
Diversity and Biological Activities of Endophytic Fungi from the Flowers of the Medicinal Plant Vernonia anthelmintica

 ,
, 
Abstract
:
1. Introduction
2. Results
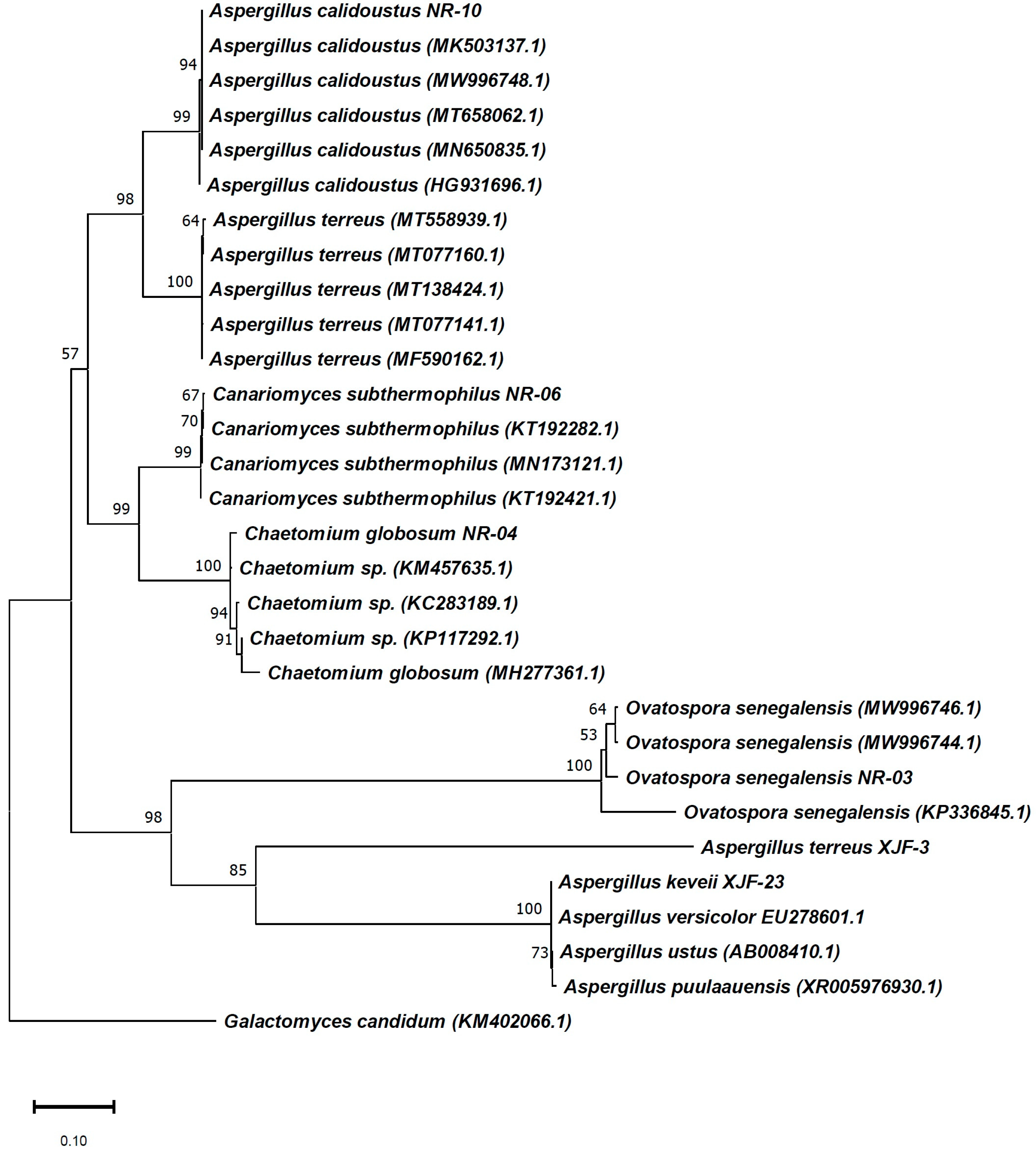
2.1. Isolation and Identification of Endophytic Fungi from the Flowers of V. anthelmintica
2.2. Antimicrobial Activity of the Crude Extracts of Endophytic Fungi
2.3. Determination of the MIC
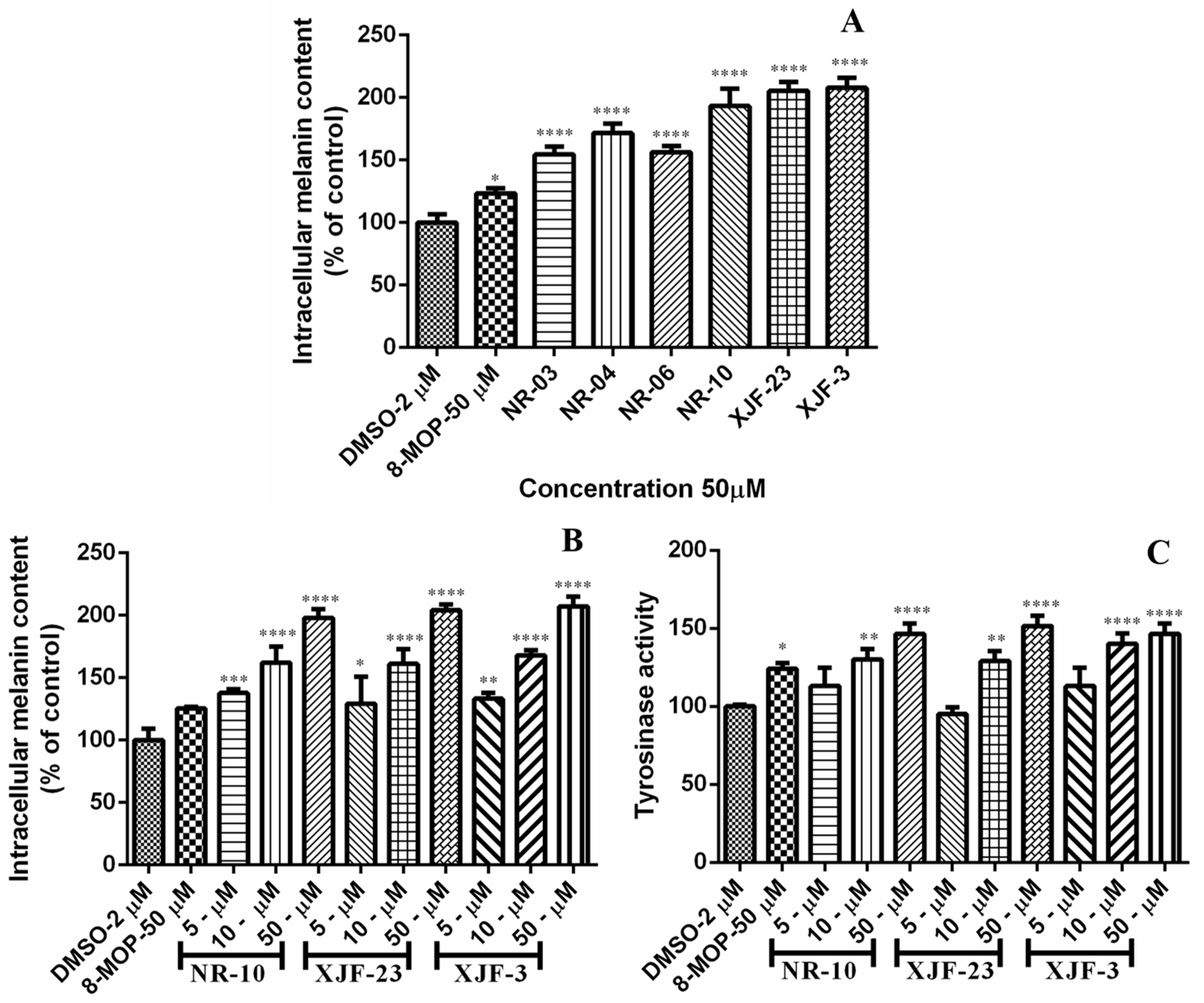
2.4. Effect of the Crude Extracts of Endophytic Fungi Isolated from V. anthelmintica Flowers on Melanin Content and Tyrosinase Activity in B16 Cells
2.5. Cytotoxic Activity of Fungal Endophytes
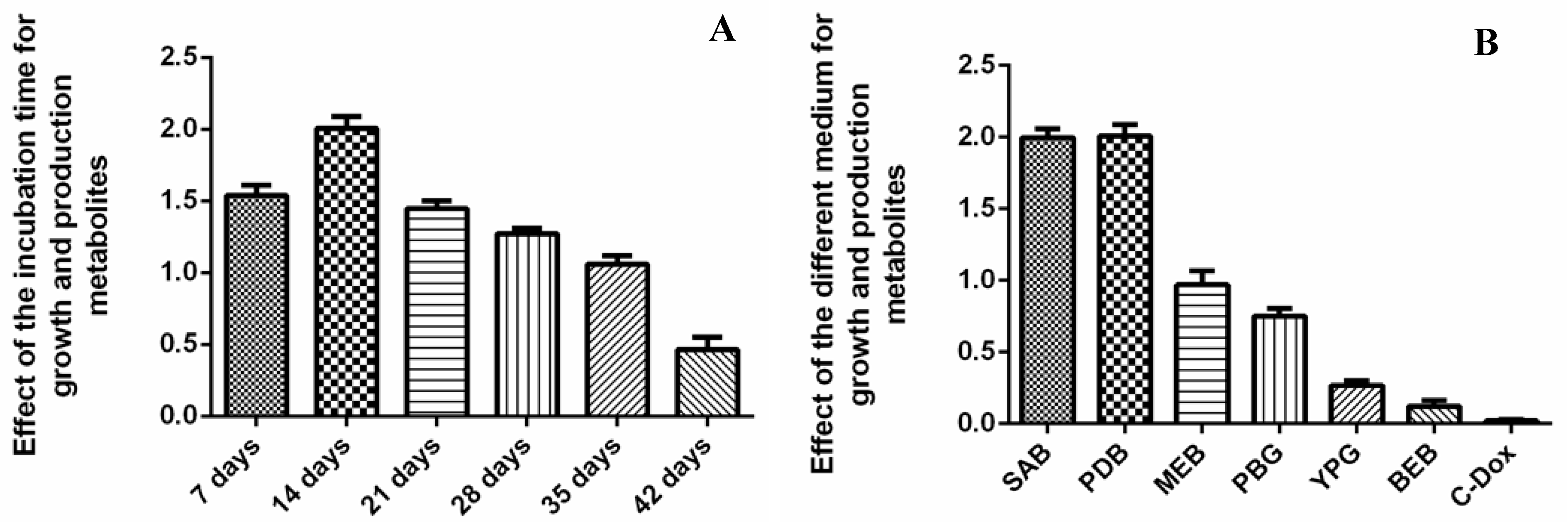
2.6. The Effects of Different Media and Incubation Times on the Growth and Production of Metabolites from Endophytic Fungi
2.7. Scanning Electron Microscopy (SEM) Analysis of O. senegalensis NR-03
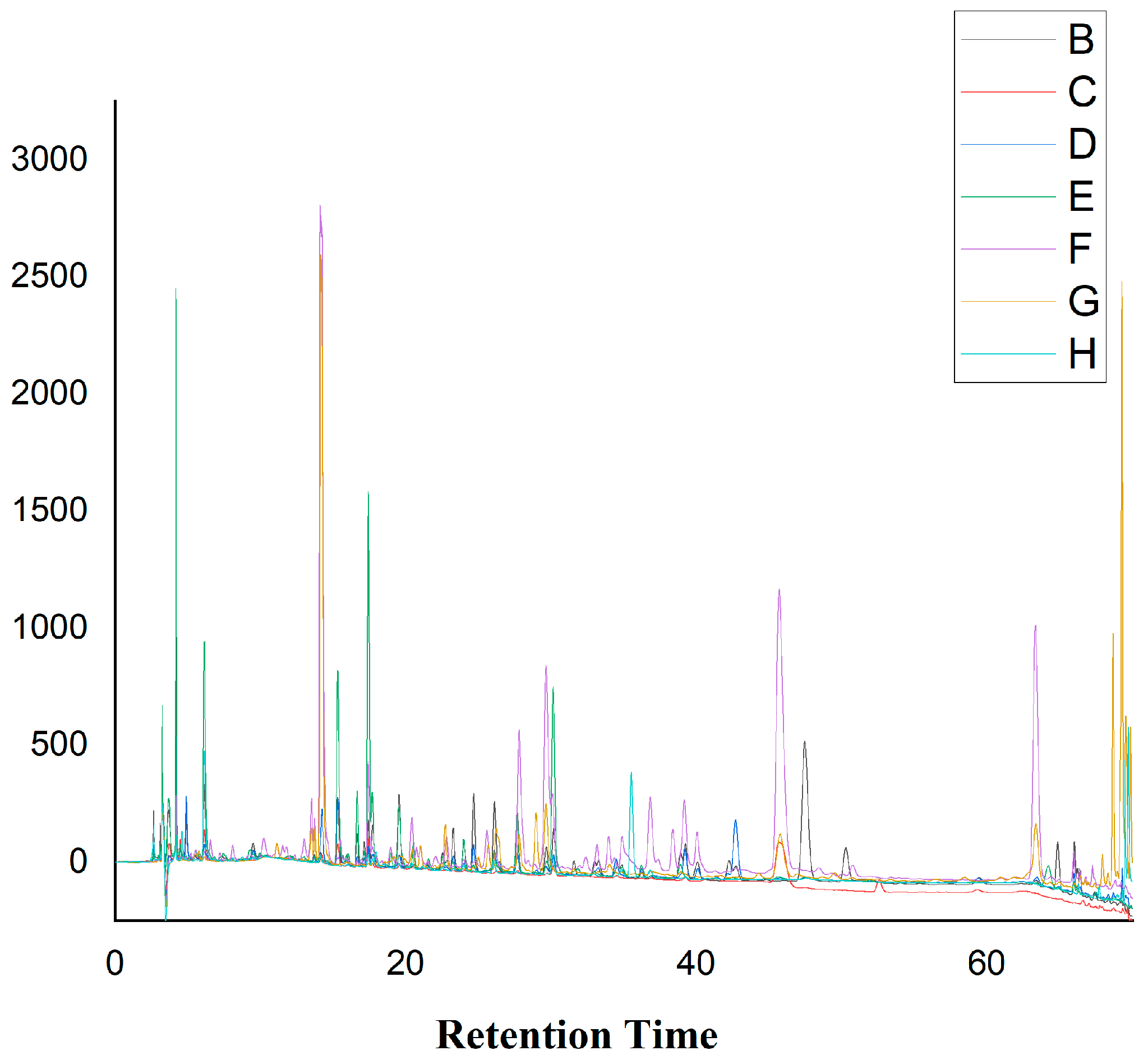
2.8. HPLC Analysis of the Crude Extracts from Different Media
2.9. Chemical Composition of the Endophyic Fungus O. senegalensis NR-03 Analyzed by GC-MS
3. Discussion
4. Materials and Methods
4.1. Plant Samples Collection
4.2. Isolation of Endophytic Fungi Producing Bioactive Secondary Metabolites
4.3. Identification of Endophytic Fungi Producing Bioactive Secondary Metabolites
4.4. Fermentation and Crude Extract Preparation
4.5. Antimicrobial Activity of Total Extracts of the Endophytic Fungi
4.6. Determination of MIC
4.7. Melanin Content Assay
4.8. Cytotoxic Activity (MTT Assay)
4.9. Optimization of the Optimal Culture Medium for the Production of Biologically Active Secondary Metabolites from Endophytic Fungi
4.10. The Effect of Cultivation Time on the Secondary Metabolite Production of Endophytic Fungi
4.11. Scanning Electron Microscopy Analysis of Endophytic Fungi
4.12. HPLC Analysis
4.13. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rajani, P.; Rajasekaran, C.; Vasanthakumari, M.M.; Olsson, S.B.; Ravikanth, G.; Uma Shaanker, R. Inhibition of plant pathogenic fungi by endophytic Trichoderma spp. through mycoparasitism and volatile organic compounds. Microbiol. Res. 2021, 242, 126595. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Raviraja, N.S. Fungal endophytes in five medicinal plant species from Kudremukh Range, Western Ghats of India. J. Basic Microbiol. 2005, 45, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Rustamova, N.; Bozorov, K.; Efferth, T.; Yili, A. Novel secondary metabolites from endophytic fungi: Synthesis and biological properties. Phytochem. Rev. 2020, 19, 425–448. [Google Scholar] [CrossRef]
- Dhayanithy, G.; Subban, K.; Chelliah, J. Diversity and biological activities of endophytic fungi associated with Catharanthus roseus. BMC Microbiol. 2019, 19, 22. [Google Scholar] [CrossRef]
- Zheng, R.; Li, S.; Zhang, X.; Zhao, C. Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study. Int. J. Mol. Sci. 2021, 22, 959. [Google Scholar] [CrossRef]
- Zhang, J.; Islam, M.S.; Wang, J.; Zhao, Y.; Dong, W. Isolation of Potato Endophytes and Screening of Chaetomium globosum Antimicrobial Genes. Int. J. Mol. Sci. 2022, 23, 4611. [Google Scholar] [CrossRef] [PubMed]
- Rustamova, N.; Litao, N.; Bozorov, K.; Sayyed, R.; Aisa, H.A.; Yili, A. Plant-associated endophytic fungi: A source of structurally diverse and bioactive natural products. Plant Cell Biotechnol. Mol. Biol. 2022, 23, 1–19. [Google Scholar]
- Santos, I.P.d.; Silva, L.C.N.d.; Silva, M.V.d.; Araújo, J.M.d.; Cavalcanti, M.d.S.; Lima, V.L.d.M. Antibacterial activity of endophytic fungi from leaves of Indigofera suffruticosa Miller (Fabaceae). Front. Microbiol. 2015, 6, 350. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.-K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites Paper presented at the British Mycological Society symposium on Fungal Bioactive Compounds, held at the University of Wales Swansea on 22–27 April 2001. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Ancheeva, E.; Daletos, G.; Proksch, P. Bioactive Secondary Metabolites from Endophytic Fungi. Curr. Med. Chem. 2020, 27, 1836–1854. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Medicinal Plants: A Source of Anti-Parasitic Secondary Metabolites. Molecules 2012, 17, 12771–12791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorrilla, J.G.; D’Addabbo, T.; Roscetto, E.; Varriale, C.; Catania, M.R.; Zonno, M.C.; Altomare, C.; Surico, G.; Nimis, P.L.; Evidente, A. Antibiotic and Nematocidal Metabolites from Two Lichen Species Collected on the Island of Lampedusa (Sicily). Int. J. Mol. Sci. 2022, 23, 8471. [Google Scholar] [CrossRef]
- Yasser, M.M.; Marzouk, M.A.; El-Shafey, N.M.; Shaban, S.A. Diversity and Antimicrobial Activity of Endophytic Fungi from the Medicinal Plant Pelargonium graveolens (geranium) in Middle Egypt. Jordan J. Biol. Sci. 2020, 13, 2. [Google Scholar]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Sederoff, H.; Chiang, V.L.; Borriss, R. Microbial Interactions Within Multiple-Strain Biological Control Agents Impact Soil-Borne Plant Disease. Front. Microbiol 2020, 11, 2452. [Google Scholar] [CrossRef]
- Shah, A.; Hassan, Q.P.; Mushtaq, S.; Shah, A.M.; Hussain, A. Chemoprofile and functional diversity of fungal and bacterial endophytes and role of ecofactors—A review. J. Basic Microbiol. 2017, 57, 814–826. [Google Scholar] [CrossRef]
- Shi, X.-S.; Meng, L.-H.; Li, X.; Wang, D.-J.; Zhou, X.-W.; Du, F.-Y.; Wang, B.-G.; Li, X.-M. Polyketides and Terpenoids with Potent Antibacterial Activities from the Artemisia argyi-Derived Fungus Trichoderma koningiopsis QA-3. Chem. Biodivers. 2020, 17, e2000566. [Google Scholar] [CrossRef]
- Duan, R.-T.; Yang, R.-N.; Li, H.-T.; Tang, L.-H.; Liu, T.; Yang, Y.-B.; Zhou, H.; Ding, Z.-T. Peniterester, a carotane-type antibacterial sesquiterpene from an artificial mutant Penicillium sp. T2-M20. Fitoterapia 2020, 140, 104422. [Google Scholar] [CrossRef]
- Xu, K.; Li, G.; Zhu, R.; Xie, F.; Li, Y.; Yang, W.; Xu, L.; Shen, T.; Zhao, Z.; Lou, H. Polyketides from the endolichenic fungus Eupenicillium javanicum and their anti-inflammatory activities. Phytochemistry 2020, 170, 112191. [Google Scholar] [CrossRef]
- Cheng, T.; Chepkirui, C.; Decock, C.; Matasyoh, J.C.; Stadler, M. Heimiomycins A–C and Calamenens from the African Basidiomycete Heimiomyces sp. J. Nat. Prod. 2020, 83, 2501–2507. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Ding, G.; Wu, G.; Yang, J.; Zhang, M.; Wang, H.; Wei, D.; Qin, J.; Guo, L. Identification of a Unique Azaphilone Produced by Chaetomium globosum Isolated from Polygonatum sibiricum. Chem. Biodivers. 2020, 17, e1900744. [Google Scholar] [CrossRef] [PubMed]
- Abdel Razek, M.M.M.; Moussa, A.Y.; El-Shanawany, M.A.; Singab, A.N.B. A New Phenolic Alkaloid from Halocnemum strobilaceum Endophytes: Antimicrobial, Antioxidant and Biofilm Inhibitory Activities. Chem. Biodivers. 2020, 17, e2000496. [Google Scholar] [CrossRef] [PubMed]
- Akshatha, V.J.; Nalini, M.S.; D’Souza, C.; Prakash, H.S. Streptomycete endophytes from anti-diabetic medicinal plants of the Western Ghats inhibit alpha-amylase and promote glucose uptake. Lett. Appl. Microbiol. 2014, 58, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Biasetto, C.R.; Somensi, A.; Sordi, R.; Chapla, V.M.; Ebrahimi, S.N.; Silva, G.H.; Teles, H.L.; Bolzani, V.d.S.; Young, M.C.M.; Pfenning, L.H.; et al. The new koninginins T-U from Phomopsis stipata, an endophytic fungus isolated from Styrax camporum Pohl. Phytochem. Lett. 2020, 36, 106–110. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.M.; Asfour, H.Z. Antimicrobial metabolites from the endophytic fungus Aspergillus versicolor. Phytochem. Lett. 2020, 35, 152–155. [Google Scholar] [CrossRef]
- Li, H.; Zhang, R.; Cao, F.; Wang, J.; Hu, Z.; Zhang, Y. Proversilins A–E, Drimane-Type Sesquiterpenoids from the Endophytic Aspergillus versicolor. J. Nat. Prod. 2020, 83, 2200–2206. [Google Scholar] [CrossRef]
- Li, W.-Y.; Liu, Y.; Lin, Y.-T.; Liu, Y.-C.; Guo, K.; Li, X.-N.; Luo, S.-H.; Li, S.-H. Antibacterial harziane diterpenoids from a fungal symbiont Trichoderma atroviride isolated from Colquhounia coccinea var. mollis. Phytochemistry 2020, 170, 112198. [Google Scholar] [CrossRef]
- He, J.; Pu, C.-J.; Wang, M.; Li, Z.-H.; Feng, T.; Zhao, D.-K.; Liu, J.-K. Conosiligins A–D, Ring-Rearranged Tremulane Sesquiterpenoids from Conocybe siliginea. J. Nat. Prod. 2020, 83, 2743–2748. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, Y.; Chen, S.; Ji, Y.; Ding, W.; Ma, Z. Strepolyketides A-C, three novel SEK15-derived polyketides from Streptomyces sp. HN2A53. Tetrahedron Lett. 2020, 61, 151996. [Google Scholar] [CrossRef]
- Li, W.; Shao, Y.-T.; Yin, T.-P.; Yan, H.; Shen, B.-C.; Li, Y.-Y.; Xie, H.-D.; Sun, Z.-W.; Ma, Y.-L. Penisarins A and B, Sesquiterpene Coumarins Isolated from an Endophytic Penicillium sp. J. Nat. Prod. 2020, 83, 3471–3475. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Jiao, F.W.; Li, W.; Zhang, X.; Yan, W.; Jiao, R.H. Cytotoxic Xanthone Derivatives from the Mangrove-Derived Endophytic Fungus Peniophora incarnata Z4. J. Nat. Prod. 2020, 83, 2976–2982. [Google Scholar] [CrossRef] [PubMed]
- Turak, A.; Liu, Y.; Aisa, H.A. Elemanolide dimers from seeds of Vernonia anthelmintica. Fitoterapia 2015, 104, 23–30. [Google Scholar] [CrossRef]
- Turak, A.; Maimaiti, Z.; Ma, H.; Aisa, H.A. Pseudo-disesquiterpenoids from seeds of Vernonia anthelmintica and their biological activities. Phytochem. Lett. 2017, 21, 163–168. [Google Scholar] [CrossRef]
- Rustamova, N.; Wubulikasimu, A.; Mukhamedov, N.; Gao, Y.; Egamberdieva, D.; Yili, A. Endophytic Bacteria Associated with Medicinal Plant Vernonia anthelmintica: Diversity and Characterization. Curr. Microbiol. 2020, 77, 1457–1465. [Google Scholar] [CrossRef]
- Rustamova, N.; Bobakulov, K.; Begmatov, N.; Turak, A.; Yili, A.; Aisa, H.A. Secondary metabolites produced by endophytic Pantoea ananatis derived from roots of Baccharoides anthelmintica and their effect on melanin synthesis in murine B16 cells. Nat. Prod. Res. 2021, 35, 796–801. [Google Scholar] [CrossRef]
- Rustamova, N.; Gao, Y.; Zhang, Y.; Yili, A. Biological Activity of Endophytic Fungi from the Roots of the Medicinal Plant Vernonia anthelmintica. Microorganisms 2020, 8, 586. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Jiao, F.-W.; Wang, J.-Q.; Shi, J.; Wang, T.-T.; Khan, S.; Jiao, R.-H.; Tan, R.-X.; Ge, H.-M. Unguisin G, a new kynurenine-containing cyclic heptapeptide from the sponge-associated fungus Aspergillus candidus NF2412. Tetrahedron Lett. 2020, 61, 152322. [Google Scholar] [CrossRef]
- Bunbamrung, N.; Intaraudom, C.; Dramae, A.; Komwijit, S.; Laorob, T.; Khamsaeng, S.; Pittayakhajonwut, P. Antimicrobial, antimalarial and anticholinesterase substances from the marine-derived fungus Aspergillus terreus BCC51799. Tetrahedron 2020, 76, 131496. [Google Scholar] [CrossRef]
- Turak, A.; Aisa, H.A. Oxygen heterocyclic Diels-Alder-Type sesquiterpenoid dimers from Vernonia anthelmintica. Phytochemistry 2022, 203, 113386. [Google Scholar] [CrossRef]
- Maimaiti, Z.; Turak, A.; Ma, Q.L.; Liu, G.; Aisa, H.A. Quantitative Determination of Marker Compounds and Fingerprint Analysis of the Seeds of Vernonia anthelmintica. Int. J. Anal. Chem. 2020, 2020, 8859425. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Y.; Li, S.; Zhang, W.; Liu, Z.; Tan, H.; Zhang, W. Trichothecene macrolides from the endophytic fungus Paramyrothecium roridum and their cytotoxic activity. Fitoterapia 2020, 147, 104768. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.-J.; Li, Y.-s.; Xia, B.; Li, W.-p.; Pang, L.; Tong, Z. Optimization of medium composition and culture conditions for antifungal activity of a tomato endophytic bacterium. Biol. Control 2015, 82, 69–75. [Google Scholar] [CrossRef]
- Kumar, V.; Sahai, V.; Bisaria, V.S. High-density spore production of Piriformospora indica, a plant growth-promoting endophyte, by optimization of nutritional and cultural parameters. Bioresour. Technol. 2011, 102, 3169–3175. [Google Scholar] [CrossRef]
- He, J.; Zou, J.; Shao, Z.; Zhang, J.; Liu, Z.; Yu, Z. Characteristics and flocculating mechanism of a novel bioflocculant HBF-3 produced by deep-sea bacterium mutant Halomonas sp. V3a’. World J. Microbiol. Biotechnol. 2010, 26, 1135–1141. [Google Scholar] [CrossRef]
- Ezhilan, B.P.; Neelamegam, R. GC-MS analysis of phytocomponents in the ethanol extract of Polygonum chinense L. Pharmacogn. Res. 2012, 4, 11–14. [Google Scholar]
- Gomathi, D.; Kalaiselvi, M.; Ravikumar, G.; Devaki, K.; Uma, C. GC-MS analysis of bioactive compounds from the whole plant ethanolic extract of Evolvulus alsinoides (L.) L. J. Food Sci. Technol. 2015, 52, 1212–1217. [Google Scholar] [CrossRef] [Green Version]
- Maimaiti, Z.; Turak, A.; Aisa, H.A. Two new compounds from the seeds of Vernonia anthelmintica. J. Asian Nat. Prod. Res. 2017, 19, 862–868. [Google Scholar] [CrossRef]
- Zhou, G.; Chen, X.; Zhang, X.; Che, Q.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Prenylated p-Terphenyls from a Mangrove Endophytic Fungus, Aspergillus candidus LDJ-5. J. Nat. Prod. 2020, 83, 8–13. [Google Scholar] [CrossRef]
- Chi, L.P.; Yang, S.Q.; Li, X.M.; Li, X.D.; Wang, B.G.; Li, X. A new steroid with 7β,8β-epoxidation from the deep sea-derived fungus Aspergillus penicillioides SD-311. J. Asian Nat. Prod. Res. 2021, 23, 884–891. [Google Scholar] [CrossRef]
- Yang, Z.; Kaliaperumal, K.; Zhang, J.; Liang, Y.; Guo, C.; Zhang, J.; Yang, B.; Liu, Y. Antifungal fatty acid derivatives against Penicillium italicum from the deep-sea fungus Aspergillus terreus SCSIO 41202. Nat. Prod. Res. 2021, 35, 4394–4401. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Liu, Y.; Huang, Y.; Yin, X.; Zhou, Y.; Duan, Y.; Xie, S.; Guo, Y.; Qiao, Y.; Shi, Z.; et al. New bioactive secondary metabolites from the Anoectochilus roxburghii endophytic fungus Aspergillus versicolor. Fitoterapia 2020, 143, 104532. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, Q.; Sun, W.; Zhang, R.; Wang, J.; Lai, Y.; Hu, Z.; Zhang, Y. 21-Epi-taichunamide D and (±)-versicaline A, three unusual alkaloids from the endophytic Aspergillus versicolor F210. Tetrahedron Lett. 2020, 61, 152219. [Google Scholar] [CrossRef]
- Hashem, A.H.; Shehabeldine, A.M.; Abdelaziz, A.M.; Amin, B.H.; Sharaf, M.H. Antifungal Activity of Endophytic Aspergillus terreus Extract Against Some Fungi Causing Mucormycosis: Ultrastructural Study. Appl. Biochem. Biotechnol. 2022, 194, 3468–3482. [Google Scholar] [CrossRef]
- Sharaf, M.H.; Abdelaziz, A.M.; Kalaba, M.H.; Radwan, A.A.; Hashem, A.H. Antimicrobial, Antioxidant, Cytotoxic Activities and Phytochemical Analysis of Fungal Endophytes Isolated from Ocimum Basilicum. Appl. Biochem. Biotechnol. 2022, 194, 1271–1289. [Google Scholar] [CrossRef]
- Tantapakul, C.; Promgool, T.; Kanokmedhakul, K.; Soytong, K.; Song, J.; Hadsadee, S.; Jungsuttiwong, S.; Kanokmedhakul, S. Bioactive xanthoquinodins and epipolythiodioxopiperazines from Chaetomium globosum 7s-1, an endophytic fungus isolated from Rhapis cochinchinensis (Lour.) Mart. Nat. Prod. Res. 2020, 34, 494–502. [Google Scholar] [CrossRef]
- Wang, W.-X.; Zheng, M.-J.; Li, J.; Feng, T.; Li, Z.-H.; Huang, R.; Zheng, Y.-S.; Sun, H.; Ai, H.-L.; Liu, J.-K. Cytotoxic polyketides from endophytic fungus Phoma bellidis harbored in Ttricyrtis maculate. Phytochem. Lett. 2019, 29, 41–46. [Google Scholar] [CrossRef]
- Pan, Y.; Jin, H.; Yang, S.; Liu, H. Changes of volatile organic compounds and bioactivity of Alternaria brassicae GL07 in different ages. J. Basic Microbiol. 2019, 59, 713–722. [Google Scholar] [CrossRef]
- Nei, M.; Saitou, N. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nischitha, R.; Shivanna, M.B. Antimicrobial activity and metabolite profiling of endophytic fungi in Digitaria bicornis (Lam) Roem. and Schult. and Paspalidium flavidum (Retz.) A. Camus. 3 Biotech 2021, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.; Chang, C.-F.; Chi, W.-C. Isolation of endophytic fungi with antimicrobial activity from medicinal plant Zanthoxylum simulans Hance. Folia Microbiol. 2021, 66, 385–397. [Google Scholar] [CrossRef]
- Rasulov, B.; Rustamova, N.; Yili, A.; Zhao, H.-Q.; Aisa, H.A. Synthesis of silver nanoparticles on the basis of low and high molar mass exopolysaccharides of Bradyrhizobium japonicum 36 and its antimicrobial activity against some pathogens. Folia Microbiol. 2016, 61, 283–293. [Google Scholar] [CrossRef]
- Omara, S.T. MIC and MBC of Honey and Gold Nanoparticles against methicillin-resistant (MRSA) and vancomycin-resistant (VRSA) coagulase-positive S. aureus isolated from contagious bovine clinical mastitis. J. Genet. Eng. Biotechnol. 2017, 15, 219–230. [Google Scholar] [CrossRef]
- Saboora, A.; Sajjadi, S.-T.; Mohammadi, P.; Fallahi, Z. Antibacterial activity of different composition of aglycone and glycosidic saponins from tuber of Cyclamen coum Miller. Ind. Crops Prod. 2019, 140, 111662. [Google Scholar] [CrossRef]
- Niu, C.; Yin, L.; Aisa, H.A. Novel Furocoumarin Derivatives Stimulate Melanogenesis in B16 Melanoma Cells by Up-Regulation of MITF and TYR Family via Akt/GSK3β/β-Catenin Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 746. [Google Scholar] [CrossRef] [Green Version]
- Fei Nie, L.; Huang, G.; Bozorov, K.; Zhao, J.; Niu, C.; Sagdullaev, S.S.; Aisa, H.A. Corrigendum to: Diversity-oriented synthesis of amide derivatives of tricyclic thieno[2,3-d] pyrimidin-4(3H)-ones and evaluation of their influence on melanin synthesis in murine B16 cells. Heterocycl. Commun. 2018, 24, 241. [Google Scholar] [CrossRef]
- Li, H.; Qing, C.; Zhang, Y.; Zhao, Z. Screening for Endophytic Fungi with Antitumour and Antifungal Activities from Chinese Medicinal Plants. World J. Microbiol. Biotechnol. 2005, 21, 1515–1519. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, C.; Chen, H.; Zhang, J.; Chen, W. Isolation and Identification of Endophytic Fungi from Actinidia macrosperma and Investigation of Their Bioactivities. J. Evid. Based. Complementary Altern. Med. 2012, 2012, 382742. [Google Scholar]
- Mukhamedov, N.; Wubulikasimu, A.; Rustamova, N.; Nuerxiati, R.; Mirzaakhmedov, S.; Ishimov, U.; Ziyavitdinov, J.; Yili, A.; Aisa, H.A. Synthesis and Characterization of Novel Chickpea Protein Hydrolysate-Vanadium Complexes Having Cell Inhibitory Effects on Lung Cancer A549 Cells Lines. Protein J. 2021, 40, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Chen, X.; Shi, Z.; Xie, S.; Qiao, Y.; Chen, G.; Tan, X.; Lu, Y.; Qi, C.; Zhang, Y. New immunosuppressive secondary metabolites from the endophytic fungus Aspergillus sp. Fitoterapia 2021, 151, 104882. [Google Scholar] [CrossRef] [PubMed]
- Nishanbaev, S.; Bobakulov, K.; Okhundedaev, B.; Sasmakov, S.; Yusupova, E.; Azimova, S.; Abdullaev, N. Component composition of the extracts and essential oils from the Alhagi canescens, growing in Uzbekistan and their antimicrobial activity. Nat. Prod. Res. 2019, 33, 3417–3420. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolated Strains Sequences Deposited in GenBank | Closest Match among Bacteria (18S rRNA Genes) (GenBank) | |||
|---|---|---|---|---|
| Strains | Accession Number | Species | Source | ID% |
| NR-03 | MW996742 | O. senegalensis | flower | 100 |
| NR-04 | MW996743 | Ch. globosum | flower | 99.04 |
| NR-06 | MW996745 | T. subthermophila | flower | 100 |
| NR-10 | MW996749 | A. calidoustus | flower | 100 |
| XJF-23 | MW881461 | A. keveii | flower | 99.82 |
| XJF-3 | MW881454 | A. terreus | flower | 99.94 |
| Sample | Sample Concentration | Sample Amount (μL) | C. albicans (mm ZOI) | E. coli (mm ZOI) | S. aureus (mm ZOI) |
|---|---|---|---|---|---|
| Ampicillin | 312.5 μg/mL | 20 | - | 11 | - |
| Ampicillin | 4.44 μg/mL | 20 | - | - | 18.5 |
| Amphotericin B | 5 μg/mL | 20 | 12 | - | - |
| O. senegalensis NR-03 | 50 μg/mL | 20 | 7.5 | 26 | 19 |
| Ch. Globosum NR-04 | 50 μg/mL | 20 | 12 | - | 20 |
| T. Subthermophila NR-06 | 50 μg/mL | 20 | 9.5 | 9 | 11 |
| A. calidoustus NR-10 | 50 μg/mL | 20 | 10 | 13 | 15 |
| A. keveii XJF-23 | 50 μg/mL | 20 | 8 | 10 | 15 |
| A. terreus XJF-3 | 50 μg/mL | 20 | 10 | 11 | 16 |
| Sample | C. albicans | E. coli | S. aureus |
|---|---|---|---|
| MIC μg/mL | MIC μg/mL | MIC μg/mL | |
| O. senegalensis NR-03 | ND | 2000 | 62.5 |
| Ch. Globosum NR-04 | 31.25 | ND | 125 |
| T. Subthermophila NR-06 | ND | ND | ND |
| A. calidoustus NR-10 | ND | 125 | ND |
| A. keveii XJF-23 | ND | 250 | ND |
| A. terreus XJF-3 | 250 | 62.5 | 250 |
| Ampicillin 100 mg/mL | - | 0.98 | 62.5 |
| Amphotericin B 50 mg/mL | 0.98 | - | - |
| Sample | Concentration (µM) | Intracellular Melanin Content (% of Control) |
|---|---|---|
| Control | 2µL-DMSO | 100.00 |
| 8-MOP | 50μM | 123.34 |
| O. senegalensis NR-03 | 50μg/mL | 154.49 |
| Ch. globosum NR-04 | 50μg/mL | 171.58 |
| T. subthermophila NR-06 | 50μg/mL | 156.27 |
| A. calidoustus NR-10 | 50μg/mL | 193.31 |
| A. keveii XJF-23 | 50μg/mL | 205.35 |
| A. terreus XJF-3 | 50μg/mL | 208.46 |
| Sample | Concentration | Intracellular Melanin Content (% of Control) | Tyrosinase Activity |
|---|---|---|---|
| Control | DMSO | 100.00 | 100.00 |
| 8-MOP | 50 µM | 125.52 | 124.8 |
| A. calidoustus NR-10 | 5 µM | 137.91 | 113.12 |
| 10 µM | 162.17 | 130.02 | |
| 50 µM | 197.98 | 146.63 | |
| A. keveii XJF-23 | 5 µM | 96.57 | 95.05 |
| 10 µM | 161.18 | 129.02 | |
| 50 µM | 204.02 | 151.63 | |
| A. terreus XJF-3 | 5 µM | 133.24 | 114.06 |
| 10 µM | 167.85 | 140.06 | |
| 50 µM | 207.12 | 146.63 |
| Samples | Cell Lines | ||
|---|---|---|---|
| IC50 (μg/mL) | |||
| HT-29 | MCF-7 | HeLa | |
| O. senegalensis NR-03 | 0.10 μg/mL ± 0.004 | Not active | 0.09 μg/mL ± 0.005 |
| Ch. Globosum NR-04 | 32.41 μg/mL ± 2.20 | Not active | 29.38 μg/mL ± 1.27 |
| T. Subthermophila NR-06 | 3.85 μg/mL ± 0.15 | 9.99 μg/mL ± 0.69 | 5.89 μg/mL ± 0.35 |
| A. calidoustus NR-10 | 16.44 μg/mL ± 0.85 | 19.55 μg/mL ± 1.03 | 13.59 μg/mL ± 0.63 |
| A. keveii XJF-23 | 31.37 μg/mL ± 1.82 | Not active | 17.81 μg/mL ± 0.78 |
| A. terreus XJF-3 | Not active | Not active | 0.10 μg/mL ± 0.005 |
| DOX | 0.82 μg/mL ± 0.041 | 0.17 μg/mL ± 0.006 | 0.11 μg/mL ± 0.005 |
| № | Composition | tR (min) | Relative Peak Area % | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BEB | SAB | YPD | MEB | PBG | CDM | PDB | |||
| 1 | 3-Methyl-butanoic acid | 3.418 | - | - | - | 1.40 | - | - | - |
| 2 | 2-Methyl-butanoic acid | 3.520 | - | - | - | 0.48 | - | - | 0.78 |
| 3 | p-Xylene | 3.741 | - | - | - | - | - | 1.71 | - |
| 4 | 3-methyl-1,2-cyclopentanedione | 5.848 | - | - | - | - | - | - | 1.41 |
| 5 | 2-Methyl-phenol | 6.247 | - | 0.21 | - | 0.12 | - | - | 0.22 |
| 6 | p-Cresol | 6.553 | - | 1.96 | - | 0.28 | - | - | - |
| 7 | Tetramethyl-pyrazine | 6.714 | - | - | 0.26 | - | - | - | - |
| 8 | Maltol | 7.105 | - | - | - | 0.57 | - | - | - |
| 9 | Phenylethyl alcohol | 7.139 | - | - | 0.13 | - | - | - | - |
| 10 | N-(3-Methylbutyl)acetamide | 7.402 | - | - | 0.16 | - | - | - | - |
| 11 | Benzeneacetic acid | 9.136 | - | 3.02 | - | - | - | - | 1.00 |
| 12 | Indole | 9.756 | - | - | 0.56 | - | - | - | - |
| 13 | Orcinol | 10.801 | - | 1.66 | - | 1.11 | - | - | - |
| 14 | 4-Hydroxy-2-methylbenzaldehyde | 11.157 | - | - | - | - | - | - | 1.11 |
| 15 | 3,4-Bis(methylene)-cyclopentanone | 11.480 | - | - | 0.36 | - | - | - | - |
| 16 | 4-Hydroxy-benzeneethanol | 11.489 | - | - | - | 0.99 | - | - | - |
| 17 | 3-Ethyl-2,5-dimethyl-pyrazine | 11.795 | - | - | 0.53 | - | - | - | - |
| 18 | Terrein | 11.948 | - | - | 0.92 | - | - | - | 33.71 |
| 19 | 3,5-Diethyl-2-methyl-pyrazine | 12.474 | - | - | 0.30 | - | - | - | - |
| 20 | Dimethyl(3-methylphenoxy)ethoxy-silane | 12.525 | - | - | - | - | - | - | 1.09 |
| 21 | N-(2-Phenylethyl)-acetamide | 12.542 | - | - | 0.76 | - | - | - | - |
| 22 | Thymine | 13.587 | - | - | 1.13 | - | - | - | - |
| 23 | 3-Mercaptobenzoic acid, S-methyl-, methyl ester | 14.258 | - | - | - | 0.44 | - | - | - |
| 24 | Tetradecanoic acid | 15.278 | - | - | - | - | - | - | 0.22 |
| 25 | Hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione | 15.337 | - | - | 0.41 | - | - | - | - |
| 26 | Pentadecanoic acid | 16.008 | - | 0.17 | - | - | - | - | 1.10 |
| 27 | N,N-Dimethyl-1H-purin-6-amine | 16.153 | - | - | 0.30 | - | - | - | - |
| 28 | N-Acetyltyramine | 16.280 | - | - | 2.21 | - | - | - | - |
| 29 | Hexadecanoic acid, methyl ester | 17.223 | - | - | - | - | - | - | 1.94 |
| 30 | Hexahydro-3-(2-methylpropyl)-pyrrolo[1,2-a]pyrazine-1,4-dione | 17.393 | 36.59 | - | - | - | - | 7.90 | - |
| 31 | 3,4-dihydro-6,8-dihydroxy-3-methyl-1H-2-benzopyran-1-one | 17.546 | - | - | - | 0.44 | - | - | - |
| 32 | n-Hexadecanoic acid | 17.656 | - | - | - | - | - | - | 0.60 |
| 33 | Eicosamethyl-cyclodecasiloxane, | 17.903 | - | - | - | - | - | - | 0.31 |
| 34 | Hexadecanoic acid, ethyl ester | 18.081 | - | - | - | - | - | - | 0.34 |
| 35 | 9,12-Octadecadienoic acid (Z,Z)-, methyl ester | 19.449 | - | - | - | - | - | - | 6.39 |
| 36 | (E)-9-Octadecenoic acid, methyl ester | 19.525 | - | - | - | - | - | - | 6.15 |
| 37 | Methyl stearate | 19.848 | - | - | - | - | - | - | 0.87 |
| 38 | Linoleic acid ethyl ester | 20.358 | - | - | - | - | - | - | 0.78 |
| 39 | (E)-9-Octadecenoic acid ethyl ester | 20.434 | - | - | - | - | - | - | 0.62 |
| 40 | N-[2-(1H-indol-3-yl)ethyl]-acetamide | 20.859 | - | - | 0.76 | - | - | - | - |
| 41 | 3-Benzyl-6-isopropyl-2,5-piperazinedione | 21.845 | - | - | 0.22 | - | - | - | - |
| 42 | Octadecamethyl-cyclononasiloxane | 21.989 | - | - | - | - | - | - | 0.36 |
| 43 | Hexadecane | 22.244 | - | - | - | - | - | 3.05 | |
| 44 | Hexahydro-3-(phenylmethyl)-pyrrolo[1,2-a]pyrazine-1,4-dione | 22.915 | 14.44 | 1.47 | 21.64 | - | 14.47 | 18.13 | 0.56 |
| 45 | Tetracosane | 23.671 | - | - | - | - | - | 4.89 | - |
| 46 | Eicosane | 25.098 | - | - | - | - | - | 5.60 | - |
| 47 | Rosenonolactone | 26.152 | - | - | - | - | - | - | 2.11 |
| 48 | (5.beta.)-chol-7-ene | 27.129 | - | - | - | 0.81 | - | - | - |
| 49 | 1,8-dihydroxy-3-methoxy-6-methyl-9,10 anthracenedione | 27.256 | - | - | - | 0.22 | - | - | - |
| 50 | 3,4-dihydro-3-hydroxy-2,2-dimethyl-, (R)-2H-naphtho[1,2-b]pyran-5,6-dione | 31.087 | - | - | - | - | 0.83 | - | - |
| 51 | Simvastatin | 31.391 | - | - | - | 14.38 | 15.68 | - | 1.60 |
| 52 | Lovastatin | 34.307 | - | - | - | - | - | - | 0.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, L.; Rustamova, N.; Ning, H.; Paerhati, P.; Lu, C.; Yili, A. Diversity and Biological Activities of Endophytic Fungi from the Flowers of the Medicinal Plant Vernonia anthelmintica. Int. J. Mol. Sci. 2022, 23, 11935. https://doi.org/10.3390/ijms231911935
Niu L, Rustamova N, Ning H, Paerhati P, Lu C, Yili A. Diversity and Biological Activities of Endophytic Fungi from the Flowers of the Medicinal Plant Vernonia anthelmintica. International Journal of Molecular Sciences. 2022; 23(19):11935. https://doi.org/10.3390/ijms231911935
Chicago/Turabian StyleNiu, Litao, Nigora Rustamova, Huxia Ning, Paiziliya Paerhati, Chunfang Lu, and Abulimiti Yili. 2022. "Diversity and Biological Activities of Endophytic Fungi from the Flowers of the Medicinal Plant Vernonia anthelmintica" International Journal of Molecular Sciences 23, no. 19: 11935. https://doi.org/10.3390/ijms231911935