
A Systematical Survey on the TRP Channels Provides New Insight into Its Functional Diversity in Zhikong Scallop (Chlamys farreri)

Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of TRP Genes in C. farreri
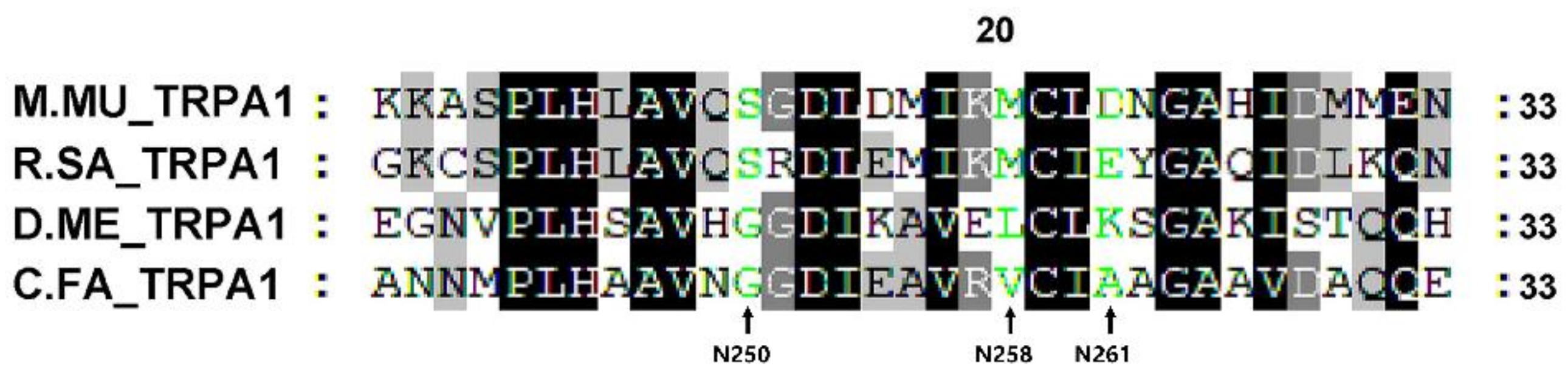
2.2. Phylogenetic Analysis and Interspecies Comparison of TRP Proteins
2.3. Spatiotemporal Expressions of C.FA TRP Genes
2.4. Expression of C.FA TRP Genes in Response to Heat Stress
3. Discussion
4. Materials and Methods
4.1. Genome-Wide Identification and Sequence Analysis of TRP Genes in C. farreri
4.2. Multiple Alignment and Phylogenetic Analysis
4.3. Spatiotemporal Expression Profiles of C.FA TRP Genes
4.4. Expression of C.FA TRP Genes Under Heat Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Minke, B.; Cook, B. TRP Channel Proteins and Signal Transduction. Physiol. Rev. 2002, 82, 429–472. [Google Scholar] [CrossRef] [Green Version]
- Mahieu, F.; Nilius, B. A road map for TR(I)Ps. Mol. Cell. 2006, 22, 297–307. [Google Scholar]
- Julius, D. From Peppers to Peppermints: Natural Products as Probes of the Pain Pathway. Harvey Lect. 2007, 101, 89–115. [Google Scholar]
- Nishida, M. TRP Channels: Molecular Diversity and Physiological Function. Microcirculation 2010, 13, 535–550. [Google Scholar] [CrossRef]
- Bowen, S.T. The Genetics of Artemia Salina. Biol. Bull. 1962, 125, 431–440. [Google Scholar] [CrossRef]
- Minke, B.; Wu, C. Induction of photoreceptor voltage noise in the dark in Drosophila mutant. Nature 1975, 258, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Rubin, G.M. Molecular characterization of the Drosophila TRP locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef]
- Montell, C. DrosophilaTRP channels. Pflüg. Arch. Eur. J. Physiol. 2005, 451, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Okamura, Y.; Nishino, A.; Murata, Y.; Nakajo, K.; Iwasaki, H.; Ohtsuka, Y.; Tanaka-Kunishima, M.; Takahashi, N.; Hara, Y.; Yoshida, T. Comprehensive analysis of the ascidian genome reveals novel insights into the molecular evolution of ion channel genes. Physiol. Genom. 2005, 22, 269–282. [Google Scholar] [CrossRef]
- Shin, J.; Adams, D.; Paukert, M.; Siba, M.; Sidi, S.; Levin, M.; Gillespie, P.; Grunder, S. Xenopus TRPN1 (NOMPC) localizes to microtubule-based cilia in epithelial cells, including inner-ear hair cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12572–12577. [Google Scholar] [CrossRef] [Green Version]
- Venkatachalam, K.; Montell, C. TRP Channels. Ann. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, X.; Xu, X.Z.S. Function and regulation of TRP family channels in C. elegans. Pflüg. Arch. Eur. J. Physiol. 2009, 458, 851–860. [Google Scholar]
- Wheeler, G.L.; Brownlee, C. Ca2+ signalling in plants and green algae—Changing channels. Trends Plant Sci. 2008, 13, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Zhu, M.X. A Unified Nomenclature for the Superfamily of TRP Cation Channels. Mol. Cell. 2002, 9, 229–231. [Google Scholar] [CrossRef]
- Palmer, C.P.; Aydar, E.; Djamgoz, M.B.A. A microbial TRP-like polycystic-kidney-disease-related ion channel gene. Biochem. J. 2005, 387, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Zhang, C. Advances in the study of the transient receptor potential channel protein (TRP) family, which mediates sensory physiological functions. Mil. Med. 2007, 73–75. [Google Scholar]
- Li, H. TRP Channel Classification; Springer Netherlands: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Sedgwick, S.G.; Smerdon, S.J. The ankyrin repeat: A diversity of interactions on a common structural framework. Trends Biochem. Sci. 1999, 24, 311–316. [Google Scholar] [CrossRef]
- Liu, S.; Wang, K. Research progress on heterologous assembly of TRP channel protein. J. Biophys. 2009, 19–25. [Google Scholar]
- Perraud, A.L.; Fleig, A.; Dunn, C.A.; Bagley, L.A.; Scharenberg, A.M. ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nature 2001, 411, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Sarna, S.K. Physiology and pathophysiology of colonic motor activity. Dig. Dis. Sci. 1991, 36, 827–862. [Google Scholar] [CrossRef]
- Montell, C. Physiology, phylogeny, and functions of the TRP superfamily of cation channels. Sci. STKE 2001, 2001, re1. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T. TRP channels: A TR(I)P through a world of multifunctional cation channels. Pflüg. Arch. Eur. J. Physiol. 2005, 451, 1–10. [Google Scholar] [CrossRef]
- Miller, B.A. The Role of TRP Channels in Oxidative Stress-induced Cell Death. J. Membr. Biol. 2006, 209, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Dadon, D.; Minke, B. Cellular functions of Transient Receptor Potential channels. Int. J. Biochem. Cell Biol. 2010, 42, 1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voets, T.; Droogmans, G.; Wissenbach, U.; Janssens, A.; Flockerzi, V.; Nilius, B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature 2004, 430, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Martínez-López, P. TRPM8 in mouse sperm detects temperature changes and may influence the acrosome reaction. J. Cell. Physiol. 2011, 226, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, H.; Ugawa, S.; Ueda, T.; Morita, A.; Shimada, S. TRPM8 activation suppresses cellular viability in human melanoma. Am. J. Physiol. Cell. Physiol. 2008, 295, C296–C301. [Google Scholar] [CrossRef] [Green Version]
- Nadler, M.J.S.; Hermosura, M.C.; Inabe, K.; Perraud, A.L.; Fleig, A. LTRPC7 is a Mg.ATP-regulated divalent cation channel required for cell viability. Nature 2001, 411, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, K.P.; Weber, S.; Peters, M.; Niemann Nejsum, L.; Vitzthum, H.; Klingel, K.; Kratz, M.; Haddad, E.; Ristoff, E.; Dinour, D. Hypomagnesemia with secondary hypocalcemia is caused by mutations in TRPM6, a new member of the TRPM gene family. Nat. Genet. 2002, 31, 166–170. [Google Scholar] [CrossRef]
- Runnels, L.W.; Yue, L.; Clapham, D.E. The TRPM7 channel is inactivated by PIP2 hydrolysis. Nat. Cell Biol. 2002, 4, 329–336. [Google Scholar] [CrossRef] [PubMed]
- West, R.J.; Sun, A.Y.; Church, D.L.; Lambie, E.J. The C-elegans gon-2 gene encodes a putative TRP cation channel protein required for mitotic cell cycle progression. Gene 2001, 266, 103–110. [Google Scholar] [CrossRef]
- Laing, R.J.; Dhaka, A. ThermoTRPs and Pain. Neuroscientist 2016, 22, 171–187. [Google Scholar] [CrossRef] [Green Version]
- Almaraz, L.; Manenschijn, J.A.; Peña, E.D.L.; Viana, F. TRPM8. Handb. Exp. Pharmacol. 2014, 222, 547–579. [Google Scholar]
- Zimmermann, K.; Lennerz, J.K.; Hein, A.; Link, A.S.; Kaczmarek, J.S.; Delling, M.; Uysal, S.; Pfeifer, J.D.; Riccio, A.; Clapham, D.E. Transient receptor potential cation channel, subfamily C, member 5 (TRPC5) is a cold-transducer in the peripheral nervous system. Proc. Natl. Acad. Sci. USA 2011, 108, 18114–18119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zygmunt, P.M.; Högestätt, E.D. TRPA1. Handb. Exp. Pharmacol. 2014, 222, 583–630. [Google Scholar]
- Caterina, M.J.; Schumacher, M.A. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, Y.; Lee, J.; Bang, S.; Hyun, S.; Kang, J.; Hong, S.T.; Bae, E.; Kaang, B.K.; Kim, J. Pyrexia is a new thermal transient receptor potential channel endowing tolerance to high temperatures in Drosophila melanogaster. Nat. Genet. 2005, 37, 305–310. [Google Scholar] [CrossRef]
- Gregory, N.G.; Keene, A.C.; Peter, D.; Chang, E.C.; Wang, Q.P.; Alp, A.Y.; Mark, R.; Michael, C.; Woolf, C.J.; Garrity, P.A. TrpA1 Regulates Thermal Nociception in Drosophila. PLoS ONE 2011, 6, e24343. [Google Scholar]
- Wang, M.; Zheng, J.; Yu, H. Chlamys Farreri (Jones & Preston, 1904) karyotype. J. Ocean Univ. China 1990, 85–89. [Google Scholar]
- Navarro, J.M. The effects of salinity on the physiological ecology of Choromytilus chorus (Molina, 1782) (Bivalvia: Mytilidae). J. Exp. Marine Biol. Ecol. 1988, 122, 1–33. [Google Scholar] [CrossRef]
- Laing, I. Effect of salinity on growth and survival of king scallop spat (Pecten maximus). Aquaculture 2002, 205, 171–181. [Google Scholar] [CrossRef]
- Jeon, C.Y.; Hur, Y.B.; Cho, K.C. The Effect of Water Temperature and Salinity on Settlement of Pacific Oyster, Crassostrea gigas Pediveliger Larvae. Korean J. Malacol. 2012, 28, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.; Foo, S.A.; Ross, P.M.; Putnam, H.M. Limitations of cross- and multigenerational plasticity for marine invertebrates faced with global climate change. Glob. Chang. Biol. 2020, 26, 80–102. [Google Scholar] [CrossRef] [Green Version]
- Wang, R. Aquaculture of Marine Shellfish; Qingdao Ocean University Press: Qingdao, China, 1993. [Google Scholar]
- Venkatachalam, K.; Luo, J.; Montell, C. Evolutionarily Conserved, Multitasking TRP Channels: Lessons from Worms and Flies. Handb. Exp. Pharmacol. 2014, 223, 937–962. [Google Scholar] [PubMed] [Green Version]
- Shi, J.; Li, J. TRP ion channels. J. Neuroanat. 2004, 2004, 1–15. [Google Scholar]
- Samuel, S.; Friedrich, R.W.; Nicolson, T. NompC TRP Channel Required for Vertebrate Sensory Hair Cell Mechanotransduction. Science 2003, 301, 96–99. [Google Scholar]
- Zou, W.; Huang, G.; Kang, L. Function and mechanism of TRP channels in response to mechanical stimuli. J. Zhejiang Univ. 2012, 16, 104–110. [Google Scholar]
- Liu, C.; Zwiebel, L.J. Molecular Characterization of Larval Peripheral Thermosensory Responses of the Malaria Vector Mosquito Anopheles gambiae. PLoS ONE 2013, 8, e72595. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wetzel, S.; Plückthun, A.; Fernandez, J.M. Stepwise Unfolding of Ankyrin Repeats in a Single Protein Revealed by Atomic Force Microscopy. Biophys. J. 2006, 90, L30–L32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcgrath, J.; Somlo, S.; Makova, S.; Tian, X.; Brueckner, M. Two Populations of Node Monocilia Initiate Left-Right Asymmetry in the Mouse. Cell 2003, 114, 1–73. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.-Z.S.; Sternbergm, P.W. A C. elegans Sperm TRP Protein Required for Sperm-Egg Interactions during Fertilization. Cell 2003, 114, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Tominaga, M. Functional diversity and evolutionary dynamics of thermoTRP channels. Cell Calcium. 2015, 57, 214–221. [Google Scholar] [CrossRef]
- Vriens, J.; Voets, T. Sensing the heat with TRPM3. Pflüg. Arch. Eur. J. Physiol. 2018, 470, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Jiao, Z.; Li, Y.; Tian, J.; Ren, L.; Zhang, F.; Li, Q.; Liu, S. Transient Receptor Potential (TRP) Channels in the Pacific Oyster (Crassostrea gigas): Genome-Wide Identification and Expression Profiling after Heat Stress between C. gigas and C. angulata. Int. J. Mol. Sci. 2021, 22, 3222. [Google Scholar] [CrossRef]
- Laursen, W.J.; Schneider, E.R.; Merriman, D.K.; Bagriantsev, S.N.; Gracheva, E.O. Low-cost functional plasticity of TRPV1 supports heat tolerance in squirrels and camels. Proc. Natl. Acad. Sci. USA 2016, 113, 11342–11347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandl, J.; Hu, H.; Bandell, M.; Bursulaya, B.; Patapoutian, A. Pore region of TRPV3 ion channel is specifically required for heat activation. Nat. Neurosci. 2008, 11, 1007–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, H.; Jara-Oseguera, A.; Hernández-García, E.; Llorente, I.; Rosenbaum, T. Structural determinants of gating in the TRPV1 channel. Nat. Struct. Mol. Biol. 2009, 16, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Grandl, J.R.; Kim, S.E.; Uzzell, V.; Bursulaya, B.; Petrus, M.; Bandell, M.; Patapoutian, A. Temperature-induced opening of TRPV1 ion channel is stabilized by the pore domain. Nat. Neurosci. 2010, 13, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.E.; Patapiutian, A.; Grandl, J. Arthur. Single Residues in the Outer Pore of TRPV1 and TRPV3 Have Temperature-Dependent Conformations. PLoS ONE 2013, 8, e59593. [Google Scholar]
- Yang, S.; Lu, X.; Wang, Y.; Xu, L.; Lai, R. A paradigm of thermal adaptation in penguins and elephants by tuning cold activation in TRPM8. Proc. Natl. Acad. Sci. USA 2020, 117, 8633–8638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa-Pagán, J.O.; Iversen, E.S.; Grandl, J. TRPV1 temperature activation is specifically sensitive to strong decreases in amino acid hydrophobicity. Sci. Rep. 2017, 7, 549. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Liu, B.; Qin, F. Modular thermal sensors in temperature-gated transient receptor potential (TRP) channels. Proc. Natl. Acad. Sci. USA 2011, 108, 11109–11114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero-Morales, J.F.; Gracheva, E.O.; Julius, D. Cytoplasmic Ankyrin Repeats of Transient Receptor Potential A1 (TRPA1) Dictate Sensitivity to Thermal and Chemical Stimuli. Biophys. J. 2012, 102, 23a. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Panzano, V.C.; Chang, E.C.; Ni, L.; Dainis, A.M.; Jenkins, A.M.; Regna, K.; Muskavitch, M.A.T.; Garrity, P.A. Modulation of TRPA1 thermal sensitivity enables sensory discrimination in Drosophila. Nature 2012, 481, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Bellemer, A.; Haidun, Y.; Ken, H.; Jessica, R.; Hwang, R.J.; Pitt, G.S.; Tracey, W.D. Thermosensory and Nonthermosensory Isoforms of Drosophila melanogaster TRPA1 Reveal Heat-Sensor Domains of a ThermoTRP Channel. Cell Rep. 2012, 1, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Dhaka, A.; Viswanath, V.; Patapoutian, A. Trpion channels and temperature sensation. Annu. Rev. Neurosci. 2006, 29, 135–161. [Google Scholar] [CrossRef] [Green Version]
- Jabba, S.; Goyal, R.; Sosa-Pagán, J.O.; Moldenhauer, H.; Wu, J.; Kalmeta, B.; Bandell, M.; Latorre, R.; Patapoutian, A.; Grandl, J. Directionality of Temperature Activation in Mouse TRPA1 Ion Channel Can Be Inverted by Single-Point Mutations in Ankyrin Repeat Six. Neuron 2014, 82, 1017–1031. [Google Scholar] [CrossRef] [Green Version]
- Philipp, E.; Portner, H.-O.; Abele, D. Mitochondrial ageing of a polar and a temperate mud clam. Mech. Ageing Dev. 2005, 126, 610–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matozzo, V.; Monari, M.; Foschi, J.; Papi, T.; Cattani, O.; Marin, M.G. Exposure to anoxia of the clam Chamelea gallina: I: Effects on immune responses. J. Exp. Mar. Biol. Ecol. 2005, 325, 163–174. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Hu, X.; Xun, X.; Zhang, J.; Guo, X.; Jiao, W.; Zhang, L.; Liu, W.; Wang, J. Scallop genome reveals molecular adaptations to semi-sessile life and neurotoxins. Nature Commun. 2017, 8, 1721. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Peng, N.; Zuo, Z.; Urs, L.; Qi, Z.; Zheng, Y.; Yong, Z. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallaace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, K.B. GeneDoc: Analysis and visualization of genetic variation. EMBNEW News 1997, 4, 14. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | TRPC | TRPM | TRPV | TRPA | TRPP | TRPML | TRPN | Total | References |
|---|---|---|---|---|---|---|---|---|---|
| Homo sapiens | 6 | 8 | 6 | 1 | 3 | 3 | 0 | 27 | [11] |
| Mus musculus | 7 | 8 | 6 | 1 | 3 | 3 | 0 | 28 | [11] |
| Danio rerio | 8 | 6 | 4 | 2 | 4 | 2 | 1 | 27 | [10] |
| Ciona intestinalis | 8 | 2 | 2 | 4 | 1 | 9 | 1 | 27 | [9] |
| Chlamys farreri | 4 | 4 | 4 | 2 | 1 | 1 | 1 | 17 | in this article |
| Drosophila melanogaste | 3 | 1 | 2 | 4 | 1 | 1 | 1 | 13 | [8] |
| Caenorhabditis elegans | 3 | 4 | 5 | 2 | 1 | 1 | 1 | 17 | [12] |
| Gene Name | Gene ID | cDNA Length (bp) | ORF Length (bp) | Exons No. | Introns No. | Amino Acid No. | Molecular Weight (kDa) | Theoretical pI | AlpHa No. | Beta No. | Colis No. | Turns No. | GRAVY of PD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CfTRPC-α | scaffold62023.33.1 | 3103 | 2847 | 15 | 14 | 948 | 109.29 | 5.42 | 44 | 47 | 63 | 68 | 0.601 |
| CfTRPC-α1 | scaffold51977.10.1 | 3131 | 2493 | 13 | 12 | 830 | 96.25 | 7.3 | 40 | 46 | 53 | 61 | 0.612 |
| CfTRPC-γ | scaffold17795.40.2 | 2721 | 2592 | 10 | 9 | 863 | 99.15 | 6.85 | 40 | 44 | 57 | 62 | 0.355 |
| CfTRPC-like | scaffold53059.6 | 3295 | 2724 | 10 | 9 | 907 | 104.40 | 7.95 | 36 | 46 | 69 | 68 | 0.526 |
| CfTRPM2a | scaffold28131.6 | 4248 | 4248 | 27 | 26 | 1415 | 161.45 | 8.48 | 71 | 76 | 104 | 103 | 0.656 |
| CfTRPM2b | scaffold64377.34 | 4875 | 4875 | 34 | 33 | 1624 | 184.87 | 5.98 | 74 | 85 | 119 | 116 | 0.764 |
| CfTRPM1 | scaffold1.4.1 | 5847 | 3072 | 17 | 16 | 1023 | 116.70 | 6.75 | 54 | 52 | 61 | 61 | 0.56 |
| CfTRPM3 | scaffold52123.6.1 | 5373 | 5052 | 25 | 24 | 1683 | 191.64 | 6.67 | 83 | 87 | 113 | 112 | 0.572 |
| CfTRPA1 | scaffold61233.39 | 3453 | 3453 | 26 | 25 | 1150 | 128.73 | 7.05 | 62 | 63 | 64 | 70 | 1.184 |
| CfTRPA1-homolog | scaffold63953.15 | 5170 | 3564 | 30 | 29 | 1187 | 133.42 | 6.5 | 63 | 60 | 68 | 65 | 0.994 |
| CfTRPN | scaffold29423.13 | 4716 | 4716 | 26 | 25 | 1571 | 172.82 | 7.87 | 82 | 78 | 99 | 86 | 0.745 |
| CfTRPV-Nan | scaffold63985.6 | 3322 | 1980 | 12 | 11 | 659 | 75.07 | 6.03 | 22 | 32 | 40 | 43 | 0.503 |
| CfTRPV1 | scaffold57527.16 | 2487 | 2487 | 17 | 16 | 828 | 95.56 | 8.71 | 43 | 45 | 47 | 55 | 0.623 |
| CfTRPV-Lav1 | scaffold24983.12 | 3473 | 3072 | 16 | 15 | 1023 | 116.59 | 8.86 | 50 | 52 | 67 | 68 | 0.75 |
| CfTRPV-Lav2 | scaffold64873.6 | 2283 | 1953 | 1 | 0 | 650 | 73.75 | 7.57 | 33 | 36 | 45 | 37 | 0.634 |
| CfTRPP | scaffold64453.6.1 | 3767 | 2571 | 15 | 14 | 856 | 98.04 | 5.36 | 32 | 41 | 73 | 70 | −0.265 |
| CfTRPML | scaffold5673.1 | 2257 | 1614 | 13 | 12 | 537 | 60.54 | 6.37 | 28 | 36 | 27 | 35 | 1.162 |
| Gene | H. sapiens | M. musculus | X. tropicalis | D. rerio | C. gigas | D. melanogaster | C. elegans |
|---|---|---|---|---|---|---|---|
| CfTRPC-α | 29.05% | 29.05% | 28.22% | 28.50% | 48.07% | 32.92% | 26.98% |
| CfTRPC-α1 | 32.82% | 32.82% | 31.75% | 31.28% | 69.28% | 34.21% | 26.37% |
| CfTRPC-γ | 40.52% | 40.39% | 40.62% | 40.03% | 77.74% | 57.09% | 52.68% |
| CfTRPC-like | 35.93% | 35.80% | 35.88% | 35.34% | 66.15% | 40.53% | 50.41% |
| CfTRPM2a | 36.70% | 37.04% | 32.18% | 31.91% | 45.35% | 34.33% | 27.42% |
| CfTRPM2b | 31.00% | 31.50% | 31.47% | 31.80% | 46.90% | 32.06% | 34.53% |
| CfTRPM1 | 41.77% | 41.55% | 38.29% | 38.99% | 53.39% | 39.16% | 30.95% |
| CfTRPM3 | 49.45% | 49.69% | 50.29% | 47.41% | 66.51% | 51.36% | 54.58% |
| CfTRPA1 | 37.25% | 36.80% | 36.62% | 36.56% | 55.41% | 40.56% | 24.39% |
| CfTRPA1-homolog | 27.72% | 27.65% | 24.63% | 26.11% | 25.57% | 26.18% | 23.62% |
| CfTRPN | / | / | / | 57.13% | 79.95% | 50.55% | 49.86% |
| CfTRPV-Nan | 27.47% | 26.93% | 27.74% | 27.29% | 69.54% | 47.57% | 39.69% |
| CfTRPV1 | 19.80% | 19.69% | 20.23% | 19.12% | 56.05% | 28.12% | 21.70% |
| CfTRPV-Lav1 | 28.62% | 28.45% | 29.61% | 25.87% | 57.25% | 45.91% | 45.64% |
| CfTRPV-Lav2 | 25.47% | 25.66% | 25.71% | 23.46% | 53.82% | 43.07% | 42.00% |
| CfTRPP | 50.55% | 50.19% | 50.06% | 52.53% | 76.98% | 37.88% | 37.03% |
| CfTRPML | 26.59% | 25.95% | 26.93% | 26.10% | 28.87% | 27.41% | 23.40% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, C.; Yang, Z.; Liu, Z.; Wang, S.; Yu, H.; Cui, C.; Hu, Y.; Xing, Q.; Hu, J.; Huang, X.; et al. A Systematical Survey on the TRP Channels Provides New Insight into Its Functional Diversity in Zhikong Scallop (Chlamys farreri). Int. J. Mol. Sci. 2021, 22, 11075. https://doi.org/10.3390/ijms222011075
Peng C, Yang Z, Liu Z, Wang S, Yu H, Cui C, Hu Y, Xing Q, Hu J, Huang X, et al. A Systematical Survey on the TRP Channels Provides New Insight into Its Functional Diversity in Zhikong Scallop (Chlamys farreri). International Journal of Molecular Sciences. 2021; 22(20):11075. https://doi.org/10.3390/ijms222011075
Chicago/Turabian StylePeng, Cheng, Zujing Yang, Zhi Liu, Shenhai Wang, Haitao Yu, Chang Cui, Yuqing Hu, Qiang Xing, Jingjie Hu, Xiaoting Huang, and et al. 2021. "A Systematical Survey on the TRP Channels Provides New Insight into Its Functional Diversity in Zhikong Scallop (Chlamys farreri)" International Journal of Molecular Sciences 22, no. 20: 11075. https://doi.org/10.3390/ijms222011075