
Dissecting the Molecular Regulation of Natural Variation in Growth and Senescence of Two Eutrema salsugineum Ecotypes

 ,
,
Abstract
:1. Introduction
2. Results
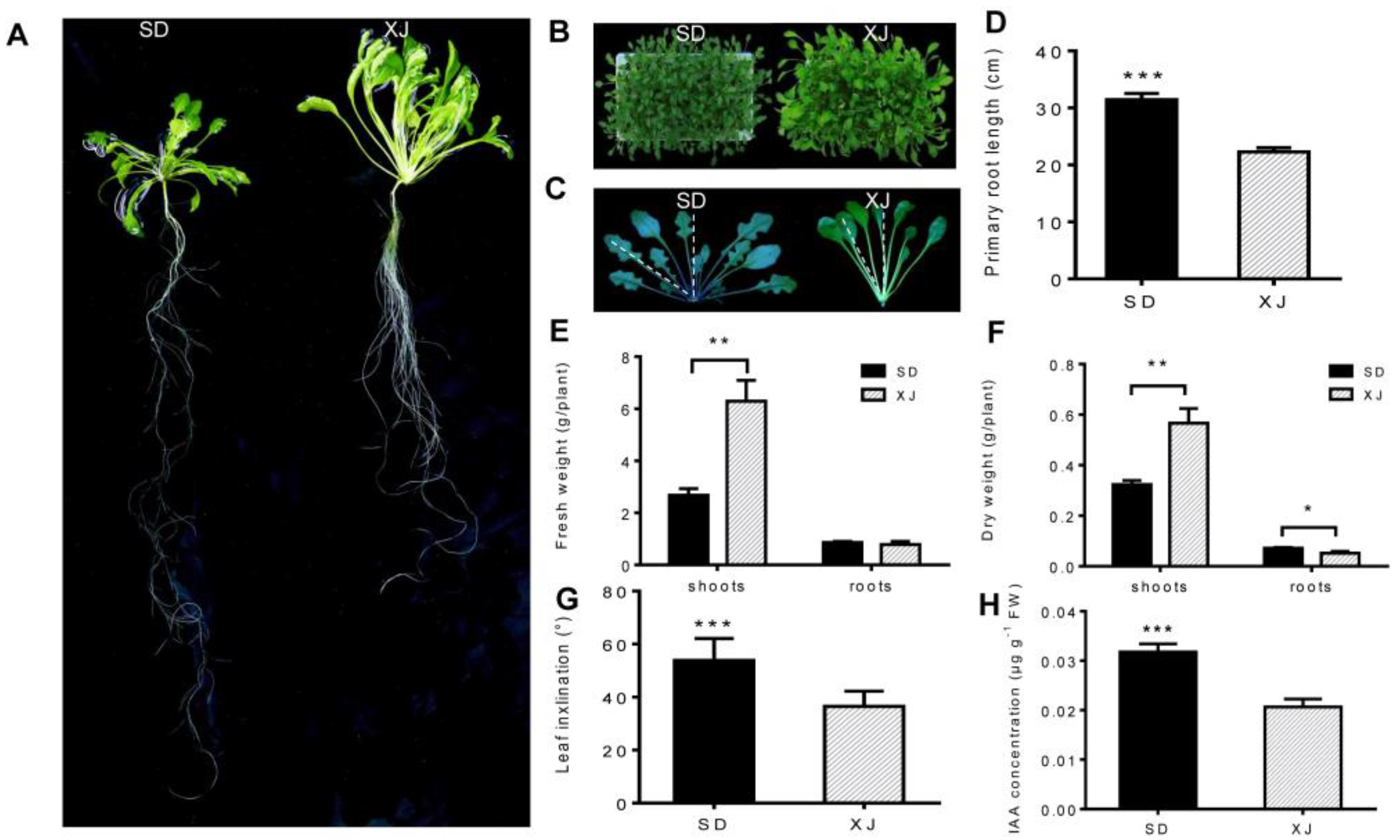
2.1. Morphological and Physiological Comparison
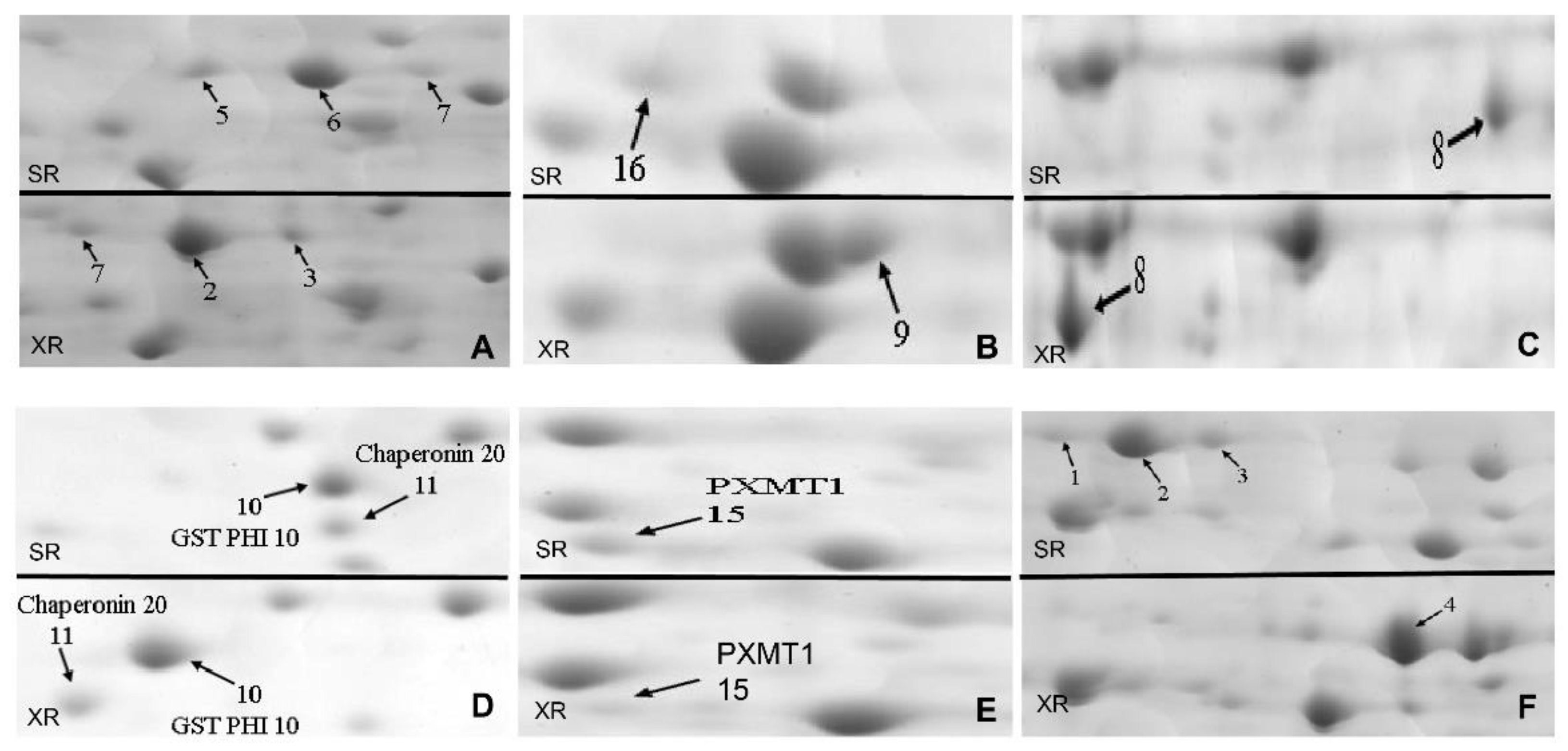
2.2. Identification of Differently Expressed Protein Spots from Two Ecotypes by 2-DE and ESI-MS/MS
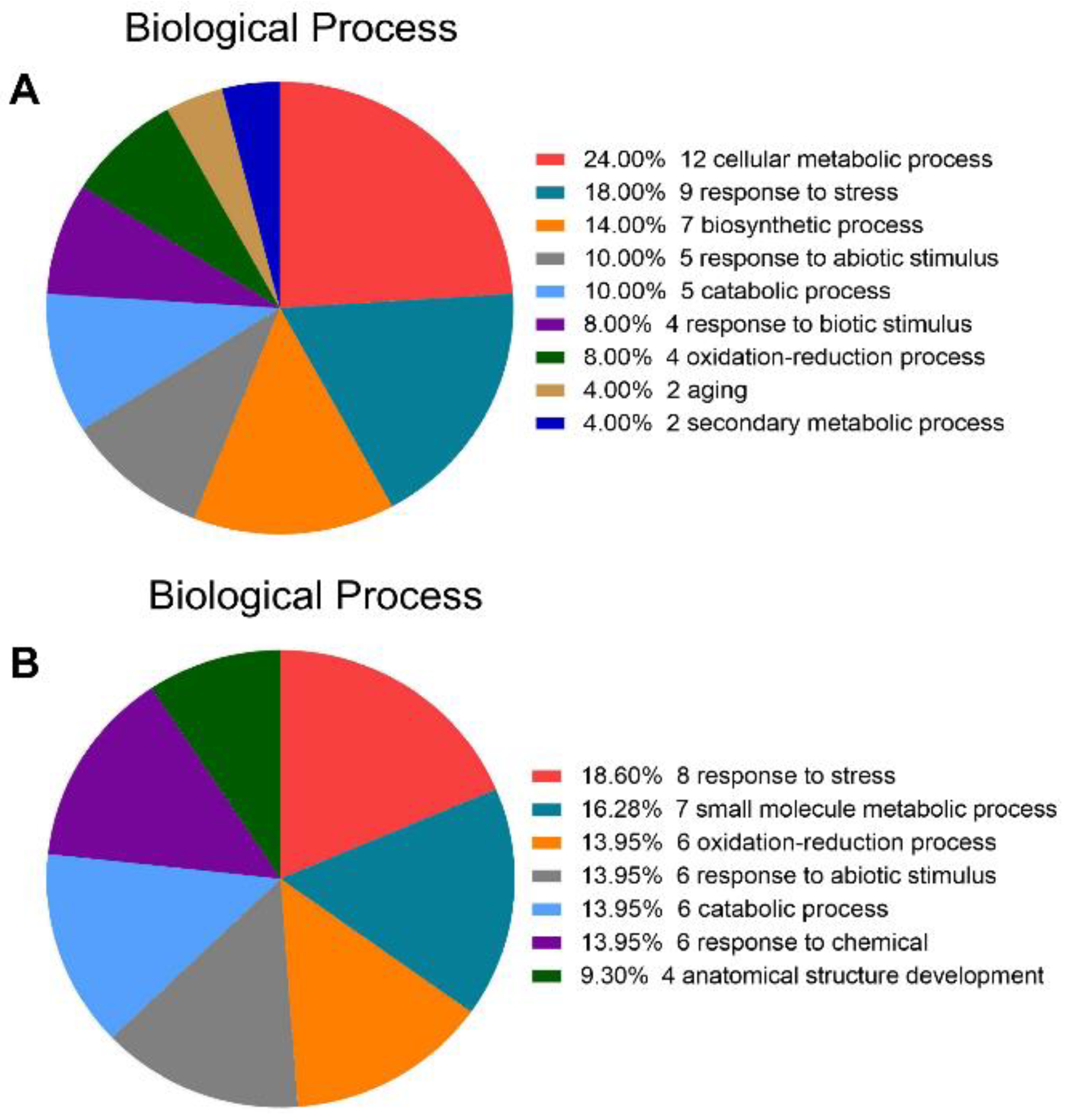
2.3. Functional Annotation of Different Proteins
2.4. Analysis of Differentially Expressed Protein Spots from Two Ecotypes
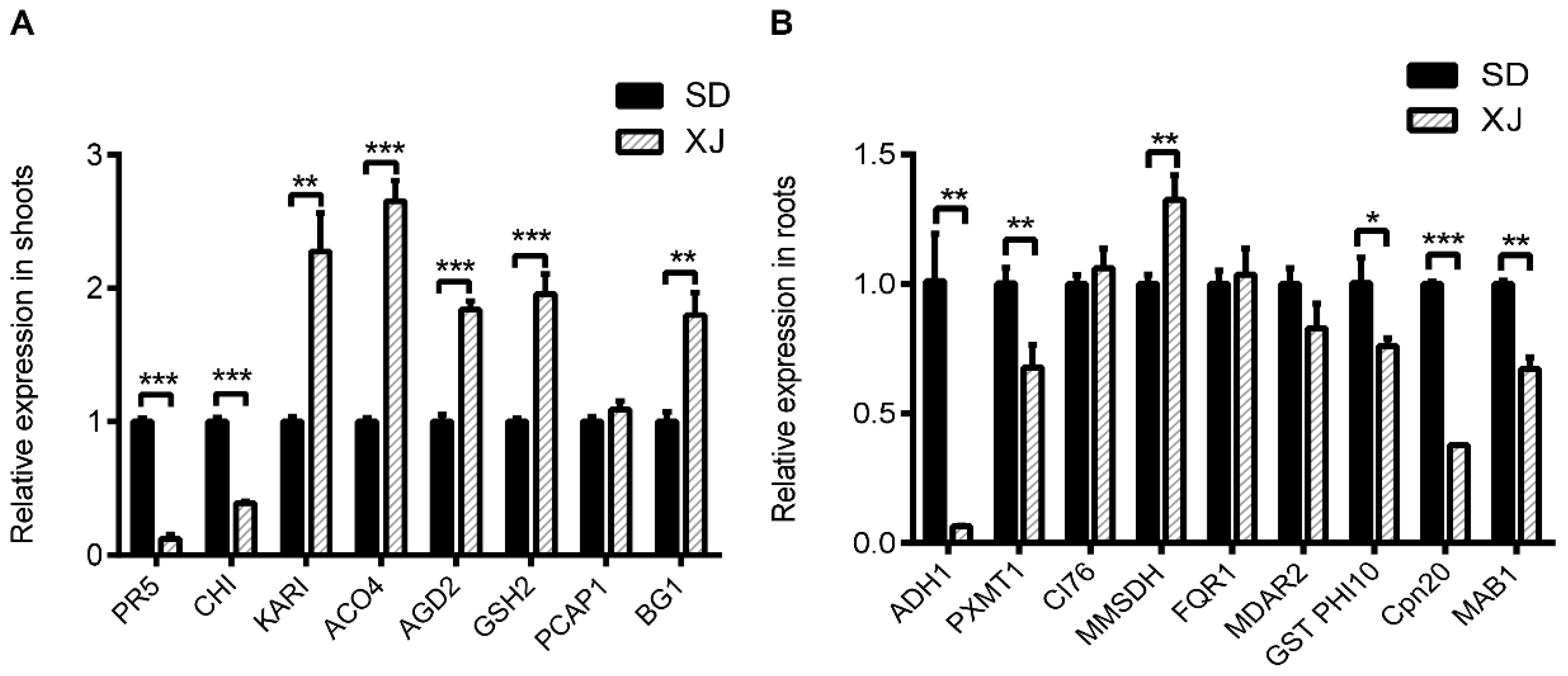
2.5. Comparison the Expression Patterns of Genes Encoding Some Differential Proteins
3. Discussion
3.1. Variations in Morpho-Physiological Traits
3.2. Stress and Defense-Related Proteins in Shoots
3.3. The Mechanism of Natural Variation of Leaf Senescence between Two Eutrema Ecotypes
3.4. Proteins Related to Energy Metabolism in Roots
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Growth Parameter Measurements and Morphological Analysis
4.3. Determination of Chlorophyll Content
4.4. Phytohormone Analysis
4.5. The Measurement of ACO Concentration
4.6. Protein Extraction and 2-DE Analysis
4.7. Protein Digestion and Identification via ESI-MS/MS
4.8. Quantitative Real Time PCR
4.9. Functional Annotation
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bressan, R.A.; Zhang, C.; Zhang, H.; Hasegawa, P.M.; Bohnert, H.J.; Zhu, J.-K. Learning from the Arabidopsis experience. Plant Physiol. 2001, 127, 1354–1360. [Google Scholar] [CrossRef]
- Inan, G.; Zhang, Q.; Li, P.; Wang, Z.; Cao, Z.; Zhang, H.; Zhang, C.; Quist, T.M.; Goodwin, S.M.; Zhu, J.; et al. Salt cress. A halophyte and cryophyte Arabidopsis relative model system and its applicability to molecular genetic analyses of growth and development of extremophiles. Plant Physiol. 2004, 135, 1718–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taji, T.; Seki, M.; Satou, M.; Sakurai, T.; Kobayashi, M.; Ishiyama, K.; Narusaka, Y.; Narusaka, M.; Zhu, J.K.; Shinozaki, K. Comparative genomics in salt tolerance between Arabidopsis and Arabidopsis-related halophyte salt cress using Arabidopsis microarray. Plant Physiol. 2004, 135, 1697–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Q.; Li, P.; Ma, S.; Indu Rupassara, S.; Bohnert, H.J. Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 2005, 44, 826–839. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Mane, S.P.; Sioson, A.A.; Robinet, C.V.; Heath, L.S.; Bohnert, H.J.; Grene, R. Effects of chronic ozone exposure on gene expression in Arabidopsis thaliana ecotypes and in Thellungiella halophila. Plant Cell Environ. 2006, 29, 854–868. [Google Scholar] [CrossRef] [Green Version]
- Griffith, M.; Timonin, M.; Wong, A.C.E.; Gray, G.R.; Akhter, S.R.; Saldanha, M.; Rogers, M.A.; Weretilnyk, E.A.; Moffatt, B. Thellungiella: An Arabidopsis-related model plant adapted to cold temperatures. Plant Cell Environ. 2007, 30, 529–538. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.-M.; Weretilnyk, E.; Barak, S.; Rothstein, S.J. The Arabidopsis halophytic relative Thellungiella halophila tolerates nitrogen-limiting conditions by maintaining growth, nitrogen uptake, and assimilation. Plant Physiol. 2008, 147, 1168–1180. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Wang, Z.-L.; Li, P.-H.; Fredricksen, M.; Gong, Z.-Z.; Kim, C.S.; Zhang, C.; Bohnert, H.J.; Zhu, J.-K.; Bressan, R.A.; Hasegawa, P.M.; et al. Expressed sequence tags from Thellungiella halophila, a new model to study plant salt-tolerance. Plant Sci. 2004, 166, 609–616. [Google Scholar] [CrossRef]
- Wong, C.E.; Li, Y.; Labbe, A.; Guevara, D.; Nuin, P.; Whitty, B.; Diaz, C.; Golding, G.B.; Gray, G.R.; Weretilnyk, E.A.; et al. Transcriptional profiling implicates novel interactions between abiotic stress and hormonal responses in Thellungiella, a close relative of Arabidopsis. Plant Physiol. 2006, 140, 1437–1450. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.B.; Caldwell, M.M. The scale of nutrient heterogeneity around individual plants and its quantification with geostatistics. Ecology 1993, 74, 612–614. [Google Scholar] [CrossRef]
- Yu, F.C.; Fei Hai, Y.; Ming, D. Scale-dependent spatial heterogeneity of vegetation in Mu Us sandy land, a semi-arid area of China. Plant Ecol. 2002, 162, 135–142. [Google Scholar]
- Mal, T.K.; Lovett-Doust, J. Phenotypic plasticity in vegetative and reproductive traits in an invasive weed, Lythrum salicaria (Lythraceae), in response to soil moisture. Am. J. Bot. 2005, 92, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Turesson, G. The species and the variety as ecological units. Hereditas 1922, 3, 100–113. [Google Scholar] [CrossRef]
- Savolainen, O.; Lascoux, M.; Merila, J. Ecological genomics of local adaptation. Nat. Rev. Genet. 2013, 14, 807–820. [Google Scholar] [CrossRef]
- Guevara, D.R.; Champigny, M.J.; Tattersall, A.; Dedrick, J.; Wong, C.E.; Li, Y.; Labbe, A.; Ping, C.-L.; Wang, Y.; Nuin, P. Transcriptomic and metabolomic analysis of Yukon Thellungiella plants grown in cabinets and their natural habitat show phenotypic plasticity. BMC Plant Biol. 2012, 12, 175. [Google Scholar] [CrossRef] [Green Version]
- Amtmann, A. Learning from evolution: Thellungiella generates new knowledge on essential and critical components of abiotic stress tolerance in plants. Mol. Plant. 2009, 2, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Bar, M.; Ori, N. Leaf development and morphogenesis. Development 2014, 141, 4219–4230. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Yoshimura, K.; Tamoi, M.; Takeda, T.; Shigeoka, S. Alternative mRNA splicing of 3′-terminal exons generates ascorbate peroxidase isoenzymes in spinach (Spinacia oleracea) chloroplasts. Biochem. J. 1997, 328, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Ahn, H.J.; Eom, S.J.; Yoon, H.-J.; Lee, B.I.; Cho, H.; Suh, S.W. Crystal structure of class I acetohydroxy acid isomeroreductase from Pseudomonas aeruginosa. J. Mol. Biol. 2003, 328, 505–515. [Google Scholar] [CrossRef]
- Cummins, I.; Dixon, D.P.; Freitag-Pohl, S.; Skipsey, M.; Edwards, R. Multiple roles for plant glutathione transferases in xenobiotic detoxification. Drug Metab. Rev. 2011, 43, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Considine, M.J.; Foyer, C.H. Redox regulation of plant development. Antioxid. Redox Signal. 2014, 21, 1305–1326. [Google Scholar] [CrossRef] [Green Version]
- Urbancsok, J.; Bones, A.M.; Kissen, R. Arabidopsis mutants impaired in glutathione biosynthesis exhibit higher sensitivity towards the glucosinolate hydrolysis product allyl-isothiocyanate. Sci. Rep. 2018, 8, 9809. [Google Scholar] [CrossRef]
- Hudson, A.O.; Singh, B.K.; Leustek, T.; Gilvarg, C. An LL-diaminopimelate aminotransferase defines a novel variant of the lysine biosynthesis pathway in plants. Plant Physiol. 2006, 140, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Anjum, N.A.; Gill, R.; Kaushik, M.; Hasanuzzaman, M.; Pereira, E.; Ahmad, I.; Tuteja, N.; Gill, S.S. ATP-sulfurylase, sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 2015, 6, 210. [Google Scholar] [CrossRef] [Green Version]
- Ide, Y.; Nagasaki, N.; Tomioka, R.; Suito, M.; Kamiya, T.; Maeshima, M. Molecular properties of a novel, hydrophilic cation-binding protein associated with the plasma membrane. J. Exp. Bot. 2007, 58, 1173–1183. [Google Scholar] [CrossRef]
- Stintzi, A.; Heitz, T.; Prasad, V.; Wiedemann-Merdinoglu, S.; Kauffmann, S.; Geoffroy, P.; Legrand, M.; Fritig, B. Plant "pathogenesis-related" proteins and their role in defense against pathogens. Biochimie 1993, 75, 687–706. [Google Scholar] [CrossRef]
- Mishina, T.E.; Zeier, J. Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in Arabidopsis. Plant J. 2007, 50, 500–513. [Google Scholar] [CrossRef]
- Kuo, W.Y.; Huang, C.H.; Liu, A.C.; Cheng, C.P.; Li, S.H.; Chang, W.C.; Weiss, C.; Azem, A.; Jinn, T.L. CHAPERONIN 20 mediates iron superoxide dismutase (FeSOD) activity independent of its co-chaperonin role in Arabidopsis chloroplasts. New Phytol. 2013, 197, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.Y.; Huang, C.H.; Jinn, T.L. Chaperonin 20 might be an iron chaperone for superoxide dismutase in activating iron superoxide dismutase (FeSOD). Plant Signal. Behav. 2013, 8, e23074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-Carboxylic Acid Oxidase (ACO): The enzyme that makes the plant hormone ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeles, F.B.; Morgan, P.W.; Saltveit, M.E. Ethylene in Plant Biology; Academic Press: London, UK, 1992. [Google Scholar]
- Johnsen, U.; Schonheit, P. Characterization of cofactor-dependent and cofactor-independent phosphoglycerate mutases from Archaea. Extremophiles 2007, 11, 647–657. [Google Scholar] [CrossRef]
- Zhao, Z.; Assmann, S.M. The glycolytic enzyme, phosphoglycerate mutase, has critical roles in stomatal movement, vegetative growth, and pollen production in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 5179–5189. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Klodmann, J.; Braun, H.P. Proteomic approach to characterize mitochondrial complex I from plants. Phytochemistry 2011, 72, 1071–1080. [Google Scholar] [CrossRef]
- Laskowski, M.J.; Dreher, K.A.; Gehring, M.A.; Abel, S.; Gensler, A.L.; Sussex, I.M. FQR1, a novel primary auxin-response gene, encodes a flavin mononucleotide-binding quinone reductase. Plant Physiol. 2002, 128, 578–590. [Google Scholar] [CrossRef]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross talk between H2O2 and interacting signal molecules under plant stress response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [Green Version]
- Stiti, N.; Missihoun, T.D.; Kotchoni, S.O.; Kirch, H.H.; Bartels, D. Aldehyde dehydrogenases in Arabidopsis thaliana: Biochemical requirements, metabolic pathways, and functional analysis. Front. Plant Sci. 2011, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.C.; Bartels, D. Comparative study of the aldehyde dehydrogenase (ALDH) gene superfamily in the glycophyte Arabidopsis thaliana and Eutrema halophytes. Ann. Bot. 2015, 115, 465–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, N.; Takahashi, H.; Kitano, H.; Matsuoka, M.; Akao, S.; Uchimiya, H.; Komatsu, S. Proteome approach to characterize the methylmalonate-semialdehyde dehydrogenase that is regulated by gibberellin. J. Proteome Res. 2005, 4, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Liu, W.; Yao, Y.; Wei, Y.; Chan, Z. Alcohol dehydrogenase 1 (ADH1) confers both abiotic and biotic stress resistance in Arabidopsis. Plant Sci. 2017, 262, 24–31. [Google Scholar] [CrossRef]
- Dixon, D.P.; Adrian, L.; Robert, E. Plant glutathione transferases. Genome Biol. 2002, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente van Bentem, S.; Anrather, D.; Dohnal, I.; Roitinger, E.; Csaszar, E.; Joore, J.; Buijnink, J.; Carreri, A.; Forzani, C.; Lorkovic, Z.J.; et al. Site-specific phosphorylation profiling of Arabidopsis proteins by mass spectrometry and peptide chip analysis. J. Proteome Res. 2008, 7, 2458–2470. [Google Scholar] [CrossRef]
- Song, M.; Xu, W.; Xiang, Y.; Jia, H.; Zhang, L.; Ma, Z. Association of jacalin-related lectins with wheat responses to stresses revealed by transcriptional profiling. Plant Mol.Biol. 2013, 84, 95–110. [Google Scholar] [CrossRef]
- Li, J.; Steen, H.; Gygi, S.P. Protein profiling with cleavable isotope-coded affinity tag (cICAT) reagents: The yeast salinity stress response. Mol. Cell. Proteomics 2003, 2, 1198–1204. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, A.J.; Yanovsky, M.J.; Botto, J.F. Germination variation in Arabidopsis thaliana accessions under moderate osmotic and salt stresses. Ann. Bot. 2010, 106, 833–842. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Wang, Y.; Cheng, Z.; Ye, T.; Chan, Z. Analysis of natural variation in bermudagrass (Cynodon dactylon) reveals physiological responses underlying drought tolerance. PLoS ONE 2012, 7, e53422. [Google Scholar] [CrossRef] [Green Version]
- Millenaar, F.F.; Cox, M.C.; van Berkel, Y.E.; Welschen, R.A.; Pierik, R.; Voesenek, L.A.; Peeters, A.J. Ethylene-induced differential growth of petioles in Arabidopsis. Analyzing natural variation, response kinetics, and regulation. Plant Physiol. 2005, 137, 998–1008. [Google Scholar] [CrossRef] [Green Version]
- Nagasaki-Takeuchi, N.; Miyano, M.; Maeshima, M. A plasma membrane-associated protein of Arabidopsis thaliana AtPCaP1 binds copper ions and changes its higher order structure. J. Biochem. 2008, 144, 487–497. [Google Scholar] [CrossRef]
- Nagasaki, N.; Tomioka, R.; Maeshima, M. A hydrophilic cation-binding protein of Arabidopsis thaliana, AtPCaP1, is localized to plasma membrane via N-myristoylation and interacts with calmodulin and the phosphatidylinositol phosphates PtdIns(3,4,5)P3 and PtdIns(3,5)P2. FEBS J. 2008, 275, 2267–2282. [Google Scholar] [CrossRef]
- Kato, M.; Nagasaki-Takeuchi, N.; Ide, Y.; Tomioka, R.; Maeshima, M. PCaPs, possible regulators of PtdInsP signals on plasma membrane. Plant Signal. Behav. 2010, 5, 848–850. [Google Scholar] [CrossRef] [Green Version]
- Nagata, C.; Miwa, C.; Tanaka, N.; Kato, M.; Suito, M.; Tsuchihira, A.; Sato, Y.; Segami, S.; Maeshima, M. A novel-type phosphatidylinositol phosphate-interactive, Ca-binding protein PCaP1 in Arabidopsis thaliana: Stable association with plasma membrane and partial involvement in stomata closure. J. Plant Res. 2016, 129, 539–550. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Qin, T.; Zhang, Y.; Liu, X.; Sun, J.; Zhou, Y.; Zhu, L.; Zhang, Z.; Yuan, M.; et al. MDP25, a novel calcium regulatory protein, mediates hypocotyl cell elongation by destabilizing cortical microtubules in Arabidopsis. Plant Cell 2011, 23, 4411–4427. [Google Scholar] [CrossRef] [Green Version]
- Mauch, F.; Mauch-Mani, B.; Boller, T.J.P. Antifungal hydrolases in pea tissue: II. Inhibition of fungal growth by combinations of chitinase and β-1,3-glucanase. Plant Physiol. 1988, 88, 936–942. [Google Scholar] [CrossRef] [Green Version]
- Mauch, F.; Hadwiger, L.A.; Boller, T. Antifungal hydrolases in pea tissue: I. Purification and characterization of two chitinases and two β-1,3-glucanases differentially regulated during development and in response to fungal infection. Plant Physiol. 1988, 87, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Bolar, J.P.; Norelli, J.L.; Wong, K.-W.; Hayes, C.K.; Harman, G.E.; Aldwinckle, H.S. Expression of endochitinase from Trichoderma harzianum in transgenic apple increases resistance to apple scab and reduces vigor. Phytopathology 2000, 90, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Emani, C.; Garcia, J.M.; Lopata-Finch, E.; Pozo, M.J.; Uribe, P.; Kim, D.J.; Sunilkumar, G.; Cook, D.R.; Kenerley, C.M.; Rathore, K.S. Enhanced fungal resistance in transgenic cotton expressing an endochitinase gene from Trichoderma virens. Plant Biotechnol. J. 2003, 1, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Gentile, A.; Deng, Z.; La Malfa, S.; Distefano, G.; Domina, F.; Vitale, A.; Polizzi, G.; Lorito, M.; Tribulato, E. Enhanced resistance to Phoma tracheiphila and Botrytis cinerea in transgenic lemon plants expressing a Trichoderma harzianum chitinase gene. Plant Breed. 2007, 126, 146–151. [Google Scholar] [CrossRef]
- Cao, S.; Wang, Y.; Li, Z.; Shi, W.; Gao, F.; Zhou, Y.; Zhang, G.; Feng, J. Genome-wide identification and expression analyses of the chitinases under cold and osmotic stress in Ammopiptanthus nanus. Genes 2019, 10, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Anderson, J.M.; Ohm, H.W. Induction of wheat defense and stress-related genes in response to Fusarium graminearum. Genome 2005, 48, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Zhang, C.; Ge, W.; Zhang, Y.; Burlingame, A.L.; Guo, Y. Identification of NaCl stress-responsive apoplastic proteins in rice shoot stems by 2D-DIGE. J. Proteomics 2011, 74, 1045–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budak, H.; Akpinar, B.A.; Unver, T.; Turktas, M. Proteome changes in wild and modern wheat leaves upon drought stress by two-dimensional electrophoresis and nanoLC-ESI–MS/MS. Plant Mol.Biol. 2013, 83, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China-Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Koyama, T. A hidden link between leaf development and senescence. Plant Sci. 2018, 276, 105–110. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Lycett, G.W.; Grierson, D. Antisense gene that inhibits synthesis of the hormone ethylene in transgenic plants. Nature 1990, 346, 284–287. [Google Scholar] [CrossRef]
- Ververidis, P.; John, P. Complete recovery in vitro of ethylene-forming enzyme activity. Phytochemistry 1991, 30, 725–727. [Google Scholar] [CrossRef]
- Jing, H.C.; Sturre, M.J.G.; Hille, J.; Dijkwel, P.P. Arabidopsis onset of leaf death mutants identify a regulatory pathway controlling leaf senescence. Plant J. 2002, 32, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.-C.; Schippers, J.H.M.; Jacques, H.; Dijkwel, P.P. Ethylene-induced leaf senescence depends on age-related changes and OLD genes in Arabidopsis. J. Exp. Bot. 2005, 56, 421. [Google Scholar] [CrossRef]
- Datta, R.; Kumar, D.; Sultana, A.; Hazra, S.; Bhattacharyya, D.; Chattopadhyay, S. Glutathione regulates 1-aminocyclopropane-1-carboxylate synthase transcription via WRKY33 and 1-aminocyclopropane-1-carboxylate oxidase by modulating messenger RNA stability to induce ethylene synthesis during stress. Plant Physiol. 2015, 169, 2963. [Google Scholar] [PubMed]
- Gan, S.; Amasino, R.M. Making sense of senescence. Plant Physiol. 1997, 113, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucerova, Z.; Rac, M.; Mikulik, J.; Plihal, O.; Pospisil, P.; Bryksova, M.; Sedlarova, M.; Dolezal, K.; Spundova, M. The Anti-Senescence Activity of Cytokinin Arabinosides in Wheat and Arabidopsis Is Negatively Correlated with Ethylene Production. Int. J. Mol. Sci. 2020, 21, 8109. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Zhang, L.; Mei, J.; Chen, J.; Piao, Z.; Lee, G.; Dong, Y. Mutation of the rice TCM12 gene encoding 2,3-bisphosphoglycerate-independent phosphoglycerate mutase affects chlorophyll synthesis, photosynthesis and chloroplast development at seedling stage at low temperatures. Plant Biol. 2019, 21, 585–594. [Google Scholar] [CrossRef]
- James, M.; Poret, M.; Masclaux-Daubresse, C.; Marmagne, A.; Coquet, L.; Jouenne, T.; Chan, P.; Trouverie, J.; Etienne, P. SAG12, a major cysteine protease involved in nitrogen allocation during senescence for seed production in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 2052–2063. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Gan, S.-S. An abscisic acid-AtNAP transcription factor-SAG113 protein phosphatase 2C regulatory chain for controlling dehydration in senescing Arabidopsis leaves. Plant Physiol. 2012, 158, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Colombi, T.; Herrmann, A.M.; Vallenback, P.; Keller, T. Cortical cell diameter is key to energy costs of root growth in wheat. Plant Physiol. 2019, 180, 2049–2060. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.R.; Johnson, T.K.; Desler, M.; Luster, T.A.; Nowling, T.; Lewis, R.E.; Rizzino, A. Effects of B-Myb on gene transcription. J. Biol. Chem. 2002, 277, 4088–4097. [Google Scholar] [CrossRef] [Green Version]
- Grana, X.; De Lecea, L.; El-Maghrabi, M.R.; Urena, J.; Caellas, C.; Carreras, J.; Puigdomenech, P.; Pilkis, S.; Climent, F. Cloning and sequencing of a cDNA encoding 2,3-bisphosphoglycerate-independent phosphoglycerate mutase from maize. Possible relationship to the alkaline phosphatase family. J. Biol. Chem. 1992, 267, 12797–12803. [Google Scholar] [CrossRef]
- Graña, X.; de la Ossa, P.P.; Broceño, C.; Stöcker, M.; Garriga, J.; Puigdomènech, P.; Climent, F. 2,3-Bisphosphoglycerate-independent phosphoglycerate mutase is conserved among different phylogenic kingdoms. Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 1995, 112, 287–293. [Google Scholar] [CrossRef]
- Steffens, B.; Steffen-Heins, A.; Sauter, M. Reactive oxygen species mediate growth and death in submerged plants. Front. Plant Sci. 2013, 4, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israelsen, W.J.; Dayton, T.L.; Davidson, S.M.; Fiske, B.P.; Hosios, A.M.; Bellinger, G.; Li, J.; Yu, Y.; Sasaki, M.; Horner, J.W.; et al. PKM2 isoform-specific deletion reveals a differential requirement for pyruvate kinase in tumor cells. Cell 2013, 155, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaxton, W.C. The organization and regulation of plant glycolysis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xiao, W.; Luo, L.; Pang, J.; Rong, W.; He, C. Downregulation of OsPK1, a cytosolic pyruvate kinase, by T-DNA insertion causes dwarfism and panicle enclosure in rice. Planta 2012, 235, 25–38. [Google Scholar] [CrossRef]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth. Res. 2002, 73, 149–156. [Google Scholar] [CrossRef]
- Giavalisco, P.; Nordhoff, E.; Lehrach, H.; Gobom, J.; Klose, J. Extraction of proteins from plant tissues for two-dimensional electrophoresis analysis. Electrophoresis 2003, 24, 207–216. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Wang, B.; Wang, D.; Li, P.; Wang, L.; Yi, X.; Huang, Q.; Peng, M.; Guo, A. Comparative proteomics of Thellungiella halophila leaves from plants subjected to salinity reveals the importance of chloroplastic starch and soluble sugars in halophyte salt tolerance. Mol. Cell. Proteomics 2013, 12, 2174–2195. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Schwartz, D.; Elias, J.E.; Thoreen, C.C.; Cheng, D.; Marsischky, G.; Roelofs, J.; Finley, D.; Gygi, S.P. A proteomics approach to understanding protein ubiquitination. Nat. Biotechnol. 2003, 21, 921–926. [Google Scholar] [CrossRef]
- Henzel, W.J.; Billeci, T.M.; Stults, J.T.; Wong, S.C.; Grimley, C.; Watanabe, C. Identifying proteins from two-dimensional gels by molecular mass searching of peptide fragments in protein sequence databases. Proc. Natl. Acad. Sci. USA 1993, 90, 5011–5015. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Shamustakimova, A.O.; Leonova, T.G.; Taranov, V.V.; de Boer, A.H.; Babakov, A.V. Cold stress increases salt tolerance of the extremophytes Eutrema salsugineum (Thellungiella salsuginea) and Eutrema (Thellungiella) botschantzevii. J. Plant Physiol. 2017, 208, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Klopfenstein, D.V.; Zhang, L.; Pedersen, B.S.; Ramírez, F.; Warwick Vesztrocy, A.; Naldi, A.; Mungall, C.J.; Yunes, J.M.; Botvinnik, O.; Weigel, M.; et al. GOATOOLS: A python library for gene ontology analyses. Sci. Rep. 2018, 8, 10872. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | NCBI Accession | Exper. e | Theor. f | |||||
|---|---|---|---|---|---|---|---|---|
| No. a | Locus. b | No. c | Protein Name. d | pI/Mr | pI/Mr | Score. g | NP. h | Pattern. i |
| Cellular metabolic process | ||||||||
| XS4 | At3g58610 | XP_006402768 | ketol-acid reductoisomerase | 5.8/56 | 5.64/57.3 | 20 | 2 | appear |
| XS19 | At5g27380 | XP_024007192 | glutathione synthetase | 5.8/56 | 5.52/53.9 | 20 | 2 | appear |
| XS8 | At4g33680 | XP_024005372 | LL-diaminopimelate aminotransferase | 5.6/44 | 6.38/46.5 | 20 | 2 | appear |
| SS4 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.45/59 | 5.32/60.5 | 78 | 7 | decrease |
| SS5 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.6/59 | 5.32/60.5 | 384 | 36 | appear |
| SS27 | At3g22890 | XP_006406118 | ATP sulfurylase 1 | 6/44 | 6.34/51.5 | 70 | 7 | increase |
| XS28 | At3g22890 | XP_006406118 | ATP sulfurylase 1 | 6/44 | 6.34/51.5 | 60 | 6 | decrease |
| SS21 | At1g12050 | XP_006417270 | fumarylacetoacetase | 5.6/44 | 5.23/46.1 | 40 | 4 | appear |
| XS33 | At2g40010 | XP_006411245 | 60S acidic ribosomal protein P0-1 | 5.3/34 | 5.19/33.7 | 100 | 9 | equal |
| Response to stress | ||||||||
| SS6 | At4g20260 | XP_006413890 | plasma membrane-associated cation-binding protein 1 | 5/36 | 4.65/24.7 | 98 | 8 | appear |
| SS7 | At4g20260 | XP_006413890 | plasma membrane-associated cation-binding protein 1 | 5/36 | 4.65/24.7 | 66 | 5 | appear |
| SS8 | At4g20260 | XP_006413890 | plasma membrane-associated cation-binding protein 1 | 5/37 | 4.65/24.7 | 134 | 9 | appear |
| SS10 | At1g75040 | XP_006390358 | pathogenesis-related protein 5 | 5/25 | 4.65/22.8 | 200 | 20 | increase |
| SS14 | At2g43570 | XP_006397529 | endochitinase CHI | 5.3/27 | 5.84/29.8 | 40 | 4 | increase |
| SS29 | At3g57270 | XP_006409034 | probable glucan endo-1,3-beta-glucosidase BG1 | 5.3/35 | 8.94/37.7 | 20 | 2 | increase |
| XS15 | At2g30860 | XP_024004280 | glutathione S-transferase F9 | 6.7/25 | 6.17/24.1 | 30 | 3 | decrease |
| Response to abiotic stimulus | ||||||||
| XS12 | At5g20720 | XP_024011493 | 20 kDa chaperonin | 5.3/25 | 5.23/21.4 | 170 | 16 | increase |
| SS11 | At3g11630 | XP_006407428 | 2-Cys peroxiredoxin BAS1 | 5/23 | 5.01/22.4 | 460 | 46 | increase |
| SS12 | At3g11630 | XP_006407428 | 2-Cys peroxiredoxin BAS1 | 5.3/24 | 5.01/22.4 | 30 | 3 | decrease |
| XS11 | At3g11630 | XP_006407428 | 2-Cys peroxiredoxin BAS1 | 5/24 | 5.01/22.4 | 88 | 9 | decrease |
| Aging | ||||||||
| XS6 | At1g05010 | XP_006418070 | 1-aminocyclopropane-1-carboxylate oxidase 4 | 5/37 | 5.24/36.7 | 60 | 4 | appear |
| Unknown | ||||||||
| SS26 | At3g01500 | XP_006408514 | beta carbonic anhydrase 1 | 6.3/33 | 6.14/25.6 | 80 | 7 | equal |
| XS27 | At3g01500 | XP_006408514 | beta carbonic anhydrase 1 | 6.3/26.3 | 6.14/25.6 | 66 | 2 | equal |
| SS30 | At1g55480 | XP_006392635 | protein MET1 | 5.3/34 | 8.36/37.4 | 30 | 3 | equal |
| XS32 | At1g55480 | XP_006392635 | protein MET1 | 5.3/35 | 8.19/37.4 | 256 | 23 | equal |
| Spot | NCBI Accession | Exper. e | Theor. f | |||||
|---|---|---|---|---|---|---|---|---|
| No. a | Locus. b | No. c | Protein Name. d | pI/Mr | pI/Mr | Score. g | NP. h | Pattern. i |
| Small molecule metabolic process | ||||||||
| SR5 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.68/65 | 5.32/60.6 | 68 | 7 | basic |
| SR6 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.72/65 | 5.32/60.6 | 490 | 42 | basic |
| SR7 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 6/65 | 5.32/60.6 | 80 | 7 | basic |
| XR2 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.5/65 | 5.27/60.5 | 128 | 12 | acid |
| XR3 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.6/65 | 5.27/60.5 | 154 | 14 | acid |
| XR7 | At1g09780 | XP_006417539 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase 1 | 5.4/65 | 5.27/60.5 | 80 | 6 | acid |
| XR5 | At5g08570 | XP_006399365 | pyruvate kinase | 6.3/64 | 5.93/55 | 70 | 6 | appear |
| SR16 | At5g50850 | XP_006402055 | pyruvate dehydrogenase E1 component subunit beta-1 | 5.1/38 | 5.11/35.9 | 32 | 4 | decrease acid |
| XR9 | At5g50850 | XP_006402055 | pyruvate dehydrogenase E1 component subunit beta-1 | 5.3/36 | 5.11/35.9 | 58 | 5 | increase basic |
| SR4 | At2g45290 | XP_024010889 | transketolase-2 | 5.7/67 | 5.64/68.9 | 50 | 4 | appear |
| Oxidation-reduction process | ||||||||
| XR1 | At5g37510 | XP_006405867 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 1 | 6/68 | 5.72/77.9 | 70 | 6 | appear |
| XR13 | At5g54500 | XP_006401566 | NAD(P)H dehydrogenase (quinone) FQR1 | 6.3/25 | 5.96/21.8 | 136 | 11 | increase |
| SR8 | At5g03630 | XP_024011589 | monodehydroascorbate reductase 2 | 6.3/45 | 5.25/47.5 | 166 | 12 | decrease basic |
| XR8 | At5g03630 | XP_024011589 | monodehydroascorbate reductase 2 | 5.7/43 | 5.25/47.5 | 108 | 9 | increase acid |
| XR6 | At2g14170 | XP_006409665 | methylmalonate-semialdehyde dehydrogenase [acylating] | 6.3/64 | 8.97/64.7 | 82 | 7 | appear |
| Response to abiotic stimulus | ||||||||
| SR9 | At1g77120 | XP_006390122 | alcohol dehydrogenase class-P | 6.5/45 | 5.83/41.2 | 120 | 12 | increase |
| SR10 | At2g30870 | XP_006410185 | glutathione S-transferase F10 | 5.5/25 | 5.49/24.1 | 50.11 | 5 | decrease basic |
| XR10 | At2g30870 | XP_006410185 | glutathione S-transferase F10 | 5.3/25 | 5.49/24.1 | 148 | 15 | increase acid |
| SR11 | At5g20720 | XP_024011493 | 20 kDa chaperonin | 5.5/25 | 5.23/21.4 | 284 | 26 | decrease basic |
| XR11 | At5g20720 | XP_024011493 | 20 kDa chaperonin | 5.2/25 | 5.23/21.4 | 262 | 21 | increase acid |
| SR15 | At1g66700 | XP_006391421 | paraxanthine methyltransferase 1 | 5/42 | 5.34/39.8 | 68 | 6 | increase |
| SR1 | At3g16460 | XP_006406833 | jacalin-related lectin 34 | 5.3/68 | 5.31/72.5 | 50 | 5 | acid |
| SR2 | At3g16460 | XP_006406833 | jacalin-related lectin 34 | 5.4/68 | 5.31/72.5 | 148 | 14 | acid |
| SR3 | At3g16460 | XP_006406833 | jacalin-related lectin 34 | 5.5/68 | 5.31/72.5 | 40 | 4 | acid |
| XR4 | At3g16460 | XP_006406833 | jacalin-related lectin 34 | 5.6/67 | 5.31/72.5 | 240 | 23 | basic |
| Unknown | ||||||||
| SR18 | At1g48090 | XP_006393484 | uncharacterized LOC18010485 | 5.3/26 | 6.33/46.1 | 42 | 4 | appear |
| XR14 | At1g28680 | XP_006415631 | spermidine sinapoyl-CoA acyltransferase | 6.1/56 | 5.65/49.7 | 34 | 3 | appear |
| XR16 | At5g43060 | XP_006403303 | probable cysteine protease RD21B | 4.6/50 | 5.89/32.5 | 140 | 13 | equal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Sun, Z.; Zhu, M.; Zhang, Q.; Sun, Y.; Sun, W.; Wu, C.; Li, T.; Zhao, Y.; Ma, C.; et al. Dissecting the Molecular Regulation of Natural Variation in Growth and Senescence of Two Eutrema salsugineum Ecotypes. Int. J. Mol. Sci. 2022, 23, 6124. https://doi.org/10.3390/ijms23116124
Wang F, Sun Z, Zhu M, Zhang Q, Sun Y, Sun W, Wu C, Li T, Zhao Y, Ma C, et al. Dissecting the Molecular Regulation of Natural Variation in Growth and Senescence of Two Eutrema salsugineum Ecotypes. International Journal of Molecular Sciences. 2022; 23(11):6124. https://doi.org/10.3390/ijms23116124
Chicago/Turabian StyleWang, Fanhua, Zhibin Sun, Min Zhu, Qikun Zhang, Yufei Sun, Wei Sun, Chunxia Wu, Tongtong Li, Yiwu Zhao, Changle Ma, and et al. 2022. "Dissecting the Molecular Regulation of Natural Variation in Growth and Senescence of Two Eutrema salsugineum Ecotypes" International Journal of Molecular Sciences 23, no. 11: 6124. https://doi.org/10.3390/ijms23116124