
Bioactive Secondary Metabolites from Fungi of the Genus Cytospora Ehrenb. (Ascomycota)

 , , , and
, , , and
Abstract
:
1. Introduction
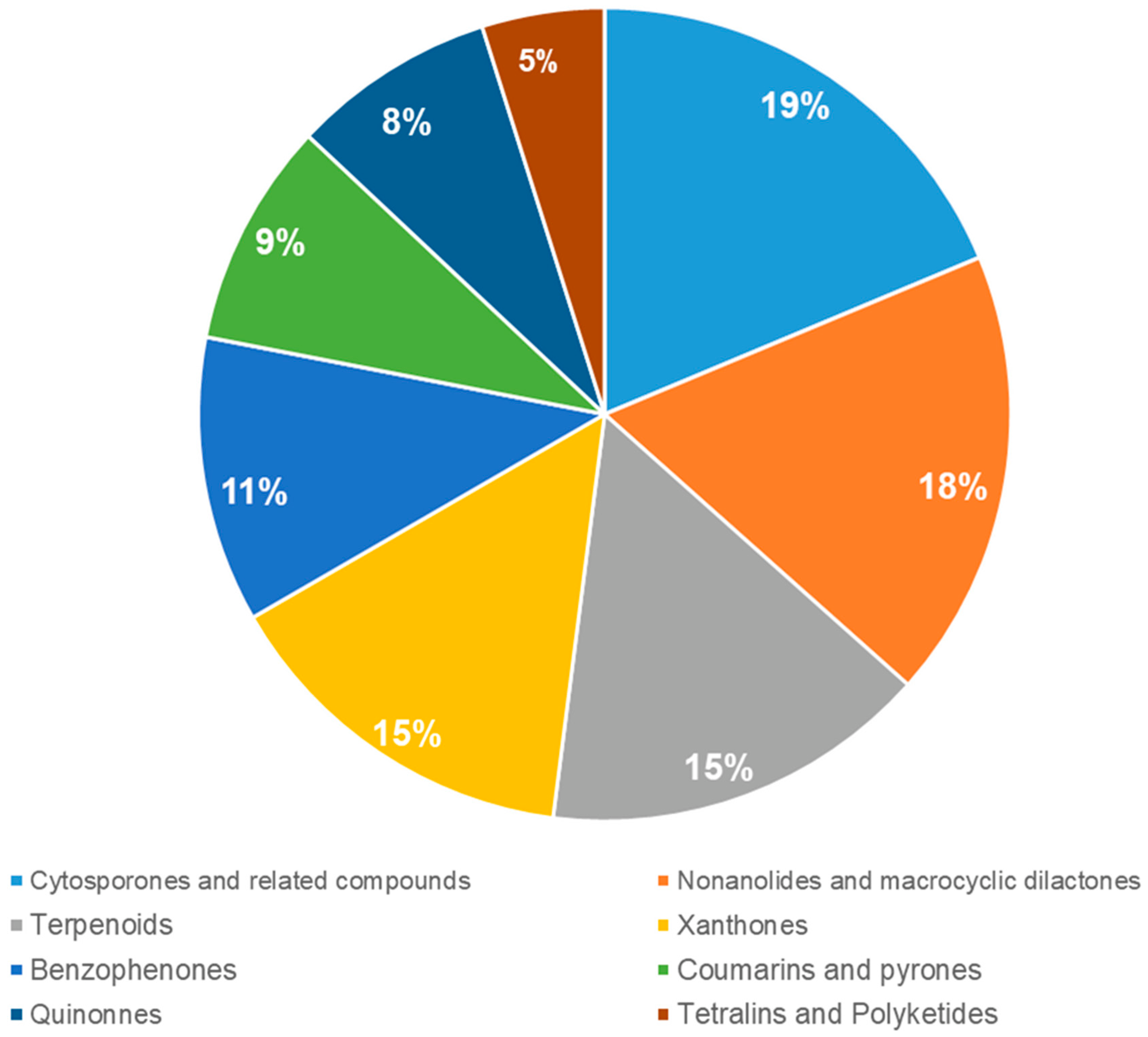
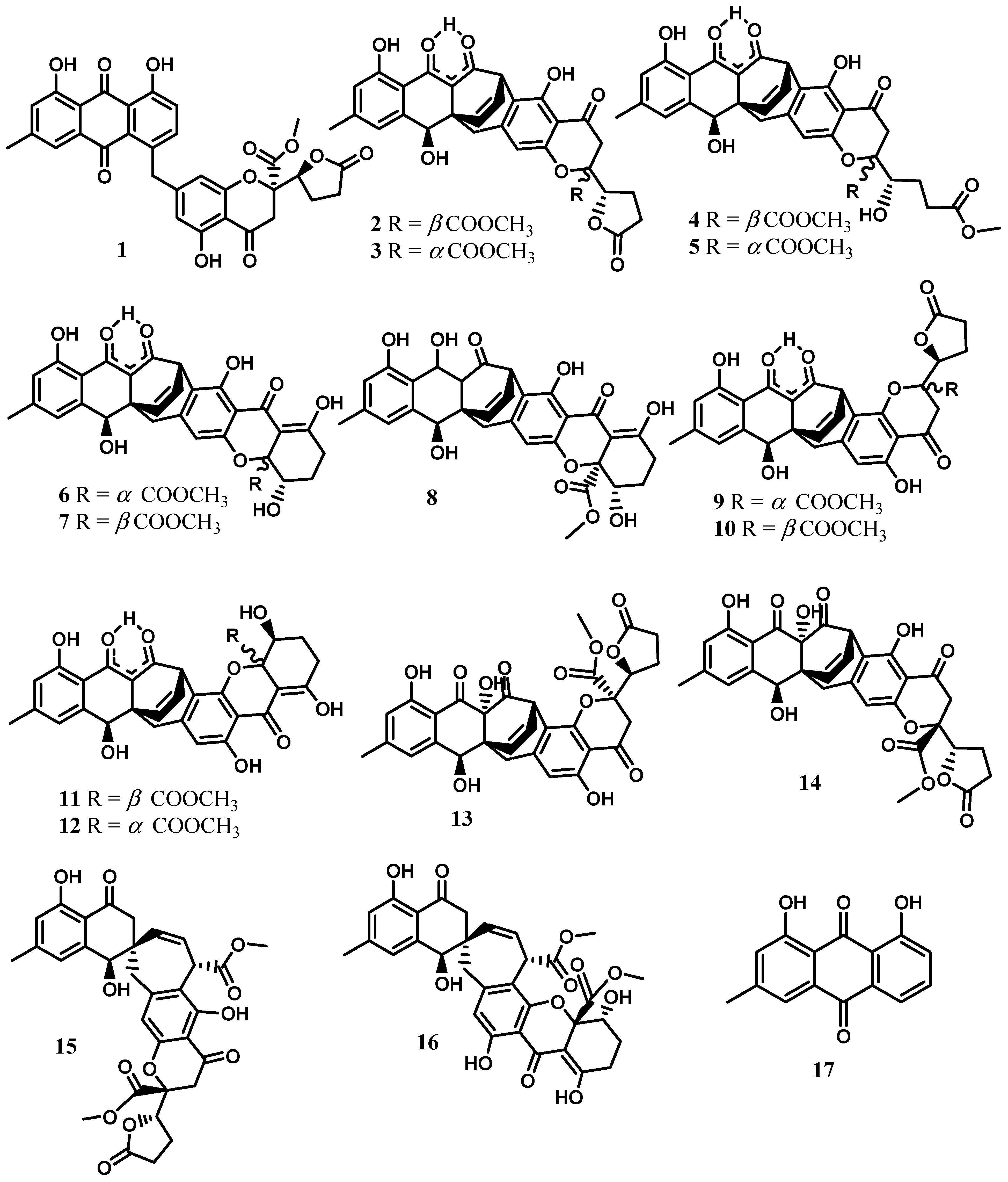
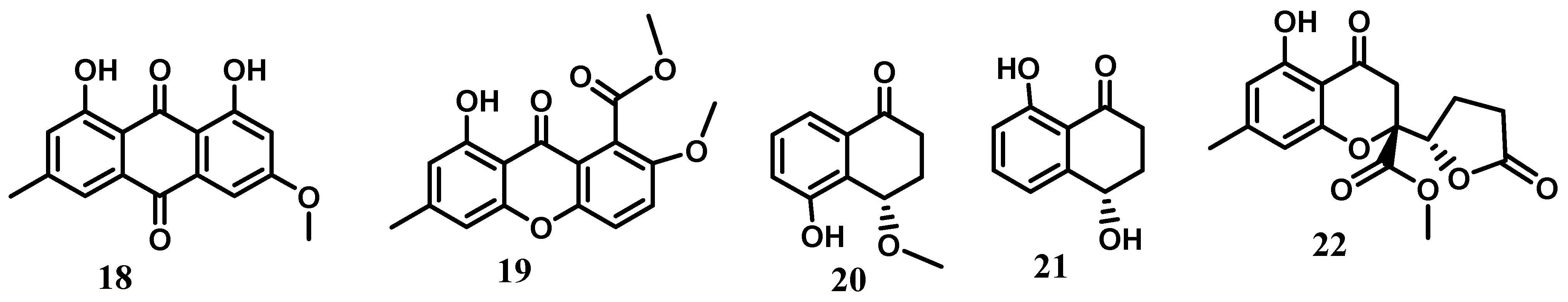
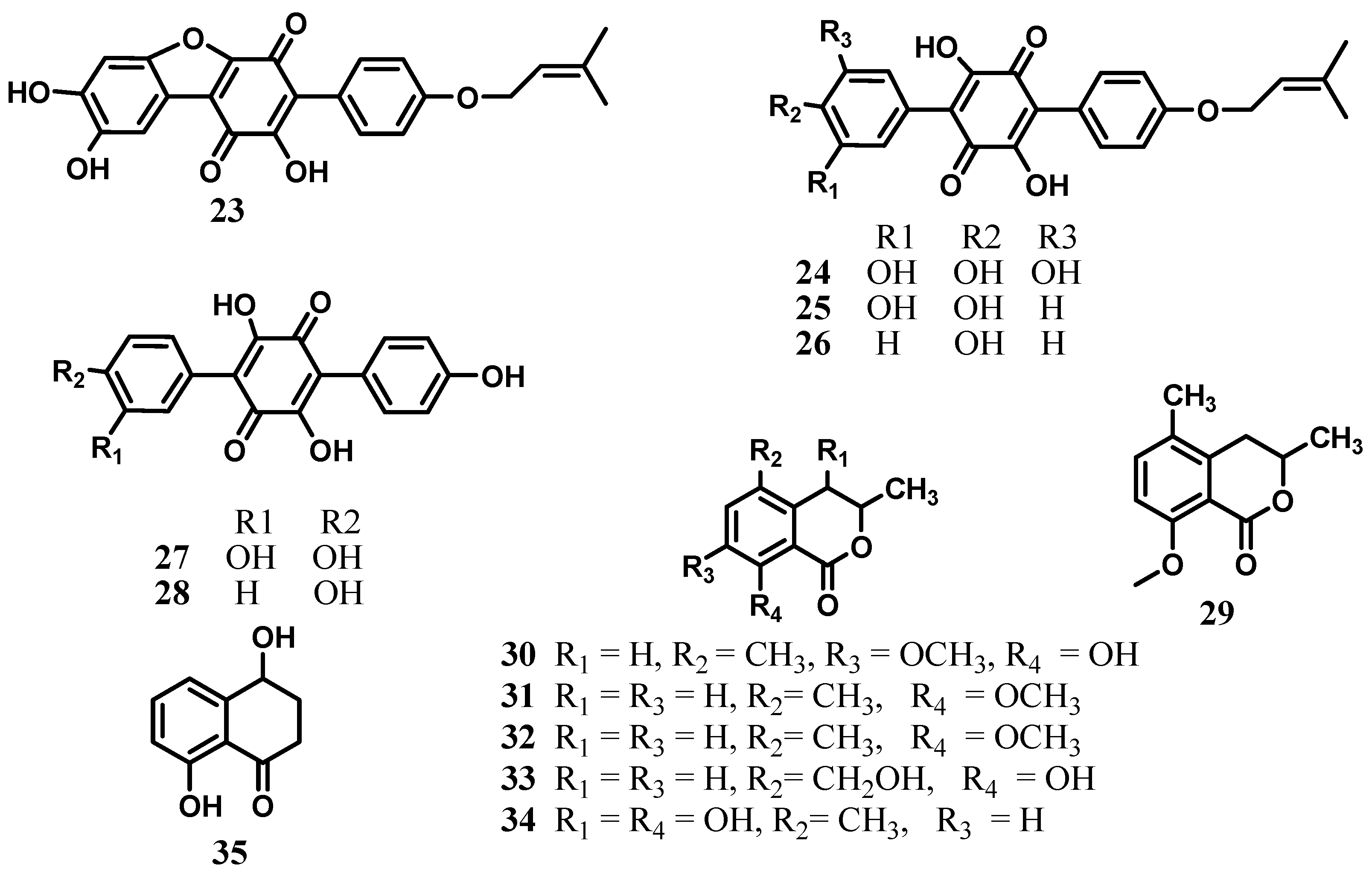
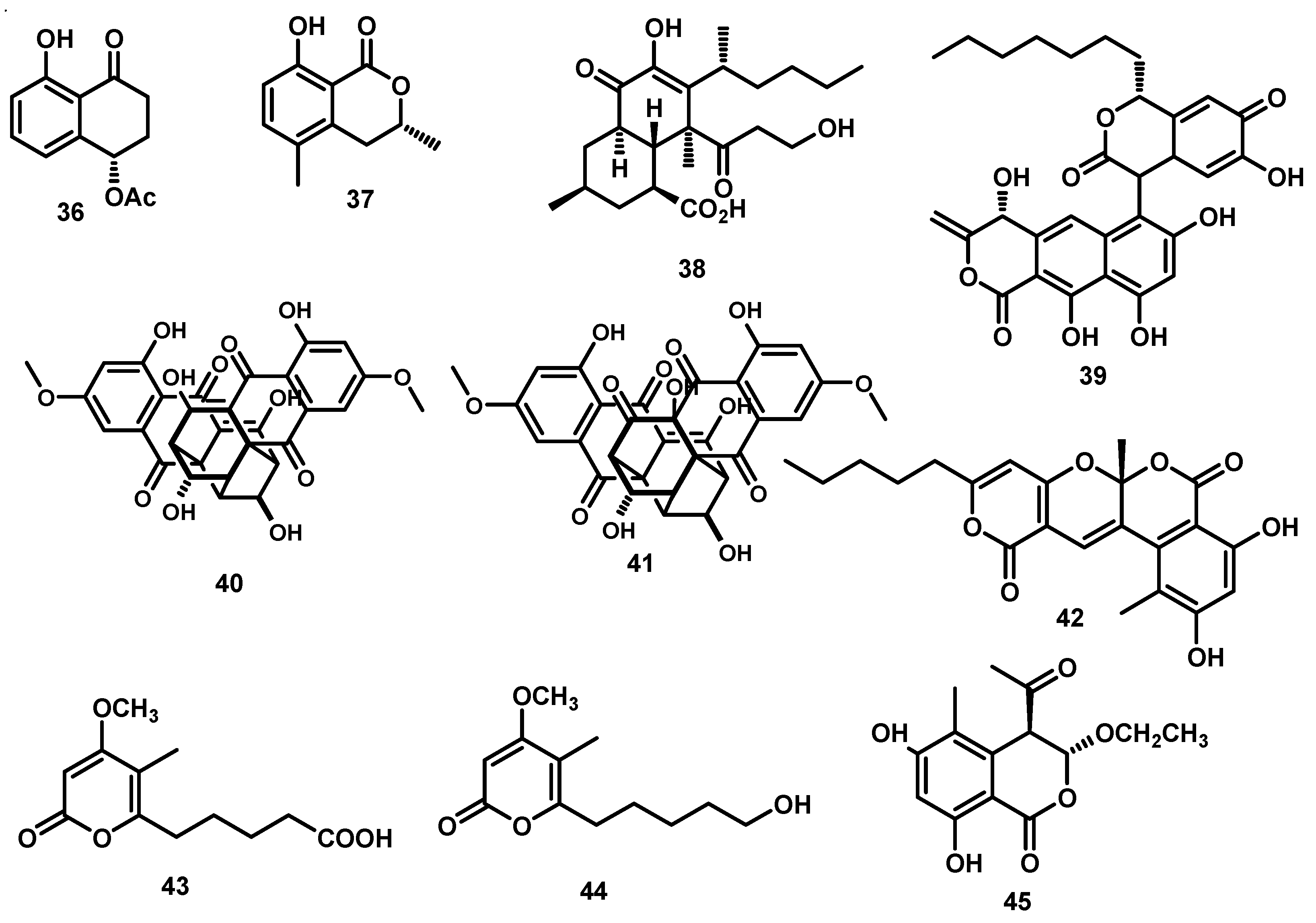
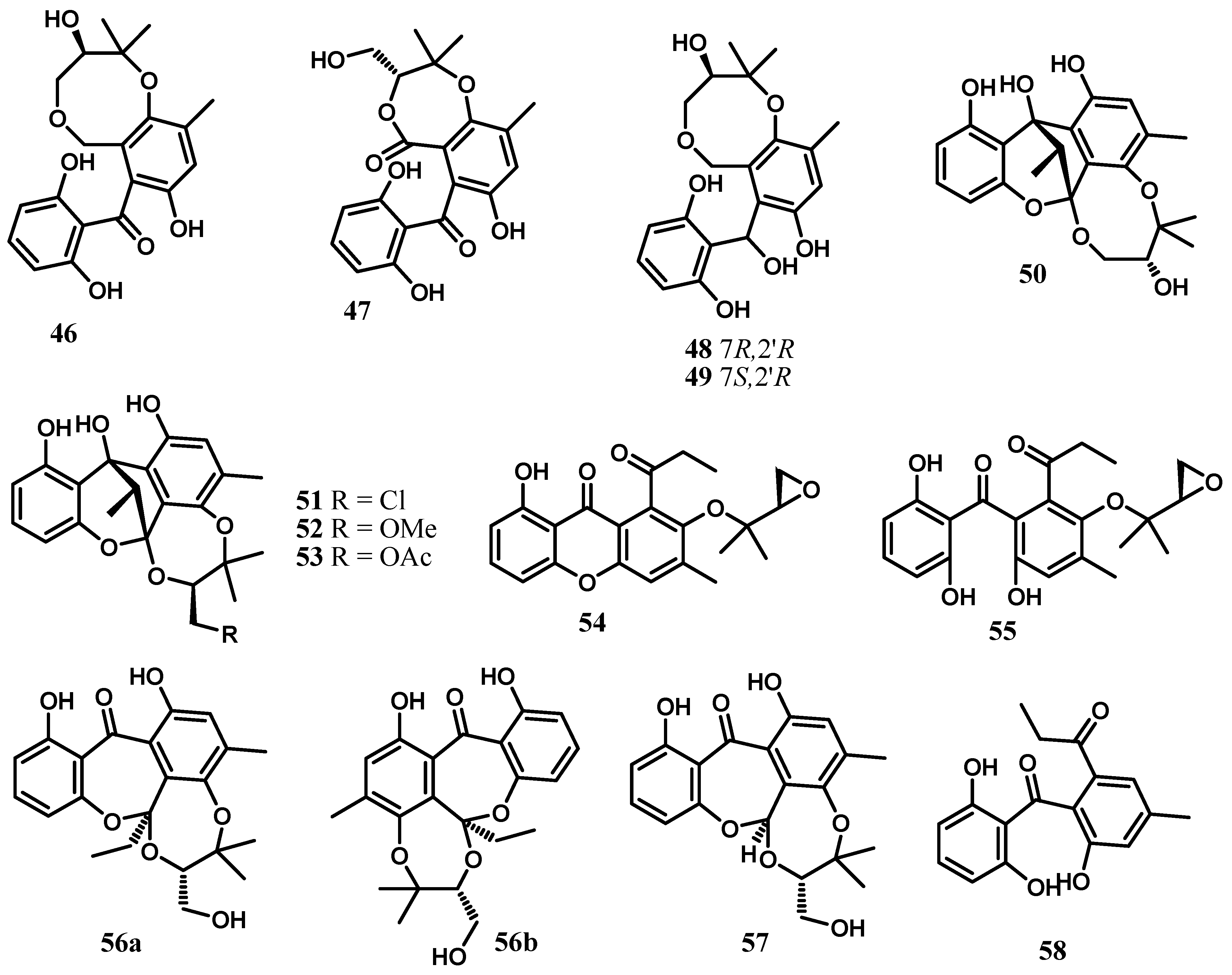
2. Xanthones, Quinones, and Coumarins
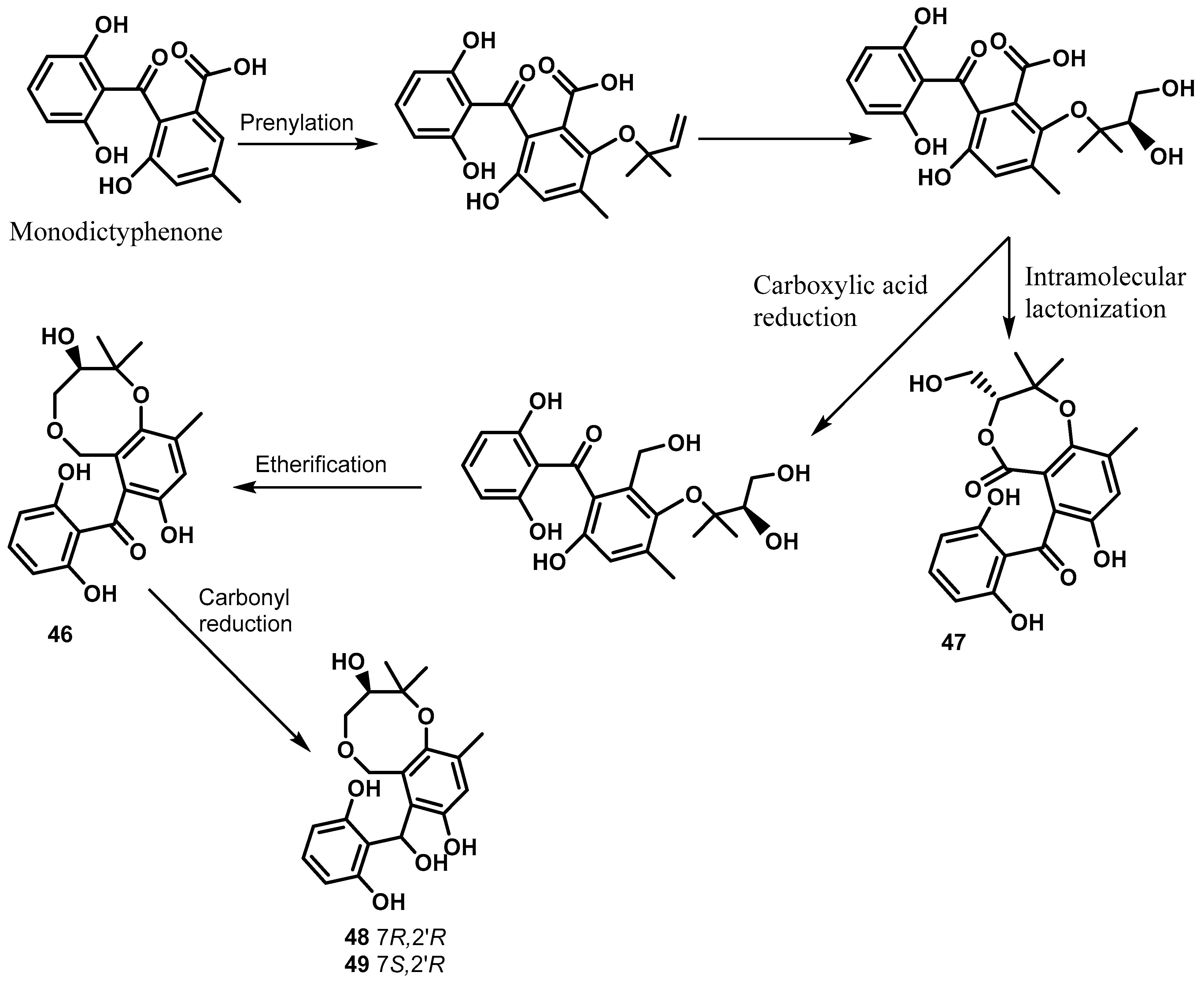
3. Benzophenones
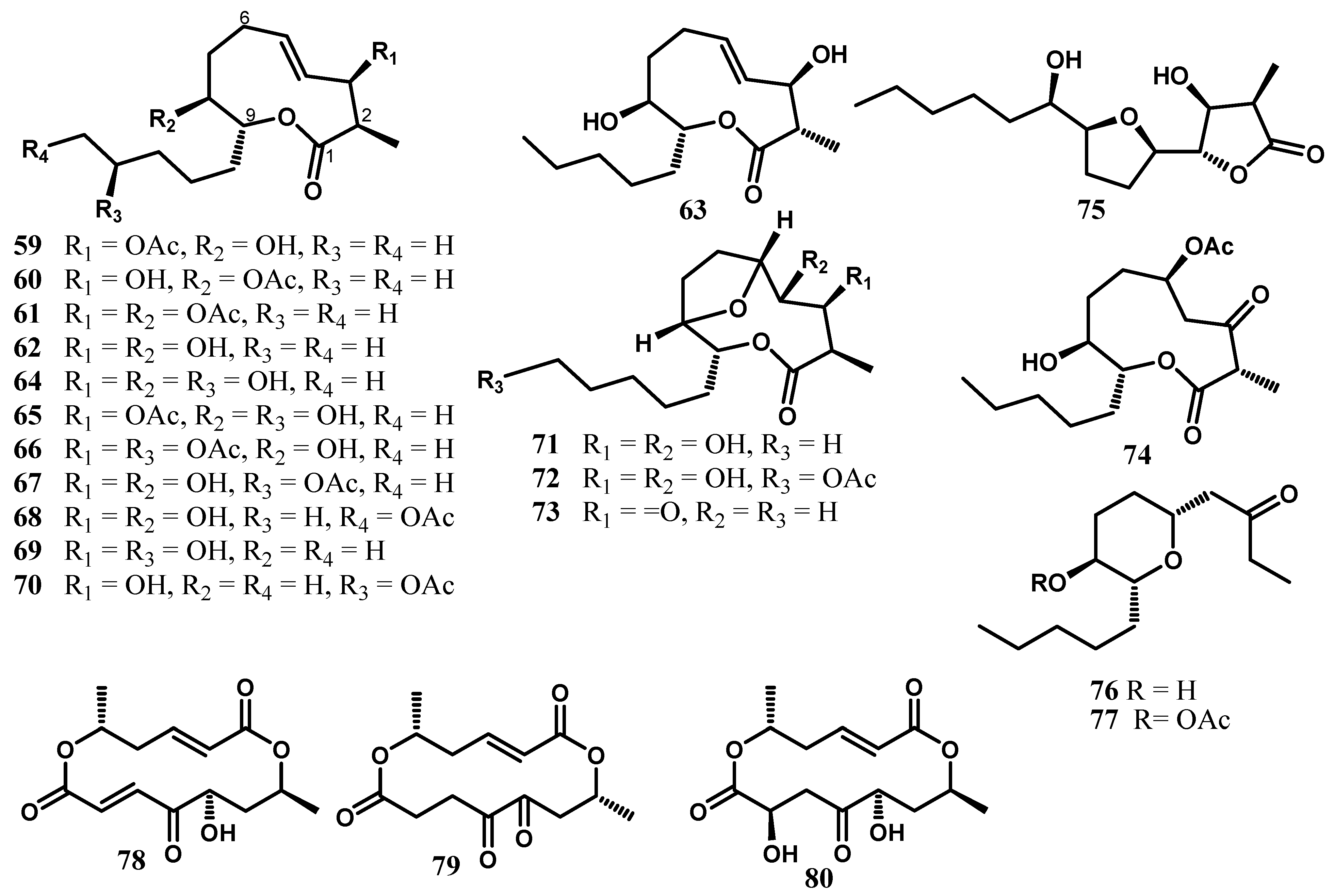
4. Nonanolides and Macrocyclic Dilactones
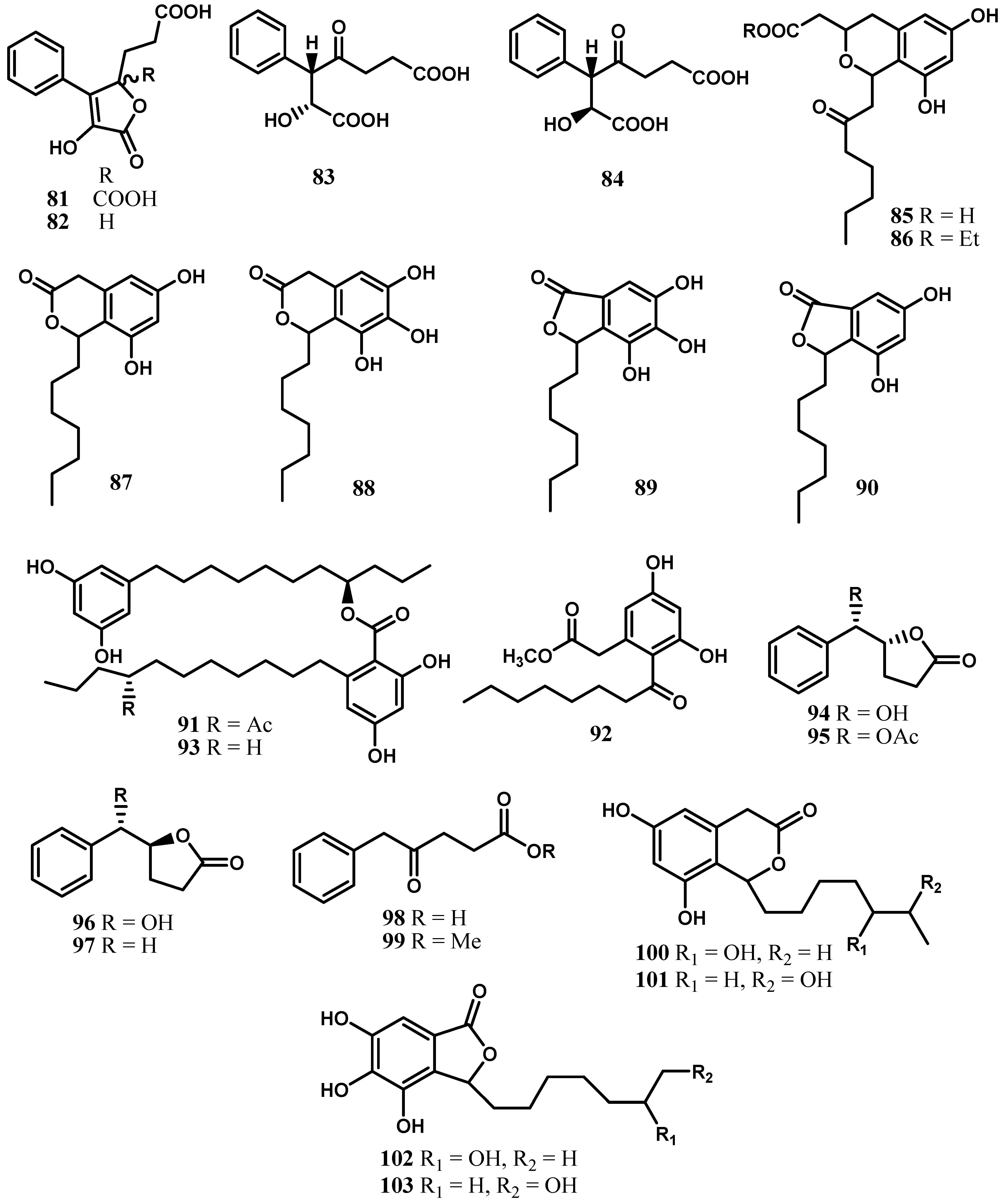
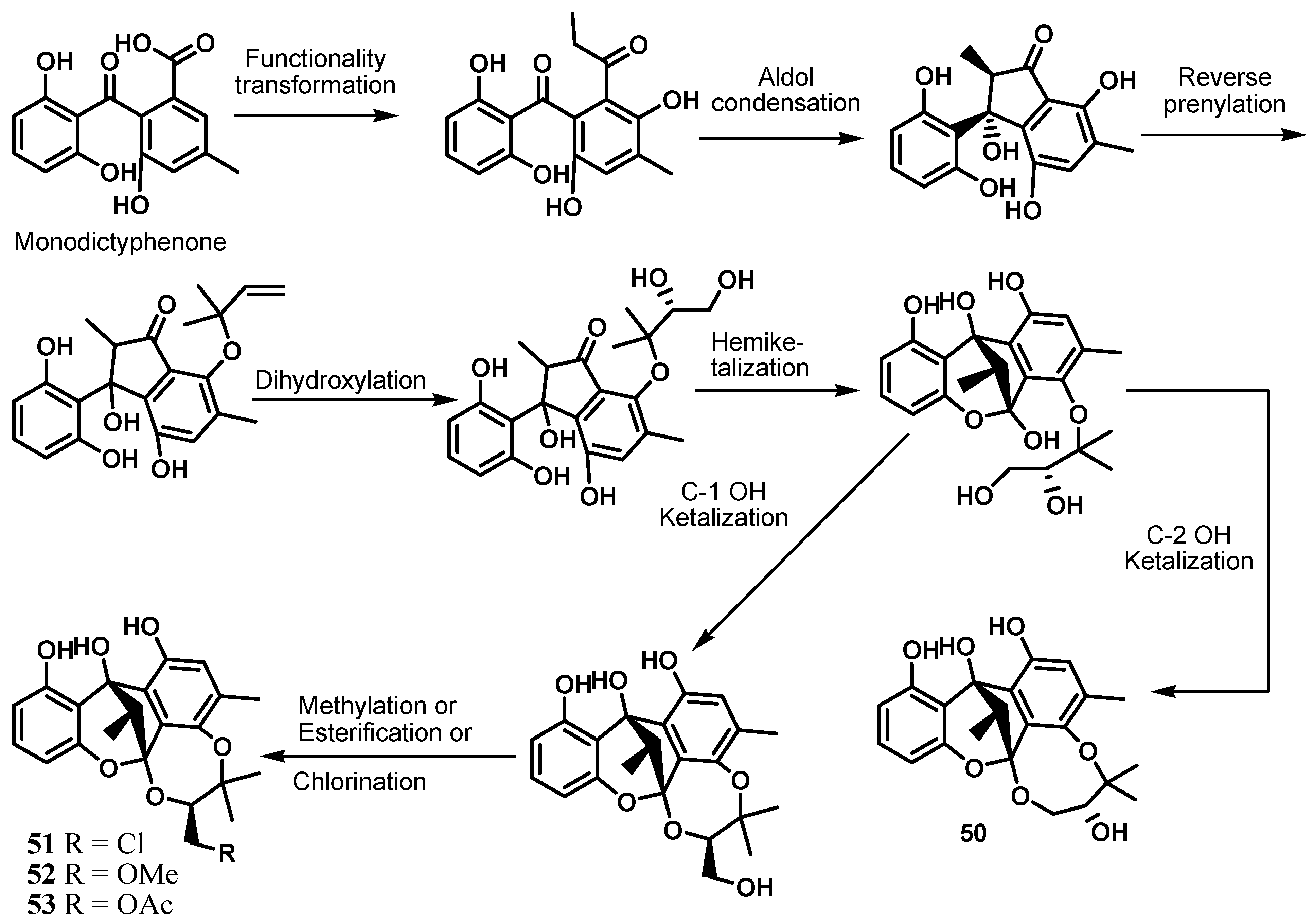
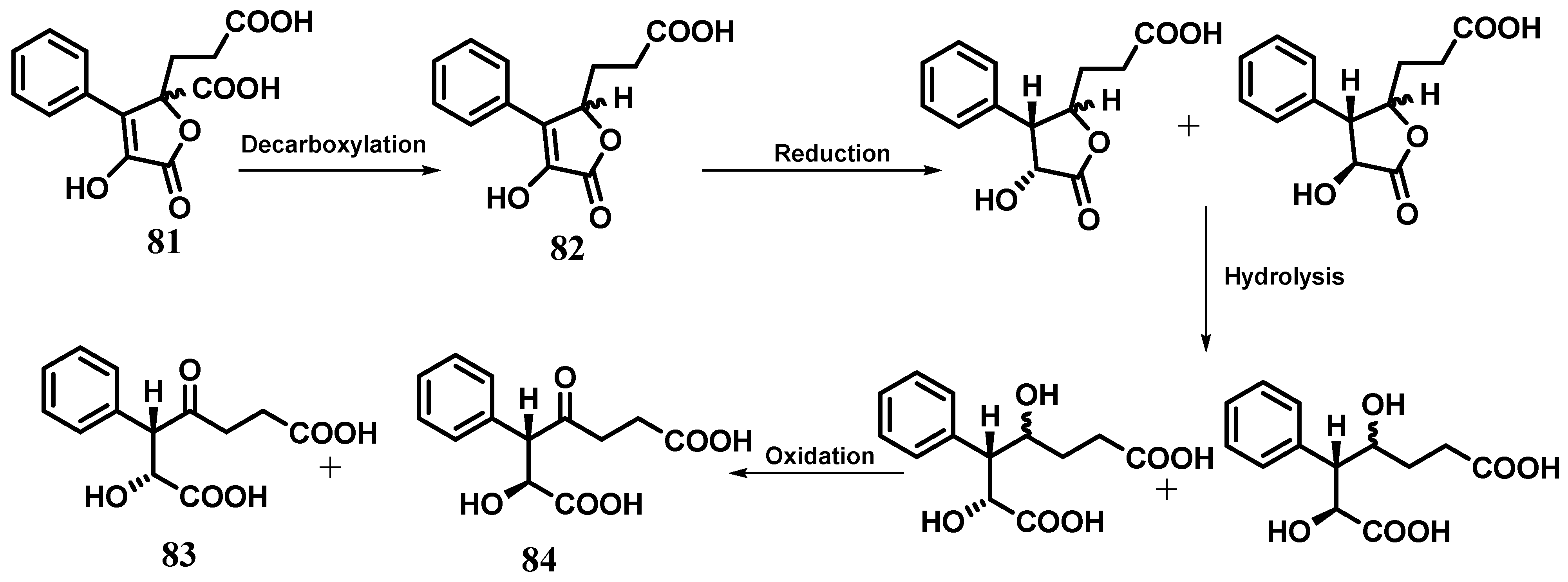
5. Cytosporones and Related Compounds
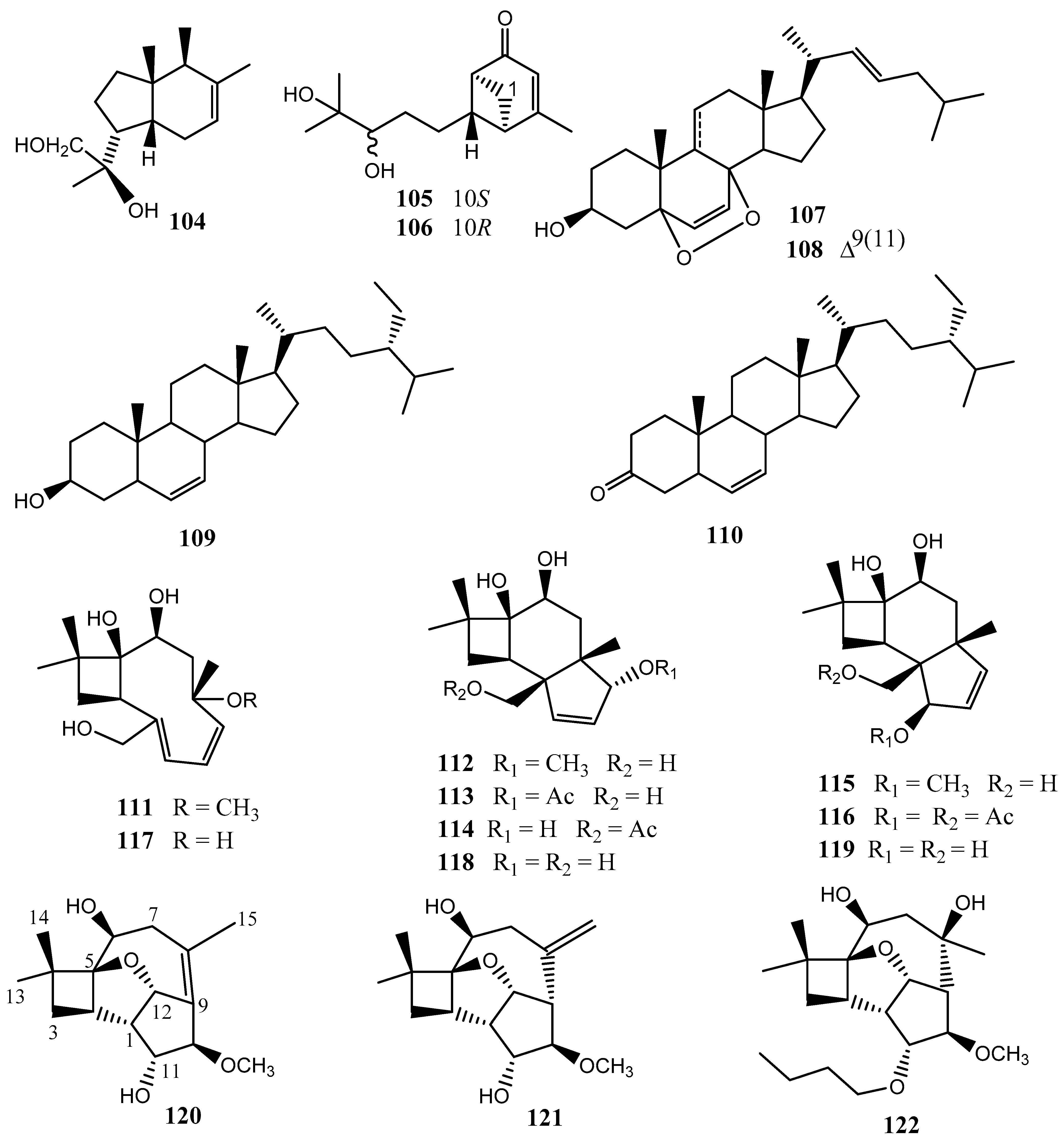
6. Terpenoids
7. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef] [PubMed]
- Knowles, S.L.; Raja, H.A.; Roberts, C.D.; Oberlies, N.H. Fungal–fungal co-culture: A primer for generating chemical diversity. Nat. Prod. Rep. 2022, 39, 1557–1573. [Google Scholar] [CrossRef] [PubMed]
- Jayasuriya, H.; Guan, Z.; Polishook, J.D.; Dombrowski, A.W.; Felock, P.J.; Hazuda, D.J.; Singh, S.B. Isolation, structure, and HIV-1 integrase inhibitory activity of cytosporic acid, a fungal metabolite produced by a Cytospora sp. J. Nat. Prod. 2003, 66, 551–553. [Google Scholar] [CrossRef]
- Sadorn, K.; Saepua, S.; Boonyuen, N.; Boonruangprapa, T.; Rachtawee, P.; Pittayakhajonwut, P. Antimicrobial activity and cytotoxicity of xanthoquinodin analogs from the fungus Cytospora eugeniae BCC42696. Phytochemistry 2018, 151, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, C.G. Sylvae Mycologicae Berolinenses; Formis Theophili Bruschcke: Berlin, Germany, 1818. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.N.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Chen, Y.; Liu, Z.; Tan, H.; Zhang, W. Pyrone and isocoumarin derivatives from the endophytic fungus Cytospora rhizophorae. Org. Biomol. Chem. 2022, 20, 4900–4904. [Google Scholar] [CrossRef] [PubMed]
- Index Fungorum Home Page. Available online: https://www.indexfungorum.org/ (accessed on 2 February 2023).
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Pers.-Mol. Phylogeny Evol. Fungi 2020, 45, 1–45. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Adams, G.C.; Cannon, P.F.; Castlebury, L.A.; Crous, P.W.; Gryzenhout, M.; Jaklitsch, W.M.; Mejia, L.C.; Stoykov, D.; Udayanga, D.; et al. Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 2015, 6, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Jeewon, R.; Chomnunti, P.; Wanasinghe, D.N.; Norphanphoun, C.; Karunarathna, A.; Pem, D.; Perera, R.H.; Camporesi, E.; McKenzie, E.H.C.; et al. Taxonomic circumscription of diaporthales based on multigene phylogeny and morphology. Fungal Divers. 2018, 93, 241–443. [Google Scholar] [CrossRef]
- Kalkanci, A.; Kustimur, S.; Turkoz Sucak, G.; Senol, E.; Sugita, T.; Adams, G.; Verkley, G.; Summerbell, R. Fulminating fungal sinusitis caused by Valsa sordida, a plant pathogen, in a patient immunocompromised by acute myeloid leukemia. Med. Mycol. 2006, 44, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Hanifeh, S.; Zafari, D.; Soleimani, M.-J.; Arzanlou, M. Multigene phylogeny, morphology, and pathogenicity trials reveal novel Cytospora species involved in perennial canker disease of apple trees in Iran. Fungal Biol. 2022, 126, 707–726. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Pan, M.; Tian, C.; Fan, X. Fungal richness of Cytospora species associated with willow canker disease in China. J. Fungi 2022, 8, 377. [Google Scholar] [CrossRef] [PubMed]
- Arzanlou, M.; Narmani, A. ITS Sequence data and morphology differentiate Cytospora chrysosperma associated with trunk disease of grapevine in Northern Iran. J. Plant Prot. Res. 2015, 55, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Chen, L.; Ge, Q.; Tian, C.; Huang, J. Spatiotemporal pattern and aggregation effects of poplar canker in Northeast China. Forests 2020, 11, 454. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Zhu, H.; Liang, L.; Tian, C.; Fan, X. Studies of canker and dieback of oak tree in China, with two Cytospora species described. Plant Pathol. 2021, 70, 2005–2015. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Lawrence, D.P.; Hand, F.P.; Trouillas, F.P. Olive twig and branch dieback in California caused by Cytospora oleicola and the newly described species Cytospora olivarum sp. Nov. Plant Dis. 2020, 104, 1908–1917. [Google Scholar] [CrossRef]
- Bills, G.F. Isolation and analysis of endophytic fungal communities from woody plants. In Endophytic Fungi in Grasses and Woody Plants: Systematics, Ecology, and Evolution; Redlin, S.C., Carris, L.M., Eds.; APS Press: College Park, MD, USA, 1996; pp. 31–65. [Google Scholar]
- González, V.; Tello, M.L. The endophytic mycota associated with Vitis vinifera in central Spain. Fungal Divers. 2011, 47, 29–42. [Google Scholar] [CrossRef]
- Singh, M.P.; Janso, J.E.; Brady, S.F. Cytoskyrins and cytosporones produced by Cytospora sp. CR200: Taxonomy, fermentation and biological activities. Mar. Drugs 2007, 5, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Narmani, A.; Teponno, R.B.; Arzanlou, M.; Surup, F.; Helaly, S.E.; Wittstein, K.; Praditya, D.F.; Babai-Ahari, A.; Steinmann, E.; Stadler, M. Cytotoxic, antimicrobial and antiviral secondary metabolites produced by the plant pathogenic fungus Cytospora sp. CCTU A309. Fitoterapia 2019, 134, 314–322. [Google Scholar] [CrossRef]
- Liu, H.-X.; Tan, H.-B.; Chen, K.; Zhao, L.-Y.; Chen, Y.-C.; Li, S.-N.; Li, H.-H.; Zhang, W.-M. Cytosporins A–D, novel benzophenone derivatives from the endophytic fungus Cytospora rhizophorae A761. Org. Biomol. Chem. 2019, 17, 2346–2350. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tan, H.; Chen, Y.; Guo, X.; Wang, W.; Guo, H.; Liu, Z.; Zhang, W. Cytorhizins A–D, four highly structure-combined benzophenones from the endophytic fungus Cytospora rhizophorae. Org. Lett. 2019, 21, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tan, H.; Chen, K.; Chen, Y.; Zhang, W.; Chen, S.; Liu, H.; Zhang, W. Rhizophols A and B, antioxidant and axially chiral benzophenones from the endophytic fungus Cytospora rhizophorae. Org. Biomol. Chem. 2019, 17, 10009–10012. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tan, H.; Wang, W.; Zhang, W.; Chen, Y.; Li, S.; Liu, Z.; Li, H.; Zhang, W. Cytorhizophins A and B, benzophenone-hemiterpene adducts from the endophytic fungus Cytospora rhizophorae. Org. Chem. Front. 2019, 6, 591–596. [Google Scholar] [CrossRef]
- Lu, S.; Kurtán, T.; Yang, G.; Sun, P.; Mándi, A.; Krohn, K.; Draeger, S.; Schulz, B.; Yi, Y.; Li, L.; et al. Cytospolides A–E, new nonanolides from an endophytic fungus, Cytospora sp. Eur. J. Org. Chem. 2011, 2011, 5452–5459. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, C.-W.; Cui, C.-B.; Liu, X.-Z.; Che, Y.-S. cytosporinols A-C, new caryophyllene sesquiterpenoids from Cytospora sp. Nat. Prod. Bioprospect. 2012, 2, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Li, G.; Sun, M.; Yang, X.; Xu, J. A new antimicrobial sesquiterpene isolated from endophytic fungus Cytospora Sp. from the Chinese mangrove plant Ceriops Tagal. Nat. Prod. Res. 2018, 34, 1404–1408. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; Collemare, J.; Mehrabi, R.; De Wit, P.J.G.M. Phytotoxic secondary metabolites and peptides produced by plant pathogenic dothideomycete fungi. FEMS Microbiol. Rev. 2013, 37, 67–93. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-D.; Chen, Y.; Gao, H.; Shen, L.-Q.; Wu, Y.; Li, X.-X.; Li, Y.; Guo, L.-D.; Cen, Y.-Z.; Yao, X.-S. Xanthoquinodins from the endolichenic fungal strain Chaetomium elatum. J. Nat. Prod. 2013, 76, 702–709. [Google Scholar] [CrossRef]
- Zhao, Y.-L.; Wang, J.-B.; Zhou, G.-D.; Shan, L.-M.; Xiao, X.-H. Investigations of free anthraquinones from Rhubarb against α-naphthylisothiocyanate-induced cholestatic liver injury in rats. Basic Clin. Pharmacol. Toxicol. 2009, 104, 463–469. [Google Scholar] [CrossRef]
- Singh, P.; Nee Pant, G.J.; Rawat, M. Phytochemistry and biological activity perspectives of Rheum species. Nat. Prod. J. 2016, 6, 84–93. [Google Scholar] [CrossRef]
- Hillis, W.E. The isolation of chrysophanic acid and physcion from Rumex hymenosepalus. Torr. Aust. J. Chem. 1955, 8, 290–292. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Feng, B.; Zhu, R.; Ma, J.; Wang, W. Preparative isolation of three anthraquinones from Rumex japonicus by high-speed counter-current chromatography. Molecules 2011, 16, 1201–1210. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, X.; Liu, X.; Wang, H.; Xue, J.; Yu, J.; Kang, N.; Wang, X. Chrysophanol inhibits NALP3 inflammasome activation and ameliorates cerebral ischemia/reperfusion in mice. Mediat. Inflamm. 2014, 2014, e370530. [Google Scholar] [CrossRef] [Green Version]
- Tamokou, J.D.D.; Tala, M.F.; Wabo, H.K.; Kuiate, J.R.; Tane, P. Antimicrobial activities of methanol extract and compounds from stem bark of Vismia rubescens. J. Ethnopharmacol. 2009, 124, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Das Sarma, M.; Patra, A.; Hazra, B. Anti-inflammatory and anticancer compounds isolated from Ventilago madraspatana Gaertn, Rubia cordifolia Linn, and Lantana camara Linn. J. Pharm. Pharmacol. 2010, 62, 1158–1166. [Google Scholar] [CrossRef]
- Kim, S.-J.; Kim, M.-C.; Lee, B.-J.; Park, D.-H.; Hong, S.-H.; Um, J.-Y. Anti-inflammatory activity of chrysophanol through the suppression of NF- kB/caspase-1 activation in vitro and in vivo. Molecules 2010, 15, 6436–6451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellado, M.; Madrid, A.; Peña-Cortés, H.; López, R.; Jara, C.; Espinoza, L. Antioxidant activity of anthraquinones isolated from leaves of Muehlenbeckia hastulata (J.E. Sm.) Johnst. (Polygonaceae). J. Chil. Chem. Soc. 2013, 58, 1767–1770. [Google Scholar] [CrossRef] [Green Version]
- Machida, K.; Matsuoka, E.; Kasahara, T.; Kikuchi, M. Studies on the constituents of Juglans species. Structural determination of (4S)- and (4R)-4-hydroxy-α-tetralone derivatives from the fruit of Juglans mandshurica Maxim. var. sieboldiana Makino. Chem. Pharm. Bull. 2005, 53, 934–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Li, G.; Deng, Q.; Zheng, D.; Yang, X.; Xu, J. Cytotoxic constituents from the mangrove endophytic Pestalotiopsis sp. induce G0/G1 cell cycle arrest and apoptosis in human cancer cells. Nat. Prod. Res. 2017, 32, 2968–2972. [Google Scholar] [CrossRef]
- Morita, T.; Aoki, H. Isosclerone, a new metabolite of Sclerotinia sclerotiorum (Lib.) De Bary. Agric. Biol. Chem. 1974, 38, 1501–1505. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Andolfi, A.; Bruno, G. Two naphthalenone pentaketides from liquid cultures of Phaeoacremonium aleophilum, a fungus associated with esca of grapevine. Phytopathol. Mediterr. 2000, 39, 162–168. [Google Scholar] [CrossRef]
- Zheng, C.-J.; Sohn, M.-J.; Kim, W.-G. Atromentin and leucomelone, the first inhibitors specific to enoyl-ACP reductase (FabK) of Streptococcus pneumoniae. J. Antibiot. 2006, 59, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Kokubun, T.; Veitch, N.C.; Bridge, P.D.; Simmonds, M.S.J. Dihydroisocoumarins and a tetralone from Cytospora eucalypticola. Phytochemistry 2003, 62, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Draeger, S.; Schulz, B.; Krohn, K.; Ahmed, I.; Hussain, H.; Yi, Y.; Li, L.; Zhang, W. Bioactive aromatic derivatives from endophytic fungus, Cytospora sp. Nat. Prod. Comm. 2011, 6, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Ballio, A.; Barcellona, S.; Santurbano, B. 5-Methylmellein, a new natural dihydroisocoumarin. Tetrahedron Lett. 1966, 31, 3723–3726. [Google Scholar] [CrossRef]
- Sumarah, M.W.; Puniani, E.; Blackwell, B.A.; Miller, J.D. Characterization of polyketide metabolites from foliar endophytes of Picea glauca. J. Nat. Prod. 2008, 71, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Janso, J.E.; Williamson, R.T.; Yang, H.Y.; Carter, G.T. Cytosporacin, a highly unsaturated polyketide: Application of the ACCORD-ADEQUATE experiment to the structural determination of natural products. J. Org. Chem. 2003, 68, 6079–6082. [Google Scholar] [CrossRef]
- Agusta, A.; Ohashi, K.; Shibuya, H. Bisanthraquinone metabolites produced by the endophytic fungus Diaporthe sp. Chem. Pharm. Bull. 2006, 54, 579–582. [Google Scholar] [CrossRef] [Green Version]
- Bouhet, J.C.; Chuong, P.P.V.; Toma, F.; Kirszenbaum, M.; Fromageot, P. Isolation and characterization of luteoskyrin and rugulosin, two hepatotoxic anthraquinonoids from Penicillium islandicum Sopp. and Penicillium rugulosum Thom. J. Agric. Food Chem. 1976, 24, 964–972. [Google Scholar] [CrossRef]
- Lu, S.; Sun, P.; Li, T.; Kurtan, T.; Mandi, A.; Antus, S.; Krohn, S.; Krohn, K.; Draeger, S.; Schulz, B.; et al. Bioactive nonanolide derivatives isolated from the endophytic fungus Cytospora sp. J. Org. Chem. 2011, 76, 9699–9710. [Google Scholar] [CrossRef]
- Gurusiddaiah, S.; Ronald, R.C. Grahamimycins: Antibiotics from Cytospora sp. Ehrenb.W.F.P.L. 13A. Antimicrob. Agents Chemother. 1981, 19, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höller, U.; König, G.M.; Wright, A.D. A new tyrosine kinase inhibitor from a marine isolate of Ulocladium botrytis and new metabolites from the marine fungi Asteromyces cruciatus and Varicosporina ramulosa. Eur. J. Org. Chem. 1999, 11, 2949–2955. [Google Scholar] [CrossRef]
- Küppers, L.; Ebrahim, W.; El-Neketi, M.; Özkaya, F.C.; Mándi, A.; Kurtán, T.; Orfali, R.S.; Müller, W.E.G.; Hartmann, R.; Lin, W.; et al. Lactones from the sponge-derived fungus Talaromyces rugulosus. Mar. Drugs 2017, 15, 359. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Koseki, T.; Koyama, H.; Shiono, Y. A new cytosporone derivative from the endophytic fungus Cytospora sp. Nat. Prod. Comm. 2014, 9, 973–975. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.B.; Zink, D.L.; Bills, G.F.; Pelaez, F.; Teran, A.; Collado, J.; Silverman, K.C.; Lingham, R.B.; Felock, P.; Hazuda, D.J. Discovery, structure and HIV-1 integrase inhibitory activities of integracins, novel dimeric alkyl aromatics from Cytonaema sp. Tetrahedron Lett. 2002, 43, 1617–1620. [Google Scholar] [CrossRef]
- Abreu, L.M.; Phipps, R.K.; Pfennig, L.H.; Gotfredsen, C.H.; Takahashi, J.A.; Larsen, T.O. Cytosporones O, P and Q from an endophytic Cytospora sp. Tetrahedron Lett. 2010, 51, 1803–1805. [Google Scholar] [CrossRef]
- Bishayee, A.; Ahmed, S.; Brankov, N.; Perloff, M. Triterpenoids as potential agents for the chemoprevention and therapy of breast cancer. Front. Biosci. 2011, 16, 980–996. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Xie, C.-L.; Zhong, T.; Xu, W.; Luo, Z.-H.; Shao, Z.; Yang, X.-W. Sesquiterpenes from a deep-sea-derived fungus Graphostroma sp. MCCC 3A00421. Tetrahedron 2017, 73, 7267–7273. [Google Scholar] [CrossRef]
- Ioannou, E.; Abdel-Razik, A.F.; Zervou, M.; Christofidis, D.; Alexi, X.; Vagias, C.; Alexis, M.N.; Roussis, V. 5α,8α-epidioxysterols from the gorgonian Eunicella cavolini and the ascidian Trididemnum inarmatum: Isolation and evaluation of their antiproliferative activity. Steroids 2009, 74, 73–80. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Q.; Liu, X.; Che, Y. Punctaporonins N–S, new caryophyllene sesquiterpenoids from Cytospora sp. BioMed. Res. Inter. 2017, 2017, 7871459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayyangar, N.R.; Bapat, D.S.; Joshi, B.S. A new synthesis of chrysophanol, rhein, islandicin, emodin and physcion. J. Sci. Ind. Res. 1961, 20B, 493–497. [Google Scholar]
- Prado, S.; Buisson, D.; Ndoye, I.; Vallet, M.; Nay, B. One-step enantioselective synthesis of (4S)-isosclerone through biotranformation of juglone by an endophytic fungus. Tetrahedron Lett. 2012, 54, 1189–1191. [Google Scholar] [CrossRef]
- Sudhakar, G.; Bayya, S.; Kadam, V.D.; Nanubolu, J.B. Total synthesis of gonytolides C and G, lachnone C, and formal synthesis of blennolide C and diversonol. Org. Biomol. Chem. 2014, 12, 5601–5610. [Google Scholar] [CrossRef] [PubMed]
- Gan, X.; Jiang, W.; Wang, W.; Hu, L. An Approach to 3,6-disubstituted 2,5-dioxybenzoquinones via two sequential Suzuki couplings. Three-step synthesis of leucomelone. Org. Lett. 2009, 11, 589–592. [Google Scholar] [CrossRef]
- Ye, Y.Q.; Koshino, H.; Abe, N.; Nakamura, T.; Hashizume, D.; Takahashi, S. Synthesis of atromentin and its O-alkylated natural products. Biosci. Biotechnol. Biochem. 2010, 74, 2342–2344. [Google Scholar] [CrossRef] [Green Version]
- Nicolaou, K.C.; Lim, Y.H.; Piper, J.L.; Papageorgiou, C.D. Total syntheses of 2,2′-epi-cytoskyrin A, rugulosin, and the alleged structure of rugulin. J. Am. Chem. Soc. 2007, 129, 4001–4013. [Google Scholar] [CrossRef]
- Kamal, A.; Balakrishna, M.; Reddy, P.V.; Rahim, A. First total synthesis of the E- and Z-isomers of cytospolide-D. Tetrahedron Asymmetry 2014, 25, 148–155. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Matsuumi, M. First total synthesis of grahamimycin A. J. Org. Chem. 2000, 65, 7221–7224. [Google Scholar] [CrossRef] [PubMed]
- Hillis, L.R.; Ronald, R.C. Total synthesis of (−)-grahamimycin A1. J. Org. Chem. 1985, 50, 470–473. [Google Scholar] [CrossRef]
- He, H.; Lei, Z.; Xiaodong, Z.; Xun, J.; Xiao, D.; Xu, S.; Hualiang, J.; Hong, L. Total synthesis of cytosporone B. Chin. J. Chem. 2010, 28, 1041–1043. [Google Scholar] [CrossRef]
- Hall, J.D.; Duncan-Gould, N.W.; Siddiqi, N.A.; Kelly, J.N.; Hoeferlin, L.A.; Morrison, S.J.; Wyatt, J.K. Cytosporone E: Racemic synthesis and preliminary antibacterial testing. Bioorg. Med. Chem. 2005, 13, 1409–1413. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Reactions | References |
|---|---|---|
| Chrysophanol (17) Physcion (18) | Synthesis from 1-amino-5-chloroanthraquinone and 2-methylanthraquinone | [64] |
| Isosclerone (21) | Biotransformation of the commercially available 5-hydroxy-1,4-naphthalenedione using endophytic fungus Paraconiothyrium variabile | [65] |
| Gonytolide C (22) | Stereoselective total synthesis by aldol reaction between acetophenone and butyrolactone containing α-keto ester, followed by diastereoselective intramolecular cyclization | [66] |
| Leucomelone (27) | Synthesis from 2,5-dibromo-3,6-dimethoxy-1,4-benzoquinone | [67] |
| Atromentin (28) | Total synthesis based on double Suzuki–Miyaura coupling | [68] |
| Cytoskyrin A (40) | Total synthesis of 2,2′-epi-cytoskyrin A | [69] |
| Cytospolides D (62) | Total synthesis from L-malic acid and D-mannitol | [70] |
| Grahamimycin A (78) | Total synthesis via Yamaguchi macrocyclization | [71] |
| Grahamimycin A1 (79) | Total synthesis from 5(R)-hydroxy-2(E)-hexenoate and 7(R)-octynoic acid | [72] |
| Cytosporone B (86) | Total synthesis from 1-bromo-3,5-dimethoxybenzene | [73] |
| Cytosporone E (89) | Synthesis via the Meyers ortho-alkylation of a chiral aromatic oxazoline | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kianfé, B.Y.; Tchamgoue, J.; Narmani, A.; Teponno, R.B.; Njouonkou, A.-L.; Stadler, M.; Fogue Kouam, S. Bioactive Secondary Metabolites from Fungi of the Genus Cytospora Ehrenb. (Ascomycota). Molecules 2023, 28, 3120. https://doi.org/10.3390/molecules28073120
Kianfé BY, Tchamgoue J, Narmani A, Teponno RB, Njouonkou A-L, Stadler M, Fogue Kouam S. Bioactive Secondary Metabolites from Fungi of the Genus Cytospora Ehrenb. (Ascomycota). Molecules. 2023; 28(7):3120. https://doi.org/10.3390/molecules28073120
Chicago/Turabian StyleKianfé, Boris Yotta, Joseph Tchamgoue, Abolfazl Narmani, Rémy Bertrand Teponno, André-Ledoux Njouonkou, Marc Stadler, and Simeon Fogue Kouam. 2023. "Bioactive Secondary Metabolites from Fungi of the Genus Cytospora Ehrenb. (Ascomycota)" Molecules 28, no. 7: 3120. https://doi.org/10.3390/molecules28073120