
Long-Chain Molecules with Agro-Bioactivities and Their Applications


College of Science, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Molecules 2023, 28(15), 5880; https://doi.org/10.3390/molecules28155880
Submission received: 10 July 2023
/
Revised: 31 July 2023
/
Accepted: 31 July 2023
/
Published: 4 August 2023
Abstract
:Long-chain molecules play a vital role in agricultural production and find extensive use as fungicides, insecticides, acaricides, herbicides, and plant growth regulators. This review article specifically addresses the agricultural biological activities and applications of long-chain molecules. The utilization of long-chain molecules in the development of pesticides is an appealing avenue for designing novel pesticide compounds. By offering valuable insights, this article serves as a useful reference for the design of new long-chain molecules for pesticide applications.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
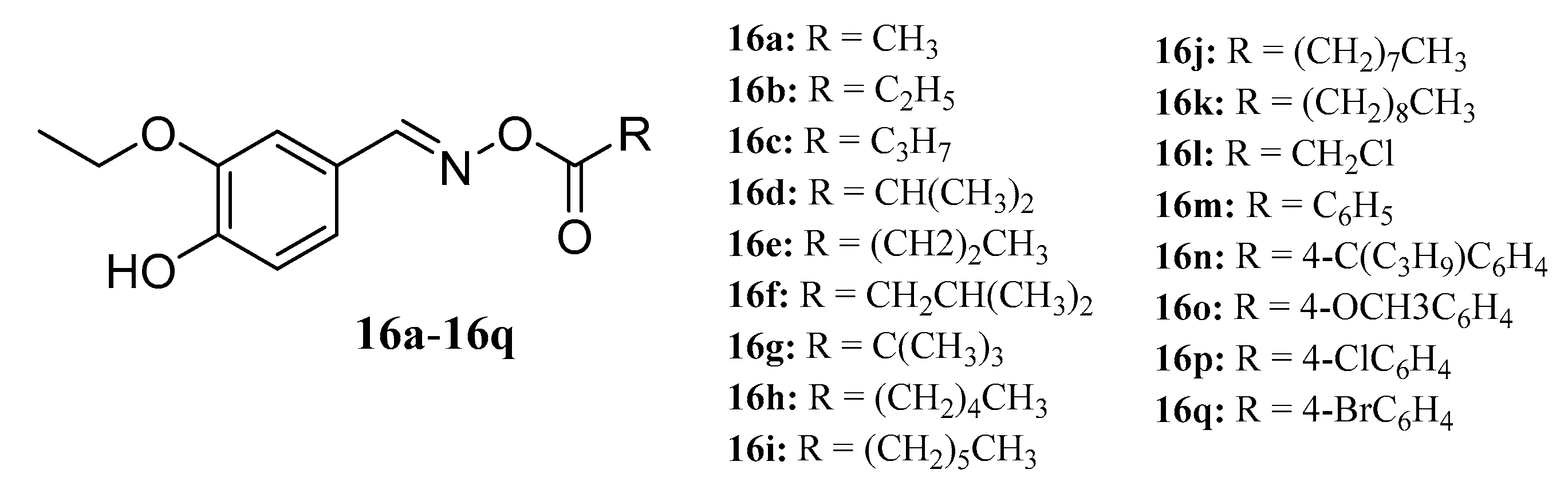{kind=link}
{kind=link}
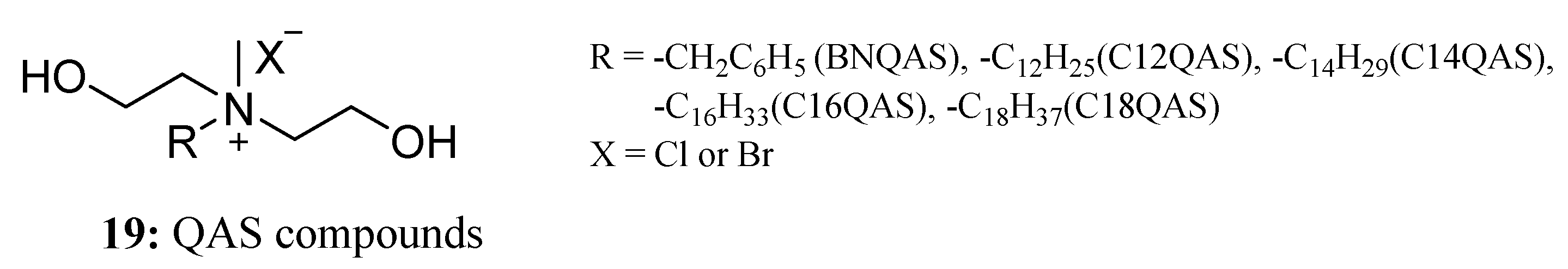{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Plant pests, diseases, and weeds are significant challenges in agriculture, as they can lead to decreased crop yields and pose threats to food safety. While resistant crop varieties and other biological strategies have been employed to combat these issues, pesticides remain the primary means of control [1,2,3]. Among the various types of pesticides, long-chain molecules—both natural and synthetic—are widely used for their efficacy as fungicides, insecticides, acaricides, herbicides, and plant growth regulators (Figure 1). For the purposes of this review, we define long-chain molecules as compounds with five or more atoms (C, O, and S) in their chains. We exclude long-chain molecules containing nitrogen atoms due to their unique effects on the biological activity of the compounds, such as their tendency to form hydrogen bonds. In this article, we primarily focus on the agricultural biological activities and applications of long-chain molecules.
2. Long-Chain Molecules Exist Widely in Nature
Long-chain molecules are prevalent in nature and are involved in vital physiological processes in living organisms. These molecules exhibit diverse biosynthetic pathways and possess remarkable biological properties. This section provides a concise overview of various natural long-chain molecules, focusing on their biosynthetic pathways and biological activities.
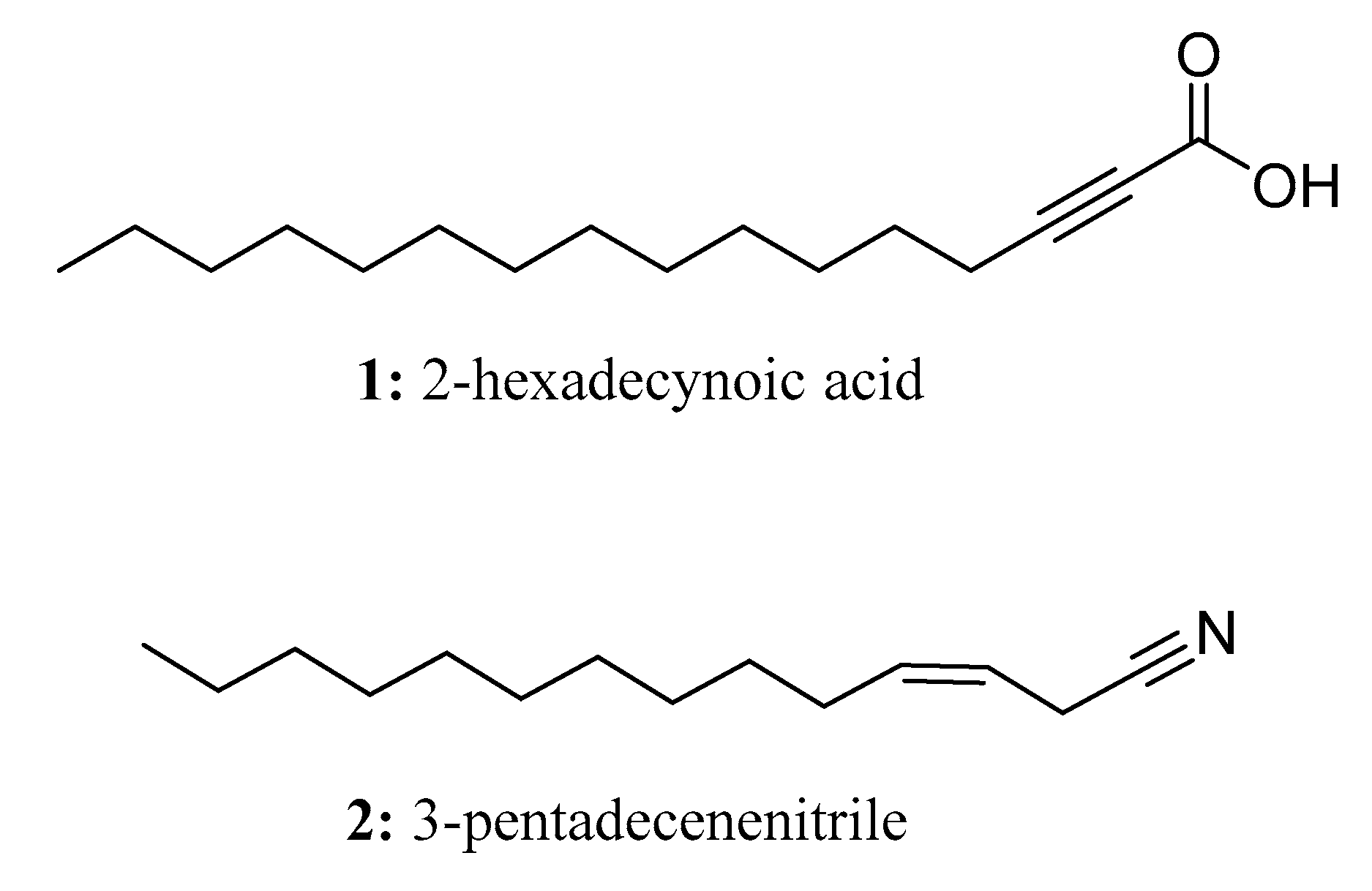
In nature, fatty acids (FAs) have wide-ranging functions and are found in abundance. They play important roles in energy storage, structural organization, and signaling processes [4,5]. Plants utilize a complex enzymatic system called the plastid-localized fatty acid synthase complex to synthesize fatty acids. This complex acts on acyl-intermediates that are attached to acyl carrier proteins (ACPs). The synthesis process starts with acetyl-CoA, and carbon dioxide is released through the condensation reaction between acetyl-CoA and malonyl-ACP. Subsequent reactions lead to the formation of carbon-expanded, saturated hydrocarbon chains from the intermediate 3-ketone. As a result of the fatty acid synthase cycles, saturated C16 and C18 fatty acids are produced [6,7]. It is noteworthy that many long-chain fatty acids exhibit significant biological activity. For instance, scleropyric acid demonstrates anti-plasmodial activity, 2-hexadecynoic acid (1) (Figure 2) and 2-alkynoic fatty acid have been found to exhibit antibacterial activity against mycobacterium tuberculosis, and linoleic acid exhibits antifungal activity [8,9,10]. Furthermore, oleic acid has been shown to possess antifungal effects against Rhizoctonia solani, Pythium ultimum, Pyrenophora avenae, and Crinipellis perniciosa [5,11].
Natural products containing long-chain aliphatic nitriles are produced by two types of bacteria: Gram-positive Micromonospora echinospora and Gram-negative Pseudomonas veronii [12]. Some of these nitriles have antimicrobial activity. For example, 3-pentadecenenitrile (2) (Figure 2) has been found to be effective against bacteria such as Bacillus subtilis, Micrococcus luteus, and particularly Staphylococcus aureus, including drug-resistant strains. Nitriles are synthesized from fatty acids, which are first converted into amides and then dehydrated. The process of chain elongation and dehydration is variable during fatty acid biosynthesis, resulting in the formation of unbranched saturated or unsaturated nitriles with an ω-7 double bond. For instance, (Z)-11-octadeconitrile is an example of an unbranched nitrile, while (Z)-13-methyl-tetracylcarbon-3-nitrile is an example of a methyl-branched unsaturated nitrile with the double bond located at C-3.
Yaoshanenolides A (3a) and B (3b) are novel tricyclic spirolactones isolated from the bark of Machilus yaoshansis [13]. These compounds contain long linear chains and have exhibited non-selective cytotoxic activity against various human cancer cell lines. Co-occurring obtusilactone A and/or isoobtusilactone A and dihydroisoobtusilactone are believed to be biosynthetic precursors of yaoshanenolides A and B, both of which were isolated from the Machilus genus. The biosynthesis of yaoshanenolides A and B involves an enzyme-catalyzed Diels–Alder [4 + 2] cycloaddition between a molecule of the precursors and a molecule of β-phellandrene, which has also been found in Machilus genus. This is followed by either simultaneous or sequential allylic hydroxy rearrangement [13,14,15] (Figure 3).
Isoprenoid quinones, which contain a quinone head group and a poly-pentenyl tail of varying length play a crucial role in bioenergetics as electron and proton carriers in the respiratory chain of most organisms [16]. They can be categorized into naphthoquinones and benzoquinones based on the characteristics of their head groups and midpoint redox potentials. Menaquinone (MK, 4d) belongs to the naphthoquinones, while ubiquinone (UQ, 4a), plastoquinone (PQ, 4b), and rhodoquinone (RQ, 4c) belong to the benzoquinones [17] (Figure 4). The biosynthetic pathway of isoprenoid quinones varies across species. For example, in Escherichia coli, the biosynthesis of UQ8 (ubiquinone 8) initiates with 4-hydroxybenzoic acid (4-HB) and involves twelve proteins (UbiA to UbiK and UbiX). First, UbiC removes pyruvate from chorismite to form 4-HB [18]. Then, the membrane-bound UbiA prenylates 4-HB using octaprenyl diphosphate as a precursor for the side chain. After prenylation, the UbiD–UbiX system decarboxylates 4-HB [19]. UQ8 biosynthesis involves two O-methylation reactions catalyzed by the S-adenosyl-L-methionine-dependent UbiG protein [20,21]. In this pathway, UbiE catal00yzes the C-methylation reaction, while three class A flavoprotein monooxygenases (FMOs)—UbiH, UbiI, and UbiF—catalyze the hydroxylation of carbon atoms C-1, C-5, and C-6 on the aromatic rings, respectively [22,23].
3. Long-Chain Molecules with Agro-Bioactivities
Long-chain molecules exhibit a diverse assay of biological activities and are frequently utilized as fungicides, insecticides, herbicides, and plant growth regulators in agricultural settings. Despite sharing the common feature of a long-chain molecular structure, these compounds operate through a multitude of different mechanisms. Among these fungicides, compounds can be classified based on their mechanism of action, which includes respiratory chain inhibitors, 14α-demethylation inhibitors, and DNA/RNA synthesis inhibitors. Insecticides, on the other hand, include respiratory chain inhibitors, acetylcholinesterase inhibitors, and sodium channel inhibitors. Herbicides, which are made up of long-chain molecules, exhibit different mechanisms of action. For example, acetanilide compounds inhibit the biosynthesis of long-chain fatty acids in plants. Additionally, some compounds inhibit 4-HPPD, resulting in plant albinism and eventually death. In conclusion, long-chain pesticide compounds are not only widely used but also complex. In addition to commercial pesticides, there are many natural products or long-chain derivatives with pesticide activity. In this section, we provide some examples of such compounds.
3.1. Phytopathogenic Fungicides Containing a Long Chain
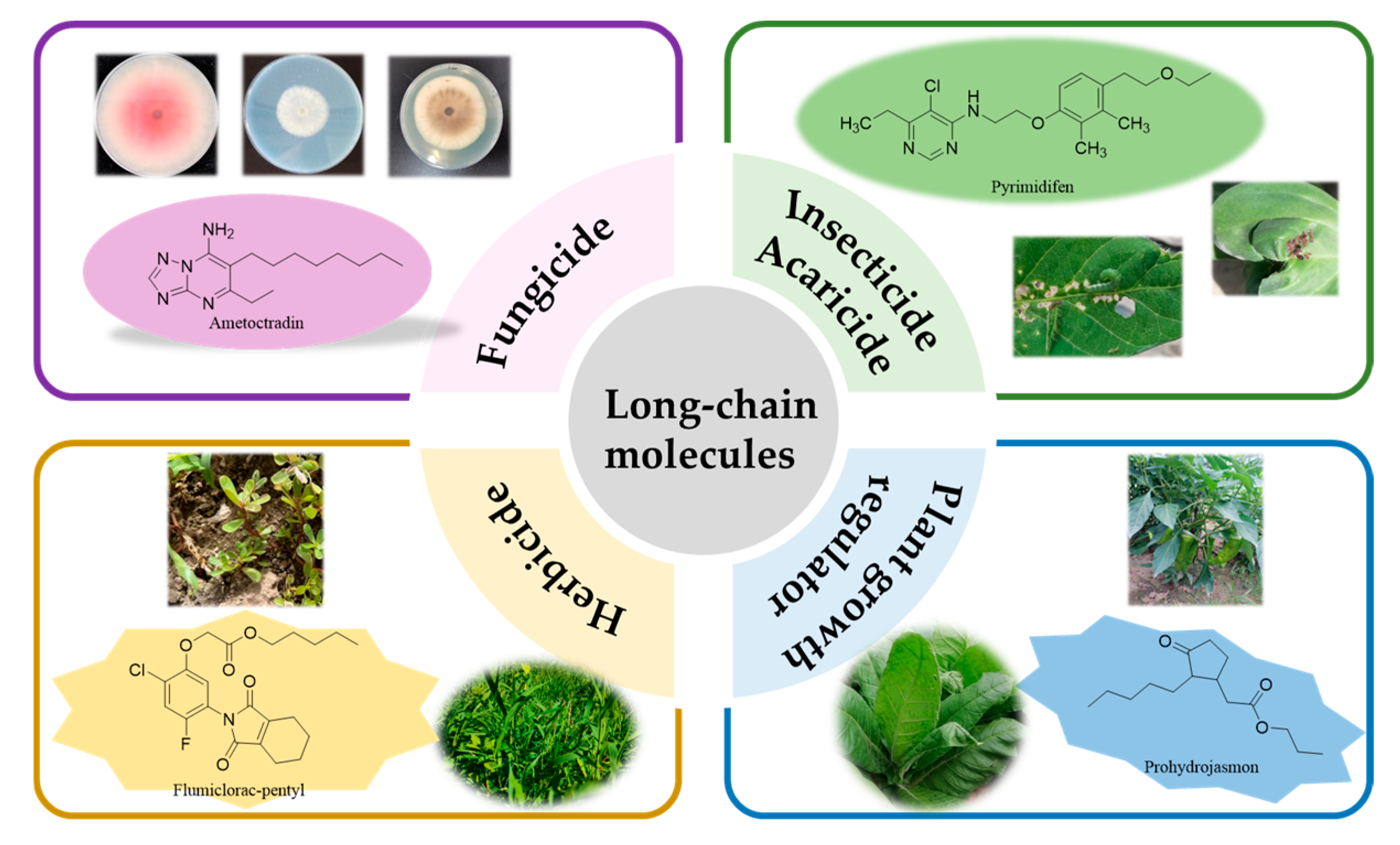
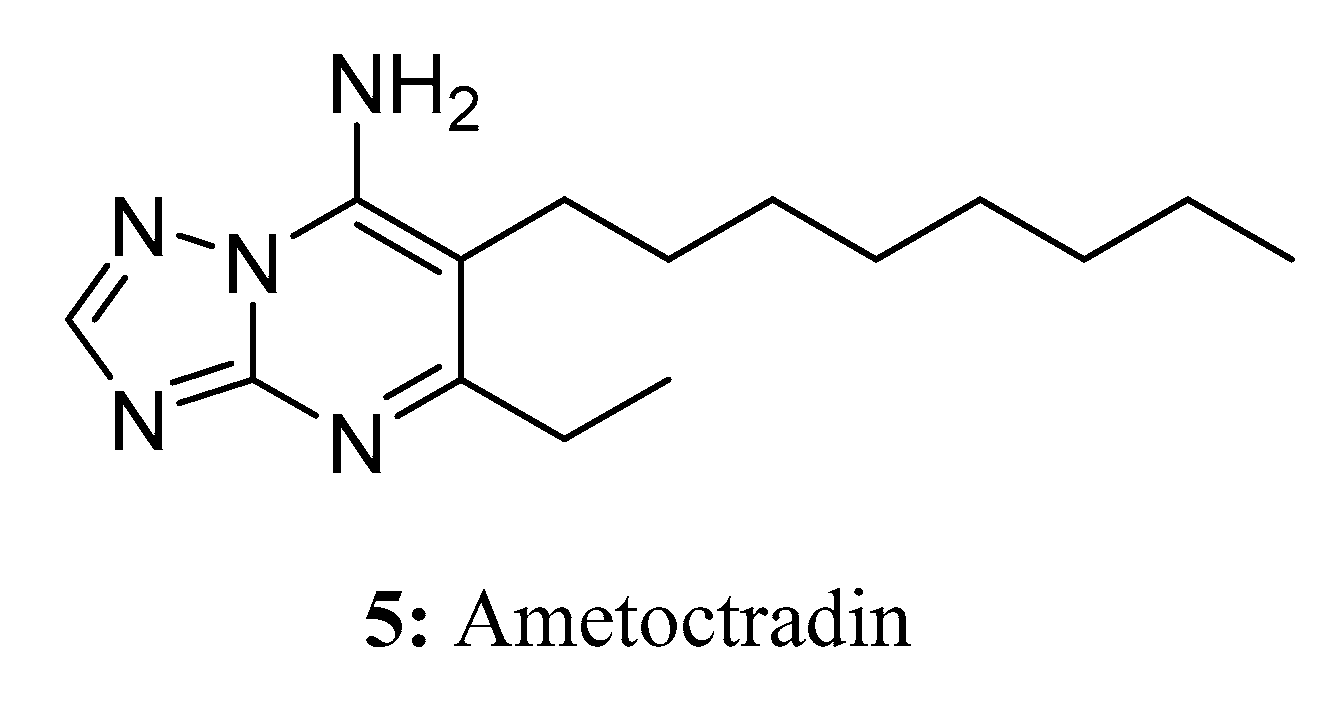
Ametoctradin (5) [24,25] is a novel oomycete inhibitor with a unique chemical scaffold. Spray applications of ametoctradin, which belongs to the triazolopyrimidine fungicide class, can bring extensive benefits to various special crops. Field trials have demonstrated that ametoctradin is highly selective and effective in preventing late blight and downy mildew (Figure 5).
On the molecular level, ametoctradin functions as an inhibitor of the respiratory bc1 complex found in oomycete crop pathogens. It achieves this by binding to either the Qo-stigmatellin subsite or the Qi-site of cytochrome b within complex III of the respiratory electron transport chain, thereby impeding electron transfer. As a consequence of its action, the consumption of oxygen and the levels of intracellular ATP diminish rapidly after ametoctradin treatment in oomycete pathogens. Due to its ability to hinder multiple energy-intensive processes, such as zoospore formation and release, zoosporangia release, germination, and motility, ametoctradin has been designated as a fungicide belonging to the category of QoSI (quinone outside inhibitors) [24,26,27].
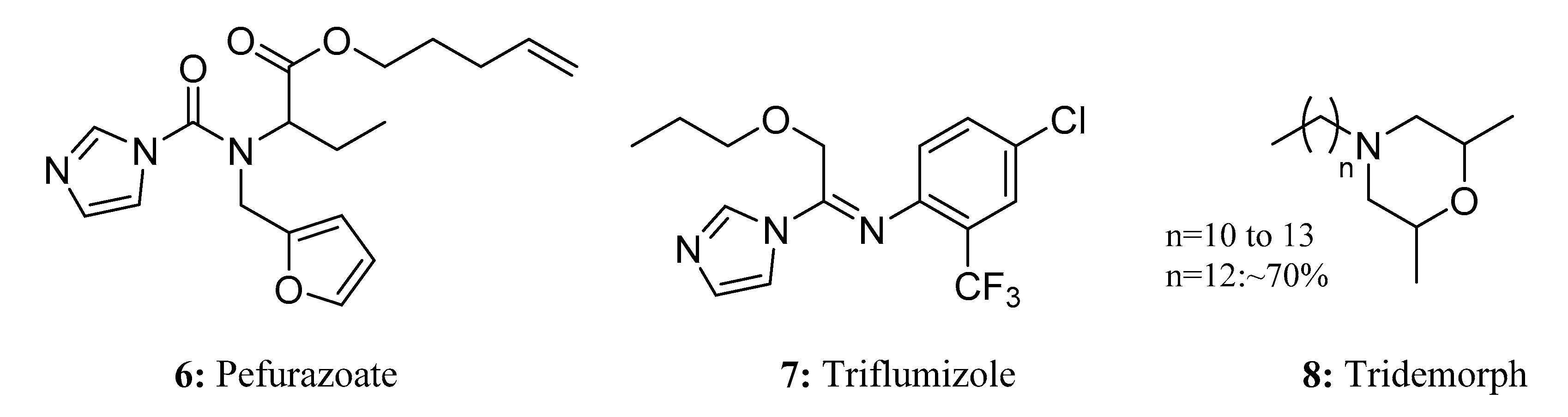
Pefurazoate (6), a novel imidazole compound, is a rice-seed disinfectant. As a fungitoxic agent, it is effective against seed-borne pathogenic fungi such as Gibbeyella fujikuyoi, Pyricularia oryzae, and Cochliobolus miyabeanus [28]. Similar to other azole fungicides, pefurazoate acts as by inhibiting the 14α-demethylation of 24-methylene dihydro lanosterol [28,29]. In addition to its sterilization and disease-control effects, pefurazoate also has a positive effect on promoting seed germination and seedling growth, thereby promoting healthy rice plant development (Figure 6).
Triflumizole (7) is one of the imidazole fungicides [30]. This fungicide is known for its high efficiency, low toxicity, and low residual properties. It contains fluorimidazole heterocyclic and exhibits both protective and therapeutic effects. Similar to pefurazoate, triflumizole works by inhibiting ergosterol biosynthesis through interference with the demethylation of the ergosterol skeleton at C-14 [31]. Triflumizole is widely used in various fruit and vegetable production areas, such as those for rice, sweet corn, apples, grapes, pears, and cherries, to control the growth and spread of powdery mildew [30,32] (Figure 6).
Tridemorph (8) belongs to the morpholine fungicides [33]; it is a systemic fungicide, as it is taken up by the leaves and roots. It acts as an inhibitor of ergosterol biosynthesis, especially through the inhibition of sterol reduction (sterol-Δ14-reductase) and isomerization (Δ8 to Δ7-isomerase) [34]. This fungicide has a broad spectrum of activity and is commonly used for the prevention and control of grain powdery mildew and Mycosphaerella spp. in bananas [34,35] (Figure 6).
Dodine (9), commonly used as a protective fungicide, contains a guanidyl headgroup and a dodecyl tail [36]. A variety of major mold diseases can be controlled by dodine on fruit trees, vegetables, nuts, ornamental plants, and shade trees [37]. As a dodecyl derivative of guanidinium salt, dodine differs from guanidinium salt because of the presence of a long hydrocarbon chain [38]. It attaches readily to the surfaces of negatively charged microorganisms and can penetrate their membranes, thereby damaging cellular structures [37] (Figure 7).
Seboctylamine (10) is a kind of high-efficiency, low-toxicity, and spectral fungicide developed in China [39]. Seboctylamine exhibits a pronounced killing and inhibitory effect on various types of fungi, bacteria, and viruses. Moreover, it demonstrates a potent inhibition effect specifically on the growth of mycelium and the germination of spores [40]. The mechanism of seboctylamine can produce ionization in aqueous solutions; the hydrophilic group part is strongly electropositive, while the adsorption usually contains all kinds of negatively charged bacteria and viruses. It inhibits the multiplication of bacteria and viruses by causing bacterial proteins to coagulate and the polymer to form a film that blocks the ion channels of these microorganisms, causing them to suffocate and die immediately, thus achieving optimal bactericidal effects [39,40,41] (Figure 7).
Iminoctadine (11) is a non-systemic aliphatic nitrogen contact fungicide that exhibits preventive effects [42]. It is highly effective against a wide range of fungal diseases caused by ascomycetes, including gray mold, powdery mildew, sclerotinia, and others [43,44]. Iminoctadine affects the biosynthesis of fungal lipids, disrupts the function of fungal cell membranes, and suppresses the formation of appressorium and mycelium through inhibition of spore germination. Iminoctadine tris, on the other hand, is a compound resulting from the combination of iminoctadine with three alkylbenzene sulfonates. Likewise, iminoctadine triacetate refers to the triacetate salt of iminoctadine. This fungicide works by disrupting the membrane function of the pathogen’s cells [45] (Figure 8).
Copper octanoate (12) [46,47] is a kind of saturated fatty acid that combines copper ions and caprylic acid, which has the action of contact. Copper octanoate can be used as a fungicide and bactericide for leaf surfaces to control a variety of plant diseases in various crops and landscape plants (Figure 8).
Octhilinone (13) is a DNA/RNA synthesis inhibitor that is effective against apple and pear canker (Nectria galligena) as well as other bacterial and fungal diseases (Ceratocystis spp.) of top fruits and citrus fruits [48]. Octhilinone is also one of the most frequently used biocides in construction materials as an in-can or film preservative [49]. In addition to being used as a preservative in many industrial applications, including cooling lubricants, sealants, and adhesives, 2-octyl-2H-isothiazol-3-one (OIT, 14) is a well-known indoor surface fungicide [50] with a strong ability to control mold. Debacarb (15) is used via injection for the control of various fungal diseases of trees [51] (Figure 9).
Ahluwalia et al. synthesized a series of oxime esters (16a−16q) with 3-ethoxy-4-benzaldehyde oxime in the presence of triethylamine and acid chloride. The antibacterial activity of three plant pathogens (Rhizoctonia bataticola, Fusarium udum, and Alternaria porii) was evaluated in vitro. Compounds containing medium-long alkyl chains showed higher activity than those containing long alkyl chains [52] (Figure 10).
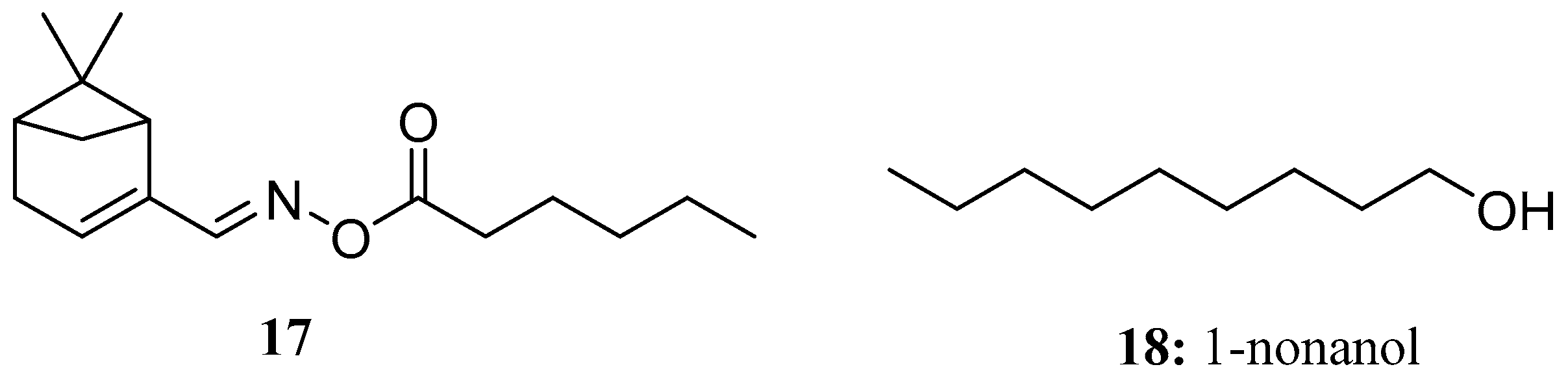
Compound 17 is a novel myrtenal oxime ester molecule designed and synthesized by Lin et al. using ketol-acid reductoisomerase as the target enzyme. The results showed that the growth inhibition rate of compound 17 on Brassica campestris was 64.2% and that on Echinochloa crusgalli L. was 81.8% at 100 μg/mL. Meanwhile, compound 17 exhibited certain in vitro antifungal activity against all tested fungi, such as Fusarium oxysporum f. cummerinum, Physalospora piricola, Cercospora arachidicola, and Gibberella zeae [53] (Figure 11).
1-Nonanol (18), which is the main component of cereal volatiles, has potential antifungal activity against Aspergillus flavus. The damaging effect of 1-nonanol on the growth of Aspergillus flavus is manifested by intracellular electrolyte leakage; reduced succinate dehydrogenase, mitochondrial dehydrogenase, and ATPase activities; and reactive oxygen species accumulation. We speculated that 1-nonanol can damage the membrane integrity and mitochondrial function of Aspergillus flavus and may lead to the apoptosis of Aspergillus flavus [54] (Figure 11).
Liu et al. prepared five quaternary ammonium salt (QAS) compounds (19) (R = -benzyl (chloride, BNQAS), -dodecyl (C12QAS), -tetradecyl (C14QAS), hexadecyl (C16QAS), and -octadecyl (C18QAS)), and their antifungal properties were tested (Figure 12). The results indicated that C12QAS is effective against several apple fungi, including Cytospora mandshuria, Botryosphaeria ribis, Physalospora piricola, and Glomerella Cingulata. The antifungal activity of QAS is mainly related to the introduction of a long-chain alkyl in the molecule. The long alkyl chains in QAS can change the geometry of QAS, which can help the two flexible hydroxyl groups of QAS cross the cell membrane and enter the cell to cause damage [55].
Muhizi et al. synthesized different N-alkyl-β-d-glucosylamines (20a−20i) and evaluated their antifungal activities. The results showed that these compounds have different biological activities and that the antifungal activity of glucosylamines increases with the length of the alkyl chain. DoGPA was more bioactive against all target strains than other N-alkyl glucosamines and could be used to inhibit Fusarium proliferatum [56] (Figure 13).
3.2. Insecticides/Acaricides Containing a Long Chain
Dinocap is a mixture of six isomers of dinitrooctylphenyl crotonate (2,4-DNOPC), including ortho and para methylheptyl, ethylhexyl, and propylpentyl crotonate isomers [57]. It is a contact fungicide and acaricide that has been used to control mites in apple crops and powdery mildew in orchard fruits, vegetables, and ornamental crops [58]. The new meptyldinocap (21) is an improved version of the single 2,4-DNOPC methylheptyl isomer with a better toxicological profile compared to the old dinocap. It is a non-inhalant acaricide and a powdery mildew (Erysiphe necator) fungicide that shows both protectant and post-infection activity [57] (Figure 14).
Sulfuramid (22) is a potential bait agent that belongs to a novel class of insecticides known as fluoroaliphatic sulfonamides [59]. It acts as a delayed-acting insecticide by uncoupling oxidative phosphorylation and exhibits chronic toxicity. When insects consume sulfuramid, it acts as a stomach poison, inhibiting their energy metabolism. Sulfuramid has proven effective in controlling leaf-cutting ants (Atta spp. and Acromyrmex spp.) in eucalyptus and pine plantations [60] (Figure 14).
Acequinocyl (23) is a commercially available acaricide that belongs to the naphthoquinone analog group. Acequinocyl acts by inhibiting mitochondrial respiratory complex III. Its deacetylated metabolite contains free hydroxyl groups, which function as ubiquitin analogs, and it is a powerful inhibitor of the Qo center [61]. Acequinocyl is commonly used to control various herbivorous mites, including Tetranychus urticae [62]. Acequinocyl exhibits a stronger killing effect on immature spider mites, while its toxicity to mammals is relatively low. Additionally, it has a shorter persistence in the environment [63] (Figure 15).
Piericidins are a group of compounds isolated from actinomyces [64], particularly from the genus Streptomyces. One of these compounds, called Piericidin A (24), is a natural insecticide that is highly active against lepidoptera larvae. It was first isolated in the 1960s from Streptomyces mobaraensis [65,66]. Piericidin A has a similar structure to coenzyme Q, with a head group that resembles ubiquinone and a nitrogen atom in place of one of the carbonyl groups. The hydrophobic tail of Piericidin A contains an isoprenoid group with a hydroxyl group at its end [65]. Piericidin A acts as an antagonist to coenzyme Q, inhibiting the activity of complex I [64] (Figure 15).
Pyrimidifen (25) is a potent insecticide that has proven effective in combatting various mite species that pose a threat to fruits and vegetables. It exhibits efficacy against all life stages of spider mites and is also capable of managing the population of diamondback moths (Plutella xylostella L.). Pyrimidifen operates through the same mechanism of action as other pyrimidine acaricides: inhibiting the electron transfer on complex Ι [67,68] (Figure 15).
The main structural feature of ACG (26) is an alkyl chain containing a methyl γ-lactone ring at the end, 0~3 tetrahydrofuran rings on the alkyl chain, and a certain number of oxygen-containing functional groups (such as hydroxyl, acetoxy, ketone, epoxide, etc.) or a double bond; the number of carbon atoms is 35 or 37 [69]. According to the number and spatial arrangement of THF in ACG, ACG can be divided into various types, such as mono-THF acetogenins, adjacent bis-THF acetogenins, non-adjacent bis-THF acetogenins, non-THF acetogenins, and so on. Due to the strong inhibitory effect of ACG on the mitochondrial respiratory chain complex Ι of cells, most ACGs have good insecticidal effects, and adjacent bis-THF acetogenins have the most obvious insecticidal effect [70,71] (Figure 15).Demeton-S-methyl (27) is a widely used systemic organophosphorus insecticide with a certain toxicity that is used to control aphids and red spiders in various crops. Demeton-S-methyl irreversibly inhibits cholinesterases by phosphorylating their catalytic serine [72,73] (Figure 16).
Disulfoton (28), a kind of organophosphorus pesticide, is a systemic insecticide and acaricide [74,75,76,77] that is normally applied as granules with the seed at sowing [78]. Disulfoton has been extensively used across a wide range of crops to manage sap-feeding insects, including aphids, mites, and thrips [76]. Its mode of action involves inhibiting the enzyme acetylcholinesterase (AChE) [77,79], which leads to the accumulation of acetylcholine in nerve endings in both the peripheral and central nervous systems. Consequently, disulfoton poses a high level of toxicity to humans [80,81,82]. The World Health Organization (WHO) has classified disulfoton as “extremely hazardous” (Figure 16).
AMTSn (29), a small molecule containing methanethiosulfonate, was developed by Pang as an inhibitor of AChE activity. It was found to irreversibly inhibit 99% of the AChE derived from the green insect aphid (Schizaphis graminum) at a concentration of 6.0 mM. Interestingly, there was no measurable inhibition of human AChE. Reactivation studies using β-mercaptoethanol confirmed that this irreversible inhibition occurs by binding AMTSn to a unique Cys residue found at the active site of AChE in aphids and other insects but not in mammals. This discovery suggests that targeting this specific Cys residue could potentially be used to develop insect-selective insecticides [83,84,85,86] (Figure 16).
Empenthrin (30) is a highly volatile synthetic pyrethroid with potent insecticidal activity against houseflies and textile pests and a low toxicity to mammals [87,88]. The efficacy of empenthrin is primarily attributed to the unique structure of its alcoholic part and its high vapor pressure. This pyrethroid is formed through the combination of synthetic ethynyl alcohol and chrysanthemic acid, in part mimicking the rethrolone skeleton found in natural pyrethroids [89] (Figure 17).
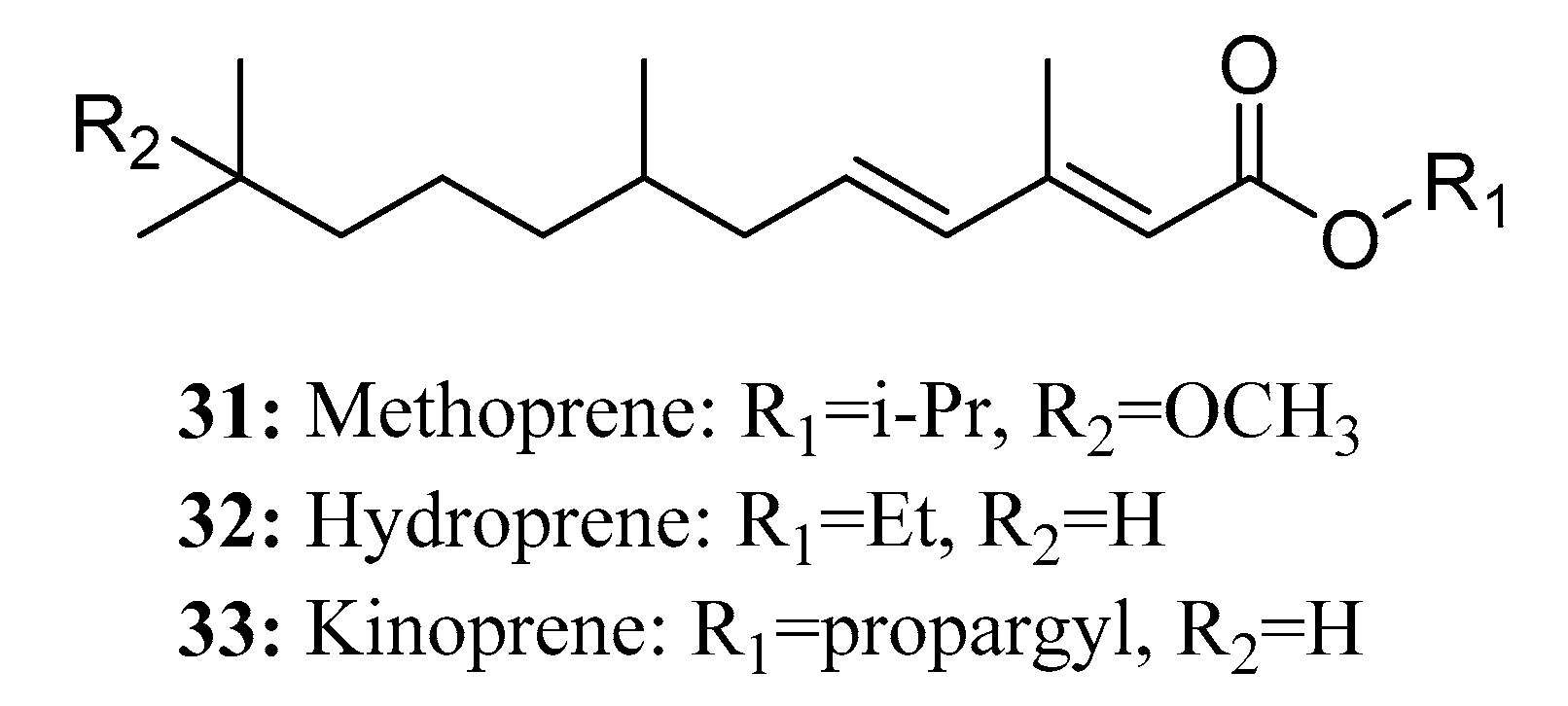
Juvenile hormone (JH) analogs, namely methoprene (31), hydroprene (32), and kinoprene (33), are widely used in pest control due to their non-toxicity toward vertebrates and other non-target organisms [90] as well as their rapid degradation after application (Figure 18).
Methoprene, the first synthetic insect juvenile hormone analog, is known for its efficacy in pest control. In particular, S-methoprene is a highly effective form of this compound. Acting as a synthetic insect juvenile hormone analog, S-methoprene operates through a mechanism similar to that of natural juvenile hormones. It interferes with the normal growth and development process of insects, causing diapause to occur, and disrupts the development of larvae to adults, thereby inhibiting the reproduction of pests [91,92]. Methoprene has been proven to be particularly effective against harmful species of Diptera and Coleoptera [93,94]. Furthermore, it has been applied in the prevention and control of ants and animal fleas [95,96].
Hydroprene is effective against certain lepidopteran pests by imitating the actions of naturally occurring JHs [97]. Application of hydroprene at the larval stage limits and suppresses normal larval development. Larvae treated with hydroprene either fail to reach adulthood or become abnormally sterile adults [98]. As an active ingredient, Kinoprene exhibits particularly high activity against Homopteran insects.
In addition, Mori et al. prepared and bioassayed twenty-six juvenile hormone analogs with varying molecular chain lengths using allatectomized fourth instar larvae of Bombyx mori L. Their findings revealed that among methyl or ethyl esters, the optimal chain length for the high juvenile hormone activity on the silkworm is 17 atoms [99]. Additionally, Kisida et al. reported the discovery of thiolcarbamates and their derivatives, which showed strong larvicidal activities against the larvae of C. pipiens and A. aegypti, similar to JHs [100].
Oleic acid (34) can destroy insect cuticles, and its potassium and sodium salts can be used as an insecticide against soft-bodied pests (aphids, whiteflies, and spider mites) on vegetables, fruits, and ornamentals, as well as a fungicide against powdery mildew, while oleic acid can also be used for weed control on uncultivated land [101,102,103]. In addition, oleic esters are also widely used in the field of pesticides. For example, methyl oleate is usually used as emulsifier and auxiliary for pesticides, which can increase the degree of dispersion of the agent and improve the utilization rate of the pesticide [104,105,106], while ethyl oleate can be used for the preparation of fungicides, acaricides [107,108], etc., which have the advantages of high efficiency, no residue, a low production cost, environmental protection, and excellent comprehensive performance (Figure 19).
Tyclopyrazoflor (35) is a pyridyl-pyrazole insecticide with a good killing effect against sap-feeding insects. This compound belongs to a novel category of chemicals that effectively manage infestations caused by Myzus persicae and sweet potato whitefly crawlers [109,110,111,112]. Unlike conventional ryanodine receptor insecticides, Tyclopyrazoflor exhibits a distinctive structure characterized by a unique amide group (Figure 19). Capsaicin (36) is a natural alkaloid obtained from the mature fruit of the capsicum [113]. Studies have shown that capsaicin has a stimulating effect in various species, can function as a repellent, and can also influence the egg-laying decisions of various insects [113,114,115]. As a botanical pesticide, capsaicin exhibits several desirable characteristics, including high effectiveness, prolonged duration, degradability, and non-toxicity (Figure 19).
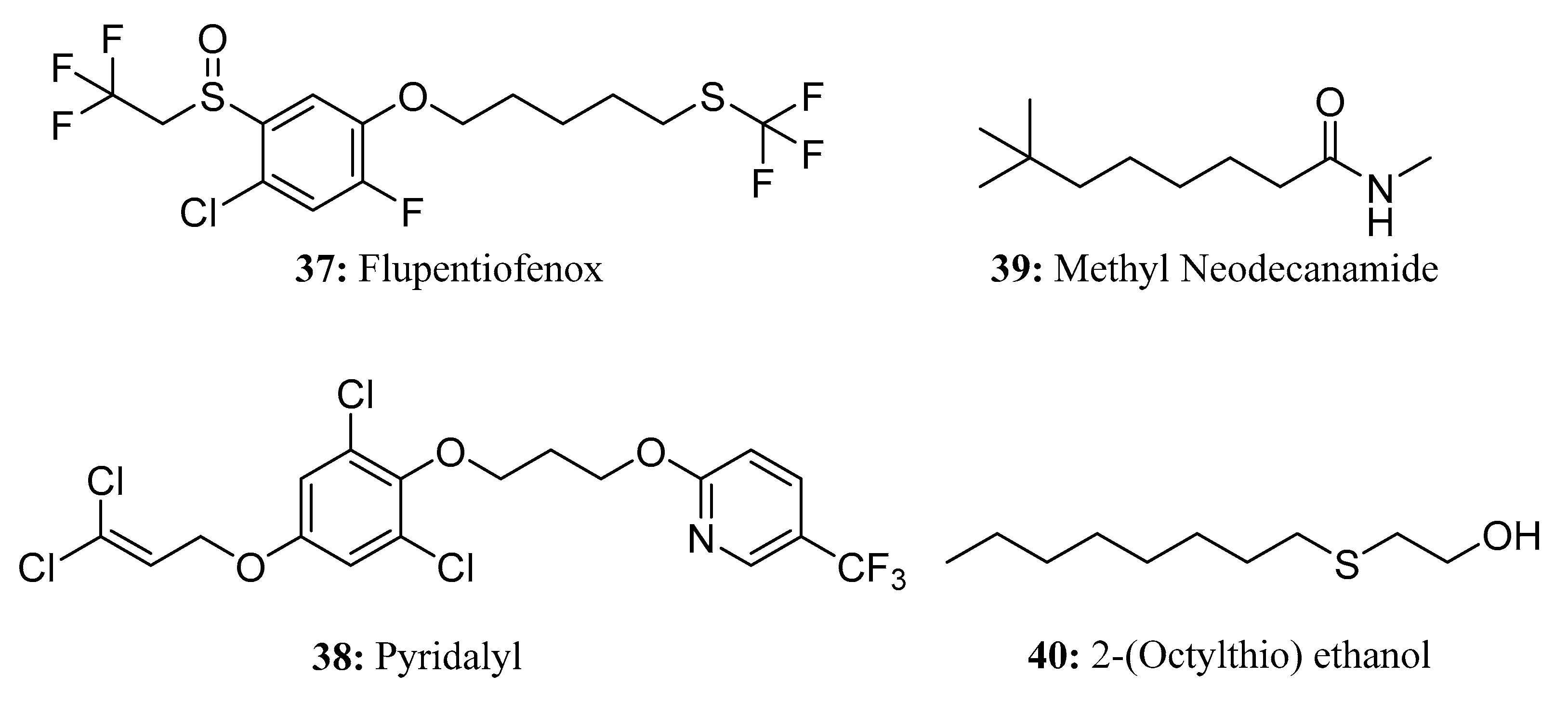
Flupentiofenox (37) is a new trifluoromethyl thioether acaricide. It is a racemate that contains chiral sulfur atoms. It has a novel structure with an unknown mechanism of action. Notably, flupentiofenox demonstrates excellent activity against adult tetranychus and larvae of the brown planthopper, even at low concentrations. As such, it can be used to control harmful arthropods, including mites on fruits and vegetables and planthoppers on rice [116] (Figure 20).
Pyridalyl (38) is a highly efficient and low-toxicity insecticide with excellent control efficacy against Lepidoptera and Thysanoptera pests [117]. Due to its unique chemical structure and characteristic insecticidal profile [118], it has no cross-resistance with various currently used insecticides in Plutella xylostella or Heliothis virescens, inferring that it may have a new mechanism of action [119,120,121] (Figure 20).
Methyl neodecanamide(MNDA, 39) is an isomeric distribution of secondary amides that has shown efficacy as an insect repellent [122,123]. MNDA can be used as insect repellent for cockroaches, mosquitoes, moths, flies, fleas, ants, lice, spiders, ticks, and mites; for this reason, it is a useful ingredient in household cleaners [122,123] (Figure 20).
2-(Octylthio)ethanol (40) [124] is a new microtoxic (nearly non-toxic) and highly effective insect repellent that exhibits a strong ability to repel mosquitoes, flies, reptiles, ants, bedbugs, and other insects (Figure 20).
Compound 41 (Figure 21) belongs to a series of C7-oxime ester derivatives (n = 0~6,8,9,14,16) of obacunone prepared by Xiang Yu; its insecticidal activity against the pre-third-instar larvae of oriental armyworm (Mythimna separata Walker) was evaluated. A comparison of compound 41 (n = 4) with the precursor obacunone showed that compound 41 (n = 4) displayed greater insecticidal activity with final mortality rates greater than 60% [125].
Fulde reported a series of dialkyl 2-bromo-l-(2,4-dichlorophenyl) vinyl phosphates (42) that exhibited biological activity against the house fly (Musca domestica L.). The toxicity of these compounds to flies was found to decrease with an increase in the length of the alkyl chain. The methyl ester was identified as the most active compound, while the 2-methoxyethyl ester was the least active [126]. This suggests that the biological activity of these compounds is not solely dependent on the length of the alkyl chain (Figure 21).
Escriba et al. synthesized a group of allyl esters of fatty acids (43a–43h) (Figure 21) from glycerol. These compounds showed ovicidal activity against codling moth eggs, and the alkyl chain length was inversely related to the ovicidal activity of the allyl esters. Notably, the two compounds with longer alkyl chains showed significantly lower ovicidal activity compared to the other compounds [127].
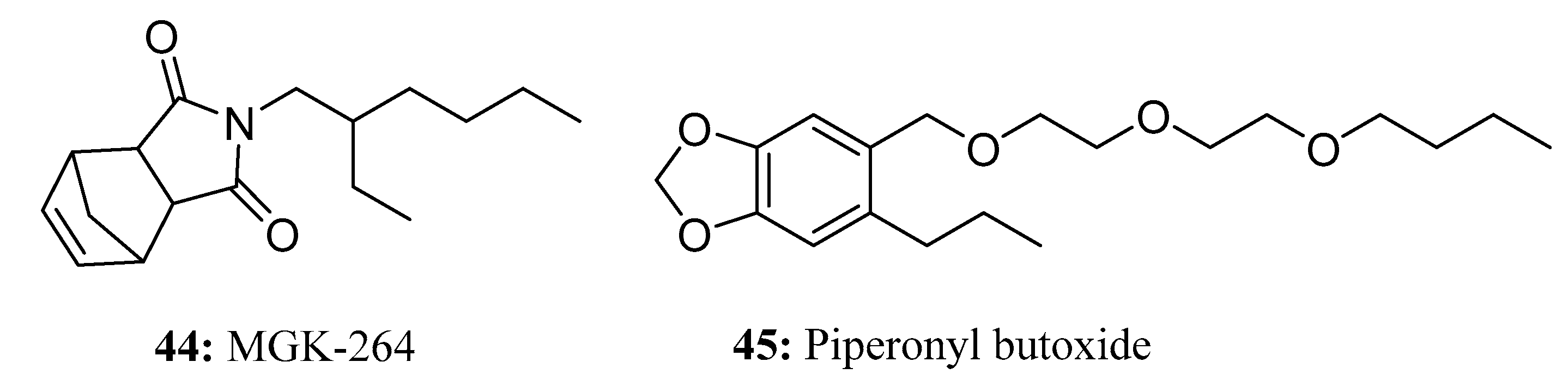
MGK-264 (44) [128,129,130] has been shown to enhance the insecticidal activity of pyrethrin, pyrethroid, and carbamate insecticides. When used in conjunction with pyrethrin or allethrin, this synergist is especially effective in preventing and controlling cockroach infestations. Importantly, MGK-264 does not possess any inherent toxicity in insects (Figure 22).
The semi-synthetic compound piperonyl butoxide (45) (Figure 22) is a methylenedioxyphenyl (MDP) compound derived from natural safrole extracted from sassafras oil [131,132]. By inhibiting insect cytochrome P450 (CYP) and esterase enzymes, piperonyl butoxide enhances the insecticidal activity of natural and synthetic pyrethroids and other insecticides [133]. Piperonyl butoxide exhibits characteristics such as prolonging the drug-holding time, broadening the insecticidal spectrum, reducing pesticide dosages, lowering costs, and being safe and non-toxic.Insect pheromones and their analogs are a very large family containing a vast number of long-chain molecules. These pheromones serve various highly specific functions, such as attracting, stimulating, facilitating or inhibiting feeding, promoting egg laying, facilitating mating, organizing assembly, alarming, and defending. Due to the wide variety of insect pheromones (both commercialized and analogues) under development, they were not reviewed in this paper.
3.3. Herbicides Containing a Long Chain
Flumiclorac-pentyl (46) is a post-emergence herbicide that exhibits selective efficacy against broadleaf weeds in soybeans. Problematic weeds such as velvetleaf, prickly sida, jimsonweed, and common lambsquarters can be effectively controlled with flumiclorac-pentyl [134]. The herbicide is absorbed by the leaves and sheaths of plants, transmitted via the phloem, and accumulates in the meristem region. Flumiclorac-pentyl demonstrates resilience against acetyl-CoA carboxylase by hindering fatty acid synthesis, disrupting cellular growth and division, and impairing lipid structures like membrane systems, ultimately inducing plant death. Remarkably, flumiclorac-pentyl stands out among aromatic oxyphenoxypropionic acid herbicides due to its exceptional safety profile for rice cultivation (Figure 23).
Pethoxamid (47) (Figure 24) is a novel chloroacetamide herbicide that was developed by Tokuyama in Japan through modification and repeated screening of the chemical structure and biological activity of the rice field herbicide thenylchlor. Pethoxamid is typically applied to the soil as a pre-plant, pre-plant-incorporated, or pre-emergence herbicide [135]. It has shown effective weed control against broadleaved weeds such as redroot pigweed (Amaranthus retroflexus L.), common lambsquarters (Chenopodium album L.), and ladysthumb (Polygonum persicaria L.) as well as annual grasses that include species from the foxtail species (Setaria spp.) and large crabgrass (Digitaria spp.) [136,137]. Similar to pretilachlor, pethoxamid inhibits the formation of very-long-chain fatty acids in responsive weeds and is generally used to control weeds before they emerge [137,138].
Pretilachlor (48) is a selective pre- or post-emergence herbicide that belongs to the chloroacetanilide class (Figure 24). It is widely used to control broadleaved weeds, various grasses, and sedges in transplanted and directly seeded paddy fields [139,140,141,142,143,144,145,146,147,148]. Pretilachlor works by inhibiting the synthesis of very-long-chain fatty acids (VLCFAs) and cell division.
Butachlor (49) is one of the widely and extensively used herbicides that is primarily used for controlling grass weeds and various broadleaved weeds in crops such as wheat, rice, and other cereal crops [149,150] (Figure 24). It is a chloroacetamide herbicide that inhibits early plant development by inhibiting the biosynthesis of long-chain fatty acids in microsomes [151,152].
Monalide (50), which belongs to the class of aniline herbicides, is an important selective herbicide for controlling weeds in vegetable crops. It is applied after the emergence of both weeds and crops [153,154] (Figure 24).
Pyridate (51) is a common single-dose rice field herbicide that belongs to the thiocarbamate herbicides and is a selective post-seedling herbicide (Figure 25). It is suitable for controlling broadleaf weeds of wheat, rice, corn, and other cereal crops, especially for porcine amaranth and some gramineous weeds. It works by blocking the weed’s photosystem II process, leading to rapid degradation of the weed’s cell walls [155].
Pentanochlor (52) is a selective, contact herbicide that is absorbed through leaves [156] (Figure 25). Used for selective weeding of carrot, celeriac, celery, fennel, parsley, parsnips, and other plants before and after seeding as well as for the pre-seeding of tomato and some flower crops.
Bicyclopyrone (53) is a highly effective broad-spectrum herbicide that demonstrates exceptional selectivity for use in corn, sugar beet, and grain fields (Figure 26). As a member of the 4-hydroxyphenylpyruvate dioxygenase (4-HPPD) inhibitor herbicide family, bicyclopyrone is responsible for inhibiting the activity of 4-HPPD, a non-heme iron (II)-dependent dioxygenase. Consequently, the biosynthesis of carotenoids is impeded, leading to albinism in the plant meristem and ultimately causing plant death [157,158,159,160,161,162,163]. The application of bicyclopyrone is very flexible; it can be used from pre-sowing to post-emergence, and it also works well in different environmental conditions and different planting methods.
Tolpyralate (54) is a novel selective herbicide for controlling weeds in corn crops. It demonstrates effective control over a broad spectrum of grasses and broadleaves while exhibiting excellent selectivity for corn [164] (Figure 26). Tolpyralate is a new 4-HPPD-inhibitor that belongs to the benzoylpyrazole family that affects sensitive weeds by interrupting the biosynthesis of plastoquinone and tocopherol and stopping the synthesis of carotenoid pigments [163,165]. In comparison to other herbicides of the same class, tolpyralate delivers comparable or superior weed control in annual broadleaves and grasses [166,167].
Syringomycin (55), which is derived from Pseudomonas syringae, is a large amphiphilic molecule with a polar peptide head and a hydrophobic 3-hydroxy fatty acid tail of variable length (from C10 to C14) that binds to N-terminal serine residues via an amide bond [168] (Figure 27). The mode of action of syringomycin is assembly into macromolecules and insertion into the cell membrane to form pores that can freely permeate cations, resulting in the rapid necrosis of plant tissues [169].
Sarmentine (56) is a natural product from Piper longum L. that has been patented as an herbicide (Figure 27). Sarmentine destroys the plant cuticle, leading to cell membrane damage followed by rapid drying and eventually tissue death [168].
Citral (57) is a diterpene component in many plant essential oils that also has been patented as an herbicide (Figure 27). Citral vapor disrupts microtubule polymerization in Arabidopsis seedlings [170], but it may have a new target for disrupting mitosis because citral affects microtubule polymerization differently than other mitotic inhibitors used as herbicides [168].
Cloquintocet-mexyl (CLM, 58) is a quinoline-based herbicide safety agent that selectively protects certain crops from herbicide injury without reducing the biological activity of target weeds [171,172,173]. CLM is always used in conjunction with clodinafop-propargyl for the effective control of isoproturon-resistant small-seed canary grass biotypes (Phalaris minor Retz.) as well as other broad-leaved weeds (Triticum aestivum) [174] while protecting crops from herbicide damage (Figure 28).
3.4. Plant Growth Regulators Containing a Long Chain
Plant growth regulators are defined as natural or chemically synthesized substances that play a role in plant development or metabolism.
In 1964, C6-C18 high-carbon primary alcohol was identified as a promising tobacco bud suppressor. Extensive research conducted by tobacco chemists and growers from various countries revealed that n-decanol (59) exhibited the most effective suppression, followed by n-octanol (60) and n-dodecanol (61). Due to its excellent biodegradability and composition of only carbon, hydrogen, and oxygen, high-carbon primary alcohol has gained significant popularity within the international tobacco industry for its non-toxic and non-residual characteristics [175,176] (Figure 29).
TRIA (62), a primary saturated alcohol, is classified as a plant growth regulator found in the epicuticular waxes of upper plant surfaces that stimulates physiological and biochemical processes in many crops [177,178] (Figure 29). Relatively low concentrations of triacontanol promote the growth of most crops, including rice (O. sativa L.) and maize (Zea Mays L.) [179,180]. Currently, TRIA has been used to improve plant tolerance to abiotic stresses such as low temperature, drought, salt stress, and heavy metals [179,181,182]. It is noteworthy that under abiotic stress, exogenous application of TRIA can promote plant growth, increase photosynthetic pigment content, and increase compatible osmolyte accumulation [183,184]. TRIA can also reduce stress by regulating the expression of certain genes [182,185].
Pelargonic acid (63) is a fatty acid widely found in nature that occurs not only in animals and plants but also in volatile organic compounds in soil [186,187] (Figure 29). It can be used as a blossom-thinning agent to prevent alternate-year cropping of apples [188]. Pelargonic acid, in the form of its salts and formulations, is also utilized as a non-selective herbicide suitable for gardens [186,189]. As a contact non-selective herbicide, it functions by attacking the cell membrane, resulting in cell leakage and the subsequent breakdown of membrane acyl lipids [187,190].
Prohydrojasmon (64) is a synthetic analog of jasmonic acid (JA) developed as a plant growth regulator [191]. Jasmonic acid (JA) and its derivatives are higher plant growth regulators that are important in regulating physiological processes such as senescence, fruit ripening, coloration, and pigment accumulation. It is believed that these compounds can improve crop quality by increasing anthocyanin, glucosinolate, terpene, and phenolic levels, all of which contribute to the quality of crops [192,193,194,195]. The application of PDJ as a plant growth regulator is mainly focused on fruits, including apples, oranges, mangoes and grapes [191,196,197]. PDJ has similar effects to endogenous JAs and has been shown to affect a variety of physiological processes, including senescence, leaf abscission, fruit ripening, coloration, and pigment accumulation. Other studies have shown that PDJ can also induce bioaccumulation of the above-mentioned secondary compounds, thus improving crop quality [191] (Figure 30).
3.5. Long-Chain Molecules with Rodenticidal Activity
Vitamin D, also known as calciferol, is a complex lipophilic molecule. Biologically, it is similar to other pseudo-steroids. Ergocalciferol (vitamin D2, 65) and cholecalciferol (vitamin D3, 66) are two of the most important and major examples; they are prohormones with very similar chemical structures but different side chains that are involved in calcium and phosphorus metabolism in animals [198]. Ergocalciferol can be used either alone for rodent control or in combination with 0.025% warfarin and 0.005% difenacoum, which are effective combinations for controlling anticoagulant-resistant or -susceptible rats and mice [199]. Ergocalciferol promotes bone calcification and tooth growth in animals and is not harmful to individuals when consumed in small amounts, but if an excessive amount is consumed, calcium in the blood can rise rapidly, leading to tissue calcification and eventual death. Cholecalciferol is also a rodenticidal agent with similar properties (Figure 31).
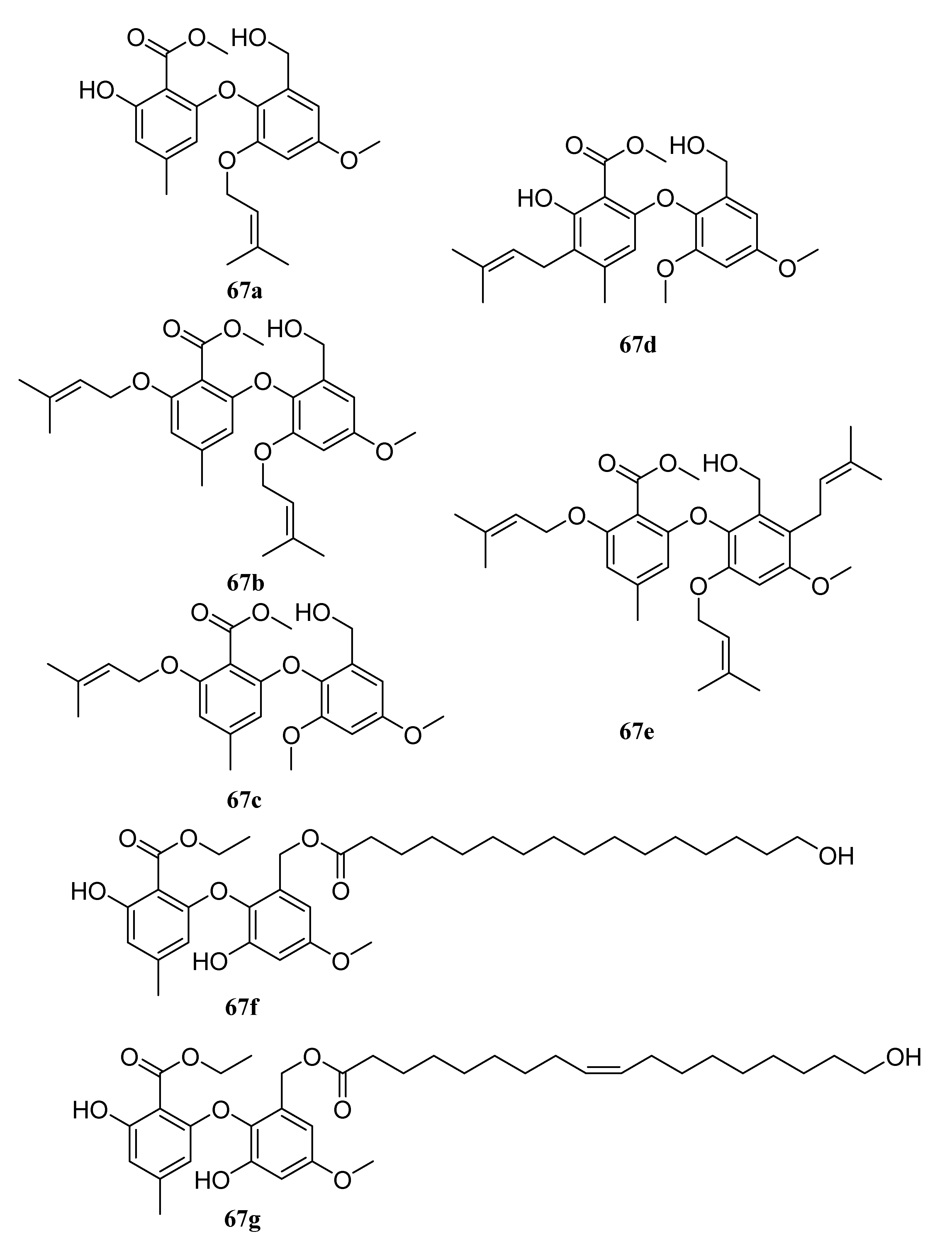
In addition, there are other kinds of long-chain molecules with agro-bioactivities, such as some organic sulfites [200] and non-ionic surfactants [201] like ammonium lauryl sulfate and triethanolamine lauryl sulfate, etc. Natural extracts are also an important source of long-chain molecules with agro-bioactivities [202,203,204]. For example, the crude plant extracts of Chenopodium ambrosioides, Conyza dioscordisis, and Convolvulus arvensis are effective against stored grain pests; the extract components are mainly long-chain fatty acid esters of hexadecanoic, arachidonic, and octadecanoic acids [203]. The hexane (HE) extract of D. insularis showed high acaricidal activity against the bovine tick by reducing oviposition and hatching rates [202], which may be related to chemical compounds such as palmitic acid, ethyl hexadecanoate, linolenic acid, and ethyl linolenoate. Zhu et al. purified nine new diphenyl ethers via fermentation using the endophytic fungus Epicoccum sorghinum L28. Compounds 67a–67e are long-chain diphenyl ethers, and compounds 67f and 67g contain ester segments formed by natural long-chain fatty acids and hydroxyl groups in diphenyl ether benzyl alcohol units (Figure 32). They have strong inhibitory activities against F. oxysporum and C. musa, which may be a defense tool induced by host chemicals. It can help hosts resist Fusarium and Colletotrichum, which widely infect mangrove plants [205].
4. Conclusions and Prospects
In summary, this paper focused on long-chain molecules with pesticide activity, highlighting the importance of the long chain in determining biological activity. The antifungal activity of glucosamine analogs increases with longer alkyl chains, while the ovicidal activity of allyl esters of fatty acids decreases with longer alkyl chains. Long-chain fatty acids with carbon chain lengths of C9~C11 exhibit the best herbicidal activity, indicating variable effects of long-chain tails on biological activity in different molecules. The introduction of long-chains in molecules can alter their geometry and physicochemical properties, offering opportunities for pesticide innovation and agricultural development. Designing long-chain compounds with these properties is an area of interest in pesticide research.
In addition, long-chain molecules exhibit a diverse range of biological activities in agricultural production, making significant contributions to the effective control of agricultural diseases, pests, and weeds while also having a strong economic value. Many agricultural bioactive long-chain molecules possess the advantages of high efficiency and low toxicity. In recent years, several new pesticide products such as seboctylamine, tyclopyrazoflor, and flupentiofenox have been continuously introduced. Furthermore, further research is needed to better understand the mechanism of action and structure–activity relationship of certain long-chain molecules. This will facilitate the development of innovative modifications and compound synthesis, ultimately promoting the wider application of long-chain molecules in agricultural production. The contribution of long chains to the biological activity of these molecules will be a subject that warrants special attention and further research.
Author Contributions
F.Y.; writing—original draft preparation, Z.Q.; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants from the National Key R&D Program of China (2021YFD1700102).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Long, Z.-Q.; Yang, L.-L.; Zhang, J.-R.; Liu, S.-T.; Xie, J.; Wang, P.-Y.; Zhu, J.-J.; Shao, W.-B.; Liu, L.-W.; Yang, S. Fabrication of Versatile Pyrazole Hydrazide Derivatives Bearing a 1,3,4-Oxadiazole Core as Multipurpose Agricultural Chemicals against Plant Fungal, Oomycete, and Bacterial Diseases. J. Agric. Food Chem. 2021, 69, 8380–8393. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.-Z.; Zhu, J.-K.; Yin, X.-D.; Yan, Y.-F.; Wang, Y.-L.; Shang, X.-F.; Liu, Y.-Q.; Zhao, Z.-M.; Peng, J.-W.; Liu, H. Design, Synthesis, and Antifungal Evaluation of Novel Quinoline Derivatives Inspired from Natural Quinine Alkaloids. J. Agric. Food Chem. 2019, 67, 11340–11353. [Google Scholar] [CrossRef] [PubMed]
- Pahutski, T.F.; Ahmad, O.K.; Marshall, E.A.; Joraski, K.; Barry, J.D.; Keathley, C.; Cordova, D.; Benner, E.; Nesnow, D.; Christianson, L.; et al. Discovery of novel (N-aryl-4-methylpiperidinyl)pyrazoles: A new class of potent lepidopteran insecticides. Pest Manag. Sci. 2023, 79, 1743–1749. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological Control of Phytopathogenic Fungi by Fatty Acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, M.; Saikia, S.; Bordoloi, P.K.; Kolita, B.; Dutta, P.P.; Bhuyan, P.D.; Dutta, S.C.; Rao, P.G. Isolation, characterization and antifungal activity of very long chain alkane derivatives from Cinnamomum obtusifolium, Elaeocarpus lanceifolius and Baccaurea sapida. J. Mol. Struct. 2017, 1142, 200–210. [Google Scholar] [CrossRef]
- Scott, S.; Cahoon, E.B.; Busta, L. Variation on a theme: The structures and biosynthesis of specialized fatty acid natural products in plants. Plant J. 2022, 111, 954–965. [Google Scholar] [CrossRef]
- He, M.; Qin, C.-X.; Wang, X.; Ding, N.-Z. Plant Unsaturated Fatty Acids: Biosynthesis and Regulation. Front. Plant Sci. 2020, 11, 390. [Google Scholar] [CrossRef] [Green Version]
- Stoyanova-Ivanova, B.; Khadzhieva, P.; Popov, S. Composition, structure and biogenesis of the ketones in rose flower wax. Phytochemistry 1969, 8, 1549. [Google Scholar] [CrossRef]
- Kokpol, U.; Chavasiri, W.; Chittawong, V.; Bruce, M.; Cunningham, G.; Miles, D. Long chain aliphatic alcohols and saturated carboxylic acids from heartwood of Rhizophora apiculata. Phytochemistry 1993, 33, 1129–1131. [Google Scholar] [CrossRef]
- Misra, T.N.; Singh, R.S.; Pandey, H.S.; Prasad, C.; Singh, B.P. Antifungal essential oil and a long chain alcohol from Achyranthes aspera. Phytochemistry 1992, 31, 1811–1812. [Google Scholar] [CrossRef]
- Raynor, L.; Mitchell, A.; Walker, R.; Walker, K. Antifungal Activities of Four Fatty Acids against Plant Pathogenic Fungi. Mycopathologia 2004, 157, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Vidal, D.M.; von Rymon-Lipinski, A.-L.; Ravella, S.; Groenhagen, U.; Herrmann, J.; Zaburannyi, N.; Zarbin, P.H.G.; Varadarajan, A.R.; Ahrens, C.H.; Weisskopf, L.; et al. Long-Chain Alkyl Cyanides: Unprecedented Volatile Compounds Released by Pseudomonas and Micromonospora Bacteria. Angew. Chem. Int. Ed. Engl. 2017, 56, 4342–4346. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Lin, S.; Gan, M.; Chen, M.; Li, L.; Wang, S.; Zi, J.; Fan, X.; Liu, Y.; Si, Y.; et al. Yaoshanenolides A and B: New Spirolactones from the Bark of Machilus yaoshansis. Org. Lett. 2012, 14, 1004–1007. [Google Scholar] [CrossRef]
- Komae, H.; Hayashi, N.; Kosela, S. Palmitone from the fruits of Lindera citriodora. Phytochemistry 1971, 10, 3311. [Google Scholar] [CrossRef]
- Ho, C.-L.; Hsu, K.-P.; Wang, E.I.-C.; Su, Y.-C. Composition and Antimicrobial Activity of the Leaf Essential Oil of Machilus obovatifolia from Taiwan. J. Essent. Oil Res. 2009, 21, 471–475. [Google Scholar] [CrossRef]
- Nowicka, B.; Kruk, J. Occurrence, biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1587–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abby, S.S.; Kazemzadeh, K.; Vragniau, C.; Pelosi, L.; Pierrel, F. Advances in bacterial pathways for the biosynthesis of ubiquinone. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148259. [Google Scholar] [CrossRef]
- Siebert, M.; Severin, K.; Heide, L. Formation of 4-hydroxybenzoate in Escherichia coli: Characterization of the ubiC gene and its encoded enzyme chorismate pyruvate-lyase. Microbiology 1994, 140, 897–904. [Google Scholar] [CrossRef] [Green Version]
- White, M.D.; Payne, K.A.P.; Fisher, K.; Marshall, S.A.; Parker, D.; Rattray, N.J.W.; Trivedi, D.K.; Goodacre, R.; Rigby, S.E.J.; Scrutton, N.S.; et al. UbiX is a flavin prenyltransferase required for bacterial ubiquinone biosynthesis. Nature 2015, 522, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wu, B.; Zhang, X.; Fan, X.; Niu, L.; Li, X.; Wang, J.; Teng, M. Structural and biochemical studies reveal UbiG/Coq3 as a class of novel membrane-binding proteins. Biochem. J. 2015, 470, 105–114. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, X.; Wang, C.; Liu, Y.; Fan, X.; Zhang, L.; Niu, L.; Teng, M.; Li, X. Structural insights into the methyl donor recognition model of a novel membrane-binding protein UbiG. Sci. Rep. 2016, 6, 23147. [Google Scholar] [CrossRef]
- Kwon, O.; Kotsakis, A.; Meganathan, R. Ubiquinone (coenzyme Q) biosynthesis in Escherichia coli: Identification of the ubiF gene. FEMS Microbiol. Lett. 2000, 186, 157–161. [Google Scholar] [CrossRef]
- Chehade, M.H.; Loiseau, L.; Lombard, M.; Pecqueur, L.; Ismail, A.; Smadja, M.; Golinelli-Pimpaneau, B.; Mellot-Draznieks, C.; Hamelin, O.; Aussel, L.; et al. ubiI, a New Gene in Escherichia coli Coenzyme Q Biosynthesis, Is Involved in Aerobic C5-hydroxylation. J. Biol. Chem. 2013, 288, 20085–20092. [Google Scholar] [CrossRef] [Green Version]
- Fehr, M.; Wolf, A.; Stammler, G. Binding of the respiratory chain inhibitor ametoctradin to the mitochondrial bc1 complex. Pest Manag. Sci. 2016, 72, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Chai, B.-S.; Fu, X.-C.; Sun, X.-F.; Li, Q.; Li, Z.-N. Synthesis of ametoctradin and its bioactivity. Nongyao 2012, 51, 645-646–674. [Google Scholar]
- Gao, X.; Hu, S.; Liu, Z.; Zhu, H.; Yang, J.; Han, Q.; Fu, Y.; Miao, J.; Gu, B.; Liu, X. Analysis of resistance risk and resistance-related point mutations in Cyt b of QioI fungicide ametoctradin in Phytophthora litchii. Pest Manag. Sci. 2022, 78, 2921–2930. [Google Scholar] [CrossRef]
- Dreinert, A.; Wolf, A.; Mentzel, T.; Meunier, B.; Fehr, M. The cytochrome bc complex inhibitor Ametoctradin has an unusual binding mode. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 567–576. [Google Scholar] [CrossRef]
- Takenaka, M.; Kimura, S.; Tanaka, T.; Wada, T. Chiral Synthesis of Pefurazoate Enantiomers and Their Antifungal Activity to Gibberella fujikuroi. J. Pestic. Sci. 1992, 17, 205–211. [Google Scholar] [CrossRef]
- Takenaka, M.; Nishimura, T.; Hayashi, K. Enantioselective Antifungal Activity of Pefurazoate against Pathogens of Rice Seed Diseases. Nippon. Noyaku Gakkaishi 2001, 26, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Xi, J.; Shao, J.; Wang, Y.; Wang, X.; Yang, H.; Zhang, X.; Xiong, D. Acute toxicity of triflumizole to freshwater green algae Chlorella vulgaris. Pestic. Biochem. Physiol. 2019, 158, 135–142. [Google Scholar] [CrossRef]
- Nakata, A.; Hashimoto, S.; Ikura, K.; Katsuura, K. Development of a New Fungicide, Triflumizole. Nippon. Noyaku Gakkaishi 1991, 16, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Song, J.-G.; Lee, I.-K.; Yeo, W.-H.; Yun, B.-S. Bacillus sp. BS061 Suppresses Powdery Mildew and Gray Mold. Mycobiology 2013, 41, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markoglou, A.N.; Ziogas, B.N. Genetic control of resistance to tridemorph inustilago maydis. Phytoparasitica 2000, 28, 349–360. [Google Scholar] [CrossRef]
- Wang, S.; Sun, H.; Liu, Y. Residual behavior and risk assessment of tridemorph in banana conditions. Food Chem. 2018, 244, 71–74. [Google Scholar] [CrossRef]
- Kerkenaar, A.; Uchiyama, M.; Versluis, G. Specific effects of tridemorph on sterol biosynthesis in Ustilago maydis. Pestic. Biochem. Physiol. 1981, 16, 97–104. [Google Scholar] [CrossRef]
- Guin, D.; Mittal, S.; Bozymski, B.; Shukla, D.; Gruebele, M. Dodine as a Kosmo-Chaotropic Agent. J. Phys. Chem. Lett. 2019, 10, 2600–2605. [Google Scholar] [CrossRef]
- Lian, C.; Xu, W.; Luo, Y.; Zhu, X.; Fan, Y.; Redshaw, C.; Tao, Z.; Xiao, X. Detection of the pesticide dodine using a cucurbit [10] uril-based fluorescent probe. Microchem. J. 2021, 167, 106309. [Google Scholar] [CrossRef]
- Biswas, B.; Singh, P.C. Does Fungicide “Dodine” Unfold Protein like Kosmo-Chaotropic Agent? J. Phys. Chem. B 2019, 123, 8240–8246. [Google Scholar] [CrossRef]
- Feng, Y.; Han, J.; Liang, L.; Ma, X.; Zhang, A.; Bian, Y. Residues, storage stability, and long-term dietary risk assessment of Xinjunan in cucumber, tomato, and citrus. Int. J. Environ. Anal. Chem. 2022. ahead of print. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.; Li, Y.; Lai, Q. The Development and Application of Novel Technology of Xinjunan in the Soil-borne Disease Control. Shijie Nongyao 2020, 42, 46–48. [Google Scholar] [CrossRef]
- Han, C.; Hu, B.; Chen, S.; Wang, N.; Hou, J.; Jin, N.; Shen, Y. Determination of Xinjunan pesticide residue in foodstuffs of plant origin by a modified QuEChERS method and ultra performance liquid chromatography-tandem mass spectrometry. LWT 2021, 151, 112101. [Google Scholar] [CrossRef]
- Smarzewska, S.; Metelka, R.; Guziejewski, D.; Skowron, M.; Skrzypek, S.; Brycht, M.; Ciesielski, W. Voltammetric behaviour and quantitative determination of pesticide iminoctadine. Anal. Methods 2014, 6, 1884–1889. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment of the active substance guazatine (variant assessed guazatine acetates). EFSA J. 2010, 8, 1708. [Google Scholar] [CrossRef]
- Brown, G.E. Efficacy of Guazatine and Iminoctadine for Control of Postharvest Decays of Oranges. Plant Dis. 1988, 72, 906. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, L.; Ma, D.; Gao, Y.; Mu, W.; Liu, F. A bioactivity and biochemical analysis of iminoctadine tris (albesilate) as a fungicide against Corynespora cassiicola. Pestic. Biochem. Physiol. 2019, 158, 121–127. [Google Scholar] [CrossRef]
- Abrahamian, P.; Jones, J.B.; Vallad, G.E. Efficacy of copper and copper alternatives for management of bacterial spot on tomato under transplant and field production. Crops Prot. 2019, 126, 104919. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Bika, R.; Jennings, C.; Palmer, C.; Simmons, T. Comparative Performance of Chemical and Biological-based Products in Management of Algal Leaf Spot on Magnolia. Horttechnology 2020, 30, 733–740. [Google Scholar] [CrossRef]
- Clifford, D.R.; Gendle, P.; Holgate, M.E.; Hunter, T. Comparison of paint and gel formulations for the treatment of Nectria cankers on apple trees. Ann. Appl. Biol. 1987, 110, 471–487. [Google Scholar] [CrossRef]
- Kresmann, S.; Arokia, A.H.R.; Koch, C.; Sures, B. Ecotoxicological potential of the biocides terbutryn, octhilinone and methylisothiazolinone: Underestimated risk from biocidal pathways? Sci. Total. Environ. 2018, 625, 900–908. [Google Scholar] [CrossRef]
- Nagorka, R.; Gleue, C.; Scheller, C.; Moriske, H.-J.; Straff, W. Isothiazolone emissions from building products. Indoor Air 2015, 25, 68–78. [Google Scholar] [CrossRef]
- Mayorquin, J.S.; Carrillo, J.D.; Twizeyimana, M.; Peacock, B.B.; Sugino, K.Y.; Na, F.; Wang, D.H.; Kabashima, J.N.; Eskalen, A. Chemical Management of Invasive Shot Hole Borer and Fusarium Dieback in California Sycamore (Platanus racemosa) in Southern California. Plant Dis. 2018, 102, 1307–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahluwalia, V.; Kumar, J.; Rana, V.S.; Singh, R.; Sati, O.P.; Walia, S.; Garg, N. Synthesis and antimicrobial activity of esters of 3-ethoxy-4-hydroxybenzaldehyde oxime. Toxicol. Environ. Chem. 2016, 99, 1–9. [Google Scholar] [CrossRef]
- Lin, G.-S.; Bai, X.; Duan, W.; Cen, B.; Huang, M.; Lu, S. High Value-Added Application of Sustainable Natural Forest Product α-Pinene: Synthesis of Myrtenal Oxime Esters as Potential KARI Inhibitors. ACS Sustain. Chem. Eng. 2019, 7, 7862–7868. [Google Scholar] [CrossRef]
- Zhang, S.-B.; Qin, Y.-L.; Li, S.-F.; Lv, Y.-Y.; Zhai, H.-C.; Hu, Y.-S.; Cai, J.-P. Antifungal mechanism of 1-nonanol against Aspergillus flavus growth revealed by metabolomic analyses. Appl. Microbiol. Biotechnol. 2021, 105, 7871–7888. [Google Scholar] [CrossRef]
- Liu, W.-S.; Wang, C.-H.; Sun, J.-F.; Hou, G.-G.; Wang, Y.-P.; Qu, R.-J. Synthesis, Characterization and Antibacterial Properties of Dihydroxy Quaternary Ammonium Salts with Long Chain Alkyl Bromides. Chem. Biol. Drug Des. 2015, 85, 91–97. [Google Scholar] [CrossRef]
- Muhizi, T.; Grelier, S.; Coma, V. Synthesis of N-Alkyl-β-d-glucosylamines and Their Antimicrobial Activity against Fusarium proliferatum, Salmonellatyphimurium, and Listeria innocua. J. Agric. Food Chem. 2009, 57, 11092–11099. [Google Scholar] [CrossRef]
- Mandal, S.; Kanrar, B.; Das, S.; Bhattacharyya, A. Analytical Method Validation for the Determination of Meptyldinocap As 2,4-Dinitrooctylphenol Metabolite in Mango and Soil Using LC−MS/MS and Dissipation Study of the Fungicide in Indian Mango Field Ecosystem. J. Agric. Food Chem. 2010, 58, 8911–8917. [Google Scholar] [CrossRef]
- Černohlávková, J.; Jarkovský, J.; Hofman, J. Effects of fungicides mancozeb and dinocap on carbon and nitrogen mineralization in soils. Ecotoxicol. Environ. Saf. 2009, 72, 80–85. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Lofgren, C.S.; Williams, D.F. Fluoroaliphatic Sulfones: A New Class of Delayed-action Insecticides for Control of Solenopsis invicta (Hymenoptera: Formicidae). J. Econ. Entomol. 1985, 78, 1190–1197. [Google Scholar] [CrossRef]
- Nascimento, R.A.; Nunoo, D.B.; Bizkarguenaga, E.; Schultes, L.; Zabaleta, I.; Benskin, J.P.; Spanó, S.; Leonel, J. Sulfluramid use in Brazilian agriculture: A source of per- and polyfluoroalkyl substances (PFASs) to the environment. Environ. Pollut. 2018, 242, 1436–1443. [Google Scholar] [CrossRef]
- Kinoshita, S.; Koura, Y.; Kariya, H.; Ohsaki, N.; Watanabe, T. AKD-2023: A novel miticide. Biological activity and mode of action. Pestic. Sci. 1999, 55, 659–660. [Google Scholar] [CrossRef]
- Van Nieuwenhuyse, P.; Van Leeuwen, T.; Khajehali, J.; Vanholme, B.; Tirry, L. Mutations in the mitochondrial cytochromeb of Tetranychus urticae Koch (Acari: Tetranychidae) confer cross-resistance between bifenazate and acequinocyl. Pest Manag. Sci. 2009, 65, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Dekeyser, M. Acaricide mode of action. Pest Manag. Sci. 2005, 61, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Fenical, W. The unique chemistry and biology of the piericidins. J. Antibiot. 2016, 69, 582–593. [Google Scholar] [CrossRef]
- Bridges, H.R.; Fedor, J.G.; Blaza, J.N.; Di Luca, A.; Jussupow, A.; Jarman, O.D.; Wright, J.J.; Agip, A.-N.A.; Gamiz-Hernandez, A.P.; Roessler, M.M.; et al. Structure of inhibitor-bound mammalian complex I. Nat. Commun. 2020, 11, 5261. [Google Scholar] [CrossRef]
- Tamura, S.; Takahashi, N.; Miyamoto, S.; Mori, R.; Suzuki, S.; Nagatsu, J. Isolation and Physiological Activities of Piericidin A, A Natural Insecticide Produced by Streptomyces. Agric. Biol. Chem. 1963, 27, 576–582. [Google Scholar] [CrossRef]
- Eisinger, M.; Almog, Y. Pyrimidifen intoxication. Ann. Emerg. Med. 2003, 42, 289–291. [Google Scholar] [CrossRef]
- Bai, Y.-l. Acaricides with different modes of action. Xiandai Nongyao 2005, 4, 27–30. [Google Scholar]
- Alali, F.Q.; Liu, X.-X.; McLaughlin, J.L. Annonaceous Acetogenins: Recent Progress. J. Nat. Prod. 1999, 62, 504–540. [Google Scholar] [CrossRef]
- Zafra-Polo, M.C.; González, M.C.; Estornell, E.; Sahpaz, S.; Cortes, D. Acetogenins from annonaceae, inhibitors of mitochondrial complex I. Phytochemistry 1996, 42, 253–271. [Google Scholar] [CrossRef]
- Lümmen, P. Complex I inhibitors as insecticides and acaricides. Biochim. Biophys. Acta Bioenerg. 1998, 1364, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazire, A.; Gillon, E.; Lockridge, O.; Vallet, V.; Nachon, F. The kinetic study of the inhibition of human cholinesterases by demeton-S-methyl shows that cholinesterase-based titration methods are not suitable for this organophosphate. Toxicol. Vitr. 2011, 25, 754–759. [Google Scholar] [CrossRef]
- Dubois, K.P.; Plazak, G.J. The acute toxicity and anticholinesterase action of O,O-dimethyl S-ethyl-2-sulfinylethyl phosphorothioate (Meta-Systox R) and related compounds. Toxicol. Appl. Pharmacol. 1962, 4, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Szeto, S.Y.; Vernon, R.S.; Brown, M.J. Degradation of disulfoton in soil and its translocation into asparagus. J. Agric. Food Chem. 1983, 31, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Futagami, K.; Otsubo, K.; Nakao, Y.; Aoyama, T.; Iimori, E.; Urakami, S.; Ide, M.; Oishi, R. Acute Organophosphate Poisoning After Disulfoton Ingestion. J. Toxicol. Clin. Toxicol. 1995, 33, 151–155. [Google Scholar] [CrossRef]
- Usui, K.; Hayashizaki, Y.; Minagawa, T.; Hashiyada, M.; Nakano, A.; Funayama, M. Rapid determination of disulfoton and its oxidative metabolites in human whole blood and urine using QuEChERS extraction and liquid chromatography–tandem mass spectrometry. Leg. Med. 2012, 14, 309–316. [Google Scholar] [CrossRef]
- Qader, B.; Baron, M.; Hussain, I.; Sevilla, J.; Johnson, R.P.; Gonzalez-Rodriguez, J. Electrochemical determination of disulfoton using a molecularly imprinted poly-phenol polymer. Electrochimica Acta 2018, 295, 333–339. [Google Scholar] [CrossRef]
- Lammerink, J.; Banfield, R.A. Effect of disulfoton on growth of aphid-free oilseed rape. N. Z. J. Exp. Agric. 1979, 7, 221–223. [Google Scholar] [CrossRef]
- Li, H.; Pan, L.; Yu, C.; Zhang, X.; Cui, X.; Luo, T.; Cao, Z.; Wang, J.; Li, Q. Development and Validation for Simultaneous Determination of Disulfoton and Its Five Metabolites in Seven Agro-Products Using Liquid Chromatography-Tandem Mass Spectrometry Combined with QuEChERS Extraction Method. Chromatographia 2022, 85, 529–537. [Google Scholar] [CrossRef]
- Tabrizi, A.B.; Abdollahi, A. Determination of Organothiophosphate Insecticides in Environmental Water Samples by a Very Simple and Sensitive Spectrofluorimetric Method. Bull. Environ. Contam. Toxicol. 2015, 95, 536–541. [Google Scholar] [CrossRef]
- Parham, H.; Saeed, S. Resonance Rayleigh scattering method for determination of ethion using silver nanoparticles as probe. Talanta 2015, 131, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Khaled, E.; Kamel, M.S.; Hassan, H.; Abdel-Gawad, H.; Aboul-Enein, H.Y. Performance of a portable biosensor for the analysis of ethion residues. Talanta 2014, 119, 467–472. [Google Scholar] [CrossRef]
- Pang, Y.-P.; Singh, S.K.; Gao, Y.; Lassiter, T.L.; Mishra, R.K.; Zhu, K.Y.; Brimijoin, S. Selective and Irreversible Inhibitors of Aphid Acetylcholinesterases: Steps Toward Human-Safe Insecticides. PLoS ONE 2009, 4, e4349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryger, G.; Harel, M.; Giles, K.; Toker, L.; Velan, B.; Lazar, A.; Kronman, C.; Barak, V.; Ariel, N.; Shafferman, A.; et al. Structures of recombinant native and E202Q mutant human acetylcholinesterase complexed with the snake-venom toxin fasciculin-II. Acta Crystallogr. Sect. D Biol. Crystallogr. 2000, 56, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.-P. Novel Acetylcholinesterase Target Site for Malaria Mosquito Control. PLoS ONE 2006, 1, e58. [Google Scholar] [CrossRef] [Green Version]
- Pezzementi, L.; Rowland, M.; Wolfe, M.; Tsigelny, I. Inactivation of an invertebrate acetylcholinesterase by sulfhydryl reagents: The roles of two cysteines in the catalytic gorge of the enzyme. Invertebr. Neurosci. 2006, 6, 47–55. [Google Scholar] [CrossRef]
- Isobe, N.; Suzuki, T.; Nishikawa, J.-I.; Kaneko, H.; Nakatsuka, I.; Yoshitake, A. Metabolism of Empenthrin Isomers in Rats. J. Pestic. Sci. 1992, 17, 27–37. [Google Scholar] [CrossRef]
- Yoshida, K.; Tsuda, S.; Okuno, Y. Practical application of empenthrin as a mothproofer of textile. Sen’i Gakkaishi 1984, 40, T254–T262. [Google Scholar] [CrossRef] [Green Version]
- Kanamaru, H.; Kawahara, K.; Nishioka, K. Stereoselective synthesis of empenthrin, a novel insecticide against fabric pests, in a regioselectively 14C-labeled form. Radioisotopes 1991, 40, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Degitz, S.J.; Durhan, E.J.; Tietge, J.E.; Kosian, P.A.; Holcombe, G.W.; Ankley, G.T. Developmental toxicity of methoprene and several degradation products in Xenopus laevis. Aquat. Toxicol. 2003, 64, 97–105. [Google Scholar] [CrossRef]
- Naruse, S.; Washidu, Y.; Miura, K.; Shinoda, T.; Minakuchi, C. Methoprene-tolerant is essential for embryonic development of the red flour beetle Tribolium castaneum. J. Insect Physiol. 2020, 121, 104017. [Google Scholar] [CrossRef] [PubMed]
- Wijayaratne, L.; Fields, P.G. Effect of methoprene on the heat tolerance and cold tolerance of Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2010, 46, 166–173. [Google Scholar] [CrossRef]
- Bai, H.; Gelman, D.B.; Palli, S.R. Mode of action of methoprene in affecting female reproduction in the African malaria mosquito, Anopheles gambiae. Pest Manag. Sci. 2010, 66, 936–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daglish, G.J.; Holloway, J.C.; Nayak, M.K. Implications of methoprene resistance for managing Rhyzopertha dominica (F.) in stored grain. J. Stored Prod. Res. 2013, 54, 8–12. [Google Scholar] [CrossRef]
- Aubuchon, M.D.; Mullen, G.R.; Eubanks, M.D. Efficacy of broadcast and perimeter applications of S-methoprene bait on the red imported fire ant in grazed pastures. J. Econ. Entomol. 2006, 99, 621–625. [Google Scholar] [CrossRef]
- Moser, B.A.; Koehler, P.G.; Patterson, R.S. Effect of Methoprene and Diflubenzuron on Larval Development of the Cat Flea (Siphonaptera: Pulicidae). J. Econ. Entomol. 1992, 85, 112–116. [Google Scholar] [CrossRef]
- Wilson, T.G. The molecular site of action of juvenile hormone and juvenile hormone insecticides during metamorphosis: How these compounds kill insects. J. Insect Physiol. 2004, 50, 111–121. [Google Scholar] [CrossRef]
- Mohandass, S.; Arthur, F.; Zhu, K.; Throne, J. Hydroprene: Mode of action, current status in stored-product pest management, insect resistance, and future prospects. Crops Prot. 2006, 25, 902–909. [Google Scholar] [CrossRef]
- Mori, K.; Takigawa, T.; Manabe, Y.; Tominaga, M.; Matsui, M.; Kiguchi, K.; Akai, H.; Ohtaki, T. Synthesis of compounds with juvenile hormone activity. XXI. Effect of the molecular chain length on biological activity of juvenile hormone analogs. Agric. Biol. Chem. 1975, 39, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Kisida, H.; Hatakoshi, M.; Itaya, N.; Nakayama, I. New Insect Juvenile Hormone Mimics: Thiolcarbamates. Agric. Biol. Chem. 1984, 48, 2889–2891. [Google Scholar] [CrossRef]
- Jones, K.A. Environmentally-Safe Bicarbonate-Containing Pesticide Compositions. WO9400982A1, 20 January 1994. [Google Scholar]
- Serre, I.; Cabanne, F.; Gauvrit, C. seed oil derivatives as adjuvants: Influence of methyl to octadecyl oleates on the penetration of herbicides through various plant cuticles. Meded.-Fac. Landbouwkd. Toegepaste Biol. Wet. 1993, 58, 795–802. [Google Scholar]
- Davidson, W.S.; Saxena, R.K.; Gupta, R. The fungistatic action of oleic acid. Curr. Sci. 1999, 76, 1137–1140. [Google Scholar]
- Duan, W.; Zhao, B.; Liu, Y. Cyhalothrin Emulsified Concentrate with Methyl Oleate as Solvent and Its Preparation Method. CN105532652, 4 May 2016. [Google Scholar]
- Li, X.; Jiang, J.; Wang, Q.; Qi, P.; Liu, X.; Li, R.; Zhou, K.; Li, H.; Chen, Z.; Jin, L. Tolfenpyrad Electrostatic Spray Liquid and Its Application in Controlling Tea Leafhopper. CN111357749, 3 July 2020. [Google Scholar]
- Wang, Y.; Hu, X.; Wang, L. Methyl Oleate Emulsifier for Pesticide and Preparation Method Thereof. CN110558315, 13 December 2019. [Google Scholar]
- Dai, G.; Chen, Y. Application of Ethyl oleate and Ethyl Oleate Fungicide. CN104957135, 7 October 2015. [Google Scholar]
- Dai, G.; Chen, Y. Application of Ethyl Oleate and Ethyl Oleate Acaricide. CN104996423, 28 October 2015. [Google Scholar]
- Gray, K.C.; Heider, P.; McGough, P.; Ondari, M.; Devaraj, J.; Yang, Q.; Frycek, G.; Graham, B.; Neuman, J.; Lorsbach, B.A.; et al. Development of a Scalable Process for the Insecticidal Candidate Tyclopyrazoflor. Part 3. A Scalable Synthesis of Methyl 3-((3,3,3-Trifluoropropyl)thio)propanoate via Thiol–Ene Chemistry. Org. Process. Res. Dev. 2019, 23, 2142–2147. [Google Scholar] [CrossRef]
- Buysse, A.M.; Niyaz, N.M.; Zhang, Y.; Walsh, M.J.; Kubota, A.; Hunter, R.; Trullinger, T.K.; Lowe, C.T.; Knueppel, D.; Demeter, D.A.; et al. Preparation of Pyridinylpyrazolamine Derivatives as Pesticides and Their Pesticidal Compositions. WO2013162715A2, 31 October 2013. [Google Scholar]
- Yang, Q.; Li, X.; Lorsbach, B.A.; Muhuhi, J.M.; Roth, G.A.; Gray, K.; Podhorez, D.E. Development of a Scalable Process for the Insecticidal Candidate Tyclopyrazoflor. Part 2. Fit-for-Purpose Optimization of the Route to Tyclopyrazoflor Featuring [3 + 2] Cyclization of 3-Hydrazinopyridine·2HCl and Methyl Acrylate. Org. Process. Res. Dev. 2019, 23, 2133–2141. [Google Scholar] [CrossRef]
- Chen, M.; Li, Z.; Shao, X.; Maienfisch, P. Bioisosteric-Replacement-Driven Lead Optimization of Tyclopyrazoflor. J. Agric. Food Chem. 2022, 70, 11123–11137. [Google Scholar] [CrossRef]
- Wang, J.; Yu, S.; Wang, L.; Liu, T.; Yang, X.; Hu, X.; Wang, Y. Capsaicin decreases fecundity in the Asian malaria vector Anopheles stephensi by inhibiting the target of rapamycin signaling pathway. Parasites Vectors 2022, 15, 458. [Google Scholar] [CrossRef] [PubMed]
- Edelson, J.V.; Duthie, J.; Roberts, W. Toxicity of biorational insecticides: Activity against the green peach aphid, Myzus persicae (Sulzer). Pest Manag. Sci. 2002, 58, 255–260. [Google Scholar] [CrossRef]
- Li, Y.; Bai, P.; Wei, L.; Kang, R.; Chen, L.; Zhang, M.; Tan, E.K.; Liu, W. Capsaicin Functions as Drosophila Ovipositional Repellent and Causes Intestinal Dysplasia. Sci. Rep. 2020, 10, 9963. [Google Scholar] [CrossRef]
- Domon, K.; Toriyabe, K.; Ogawa, Y.; Bessho, J.; Kawamoto, K.; Watanabe, A.; Komatsu, M.; Matsuda, T.; Ito, S. Preparation of Alkylphenylsulphide Derivatives as Pest Control Agents. WO2013157229A1, 24 October 2013. [Google Scholar]
- Powell, G.F.; Ward, D.A.; Prescott, M.C.; Spiller, D.G.; White, M.R.; Turner, P.C.; Earley, F.G.; Phillips, J.; Rees, H.H. The molecular action of the novel insecticide, Pyridalyl. Insect Biochem. Mol. Biol. 2011, 41, 459–469. [Google Scholar] [CrossRef]
- Sakamoto, N.; Hirose, T.; Saito, S.; Umeda, K. Discovery and development of pyridalyl. J. Pestic. Sci. 2012, 37, 265–266, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, N.; Matsuo, S.; Suzuki, M.; Hirose, T.; Tsushima, K.; Umeda, K. Preparation of Dihalopropene Insecticides and Acaricides. WO9611909A1, 25 April 1996. [Google Scholar]
- Isayama, S.; Saito, S.; Kuroda, K.; Umeda, K.; Kasamatsu, K. Pyridalyl, a novel insecticide: Potency and insecticidal selectivity. Arch. Insect Biochem. Physiol. 2005, 58, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, N.; Saito, S.; Hirose, T.; Suzuki, M.; Matsuo, S.; Izumi, K.; Nagatomi, T.; Ikegami, H.; Umeda, K.; Tsushima, K.; et al. The discovery of pyridalyl: A novel insecticidal agent for controlling lepidopterous pests. Pest Manag. Sci. 2003, 60, 25–34. [Google Scholar] [CrossRef]
- Steltenkamp, R.J.; Hamilton, R.L.; Cooper, R.A.; Schal, C. Alkyl and Aryl Neoalkanamides: Highly Effective Insect Repellents. J. Med. Entomol. 1992, 29, 141–149. [Google Scholar] [CrossRef]
- Rasmussen, H.T.; Friedman, S.K.; Mustilli, A.J.; McDonough, R.; McPherson, B.P. Analysis of Methyl Neodecanamide in Lake Water by Reversed-Phase High Performance Liquid Chromatography and Gas Chromatography-Mass Spectrometry. J. Liq. Chromatogr. 1994, 17, 589–601. [Google Scholar] [CrossRef]
- Campbell, M.M. A test for repellency to non-biting flies and a comparison of repellents using Musca domestica L. Pestic. Sci. 1983, 14, 199–212. [Google Scholar] [CrossRef]
- Yu, X.; Shi, D.; Zhi, X.; Li, Q.; Yao, X.; Xu, H. Synthesis and quantitative structure–activity relationship (QSAR) study of C7-oxime ester derivatives of obacunone as insecticidal agents. RSC Adv. 2015, 5, 31700–31707. [Google Scholar] [CrossRef]
- Fulde, S.; Kroczynaski, J.; Malinowski, H. Structure-activity relationship for some substituted dialkyl vinyl phosphates. Pestic. Sci. 1980, 11, 20–22. [Google Scholar] [CrossRef]
- Escribà, M.; Barbut, M.; Eras, J.; Canela, R.; Avilla, J.; Balcells, M. Synthesis of Allyl Esters of Fatty Acids and Their Ovicidal Effect on Cydia pomonella (L.). J. Agric. Food Chem. 2009, 57, 4849–4853. [Google Scholar] [CrossRef]
- Sahay, N.; Agarwal, R. MGK-264- pyrethroid synergism against Lymnaea acuminata. Chemosphere 1997, 35, 1011–1021. [Google Scholar] [CrossRef]
- Singh, K.; Singh, A.; Singh, D.K. The use of piperonyl butoxide and MGK-264 to improve the efficacy of some plant-derived molluscicides. Pestic. Sci. 1998, 54, 145–149. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, Y. Synergistic effects of MGK-264. Zhonghua Weisheng Shachong Yaoxie 2007, 13, 468–469. [Google Scholar]
- Bae, J.-W.; Kwon, W.-S. Piperonyl butoxide, a synergist of pesticides can elicit male-mediated reproductive toxicity. Reprod. Toxicol. 2021, 100, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Marchand, P.A.; Dimier-Vallet, C.; Vidal, R. Biorational substitution of piperonyl butoxide in organic production: Effectiveness of vegetable oils as synergists for pyrethrums. Environ. Sci. Pollut. Res. 2018, 25, 29936–29942. [Google Scholar] [CrossRef]
- Lake, B.G.; Price, R.J.; Scott, M.P.; Chatham, L.R.; Vardy, A.; Osimitz, T.G. Piperonyl butoxide: Mode of action analysis for mouse liver tumour formation and human relevance. Toxicology 2020, 439, 152465. [Google Scholar] [CrossRef]
- Matsunaga, H.; Tomigahara, Y.; Kaneko, H.; Nakatsuka, I.; Yamane, S. Metabolism of pentyl 2-chloro-4-fluoro-5-(3,4,5,6-tetrahydrophthalimido)phenoxyacetate (flumiclorac pentyl, S-23031) in rats. 3. Identification of a reduced form metabolite of flumiclorac pentyl (S-23031) in rats. Nippon. Noyaku Gakkaishi 1997, 22, 133–135. [Google Scholar] [CrossRef] [Green Version]
- Soltani, N.; Brown, L.R.; Sikkema, P.H. Weed Control in White Bean with Pethoxamid Tank-Mixes Applied Preemergence. Int. J. Agron. 2018, 2018, 2402696. [Google Scholar] [CrossRef] [Green Version]
- Schlosser, H.G.; Hunt, B.; Teicher, H.B. Herbicidal Combination of Pethoxamid and Picloram. WO2014202092A1, 24 December 2014. [Google Scholar]
- Okamoto, H.; Kato, S.; Kobutani, T.; Ogasawara, M.; Konnai, M.; Takematsu, T. Herbicidally Active N-(1-Arylethenyl)-2-chloroacetamides Bearing an Alkyloxyalkyl Moiety. Agric. Biol. Chem. 1991, 55, 2737–2743. [Google Scholar] [CrossRef]
- Dhareesank, A.; Kobayashi, K.; Usui, K. Phytotoxic activity of pethoxamid in soil under different moisture conditions. Weed Biol. Manag. 2005, 5, 197–202. [Google Scholar] [CrossRef]
- Kumar, J.; Patel, A.; Tiwari, S.; Tiwari, S.; Srivastava, P.K.; Prasad, S.M. Pretilachlor toxicity is decided by discrete photo-acclimatizing conditions: Physiological and biochemical evidence from Anabaena sp. and Nostoc muscorum. Ecotoxicol. Environ. Saf. 2018, 156, 344–353. [Google Scholar] [CrossRef]
- Wu, C.; Lou, X.; Xu, X.; Huang, A.; Zhang, M.; Ma, L. Thermodynamics and Kinetics of Pretilachlor Adsorption on Organobentonites for Controlled Release. ACS Omega 2020, 5, 4191–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Lou, X.; Huang, A.; Zhang, M.; Ma, L. Thermodynamics and kinetics of pretilachlor adsorption: Implication to controlled release from organobentonites. Appl. Clay Sci. 2020, 190, 105566. [Google Scholar] [CrossRef]
- Swatch, G.K.; Singh, D.P.; Khattar, J.S.; Mohapatra, P.K. Interaction of pretilachlor with PS-II activity of the cyanobacterium Desmonostoc muscorum PUPCCC 405.10. J. Basic Microbiol. 2020, 60, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Kaur, P.; Bhullar, M.S. Persistence behaviour of pretilachlor in puddled paddy fields under subtropical humid climate. Environ. Monit. Assess. 2015, 187, 524. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, Y.; Yu, R.; Zhao, X.; Wang, Q.; Cai, L. Pretilachlor has the potential to induce endocrine disruption, oxidative stress, apoptosis and immunotoxicity during zebrafish embryo development. Environ. Toxicol. Pharmacol. 2016, 42, 125–134. [Google Scholar] [CrossRef]
- Diyanat, M.; Saeidian, H.; Baziar, S.; Mirjafary, Z. Preparation and characterization of polycaprolactone nanocapsules containing pretilachlor as a herbicide nanocarrier. Environ. Sci. Pollut. Res. 2019, 26, 21579–21588. [Google Scholar] [CrossRef]
- Chen, H.; Liu, X.; Wang, H.; Wu, S.; Li, J.; Jin, C.; Xu, H. Polyurea microencapsulate suspension: An efficient carrier for enhanced herbicidal activity of pretilachlor and reducing its side effects. J. Hazard. Mater. 2021, 402, 123744. [Google Scholar] [CrossRef]
- Chen, H.; Liu, X.; Deng, S.; Wang, H.; Ou, X.; Huang, L.; Li, J.; Jin, C. Pretilachlor Releasable Polyurea Microcapsules Suspension Optimization and Its Paddy Field Weeding Investigation. Front. Chem. 2020, 8, 826. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Ngoc, S.T.T.; Duong, D.; Le Ngoc, P. Effect of Pretilachlor on Weedy Rice and Other Weeds in Wet-Seeded Rice Cultivation in South Vietnam. Plant Prod. Sci. 2015, 17, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Liu, Y.; Li, Y.; Zhu, S.; Li, Y.; Yi, J.; Ouyang, Z.; Liu, B.; Mehmood, K.; Hussain, R.; et al. Exposure to the herbicide butachlor activates hepatic stress signals and disturbs lipid metabolism in mice. Chemosphere 2021, 283, 131226. [Google Scholar] [CrossRef]
- Lin, Z.; Pang, S.; Zhou, Z.; Wu, X.; Bhatt, P.; Chen, S. Current insights into the microbial degradation for butachlor: Strains, metabolic pathways, and molecular mechanisms. Appl. Microbiol. Biotechnol. 2021, 105, 4369–4381. [Google Scholar] [CrossRef] [PubMed]
- Matthes, B.; Böger, P. Chloroacetamides Affect the Plasma Membrane. Z. Naturforsch. C J. Biosci. 2002, 57, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Trenkamp, S.; Martin, W.; Tietjen, K. Specific and differential inhibition of very-long-chain fatty acid elongases from Arabidopsis thaliana by different herbicides. Proc. Natl. Acad. Sci. USA 2004, 101, 11903–11908. [Google Scholar] [CrossRef] [PubMed]
- Beste, C.E. Herbicide Handbook of the Weed Science Society of America, 5th ed.; Weed Science Society of America: Champaign, IL, USA, 1983. [Google Scholar]
- Reddy, S.R.; Babu, T.R.; Sreenivasulu, B. Electrochemical Investigation of carbonyl group containing pesticide monalide in water samples by using differential pulse adsorptive stripping voltammetry. Int. J. Res. Pharm. Life Sci. 2013, 1, 43–47. [Google Scholar]
- Li, L.; Ye, F. Photosynthesis inhibitor herbicides. Pestic. Sci. Adm. 2010, 31, 25–28. [Google Scholar]
- Morgan, W.C.; Stevenson, B.C.; Morgans, A.S. Selective post-emergence chemical control of Solanum nigrum L. and other broad leaved weeds in direct seeded processing tomatoes. J. Hortic. Sci. 1991, 66, 737–746. [Google Scholar] [CrossRef]
- Wu, C.-S.; Huang, J.-L.; Sun, Y.-S.; Yang, D.-Y. Mode of Action of 4-Hydroxyphenylpyruvate Dioxygenase Inhibition by Triketone-type Inhibitors. J. Med. Chem. 2002, 45, 2222–2228. [Google Scholar] [CrossRef]
- Raspail, C.; Graindorge, M.; Moreau, Y.; Crouzy, S.; Lefèbvre, B.; Robin, A.Y.; Dumas, R.; Matringe, M. 4-Hydroxyphenylpyruvate Dioxygenase Catalysis: Identification of catalytic residues and production of a hydroxylated intermediate shared with a structurally unrelated enzyme. J. Biol. Chem. 2011, 286, 26061–26070. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.L.; Knudsen, C.G.; Michaely, W.J.; Chin, H.-L.; Nguyen, N.H.; Carter, C.G.; Cromartie, T.H.; Lake, B.H.; Shribbs, J.M.; Fraser, T. The structure-activity relationships of the triketone class of p-hydroxyphenylpyruvate dioxygenase inhibiting herbicides. Pestic. Sci. 1998, 54, 377–384. [Google Scholar] [CrossRef]
- Kovaleva, E.G.; Lipscomb, J.D. Versatility of biological non-heme Fe(II) centers in oxygen activation reactions. Nat. Chem. Biol. 2008, 4, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, J.M.; Johnson-Winters, K.; Harrison, D.H.T.; Moran, G.R. Structure of the Ferrous Form of (4-Hydroxyphenyl)pyruvate Dioxygenase from Streptomyces avermitilis in Complex with the Therapeutic Herbicide, NTBC. Biochemistry 2004, 43, 6370–6377. [Google Scholar] [CrossRef]
- Beaudegnies, R.; Edmunds, A.J.; Fraser, T.E.; Hall, R.G.; Hawkes, T.R.; Mitchell, G.; Schaetzer, J.; Wendeborn, S.; Wibley, J. Herbicidal 4-hydroxyphenylpyruvate dioxygenase inhibitors—A review of the triketone chemistry story from a Syngenta perspective. Bioorg. Med. Chem. 2009, 17, 4134–4152. [Google Scholar] [CrossRef]
- Ahrens, H.; Lange, G.; Müller, T.; Rosinger, C.; Willms, L.; van Almsick, A. 4-Hydroxyphenylpyruvate Dioxygenase Inhibitors in Combination with Safeners: Solutions for Modern and Sustainable Agriculture. Angew. Chem. Int. Ed. 2013, 52, 9388–9398. [Google Scholar] [CrossRef]
- Shimoharada, H.; Tsukamoto, M.; Ikeguchi, M.; Kikugawa, H.; Sano, M.; Kitahara, Y.; Kominami, H.; Okita, T. Preparation of Benzoylpyrazole Derivatives as Herbicides. WO2007069771, 21 June 2007. [Google Scholar]
- Langdon, N.M.; Soltani, N.; Raedar, A.J.; Robinson, D.E.; Hooker, D.C.; Sikkema, P.H. Influence of Adjuvants on the Efficacy of Tolpyralate plus Atrazine for the Control of Annual Grass and Broadleaf Weeds in Corn with and without Roundup WeatherMAX. Am. J. Plant Sci. 2020, 11, 465–495. [Google Scholar] [CrossRef] [Green Version]
- Langdon, N.M.; Soltani, N.; Raedar, A.J.; Hooker, D.C.; Robinson, D.E.; Sikkema, P.H. Tolpyralate + Atrazine Applied Preemergence Provides Residual GR Canada Fleabane [Conyza canadensis (L.) Cronq.] Control Similar to Current Industry Standards. Agric. Sci. 2020, 11, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Metzger, B.A.; Soltani, N.; Raeder, A.J.; Hooker, D.C.; Robinson, D.E.; Sikkema, P.H. Effect of hybrid varieties, application timing, and herbicide rate on field corn tolerance to tolpyralate plus atrazine. Weed Sci. 2019, 67, 475–484. [Google Scholar] [CrossRef]
- Dayan, F.; Owens, D.K.; Duke, S. Rationale for a natural products approach to herbicide discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef]
- Malev, V.V.; Kaulin, Y.; Bezrukov, S.M.; Gurnev, P.; Takemoto, J.Y.; Shchagina, L.V. Kinetics of opening and closure of syringomycin E channels formed in lipid bilayers. Membr. Cell Biol. 2001, 14, 813–829. [Google Scholar]
- Chaimovitsh, D.; Abu-Abied, M.; Belausov, E.; Rubin, B.; Dudai, N.; Sadot, E. Microtubules are an intracellular target of the plant terpene citral. Plant J. 2010, 61, 399–408. [Google Scholar] [CrossRef]
- Li, M.; Di, X.; Jiang, Z. Enantioselective separation, analysis and stereoselective dissipation of the chiral pesticide cloquintocet-mexyl using a modified QuEChERS method by high-performance liquid chromatography tandem mass spectrometry. Chemosphere 2022, 291, 133084. [Google Scholar] [CrossRef]
- Dzardanov, D.V.; Elinevskaya, L.S.; Roldughin, V.I. The influence of the nature and composition of mixed surfactants on the stability of herbicide emulsions based on fenoxaprop-P-ethyl and cloquintocet-mexyl. Colloid J. 2014, 76, 675–682. [Google Scholar] [CrossRef]
- Taylor, V.L.; Cummins, I.; Brazier-Hicks, M.; Edwards, R. Protective responses induced by herbicide safeners in wheat. Environ. Exp. Bot. 2013, 88, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Zhu, W.; Xu, X.; Zhou, Z.; Liu, D. Direct chiral resolution of cloquintocet-mexyl and its application to in vitro degradation combined with clodinafop-propargyl. Biomed. Chromatogr. 2011, 26, 1058–1061. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, H. International Tobacco Bud Inhibitors Overview. Tob. Sci. Technol. 1991, 38–39. [Google Scholar]
- Cai, K.; Li, Z.; Jin, A. Preliminary report of n-decanol inhibition test on tobacco buds. Tob. Sci. Technol. 1986, 16. [Google Scholar] [CrossRef]
- Karam, E.A.; Keramat, B. Foliar spray of triacontanol improves growth by alleviating oxidative damage in coriander under salinity. Indian J. Plant Physiol. 2017, 22, 120–124. [Google Scholar] [CrossRef]
- Ahmad, J.; Ali, A.A.; Al-Huqail, A.A.; Qureshi, M.I. Triacontanol attenuates drought-induced oxidative stress in Brassica juncea L. by regulating lignification genes, calcium metabolism and the antioxidant system. Plant Physiol. Biochem. 2021, 166, 985–998. [Google Scholar] [CrossRef]
- Naeem, M.; Khan, M.M.A.; Moinuddin. Triacontanol: A potent plant growth regulator in agriculture. J. Plant Interact. 2012, 7, 129–142. [Google Scholar] [CrossRef]
- Ali, H.M.M.; Perveen, S. Effect of foliar applied triacontanol on wheat (Triticum aestivum L.) under arsenic stress: A study of changes in growth, yield and photosynthetic characteristics. Physiol. Mol. Biol. Plants 2020, 26, 1215–1224. [Google Scholar] [CrossRef]
- Zaid, A.; Mohammad, F.; Fariduddin, Q. Plant growth regulators improve growth, photosynthesis, mineral nutrient and antioxidant system under cadmium stress in menthol mint (Mentha arvensis L.). Physiol. Mol. Biol. Plants 2020, 26, 25–39. [Google Scholar] [CrossRef]
- Islam, S.; Zaid, A.; Mohammad, F. Role of Triacontanol in Counteracting the Ill Effects of Salinity in Plants: A Review. J. Plant Growth Regul. 2020, 40, 1–10. [Google Scholar] [CrossRef]
- Borowski, E.; Blamowski, Z.K. The effects of triacontanol ‘TRIA’ and Asahi SL on the development and metabolic activity of sweet basil (Ocimum basilicum L.) plants treated with chilling. Folia Hortic. 2009, 21, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Perveen, S.; Shahbaz, M.; Ashraf, M. Influence of foliar-applied triacontanol on growth, gas exchange characteristics, and chlorophyll fluorescence at different growth stages in wheat under saline conditions. Photosynthetica 2013, 51, 541–551. [Google Scholar] [CrossRef]
- Perveen, S.; Iqbal, M.; Parveen, A.; Akram, M.S.; Shahbaz, M.; Akber, S.; Mehboob, A. Exogenous triacontanol-mediated increase in phenolics, proline, activity of nitrate reductase, and shoot k+ confers salt tolerance in maize (Zea mays L.). Braz. J. Bot. 2016, 40, 1–11. [Google Scholar] [CrossRef]
- Coleman, R.; Penner, D. Organic Acid Enhancement of Pelargonic Acid. Weed Technol. 2008, 22, 38–41. [Google Scholar] [CrossRef]
- Ciriminna, R.; Fidalgo, A.; Ilharco, L.M.; Pagliaro, M. Herbicides based on pelargonic acid: Herbicides of the bioeconomy. Biofuels, Bioprod. Biorefining 2019, 13, 1476–1482. [Google Scholar] [CrossRef]
- Williams, M.W. New Chemical Approaches for Control of Biennial Bearing of Apples. ACS Symp. Ser. 1994, 557, 16–25. [Google Scholar] [CrossRef]
- Muñoz, M.; Torres-Pagán, N.; Peiró, R.; Guijarro, R.; Sánchez-Moreiras, A.M.; Verdeguer, M. Phytotoxic Effects of Three Natural Compounds: Pelargonic Acid, Carvacrol, and Cinnamic Aldehyde, against Problematic Weeds in Mediterranean Crops. Agronomy 2020, 10, 791. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural Compounds as Next-Generation Herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [Green Version]
- Azis, H.R.; Takahashi, S.; Koshiyama, M.; Fujisawa, H.; Isoda, H. Effect of Prohydrojasmon on the Growth of Eggplant and Komatsuna. Plants 2020, 9, 1368. [Google Scholar] [CrossRef]
- Azis, H.R.; Etteieb, S.; Takahashi, S.; Koshiyama, M.; Fujisawa, H.; Isoda, H. Effect of prohydrojasmon on total phenolic content, anthocyanin accumulation and antioxidant activity in komatsuna and lettuce. Biosci. Biotechnol. Biochem. 2020, 84, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Pirbalouti, A.G.; Sajjadi, S.E.; Parang, K. A Review (Research and Patents) on Jasmonic Acid and Its Derivatives. Arch. Pharm. 2014, 347, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, C.L.; Erwin, J.E. Horticultural applications of jasmonates: A review. J. Hortic. Sci. Biotechnol. 2008, 83, 283–304. [Google Scholar] [CrossRef]
- Creelman, R.; Mullet, J. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef]
- Takahashi, S.; Namioka, Y.; Azis, H.R.; Sano, T.; Aono, M.; Koshiyama, M.; Fujisawa, H.; Isoda, H. Prohydrojasmon Promotes the Accumulation of Phenolic Compounds in Red Leaf Lettuce. Plants 2021, 10, 1920. [Google Scholar] [CrossRef]
- Sato, K.; Ikoma, Y. Improvement in Handpicking Efficiency of Satsuma Mandarin Fruit with Combination Treatments of Gibberellin, Prohydrojasmon and Ethephon. Hortic. J. 2017, 86, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Román-Hidalgo, C.; Villar-Navarro, M.; Falcón-García, G.; Carbonero-Aguilar, M.; Bautista-Palomas, J.; Bello-López, M.; Martín-Valero, M.; Fernández-Torres, R. Selective, rapid and simultaneous determination of ergosterol and ergocalciferol in mushrooms by UPLC-Q-TOF-MS. J. Pharm. Biomed. Anal. 2020, 194, 113748. [Google Scholar] [CrossRef]
- Greaves, J.H.; Redfern, R.; King, R.E. Some properties of calciferol as a rodenticide. Epidemiol. Infect. 1974, 73, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Harris, W.D.; Zukel, J.W. Chemical Structure-Activity Relationship, Chemical Structure of Series of Organic Sulfites and Its Toxicity to Two-Spotted Spider Mite. J. Agric. Food Chem. 1954, 2, 140–142. [Google Scholar] [CrossRef]
- Maxwell, K.E.; Piper, V.D. Molecular Structure of Nonionic Surfactants in Relation to Laboratory Insecticidal Activity. J. Econ. Entomol. 1968, 61, 1633–1636. [Google Scholar] [CrossRef]
- Santos, F.O.; Lima, H.G.; Rosa, S.D.S.S.; das Mercês, N.B.; Serra, T.M.; Uzeda, R.S.; Reis, I.M.A.; Botura, M.B.; Branco, A.; Batatinha, M.J.M. In vitro acaricide and anticholinesterase activities of digitaria insularis (Poaceae) against Rhipicephalus (Boophilus) microplus. Vet. Parasitol. 2018, 255, 102–106. [Google Scholar] [CrossRef]
- Peterson, G.S.; Kandil, M.A.; Abdallah, M.D.; Farag, A.-A.A. Isolation and characterisation of biologically-active compounds from some plant extracts. Pestic. Sci. 1989, 25, 337–342. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Molina, S.C.; Muñoz, E.; Lamilla, C.; Alarcon, J.; Palacios, S.M.; Carpinella, M.C.; Avila, J.G. The insecticidal, molting disruption and insect growth inhibitory activity of extracts from Condalia microphylla Cav. (Rhamnaceae). Ind. Crops Prod. 2013, 42, 78–86. [Google Scholar] [CrossRef]
- Zhu, J.; Li, Z.; Lu, H.; Liu, S.; Ding, W.; Li, J.; Xiong, Y.; Li, C. New diphenyl ethers from a fungus Epicoccum sorghinum L28 and their antifungal activity against phytopathogens. Bioorg. Chem. 2021, 115, 105232. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Classification of long-chain molecules in this article.

Figure 2.
The structures of compounds 1 and 2.

Figure 3.
The structures of compounds 3a and 3b.

Figure 4.
The structures of compounds 4a–4d.

Figure 5.
The structure of compound 5.

Figure 6.
The structures of compounds 6–8.

Figure 7.
The structures of compounds 9 and 10.

Figure 8.
The structures of compounds 11 and 12.

Figure 9.
The structures of compounds 13–15.

Figure 10.
The structures of compounds 16a–16q.

Figure 11.
The structures of compounds 17 and 18.

Figure 12.
The structure of QAS compounds.

Figure 13.
The structures of compounds 20a–20i.

Figure 14.
The structures of compounds 21 and 22.

Figure 15.
The structures of compounds 23–26.

Figure 16.
The structures of compounds 27–29.

Figure 17.
The structure of compound 30.

Figure 18.
The structures of compounds 31–33.

Figure 19.
The structures of compounds 34–36.

Figure 20.
The structures of compounds 37–40.

Figure 21.
The structures of compounds 41–43.

Figure 22.
The structures of compounds 44 and 45.

Figure 23.
The structure of compound 46.

Figure 24.
The structures of compounds 47–50.

Figure 25.
The structures of compounds 51 and 52.

Figure 26.
The structures of compounds 53 and 54.

Figure 27.
The structures of compounds 55–57.

Figure 28.
The structure of compound 58.

Figure 29.
The structures of compounds 59–63.

Figure 30.
The structure of compound 64.

Figure 31.
The structures of compounds 65 and 66.

Figure 32.
The structures of compounds 67a–67g.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yin, F.; Qin, Z. Long-Chain Molecules with Agro-Bioactivities and Their Applications. Molecules 2023, 28, 5880. https://doi.org/10.3390/molecules28155880
AMA Style
Yin F, Qin Z. Long-Chain Molecules with Agro-Bioactivities and Their Applications. Molecules. 2023; 28(15):5880. https://doi.org/10.3390/molecules28155880
Chicago/Turabian StyleYin, Fahong, and Zhaohai Qin. 2023. "Long-Chain Molecules with Agro-Bioactivities and Their Applications" Molecules 28, no. 15: 5880. https://doi.org/10.3390/molecules28155880