560