
Antispasmodic Effect of Alstonia boonei De Wild. and Its Constituents: Ex Vivo and In Silico Approaches

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
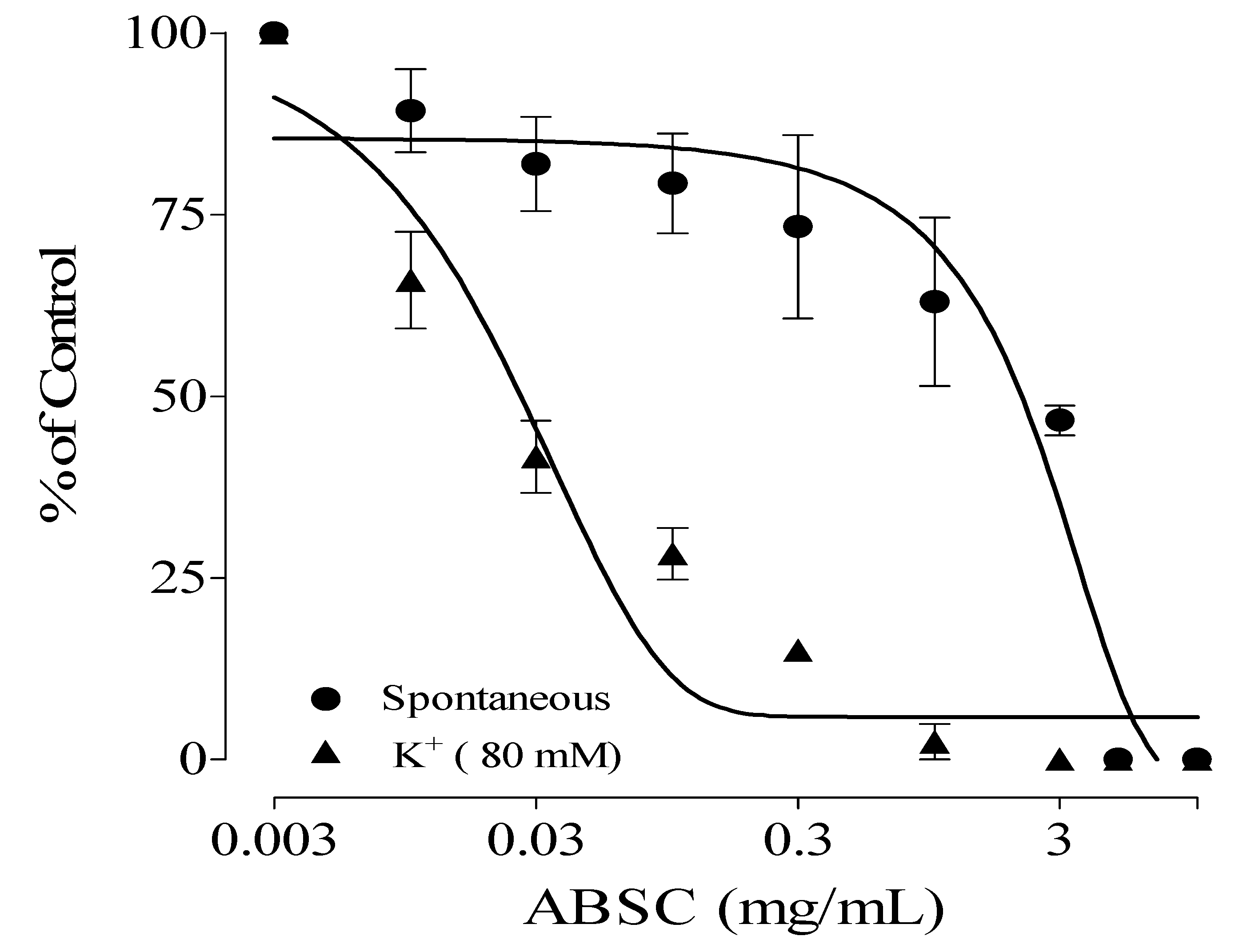
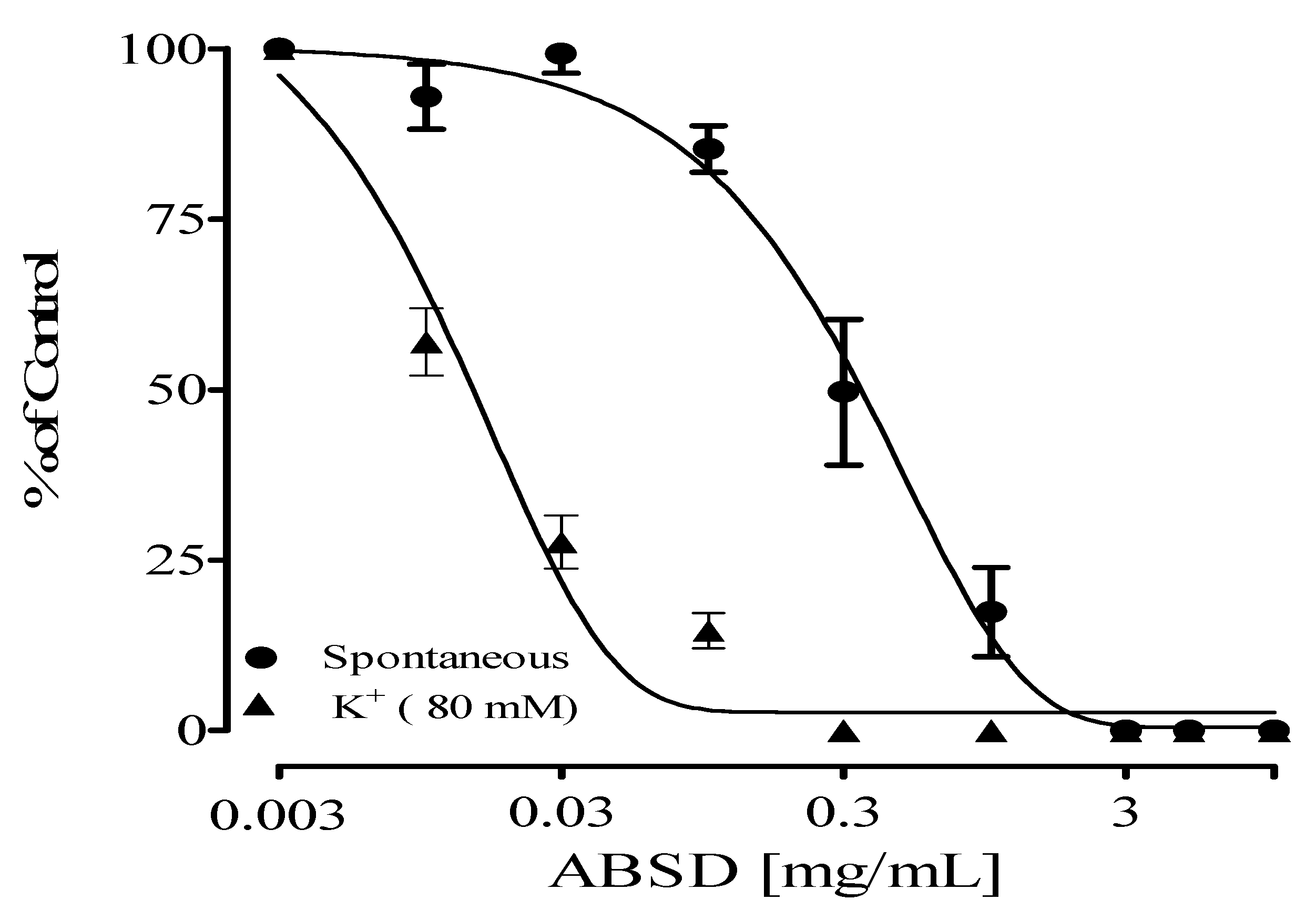
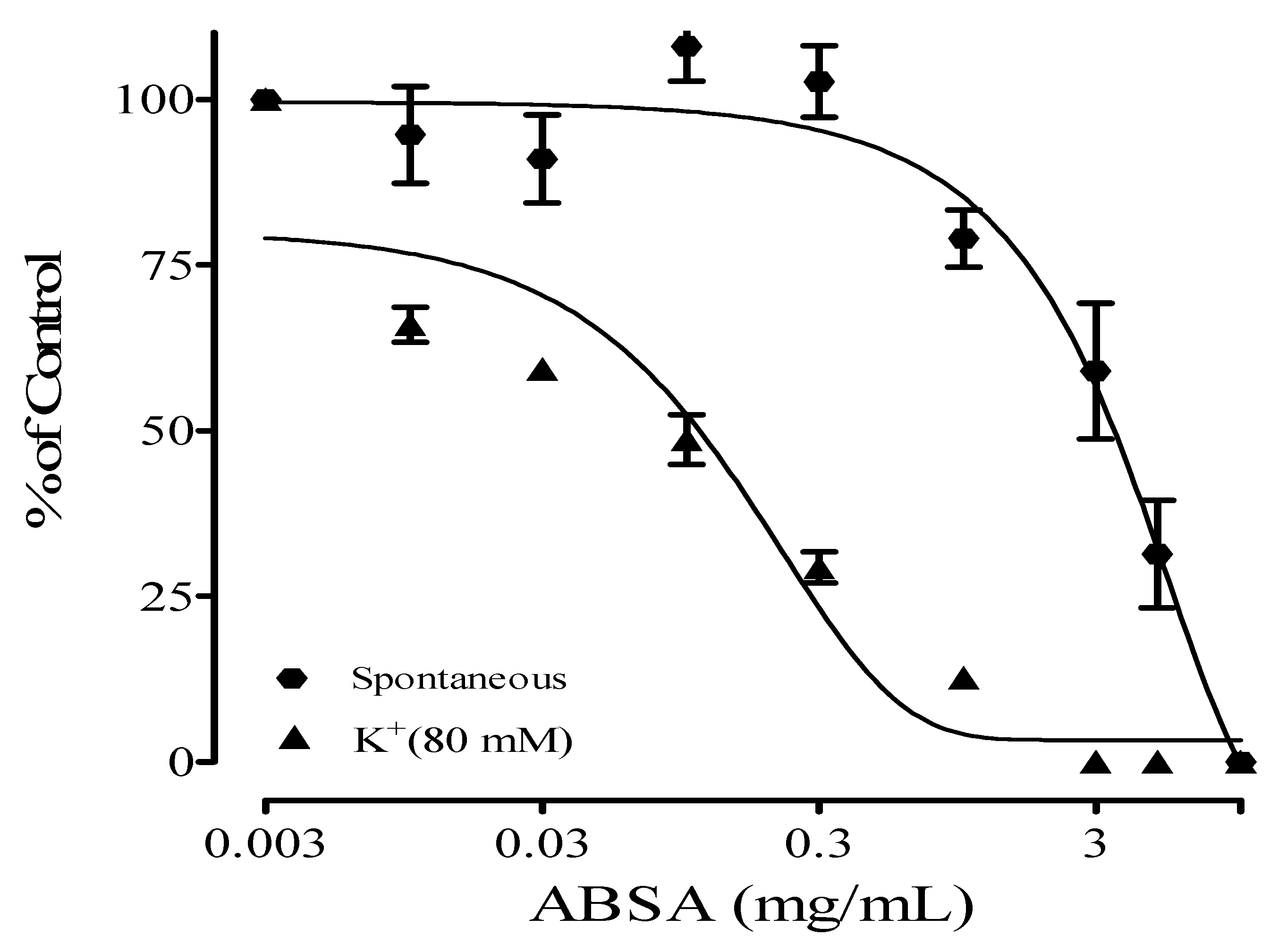
2.1. Antispasmodic Effect of Crude Extract and Fractions of Alstonia boonei
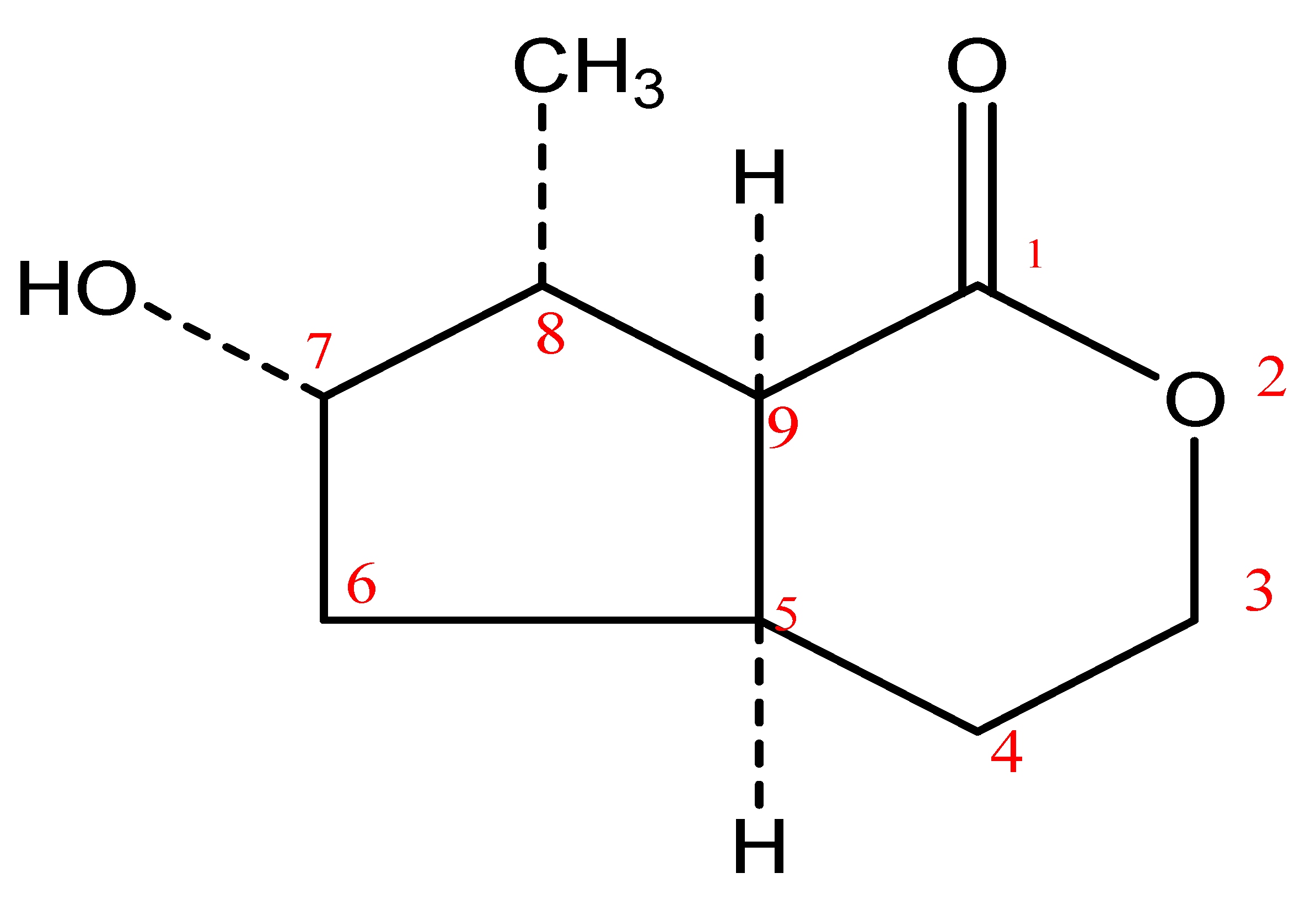
2.2. Structural Elucidation of Compounds 1 and 2
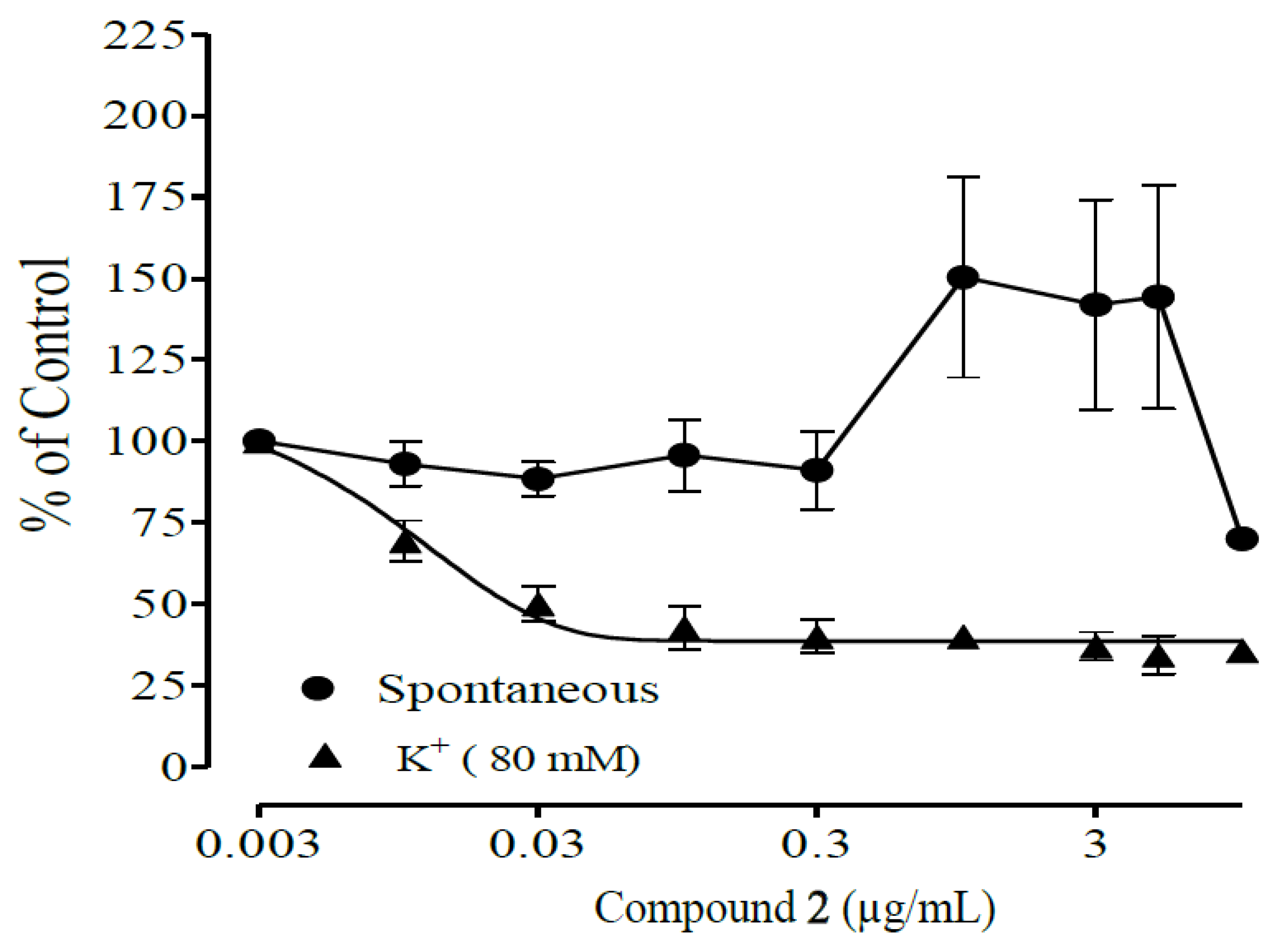
2.3. Antispasmodic Studies on Compounds 1 and 2
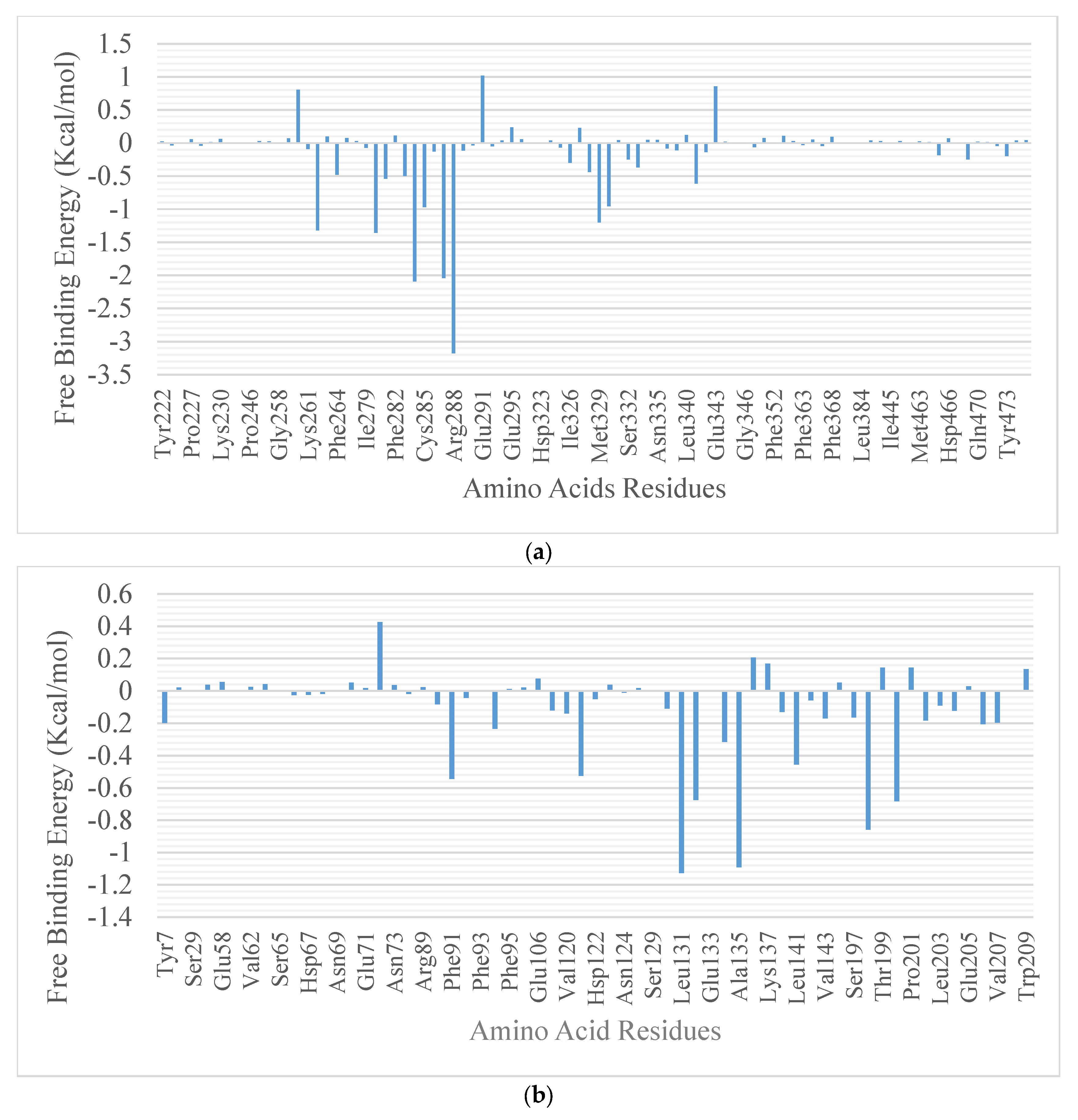
2.4. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plant Material, Extraction, and Partitioning
4.2. Ex Vivo Antispasmodic Assay
4.2.1. Experimental Animals for Antispasmodic Assay
4.2.2. Antispasmodic Effect of Crude Extract and Fractions
4.3. Isolation of Compounds 1 and 2 from A. boonei
4.4. Structure Elucidation of Compounds 1 and 2
4.5. In Silico Studies
4.5.1. Retrieval and Preparation of Proteins
4.5.2. Ligands Preparation
4.5.3. Targeted Molecular Docking
4.5.4. Molecular Dynamics
4.5.5. Binding Free Energy Calculation Using MM-GBSA
4.5.6. Clustering of Molecular Dynamic Trajectory
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Osim, E.E. Elements of Gastrointestinal Tract Physiology; Helimo Associates Publishers: Calabar, Nigeria, 2002. [Google Scholar]
- Jackquelyn, C. Anticholinergics. Health Line Reference Library Publication. 2013. Available online: www.healthline.com (accessed on 12 December 2022).
- Choung, R.S.; Rey, E.; Richard Locke, G., III; Schleck, C.D.; Baum, C.; Zinsmeister, A.R.; Talley, N.J. Chronic constipation and co-morbidities: A prospective population-based nested case-control study. United Eur. Gastroenterol. J. 2016, 4, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Ibeh, R.C.; Ikechukwu, G.C.; Ijioma, S.N.; Singh, A.K. Antispasmodic and antidiarrhoeal activity of Newbouldia laevis, Cola nitida and Acanthus montanus leaf extracts on gastrointestinal smooth muscle. J. Pharmacol. Toxicol. 2018, 14, 1–8. [Google Scholar] [CrossRef]
- Alam, F.; Najam, Q.S.; Shah, A.J.; Ashraf, M.; Al Ain, Q. Gut modulatory and butyrylcholinesterase inhibitory activities of Gaultheria trichophylla. Pharm. Biol. 2016, 54, 2917–2921. [Google Scholar] [CrossRef]
- Khana, A.U.; Gilania, A.H. Antispasmodic and bronchodilator activities of Artemisia vulgaris are mediated through dual blockade of muscarinic receptors and calcium influx. J. Ethnopharmacol. 2009, 126, 480–486. [Google Scholar]
- Kitazawa, T.; Kaiya, H. Regulation of gastrointestinal motility by motilin and ghrelin in vertebrates. Front. Endocrinol. 2019, 10, 278. [Google Scholar] [CrossRef] [PubMed]
- Muralikumar, S.; Vetrivel, U.; Narayanasamy, A.; Das, U.N. Probing the intermolecular interactions of PPARγ-LBD with polyunsaturated fatty acids and their anti-inflammatory metabolites to infer most potential binding moieties. Lipids Health Dis. 2017, 16, 1–11. [Google Scholar] [CrossRef]
- Christofides, A.; Konstantinidou, E.; Jani, C.; Boussiotis, V.A. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism 2021, 114, 154338. [Google Scholar] [CrossRef]
- Laganà, A.S.; Vitale, S.G.; Nigro, A.; Sofo, V.; Salmeri, F.M.; Rossetti, P.; Rapisarda, A.M.C.; La Vignera, S.; Condorelli, R.A.; Rizzo, G.; et al. Pleiotropic Actions of Peroxisome Proliferator-Activated Receptors (PPARs) in Dysregulated Metabolic Homeostasis, Inflammation and Cancer: Current Evidence and Future Perspectives. Int. J. Mol. Sci. 2016, 17, 999. [Google Scholar] [CrossRef] [PubMed]
- Chadt, A.; Al-Hasani, H. Glucose transporters in adipose tissue, liver, and skeletal muscle in metabolic health and disease. Pflug. Arch. Eur. J. Physiol. 2020, 472, 1273–1298. [Google Scholar] [CrossRef]
- Decara, J.; Rivera, P.; López-Gambero, A.J.; Serrano, A.; Pavón, F.J.; Baixeras, E.; De Fonseca, F.R.; Suárez, J. Peroxisome Proliferator-Activated Receptors: Experimental Targeting for the Treatment of Inflammatory Bowel Diseases. Front. Pharmacol. 2020, 11, 730. [Google Scholar] [CrossRef]
- Nadeem, M.S.; Kumar, V.; Al-Abbasi, F.A.; Kamal, M.A.; Anwar, F. Risk of colorectal cancer in inflammatory bowel diseases. Semin. Cancer Biol. 2020, 64, 51–60. [Google Scholar]
- Ning, L.; Lou, X.; Zhang, F.; Xu, G. Nuclear Receptors in the Pathogenesis and Management of Inflammatory Bowel Disease. Mediat. Inflamm. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Klepsch, V.; Moschen, A.R.; Tilg, H.; Baier, G.; Hermann-Kleiter, N. Nuclear receptors regulate intestinal inflammation in the context of IBD. Front. Immunol. 2019, 10, 1070. [Google Scholar] [PubMed]
- Lian, J.; Casari, I.; Falasca, M. Modulatory role of the endocannabinoidome in the pathophysiology of the gastrointestinal tract. Pharmacol. Res. 2022, 175, 106025. [Google Scholar]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Angeli, A.; Carta, F.; Supuran, C.T. Carbonic Anhydrases: Versatile and Useful Biocatalysts in Chemistry and Biochemistry. Catalysts 2020, 10, 1008. [Google Scholar] [CrossRef]
- Nocentini, A.; Donald, W.A.; Supuran, C.T. Human carbonic anhydrases: Tissue distribution, physiological role, and druggability. In Carbonic Anhydrases; Academic Press: Cambridge, MA, USA, 2019; pp. 151–185. [Google Scholar]
- Bua, S.; Haapanen, S.; Kuuslahti, M.; Parkkila, S.; Supuran, C.T. Activation Studies of the β-Carbonic Anhydrase from the Pathogenic Protozoan Entamoeba histolytica with Amino Acids and Amines. Metabolites 2019, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Kakakhan, C.; Türkeş, C.; Güleç, Ö.; Demir, Y.; Arslan, M.; Özkemahlı, G.; Beydemir, Ş. Exploration of 1, 2, 3-triazole linked benzenesulfonamide derivatives as isoform selective inhibitors of human carbonic anhydrase. Bioorg. Med. Chem. 2023, 77, 117111. [Google Scholar] [CrossRef]
- Aspatwar, A.; Supuran, C.T.; Waheed, A.; Sly, W.S.; Parkkila, S. Mitochondrial carbonic anhydrase VA and VB: Prop-erties and roles in health and disease. J. Physiol. 2023, 601, 257–274. [Google Scholar] [CrossRef]
- Albrahim, T.; Alonazi, M.A. Lycopene corrects metabolic syndrome and liver injury induced by high fat diet in obese rats through antioxidant, anti-inflammatory, antifibrotic pathways. BioMedicine 2021, 141, 111831. [Google Scholar] [CrossRef]
- Hu, T.; Zhang, J.; Wang, J.; Sha, L.; Xia, Y.; Ortyl, T.C.; Tian, X.; Chen, L. Advances in Epilepsy: Mechanisms, Clinical Trials, and Drug Therapies. J. Med. Chem. 2023, 66, 4434–4467. [Google Scholar] [CrossRef] [PubMed]
- Sanghavi, S.F.; Swenson, E.R. Arterial Blood Gases and Acid-Base Regulation. In Seminars in Respiratory and Critical Care Medicine; Thieme Medical Publishers, Inc.: New York, NY, USA, 2023. [Google Scholar]
- Kaczmarczyk, O.; Dąbek-Drobny, A.; Piątek-Guziewicz, A.; Woźniakiewicz, M.; Paśko, P.; Dobrowolska-Iwanek, J.; Woźniakiewicz, A.; Targosz, A.; Ptak-Belowska, A.; Szczyrk, U.; et al. The Importance of Nutritional Aspects in the Assessment of In-flammation and Intestinal Barrier in Patients with Inflammatory Bowel Disease. Nutrients 2022, 14, 4622. [Google Scholar] [CrossRef] [PubMed]
- Hadley, S.K.; Gaarder, S.M. Treatment of irritable bowel syndrome. Am. Fam. Physician 2005, 72, 2501–2506. [Google Scholar] [PubMed]
- Bayad, A.E. The antidiarrhoeal activity and phytoconstituents of the methanol extracts of Tecurium oliverianum. Glob. Vet. 2016, 16, 93–99. [Google Scholar]
- Sadraei, H.; Ghannadi, A.; Malekshahi, K. Relaxant effect of essential oil of Melissa officinalis and citral oil on rat ileum contractions. Fitoterapia 2003, 74, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Broadley, K.J.; Kelly, D.R. Muscarinic Receptor Agonists and Antagonists. Molecules 2001, 6, 142–193. [Google Scholar] [CrossRef]
- Cai, X.-H.; Du, Z.-Z.; Luo, X.-D. Unique Monoterpenoid Indole Alkaloids from Alstonia scholaris. Org. Lett. 2007, 9, 1817–1820. [Google Scholar] [CrossRef]
- Cai, X.-H.; Tan, Q.-G.; Liu, Y.-P.; Feng, T.; Du, Z.-Z.; Li, W.-Q.; Luo, X.-D. ChemInform Abstract: A Cage-Monoterpene Indole Alkaloid from Alstonia scholaris. Org. Lett. 2008, 10, 577–580. [Google Scholar] [CrossRef]
- Wang, F.; Ren, F.-C.; Liu, J.-K. Alstonic acids A and B, unusual 2,3-secofernane triterpenoids from Alstonia scholaris. Phytochemistry 2009, 70, 650–654. [Google Scholar] [CrossRef]
- Zhou, H.; He, H.-P.; Luo, X.-D.; Wang, Y.-H.; Yang, X.-W.; Di, Y.-T.; Hao, X.-J. Three New Indole Alkaloids from the Leaves of Alstonia scholaris. Helvetica Chim. Acta 2005, 88, 2508–2512. [Google Scholar] [CrossRef]
- Idowu, O.A.; Soniran, O.T.; Ajana, O.; Aworinde, D.O. Ethnobotanical survey of antimalarial plants used in Ogun State, Southwest Nigeria. Afr. J. Pharm. Pharmacol. 2010, 4, 055–060. [Google Scholar]
- Olajide, O.A.; Awe, S.; Makinde, J.; I Ekhelar, A.; Olusola, A.; Morebise, O.; Okpako, D.T. Studies on the anti-inflammatory, antipyretic and analgesic properties of Alstonia boonei Stem back. J. Ethnopharmacol. 2000, 71, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Majekodunmi, S.; Adegoke, O.; Odeku, O. Formulation of the extract of the Stem back of Alstonia boonei as tablet dosage form. Trop. J. Pharm. Res. 2008, 7, 987–994. [Google Scholar] [CrossRef]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Anthony, S. Agroforestree Database: A Tree Reference and Selection Guide Version 402009. 2019. Available online: http://www.worldagroforestry.org/publication/agroforestree-database-tree-reference-andselection-guide-version-40 (accessed on 12 December 2022).
- Science Technology and Policy Research Institute (STEPRI); Centre for Scientific and Industrial Research (CSIR). Ghana Herbal Pharmacopoeia, 2nd ed.; CSIR-INSTI: Accra, Ghana, 2015. [Google Scholar]
- Fakae, B.B.; Campbell, A.M.; Barrett, J.; Scott, I.M.; Teesdale-Spittle, P.H.; Liebau, E.; Brophy, P.M. Inhibition of glutathione S-transferases (GSTs) from parasitic nematodes by extracts from traditional Nigerian medicinal plants. Phytother. Res. 2000, 14, 630–634. [Google Scholar] [CrossRef]
- Marini-Bettolo, G.B.; Nicoletti, M.; Messana, I.; Patamia, M.; Galeffi, C.; Oguakwa, J.U.; Portalone, G.; Vaciago, A. Re-search on African medicinal plants—IV: Boonein, A new C-9 terpenoid lactone from Alstonia boonei: A possible precursor in the indole alkaloid biogenesis. Tetrahedron 1983, 39, 323–329. [Google Scholar] [CrossRef]
- Okoye, N.N.; Ajaghaku, D.L.; Okeke, H.N.; Ilodigwe, E.E.; Nworu, C.S.; Okoye, F.B.C. Beta-amyrin and Al-pha-amyrin acetate isolated from the Stem back of Alstonia boonei display profound anti-inflammatory activity. Pharm. Biol. 2014, 52, 1478–1486. [Google Scholar] [CrossRef] [PubMed]
- Makrane, H.; Aziz, M.; Mekhfi, H.; Ziyyat, A.; Bnouham, M.; Legssyer, A.; Gressier, B.; Eto, B. Antispasmodic and Myorelaxant Activity of Organic Fractions from Origanum majorana L. on Intestinal Smooth Muscle of Rodents. Eur. J. Med. Plants 2018, 23, 1–11. [Google Scholar] [CrossRef]
- Bashir, S.; Janbaz, K.H.; Jabeen, Q.; Gilani, A.H. Studies on spasmogenic and spasmolytic activities of Calendula officinalis flowers. Phytother. Res. 2006, 20, 906–910. [Google Scholar] [CrossRef]
- Taiwo, O.B.; Kroes, B.H.; Beukelman, C.J.; Horsten ST, A.J.; Makinde, J.M.; Labadie, R.P. Activity of the stem-bark extract of Alstonia boonei De-Wild. (Apocynaceae) on human complement and polymorphonuclear leukocytes. J. Ethnopharmacol. 1998, 17, 13–15. [Google Scholar]
- Karaki, H.; Weiss, G.B. Calcium release in smooth muscle. Life Sci. 1988, 42, 111–122. [Google Scholar] [CrossRef]
- Godfraind, T.; Robert, M.; Maurice, W. Calcium antagonism and calcium entry blockade. Pharmacol. Rev. 1986, 38, 321–416. [Google Scholar] [PubMed]
- Ojewole, J.A.O. Studies on the pharmacology of echitamine, an alkaloid from the Stem back of Alstonia boonei L. (Apocynaceae). Int. J. Crude Drug Res. 1984, 22, 121–143. [Google Scholar] [CrossRef]
- Martin, M.L.; Diaz, M.T.; Montero, M.J.; Prieto, P.; San Roman, L.; Cortes, D. Antispasmodic activity of benzylisoquinoline alkaloids analogous to papaverine. Planta Medica 1993, 59, 63–67. [Google Scholar] [CrossRef]
- De Urbina, A.O.; Martin, M.L.; Fernandez, B.; San Roman, L.; Cubillo, L. In vitro antispasmodic activity of peracetylated penstemonoside, aucubin and catalpol. Planta Medica 1994, 60, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Hiller, K.O.; Ghorbani, M.; Schilcher, H. Antispasmodic and relaxant activity of chelidonine, protopine, coptisine, and chelidonium majus extracts on isolated Guinea-pig lleum. Planta Medica 1998, 64, 758–760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xian, Y.-F.; Ip, S.-P.; Fong, H.H.; Che, C.-T. A new and weakly antispasmodic protoberberine alkaloid from Rhizoma Coptidis. Phytotherapy Res. 2010, 24, 1414–1416. [Google Scholar] [CrossRef]
- Pongprayoon, U.; Baeckström, P.; Jacobsson, U.; Lindström, M.; Bohlin, L. Antispasmodic activity of β-damascenone and E-phytol isolated from Ipomoea pes-caprae. Planta Medica 1992, 58, 19–21. [Google Scholar] [CrossRef]
- Corea, G.; Fattorusso, E.; Lanzotti, V.; Capasso, R.; Izzo, A.A. Antispasmodic Saponins from Bulbs of Red Onion, Allium cepa L. Var. Tropea. J. Agric. Food Chem. 2005, 53, 935–940. [Google Scholar] [CrossRef]
- Fleer, H.; Verspohl, E. Antispasmodic activity of an extract from Plantago lanceolata L. and some isolated compounds. Phytomedicine 2007, 14, 409–415. [Google Scholar] [CrossRef]
- Zoofishan, Z.; Kúsz, N.; Csorba, A.; Tóth, G.; Hajagos-Tóth, J.; Kothencz, A.; Gáspár, R.; Hunyadi, A. Antispasmodic Activity of Prenylated Phenolic Compounds from the Root Bark of Morus nigra. Molecules 2019, 24, 2497. [Google Scholar] [CrossRef]
- Karplus, M.; McCammon, J.A. Molecular dynamics simulations of biomolecules. Nat. Struct. Mol. Biol. 2002, 9, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ivanov, I. Molecular dynamics. Comput. Toxicol. 2012, 1, 243–285. [Google Scholar]
- Dong, Y.W.; Liao, M.I.; Meng, X.I.; Somero, G.N. Structural flexibility and protein adaptation to temperature: Molecular dynamics analysis of malate dehydrogenases of marine molluscs. Proc. Natl. Acad. Sci. USA 2018, 115, 1274–1279. [Google Scholar] [CrossRef]
- Sinha, S.; Wang, S.M. Classification of VUS and unclassified variants in BRCA1 BRCT repeats by molecular dynamics simulation. Comput. Struct. Biotechnol. J. 2020, 18, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Morrone, J.A.; Simmerling, C.; A Dill, K. Advances in free-energy-based simulations of protein folding and ligand binding. Curr. Opin. Struct. Biol. 2016, 36, 25–31. [Google Scholar] [CrossRef]
- Gilani, S.A.; Kikuchi, A.; Shinwari, Z.K.; Khattak, Z.I.; Watanabe, K.N. Phytochemical, pharmacological and ethnobotanical studies of Rhazya stricta Decne. Review article. Phytother. Res. 2007, 4, 301–307. [Google Scholar] [CrossRef]
- Farre, A.; Colombo, M.; Fort, M.; Gutierrez, B. Differential effects of various Ca2+ antagonists. Gen. Pharmacol. Vasc. Syst. 1991, 22, 177–181. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and Auto-DockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Istyastono, E.P.; Radifar, M.; Yuniarti, N.; Prasasty, V.D.; Mungkasi, S. PyPLIF HIPPOS: A molecular interaction fingerprinting tool for docking results of AutoDock Vina and plants. J. Chem. Inf. Model. 2020, 60, 3697–3702. [Google Scholar] [CrossRef]
- Bekker, H.; Berendsen, H.J.C.; Dijkstra, E.J.; Achterop, S.; Vondrumen, R.V.; Vanderspoel, D.; Sijbers, A.; Keegstra, H.; Renardus, M.K.R. Gromacs-a parallel computer for molecular-dynamics simulations. In 4th International Conference on Computational Physics (PC 92), June 1992; World Scientific Publishing: Singapore, 1993; pp. 252–256. [Google Scholar]
- Oostenbrink, C.; Villa, A.; Mark, A.E.; Van Gunsteren, W.F. A biomolecular force field based on the free enthalpy of hydration and solvation: The GROMOS force-field parameter sets 53A5 and 53A6. J. Comput. Chem. 2004, 25, 1656–1676. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Tresanco, M.E.; Valiente, P.A.; Moreno, E. gmx_MMPBSA: A New Tool to Perform End-State Free Energy Calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef]
- Xue, W.; Yang, F.; Wang, P.; Zheng, G.; Chen, Y.; Yao, X.; Zhu, F. What contributes to serotonin–norepinephrine reuptake inhibitors’ dual-targeting mechanism? The key role of transmembrane domain 6 in human serotonin and norepinephrine transporters revealed by molecular dynamics simulation. ACS Chem. Neurosci. 2018, 9, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Tuccinardi, T. What is the current value of MM/PBSA and MM/GBSA methods in drug discovery? Expert Opin. Drug Discov. 2021, 16, 1233–1237. [Google Scholar] [CrossRef]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | IC50 (mg/mL) | |
|---|---|---|
| Spontaneous | K+ (80 mM) | |
| ABSC | 1.15 ± 0.10 | 0.03 ± 0.20 |
| ABSD | 0.31 ± 0.02 | 0.02 ± 0.05 |
| ABSE | 0.35 ± 0.03 | 0.03 ± 0.14 |
| ABSA | 2.38 ± 0.65 | 0.90 ± 0.06 |
| Compounds | IC50 (µg/mL) | |
|---|---|---|
| Spontaneous | K+ (80 mM) | |
| 1 | 0.29 ± 0.05 | 0.09 ± 0.01 |
| 2 | 2.20 ± 0.70 | 0.90 ± 0.10 |
| Compounds | Protein | Hydrogen Bond Distance (Å) | Hydrophobic Interaction | ||
|---|---|---|---|---|---|
| Numbers | Interacted Residues | Numbers | Interacted Residues | ||
| Ref. inhibitor | 2 | Lys263 Cys285 Arg280 | 2 | Val339 Met364 Leu330 Phe363 Arg288 Leu353 Ile341 Gly284 Ile281 Met348 Ile262 | |
| β-amyrin | 4F9M | 5 | 3 | Phe287 Lys263 Ile281 Ile262 Leu255 Met348 Ile341 Arg288 Phe264 | |
| Boonein | 4 | Ser324 Ile281 | 8 | Arg288 Ile341 Met348(2) Ile262(3) Cys285 | |
| Ref. Inhibitor | 6 | Glu106 His119 Thr199 His96 Gn92 | 7 | Trp209 His69 Leu198 Leu131 Ala135 His67 Pro201 His64 | |
| β-amyrin | 5E2M | 3 | Ala135 Leu131 Phe91 His119 Val143 His94 Ala121 Val207 Trp209 His67 Leu198 Pro292 | ||
| Boonein | 4 | Thr199 His200 leu203 Gln92 | Leu198 Ala121 His119 Val143 Trp209 | ||
| SYSTEM | ΔVDWAALS | ΔEEL | ΔEGB | ΔESURF | ΔGGAS | ΔGSOLV | ΔTOTAL |
|---|---|---|---|---|---|---|---|
| 4F9M—β AMYRIN | −46.56 ± 3.18 | 3.66 ± 2.77 | 21.7 ± 3.39 | −5.82 ± 0.48 | −42.9 ± 4.18 | 15.88 ± 3.18 | −27.02 ± 2.61 |
| 4F9M—BOONEIN | −1.78 ± 3.65 | −1.41 ± 4.99 | 2.62 ± 6.24 | −0.29 ± 0.6 | −3.19 ± 7.48 | 2.34 ± 5.79 | −0.86 ± 2.33 |
| 5E2M—β AMYRIN | −26.86 ± 4.66 | −1.24 ± 6.84 | 18.83 ± 6.46 | −3.61 ± 0.66 | −28.1 ± 8.07 | 15.22 ± 6.29 | −12.88 ± 4.06 |
| 5E2M—BOONEIN | −6.3 ± 6.94 | −3.86 ± 7.56 | 6.89 ± 8.92 | −0.92 ± 0.99 | −10.17 ± 12.36 | 5.97 ± 8.19 | −4.19 ± 5.22 |
| Compound | Salt Bridges | Hydrophobic Interactions | Hydrogen Bonds | |||||
|---|---|---|---|---|---|---|---|---|
| Cluster Number | Number | Amino Acids | Number | Amino Acids | Number | Amino Acids | ||
| 4F9M | Boonein | C1 | 1 | K373 | 0 | None | 0 | None |
| C2 | 2 | R397–R443 | 0 | None | 2 | S394–R397 | ||
| C3 | 0 | None | 1 | V277 | 0 | None | ||
| C4 | 0 | None | 2 | P304–I409 | 0 | None | ||
| β-amyrin | C1 | 6 | I262–M329–L330–L333–I341–Y473 | 1 | Y473 | |||
| C2 | 5 | R280–F287–M329–L330 (2) | 0 | None | ||||
| C3 | 10 | I262–F264 (2)–I281–F287 (2)–I326–L330–L333–I341 | 0 | None | ||||
| C4 | 9 | I262 (2)–F264 (2)–R288 (2)–I341 (2)–L465 | 0 | None | ||||
| 5E2M | boonein | C1 | 0 | None | 0 | None | 0 | None |
| C2 | 0 | None | 2 | P175–F176 | 0 | None | ||
| C3 | 1 | K159 | 1 | F176 | 1 | N178 | ||
| C4 | 0 | None | 1 | F176 | 1 | T177 | ||
| β-amyrin | C1 | 2 | F91–L198 | |||||
| C2 | 5 | F91–L131–A135–L141–V207 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akinmurele, O.J.; Sonibare, M.A.; Elujoba, A.A.; Ogunlakin, A.D.; Yeye, O.E.; Gyebi, G.A.; Ojo, O.A.; Alanzi, A.R. Antispasmodic Effect of Alstonia boonei De Wild. and Its Constituents: Ex Vivo and In Silico Approaches. Molecules 2023, 28, 7069. https://doi.org/10.3390/molecules28207069
Akinmurele OJ, Sonibare MA, Elujoba AA, Ogunlakin AD, Yeye OE, Gyebi GA, Ojo OA, Alanzi AR. Antispasmodic Effect of Alstonia boonei De Wild. and Its Constituents: Ex Vivo and In Silico Approaches. Molecules. 2023; 28(20):7069. https://doi.org/10.3390/molecules28207069
Chicago/Turabian StyleAkinmurele, Opeyemi Josephine, Mubo Adeola Sonibare, Anthony A. Elujoba, Akingbolabo Daniel Ogunlakin, Oloruntoba Emmanuel Yeye, Gideon Ampoma Gyebi, Oluwafemi Adeleke Ojo, and Abdullah R. Alanzi. 2023. "Antispasmodic Effect of Alstonia boonei De Wild. and Its Constituents: Ex Vivo and In Silico Approaches" Molecules 28, no. 20: 7069. https://doi.org/10.3390/molecules28207069