
Transcriptomic Analyses Reveal 2 and 4 Family Members of Cytochromes P450 (CYP) Involved in LPS Inflammatory Response in Pharynx of Ciona robusta

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Next-Generation Sequencing Transcriptomics Analysis of Ciona robusta Highlights the Effects of LPS on Cytochrome P450 Transcripts Involved in Inflammation
2.2. Alignment and Phylogenetic Analysis of C. robusta Cytochrome P450 LPS Deregulated Genes
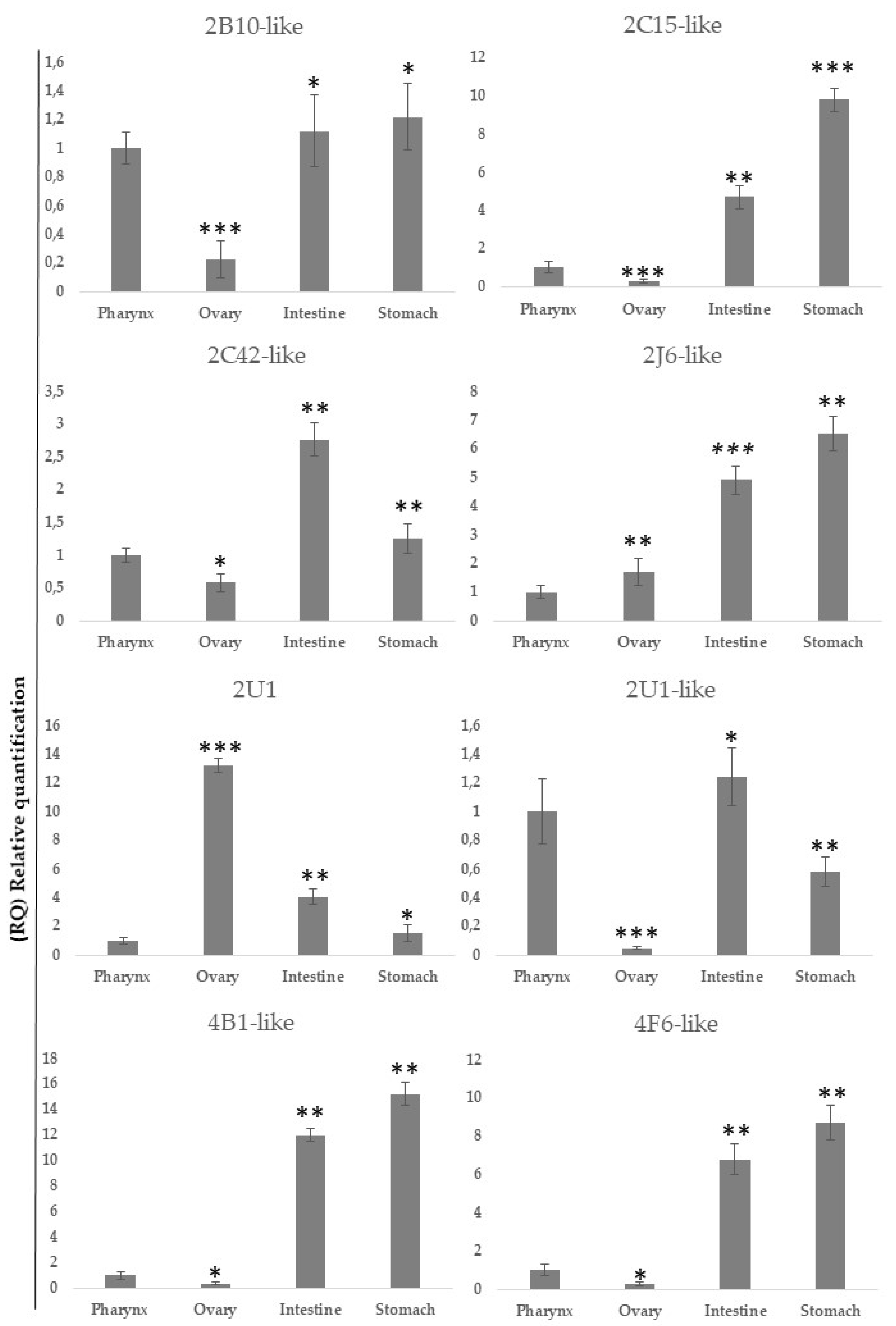
2.3. Differential Gene Expression of Cytochrome P450 Genes in Different Tissues
2.4. Analyses of the Expression of Cytochrome 450 and Cytokines Genes under LPS Exposure
2.5. miRNA-Target Interaction Prediction
2.6. Cytochrome and Inflammation miRNA-Target Network Reconstruction
2.7. Transcription Factor Orthologue Identification of Cyp450 Response Elements
3. Discussion
4. Materials and Methods
4.1. Tunicates and LPS Injection
4.2. RNA Sequencing (RNA-Seq)
4.3. Alignments and Phylogenetic Analyses
4.4. qRT-PCR
4.5. Functional Enrichment Analysis
4.6. miRNA–mRNA Target Interaction Prediction
4.7. Study of Evolution Pattern of C. robusta Homologues microRNA in Animal Genome
4.8. Network Construction and Visualisation
4.9. Transcription Factor Orthologue Identification and TF Binding Site Prediction
4.10. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ortiz de Montellano, P.R. (Ed.) Substrate oxidation by cytochrome P450 enzymes. In Cytochrome P450: Structure, Mechanism and Biochemistry, 4th ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2015; pp. 111–1762. [Google Scholar]
- Palrasu, M.; Siddavaram, N. Cytochrome P450 Structure, Function and Clinical Significance: A Review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar]
- Guengerich, F.P.; Munro, A.W. Unusual cytochrome P450 enzymes and reactions. J. Biol. Chem. 2013, 288, 17065–17073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guengerich, F.P. Human cytochrome P450 enzymes. In Cytochrome P450: Structure, Mechanism and Biochemistry, 4th ed.; Ortiz de Montellano, P.R., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2015; pp. 523–785. [Google Scholar]
- Gotoh, O. Evolution of Cytochrome P450 Genes from the Viewpoint of Genome Informatics. Biol. Pharm. Bull. 2012, 35, 812–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.R.; Goldstone, J.V.; Stegeman, J.J. The cytochrome P450 genesis locus: The origin and evolution of animal cytochrome P450s. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, S. Evolution by Gene Duplication; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.K.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.-K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 1970, 453, 1064–1071. [Google Scholar] [CrossRef] [Green Version]
- Kuraku, S. Insights into cyclostome phylogenomics: Pre-2R or post-2R. Zool. Sci. 2008, 25, 960–968. [Google Scholar] [CrossRef]
- Kuraku, S.; Meyer, A.; Kuratani, S. Timing of genome duplications relative to the origin of the vertebrates: Did cyclostomes diverge before or after? Mol. Biol. Evol. 2009, 26, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Van de Peer, Y. From 2R to 3R: Evidence for a fish-specific genome duplication (FSGD). BioEssays 2005, 27, 937–945. [Google Scholar] [CrossRef] [Green Version]
- Siegel, N.; Hoegg, S.; Salzburger, W.; Braasch, I.; Meyer, A. Comparative genomics of ParaHox clusters of teleost fishes: Gene cluster breakup and the retention of gene sets following whole genome duplications. BMC Genom. 2007, 8, 312. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.B.; Feyereisen, R.A. Cluster of cytochrome P450 genes of the CYP6 family in the house fly. DNA Cell Biol. 1995, 14, 73–82. [Google Scholar] [CrossRef]
- Nelson, D.R.; Zeldin, D.C.; Hoffman, S.M.G.; Maltais, L.J.; Wain, H.M.; Nebert, D.W. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 2004, 14, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Chen, S. Epigenetic regulation of cytochrome P450 enzymes and clinical implication. Curr. Drug Metab. 2015, 16, 86–96. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Moriya, N.; Kataoka, H.; Fujino, H.; Nishikawa, J.; Kugawa, F. Effect of lipopolysaccharide on the xenobiotic-induced expression and activity of hepatic cytochrome P450 in mice. Biol. Pharm. Bull. 2012, 35, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Y.; Zhang, C.; Wang, H.; Ji, Y.L.; Wang, S.F.; Zhao, L.; Chen, X.; Xu, D.X. Tumor necrosis factor alpha partially contributes to lipopolysaccharide induced downregulation of CYP3A in fetal liver: Its repression by a low dose LPS pretreatment. Toxicol. Lett. 2008, 179, 71–77. [Google Scholar] [CrossRef]
- Warren, G.W.; Poloyac, S.M.; Gary, D.S.; Mattson, M.P.; Blouin, R.A. Hepatic cytochrome P-450 expression in tumor necrosis factor-alpha receptor (p55/p75) knockout mice after endotoxin administration. J. Pharmacol. Exp. Ther. 1999, 288, 945–950. [Google Scholar]
- Siewert, E.; Bort, R.; Kluge, R.; Heinrich, P.C.; Castell, J.; Jover, R. Hepatic cytochrome P450 down-regulation during aseptic inflammation in the mouse is interleukin 6 dependent. Hepatology 2000, 32, 49–55. [Google Scholar] [CrossRef]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Tsagkogeorga, G.; Turon, X.; Hopcroft, R.R.; Tilak, M.-K.; Feldstein, T.; Shenkar, N.; Loya, Y.; Huchon, D.; Douzery, E.J.P.; Delsuc, F. An updated 18S rRNA phylogeny of tunicates based on mixture and secondary structure models. BMC Evol. Biol. 2009, 9, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, L.; Swalla, B.J. Molecular phylogeny of the protochordates: Chordate evolution. Can. J. Zool. 2005, 83, 24–33. [Google Scholar] [CrossRef]
- Satoh, N. The ascidian tadpole larva: Comparative molecular development and genomics. Nat. Rev. Genet. 2003, 4, 285–295. [Google Scholar] [CrossRef]
- Vizzini, A. Gene expression and regulation of molecules involved in pharynx inflammatory response induced by LPS in Ciona intestinalis. Invertebr. Surviv. J. 2017, 14, 119–128. [Google Scholar]
- Giacomelli, S.; Melillo, D.; Lambris, J.D.; Pinto, M.R. Immune competence of the Ciona intestinalis pharynx: Complement system-mediated activity. Fish Shellfish Immunol. 2012, 33, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Martinucci, G.B.; Dallai, R.; Burighel, P.; Lane, N.J. Different functions of tight junctions in the ascidian branchial basket. Tissue Cell 1988, 20, 119–132. [Google Scholar] [CrossRef]
- Bonura, A.; Vizzini, A.; Salerno, G.; Parrinello, N.; Longo, V.; Colombo, P. Isolation and expression of a novel MBL-like collectin cDNA enhanced by LPS injection in the body wall of the ascidian Ciona intestinalis. Mol. Immunol. 2009, 46, 2389–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizzini, A.; Parrinello, D.; Sanfratello, M.A.; Salerno, G.; Cammarata, M.; Parrinello, N. Inducible galectins are expressed in the inflamed pharynx of the ascidian Ciona intestinalis. Fish Shellfish Immunol. 2012, 32, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizzini, A.; Falco, F.D.; Parrinello, D.; Sanfratello, M.A.; Mazzarella, C.; Parrinello, N.; Cammarata, M. Ciona intestinalis interleukin 17-like genes expression is upregulated by LPS challenge. Dev. Comp. Immunol. 2015, 48, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Di Falco, F.; Parrinello, D.; Sanfratello, M.A.; Cammarata, M. Transforming growth factor b (CiTGF-b) gene expression is induced in the inflammatory reaction of Ciona intestinalis. Dev. Comp. Immunol. 2016, 55, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Parisi, M.G.; Di Falco, F.; Cardinale, L.; Cammarata, M.; Arizza, V. Identification of CPE and GAIT elements in 3’UTR of macrophage migration inhibitory factor (MIF) involved in inflammatory response induced by LPS in Ciona robusta. Mol. Immunol. 2018, 9, 66–74. [Google Scholar] [CrossRef]
- Vizzini, A.; Bonura, A.; La Paglia, L.; Fiannaca, A.; La Rosa, M.; Urso, A.; Arizza, V. ceRNA Network Regulation of TGF-β, WNT, FOXO, Hedgehog Pathways in the Pharynx of Ciona robusta. Int. J. Mol. Sci. 2021, 22, 3497. [Google Scholar] [CrossRef]
- Arizza, V.; Bonura, A.; La Paglia, L.; Urso, A.; Pinsino, A.; Vizzini, A. Transcriptional and in silico analyses of MIF cytokine and T LR signalling interplay in the LPS inflammatory response of Ciona robusta. Sci. Rep. 2020, 10, 11339. [Google Scholar] [CrossRef]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef]
- Nelson, D.R. The cytochrome P450 homepage. Hum. Genom. 2009, 4, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. Metazoan cytochrome P450 evolution. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1998, 121, 15–22. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Styrishave, B.; Lobner-Olsen, A.; Andersen, O. Marine invertebrate cytochrome P450: Emerging insights from vertebrate and insects analogies. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 363–381. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J.V.; Hamdoun, A.; Cole, B.J.; Howard-Ashby, M.; Nebert, D.W.; Scally, M.; Dean, M.; Epel, D.; Hahn, M.E.; Stegeman, J.J. The chemical defensome: Environmental sensing and response genes in the Strongylocentrotus purpuratus genome. Dev. Biol. 2006, 300, 366–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iber, H.; Sewer, M.B.; Barclay, T.B.; Mitchell, S.R.; Li, T.; Morgan, E.T. Modulation of drug metabolism in infectious and inflammatory diseases. Drug Metab. Rev. 1999, 31, 29–41. [Google Scholar] [CrossRef]
- Aitken, A.E.; Richardson, T.A.; Morgan, E.T. Regulation of drug metabolizing enzymes and transporters in inflammation. Annu. Rev. Pharmacol. Toxicol. 2006, 46, 123–149. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.H.; Lee, M.G. Effects of endotoxin derived from Escherichia coli lipopolysaccharide on the pharmacokinetics of drugs. Arch. Pharmacal Res. 2008, 31, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.T. Suppression of constitutive cytochrome P-450 gene expression in livers of rats undergoing an acute phase response to endotoxin. Mol. Pharmacol. 1989, 36, 699–707. [Google Scholar]
- Moriya, N.; Kataoka, H.; Fujino, H.; Nishikawa, J.; Kugawa, F. Different expression patterns of hepatic cytochrome P450s during anaphylactic or lipopolysaccharide-induced inflammation. Pharmazie 2014, 69, 142–147. [Google Scholar] [PubMed]
- Cuia, X.; Kalsotraa, A.; Robidab, A.M.; Matzilevichc, D.; Moorec, A.N.; Boehmea, C.L.; Morgan, E.T.; Dashc, P.K.; Strobela, H.W. Expression of cytochromes P450 4F4 and 4F5 in infection and injury models of inflammation . Biochim. Biophys. Acta BBA Gen. Subj. 2003, 1619, 325–331. [Google Scholar] [CrossRef]
- Zordoky, B.; El-Kadi, A. Role of NF-kB in the Regulation of Cytochrome P450 Enzymes. Curr. Drug Metab. 2009, 10, 164–178. [Google Scholar] [CrossRef] [PubMed]
- De Jong, L.M.; Jiskoot, W.; Swen, J.J.; Manson, M.L. Distinct Effects of Inflammation on Cytochrome P450 Regulation and Drug Metabolism: Lessons from Experimental Models and a Potential Role for Pharmacogenetics. Genes 2020, 11, 1509. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ferguson, S.S.; Negishi, M.; Goldstein, J.A. Identification of constitutive androstane receptor and glucocorticoid receptor binding sites in the CYP2C19 promoter. Mol. Pharmacol. 2003, 64, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.S.; Lecluyse, E.L.; Negishi, M.; Goldstein, J.A. Regulation of human CYP2C9 by the constitutive androstane receptor: Discovery of a new distal binding site. Mol. Pharmacol. 2002, 62, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Barwick, J.L.; Simon, C.M.; Pierce, A.M.; Safe, S.; Blumberg, B.; Guzelian, P.S.; Evans, R.M. Reciprocal activation of xenobiotic response genes by nuclear receptors SXR/PXR and CAR. Genes Dev. 2000, 14, 3014–3023. [Google Scholar] [CrossRef] [Green Version]
- Pascussi, J.M.; Gerbal-Chaloin, S.; Pichard-Garcia, L.; Daujat, M.; Fabre, J.M.; Maurel, P.; Vilarem, M.J. Interleukin-6 negatively regulates the expression of pregnane X receptor and constitutively activated receptor in primary human hepatocytes. Biochem. Biophys. Res. Commun. 2000, 274, 707–713. [Google Scholar] [CrossRef]
- Sun, H.Y.; Yan, Y.J.; Li, Y.H.; Lv, L. Reversing effects of ginsenosides on LPS-induced hepatic CYP3A11/3A4 dysfunction through the pregnane X receptor. J. Ethnopharmacol. 2019, 229, 246–255. [Google Scholar] [CrossRef]
- Beigneux, A.P.; Moser, A.H.; Shigenaga, J.K.; Grunfeld, C.; Feingold, K.R. Reduction in cytochrome P-450 enzyme expression is associated with repression of CAR (constitutive androstane receptor) and PXR (pregnane X receptor) in mouse liver during the acute phase response. Biochem. Biophys. Res. Commun. 2002, 293, 145–149. [Google Scholar] [CrossRef]
- Zhou, X.; Li, X.; Wang, X.; Jin, X.; Shi, D.; Wang, J.; Bi, D. Cecropin B represses CYP3A29 expression through activation of the TLR2/4-NF-_ B/PXR signaling pathway. Sci. Rep. 2016, 6, 27876. [Google Scholar] [CrossRef]
- Yagi, K.; Satou, Y.; Mazet, F.; Shimeld, S.M.; Degnan, B.; Rokhsar, D.; Levine, M.; Kohara, Y.; Satoh, N. A genomewide survey of developmentally relevant genes in Ciona intestinalis. III. Genes for Fox, ETS, nuclear receptors and NF-kB. Dev. Genes Evol. 2003, 213, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Krasowski, M.D.; Reschly, E.J.; Ekins, S. Intrinsic disorder in nuclear hormone receptors. J. Proteome Res. 2008, 7, 4359–4372. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, R.; Gissi, C.; Pennati, R.; Manni, L. Morphological evidence that the molecularly determined Ciona intestinalis type A and type B are different species: Ciona robusta and Ciona intestinalis. J. Zool. Syst. Evol. Res. 2015, 53, 186–193. [Google Scholar] [CrossRef]
- Pennati, R.; Ficetola, G.F.; Brunetti, R.; Caicci, F.; Gasparini, F.; Griggio, F.; Sato, A.; Stach, T.; Kaul-Strehlow, S.; Gissi, C.; et al. Morphological differences between larvae of the Ciona intestinalis species complex: Hints for a valid taxonomic definition of distinct species. PLoS ONE 2015, 10, e0122879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputi, L.; Andreakis, N.; Mastrototaro, F.; Cirino, P.; Vassillo, M.; Sordino, P. Cryptic speciation in a model invertebrate chordate. Proc. Natl. Acad. Sci. USA 2007, 104, 9364–9369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannelli, F.; Pesole, G.; Sordino, P.; Gissi, C. Mitogenomics reveals two cryptic species in Ciona intestinalis. Trends Genet. 2007, 23, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yu, X.; Wu, C.; Zhu, T.; Wang, W.; Zheng, X.; Jin, H. RNA sequencing-based longitudinal transcriptomic profiling gives novel insights into the disease mechanism of generalized pustular psoriasis. BMC Med. Genom. 2018, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiannaca, A.; La Rosa, M.; La Paglia, L.; Rizzo, R.; Urso, A. MiRNATIP: A SOM-based miRNA-target interactions predictor. BMC Bioinform. 2016, 17, 321. [Google Scholar] [CrossRef] [Green Version]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.S.; Tam, W.L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of microRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ENSEMBL ID | NAME | LogFC | LogCPM | p-Value | Adj p-Value | Logp Value | Chr. Position |
|---|---|---|---|---|---|---|---|
| ENSCING00000023704 | cytochrome P450 2B10 (LOC101242210) | 2.326276709 | −0.954064903 | 7.85 × 10−5 | 0.001978725 | 4.104962459 | Chromosome 11: 2,221,309–2,225,001 reverse strand. |
| ENSCING00000014704 | cytochrome P450 2C15-like (LOC100186646) | 2.08035981 | 0.149882558 | 5.87 × 10−5 | 0.001551545 | 4.23164033 | Chromosome 10: 1,471,100–1,473,817 |
| ENSCING00000017012 | cytochrome P450 2J6-like (LOC100175185) | 3.28873837 | −1.420263798 | 2.46 × 10−6 | 0.000105594 | 5.608489757 | Chromosome 8: 5,194,025–5,197,825 reverse strand. |
| ENSCING00000005903 | cytochrome P450 2U1(LOC100185251) | 1.511849854 | 0.613848759 | 0.002721531 | 0.035008689 | 2.565186709 | Scaffold HT001236.1: 8,305–10,991 |
| ENSCING00000004714 | cytochrome P450 4B1-like (LOC100182965) | 2.251530553 | 1.669810098 | 6.12 × 10−6 | 0.000231035 | 5.213430612 | Chromosome 1: 7,364,051–7,366,204 forward strand |
| ENSCING00000006567 | cytochrome P450 4F6-like (LOC100186171) | 2.228936378 | 2.87892154 | 4.97 × 10−6 | 0.000194829 | 5.303654163 | Chromosome 7: 1,548,748–1,552,535 forward strand. |
| ENSCING00000009298 | cytochrome P450 2U1-like (LOC100182684) | -2.611064465 | 5.303935694 | 7.82 × 10−8 | 5.38 × 10−6 | 7.106747277 | Chromosome 5: 2,876,599–2,880,776 forward strand. |
| ENSCING00000013919 | cytochrome P450 2C42-like (LOC100184869) | -3.458407214 | 7.389835139 | 6.28 × 10−12 | 1.17 × 10−9 | 11.20175682 | Chromosome 10: 3,837,453–3,843,016 reverse strand. |
| Cyp P450 | Lengh | I-Helix [AG]G-x-[DE]T[TS] | K-Helix E-x-x-R | Meander Coil FDDER | Heme Loop F-x-x-G-x-R-x-C-x-G |
|---|---|---|---|---|---|
| 2B10-like | 365 | AGTETS | ESLR | -------------CL- | |
| 2C15 | 509 | AGTETS | KQLL | FRPER | FSVGLRSCIG |
| 2C42 | 506 | AGVETT | DQLX | FNPHR | FSIGPRYCMG |
| 2U1 | 501 | AGTDTT | EQLF | FKPDR | FNVGQRSCLG |
| 2U1-like | 506 | AGTETT | LQLL | FKPER | FSVGPRQCLG |
| 2J6 | 504 | AGNETT | LQLC | FDPSR | FSLGPRQCIG |
| 4B1 | 439 | - | -DIE | FNPDR | FSAGSRNCIG |
| 4F6 | 538 | EGHDTT | KEIR | YDPER | FSAGPRNCIG |
| Conserved miRNAs | Target Gene ID | ||||
|---|---|---|---|---|---|
| cin-miR-196-3p | ENSCING00000004714 | ENSCING00000009298 | ENSCING00000005269 | ENSCING00000013600 | |
| (cyp450 4B1-like) | (cyp450 2U1-like) | (Il-17-3) | (Tgf-ß-na2) | ||
| cin-miR-92e-5p | ENSCING00000004714 | ENSCING00000022988 | ENSCING00000023704 | ||
| (cyp450 4B1-like) | (cyp450 2U1-like) | (cyp450 2C13, male-specific-like) | |||
| cin-let-7f-5p | ENSCING00000014704 | ENSCING00000005269 (Il-17-3) | |||
| (cyp450 4B1-like) | |||||
| cin-let-7d-5p | ENSCING00000022988 | ENSCING00000006967 | |||
| (cyp450 4F4-like) | (Il-17-1) | ||||
| cin-let-7e | ENSCING00000004714 (cyp450 4B1-like) | ENSCING00000006967 (Il-17-1) | |||
| cin-let-7d-3p | ENSCING00000013919 (cyp450 2C42-like) | ENSCING00000013600 (Tgf-ß-na2) | |||
| cin-miR-200-3p | ENSCING00000008093 | ENSCING00000023704 | |||
| (Tgf-ß-na1) | (cyp450 2C13, male-specific-like) | ||||
| Specie-Specific miRNAs | Target Gene ID | ||||
| cin-miR-5596b-3p | ENSCING00000005903 | ENSCING00000013919 | ENSCING00000014704 | ENSCING00000017012 | |
| (cyp450 2U1) | (cyp450 2C42-like) | (cyp450 4B1-like) | (cyp450 2J6-like) | ||
| cin-miR-4085-3p | ENSCING00000022988 | ENSCING00000014704 | ENSCING00000006967 | ||
| (cyp450 4F4-like) | (cyp450 4B1-like) | (Il-17-1) | |||
| cin-miR-4036-3p | ENSCING00000009298 | ENSCING00000013919 | ENSCING00000008093 | ||
| (cyp450 2U1-like) | (cyp450 2C42-like) | (Tgf-ß-na1) | |||
| cin-miR-4200-3p | ENSCING00000006567 | ENSCING00000017012 | |||
| (cyp450 4F6-like) | (cyp450 2J6-like) | ||||
| cin-miR-4148-5p | ENSCING00000005903 | ENSCING00000006567 | |||
| (cyp450 2U1) | (cyp450 4F6-like) | ||||
| cin-miR-4116-5p | ENSCING00000005903 | ENSCING00000006967 | |||
| (cyp450 2U1) | (Il-17-1) | ||||
| cin-miR-4119-3p | ENSCING00000006567 | ENSCING00000006967 | |||
| (cyp450 4F6-like) | (Il-17-1) | ||||
| cin-miR-4011a-5p | ENSCING00000006567 | ENSCING00000008093 | |||
| (cyp450 4F6-like) | (Tgf-ß-na1) | ||||
| cin-miR-4064-5p | ENSCING00000013919 | ENSCING00000008093 | |||
| (cyp450 2C42-like) | (Tgf-ß-na1) | ||||
| cin-miR-5604-3p | ENSCING00000017012 | ENSCING00000008093 | |||
| (cyp450 2J6-like) | (Tgf-ß-na1) | ||||
| cin-miR-4020b-5p | ENSCING00000005269 | ENSCING00000013600 | |||
| (Il-17-3) | (Tgf-ß-na2) | ||||
| cin-miR-4065-3p | ENSCING00000006567 | ENSCING00000013600 | |||
| (cyp450 4F6-like) | (Tgf-ß-na2) | ||||
| cin-miR-4121-3p | ENSCING00000014704 | ENSCING00000013600 | |||
| (cyp450 4B1-like) | (Tgf-ß-na2) | ||||
| cin-miR-4203-3p | ENSCING00000022988 | ENSCING00000013919 | |||
| (cyp450 4F4-like) | (cyp450 2C42-like) | ||||
| cin-miR-4083-5p | ENSCING00000005903 | ENSCING00000014704 | |||
| (cyp450 2U1) | (cyp450 4B1-like) | ||||
| cin-miR-5596a-3p | ENSCING00000005903 | ENSCING00000014704 | |||
| (cyp450 2U1) | (cyp450 4B1-like) | ||||
| cin-miR-4001b-5p | ENSCING00000014704 | ENSCING00000017012 | |||
| (cyp450 4B1-like) | (cyp450 2J6-like) | ||||
| cin-miR-5605-3p | ENSCING00000005903 | ENSCING00000017012 | |||
| (cyp450 2U1) | (cyp450 2J6-like) | ||||
| Conserved miRNAS | Species-Specific miRNAs |
|---|---|
| cin-let-7d | cin-miR-3575-3p |
| cin-let-7e | cin-miR-4001b-1-3p |
| cin-let-7f-5p | cin-miR-4009a-3p |
| cin-miR-183-5p | cin-miR-4009b-5p |
| cin-miR-196-3p | cin-miR-4011a-5p |
| cin-miR-200-5p | cin-miR-4020b-3p |
| cin-miR-672 | cin-miR-4036-3p |
| cin-miR-7-5p | cin-miR-4037-5p |
| cin-miR-92c-5p | cin-miR-4043-5p |
| cin-miR-92e-5p | cin-miR-4047-3p |
| cin-miR-4058-5p | |
| cin-miR-4064-5p | |
| cin-miR-4064-5p | |
| cin-miR-4065-3p | |
| cin-miR-4077d-5p | |
| cin-miR-4102-5p | |
| cin-miR-4115-5p | |
| cin-miR-4116-5p | |
| cin-miR-4117-5p | |
| cin-miR-4119-3p | |
| cin-miR-4121-3p | |
| cin-miR-4148-5p | |
| cin-miR-4187-5p | |
| cin-miR-4207-3p | |
| cin-miR-4219-5p | |
| cin-miR-5596a-3p | |
| cin-miR-5596b-3p | |
| cin-miR-5603-3p | |
| cin-miR-5607-3p | |
| cin-miR-5609-5p | |
| cin-miR-5612-5p | |
| cin-miR-4219-5p |
| Name | GenBank no. |
|---|---|
| Ciona robusta Cyp P450 2B10-like | XP_026692382.1 |
| Branchiostoma floridae Cyp P450 2J6-like | XP_035667220 |
| Branchiostoma floridae Cyp 450 2U1-like | XP_035667221.1 |
| Homo sapiens Cyp 450 2B6 | NP_000758.1 |
| Homo sapiens Cyp 450 1A1 | NP_000490.1 |
| Homo sapiens Cyp 450 2D6 | NP_000097.3 |
| Homo sapiens Cyp 450 26A1 | NP_000774.2 |
| Homo sapiens Cyp 450 4A22 | NP_001010969.2 |
| Homo sapiens Cyp 450 4A11 | NP_000769.2 |
| Homo sapiens Cyp 450 2C18 | NP_000763.1 |
| Homo sapiens Cyp 450 2A7 | NP_000755.2 |
| Ciona robusta Cyp 450 2C15-like | XP_002123518.1 |
| Branchiostoma floridae Cyp 450 2C15-like | XP_035694363.1 |
| Drosophila melanogaster C15 | NP_476873.2 |
| Ciona robusta Cyp 450 2C42-like | XP_018669209.2 |
| Sus scrofa Cyp 450 2C42 | NP_001161307.1 |
| Aplysia californica Cyp 450 2C42 | XP_012936173.2 |
| Crassostrea gigas Cyp 450 2C42 | XP_011452086.2 |
| Bos Taurus Cyp 450 2C42 | XP_005225699.1 |
| Ciona robusta Cyp 450 2J6-like | XP_002129285.1 |
| Mus musculus Cyp 450 2J6 | NP_034138.3 |
| Haplochromis burtoni Cyp 450 2J6 | XP_014186874.1 |
| Xenopus tropicalis Cyp 450 2J6 | XP_002935636.1 |
| Strongylocentrotus purpuratus Cyp 450 2J6 | XP_001178133.3 |
| Ciona robusta Cyp 450 2U1 | XP_002119562.1 |
| Homo sapiens Cyp 450 2U1 | NP_898898.1 |
| Mus musculus Cyp 450 2U1 | NP_082092.2 |
| Ciona robusta Cyp 450 2U1-licke | XP_026690452.1 |
| Homo sapiens Cyp 450 4F2 | sp|P78329.1| |
| Homo sapiens Cyp 450 4F3 | sp|Q08477.2| |
| Homo sapiens Cyp 450 4B1 | NP_001093242.1 |
| Mus musculus Cyp 450 4B1 | NP_031849.1 |
| Bos taurus Cyp 450 4B1 | NP_001069670.1 |
| Haplochromis burtoni Cyp 450 4B1 | XP_014190098.1 |
| Xenopus tropicalis Cyp 450 4B1 | XP_012817051.1 |
| Branchiostoma floridae Cyp 450 4B1-like | XP_035680110.1 |
| Drosophila melanogaster Cyp 450 | AAC47424.1 |
| Ciona robusta 4B1 | XP_002125043.1 |
| Ciona robusta 4F6 | XP_002123011.3 |
| Gene | Primer Sequence (5′-3′) |
|---|---|
| Mif1 | 5′-GCTTGCAGCGCTTTTGATG-3′ 5′-AAACGGGTTCCAGAAACTCCTAA-3′ |
| Mif2 | 5′-CCATGAAGCAACGAGGGAAA-3′ 5′-TTCTTGGCTGCGAGTTGGT-3′ |
| Cyp 2B10 | 5′-CAAGGCCCAGCGTTTCAG-3′ 5′-CATTGCTGTGGGCTTCGAT-3′ |
| Cyp 2C15 | 5′-CAACGACAAGCATCGAACTCA-3′ 5′-TTGGCGATAACAGGCATACCA-3′ |
| Cyp 2C42 | 5′-TCGTCATTTTAGGTCGGTGATG-3′ 5′-TCAGTCATAGCTCGATACGAATACG-3′ |
| Cyp 2J6 | 5′-TCCTAAATGGCAAGATCGCATA-3′ 5′-AAACTCCGTTCTCACCGATATTG-3′ |
| Cyp 2U1 | 5′-TGGTCGAAGATCCGAACGA-3′ 5′-ACAACTGCTCTTTGTTCCACCAT-3′ |
| Cyp 2U1-like | 5′-AATGCAAAAATGGAGCAGAAAGT-3′ 5′-CCCGGCTCCCCATACG-3′ |
| Cyp 4B1 | 5′-AAAACGAGCCCAACGTACCA-3′ 5′-TAAAGGTCCAAACCATGTTGTCA-3′ |
| Cyp 4F6 | 5′-GGAGATGGTCTGTTGACAAGCA-3′ 5′-CAGGCGTTAGAAGCCTTCTGTT-3′ |
| Tgfβ | 5′-TTTCAGGGACCCAAAAACGA-3′ 5′-GCCAGCTATAATGACATCCAAGGT-3′ |
| Tnfα | 5′-GCCTCCCATAGACCGTTGTTAA-3′ 5′-CGGGACACCTTCAGCACAT-3′ |
| Il17-1 | 5′- GCCGGGAACGTGACAGAA- 3′ 5′-GGCATGTTGATTGCGACCTT- 3′ |
| Il17-2 | 5′-GTGTAGCGGGTGCATTGCT-3′ 5′-GGCACCGACTTCCCAACA-3 |
| Il17-3 | 5′-CAAAGCGGAGCCTTCAATGT-3′ 5′-GCTTCTTTGCTCGACACTTGTG-3′ |
| Nf-kB | 5′-GCCGACGTACTGCTTTGCA-3 5′-GCCAGCCACCACGATGTT-3″ |
| Actin | 5′-TGATGTTGCCGCACTCGTA-3 5′-TCGACAATGGATCCGGT-3″ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizzini, A.; Bonura, A.; La Paglia, L.; Fiannaca, A.; La Rosa, M.; Urso, A.; Mauro, M.; Vazzana, M.; Arizza, V. Transcriptomic Analyses Reveal 2 and 4 Family Members of Cytochromes P450 (CYP) Involved in LPS Inflammatory Response in Pharynx of Ciona robusta. Int. J. Mol. Sci. 2021, 22, 11141. https://doi.org/10.3390/ijms222011141
Vizzini A, Bonura A, La Paglia L, Fiannaca A, La Rosa M, Urso A, Mauro M, Vazzana M, Arizza V. Transcriptomic Analyses Reveal 2 and 4 Family Members of Cytochromes P450 (CYP) Involved in LPS Inflammatory Response in Pharynx of Ciona robusta. International Journal of Molecular Sciences. 2021; 22(20):11141. https://doi.org/10.3390/ijms222011141
Chicago/Turabian StyleVizzini, Aiti, Angela Bonura, Laura La Paglia, Antonino Fiannaca, Massimo La Rosa, Alfonso Urso, Manuela Mauro, Mirella Vazzana, and Vincenzo Arizza. 2021. "Transcriptomic Analyses Reveal 2 and 4 Family Members of Cytochromes P450 (CYP) Involved in LPS Inflammatory Response in Pharynx of Ciona robusta" International Journal of Molecular Sciences 22, no. 20: 11141. https://doi.org/10.3390/ijms222011141