

The Potential of Magnolia spp. in the Production of Alternative Pest Control Substances


Department of Biology, Division of Natural and Exact Sciences, University of Guanajuato, Guanajuato 36050, Mexico
*
Author to whom correspondence should be addressed.
Molecules 2023, 28(12), 4681; https://doi.org/10.3390/molecules28124681
Submission received: 19 May 2023
/
Revised: 7 June 2023
/
Accepted: 8 June 2023
/
Published: 9 June 2023
(This article belongs to the Special Issue Advances in Natural Products and Their Biological Activities)
Abstract
:The irrational use of synthetic pesticides in agriculture has had negative impacts on ecosystems and contributed to environmental pollution. Botanical pesticides offer a clean biotechnological alternative to meet the agricultural challenges posed by pests and arthropods. This article proposes the use of fruit structures (fruit, peel, seed, and sarcotesta) of several Magnolia species as biopesticides. The potential of extracts, essential oils, and secondary metabolites of these structures for pest control is described. From 11 Magnolia species, 277 natural compounds were obtained, 68.7% of which were terpenoids, phenolic compounds, and alkaloids. Finally, the importance of a correct management of Magnolia species to ensure their sustainable use and conservation is stressed.
Keywords:
magnoliaceae; bioprospecting; conservation; botanical pesticides; pollyfollicle; seed; sarcotesta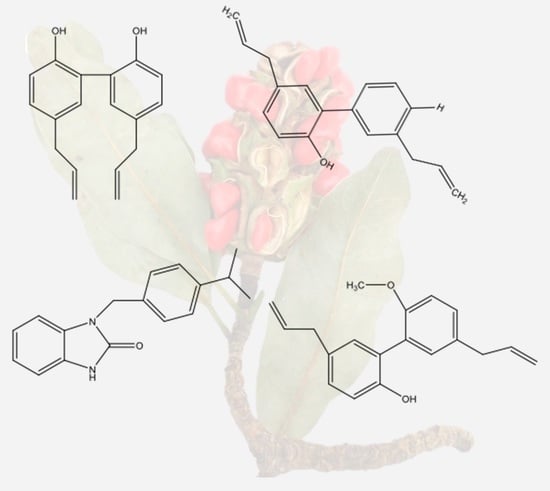
1. Introduction
One of the main problems faced in the production of fruits and vegetables around the world is pest control [1]. The Food and Agriculture Organization of the United Nations (FAO) reports that 40% of total world agricultural production is lost to pests [2], mostly of the Hexapoda class (insects) and the orders, Coleoptera, Diptera, Hemiptera, Homoptera, Hymenoptera, Lepidoptera, Orthoptera, and Thysanoptera [3,4].
Among the many crop protection methods known today, chemical control (pesticides) remains the most widely used one [5]. Applying synthetic pesticides to crops is an effective way to reduce the production losses. However, their toxicity poses serious risks. Pesticides are chemical compounds or combinations of them used to repel, destroy, and control pests [6]. Generally, they are characterized as highly effective, wide-spectrum chemical substances, but the majority of them are also highly toxic and contaminates the ecosystem. According to the target organism, pesticides are classified as insecticides, molluscicides, acaricides, fungicides, bactericides, or others [7]. In the case of insecticides, the most environmentally harmful groups are organochlorines, organophosphates, carbamates, and pyrethroids [8].
The Integral Pest Control promoted by the FAO consists of monitoring, regulating, and controlling pests via sound methods compatible with the natural environment, while reducing the use of toxic pesticides that affect the life of non-target organisms, the environment, and natural resources [9]. An alternative to synthetic pesticides is the use of pesticides of botanical origin, whose action may be comparatively slower but are safer and environmentally friendly [8,9,10]. They are also known as natural pesticides and biopesticides and may come in the form of botanical extracts, essential oils, and natural compounds [11,12].
Botanical pesticides offer several advantages over synthetic pesticides. In fact, they are as effective or even superior to their counterparts [13]. They pose minimal risks to other organisms, such as mammals (including humans), birds, reptiles, and plants. As they are of natural origin, their degree of persistence and accumulation in the environment are very low. They are photosensitive biodegradable molecules that are easily decomposed by solar rays and the action of microorganisms [14]. For example, it has been demonstrated that azadirachtin remains in the soil and cultivars for 24 to 48 h [15], while pyrethrin persists for three to five hours after application, which reduces its potential impact on natural resources (water, soil, and air) and constitutes a beneficial attribute for environmental conservation [14]. Another important characteristic of botanical insecticides is their multispecific action, which makes it more difficult for pests to develop resistance to the compound, as opposed to chemical pesticides, which regularly target a specific molecule [16]. For example, flavanones were evaluated in terms of Mycobacterium tuberculosis viability to act against protein kinase G (PknG) as a new promising drug target [17], and fukugetin, a natural flavone as an inhibitor of human tissue kallikreins [18].
In this article, the biopesticide potential of botanical extracts and essential oils obtained from the fruit (seeded and whole), seeds, and sarcotesta of several Magnolia species is analyzed in detail. To understand their effectiveness in pest control, the biological activity of their secondary metabolites is described. The bibliographic review conducted here confirms the benefits of using Magnolia species as natural biopesticides in agroecosystems and stresses the need to promote their conservation and further the study of this taxonomic group from an ecological perspective.
2. The Magnoliaceae Family
The Magnoliaceae family is among the most primitive living flower plant families. Fossil registries date from the Cretacic period (135–100 million years ago), when dinosaurs were still alive [19,20]. The family is divided into Liriodendron and Magnolia, the latter genus being the largest one, with a total of 312 species [21]. Most of the species identified (~80%) are distributed in temperate and tropical climate zones in south-east Asia, and a smaller number (~20%) are found in the American continent [22]. Magnolia species are easily identified by their morphological characteristics. They are arboreous or shrubby plants with deciduous or evergreen foliage [23]. The flowers are large and solitary, with a perianth of two or more spirals of free tepals (petaloids), many stamens, anthers with two loculaments and carpels arranged in spirals. The fruits are polyfollicles made of joint or separated carpels and they may be dehiscent, circumscissile, or indehiscent, and the seeds are long and wrapped in a crimson red sarcotesta that can be removed from the endocarp [22]. The sarcotesta is also called aril, an edible pulp that covers the seed (Figure 1) [24]. Interestingly, the seeds specifically depend on spreading birds that can detect their red color [25,26].
3. Magnolia in Traditional Medicine
Some Magnolia species have been used in traditional medicine for their therapeutic and pharmacological properties [27]. Houpu, a traditional Chinese remedy made with the bark of M. officinalis, has been used for millennia to treat “energy (qi) stagnation”, the cause of asthma and digestive afflictions, as well as to prevent stress, anxiety, and depression [28]. It has been found that the two main active components of houpu are the lignans, magnolol and honokiol [29]. Likewise, in traditional medicine in India, root bark extracts of M. champaca have been used to treat tumors, constipation, swelling, amenorrhea, and dysmenorrhea, and its flowers are used to treat chronic gastritis, fever, cough, bronchitis, and heart weakness [30]. In traditional medicine in North America, Magnolia has been used to treat several illnesses. The ethnomedical data describe that the tonic obtained from the bark of the root and stem of M. virginiana is a remedy for autumn fever, fever paroxysms, and rheumatism [31]. Native communities use an extract of M. grandiflora seeds, whose sedative and hypnotic effects help to control sleep and body temperature. This extract has also antispasmodic and anti-inflammatory properties and can eliminate the immunoresistance associated with breast and prostate cancers. It can also be used to treat convulsions and fight microbial infections [32]. In Mexico, infusions of flowers and leaves of M. yajlachhi have been used in traditional Zapotec medicine for several purposes, including the strengthening of heart rhythms, invigorating the blood, and the clearing eyes, in addition the aroma of the flowers is used to treat asthma [33]. The decoction of leaves and bark of M. dealbata is used as a tranquilizer and anticonvulsant in cases of epilepsy [34].
4. Magnolia and Sustainable Agriculture
The growing interest in the use of botanical pesticides in agriculture today constitutes a favorable scenario for the application of natural products (botanical extracts, essential oils, and others) derived from Magnolia plants. Several reports highlight the biocidal properties of these compounds on insects that afflict plants of commercial value [35,36]. For example, the raw extracts and essential oils from fruits, seeds, and sarcotesta of Magnolia spp. have been successfully used against insect infestations [37,38,39], and a large variety of phytochemicals with possible insecticidal properties have been proposed. Sarker and Maruyama [22] and Song and Fischer [40] have documented that some Magnolia spp. are rich in lignans, neolignans, alkaloids, flavonoids, and terpenoids, with different biological uses as insecticides, deterrents, repellents, and anti-nutrients [22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40]. Therefore, the use of natural products derived from Magnolia spp. is a good strategy in integral pest management and can help mitigate environmental deterioration and the accumulation of toxic residues derived from the application of synthetic insecticides [41].
5. Botanical Extracts and Essential Oils with Insecticidal Properties
The biocidal potential of natural products derived from native plants to combat different pest insects has called the attention of the scientific community [42]. Though the importance of knowing the properties (chemical composition, biomolecules such as proteins, and genes) of the species that make up the local and endemic flora has become increasingly evident, not all the taxonomic groups of the Magnolia genus have been sufficiently explored [43]. For example, while M. officinalis, an endemic species from China, has been widely studied, the biocidal capabilities of M. fragarigynandria, M. mayae, M. narinensis, and M. rzedowskiana remain scarcely known [21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44]. Moreover, phytochemical studies of the vegetative structures of Magnolia spp. have rarely paid attention to leaf, bark, flower, fruit, seed, and sarcotesta in equal proportions.
The insecticidal properties of Magnolia have been confirmed in a study by Kelm et al., [45], in which extracts of hexane and methanol from fruits of M. salicifolia, a species endemic to Japan, were given to mosquitoes. The results indicate that both of these extracts had a significant biocide potential (250 ppm in 24 h) on Ae. aegypti at the fourth larval stage [45]. Similarly, the insecticidal activity of essential oils of the mature and immature leaves, flowers, and fruits of M. grandiflora, a species endemic to the United States, were found to have the worst toxic effects (49.4 and 48.9 ppm) on Ae. aegypti larvae. Finally, an essential oil obtained from seeds showed a strong repellent effect (0.89) on adult mosquitoes [46].
Wang et al., [47] run toxicity tests of a substance obtained via the hydro-distillation of seeds of M. denudata, a species endemic to China, on larvae of Culex pipens pallens, Ae. aegypti, Ae. albopictus, and Anopheles sinensis and obtained insecticidal bioactivity values of 19.6, 19.3, 21.4, and 24.84 mg/L, respectively [47]. Recently, it was reported that the essential oil of M. grandiflora seeds is highly effective against imported hybrid fire ants (Solenopsis invicta) [48]. Vásquez-Morales et al., [38] on the other hand, reported that the ethanolic extracts of sarcotesta and seed of Magnolia schiedeana, a species endemic to Mexico, have a potential in the development of useful bioinsecticides in the control of adult specimens of Anastrepha ludens, the Mexican fruit fly. In the study, ethanolic extracts of leaves, flowers, bark, empty polyfollicles, seeds, and sarcotesta were evaluated, and it was found that only the seed and sarcotesta extracts had significant levels of insecticidal effectiveness (59.3 and 64.7%) against flies [38]. On the same insect, the ethanolic extracts of leaves, flowers, bark, seeds, and sarcotesta of M. dealbata (currently M. vovidesii, a species endemic to Mexico) were evaluated. It was reported that the ethanolic extracts of sarcotesta showed the highest insecticidal activity level (96%) against A. ludens adults [37]. Additionally, feeding bioessays showed that the sarcotesta extracts of M. perezfarrerae and M pugana, species endemic to Mexico, were 95% and 93% effective, respectively, against Anastrepha ludens adults, while the sarcotesta extracts of M. vovidesii, were 92% effective against A. obliqua, the West Indian fly [49].
6. Secondary Metabolites in Magnolia
6.1. Fruit with Seed
The fruit (polyfollicle) of Magnolias is a structure that is rich in secondary metabolites with specialized metabolic pathways, which are not involved in primary metabolism [50]. The chemical analysis of essential oils derived from fruits of 2 populations of M. ovata, a species endemic to Brazil, showed that they possess a wide diversity of metabolites, including 49 volatile constituents, such as α and β-cubebene, butyl heptanoate, and naphtalene, and 14 non-volatile constituents, such as parthenolide, michelenolide, 1-hexadecanol, as well as 3 alkaloids, lysicamine, lanuginosine and O-methylmoschatoline [51].
The phytochemicals contained in the essential oils of ripe fruits with seed of M. grandiflora comprised 49 terpenes, such as α-pinene, ethyl 2-methylbutyrate, and isobutyl isobutyrate, and 3 fatty acids, such as (Z)-9-methyl octadecanoate (=methyl oleate), (Z.Z)-9,12-methyl octadecadenoate (=methyl linoleate), and hexadecanoic acid [46]. Additionally, the phytochemical study of immature fruits with seeds of M. grandiflora reported the isolation of five chemical compounds (Figure 2) [52]. The presence of nitrile functional groups in the compound in Figure 2E suggest their diversity in terms of biological activity and use in the pharmaceutical industry [52,53]. Additionally, research with phytochemicals of fruits of M. tripetala, a species endemic to the United States, lead to the isolation of tripetalin A and B, 4′-methoxymagnaldehyde B, magnaldehyde B, magnoquinone, and magnotriol B [54].
On the other hand, in the essential oil derived from the dry and fresh fruits of M. kobus, a species endemic to China, Japan, and the Republic of Korea, 17 chemical compounds were found, including α-thujene, α-pinene, and camphene [55]. Similarly, in the fruits of M. obovata, a species endemic to Japan and the Republic of Korea, 20 neolignans (including obovatalignan A, magnolol, and honokiol), six phenylethanoid glycosides (such as (1→2)-β-D-allopyranoside, magnoloside D, and magnoloside A), and five phenylpropanoids (including obovatoside A, syringin, and pavonisol) were identified [56,57,58,59,60,61]. Finally, in the fruit of M. officinalis var. biloba, nine phenylethanoid glycosides, including magnoloside Ia and crassifolioside, were found [62].
6.2. Seedless Fruit (Peel)
As with all fruit structures of Magnolia species, the seedless fruit (or fruit follicles) contain a wide variety of natural compounds. In a recent study, the phytochemicals contained in the essential oils of ripe seedless fruits of M. grandiflora were analyzed [46]. A total of 43 compounds were identified in this structure, including α-pinene, 1,8-cineole, p-cymene, terpinolene, bornyl acetate, α-humulene, myrtenol, and T-cadinol. From the analysis of essential oils from seedless fruits of the species M. acuminata, M. grandiflora, M. fraseri, and M. tripetala, species endemic to Canada and the United States, 34 volatile compounds were identified, including α-pinene, β-myrcene, limonene, eucalyptol, borneol, and trans-nerolidol [63]. Likewise, in the seedless fruits of M. vovidesii, 15 compounds were isolated from sesquiterpene lactones, such as shizukolidol, and phenols, such as protocatechuic acid, among others [64]. In another study, from the essential oil derived from the peel of M. kwangsiensis, a species endemic to China, 21 volatile compounds (including cis-4-thujanol, borneol, and guaiol) were obtained, as well as 10 fatty acids (including heptadecanoic acid, linoleic acid, and heneicosanoic acid [39].
6.3. Seed
Magnolia seeds also contain a wealth of natural chemical compounds. Via the purpose of isolating lignans, extracts of dichloromethane from seeds of M. grandiflora, M. acuminata, and M. virginiana (species endemic to Cuba and the United States) were analyzed, including five phenylpropanoids from M. grandiflora (Figure 3), honokiol, and magnolol from M. virginiana, galgravin, and veraguensin from M. acuminata [65]. Another analysis of the compounds in the essential oil of M. grandiflora seeds reported the presence of 14 chemical compounds, such as 4-(2-propenyl)-phenol, tetradecanoic acid, eucalyptol, and 2,3-dihydroxy-anti-oleic acid ester [66]. The methanolic extract of M. grandiflora seeds presented the neolignans, honokiol and bishonokiol [67]. In a seed hydrodistillate of M. denudata, 17 chemical compounds were found, including p-cymene, 𝛽-caryophyllene, nerolidol, and ethyl palmitate [47]. The CG-MS chemical analysis of the essential oil from seeds of M. pugana obtained 33 chemical compounds, such as isovalerate isobutyl, α-bergamotene, germacrene D, cyclocolorenone, and dehydrosaussurea lactone [68].
6.4. Sarcotesta (Aril)
Since the majority of Magnolia seeds studies are conducted using whole seeds with sarcotesta, only a small amount is known about the secondary metabolites’ profile of this particular structure. The chemical analysis of an essential oil from the sarcotesta of M. kwangsiensis produced 21 terpenes, among them p-menth-2-ene, β-phellandrene, acoradiene, and guaiol, and 10 fatty acids, including pentadecanoic, linoleic, eicosanoic, and heneicosanoic, were among them [39].
A search conducted of the scientific literature obtained 277 chemical compounds in the Magnolia plant structures, attesting to the wide variety of metabolites contained in each of them. The Venn diagram (Figure 4) shows that fruit with seeds is the plant structure with the largest number of exclusive chemical compounds (122), followed by seedless fruit or peel (46), seed (42), and sarcotesta (5). Additionally, fruit with seeds was the structure that presented the largest number of chemical compounds shared with other structures: it shares 17 chemical compounds with fruit without seeds, 13 with the latter one and with seeds, and 1 compound it shares fruit without seeds with seeds. Finally, only five chemical compounds are shared by all the structures (Table S1).
A large part of these compounds has been individually evaluated for different plague insect species. Boulogne et al., [69] pointed out that terpenoids, phenolic compounds, and alkaloids are the most frequently reported compounds in relation to protection from insects. These three types of compounds are the main secondary metabolism groups involved in the ecological interactions of plants, such as competition and herbivory [70].
7. Terpenoids
Terpenoids is a group of secondary metabolites composed of isoprene molecules, units of five carbons (C5), which are known as isopentenil diphosphate (IPP), and dimetilalil diphosphate (DMPP) [71]. In plants, it is possible to synthesize IPP and DMPP following the route of cytosolic mevalonate derived from acetyl-CoA (MEV) and the route of 2-C-metil-D-eritritol-4-phosphate (MEP) plastidial derived from pivurate [72]. Terpenoids are very heterogeneous substances in structure and property, though the majority of them are fat soluble and can be found mainly in essential oils. They stand out for their various degrees of volatility and the influence they have on community and ecosystem interactions [73].
Monoterpenes (C10) and sesquiterpenes (C15) have high degrees of volatility, and they are referred as “inferior terpenoids” [74]. Plants release them in direct defense after an attack by microorganisms, insects, or mammals, but they also have the indirect defense property of attracting the natural predators of attacking herbivores [75]. C10 and C15 are released in complex blends that confuse herbivores and inhibit their capacity to develop resistance to the substances [73].
For these reasons, terpenoids have proved to have insecticidal and anti-nutrient effects on several species of plague insects. However, only a small percentage of monoterpenoids and sesquiterpenoids (approx. 16%) obtained from Magnolia fruits with and without seeds and sarcotesta have been tested on insects and other arthropods (Table 1). This small part can have an important role as bioinsecticides and replace the indiscriminate use of harmful pesticides. Unassayed terpenoids (84%) remain an opportunity for research on bioactivity against several pests [76]. Their synergic effects could be explored by trying them in different combinations. Moreover, the effectiveness of existing natural extracts and essential oils can be enhanced via adding extra terpenoids [77,78].
8. Phenols
Phenolic compounds are characterized as having at least one aromatic ring with one or more hydroxyl groups in combination. They are classified as non-flavonoids or flavonoids [98]. Non-flavonoids include free phenols, phenolic acids, and phenylpropanoids, which in turn are divided into hydroxycinnamic acids, coumarins, phenylpropanoids, lignins, lignans, and neolignans. Flavonoids, on the other hand, are classified into more than 10 classes, including flavones, isoflavones, flavonoids, flavanones, stilbenes, and anthocyanins [99]. The synthesis of phenolic compounds in superior plants happens in two ways: (1) via shikimic-phenylpropanoids (predominantly in plants) and/or (2) via malonate-polyketide-phenylpropanoids (predominantly in bacteria, fungi, and plants) [100].
Phenolic compounds have an ecological function, namely, to prevent nutrient loss in plants as a result of the feeding behavior of phytophaga [70]. They act also as protection agents against the effects of abiotic factors (sunlight and low temperatures) that indirectly modify plant growth, mineral nutrition, and pigment and aroma in flowers and fruits, and additionally, they act as natural toxic inhibitors for animals and invasive organisms [101,102].
It is evident that phenols have the capacity to act as molecules against insects and that they can also have an important role in the protection of crops. Table 2 presents the insecticidal potential of those phenolic components that have been evaluated in different plague insects, which represent approximately 18% of the total compounds reported from Magnolia fruit with and without seeds and sarcotesta. It is important to note that a high percentage (82%) of the phenolic compounds obtained have not yet been assayed (Table 2) either in the laboratory or in the field to determine their insecticidal effect on several plagues.
9. Alkaloids
Alkaloids are organic substances containing nitrogen, with a high or low degree of base quality [116]. In the case of Magnolia, alkaloids of the aporphine type belonging to the class of isoquinolines were identified in the fruit of M. ovata [51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117]. Their biosynthesis is achieved via benzylisoquinolines that give way to (s)-reticulin, an intermediary metabolite key to the formation of aporphines [118]. In general, alkaloids are part of the arsenal of chemical defense against herbivores and pathogens [119]. They can be characterized as repellents, deterrents, anti-nutrients, toxic, allelopathics, or germination inhibitors [120]. However, the study of the insecticidal interaction between lysicamine aporphines, lanuginosine, and O-methylmoschatoline has not been determined. This is an area of opportunity to ask new research questions about unassayed alkaloids and get to learn about their bioactivity against pest insects and other organisms.
10. Magnolia: Between Bioprospection and Conservation
Existing information on the biology of Magnolia species remains scarce and, in many cases, restricted to the study of taxonomic aspects [21,121]. According to the Red List of Magnoliaceae, 85% of species are at risk or have insufficient data [21]. The indiscriminate felling of trees, land use changes, and high fragmentation rates of native habitats form a scenario in which practically the totality of Magnolia species are under some degree of threat [122]. Moreover, the susceptibility of Magnolia increases as the inadequate conditions of restricted habitats and the predation from soil-bound organisms prevent the successful dispersion and germination of seeds [123].
If we add knowledge about existing lacunae and the physiological and genetic aspects of these plants, the geographical distribution patterns of plant diversity, in general, the effects and responses to anthropic processes, and the mechanisms for conservation to this picture, it becomes evident that developing strategies to help and promote the conservation of Magnolia species must be considered as a priority [43,124,125].
The bioprospection of plants for the obtention of bioinsecticides based on natural extracts and secondary metabolites can produce more information about this group of species. This would attract the attention of researchers and elicit interest in their preservation via correct methods, tactics, and planning to ensure the equilibrium and dynamics of Magnolia populations [126]. In the agricultural sector, Magnolia natural products have the potential to be part of integrated pest control management and contribute to a sustainable agriculture free from the use of dangerous synthetic pesticides [127].
The following question arises then: how can Magnolia be used to protect crops? Judging from the data obtained in the revision of scientific literature, we believe that the ideal sources of bioprospection are the sarcotesta and seedless fruit. When they are not consumed by a dispersal agent, the sarcotesta inhibits the seed germination process, and the fruit, once empty, decomposes into organic matter, a viable source of bioactive components [128,129]. Removing both seeds and sarcotesta improves the germination process and contributes to the in situ and ex situ propagation of species [43]. The implementation of a restoration strategy via reinstating individuals to their natural populations (in situ conservation) can be complemented by ex situ conservation in botanical gardens, natural protected areas, and germplasm banks [21]. These actions combined will help to reduce the risk of extinction. Finally, it has been observed that seeds are a great source of secondary metabolites, and so, it is important to isolate the existing bioactive insecticides and evaluate their effectiveness in pest control around the world.
Currently, Honokiol and Magnolol isolated from Magnolia officinalis are available on the active compounds market to treat several human diseases, respiratory diseases [22,130], cancer [131,132], obesity [133], intestinal problems, and gastric disturbances [22]. In this review, we suggest that the use of Magnolia active compounds against agriculture pests has a huge economic potential, for example: (I) Terpenoids: β-caryophyllene has an insecticide effect on Hymenoptera and Coleoptera [80,87], β-phellandrene and α-terpinene have an insecticide effect on Coleoptera [83,84], p-cymene has an insecticide effect on Diptera [85], and E-nerolidol has an insecticide effect on Hemiptera [93]. (II) Phenols: 2,4-di-tert-butylphenol has an insecticide effect on Trombidiformes [103], Protochatecuic acid has an insecticide effect on Diptera [110], Rutin has an insecticide effect on Orthoptera [113], and Estragol and Quercetin have an insecticide effect on Lepidoptera [104,111,112].
11. Conclusions
The interest in the potential of botanical pesticides in agricultural practices to reduce the use of synthetic ones is on the rise. Magnolia species are an ideal source of natural bioactive insecticides. As shown in previous studies, seedless fruit and sarcotesta constitute an excellent source for the study of phytochemicals with potential pest control properties. In this study, we considered these two plant structures and their possible role in the production of natural crop-protection ingredients. However, Magnolia species are vulnerable, and the extraction of any part of the plant for research on natural products and the obtention of raw extracts must follow a strict procedure and be managed according to local demographic conditions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules28124681/s1, Table S1: Compounds identified in Magnolia’s fruit parts.
Author Contributions
J.V.H.-R.: literature review and manuscript writing. S.G.V.-M.: manuscript review, corrections, and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
This works has been supported by the Consejo Nacional de Ciencia y Tecnología (CONACYT) in Mexico via a scholarship for a master’s degree received by the first author. We also thank to three reviews for their valuable suggestions and comments.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Not applicable.
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- FAO. Plant Health and Food Security. Food and Agriculture Organization. 2020. Available online: http://www.fao.org/3/i7829en/I7829EN.pdf (accessed on 21 April 2021).
- Altieri, M.A.; Nicholls, C.I. Biodiversidad y Manejo de Plagas en Agroecosistemas; Icaria Editorial: Capellades, Barcelona, 2007; Volume 2. [Google Scholar]
- Selfa, J. Plagas agrícolas y forestales. Artrópodos Hombre 1997, 20, 75–91. [Google Scholar]
- FAO. Codex Pesticides Residues in Food Online Database. Food and Agriculture Organization of the United Nations. 2019. Available online: https://www.fao.org/fao-who-codexalimentarius/codex-texts/dbs/pestres/en/ (accessed on 25 February 2021).
- FAO. Terms and Definitions. Pesticide Registration Toolkit. Food and Agriculture Organization. 2021. Available online: http://www.fao.org/pesticide-registration-toolkit/information-sources/terms-and-definitions/terms-and-definitions-p/en/ (accessed on 25 February 2021).
- Gilden, R.C.; Huffling, K.; Sattler, B. Pesticides and health risks. J. Obstet. Gynecol. Neonatal Nurs. 2010, 39, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Coats, J.R. Risks from natural versus synthetic insecticides. Annu. Rev. Entomol. 1994, 39, 489–515. [Google Scholar] [CrossRef]
- FAO. Quinua Manejo Integrado de Plagas. Estrategias en el Cultivo de la Quinua Para Fortalecer el Sistema Agroalimentario en la Zona Andina. Organización de Las Naciones Unidas Para La Alimentación y La Agricultura. 2016. Available online: http://www.fao.org/3/i6038s/i6038s.pdf (accessed on 2 May 2021).
- Grdiša, M.; Gršić, K. Botanical insecticides in plant protection. Agric. Conspec. Sci. Cus. 2013, 78, 85–93. [Google Scholar]
- Singh, D. Advances in Plant. In Biopesticides; Singh, D., Ed.; Springer India: Uttar Pradesh, India, 2014. [Google Scholar] [CrossRef]
- Táborsky, V. Small-Scale Processing of Microbial Pesticides; FAO Agricultural Services Bulletin No. 96. Food and Agriculture Organization of the United Nations Rome: Prague, Czech Republic, 1992. [Google Scholar]
- Riyaz, M.; Mathew, P.; Zuber, S.M.; Rather, G.A. Botanical pesticides for an eco-friendly and sustainable agriculture: New challenges and prospects. In Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2022; pp. 69–96. [Google Scholar] [CrossRef]
- Lengai, G.M.W.; Muthomi, J.W.; Mbega, E.R. Phytochemical activity and role of botanical pesticides in pest management for sustainable agricultural crop production. Sci. Afr. 2020, 7, e00239. [Google Scholar] [CrossRef]
- Yang, X.; Huang, Q.; Jiang, T.; Xu, H. Degradation dynamics of azadirachtin in cabbage and soil. J. South China Agric. Univ. 2007, 38, 37–40. [Google Scholar]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Swain, S.P.; Gupta, S.; Das, N.; Costa-Franca, T.C.; da Silva-Goncalves, A.; Castro-Ramalho, T.; Subrahmanya, S.; Narsaria, U.; Deb, D.; Mishra, N. Flavanones: A potential natural inhibitor of the ATP binding site of PknG of Mycobacterium tuberculosis. J. Biomol. Struct. Dyn. 2022, 40, 11885–11899. [Google Scholar] [CrossRef]
- Santos, J.A.N.; Kondo, M.Y.; Freitas, R.F.; dos Santos, M.H.; Ramalho, T.C.; Assis, D.M.; Juliano, L.; Juliano, A.M.; Puzer, L. The natural flavone fukugetin as a mixed-type inhibitor for human tissue kallikreins. Bioorganic Med. Chem. Lett. 2016, 26, 1485–1489. [Google Scholar] [CrossRef]
- Nelson, G. America’s magnolias. The American Gardener. Am. Hortic. Soc. Alex. 2008, 87, 38–43. [Google Scholar]
- Romanov, M.S.; Dilcher, D.L. Fruit structure in Magnoliaceae s.l. and Archaeanthus and their relationships. Am. J. Bot. 2013, 100, 1494–1508. [Google Scholar] [CrossRef] [PubMed]
- Rivers, M.; Beech, E.; Murphy, L.; Oldfield, S. The Red List of Magnoliaceae—Revised and Extended; Botanic Gardens Conservation International: Richmond, UK, 2016. [Google Scholar]
- Sarker, S.D.; Maruyama, Y. Magnolia: The Genus Magnolia; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Vázquez-García, J.; Muñiz-Castro, M.A.; Arroyo, F.; Pérez, Á.; Serna González, M.; Cuevas Guzmán, R.; Domínguez-Yescas, R.; de Castro-Arce, E.; Gurrola-Díaz, C. Novelties in Neotropical Magnolia and an Addendum Proposal to the IUCN Red List of Magnoliaceae; University of Guadalajara: Jalisco, Mexico, 2013; pp. 461–496. [Google Scholar]
- Boesewinkel, F.D.; Bouman, F. The seed: Structure. In Embryology of Angiosperms; Springer: Berlin/Heidelberg, Germany, 1984; pp. 567–610. [Google Scholar] [CrossRef]
- Bauman, A.J.; Yokoyama, H. Magnolia seed carotenoid pigments: Typical evolutionarily-static relicts? J. Theor. Biol. 1975, 53, 277–284. [Google Scholar]
- Cazetta, E.; Rubim, P.; de Oliveira, V.; Roberto-Francisco, M.; Galetti, M. Frugivoria e dispersão de sementes de Talauma ovata (Magnoliaceae) no sudeste brasileiro. Ararajuba 2002, 10, 199–206. [Google Scholar]
- Lee, Y.D. Use of Magnolia (Magnolia grandiflora) seeds in medicine, and possible mechanisms of action. In Nuts and Seeds in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2011; pp. 727–732. [Google Scholar] [CrossRef]
- Patočka, J.; Jakl, J.; Strunecká, A. Expectations of biologically active compounds of the genus Magnolia in biomedicine. J. Appl. Biomed. 2006, 4, 171–178. [Google Scholar] [CrossRef]
- Li, J.; Copmans, D.; Partoens, M.; Hunyadi, B.; Luyten, W.; de Witte, P. Zebrafish-based screening of antiseizure plants used in Traditional Chinese Medicine: Magnolia officinalis extract and its constituents magnolol and honokiol exhibit potent anticonvulsant activity in a therapy-resistant epilepsy model. ACS Chem. Neurosci. 2020, 11, 730–742. [Google Scholar] [CrossRef]
- Ramyashree, C.; Hemalatha, K. Ethnomedicinal profile on Magnolia species (Magnoliaceae): A review. Int. J. Herb. Med. 2020, 8, 39–46. Available online: https://www.florajournal.com/archives/2020/vol8issue3/PartA/7-5-101-194.pdf (accessed on 2 July 2021).
- Schühly, W.; Khan, I.; Fischer, N.H. The ethnomedicinal uses of Magnoliaceae from the southeastern United States as leads in drug discovery. Pharm. Biol. 2001, 39, 63–69. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lee, Y.M.; Lee, C.K.; Jung, J.K.; Han, S.B.; Hong, J.T. Therapeutic applications of compounds in the Magnolia family. Pharmacol. Ther. 2011, 130, 157–176. [Google Scholar] [CrossRef]
- Domínguez-Yescas, R.; Vázquez-García, J.A. Flower of the heart, Magnolia yajlachhi (subsect. Talauma, Magnoliaceae), a new species of ceremonial, medicinal, conservation and nurse tree relevance in the Zapotec culture, Sierra Norte de Oaxaca, Mexico. Phytotaxa 2019, 393, 21. [Google Scholar] [CrossRef]
- Martínez, A.L.; Domínguez, F.; Orozco, S.; Chávez, M.; Salgado, H.; González, M.; González-Trujano, M.E. Neuropharmacological effects of an ethanol extract of the Magnolia dealbata Zucc. leaves in mice. J. Ethnopharmacol. 2006, 106, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.X.; Lu, X.X.; Du, Y.S.; Zheng, Y.; Zeng, D.; Du, S.S. Sesquiterpenoid-rich essential oils from two Magnolia plants: Contact and Repellent Activity to three stored-product insects. J. Oleo Sci. 2022, 71, ess21241. [Google Scholar] [CrossRef]
- Luu-Dam, N.A.; Tabanca, N.; Estep, A.S.; Nguyen, D.H.; Kendra, P.E. Insecticidal and attractant activities of Magnolia citrata leaf essential oil against two major pests from Diptera: Aedes aegypti (Culicidae) and Ceratitis capitata (Tephritidae). Molecules 2021, 26, 2311. [Google Scholar] [CrossRef] [PubMed]
- Flores-Estévez, N.; Vasquez-Morales, S.G.; Cano-Medina, T.; Sánchez-Velásquez, L.R.; Noa-Carrazana, J.C.; Díaz-Fleischer, F. Insecticidal activity of raw ethanolic extracts from Magnolia dealbata Zucc on a tephritid pest. J. Environ. Sci. Health Part B 2013, 48, 582–586. [Google Scholar] [CrossRef]
- Vásquez-Morales, S.; Norma, F.E.; Sánchez-Velásquez, L.R.; María del Rosario, P.L.; Viveros-Viveros, H.; Díaz-Fleischer, F. Bioprospecting of botanical insecticides: The case of ethanol extracts of Magnolia schiedeana Schltl. applied to a Tephritid, fruit fly Anastrepha ludens Loew. J. Entomol. Zool. Stud. 2015, 3, 1–5. [Google Scholar]
- Zheng, Y.F.; Liu, X.M.; Zhang, Q.; Lai, F.; Ma, L. Constituents of the essential oil and fatty acid from rare and endangered plant Magnolia kwangsiensis Figlar & Noot. J. Essent. Oil Bear. Plants 2019, 22, 141–150. [Google Scholar] [CrossRef]
- Song, Q.; Fischer, N.H. Biologically active lignans and neolignans from Magnolia species. Rev. Soc. Química México 1999, 43, 211–218. [Google Scholar]
- Damalas, C.A.; Koutroubas, S.D. Botanical pesticides for eco-friendly pest management. In Pesticides in Crop Production; Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 181–193. [Google Scholar] [CrossRef]
- Karkanis, A.C.; Athanassiou, C.G. Natural insecticides from native plants of the Mediterranean basin and their activity for the control of major insect pests in vegetable crops: Shifting from the past to the future. J. Pest Sci. 2021, 94, 187–202. [Google Scholar] [CrossRef]
- Vázquez-García, J.A.; Muñiz-Castro, M.A.; Dahua-Machoa, A.; Osorio-Muñoz, E.A.; Hernández-Vera, G.; Ortega-Peña, A.S.; Romo-Campos, R.L.; Jacobo-Pereira, C. How to save endangered magnolias? From population biology to conservation action: The case of allopatric radiation in western Mexico. In Endangered Plants; Kumar, S., Ed.; IntechOpen: London, UK, 2021; pp. 13–56. [Google Scholar]
- Poivre, M.; Duez, P. Biological activity and toxicity of the Chinese herb Magnolia officinalis Rehder & E. Wilson (Houpo) and its constituents. J. Zhejiang Univ. Sci. B 2017, 18, 194–214. [Google Scholar] [CrossRef]
- Kelm, M.A.; Nair, M.G.; Schutzki, R.A. Mosquitocidal Compounds from Magnolia salicifolia. Int. J. Pharmacogn. 1997, 35, 84–90. [Google Scholar] [CrossRef]
- Ali, A.; Tabanca, N.; Demirci, B.; Raman, V.; Budel, J.M.; Baser, K.H.C.; Khan, I.A. Insecticidal and biting deterrent activities of Magnolia grandiflora essential oils and selected pure compounds against Aedes aegypti. Molecules 2020, 25, 1359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Perumalsamy, H.; Wang, M.; Shu, S.; Ahn, Y.J. Larvicidal activity of Magnolia denudata seed hydrodistillate constituents and related compounds and liquid formulations towards two susceptible and two wild mosquito species. Pest Manag. Sci. 2016, 72, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Chen, J.; Khan, I.A. Toxicity and repellency of Magnolia grandiflora seed essential oil and selected pure compounds against the workers of hybrid imported fire ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2022, 115, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Vásquez Morales, S.G.; Alvarez Vega, E.A.; Infante Rodríguez, D.A.; Huchin Mian, J.P.; Pedraza Reyes, M. Evaluación de extractos de árboles endemicos (Magnolia spp.) de México contra la plaga de la mosca de la fruta y estudio fitoquímico preliminar. Polibotánica 2022, 53, 168–182. [Google Scholar] [CrossRef]
- Chadwick, D.J.; Whelan, J. Secondary Metabolites: Their Function and Evolution; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Barros, L.F.L.; Ehrenfried, C.A.; Riva, D.; Barison, A.; de Mello-Silva, R.; Stefanello, M.E.A. Essential oil and other constituents from Magnolia ovata fruit. Nat. Prod. Commun. 2012, 7, 1934578X1200701. [Google Scholar] [CrossRef]
- Garza, B.; Echeverria, A.; Gonzalez, F.; Castillo, O.; Eubanks, T.; Bandyopadhyay, D. Phytochemical investigation of Magnolia grandiflora green seed cones: Analytical and phytoceutical studies. Food Sci. Nutr. 2019, 7, 1761–1767. [Google Scholar] [CrossRef]
- Scotti, C.; Barlow, J.W. Natural products containing the Nitrile functional group and their biological activities. Nat. Prod. Commun. 2022, 17, 1–24. [Google Scholar] [CrossRef]
- Nie, W.; Ding, L.F.; Lei, T.; Liu, Z.X.; Li, J.D.; Song, L.D.; Wu, X.D. Biphenyl-type neolignans with NO inhibitory activity from the fruits of Magnolia tripetala. Phytochem. Lett. 2021, 44, 222–226. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Cho, H.; Yu, B.; Kim, S. Comparison of essential oil compositions of fresh and dried fruits of Magnolia kobus DC. J. Appl. Pharm. Sci. 2016, 6, 146–149. [Google Scholar] [CrossRef]
- Pyo, M.K.; Lee, Y.; Yun-Choi, H.S. Anti-platelet effect of the constituents isolated from the barks and fruits of Magnolia obovata. Arch. Pharmacal Res. 2002, 25, 325–328. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, D.Y.; Lee, D.S.; Park, J.H.; Jeong, R.H.; Jung, Y.J.; Shrestha, S.; Chung, I.S.; Kim, G.S.; Kim, Y.C.; et al. Neolignans from the fruits of Magnolia obovata and their inhibition effect on NO production in LPS-induced RAW 264.7 cells. Planta Med. 2013, 79, 1335–1340. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, D.Y.; Jung, J.W.; Lee, D.S.; Kim, Y.C.; Lee, Y.H.; Baek, N.I. Neolignans from the fruits of Magnolia obovata inhibit NO production and have neuroprotective effects. Helv. Chim. Acta 2016, 99, 411–415. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, D.Y.; Lee, Y.G.; Baek, N.I. Dineolignans of 3-O-4′ diphenyl ether-type from fruits of Magnolia obovata. Phytochemistry 2017, 136, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.H.; Lee, D.Y.; In, S.J.; Lee, D.G.; Kang, H.C.; Song, M.C.; Baek, N.I. Phenylethanoid glycosides from the fruits of Magnolia obovata. Chem. Nat. Compd. 2015, 51, 660–665. [Google Scholar] [CrossRef]
- Seo, K.H.; Nam, Y.H.; Lee, D.Y.; Ahn, E.M.; Kang, T.H.; Baek, N.I. Recovery effect of phenylpropanoid glycosides from Magnolia obovata fruit on alloxan-induced pancreatic islet damage in zebrafish (Danio rerio). Carbohydr. Res. 2015, 416, 70–74. [Google Scholar] [CrossRef]
- Ge, L.; Zhang, W.; Zhou, G.; Ma, B.; Mo, Q.; Chen, Y.; Wang, Y. Nine phenylethanoid glycosides from Magnolia officinalis var biloba fruits and their protective effects against free radical-induced oxidative damage. Sci. Rep. 2017, 7, 45342. [Google Scholar] [CrossRef] [PubMed]
- Schühly, W.; Ross, S.A.; Mehmedic, Z.; Fischer, N.H. Essential oil analysis of the follicles of four North American Magnolia species. Nat. Prod. Commun. 2008, 3, 1934578X0800300. [Google Scholar] [CrossRef]
- Ramírez-Reyes, T.; Monribot-Villanueva, J.L.; Jiménez-Martínez, O.D.; Aguilar-Colorado, Á.S.; Bonilla-Landa, I.; Flores-Estévez, N.; Luna-Rodríguez, M.; Guerrero-Analco, J.A. Sesquiterpene lactones and phenols from polyfollicles of Magnolia vovidesii and their antimicrobial activity. Nat. Prod. Commun. 2018, 13, 1934578X1801300. [Google Scholar] [CrossRef]
- Schühly, W.; Khan, S.I.; Fischer, N.H. Neolignans from north American Magnolia species with cyclooxygenase 2 inhibitory Activity. Inflammopharmacology 2009, 17, 106–110. [Google Scholar] [CrossRef]
- Luo, M.; Sun, J.; Zhang, B.; Jiang, L. Chemical composition and antioxidant activity of essential oil from Magnolia grandiflora L. seed. Wuhan Univ. J. Nat. Sci. 2012, 17, 249–254. [Google Scholar] [CrossRef]
- Li, H.M.; Zhao, S.R.; Huo, Q.; Ma, T.; Liu, H.; Lee, J.K.; Hong, Y.S.; Wu, C.Z. A new dimeric neolignan from Magnolia grandiflora L. seeds. Arch. Pharmacal Res. 2015, 38, 1066–1071. [Google Scholar] [CrossRef]
- Osorio-Muñoz, E.A. Identificación Química y Perspectiva Medicinal de los aceites esenciales de hojas, semillas y flores de Magnolia pugana. Tesis de Maestría en Ciencias en Biosistemática y Manejo de Recursos Naturales y Agrícolas. Master’s Thesis, Universidad de Guadalajara, Guadalajara, México, 2020. [Google Scholar]
- Boulogne, I.; Petit, P.; Ozier-Lafontaine, H.; Desfontaines, L.; Loranger-Merciris, G. Insecticidal and antifungal chemicals produced by plants: A review. Environ. Chem. Lett. 2012, 10, 325–347. [Google Scholar] [CrossRef]
- Merillon, J.M.; Ramawat, K.G. Co-evolution of Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- Karunanithi, P.S.; Zerbe, P. Terpene synthases as metabolic gatekeepers in the evolution of plant terpenoid chemical diversity. Front. Plant Sci. 2019, 10, 1166. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczuk, A.; Skalicka-Woźniak, K.; Georgiev, M.I. Terpenoids. In Pharmacognosy; Elsevier: Amsterdam, The Netherlands, 2017; pp. 233–266. [Google Scholar] [CrossRef]
- Langenheim, J.H. Higher plant terpenoids: A phytocentric overview of their ecological roles. J. Chem. Ecol. 1994, 20, 1223–1280. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.X.; Lou, Y.G.; Mao, Y.B.; Lu, S.; Wang, L.J.; Chen, X.Y. Plant terpenoids: Biosynthesis and ecological functions. J. Integr. Plant Biol. 2007, 49, 179–186. [Google Scholar] [CrossRef]
- Mumm, R.; Posthumus, M.A.; Dicke, M. Significance of terpenoids in induced indirect plant defense against herbivorous arthropods. Plant Cell Environ. 2008, 31, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Andrés, M.F.; Rossa, G.E.; Cassel, E.; Vargas, R.M.F.; Santana, O.; Díaz, C.E.; González-Coloma, A. Biocidal effects of Piper hispidinervum (Piperaceae) essential oil and synergism among its main components. Food Chem. Toxicol. 2017, 109, 1086–1092. [Google Scholar] [CrossRef]
- Mesbah, H.A.; Saad, A.S.A.; Mourad, A.K.; Taman, F.A.; Mohamed, I.B. Joint action of quercetin with four insecticides on the cotton leaf-worm larvae, Spodoptera littoralis Boisd. (Lep.: Noctuidae) in Egypt. Commun. Agric. Appl. Biol. Sci. 2007, 72, 445–457. [Google Scholar]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Benelli, G.; Govindarajan, M.; Rajeswary, M.; Vaseeharan, B.; Alyahya, S.A.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Maggi, F. Insecticidal activity of camphene, zerumbone and α-humulene from Cheilocostus speciosus rhizome essential oil against the Old-World bollworm, Helicoverpa armigera. Ecotoxicol. Environ. Saf. 2018, 148, 781–786. [Google Scholar] [CrossRef]
- de Oliveira, B.M.S.; Melo, C.R.; Santos, A.C.C.; Nascimento, L.F.A.; Nízio, D.A.C.; Cristaldo, P.F.; Blank, A.F.; Bacci, L. Essential oils from Varronia curassavica (Cordiaceae) accessions and their compounds (E)-caryophyllene and α-humulene as an alternative to control Dorymyrmex thoracius (Formicidae: Dolichoderinae). Environ. Sci. Pollut. Res. 2019, 26, 6602–6612. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Du, S.S. Fumigant components from the essential oil of Evodia rutaecarpa hort unripe fruits. E-J. Chem. 2011, 8, 1937–1943. [Google Scholar] [CrossRef]
- Choi, W.S.; Park, B.S.; Lee, Y.H.; Jang, D.Y.; Yoon, H.Y.; Lee, S.E. Fumigant toxicities of essential oils and monoterpenes against Lycoriella mali adults. Crop Prot. 2006, 25, 398–401. [Google Scholar] [CrossRef]
- Cao, J.; Pang, X.; Guo, S.; Wang, Y.; Geng, Z.; Sang, Y.; Guo, P.; Du, S. Pinene-rich essential oils from Haplophyllum dauricum (L.) G. Don display anti-insect activity on two stored-product insects. Int. Biodeterior. Biodegrad. 2019, 140, 1–8. [Google Scholar] [CrossRef]
- Saad, M.M.G.; El-Deeb, D.A.; Abdelgaleil, S.A.M. Insecticidal potential and repellent and biochemical effects of phenylpropenes and monoterpenes on the red flour beetle, Tribolium castaneum Herbst. Environ. Sci. Pollut. Res. 2019, 26, 6801–6810. [Google Scholar] [CrossRef]
- Zhang, Z.; Xie, Y.; Wang, Y.; Lin, Z.; Wang, L.; Li, G. Toxicities of monoterpenes against housefly, Musca domestica L. (Diptera: Muscidae). Environ. Sci. Pollut. Res. 2017, 24, 24708–24713. [Google Scholar] [CrossRef] [PubMed]
- AlShebly, M.M.; AlQahtani, F.S.; Govindarajan, M.; Gopinath, K.; Vijayan, P.; Benelli, G. Toxicity of ar-curcumene and epi-β-bisabolol from Hedychium larsenii (Zingiberaceae) essential oil on malaria, chikungunya and St. Louis encephalitis mosquito vectors. Ecotoxicol. Environ. Saf. 2017, 137, 149–157. [Google Scholar] [CrossRef]
- Pang, X.; Almaz, B.; Qi, X.J.; Wang, Y.; Feng, Y.X.; Geng, Z.F.; Xi, C.; Du, S.S. Bioactivity of essential oil from Atalantia buxifolia leaves and its major sesquiterpenes against three stored-product insects. J. Essent. Oil Bear. Plants 2020, 23, 38–50. [Google Scholar] [CrossRef]
- Feng, Y.X.; Wang, Y.; Chen, Z.Y.; Guo, S.S.; You, C.X.; Du, S.S. Efficacy of bornyl acetate and camphene from Valeriana officinalis essential oil against two storage insects. Environ. Sci. Pollut. Res. 2019, 26, 16157–16165. [Google Scholar] [CrossRef]
- Chu, S.S.; Jiang, G.H.; Liu, Z.L. Insecticidal compounds from the essential oil of Chinese medicinal herb Atractylodes chinensis. Pest Manag. Sci. 2011, 67, 1253–1257. [Google Scholar] [CrossRef]
- Zhu, L.; Tian, Y. Chemical composition and larvicidal activity of essential oil of Artemisia gilvescens against Anopheles anthropophagus. Parasitol. Res. 2013, 112, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Yoon, J.S.; Jung, J.W.; Hong, K.B.; Ahn, Y.J.; Kwon, H.W. Toxicity and repellency of origanum essential oil and its components against Tribolium castaneum (Coleoptera: Tenebrionidae) adults. J. Asia-Pac. Entomol. 2010, 13, 369–373. [Google Scholar] [CrossRef]
- Govindarajan, M.; Rajeswary, M.; Benelli, G. δ-Cadinene, calarene and δ-4-carene from Kadsura heteroclita essential oil as novel larvicides against malaria, dengue and filariasis mosquitoes. Comb. Chem. High Throughput Screen. 2016, 19, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R.; Drenaggi, E.; Desneux, N.; Maggi, F. Phytol, (E)-nerolidol and spathulenol from Stevia rebaudiana leaf essential oil as effective and eco-friendly botanical insecticides against Metopolophium dirhodum. Ind. Crops Prod. 2020, 155, 112844. [Google Scholar] [CrossRef]
- Wu, M.; Xiong, Y.; Han, R.; Dong, W.; Xiao, C. Fumigant toxicity and oviposition deterrent activity of volatile constituents from Asari Radix et Rhizoma against Phthorimaea operculella (Lepidoptera: Gelechiidae). J. Insect Sci. 2020, 20, 32. [Google Scholar] [CrossRef]
- Liu, T.; Wang, C.J.; Xie, H.Q.; Mu, Q. Guaiol-a naturally occurring insecticidal sesquiterpene. Nat. Prod. Commun. 2013, 8, 1353–1354. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Zhao, N.; Du, S.; Yang, K.; Wang, C.F.; Liu, Z.L.; Qiao, Y.J. Insecticidal activity of the essential oil of Lonicera japonica flower buds and its main constituent compounds against two grain storage insects. J. Med. Plants Res. 2012, 6, 912–917. [Google Scholar] [CrossRef]
- Traboulsi, A.F.; Taoubi, K.; El-Haj, S.; Bessiere, J.; Rammal, S. Insecticidal properties of essential plant oils against the mosquito Culex pipiens molestus (Diptera: Culicidae). Pest Manag. Sci. 2002, 58, 491–495. [Google Scholar] [CrossRef]
- Crozier, A.; Clifford, M.N.; Ashihara, H. Phenols, Polyphenols, and Tannins: An Overview. Plant Secondary Metabolites: Occurrence, Structure, and role in the Human Diet. Blackwell Publishing: Tokio, Japan, 2006; Volume 1. [Google Scholar]
- de la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–271. [Google Scholar] [CrossRef]
- Azcón-Bieto, J.; Talón, M. Introducción al metabolismo secundario. In Fundamentos de Fisiología Vegetal, 2nd ed.; McGRAW-HILL—Interamericana De España: Madrid, España, 2013; pp. 326–340. [Google Scholar]
- Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1543–1580. [Google Scholar] [CrossRef]
- McClure, J.W. The physiology of phenolic compounds in plants. In Biochemistry of Plant Phenolics; Springer: Boston, MA, USA, 1979; pp. 525–556. [Google Scholar]
- Chen, Y.; Bertrand, C.; Dai, G.; Yuan, J. Biochemical mechanisms of acaricidal activity of 2,4-di-tert-butylphenol and ethyl oleate against the carmine spider mite Tetranychus cinnabarinus. J. Pest Sci. 2018, 91, 405–419. [Google Scholar] [CrossRef]
- de Menezes, C.W.G.; Carvalho, G.A.; Alves, D.S.; de Carvalho, A.A.; Aazza, S.; de Oliveira Ramos, V.; Pinto, J.E.B.P.; Bertolucci, S.K.V. Biocontrol potential of methyl chavicol for managing Spodoptera frugiperda (Lepidoptera: Noctuidae), an important corn pest. Environ. Sci. Pollut. Res. 2020, 27, 5030–5041. [Google Scholar] [CrossRef]
- Obeng-Ofori, D.; Reichmuth, C.H. Bioactivity of eugenol, a major component of essential oil of Ocimum suave (Wild.) against four species of stored-product Coleoptera. Int. J. Pest Manag. 1997, 43, 89–94. [Google Scholar] [CrossRef]
- González Armijos, M.J.; Viteri Jumbo, L.; Faroni, L.R.; Oliveira, E.E.; Flores, A.F.; Heleno, F.; Haddi, K. Fumigant toxicity of eugenol and its negative effects on biological development of Callosobruchus maculatus L. Rev. Cienc. Agrícolas 2019, 36, 5–15. [Google Scholar] [CrossRef]
- Wang, Z.; Perumalsamy, H.; Wang, X.; Ahn, Y.J. Toxicity and possible mechanisms of action of honokiol from Magnolia denudata seeds against four mosquito species. Sci. Rep. 2019, 9, 411. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.H.; Li, C.; Yang, J.; Ma, L.; Xiao, Y.; Hu, J.; Rajput, N.A.; Gao, C.F.; Zhang, Y.Y.; Wang, M.H. Construction of an immobilised acetylcholinesterase column and its application in screening insecticidal constituents from Magnolia officinalis. Pest Manag. Sci. 2015, 71, 607–615. [Google Scholar] [CrossRef] [PubMed]
- González-Coloma, A.; Escoubas, P.; Mizutani, J.; Lajide, L. Insect growth inhibitors from Machilus japonica. Phytochemistry 1994, 35, 607–610. [Google Scholar] [CrossRef]
- Amin, E.M.; Radwan, M.; El-Hawary, S.S.; Fathy, M.M.; Mohammed, R.J.; Becnel, J.; Khan, I. Potent insecticidal secondary metabolites from the medicinal plant Acanthus montanus. Rec. Nat. Prod. 2012, 6, 301–306. [Google Scholar]
- Selin-Rani, S.; Senthil-Nathan, S.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Edwin, E.S.; Ponsankar, A.; Lija-Escaline, J.; Kalaivani, K.; Abdel-Megeed, A.; Hunter, W.B.; et al. Toxicity and physiological effect of quercetin on generalist herbivore, Spodoptera litura Fab. and a non-target earthworm Eisenia fetida Savigny. Chemosphere 2016, 165, 257–267. [Google Scholar] [CrossRef]
- Shaver, T.N.; Lukefahr, M.J. Effect of flavonoid pigments and gossypol on growth and development of the Bollworm, tobacco budworm, and pink bollworm 123. J. Econ. Entomol. 1969, 62, 643–646. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, X.; Chang, B.H.; Zhang, Z. The survival, growth, and detoxifying enzyme activities of grasshoppers Oedaleus asiaticus (Orthoptera: Acrididae) exposed to toxic rutin. Appl. Entomol. Zool. 2020, 55, 385–393. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Bhakuni, R.S.; Upadhyay, S.; Gaur, R. Insect feeding deterrent and growth inhibitory activities of scopoletin isolated from Artemisia annua against Spilarctia obliqua (Lepidoptera: Noctuidae). Insect Sci. 2011, 18, 189–194. [Google Scholar] [CrossRef]
- Cis, J.; Nowak, G.; Kisiel, W. Antifeedant properties and chemotaxonomic implications of sesquiterpene lactones and syringin from Rhaponticum pulchrum. Biochem. Syst. Ecol. 2006, 34, 862–867. [Google Scholar] [CrossRef]
- Hesse, M. Alkaloids: Nature’s Curse or Blessing? 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Shamma, M. The Isoquinoline Alkaloids Chemistry and Pharmacology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 25. [Google Scholar]
- Morris, J.S.; Facchini, P.J. Isolation and characterization of reticuline N-methyltransferase involved in biosynthesis of the aporphine alkaloid magnoflorine in opium poppy. J. Biol. Chem. 2016, 291, 23416–23427. [Google Scholar] [CrossRef] [PubMed]
- Facchini, P.J. Alkaloid biosynthesis in plants: Biochemistry, cell biology, molecular regulation, and metabolic engineering applications. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 29–66. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Ecological roles of alkaloids. In Modern Alkaloids; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2008; pp. 1–24. [Google Scholar] [CrossRef]
- Figlar, R.B.; Nooteboom, H.P. Notes on Magnoliaceae IV. Blumea Biodivers. Evol. Biogeogr. Plants 2004, 49, 87–100. [Google Scholar] [CrossRef]
- Serna, L.M.; Tuberquia, D.; Velásquez, C.; Rincón, H.; Cogollo, Á. Implementación de Una Estrategia de Conservación Para las Especies de Magnoliaceae en la Jurisdicción de Corantioquia. 2002. Available online: https://www.corantioquia.gov.co/ciadoc/FLORA/AIRNR_CN_3009_2001.pdf (accessed on 10 March 2022).
- Wang, G.; Yang, Z.; Chen, P.; Tan, W.; Lu, C. Seed Dispersal of an Endangered Kmeria septentrionalis by Frugivorous Birds in a Karst Habitat. Pak. J. Zool. 2019, 51, 1195–1198. [Google Scholar] [CrossRef]
- Sánchez-Velásquez, L.; Pineda-López, M.R.; Vásquez-Morales, S.; Avendaño-Yáñez, M. Ecology and conservation of endangered species: The case of Magnolias. In Endangered Species: Threats, Conservation and Future Research; Nova Sciences Publishers Inc.: Hauppauge, NY, USA, 2016; pp. 63–84. [Google Scholar]
- Vásquez-Morales, S.G.; Sánchez-Velásquez, L.R.; Pineda-López, M.R.; Díaz-Fleischer, F.; Flores-Estévez, N.; Viveros-Viveros, H. Moderate anthropogenic disturbance does not affect the Magnolia schiedeana Schltl. Demography: An endangered species from Mexico. Flora 2017, 234, 77–83. [Google Scholar] [CrossRef]
- Tiwari, S.; Chauhan, P.S. Ecological restoration and plant biodiversity. In Bioprospecting of Plant Biodiversity for Industrial Molecules; Wiley: West Sussex, UK, 2021; pp. 91–97. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides in the twenty-first century—Fulfilling their promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef]
- Vásquez-Morales, G.S.; Sánchez-Velásquez, L. Seed ecology and pre-germinative treatments in Magnolia schiedeana Schlecht, an endangered species from Mexico. J. Food Agric. Environ. 2011, 99, 604–608. [Google Scholar]
- Zhao, W.; Zhao, Y.; Li, J.; Zhao, L. Preliminary study of volatile components and function of sarcotesta in Magnolia denudata Desr. Acta Bot. Boreali-Occident. Sin. 2015, 35, 1254–1261. [Google Scholar]
- Fontana, R.; Mattioli, L.B.; Biotti, G.; Budriesi, R.; Gotti, R.; Micucci, M.; Corazza, I.; Marconi, P.; Frosini, M.; Manfredini, S.; et al. Magnolia officinalis L. bark extract and respiratory diseases: From traditional Chinese medicine to western medicine via network target. Phytother. Res. 2023, 1–24. [Google Scholar] [CrossRef]
- Cardullo, N.; Barresi, V.; Muccilli, V.; Spampinato, G.; D’Amico, M.; Filippo-Condorelli, D.; Tringali, C. Synthesis of bisphenol neolignans inspired by Honokiol as antiproliferative agents. Molecules 2020, 25, 733. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, S.; Chen, K.; Ji, L.; Cui, S. Magnolol and 5-fluorouracil synergy inhibition of metastasis of cervical cancer cells by targeting P13K/AKT/mTOR and EMT pathways. Chin. Herb. Med. 2023. [Google Scholar] [CrossRef]
- Sciazza, C.; Cardullo, N.; Pulvirenti, L.; Di Francesco, A.; Muccilli, V. Evaluation of honokiol, magnolol and of a library of new nitrogenated neolignans as pancreatic lipase inhibitors. Bioorganic Chem. 2023, 134, 106455. [Google Scholar] [CrossRef]
Figure 1.
Morphological characteristics of Magnolia. (A) flower of M. pugana. (B) Immature fruit of M. pugana, mature fruit with exposed seeds of (C) M. pugana and (D) M. vovidesii. (E) Seeds of M. vovidesii. (F) Sarcotesta (aril) seed without sarcotesta and seedless fruit of M. vovidesii. (G) Ripening process of fruit with seeds of M. perezfarrerae. (H) Mature fruit with exposed seeds of M. perezfarrerae. Photo credit: Suria Vásquez.
Figure 1.
Morphological characteristics of Magnolia. (A) flower of M. pugana. (B) Immature fruit of M. pugana, mature fruit with exposed seeds of (C) M. pugana and (D) M. vovidesii. (E) Seeds of M. vovidesii. (F) Sarcotesta (aril) seed without sarcotesta and seedless fruit of M. vovidesii. (G) Ripening process of fruit with seeds of M. perezfarrerae. (H) Mature fruit with exposed seeds of M. perezfarrerae. Photo credit: Suria Vásquez.

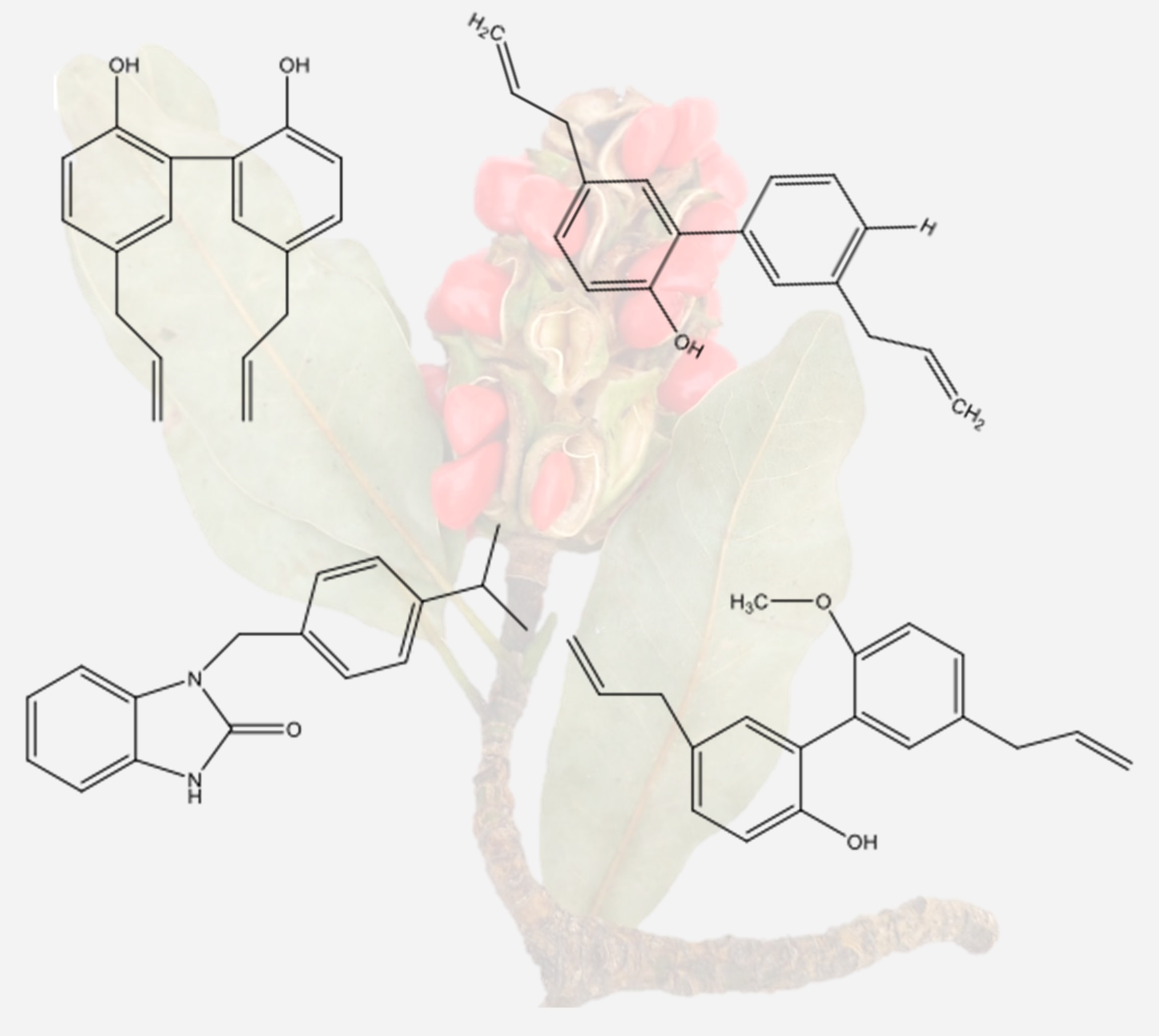
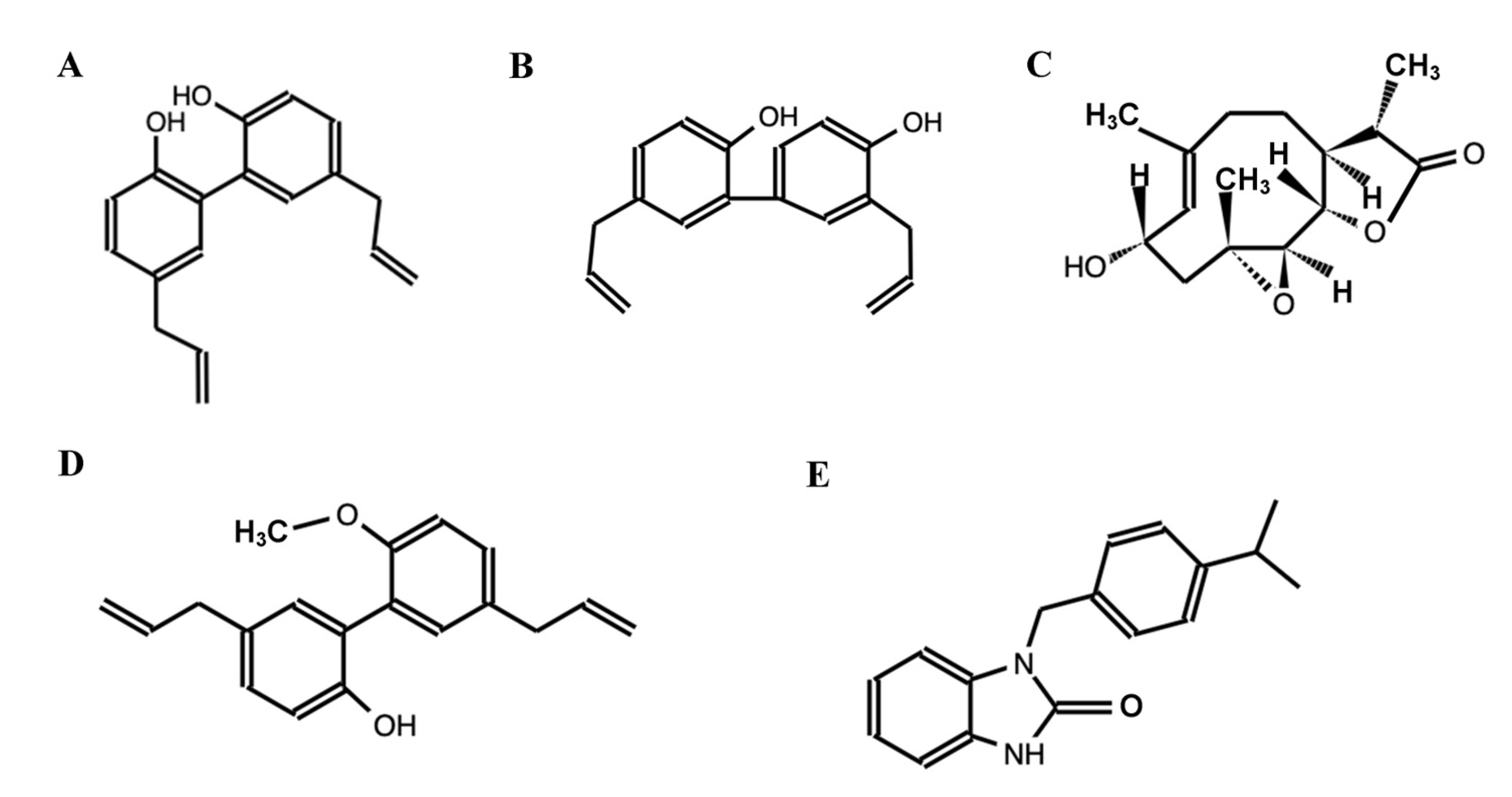
Figure 2.
Compounds obtained from Magnolia grandiflora. (A) 5,5′-diallyl-[1,1′-biphenyl]-2,2′-diol, (B) 3′,5-diallyl-[1,1′-biphenyl]-2,4′-diol, (C) (3S,3aS,8S,9aS,10aR,10bS,E)-8-hydroxy-3,6,9a-trimethyl-3a,4,5,8,9,9a,10a,10b-octahydrooxireno [2′,3′:9,10]cyclodeca [1,2-b]furan-2(3H)-one, (D) 5,5′-diallyl-2′-methoxy-[1,1′-biphenyl]-2-ol, and (E) 1-(4-isopropylbenzyl)-1,3-dihydro-2H-benzo[d]imidazol-2-one (Source: [52]).
Figure 2.
Compounds obtained from Magnolia grandiflora. (A) 5,5′-diallyl-[1,1′-biphenyl]-2,2′-diol, (B) 3′,5-diallyl-[1,1′-biphenyl]-2,4′-diol, (C) (3S,3aS,8S,9aS,10aR,10bS,E)-8-hydroxy-3,6,9a-trimethyl-3a,4,5,8,9,9a,10a,10b-octahydrooxireno [2′,3′:9,10]cyclodeca [1,2-b]furan-2(3H)-one, (D) 5,5′-diallyl-2′-methoxy-[1,1′-biphenyl]-2-ol, and (E) 1-(4-isopropylbenzyl)-1,3-dihydro-2H-benzo[d]imidazol-2-one (Source: [52]).

Figure 3.
Phenylpropanoids from the seeds of Magnolia grandiflora. (A) 1—Honokiol. (A) 3—4-O-metil-honokiol. (B) Magnolol, (C) 5,5′-di-2-propenyl-3-methoxy-[1,1′-biphenyl]-2,2′-diol. (D) 4a,9b-dihydro-8,9b-di-2-propenyl-(4H)-dibenzofuran-3-one, grandifloralignan (Source: [65]).
Figure 3.
Phenylpropanoids from the seeds of Magnolia grandiflora. (A) 1—Honokiol. (A) 3—4-O-metil-honokiol. (B) Magnolol, (C) 5,5′-di-2-propenyl-3-methoxy-[1,1′-biphenyl]-2,2′-diol. (D) 4a,9b-dihydro-8,9b-di-2-propenyl-(4H)-dibenzofuran-3-one, grandifloralignan (Source: [65]).

Figure 4.
Venn diagram with the distribution of 277 chemical compounds in each of the plant structures of 11 Magnolia species, represented by the following colors: fruit (green), peel (red), seed (blue), and sarcotesta (purple), as well as intersection percentages. The compounds were obtained from M. acuminata, M. fraseri, M. grandiflora, M. kobus, M. kwangsiensies, M. obovata, M. ovata, M. pugana, M. tripetala, M. virginiana, and M. vovidesii.
Figure 4.
Venn diagram with the distribution of 277 chemical compounds in each of the plant structures of 11 Magnolia species, represented by the following colors: fruit (green), peel (red), seed (blue), and sarcotesta (purple), as well as intersection percentages. The compounds were obtained from M. acuminata, M. fraseri, M. grandiflora, M. kobus, M. kwangsiensies, M. obovata, M. ovata, M. pugana, M. tripetala, M. virginiana, and M. vovidesii.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biocidal potential of Magnolia terpenoids against insects and other arthropods. Compounds were isolated from M. acuminata, M. fraseri, M. grandiflora, M. kobus, M. kwangsiensies, M. obovata, M. ovata, M. pugana, M. tripetala, M. virginiana, and M. vovidesii. Stages: (A) adult, (L4) fourth instar larva, (L3) third instar larva, and (L) larva.
Table 1.
Biocidal potential of Magnolia terpenoids against insects and other arthropods. Compounds were isolated from M. acuminata, M. fraseri, M. grandiflora, M. kobus, M. kwangsiensies, M. obovata, M. ovata, M. pugana, M. tripetala, M. virginiana, and M. vovidesii. Stages: (A) adult, (L4) fourth instar larva, (L3) third instar larva, and (L) larva.
| Compound | Activity | Species | Orden | Stages | LD50 | Time | Exposition | References |
|---|---|---|---|---|---|---|---|---|
| α-caryophyllene | Insecticide | Helicoverpa armigera | Lepidoptero | L3 | 20.86 µg/mL | 24 h | Intake | [79] |
| α-humulene | Insecticide | Dorymyrmex thoracicus | Hymenoptera | A | 75 μL/L | 48 h | Spraying | [80] |
| α-phellandrene | Insecticide | Sitophilus zemais | Coleoptera | A | 15.61 mg/L | 24 h | Spraying | [81] |
| α-pinene | Insecticide | Lycoriella mali | Diptera | A | 9.85 µL/L air | 24 h | Spraying | [82] |
| Insecticide | Tribolium castaneum | Coleoptera | A | 14.08 mg/L air | 24 h | Spraying | [83] | |
| α-terpinene | Insecticide | T. castaneum | Coleoptera | A | 23.70 μL/L air | 24 h | Spraying | [84] |
| Insecticide | Musca domestica | Diptera | A | 2.41 μL/L | 24 h | Spraying | [85] | |
| α-terpineol | Larvicide | Culex pipiens molestus | Diptera | L4 | 194 mg/L | 24 h | Contact | [81] |
| Insecticide | M. domestica | Diptera | A | 3.74 μL/L | 24 h | Spraying | [85] | |
| Ar-curcumene | Larvicide | An. stephensi | Diptera | L3 | 10.45 µg/mL | 24 h | Contact | [86] |
| Larvicide | Cx. quinquefasciatus | Diptera | L3 | 12.24 µg/mL | 24 h | Contact | [86] | |
| β-caryophyllene | Insecticide | D. thoracicus | Hymenoptera | A | 1.49 μL/L | 48 h | Spraying | [80] |
| Insecticide | T. castaneum | Coleoptera | A | 36.0 μg/adult | 24 h | Contact | [87] | |
| β-myrcene | Insecticide | M. domestica | Diptera | A | 4.95 μL/L | 24 h | Spraying | [85] |
| β-phellandrene | Insecticide | T. castaneum | Coleoptera | A | 22.56 mg/L | 24 h | Spraying | [83] |
| β-pinene | Insecticide | Lasioderma serricorne | Coleoptera | A | 14.66 mg/L | 24 h | Spraying | [83] |
| Insecticide | L. mali | Diptera | A | 11.85 µL/l air | 24 h | Spraying | [82] | |
| Bornyl acetate | Insecticide | Liposcelis bostrychophila | Psocoptera | A | 1.1 mg/L air | 24 h | Spraying | [88] |
| Insecticide | M. domestica | Diptera | A | 4.24 μL/L | 24 h | Spraying | [85] | |
| β-selinene | Insecticide | Drosophila melanogaster | Diptera | A | 0.55 µg/adult | 3 h | Topical application | [89] |
| Caryophyllene oxide | Larvicide | An. anthropophagus | Diptera | L4 | 49.46 mg/L | 24 h | Contact | [90] |
| Insecticide | T. castaneum | Coleoptera | A | 0.00018 mg/cm3 | 24 h | Spraying | [91] | |
| δ-cadinene | Larvicide | Anopheles stephensi | Diptera | L3 | 8.23 µg/mL | 24 h | Contact | [92] |
| Larvicide | Aedes aegypti | Diptera | L3 | 9.03 µg/mL | 24 h | Contact | [92] | |
| E-nerolidol | Insecticide | Metopolophium dirhodum | Hemiptera | A | 3.5 mL/L | 48 h | Contact | [93] |
| γ-terpinene | Insecticide | Phthorimaea operculella | Lepidoptera | A | 5.98 mg/L air | 24 h | Spraying | [94] |
| Guaiol | Insecticide | M. domestica | Diptera | A | 16.9 µL/L | 48 h | Spraying | [95] |
| Larvicide | Plutella xylostella | Lepidoptera | L3 | 8.9 mg/larva | 12 h | Contact | [95] | |
| Limonene | Insecticide | T. castaneum | Coleoptera | A | 6.79 mg/L | 24 h | Spraying | [83] |
| Insecticide | M. domestica | Diptera | A | 3.22 μL/L | 24 h | Spraying | [85] | |
| Linalool | Insecticide | Sitophilus zeamais | Coleoptera | A | 10.46 mg/L | 24 h | Spraying | [96] |
| Larvicide | Cx. pipiens molestus | Diptera | L4 | 193 mg/L | 24 h | Contact | [97] | |
| p-cymene | Insecticide | T. castaneum | Coleoptera | A | 27.01 μL/l air | 24 h | Contact | [84] |
| Insecticide | M. domestica | Diptera | A | 0.77 μL/L | 24 h | Spraying | [85] |
Table 2.
Biocidal potential of Magnolia phenols against insects and other arthropods. Compounds were isolated from M. acuminata, M. fraseri, M. grandiflora, M. kobus, M. obovata, M. ovata, M. tripetala, M. virginiana, and M. vovidesii. Stages: (A) adult, (L4) fourth instar larva, (L3) third instar larva, and (L) larva.
Table 2.
Biocidal potential of Magnolia phenols against insects and other arthropods. Compounds were isolated from M. acuminata, M. fraseri, M. grandiflora, M. kobus, M. obovata, M. ovata, M. tripetala, M. virginiana, and M. vovidesii. Stages: (A) adult, (L4) fourth instar larva, (L3) third instar larva, and (L) larva.
| Compound | Activity | Species | Orden | Stages | LD50 | Time | Exposition | References |
|---|---|---|---|---|---|---|---|---|
| 2,4-di-tert-butylphenol | Acaricide | Tetranychus cinnabarinus | Trombidiformes | A | 7.61 µM | 24 h | Spraying | [103] |
| Estragol | Larvicide | Spodoptera frugiperda | Lepidoptera | A | 0.92 mg mL | 24 h | Intake | [104] |
| Insecticide | Sitophilus zeamais | Coleoptera | A | 14.10 mg/L | 24 h | Spraying | [87] | |
| Eugenol | Insecticide | Tribolium castaneum | Coleoptera | A | 1 μg/kg | 24 h | Contact | [105] |
| Insecticide | Callosobruchus maculatus | Coleoptera | A | 24.8 μL/L | 24 h | Spraying | [106] | |
| Honokiol | Larvicide | Aedes albopictus | Diptera | L3 | 6.13 mg/L | 24 h | Contact | [107] |
| Larvicide | Anopheles sinensis | Diptera | L3 | 7.37 mg/L | 24 h | Contact | [107] | |
| Insecticide | Nilaparvata lugens | Hemiptera | A | 0.324 mM | 48 h | Topical application | [108] | |
| Licarin A | Larvicide | S. litura | Lepidoptera | L | 0.20% m/m | 7 d | Intake | [109] |
| Magnolol | Insecticide | N. lugens | Hemiptera | A | 0.137 mM | 48 h | Topical application | [108] |
| Larvicide | Culex pipiens pallens | Diptera | L3 | 26 mg/L | 24 h | Contact | [107] | |
| Protochatecuic acid | Insecticide | Ae. aegypti | Diptera | A | 1.25 µg/mg | 24 h | Contact | [110] |
| Quercetin | Larvicide | S. litura | Lepidoptera | L4 | 10.88 ppm | 24 h | Intake | [111] |
| Larvicide | Pectinophora gossypiella | Lepidoptera | L | 0.2% | Until pupae maturation | Intake | [112] | |
| Rutin | Insecticide | Oedaleus asiaticus | Orthoptera | A | 763.7 mg/L | 7 days | Intake | [113] |
| Scopoletin | Larvicide | Spilarctia obliqua | Lepidoptera | L4 | 20.9 μg/g | 24 h | Intake | [114] |
| Syringin | Anti-nutrients | S. granarius | Coleoptera | A | 134.4 μL/L | 5 d | Intake | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hernandez-Rocha, J.V.; Vásquez-Morales, S.G. The Potential of Magnolia spp. in the Production of Alternative Pest Control Substances. Molecules 2023, 28, 4681. https://doi.org/10.3390/molecules28124681
AMA Style
Hernandez-Rocha JV, Vásquez-Morales SG. The Potential of Magnolia spp. in the Production of Alternative Pest Control Substances. Molecules. 2023; 28(12):4681. https://doi.org/10.3390/molecules28124681
Chicago/Turabian StyleHernandez-Rocha, Juana Valeria, and Suria Gisela Vásquez-Morales. 2023. "The Potential of Magnolia spp. in the Production of Alternative Pest Control Substances" Molecules 28, no. 12: 4681. https://doi.org/10.3390/molecules28124681