
Fungal Effectoromics: A World in Constant Evolution
by
 , ,
, ,
Jewel Nicole Anna Todd
1,
Karla Gisel Carreón-Anguiano
1,
Ignacio Islas-Flores
2 and
Blondy Canto-Canché
1,* 1
Unidad de Biotecnología, Centro de Investigación Científica de Yucatán, A.C., Calle 43 No. 130 x 32 y 34, Colonia Chuburná de Hidalgo, Mérida C.P. 97205, Yucatán, Mexico
2
Unidad de Bioquímica y Biología Molecular de Plantas, Centro de Investigación Científica de Yucatán, A.C., Calle 43 No. 130 x 32 y 34, Colonia Chuburná de Hidalgo, Mérida C.P. 97205, Yucatán, Mexico
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(21), 13433; https://doi.org/10.3390/ijms232113433
Submission received: 28 September 2022
/
Revised: 25 October 2022
/
Accepted: 31 October 2022
/
Published: 3 November 2022
(This article belongs to the Special Issue Molecular Plant-Microbe Interactions 2.0)
Abstract
:Effectors are small, secreted molecules that mediate the establishment of interactions in nature. While some concepts of effector biology have stood the test of time, this area of study is ever-evolving as new effectors and associated characteristics are being revealed. In the present review, the different characteristics that underly effector classifications are discussed, contrasting past and present knowledge regarding these molecules to foster a more comprehensive understanding of effectors for the reader. Research gaps in effector identification and perspectives for effector application in plant disease management are also presented, with a focus on fungal effectors in the plant-microbe interaction and interactions beyond the plant host. In summary, the review provides an amenable yet thorough introduction to fungal effector biology, presenting noteworthy examples of effectors and effector studies that have shaped our present understanding of the field.
1. Introduction
Effectors are molecules that provide an evolutive advantage to organisms as they complete their life cycles and thrive in their respective niches. Effectors are traditionally defined as molecules that manipulate host cell structure and function, thereby facilitating infection (virulence factors or toxins) and/or triggering defense responses (avirulence factors or elicitors) [1]. We define effectors as molecules of diverse functionality and molecular nature that influence organisms’ interactions with each other, usually to the benefit of the organism using them. The effector gene products are produced by one organism but they mainly function in another, resulting in the alteration of host cell structure and function [1,2,3]. Effectors were first discovered in the pathogen-plant interaction between a biotrophic fungus and its plant host but have since been widely described in different pathosystems involving biotrophic, hemibiotrophic, and necrotrophic fungi [4,5,6]. Biotrophic organisms require live hosts to complete their life cycle, and their effectors allow them to stealthily enter and remain in the host while avoiding recognition and suppressing the host’s defenses [7,8,9]. Necrotrophs, which are heavily dependent on cell wall-degrading enzymes and phytotoxins to infect hosts also use effector molecules [10,11,12].
In each ecological interaction, communication exists between effectors and receptors that shape the outcome of the interaction. Although widely associated with pathogenicity, effectors have been discovered in plant-beneficial organisms such as mutualist mycorrhizal species and endophytes [13,14,15,16] and the study of their roles in microbe-microbe interactions is gaining traction in the field of effector biology [17,18]. These molecules are ubiquitous in microorganisms, and the effectorome complement of each organism is highly specialized and greatly affects its lifestyle, as both gene gain and loss of certain effectors have implications for pathogen virulence and host adaptation [19,20,21,22]. Effectors have even shed their “microbial” association and have been identified in larger organisms [23,24,25,26] and are also defense molecules used by plants in their interactions with invaders [27,28]. It is now more evident than ever before that effectors participate in diverse associations between organisms that transcend the boundaries of taxonomic kingdoms and lifestyles, making them cornerstones of biological interactions.
Effectors can be classified in many ways: according to their molecular nature, effector molecules can be proteins [29,30], secondary metabolites [31,32,33], or small RNAs [34,35]. Apart from their molecular nature, effectors can be classified based on their site of action in the host plant i.e., apoplastic (extracellular) or intracellular; the latter comprising effectors targeting host proteins in the cytoplasm and cell organelles [1,36,37,38]. Some effectors may also be race-specific i.e., only found in some strains or isolates of a species [39,40] while other effectors are common to the genomes of different closely or distantly related species and are termed “core effectors” [41,42]. Another type of classification for effectors is based on their interactions with organisms or host specificity [43] since effectors participate in more than one type of interaction, e.g., in both plant-microbe and microbe-microbe interactions, while other effectors are interaction-specific.
Since the hypothesis of pathogen Avr genes by Flor in 1942, our knowledge of these molecules has greatly expanded, with various models produced to explain effector interactions with their receptors and the application of molecular biology techniques to elucidate the functions of chosen microbial effectors (Figure 1), but the study of effectors is far from black and white. The present review gives a comprehensive summary of the pioneering literature as well as, disruptive findings regarding effector molecular nature, localization, taxonomic distribution, and the types of interactions involving effectors. The associated changes that have occurred within effector identification methods over the years and novel bioinformatic tools are also briefly discussed. Remarkable effector reviews that summarized significant findings regarding effector identification, evolution, effector functions, host targets, and Avr-R interactions were published in 2009 [3,30,44] followed by noteworthy publications in the last decade that, in large part, have explored newly discovered effector functions and how they undermine plant defense [4,5,36,37,45,46]. To our knowledge, this review is the first of its kind to present recent updates in effector biology and demonstrate how they may differ from previously established dogmas in the field. Furthermore, a wider range of effector topics is addressed than in previous publications, and interesting findings are organized according to each theme. Finally, a comprehensive table compares the “past versus present” ideas surrounding these themes. In summary, the objective of the review was to demonstrate how effector concepts have evolved over the last three decades, challenging the initial definition of an effector.
2. Fundamentals of Effector Biology
Effectors were first called ‘avirulence factors’ as described by the botanist Flor in the 1940s [47]; genes called Avr were used by pathogens for which a cognate ‘R’ or resistance gene existed in plants resistant to the particular pathogen. This was dubbed the ‘gene-for-gene hypothesis and in Flor’s work, it was applied to the fungus Melampsora lini and the plant host, the flax plant, Linum usitatissimum. Though the original discovery was made between these two organisms it was found to apply to other pathogens in interaction with their plant hosts, although it is unlikely that the gene-for-gene interaction is the most common interaction found between effectors and targets [48]. Resistant plants evolved with resistance gene products to recognize pathogen Avrs and curtail the infection; this phenomenon is discussed later in this section. Conversely, gene products belonging to the host which promote the infection are encoded by susceptibility (S) genes, where negative regulators of plant immunity can be found [49]. Triggering susceptibility is fundamental for the compatible pathogen-host interaction, ultimately resulting in the development of disease. Most effectors characterized to date, target positive regulators of host immunity, interfering with their functionality in host defense [46].
Regarding plant immunity, MAMPs and effectors are the major elicitors of host defense; conserved molecules called MAMPs are the elicitors of host defense mechanisms (deposition of callose, induction of pathogenicity-related proteins, oxidative burst) that are collectively referred to as MAMP-triggered immunity (MTI). MTI is induced upon the recognition of MAMPs by plasma membrane-localized pattern recognition receptors (PRRs). These mechanisms are also induced and potentiated during another defense tier called Effector-triggered immunity (ETI) which is based on a unique protein-protein interaction: the recognition of the ‘Avr’ protein of the pathogen by the ‘R’ protein of the host. Many R proteins are intracellular and belong to the family of nucleotide-binding and leucine-rich repeat domain (NLR) proteins [50,51]. Regarding the pathogen, microbes most likely evolved with effectors to overcome the basal immunity response of the plant; when PTI and ETI are compromised in the host due to effector deployment, this phenomenon is referred to as “Effector-triggered susceptibility” [50,52,53]. The defining characteristic of ETI is a form of programmed cell death induced in the plant called the hypersensitive response (HR) and the induction of systemic acquired resistance (SAR), which prevents further propagation of the pathogen. ETI induced by the Avr-R protein interaction is essential to the incompatible pathogen-host interaction, where the pathogen fails to cause disease. Pathogen Avr genes have virulence functions that aid in disease development, e.g., the Cladosporium fulvum Avr effectors Avr2 [54] and Avr4 [55] which suppress MTI. Avr-Pik is another example of a fungal Avr whose virulence function is known; Avr-Pik binds and stabilizes the host protein OsHIPP20, a product of a susceptibility gene that promotes Magnaporthe oryzae infection [56]. Some Avrs suppress ETI mediated by other Avrs, like the AvrLm4-7 effector of Leptoshaeria maculans that suppresses the AvrLm3-Rlm3 mediated resistance response in Brassica napus [57]. The virulence functions of most Avrs remain unknown.
The Avr and R genes of pathogens and plants are under high selective pressure to evolve; mutations and/or gene losses and gains of the Avr allow pathogens to evade recognition while the plant (at a much slower rate) evolves new R genes to maintain the ability to recognize the Avrs in order to protect itself [50]. The direct interaction between Avr-R is one of a few types of effector-target interactions in the plant. The guard hypothesis suggests that R genes exist that guard many possible targets [58], while in the decoy model the Avr protein has an indirect target, a decoy, and the R protein is alerted by this interaction [59]. The integrated decoy model explains how some R proteins (NLR receptors) have evolved to integrate additional decoy domains that act as a sensor or bait for Avrs; in this interaction, the bait binds to the effector, and another receptor interacting with the bait triggers defense signaling [60,61]. In another effector interaction theory, not every molecule that an effector will interact with is its true target; effector ‘helpers’ exist which aid in signal transduction or signal amplification like “helper” NLRs which interact with true “sensor” NLRs [62,63]. Other helpers may be cofactors, chaperones that aid in protein folding, or transporters, among other functions, that ultimately aid in host susceptibility or defense [64,65].
The strict separation of the MTI-ETI dichotomy and the zig-zag model has been challenged for various reasons: (1) the zig-zag model was established for biotrophic pathogens, (2) some effectors are broadly conserved and can therefore be considered MAMPs, and (3) the stages of MTI and ETI are not static [66]. A more inclusive model called the “invasion model” was therefore proposed to rename the players involved. All immunogenic molecules (effectors and elicitors/MAMPs) are described as invasion patterns (IPs) perceived by plant IP receptors (IPRs), leading to an IP-triggered response (IPTR) [66]. The more recent “iceberg model” suggests that both resistance (R) and susceptibility (S) targets are monitored by NLRs that act in “interaction units”; the majority of interaction units are silent (due to effectors that suppress host immunity) and make up the larger portion of the iceberg which is under the surface and unseen, while the visible tip of the iceberg consists of those interaction units (effector and receptor interactions) that are able to induce host defense resulting in the resistant phenotype [53].
The latest developments in effector biology revolve around the MTI-ETI dichotomy as it is now clearer than ever that the two are more interrelated than they are distinct. MTI and ETI share many of the same components; recently it was observed that TIR signaling mutants (domains found in NLR resistance proteins involved in ETI) displayed compromised MTI and overexpression of TIR genes amplified the MTI defense response [67]. In the same vein, another study found that ETI’s hypersensitive response is enhanced by MTI PRRs and that effective host resistance is mediated by MTI and ETI working in concert [68].
3. Effector Identification: Past and Present
The first 80 years of effectoromics were challenge-ridden due to the lack of homology present among effector molecules and also due to discrepancies in effector identification methods. Certain criteria have been established to identify effector candidates; researchers either prepare in-house tailored pipelines for effector identification or use already established programs that are based on common effector characteristics. Effector candidature is usually determined based on sequence length ≤ 300 amino acids; cysteine richness (>2% cysteine content or >4 cys residues), presence of a secretory signal peptide; absence of transmembrane domains; higher expression in interaction with the host; limited taxonomic distribution with no or limited sequence similarity to other organisms and encoded by genes with long intergenic regions or in gene-sparse, repeat-rich chromosomes [30,36,69,70,71].
The number of validated effectors in recent times has increased from 96 in 2018 to more than 300 in 2022 [69,72,73,74]. This has occurred due to advances in ‘omics’ studies and the development of bioinformatic tools. Bioinformatic tools used for the prediction of effector characteristics include SignalP (https://services.healthtech.dtu.dk/service.php?SignalP-5.0; [75]), for the detection of signal peptides; TargetP (https://services.healthtech.dtu.dk/service.php?TargetP-2.0; [76]), LOCALIZER (http://localizer.csiro.au/; [77]), and WoLF PSORT (https://wolfpsort.hgc.jp/; [78]) for the determination of effector localization and DeepTMHMM (https://dtu.biolib.com/DeepTMHMM; [79]), for the prediction of transmembrane domains where the absence of said domains is preferred. Databases of experimentally validated proteins with roles in pathogen virulence have been especially useful for effector prediction. The database PHI-base which has a compendium of genes involved in plant-pathogen interactions allows users to compare their effector candidates with those homologs in the database using PHI-BLAST (http://phi-blast.phi-base.org/; [80]). These bioinformatic prediction tools and databases have supported effector predictions on high, medium, and low-throughput screenings for effectors. In the last decade, a turning-point for effector prediction came with the creation of software based on machine-learning (ML) methods that predict effectors based on sequence characteristics shared among experimentally validated effectors. Notably, Sperschneider and colleagues created the EffectorP series, the most commonly used machine learning algorithm for fungal effector prediction [74,81,82].
EffectorP 1.0 is a Naïve Bayes machine learning predictor that was trained with 58 true fungal effectors from 16 fungal species. The negative dataset (14,143 proteins) was constructed based on the secretomes (total set of secreted proteins) of 16 fungal species, filtering the known effectors and homologs. Although EffectorP 1.0 improved fungal effector prediction from secretomes, it was trained with a negative dataset including both undiscovered effectors and non-effectors [81]. EffectorP 2.0 was then trained with 94 secreted true effectors from 23 fungal species and the negative dataset (21,840 proteins) was constructed initially as it was for EffectorP 1.0, but with 23 fungal pathogen secretomes, after which EffectorP 1.0 was applied to exclude potentially undiscovered effectors. To this output, the secretomes from 27 non-pathogenic saprophytes, and 10 animal-pathogenic fungal secretomes were added. As a result, the updated EffectorP 2.0 was successful in decreasing by 40% the number of effector candidates from the negative set that was from fungal plant symbionts and saprophytes vs. EffectorP 1.0 [82]. EffectorP 2.0 has been the most frequently used fungal effector predictor in recent years. However, reports about effectors in non-pathogenic fungi are ever increasing [83,84], and these effectors may be excluded during effector mining, even by EffectorP 2.0. The latest EffectorP algorithm, version 3.0, classifies effectors according to apoplastic and cytoplasmic localization [74].
Another recent predictor of fungal effectors, Effhunter, is a pipeline written in Perl/Bioperl language that retrieves effector candidates that strictly meet the canonical or conventional effector criteria: secreted (signal peptide presence), small size (<400 amino acids), cysteine-rich (>4 residues) and no transmembrane domain [69]. Effhunter is a suitable effector predictor, possessing the highest F1 score for identifiers of canonical fungal effectors. Regarding oomycetes, the ML algorithm EffectorO identifies proteins unique to one species or genus of oomycetes and was able to predict larger effectoromes than previously estimated for Bremia lactucae and Phytophthora infestans [73].
Many effectors evade the current predictors because of their non-conventional nature; these effectors called non-canonical effectors (NCEs) constitute approximately 90% of an organism’s effectorome according to WideEffHunter (Carreón-Anguiano et al. submitted). Since some effectors do not meet all the criteria used to define them, researchers in effectoromics face a great challenge during in silico effector identification. To help researchers prioritize the most important criteria for selecting or ranking effectors, authors of WideEffHunter in silico characterized 314 true (validated) fungal and oomycete effectors. The ranking of the criteria was as follows: the absence of glycosylphosphatidylinositol anchors (GPI), 96.5%; the absence of transmembrane domains (TMD), 91.1%; sequence length < 400 amino acids, 89.4%; the presence of a signal peptide, 85%; extracellular localization, 71.6%; >4% Cys content, 54.4%. The determination of the importance or weight of each of these effector criteria will be beneficial for future effector predictions and the selection of candidates for further characterization in the lab.
NCE characteristics have been shown to vary across proteins; some are larger than 400 amino acids (Ace1: [85]), lack a signal peptide (VdIsc1: [86]), have low cysteine contents (AvrLm1: [87]) while others show high similarity to other effector sequences or are conserved among species e.g., ceratoplatanins [88], the latter being easier to recognize by pipelines that integrate homology as one of the effector identification characteristics. These non-conventional effectors require creative strategies for their identification, like the use of effector-related motifs. The motifs RXLR, LFLAK, Y/F/WxC, and CRN motifs are commonly used for identifying effectors in oomycetes, but recently, Zhao et al., (2020) used this strategy on the fungus Puccinia graminis and found 719 RXLR, 19 CRN, and 138 Y/F/WxC new effector candidates [89]. Recently, the Predector [90] pipeline, which was created for ranking effector candidates, was able to identify MoCDIP8, a non-canonical Magnaporthe oryzae effector with two predicted transmembrane domains and no signal peptide; the predictors EffHunter, EffectorP 1.0 or EffectorP 2.0 fail to recognize this protein as an effector. Predector also recognizes MoCDIP13 which is retrieved by EffectorP 1.0 but not by Effhunter and EffectorP 2.0.
Admittedly, it would be difficult for any singular algorithm to correctly identify all the effectors of an organism’s effectorome, but this is an exciting era where improvements in the current algorithms and newer algorithms are increasing the number of true effectors identified per organism. Hundreds of effector candidates are being identified from omics projects, necessitating strategies for prioritizing candidates for functional analysis.
4. Effector Conservation: Effectors Shedding the “Species-Specific” Label
The genes that encode effectors are commonly referred to as “compartmentalized” in gene-sparse, repeat-rich regions of the accessory chromosomes of the genome which facilitate their “high-speed” evolution. These regions are characterized by high mutation rates and chromosomal rearrangements [71,91,92,93]. A “two-speed” genome model proposes that genome organization could be broadly divided into two parts: the core genome which holds essential genes that are protected from high rates of mutation while the accessory genome has the effectors which require rapid evolution [92,94,95]. This two-speed genome arrangement has been documented, for example, in the fungi Verticillium dahliae [96], Leptosphaeria maculans [97], and Colletotrichum higginsianum [98]. The accessory or “dispensable” chromosomes are not found in all races or strains of a pathogen; they are referred to as strain- or pathotype-specific chromosomes [99]. However, not all genomes exhibit compartmentalization of virulence genes, and gene distribution is rather homogenous in “one-speed” genomes [100,101].
Effectors were once commonly referred to as race-specific or lineage-specific, taking part in interactions that are unique to one host or a limited host range [40,102,103]. Some effectors are more conserved than others and can be found among all the strains of a pathogen and are referred to as “core” effectors [41,104]. The term has been extended to those effectors that are also found in related species of a taxonomic family [42]. It is hypothesized that effector genes that are conserved among pathogens and reside in core genome regions are most likely to hold indispensable virulence functions. As such, they are not under the same rate of diversifying selection as their race-specific effector counterparts [105]. Various core effectors have been identified from the biotrophic fungus, Ustilago maydis. The Pep1 effector that was found in species across the Ustilaginaceae family including Sporisorium reilianum, S. scitamineum, Melanopsichium pennsylvanicum, and other Ustilago species (45.76–62.36% identity with the U. maydis Pep1) [106]. Pep1 is indispensable for pathogen virulence and is an inhibitor of apoplastic plant peroxidases, suppressing the oxidative burst. Cce1 is another core effector of Ustilago maydis of unknown functionality that is indispensable for infection; Cce1 is conserved among other smut fungi showing a range of sequence identity between 59–65% [107]. In U. maydis, a substantial number of its effectors are core effectors; 202 of the 467 effector candidates have an ortholog in the related smut pathogens [108].
Colletotrichum fungi represent another interesting model for effector conservation. CgEP1 was initially identified in the maize anthracnose pathogen, Colletotrichum graminicola. This protein is synthesized during the early stages of disease development and targets the host’s nucleus. EP1 arose from a gene duplication event in an ancestor of Colletotrichum sp. and has been under positive selection, resulting in CgEP1 homologs in several species of Colletotrichum sp. [109]. A recent inventory of effectors in Colletotrichum sp. genomes revealed 288–608 effectors per genome; cluster analysis of the effectoromes revealed that ~20% of conserved effectors were core effectors present in all Colletotrichum species [110]. Some examples of conserved effectors are NIS1, EC92, DN3, EC2-1, CEC2-2, CEC3, and CEC6, among others [110,111,112]. The core effector, NIS1, is not only found throughout the Colletotrichum genus but is broadly conserved in Ascomycota and Basidiomycota filamentous fungi [41]. In opposition to the usual effector narrative, most effector candidates of Colletotrichum species are conserved; only 4.1–15.6% of their effectoromes consist of species-specific effectors [110], giving support to the concept that each effectorome is a function of species host range and virulence.
It appears that not all shared effector homologs among species have the same functions, rather, some have evolved to have additional or different functions to suit their organism’s needs. Avr4 is a fungal effector and chitin-binding lectin that protects fungal cell walls against plant chitinases [55]. It was first described in Cladosporium fulvum, the causal agent of the leaf mold of tomato. Homologs of this protein were identified in Pseudocercospora (previously Mycosphaerella) fijiensis, and in Cercospora beticola, C. nicotianae, C. apii, and C. Zeina [113], Dothistroma septosporum [114], and Pseudocercospora fuligena [115], all of which are related Dothideomycete fungi. In addition to its role as an avirulence protein, the Avr4 of C. flagellaris, the causal agent of Cercospora leaf blight (CLB) on soybean, was found to be involved in cercosporin biosynthesis, since the Avr4 knockout mutant had a dramatically reduced production of this toxin [116]. In P. fuligena it was found that a second copy (paralog) of Avr4, called PfAvr4-2, does not bind chitin but it binds to highly de-esterified pectin to relax the plant cell wall structure with the help of secreted endo-polygalacturonases to facilitate pathogen entry [117]. Another effector first described in C. fulvum, Ecp2, has homologs in closely related fungi, like P. fijiensis and Mycospharella graminicola [113], but the function of this effector is unknown. As more genome sequencing and effector identification studies are performed, we are likely to see more effectors being present across related and even unrelated species and genera. The previous effector families are shared among phylogenetically close relatives. The next examples have larger phylogenetic distribution.
Ecp6 is a secreted LysM-containing effector first described in C. fulvum. Ecp6 binds chitin oligosaccharides that are released upon degradation of the fungal cell wall, avoiding detection by the host chitin receptors [118]. Proteins with LysM domain are conserved throughout fungal taxa and evasion of immune detection by Ecp6s may very well be a common strategy of fungi to subvert host immunity [119]. RALPH and MAX constitute large effector families that were more recently discovered. The former, “RNAse-Like Proteins Associated with Haustoria” (RALPH), comprises 25% of the predicted effectors in Blumeria graminis [120], meanwhile MAX or “Magnaporthe Avrs and ToxB like” effectors were discovered by comparative 3D modeling of Magnaporthe oryzae effectors and effectors of the phylogenetically distant pathogen Pyrenophora tritici-repentis [121]. To date, they represent between 5 to 10% of the Magnaporthe oryzae and M. grisea effectoromes [122]. Functional analysis has validated that these proteins induce clear necrosis in Nicotiana benthamiana when co-expressed with the corresponding ‘cognate’ resistant protein.
ToxA, first identified in Pyrenophora tritici-repentis, has homologs in Parastagonospora nodorum, and Bipolaris sorokiniania, among other species, that most likely originated through horizontal transfer of these genes [123]. Similarly, AvrLm6 first reported in Leptosphaeria maculans, has been reported in Leptosphaeria biglobosa, Fusarium oxysporum, Colletotrichum sp., Venturia inaequalis and V. pirina [90]. Lastly, a Crinkler effector candidate CRN13, from the legume root pathogen Aphanomyces euteiches, is also found in the genome of the amphibian fungal pathogen, Batrachochytrium dendrobatidis. The effector was found to have detrimental effects on plants through the inhibition of root growth and produced abnormal growth in frog embryos [124]. The number of conserved effector families is still small, but the use of common domains during effector identification and 3-D homology modeling has undoubtedly improved our ability to predict effectoromes and will enable us to identify new effector families shared in even taxonomically distant microorganisms. Table 1 presents different levels of conservation found in effectors, from species-specific to widely distributed conserved effectors.
5. Effector Targets: Beyond the Apoplast
The effector leaves the organism’s secretory system to arrive at either its cell wall, as in the case of those “protective effectors” (e.g., the ortholog of Avr4, PfAvr4 [145]), or is secreted into the extracellular space where it finds its host target in the apoplast or in the cytoplasm. Inside the cell, effectors may target various intracellular organelles (Figure 2). The haustorium is associated with fungal effector delivery [146], though effectors have been found to be secreted from conidia [147], as well as appressoria [130]. In pioneer investigations between 1996 and 2008 in fungal effector biology, the effectors that were identified and characterized were mainly apoplastic, like the Avr and Ecp effectors from the biotrophic fungus Cladosporium fulvum [5,30,48]. Regarding fungi and oomycetes, 314 protein effectors have been characterized to date, 228 from fungi, and 86 from oomycetes (compilation from the literature by Carreón-Anguiano et al., submitted). Recently, 176 known effectors were classified; 64 apoplastic (50 from fungi and 14 from oomycetes), and 112 cytoplasmic effectors (77 from fungi and 35 from oomycetes) [74].
The way in which effectors arrive at their targets is an interesting and controversial facet of effector biology. Canonical secretion in eukaryotes occurs through the endoplasmic reticulum and Golgi apparatus, then across the cell membrane via exocytosis of the secreted vesicles of the Golgi. Secreted proteins commonly have signal peptides: protein-sorting signals that direct the protein to the cell’s protein secretion machinery [148]. Conversely, proteins that lack signal peptides are usually considered to have non-canonical secretion and can be secreted by vesicular and non-vesicular pathways; vesicular pathways involve autophagy-based secretion and Golgi-bypass proteins while non-vesicular secretion includes protein translocation across plasma membranes and secretion by ABC-transporters [149]. It is not well known which of these pathways, if any, are used for non-canonical secretion of effectors, only that some effectors lacking predicted N-terminal signal peptides do exist and are secreted to the extracellular space. In the oomycete, Phytophthora infestans, a cytoplasmic effector lacking a signal peptide, Pi410314, was found to exhibit non-canonical secretion as Brefeldin A inhibition assays did not inhibit the retrograde secretion of the effector, compared to an apoplastic effector studied which showed canonical ER-Golgi secretion [150]. PsIsc1 of P. sojae and VdIsch1 of Verticillium dahliae are effectors that lack predicted N-terminal signal peptides. The effectors were mutated to remove the entire N-terminal region and as a result, both were found to have markedly reduced virulence in cotton seedlings indicating that putative unconventional secretion signals located in the N-terminal region of the proteins are necessary for their secretion [86]. Probably the most well-understood non-canonical secretion occurrence is in Magnaporthe oryzae, where cytoplasmic effectors were found to be secreted non-canonically by a form of secretion involving the exocyst complex and t-SNAREs [151] while apoplastic effectors such as BAS4, BAS113, and SLP1 are secreted by the conventional ER-Golgi pathway [152]. In the case of oomycetes, motifs in their sequences like the N-terminal RxLR-(d)EER motif mediate host cell entry by binding to phosphatidylinositol phosphates on the outer surface of plant plasma membranes [153]; the RXLR motif has since been identified in some fungi [154]. Other sequence features allowing effector movements are chloroplast/mitochondrial transit peptides and nuclear localization signals. These sequence features are native to plant proteins, and it is hypothesized that through mimicry, effectors would have evolved with these transit peptides to reach organelles such as the chloroplast [155].
Large-scale effector visualization has been achieved using fluorescent tags in Colletotrichum higginsianum; three effectors were targeted to plant peroxisomes, three others to plant cortical microtubules, and one to the Golgi apparatus, along with nine nuclear-targeted effectors [156]. In Melamspora larici-populina, effectors localized to the nucleus, chloroplast, and mitochondria [155]. In the oomycete, Plasmopara viticola, twenty-nine effectors were found to localize to the nucleus, nine to the cell membrane, three to chloroplasts, and one that targeted both chloroplast and mitochondria [157]. As more localization studies are undertaken, we will most likely see more effectors targeting all organelles in the plant host although the most common organelles targeted appear to be the nucleus and the cell membrane. Table 2 presents examples of effectors which have been proven to localize to different plant cell organelles. In a comparative study of effector targets of bacteria, oomycetes, and fungi, it was found that 95% of bacterial effector targets are in the cell membrane, nucleus, and/or cytoplasm while this value decreases to 63% for oomycetes that show more diversity in their targets (inclusive of peroxisomes and endoplasmic reticula). With respect to fungi, it was found that 61% of effector targets are cytoplasmic and are involved in signaling and protein processing, while signaling and transcription are more prevalent bacterial targets. The previously mentioned targets as well as those linked to metabolism are the main targets of oomycetes [158].
With respect to effector targets, some effectors have been found to have more than one target in an organism, like AvrLm4-7 of Leptosphaeria maculans which interacts with multiple R genes in Brassica napus [187]. On the other hand, a target may interact with multiple effectors of the same pathogen like the RGA4/RGA5 receptor pair in rice that is recognized by AVR-Pia and AVR1-CO39 of Magnaporthe oryzae [139]. Furthermore, one target may have interactions with several effectors of different organisms; the Rcr3 protease of tomato is targeted by the Avr2 effector of Cladosporium fulvum and two other effectors, EPIC1 and EPIC2B of Phytophthora infestans [188]. These targets in the host that attract many effectors have been denominated ‘hubs’ and are central in plant protein-protein interaction networks [189,190]. Common hubs include the serine/threonine protein kinase and mitogen-activated protein kinase families which are actively involved in plant immune signaling, transcription factors, enzymes involved in the biosynthesis or regulation of jasmonic acid and salicylic acid pathways, and other phytoregulators involved in host responses [191]. Similarly, common proteins or hubs in pathogens have been identified in interaction networks, for example, ubiquitin-like activating enzymes, small GTPases such as Rho, Ran, and Ras, SUMO-conjugating enzymes, thioredoxin reductase, among others [192]. Table 3 highlights examples of hubs in plant hosts and pathogens. Recently, through mining deduced proteomes and conducting interactomics analyses, multiple targets of effectors from different kingdoms (bacteria, fungi, oomycetes, and nematodes) were identified in Arabidopsis thaliana [193]. The existence of these common interactors demonstrates how pathogens evolved to cleverly debilitate their hosts, and in this manner, maintain low fitness costs as compared with the individual gene-for-gene interactions [189]. More high-throughput interactomics network studies are necessary to elucidate effector targets [194,195,196].
6. Effector Nature: The Rise of Non-Proteinaceous Effectors
The majority of effectors reported are small (<300 aa), secreted proteins (SSPs) which make up around 2–3% of the total proteome of fungal organisms [10]. Secondary metabolites and small RNAs (sRNA) have also been reported to suppress PTI and thus act as effectors. Here, the roles of these molecules are explored with the spotlight on plant-pathogen interactions.
6.1. RNA Effectors
RNA effectors discovered to date belong to the class of small silencing RNAs (sRNAs). These are short, non-coding RNAs capable of gene expression regulation through binding to host Argonaute (AGO) proteins and directing the RNA-induced silencing complex (RISC) to RNAs with complementary sequences [202]. In a pioneer study involving the fungal pathogen, Botrytis cinerea, sRNAs were found to bind to the argonaute protein 1 (AGO1) of the RNAi machinery in Arabidopsis to commandeer the host’s RNA-interference machinery and induce gene silencing of host targets. These sRNAs were able to silence transcripts of mitogen-activated protein (MAP) kinases such as mitogen-activated protein kinase 2 (MPK2) and MPK1 which are responsible for cell signaling to activate defense responses against pathogens. Transgenic Arabidopsis containing these pathogen sRNA effectors were more susceptible to the disease than the wild-type control plants, proving that these sRNAs play a role in the suppression of host immunity [34]. A more recent study showed that B. cinerea sRNA, Bc-siR37, targets the transcripts of multiple host proteins in Arabidopsis to subvert plant immunity [35]. The pathogen Puccinia striiformis f. sp. tritici (Pst) through the microRNA-like RNA 1 (Pst-milR1) attenuates wheat immunity by silencing the pathogenicity-related protein 2 (PR2), a protein that contributes to wheat resistance against the virulent Pst isolate [203]. This phenomenon of sRNA effectors translocating from the invader to the plant host to induce gene silencing of one or multiple targets is referred to as cross-kingdom RNA interference [204,205] and can be bidirectional when plants, in turn, release RNAs in their defense [206,207]. Pathogens, in response, have evolved effectors to suppress the RNA-mediated immunity of plants; Arabidopsis siRNAs that silence pathogen genes are inhibited by the Phytopthora protein effector PSR2 which interferes with host dsRNA processing [208].
RNA effectors have also been identified beyond the plant-pathogen interaction. The entomopathogenic fungus, Beauveria bassiana, uses a microRNA-like effector called bba-milR1 to silence the mosquito Toll receptor ligand Spätzle 4 (Spz4) upon host penetration to facilitate infection [202] and in the same interaction, it was later discovered that mosquitoes also use microRNAs (miRNAs) that translocate to fungal hyphae and target virulence genes in a fascinating example of insect-fungus cross-kingdom RNAi [209]. In silico RNA effector prediction is likely to gain popularity among researchers with the advent of bioinformatic pipelines for RNA effector identification [210], and high-throughput small RNA sequencing for the determination of sRNAs and their targets involved in host immunity [211,212,213,214].
6.2. Secondary Metabolite Effectors
Secondary metabolites (SM) are low molecular weight compounds that are not essential for growth but are still advantageous to the producer since they indirectly contribute to the survival and adaptation of the organism in its ecological niche [215,216]. Many fungal SMs are derivatives of polyketides, non-ribosomal peptides, hybrid polyketide–non-ribosomal peptides, and terpenes [217,218]. The genes responsible for SM production are usually found together in large genomic clusters and are coregulated [219]. Some of the core SM biosynthetic enzymes are polyketide synthases (PKSs), non-ribosomal peptide synthases (NRPSs), prenyltransferases (PTs)/terpene synthases (TSs), and dimethylallyl tryptophan synthases (DMATSs) [220,221].
Effectors that are secondary metabolites have a role in disease establishment and either induce necrotrophy [host selective toxins (HSTs) and non-HSTs] or do not induce necrosis but contribute to pathogen virulence [221]. Necrotrophs tend to have a larger proportion of SM biosynthesis genes in their genomes than biotrophs [222]. Necrotroph HSTs are involved in the inverse gene-for-gene interaction: a phenomenon occurring in a susceptible host where the HST effector is recognized by the host susceptibility gene product (S), triggering host cell death which is beneficial to the necrotroph. The most characterized SM effectors have been identified from Cochliobolus sp., host-specific necrotrophic fungi that target maize and use HSTs like T-toxin, victorin, and HC-toxin to establish disease on susceptible hosts [220]. T-toxin is a polyketide produced by Cochliobolus heterostrophus which targets the mitochondrial protein, URF13, resulting in the formation of pores in the mitochondrial membrane and necrosis [223,224]. A non-HST secreted by the hemibiotroph, Pseudomonas syringae, called coronatine (COR) (a hybrid between a polyketide, coronafacic acid (CFA), and a derivative of isoleucine, coronamic acid), suppresses salicylic acid (SA)-dependent defense responses and is a virulence factor for disease development in A. thaliana. In order to facilitate colonization, the toxin mimics one or more plant jasmonates involved in jasmonate signaling which antagonizes SA-dependent defense responses [225]. Some pathogen SMs have a positive contribution to plant health in the presence of a resistant host, for example, the Ace1 polyketide produced by the hemibiotroph, Magnaporthe grisea, which triggers resistance when recognized by the rice Pi33 resistance protein [226].
In silico genomics analyses of the SM gene clusters in pathogenic fungi have revealed the proportions of these core enzymes and their homologs in other fungi [219,227] and transcriptomic studies have revealed potential SM effectors that are upregulated in interaction with the host [218,228], but the characterization of the SMs has been a challenge due to difficulties in isolating metabolites found in small quantities, the need for suitable solvents, the limited availability of standards and limited mass spectra and NMR databases [229,230]. In addition, in silico reconstructed microbial SM pathways are sometimes incomplete, either because the genome in question has not been completely sequenced or because the microorganisms share part(s) of the pathways but have different end products [231,232,233] and to investigate and clarify these possibilities is not trivial work.
7. Effectors: Not Just Plant-Pathogenic Molecules
Until recently, the effector narrative has been centered on pathogenicity; effectors commonly being described as pathogen proteins and small molecules [3,59]. Pathogenic interactions between plants and microorganisms such as Cladosporium fulvum-tomato [234,235]; Phytophthora infestans-potato [236,237], the Pseudomonas syringae species complex with various hosts [238,239], are prime examples of well-studied pathosystems utilized for fungi, oomycetes, bacteria, respectively, and are also the leading pathosystems in effector biology studies. A budding area of interest centers on effectors beyond the plant-pathogen interaction, with discoveries in non-pathogenic organisms such as mutualist mycorrhizal species [13,186,240] and endophytes [16,241,242] in interaction with the plant host. These organisms also modulate host immunity with the help of effectors to establish plant-beneficial interactions.
One of the more noteworthy discoveries in effector biology has been the role of effectors in microbial interactions. In these interactions, effectors are required to subdue microbial competitors and dominate their respective niches [243]. Literature supporting the evidence of effectors in microbial interactions can be found for bacteria, especially effectors from the type VI secretion system that antagonize other bacterial and fungal competitors [244,245,246,247]. In fungal interactions, examples of effector candidates can be found in mycoparasitic interactions involving Trichoderma sp. against Rhizoctonia solani [248], Pseudomyza flocculosa against Blumeria graminis [147], and Pythium oligandrum against Phytophthora infestans [249].
Due to effector findings in interactions beyond the plant host, it has been postulated that effectors can be additionally classified based on their host specificity: effectors that target only plants, effectors that target only microbes, and effectors that target both plants and microorganisms [43] (Figure 3). Possible broad host-range effectors were identified in Trichoderma atroviride and T. virens, where expression of candidate effector genes (LysM protein, Epl2, and a hydrophobin, Tvhydii1) was observed in the presence of the plant Arabidopsis thaliana, as well as, during in vitro interaction with the fungus Rhizoctonia solani. In the same study, a metalloprotease was found to only be induced in the interaction with R. solani but not with the plant, a possible example of a host-specific effector [248]. Conversely, T. atroviride’s Tal6 is a broad host-range effector acting in microbe-microbe and microbe-plant interactions [250]. Tal6 is an effector with LysM domains that allow it to bind to chitin from the fungal cell wall, serving as protection against plant chitinases and interfering with the perception of its chitin fragments that could trigger a host immune response. The gene was upregulated during contact with the phytopathogen R. solani and Tal6 deletions mutants showed a decrease in antagonism against R. solani, B. cinerea, Sclerotium cepivorum and C. lindemuthianum, while overexpression of Tal6 results in increased antagonistic capacity [250]. Although the exact function of the Tal6 gene in microbial interactions was not elucidated, it may similarly act in hyphal protection against microbial chitinases as it does with plant chitinases.
Another addition to the short list of effectors discovered in microbe-microbe interactions is Zt6, a ribonuclease effector of Zymoseptoria tritici. This effector is detrimental to the plant host Triticum aestivum, the non-host N. benthamiana, as well as the bacterium E. coli and the yeasts Saccharomyces cerevisiae and Pichia pastoris during in vitro assay [201]. Recently, the functions of two effectors, VdAve1 and VdAMP3 of the fungal pathogen, V. dahliae, have been characterized in microbial interactions [251]. In plant-pathogen interaction, VdAve1 is an avirulence effector that is recognized by the receptor Ve1 in tomatoes. In microbial interaction, this effector was shown to selectively inhibit the growth of gram-positive bacteria in vitro. Analysis of the bacterial communities 10 days after in planta infection with V. dahlia wildtype and VdAve1 deletion mutant strains revealed significant differences in the microbiomes present; for example, bacteria of the Sphingomonadaceae family were completely absent in cotton plants infected with the wildtype fungus, meanwhile, they were present where the mutant was used. The direct application of the purified VdAve1 protein on the bacteria produced the same result, ruling out indirect effects of the effector on the host [251]. Conversely, VdAMP3, allows the fungus to suppress other fungi in decomposing plant tissue, while it forms its resting structures called microsclerotia, the ultimate phase of its disease cycle. The effector showed potent and specific antimicrobial activity against Saccharomycete and Sordariomycete filamentous fungi and yeasts, while bacteria are suppressed to a much lesser degree [252]. However, VdAMP3 does not appear to contribute to infection of the plant, N. benthamiana, since the V. dahliae mutant lacking the effector gene remained virulent, suggesting this effector is specifically employed in microbe-microbe interactions. The identification of microbial antagonism-related effectors opens a new avenue to produce novel antimicrobials as alternatives against recalcitrant pathogens which display resistance against the current antimicrobials. These effectors may prove useful not only for plants but additionally, for human health.
8. Discussion: Coming Changes to Effectoromics
As highlighted in this review, the initial beliefs about effectors have constantly been evolving with novel effector discoveries. This is the first time such a range of effector characteristics has been comprehensibly presented in the context of past and present knowledge related to effectors. It is also uncommon to witness within the effector literature, varied types of effector classifications apart from the common classification of localization in the plant host (apoplast or cytoplasm).
The dogmas in effectoromics seem as though they are meant to be challenged (Table 4) and as such, the effector literature merits a change in how effectors are defined. Effectors are often referred to as virulence-associated molecules due to their role in disease development; they either induce susceptibility or resistance depending on the presence of related targets in the host and as such, can have a dual function as promoters of disease or health. The proverbial box in which effectors were first placed is being dismantled as these molecules have been found beyond the limits of pathogenicity and virulence, although the socioeconomic impact of disease merits the attention paid to effectors in plant-pathogen interactions. Ultimately, effectors should be referred to as interaction-associated molecules since they are used by both pathogenic and non-pathogenic organisms.
The study of effectors in microbe-microbe interactions and in tripartite plant-pathogen-mutualist interactions have revealed effectors involved in mycoparasitism and biological control of pathogens in the host [201,248,253], as well as the induction of disease resistance in plants [254,255]. Microbial effector-based screening may facilitate the selection of elite strains of antagonists that can be applied to the field. It also forms the basis for the development of novel bioproducts for exogenous application to plants. Effective effector-bioproducts may be used to trigger systemic resistance or help modulate the composition of plant microbiomes for plant health promotion. One of the biggest limitations in understanding effector molecules in microbe-microbe interactions is the technical barrier associated with elucidating their targets and their localization in vivo.
Investigations into the host-pathogen interactomes are identifying core proteins that are suitable targets for engineering disease resistance [104,105], and novel algorithms/pipelines for identification of proteinaceous and non-proteinaceous effectors are elucidating complete pathogen effectoromes providing a sound foundation for effectoromics-based pathogen control strategies [69,72,73,81,210]. To date, the number of effector families is still small, but the construction of novel algorithms that allow us to expand and compare effectoromes may result in the identification of conserved effector families. The identification and characterization of conserved effectors among pathogens (core effectors) may provide effective solutions to disease management through their inactivation in the field.
Another necessary revolution in effectoromics involves the elucidation of the true effectorome; the identification of non-canonical effectors will be essential for the expansion of in silico-identified effector sets where the implementation of common effector domains and motifs are particularly useful for this type of effector. More occurrences are likely to be revealed of domains and motifs shared between fungi and oomycetes that were previously described as unique to each kingdom e.g., the RXLR and CRN motifs that are associated with oomycetes and LysM, ceratoplatanin, ribonuclease/ribotoxin, etc. domains commonly associated with fungal effectors. Additionally, the identification of novel motifs among the different kingdoms represents a promising tool that will promote effectorome elucidation.
9. Conclusions
Locating the resistance proteins targeted by Avr effectors in plant hosts has been a priority in effectoromics research for effector-assisted plant breeding [256,257,258], but varied possibilities for disease management exist with effector molecules [259]. This review highlighted many changes which have occurred in the study of fungal effectors since the gene-for-gene model. Effector research, especially in the last 30 years, has revealed the dynamic nature of effectors in different interactions, with and beyond the plant host. There are effectors which exist of varying molecular natures, functions, localizations in the host cell, and interactions (with one or more hosts and targets). Investigations surrounding effector targets beyond the R gene and how they can be utilized in plant protection should dominate effectoromics in the coming years; whether a particular effector has the same target in different hosts is an important line of investigation to be taken into consideration. In addition to the changes to the field mentioned in this review, others are on the horizon and are sure to improve our current understanding of what effectors are and how they can be applied to our benefit. Successful times are in sight for effectors in biotechnology.
Author Contributions
B.C.-C. and J.N.A.T. conceived and drafted the review. K.G.C.-A. created the figures for the review. B.C.-C., J.N.A.T., K.G.C.-A. and I.I.-F. contributed to the writing and correction of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received funding from CONACyT-Mexico project FOP16-2021-01 No. 320993 and CONACyT-funded scholarship for doctoral student Todd J.N.A (863239).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kamoun, S. A Catalogue of the Effector Secretome of Plant Pathogenic Oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamoun, S. Groovy Times: Filamentous Pathogen Effectors Revealed. Curr. Opin. Plant Biol. 2007, 10, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging Concepts in Effector Biology of Plant-Associated Organisms. Mol. Plant-Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, M.C.; Valent, B. Filamentous Plant Pathogen Effectors in Action. Nat. Rev. Microbiol. 2013, 11, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Selin, C.; de Kievit, T.R.; Belmonte, M.F.; Fernando, W.G.D. Elucidating the Role of Effectors in Plant-Fungal Interactions: Progress and Challenges. Front. Microbiol. 2016, 7, 00600. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action Mechanisms of Effectors in Plant-Pathogen Interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef]
- Koeck, M.; Hardham, A.R.; Dodds, P.N. The Role of Effectors of Biotrophic and Hemibiotrophic Fungi in Infection. Cell. Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Yuan, Q.; Tang, J.; Huang, J.; Hsiang, T.; Wei, Y.; Zheng, L. Colletotrichum higginsianum as a Model for Understanding Host–Pathogen Interactions: A Review. Int. J. Mol. Sci. 2018, 19, 2142. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Kahmann, R. Cell Wall–Associated Effectors of Plant-Colonizing Fungi. Mycologia 2021, 113, 247–260. [Google Scholar] [CrossRef]
- Kim, K.-T.; Jeon, J.; Choi, J.; Cheong, K.; Song, H.; Choi, G.; Kang, S.; Lee, Y.-H. Kingdom-Wide Analysis of Fungal Small Secreted Proteins (SSPs) Reveals Their Potential Role in Host Association. Front. Plant Sci. 2016, 7, 00186. [Google Scholar] [CrossRef]
- Shao, D.; Smith, D.L.; Kabbage, M.; Roth, M.G. Effectors of Plant Necrotrophic Fungi. Front. Plant Sci. 2021, 12, 995. [Google Scholar] [CrossRef] [PubMed]
- Samperna, S.; Masi, M.; Vurro, M.; Evidente, A.; Marra, M. Cyclopaldic Acid, the Main Phytotoxic Metabolite of Diplodia Cupressi, Induces Programmed Cell Death and Autophagy in Arabidopsis Thaliana. Toxins 2022, 14, 474. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Aerts, A.; Ahrén, D.; Brun, A.; Danchin, E.G.J.; Duchaussoy, F.; Gibon, J.; Kohler, A.; Lindquist, E.; Pereda, V.; et al. The Genome of Laccaria bicolor Provides Insights into Mycorrhizal Symbiosis. Nature 2008, 452, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Kemppainen, M.; Kale, S.D.; Kohler, A.; Legué, V.; Brun, A.; Tyler, B.M.; Pardo, A.G.; Martin, F. A Secreted Effector Protein of Laccaria bicolor Is Required for Symbiosis Development. Curr. Biol. 2011, 21, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Voß, S.; Betz, R.; Heidt, S.; Corradi, N.; Requena, N. RiCRN1, a Crinkler Effector From the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis, Functions in Arbuscule Development. Front. Microbiol. 2018, 9, 2068. [Google Scholar] [CrossRef] [PubMed]
- Wawra, S.; Fesel, P.; Widmer, H.; Timm, M.; Seibel, J.; Leson, L.; Kesseler, L.; Nostadt, R.; Hilbert, M.; Langen, G.; et al. The Fungal-Specific β-Glucan-Binding Lectin FGB1 Alters Cell-Wall Composition and Suppresses Glucan-Triggered Immunity in Plants. Nat. Commun. 2016, 7, 13188. [Google Scholar] [CrossRef]
- Snelders, N.C.; Rovenich, H.; Thomma, B.P.H.J. Microbiota Manipulation through the Secretion of Effector Proteins Is Fundamental to the Wealth of Lifestyles in the Fungal Kingdom. FEMS Microbiol. Rev. 2022, 46, fuac022. [Google Scholar] [CrossRef] [PubMed]
- Redkar, A.; Sabale, M.; Zuccaro, A.; Di Pietro, A. Determinants of Endophytic and Pathogenic Lifestyle in Root Colonizing Fungi. Curr. Opin. Plant Biol. 2022, 67, 102226. [Google Scholar] [CrossRef]
- Hulin, M.T.; Armitage, A.D.; Vicente, J.G.; Holub, E.B.; Baxter, L.; Bates, H.J.; Mansfield, J.W.; Jackson, R.W.; Harrison, R.J. Comparative Genomics of Pseudomonas syringae Reveals Convergent Gene Gain and Loss Associated with Specialization onto Cherry (Prunus avium). New Phytol. 2018, 219, 672–696. [Google Scholar] [CrossRef] [Green Version]
- Frantzeskakis, L.; Di Pietro, A.; Rep, M.; Schirawski, J.; Wu, C.-H.; Panstruga, R. Rapid Evolution in Plant–Microbe Interactions –a Molecular Genomics Perspective. New Phytol. 2020, 225, 1134–1142. [Google Scholar] [CrossRef]
- Gómez Luciano, L.B.; Tsai, I.J.; Chuma, I.; Tosa, Y.; Chen, Y.-H.; Li, J.-Y.; Li, M.-Y.; Lu, M.-Y.J.; Nakayashiki, H.; Li, W.-H. Blast Fungal Genomes Show Frequent Chromosomal Changes, Gene Gains and Losses, and Effector Gene Turnover. Mol. Biol. Evol. 2019, 36, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, M.; Zhang, Y.; Song, Z.; Zhang, S.; Zhang, Q.; Xu, J.-R.; Liu, H. Extensive Chromosomal Rearrangements and Rapid Evolution of Novel Effector Superfamilies Contribute to Host Adaptation and Speciation in the Basal Ascomycetous Fungi. Mol. Plant Pathol. 2020, 21, 330–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leelarasamee, N.; Zhang, L.; Gleason, C. The Root-Knot Nematode Effector MiPFN3 Disrupts Plant Actin Filaments and Promotes Parasitism. PLoS Pathog. 2018, 14, e1006947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kud, J.; Wang, W.; Gross, R.; Fan, Y.; Huang, L.; Yuan, Y.; Gray, A.; Duarte, A.; Kuhl, J.C.; Caplan, A.; et al. The Potato Cyst Nematode Effector RHA1B Is a Ubiquitin Ligase and Uses Two Distinct Mechanisms to Suppress Plant Immune Signaling. PLoS Pathog. 2019, 15, e1007720. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.A.; Escudero-Martinez, C.; Bos, J.I.B. An Aphid Effector Targets Trafficking Protein VPS52 in a Host-Specific Manner to Promote Virulence. Plant Physiol. 2017, 173, 1892–1903. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.-X.; Qian, L.-X.; Wang, X.-W.; Shao, R.-X.; Hong, Y.; Liu, S.-S.; Wang, X.-W. A Salivary Effector Enables Whitefly to Feed on Host Plants by Eliciting Salicylic Acid-Signaling Pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Yin, H.; Mewalal, R.; Hu, R.; Li, T.; Ranjan, P.; Jawdy, S.; Paoli, H.C.D.; Butler, G.; Burch-Smith, T.M.; et al. Populus trichocarpa Encodes Small, Effector-like Secreted Proteins That Are Highly Induced during Mutualistic Symbiosis. Sci. Rep. 2017, 7, 382. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Liu, H.; Wafula, E.K.; Honaas, L.; de Pamphilis, C.W.; Timko, M.P. SHR4z, a Novel Decoy Effector from the Haustorium of the Parasitic Weed Striga gesnerioides, Suppresses Host Plant Immunity. New Phytol. 2020, 226, 891–908. [Google Scholar] [CrossRef] [Green Version]
- Alfano, J.R.; Collmer, A. Type III Secretion System Effector Proteins: Double Agents in Bacterial Disease and Plant Defense. Annu. Rev. Phytopathol. 2004, 42, 385–414. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal Effector Proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D.; Solomon, P.S.; Oliver, R.P. Host-Specific Toxins: Effectors of Necrotrophic Pathogenicity. Cell. Microbiol. 2008, 10, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Collemare, J.; Mehrabi, R.; De Wit, P.J.G.M. Phytotoxic Secondary Metabolites and Peptides Produced by Plant Pathogenic Dothideomycete Fungi. FEMS Microbiol. Rev. 2013, 37, 67–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collemare, J.; O’Connell, R.; Lebrun, M.-H. Nonproteinaceous Effectors: The Terra Incognita of Plant–Fungal Interactions. New Phytol. 2019, 223, 590–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Dellota, E.; Yamane, D.; Jin, H. Botrytis Small RNA Bc-SiR37 Suppresses Plant Defense Genes by Cross-Kingdom RNAi. RNA Biol. 2017, 14, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Rocafort, M.; Fudal, I.; Mesarich, C.H. Apoplastic Effector Proteins of Plant-Associated Fungi and Oomycetes. Curr. Opin. Plant Biol. 2020, 56, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Ortiz, D.; Henningsen, E.C. Tactics of Host Manipulation by Intracellular Effectors from Plant Pathogenic Fungi. Curr. Opin. Plant Biol. 2021, 62, 102054. [Google Scholar] [CrossRef]
- Schulze-Lefert, P.; Panstruga, R. A Molecular Evolutionary Concept Connecting Nonhost Resistance, Pathogen Host Range, and Pathogen Speciation. Trends Plant Sci. 2011, 16, 117–125. [Google Scholar] [CrossRef]
- Bourras, S.; Kunz, L.; Xue, M.; Praz, C.R.; Müller, M.C.; Kälin, C.; Schläfli, M.; Ackermann, P.; Flückiger, S.; Parlange, F.; et al. The AvrPm3-Pm3 Effector-NLR Interactions Control Both Race-Specific Resistance and Host-Specificity of Cereal Mildews on Wheat. Nat. Commun. 2019, 10, 2292. [Google Scholar] [CrossRef]
- Irieda, H.; Inoue, Y.; Mori, M.; Yamada, K.; Oshikawa, Y.; Saitoh, H.; Uemura, A.; Terauchi, R.; Kitakura, S.; Kosaka, A.; et al. Conserved Fungal Effector Suppresses PAMP-Triggered Immunity by Targeting Plant Immune Kinases. Proc. Natl. Acad. Sci. USA 2019, 116, 496–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, C.V.; Bhaskar, C.K.; Ma, L.-S. A Novel Core Effector Vp1 Promotes Fungal Colonization and Virulence of Ustilago maydis. J. Fungi 2021, 7, 589. [Google Scholar] [CrossRef] [PubMed]
- Snelders, N.C.; Kettles, G.J.; Rudd, J.J.; Thomma, B.P.H.J. Plant Pathogen Effector Proteins as Manipulators of Host Microbiomes? Mol. Plant Pathol. 2018, 19, 257–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, P.J.G.M.; Mehrabi, R.; Van den Burg, H.A.; Stergiopoulos, I. Fungal Effector Proteins: Past, Present and Future. Mol. Plant Pathol. 2009, 10, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Tariqjaveed, M.; Mateen, A.; Wang, S.; Qiu, S.; Zheng, X.; Zhang, J.; Bhadauria, V.; Sun, W. Versatile Effectors of Phytopathogenic Fungi Target Host Immunity. J. Integr. Plant Biol. 2021, 63, 1856–1873. [Google Scholar] [CrossRef]
- He, Q.; McLellan, H.; Boevink, P.C.; Birch, P.R.J. All Roads Lead to Susceptibility: The Many Modes of Action of Fungal and Oomycete Intracellular Effectors. Plant Commun. 2020, 1, 100050. [Google Scholar] [CrossRef]
- Flor, H.H. Inheritance of Pathogenicity in Melampsora Lini. Phytopathology 1942, 32, 653–669. [Google Scholar]
- De Wit, P.J.G.M. Apoplastic Fungal Effectors in Historic Perspective; a Personal View. New Phytol. 2016, 212, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Van Schie, C.C.N.; Takken, F.L.W. Susceptibility Genes 101: How to Be a Good Host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323. [Google Scholar] [CrossRef] [Green Version]
- Kapos, P.; Devendrakumar, K.T.; Li, X. Plant NLRs: From Discovery to Application. Plant Sci. 2019, 279, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Keller, H.; Boyer, L.; Abad, P. Disease Susceptibility in the Zig-Zag Model of Host–Microbe Interactions: Only a Consequence of Immune Suppression? Mol. Plant Pathol. 2016, 17, 475–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thordal-Christensen, H. A Holistic View on Plant Effector-Triggered Immunity Presented as an Iceberg Model. Cell. Mol. Life Sci. 2020, 77, 3963–3976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooney, H.C.E.; van’t Klooster, J.W.; van der Hoorn, R.A.L.; Joosten, M.H.A.J.; Jones, J.D.G.; de Wit, P.J.G.M. Cladosporium Avr2 Inhibits Tomato Rcr3 Protease Required for Cf-2-Dependent Disease Resistance. Science 2005, 308, 1783–1786. [Google Scholar] [CrossRef] [Green Version]
- Van Esse, H.P.; Bolton, M.D.; Stergiopoulos, I.; de Wit, P.J.G.M.; Thomma, B.P.H.J. The Chitin-Binding Cladosporium fulvum Effector Protein Avr4 Is a Virulence Factor. Mol. Plant. Microbe Interact. 2007, 20, 1092–1101. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, K.; Fujisaki, K.; Shimizu, M.; Takeda, T.; Saitoh, H.; Hirabuchi, A.; Hiraka, Y.; Białas, A.; Langner, T.; Kellner, R.; et al. The Blast Pathogen Effector AVR-Pik Binds and Stabilizes Rice Heavy Metal-Associated (HMA) Proteins to Co-Opt Their Function in Immunity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Plissonneau, C.; Blaise, F.; Ollivier, B.; Leflon, M.; Carpezat, J.; Rouxel, T.; Balesdent, M.-H. Unusual Evolutionary Mechanisms to Escape Effector-Triggered Immunity in the Fungal Phytopathogen Leptosphaeria maculans. Mol. Ecol. 2017, 26, 2183–2198. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant Pathogens and Integrated Defence Responses to Infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Van der Hoorn, R.A.L.; Kamoun, S. From Guard to Decoy: A New Model for Perception of Plant Pathogen Effectors. Plant Cell 2008, 20, 2009–2017. [Google Scholar] [CrossRef] [Green Version]
- Cesari, S.; Bernoux, M.; Moncuquet, P.; Kroj, T.; Dodds, P.N. A Novel Conserved Mechanism for Plant NLR Protein Pairs: The ‘Integrated Decoy’ Hypothesis. Front. Plant Sci. 2014, 5, 00606. [Google Scholar] [CrossRef] [Green Version]
- Kroj, T.; Chanclud, E.; Michel-Romiti, C.; Grand, X.; Morel, J.-B. Integration of Decoy Domains Derived from Protein Targets of Pathogen Effectors into Plant Immune Receptors Is Widespread. New Phytol. 2016, 210, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Jubic, L.M.; Saile, S.; Furzer, O.J.; El Kasmi, F.; Dangl, J.L. Help Wanted: Helper NLRs and Plant Immune Responses. Curr. Opin. Plant Biol. 2019, 50, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.-M.; Iswanto, A.B.B.; Son, G.H.; Kim, S.H. Recent Advances in Effector-Triggered Immunity in Plants: New Pieces in the Puzzle Create a Different Paradigm. Int. J. Mol. Sci. 2021, 22, 4709. [Google Scholar] [CrossRef] [PubMed]
- Win, J.; Chaparro-Garcia, A.; Belhaj, K.; Saunders, D.G.O.; Yoshida, K.; Dong, S.; Schornack, S.; Zipfel, C.; Robatzek, S.; Hogenhout, S.A.; et al. Effector Biology of Plant-Associated Organisms: Concepts and Perspectives. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 235–247. [Google Scholar] [CrossRef]
- Madina, M.H.; Rahman, M.S.; Huang, X.; Zhang, Y.; Zheng, H.; Germain, H. A Poplar Rust Effector Protein Associates with Protein Disulfide Isomerase and Enhances Plant Susceptibility. Biology 2020, 9, 294. [Google Scholar] [CrossRef]
- Cook, D.E.; Mesarich, C.H.; Thomma, B.P.H.J. Understanding Plant Immunity as a Surveillance System to Detect Invasion. Annu. Rev. Phytopathol. 2015, 53, 541–563. [Google Scholar] [CrossRef]
- Tian, H.; Wu, Z.; Chen, S.; Ao, K.; Huang, W.; Yaghmaiean, H.; Sun, T.; Xu, F.; Zhang, Y.; Wang, S.; et al. Activation of TIR Signalling Boosts Pattern-Triggered Immunity. Nature 2021, 598, 500–503. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual Potentiation of Plant Immunity by Cell-Surface and Intracellular Receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef]
- Carreón-Anguiano, K.G.; Islas-Flores, I.; Vega-Arreguín, J.; Sáenz-Carbonell, L.; Canto-Canché, B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules 2020, 10, 712. [Google Scholar] [CrossRef]
- Saunders, D.G.O.; Win, J.; Cano, L.M.; Szabo, L.J.; Kamoun, S.; Raffaele, S. Using Hierarchical Clustering of Secreted Protein Families to Classify and Rank Candidate Effectors of Rust Fungi. PLoS ONE 2012, 7, e29847. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Kamoun, S. Genome Evolution in Filamentous Plant Pathogens: Why Bigger Can Be Better. Nat. Rev. Microbiol. 2012, 10, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.A.B.; Rozano, L.; Debler, J.W.; Mancera, R.L.; Moolhuijzen, P.M.; Hane, J.K. An Automated and Combinative Method for the Predictive Ranking of Candidate Effector Proteins of Fungal Plant Pathogens. Sci. Rep. 2021, 11, 19731. [Google Scholar] [CrossRef] [PubMed]
- Nur, M.; Wood, K.; Michelmore, R. EffectorO: Motif-Independent Prediction of Effectors in Oomycete Genomes Using Machine Learning and Lineage Specificity. BioRxiv 2021. [Google Scholar] [CrossRef]
- Sperschneider, J.; Dodds, P.N. EffectorP 3.0: Prediction of Apoplastic and Cytoplasmic Effectors in Fungi and Oomycetes. Mol. Plant-Microbe Interact. 2021, 35, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting Sequence Signals in Targeting Peptides Using Deep Learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Sperschneider, J.; Catanzariti, A.-M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular Localization Prediction of Both Plant and Effector Proteins in the Plant Cell. Sci. Rep. 2017, 7, 44598. [Google Scholar] [CrossRef] [Green Version]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.; Cuzick, A.; Seager, J.; Wood, V.; Rutherford, K.; Venkatesh, S.Y.; De Silva, N.; Martinez, M.C.; Pedro, H.; Yates, A.D.; et al. PHI-Base: The Pathogen–Host Interactions Database. Nucleic Acids Res. 2020, 48, D613–D620. [Google Scholar] [CrossRef]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting Fungal Effector Proteins from Secretomes Using Machine Learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved Prediction of Fungal Effector Proteins from Secretomes with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dölfors, F.; Holmquist, L.; Dixelius, C.; Tzelepis, G. A LysM Effector Protein from the Basidiomycete Rhizoctonia solani Contributes to Virulence through Suppression of Chitin-Triggered Immunity. Mol. Genet. Genom. 2019, 294, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Yarden, O.; Hadar, Y. Seeking the Roles for Fungal Small-Secreted Proteins in Affecting Saprophytic Lifestyles. Front. Microbiol. 2020, 11, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhnert, H.U.; Fudal, I.; Dioh, W.; Tharreau, D.; Notteghem, J.-L.; Lebrun, M.-H. A Putative Polyketide Synthase/Peptide Synthetase from Magnaporthe grisea Signals Pathogen Attack to Resistant Rice. Plant Cell 2004, 16, 2499–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Song, T.; Zhang, X.; Yuan, H.; Su, L.; Li, W.; Xu, J.; Liu, S.; Chen, L.; Chen, T.; et al. Unconventionally Secreted Effectors of Two Filamentous Pathogens Target Plant Salicylate Biosynthesis. Nat. Commun. 2014, 5, 4686. [Google Scholar] [CrossRef] [Green Version]
- Gout, L.; Fudal, I.; Kuhn, M.-L.; Blaise, F.; Eckert, M.; Cattolico, L.; Balesdent, M.-H.; Rouxel, T. Lost in the Middle of Nowhere: The AvrLm1 Avirulence Gene of the Dothideomycete Leptosphaeria maculans. Mol. Microbiol. 2006, 60, 67–80. [Google Scholar] [CrossRef]
- Pazzagli, L.; Seidl-Seiboth, V.; Barsottini, M.; Vargas, W.A.; Scala, A.; Mukherjee, P.K. Cerato-Platanins: Elicitors and Effectors. Plant Sci. 2014, 228, 79–87. [Google Scholar] [CrossRef]
- Zhao, S.; Shang, X.; Bi, W.; Yu, X.; Liu, D.; Kang, Z.; Wang, X.; Wang, X. Genome-Wide Identification of Effector Candidates With Conserved Motifs From the Wheat Leaf Rust Fungus Puccinia triticina. Front. Microbiol. 2020, 11, 1188. [Google Scholar] [CrossRef]
- Jones, D.A.B.; Moolhuijzen, P.M.; Hane, J.K. Remote Homology Clustering Identifies Lowly Conserved Families of Effector Proteins in Plant-Pathogenic Fungi. Microb. Genomics 2021, 7, 000637. [Google Scholar] [CrossRef]
- Bertazzoni, S.; Williams, A.H.; Jones, D.A.; Syme, R.A.; Tan, K.-C.; Hane, J.K. Accessories Make the Outfit: Accessory Chromosomes and Other Dispensable DNA Regions in Plant-Pathogenic Fungi. Mol. Plant-Microbe Interact. 2018, 31, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Raffaele, S.; Kamoun, S. The Two-Speed Genomes of Filamentous Pathogens: Waltz with Plants. Curr. Opin. Genet. Dev. 2015, 35, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Win, J.; Cano, L.M.; Kamoun, S. Analyses of Genome Architecture and Gene Expression Reveal Novel Candidate Virulence Factors in the Secretome of Phytophthora infestans. BMC Genom. 2010, 11, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croll, D.; McDonald, B.A. The Accessory Genome as a Cradle for Adaptive Evolution in Pathogens. PLoS Pathog. 2012, 8, e1002608. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vallet, A.; Fouché, S.; Fudal, I.; Hartmann, F.E.; Soyer, J.L.; Tellier, A.; Croll, D. The Genome Biology of Effector Gene Evolution in Filamentous Plant Pathogens. Annu. Rev. Phytopathol. 2018, 56, 21–40. [Google Scholar] [CrossRef]
- Faino, L.; Seidl, M.F.; Shi-Kunne, X.; Pauper, M.; van den Berg, G.C.M.; Wittenberg, A.H.J.; Thomma, B.P.H.J. Transposons Passively and Actively Contribute to Evolution of the Two-Speed Genome of a Fungal Pathogen. Genome Res. 2016, 26, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Rouxel, T.; Balesdent, M.-H. Life, Death and Rebirth of Avirulence Effectors in a Fungal Pathogen of Brassica Crops, Leptosphaeria maculans. New Phytol. 2017, 214, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Plaumann, P.-L.; Schmidpeter, J.; Dahl, M.; Taher, L.; Koch, C. A Dispensable Chromosome Is Required for Virulence in the Hemibiotrophic Plant Pathogen Colletotrichum higginsianum. Front. Microbiol. 2018, 9, 1005. [Google Scholar] [CrossRef] [Green Version]
- Witte, T.E.; Villeneuve, N.; Boddy, C.N.; Overy, D.P. Accessory Chromosome-Acquired Secondary Metabolism in Plant Pathogenic Fungi: The Evolution of Biotrophs Into Host-Specific Pathogens. Front. Microbiol. 2021, 12, 664276. [Google Scholar] [CrossRef]
- Frantzeskakis, L.; Kracher, B.; Kusch, S.; Yoshikawa-Maekawa, M.; Bauer, S.; Pedersen, C.; Spanu, P.D.; Maekawa, T.; Schulze-Lefert, P.; Panstruga, R. Signatures of Host Specialization and a Recent Transposable Element Burst in the Dynamic One-Speed Genome of the Fungal Barley Powdery Mildew Pathogen. BMC Genom. 2018, 19, 381. [Google Scholar] [CrossRef] [Green Version]
- Stam, R.; Münsterkötter, M.; Pophaly, S.D.; Fokkens, L.; Sghyer, H.; Güldener, U.; Hückelhoven, R.; Hess, M. A New Reference Genome Shows the One-Speed Genome Structure of the Barley Pathogen Ramularia collo-cygni. Genome Biol. Evol. 2018, 10, 3243–3249. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Stam, R.; Cano, L.M.; Song, J.; Sklenar, J.; Yoshida, K.; Bozkurt, T.O.; Oliva, R.; Liu, Z.; Tian, M.; et al. Effector Specialization in a Lineage of the Irish Potato Famine Pathogen. Science 2014, 343, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Chavarro-Carrero, E.A.; Vermeulen, J.P.; Torres, D.E.; Usami, T.; Schouten, H.J.; Bai, Y.; Seidl, M.F.; Thomma, B.P.H.J. Comparative Genomics Reveals the in Planta-Secreted Verticillium dahliae Av2 Effector Protein Recognized in Tomato Plants That Carry the V2 Resistance Locus. Environ. Microbiol. 2021, 23, 1941–1958. [Google Scholar] [CrossRef] [PubMed]
- Chepsergon, J.; Motaung, T.E.; Moleleki, L.N. “Core” RxLR Effectors in Phytopathogenic Oomycetes: A Promising Way to Breeding for Durable Resistance in Plants? Virulence 2021, 12, 1921–1935. [Google Scholar] [CrossRef]
- Depotter, J.R.L.; Doehlemann, G. Target the Core: Durable Plant Resistance against Filamentous Plant Pathogens through Effector Recognition. Pest Manag. Sci. 2020, 76, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Hemetsberger, C.; Mueller, A.N.; Matei, A.; Herrberger, C.; Hensel, G.; Kumlehn, J.; Mishra, B.; Sharma, R.; Thines, M.; Hückelhoven, R.; et al. The Fungal Core Effector Pep1 Is Conserved across Smuts of Dicots and Monocots. New Phytol. 2015, 206, 1116–1126. [Google Scholar] [CrossRef]
- Seitner, D.; Uhse, S.; Gallei, M.; Djamei, A. The Core Effector Cce1 Is Required for Early Infection of Maize by Ustilago maydis. Mol. Plant Pathol. 2018, 19, 2277–2287. [Google Scholar] [CrossRef] [Green Version]
- Lanver, D.; Tollot, M.; Schweizer, G.; Lo Presti, L.; Reissmann, S.; Ma, L.-S.; Schuster, M.; Tanaka, S.; Liang, L.; Ludwig, N.; et al. Ustilago maydis Effectors and Their Impact on Virulence. Nat. Rev. Microbiol. 2017, 15, 409–421. [Google Scholar] [CrossRef]
- Vargas, W.A.; Sanz-Martín, J.M.; Rech, G.E.; Armijos-Jaramillo, V.D.; Rivera, L.P.; Echeverria, M.M.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. A Fungal Effector With Host Nuclear Localization and DNA-Binding Properties Is Required for Maize Anthracnose Development. Mol. Plant-Microbe Interact. 2016, 29, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Miao, J.; Shen, D.; Dou, D. Proteinaceous Effector Discovery and Characterization in Plant Pathogenic Colletotrichum Fungi. Front. Microbiol. 2022, 13, 1353–1376. [Google Scholar] [CrossRef]
- Wang, M.; Ji, Z.; Yan, H.; Xu, J.; Zhao, X.; Zhou, Z. Effector Sntf2 Interacted with Chloroplast-Related Protein Mdycf39 Promoting the Colonization of Colletotrichum gloeosporioides in Apple Leaf. Int. J. Mol. Sci. 2022, 23, 6379. [Google Scholar] [CrossRef]
- Tsushima, A.; Narusaka, M.; Gan, P.; Kumakura, N.; Hiroyama, R.; Kato, N.; Takahashi, S.; Takano, Y.; Narusaka, Y.; Shirasu, K. The Conserved Colletotrichum Spp. Effector Candidate CEC3 Induces Nuclear Expansion and Cell Death in Plants. Front. Microbiol. 2021, 12, 682155. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; van den Burg, H.A.; Ökmen, B.; Beenen, H.G.; van Liere, S.; Kema, G.H.J.; de Wit, P.J.G.M. Tomato Cf Resistance Proteins Mediate Recognition of Cognate Homologous Effectors from Fungi Pathogenic on Dicots and Monocots. Proc. Natl. Acad. Sci. USA 2010, 107, 7610–7615. [Google Scholar] [CrossRef] [Green Version]
- Mesarich, C.H.; Ökmen, B.; Rovenich, H.; Griffiths, S.A.; Wang, C.; Karimi Jashni, M.; Mihajlovski, A.; Collemare, J.; Hunziker, L.; Deng, C.H.; et al. Specific Hypersensitive Response-Associated Recognition of New Apoplastic Effectors from Cladosporium fulvum in Wild Tomato. Mol. Plant-Microbe Interact. 2018, 31, 145–162. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.C.; Chen, L.-H.; Hurlburt, N.; Salvucci, A.; Schwessinger, B.; Fisher, A.J.; Stergiopoulos, I. Structural Analysis of an Avr4 Effector Ortholog Offers Insight into Chitin Binding and Recognition by the Cf-4 Receptor. Plant Cell 2016, 28, 1945–1965. [Google Scholar] [CrossRef] [Green Version]
- Santos Rezende, J.; Zivanovic, M.; Costa de Novaes, M.I.; Chen, Z.-Y. The AVR4 Effector Is Involved in Cercosporin Biosynthesis and Likely Affects the Virulence of Cercospora cf. flagellaris on Soybean. Mol. Plant Pathol. 2020, 21, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-H.; Kračun, S.K.; Nissen, K.S.; Mravec, J.; Jørgensen, B.; Labavitch, J.; Stergiopoulos, I. A Diverse Member of the Fungal Avr4 Effector Family Interacts with De-Esterified Pectin in Plant Cell Walls to Disrupt Their Integrity. Sci. Adv. 2021, 7, eabe0809. [Google Scholar] [CrossRef]
- Bolton, M.D.; van Esse, H.P.; Vossen, J.H.; de Jonge, R.; Stergiopoulos, I.; Stulemeijer, I.J.E.; van den Berg, G.C.M.; Borrás-Hidalgo, O.; Dekker, H.L.; de Koster, C.G.; et al. The Novel Cladosporium fulvum Lysin Motif Effector Ecp6 Is a Virulence Factor with Orthologues in Other Fungal Species. Mol. Microbiol. 2008, 69, 119–136. [Google Scholar] [CrossRef]
- De Jonge, R.; van Esse, H.P.; Kombrink, A.; Shinya, T.; Desaki, Y.; Bours, R.; van der Krol, S.; Shibuya, N.; Joosten, M.H.A.J.; Thomma, B.P.H.J. Conserved Fungal LysM Effector Ecp6 Prevents Chitin-Triggered Immunity in Plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]
- Spanu, P.D. Cereal Immunity against Powdery Mildews Targets RNase-Like Proteins Associated with Haustoria (RALPH) Effectors Evolved from a Common Ancestral Gene. New Phytol. 2017, 213, 969–971. [Google Scholar] [CrossRef] [Green Version]
- De Guillen, K.; Ortiz-Vallejo, D.; Gracy, J.; Fournier, E.; Kroj, T.; Padilla, A. Structure Analysis Uncovers a Highly Diverse but Structurally Conserved Effector Family in Phytopathogenic Fungi. PLoS Pathog. 2015, 11, e1005228. [Google Scholar] [CrossRef]
- Lazar, N.; Mesarich, C.H.; Petit-Houdenot, Y.; Talbi, N.; de la Sierra-Gallay, I.L.; Zélie, E.; Blondeau, K.; Gracy, J.; Ollivier, B.; Blaise, F.; et al. A New Family of Structurally Conserved Fungal Effectors Displays Epistatic Interactions with Plant Resistance Proteins. PLoS Pathog. 2022, 18, e1010664. [Google Scholar] [CrossRef]
- Moolhuijzen, P.M.; See, P.T.; Oliver, R.P.; Moffat, C.S. Genomic Distribution of a Novel Pyrenophora tritici-repentis ToxA Insertion Element. PLoS ONE 2018, 13, e0206586. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Garcés, D.; Camborde, L.; Pel, M.J.C.; Jauneau, A.; Martinez, Y.; Néant, I.; Leclerc, C.; Moreau, M.; Dumas, B.; Gaulin, E. CRN13 Candidate Effectors from Plant and Animal Eukaryotic Pathogens Are DNA-Binding Proteins Which Trigger Host DNA Damage Response. New Phytol. 2016, 210, 602–617. [Google Scholar] [CrossRef] [Green Version]
- Frías, M.; Brito, N.; González, M.; González, C. The Phytotoxic Activity of the Cerato-Platanin BcSpl1 Resides in a Two-Peptide Motif on the Protein Surface. Mol. Plant Pathol. 2014, 15, 342–351. [Google Scholar] [CrossRef]
- Frías, M.; González, C.; Brito, N. BcSpl1, a Cerato-Platanin Family Protein, Contributes to Botrytis cinerea Virulence and Elicits the Hypersensitive Response in the Host. New Phytol. 2011, 192, 483–495. [Google Scholar] [CrossRef]
- Schouten, A.; Baarlen, P.V.; Kan, J.A.L.V. Phytotoxic Nep1-like Proteins from the Necrotrophic Fungus Botrytis cinerea Associate with Membranes and the Nucleus of Plant Cells. New Phytol. 2008, 177, 493–505. [Google Scholar] [CrossRef]
- Bae, H.; Kim, M.S.; Sicher, R.C.; Bae, H.-J.; Bailey, B.A. Necrosis- and Ethylene-Inducing Peptide from Fusarium oxysporum Induces a Complex Cascade of Transcripts Associated with Signal Transduction and Cell Death in Arabidopsis. Plant Physiol. 2006, 141, 1056–1067. [Google Scholar] [CrossRef] [Green Version]
- Bailey, B.A.; Jennings, J.C.; Anderson, J.D. The 24-KDa Protein from Fusarium oxysporum f.Sp. erythroxyli: Occurrence in Related Fungi and the Effect of Growth Medium on Its Production. Can. J. Microbiol. 1997, 43, 45–55. [Google Scholar] [CrossRef]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; Ver Loren van Themaat, E.; van Themaat, E.V.L.; van der Does, H.C.; Hacquard, S.; Stüber, K.; Will, I.; et al. Sequential Delivery of Host-Induced Virulence Effectors by Appressoria and Intracellular Hyphae of the Phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Joosten, M.H.; Vogelsang, R.; Cozijnsen, T.J.; Verberne, M.C.; De Wit, P.J. The Biotrophic Fungus Cladosporium fulvum Circumvents Cf-4-Mediated Resistance by Producing Unstable AVR4 Elicitors. Plant Cell 1997, 9, 367–379. [Google Scholar] [CrossRef]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis Effector Pep1 Suppresses Plant Immunity by Inhibition of Host Peroxidase Activity. PLoS Pathog. 2012, 8, e1002684. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, H.; Fujisawa, S.; Mitsuoka, C.; Ito, A.; Hirabuchi, A.; Ikeda, K.; Irieda, H.; Yoshino, K.; Yoshida, K.; Matsumura, H.; et al. Large-Scale Gene Disruption in Magnaporthe oryzae Identifies MC69, a Secreted Protein Required for Infection by Monocot and Dicot Fungal Pathogens. PLoS Pathog. 2012, 8, e1002711. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.G.; Casady, M.S.; Fee, R.A.; Vaughan, M.M.; Deb, D.; Fedkenheuer, K.; Huffaker, A.; Schmelz, E.A.; Tyler, B.M.; McDowell, J.M. Homologous Rxlr Effectors from Hyaloperonospora arabidopsidis and Phytophthora sojae Suppress Immunity in Distantly Related Plants. Plant J. Cell Mol. Biol. 2012, 72, 882–893. [Google Scholar] [CrossRef]
- Zhang, J.; Shao, F.; Li, Y.; Cui, H.; Chen, L.; Li, H.; Zou, Y.; Long, C.; Lan, L.; Chai, J.; et al. A Pseudomonas syringae Effector Inactivates MAPKs to Suppress PAMP-Induced Immunity in Plants. Cell Host Microbe 2007, 1, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Kelley, B.S.; Lee, S.-J.; Damasceno, C.M.B.; Chakravarthy, S.; Kim, B.-D.; Martin, G.B.; Rose, J.K.C. A Secreted Effector Protein (SNE1) from Phytophthora infestans Is a Broadly Acting Suppressor of Programmed Cell Death. Plant J. Cell Mol. Biol. 2010, 62, 357–366. [Google Scholar] [CrossRef]
- Catanzariti, A.-M.; Dodds, P.N.; Ve, T.; Kobe, B.; Ellis, J.G.; Staskawicz, B.J. The AvrM Effector from Flax Rust Has a Structured C-Terminal Domain and Interacts Directly with the M Resistance Protein. Mol. Plant-Microbe Interact. 2010, 23, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Ve, T.; Williams, S.J.; Catanzariti, A.-M.; Rafiqi, M.; Rahman, M.; Ellis, J.G.; Hardham, A.R.; Jones, D.A.; Anderson, P.A.; Dodds, P.N.; et al. Structures of the Flax-Rust Effector AvrM Reveal Insights into the Molecular Basis of Plant-Cell Entry and Effector-Triggered Immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 17594–17599. [Google Scholar] [CrossRef] [Green Version]
- Cesari, S.; Thilliez, G.; Ribot, C.; Chalvon, V.; Michel, C.; Jauneau, A.; Rivas, S.; Alaux, L.; Kanzaki, H.; Okuyama, Y.; et al. The Rice Resistance Protein Pair RGA4/RGA5 Recognizes the Magnaporthe oryzae Effectors AVR-Pia and AVR1-CO39 by Direct Binding. Plant Cell 2013, 25, 1463–1481. [Google Scholar] [CrossRef] [Green Version]
- Hammond-Kosack, K.E.; Tang, S.; Harrison, K.; Jones, J.D.G. The Tomato Cf-9 Disease Resistance Gene Functions in Tobacco and Potato to Confer Responsiveness to the Fungal Avirulence Gene Product Avr9. Plant Cell 1998, 10, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, H.; Yoshida, K.; Saitoh, H.; Fujisaki, K.; Hirabuchi, A.; Alaux, L.; Fournier, E.; Tharreau, D.; Terauchi, R. Arms Race Co-Evolution of Magnaporthe Oryzae AVR-Pik and Rice Pik Genes Driven by Their Physical Interactions. Plant J. 2012, 72, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction Transcriptome Analysis Identifies Magnaporthe oryzae BAS1-4 as Biotrophy-Associated Secreted Proteins in Rice Blast Disease. Plant Cell 2009, 21, 1273–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, L.; Wang, Y.; Wang, C.; Yan, J.; Liu, Y.; Wang, C.; Li, C. Overexpression of BAS1 in Rice Blast Fungus Can Promote Blast Fungus Growth, Sporulation and Virulence in Planta. Saudi J. Biol. Sci. 2017, 24, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Pennington, H.G.; Jones, R.; Kwon, S.; Bonciani, G.; Thieron, H.; Chandler, T.; Luong, P.; Morgan, S.N.; Przydacz, M.; Bozkurt, T.; et al. The Fungal Ribonuclease-like Effector Protein CSEP0064/BEC1054 Represses Plant Immunity and Interferes with Degradation of Host Ribosomal RNA. PLoS Pathog. 2019, 15, 1007620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos-Canul, Y.Y.; Canto-Canché, B.; Berezovski, M.V.; Mironov, G.; Loyola-Vargas, V.M.; Barba de Rosa, A.P.; Tzec-Simá, M.; Brito-Argáez, L.; Carrillo-Pech, M.; Grijalva-Arango, R.; et al. The Cell Wall Proteome from Two Strains of Pseudocercospora fijiensis with Differences in Virulence. World J. Microbiol. Biotechnol. 2019, 35, 105. [Google Scholar] [CrossRef]
- Sucheta Tripathy, S.B. A Tale of Effectors; Their Secretory Mechanisms and Computational Discovery in Pathogenic, Non-Pathogenic and Commensal Microbes. Mol. Biol. 2014, 03, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors Involved in Fungal–Fungal Interaction Lead to a Rare Phenomenon of Hyperbiotrophy in the Tritrophic System Biocontrol Agent–Powdery Mildew–Plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Rabouille, C. Pathways of Unconventional Protein Secretion. Trends Cell Biol. 2017, 27, 230–240. [Google Scholar] [CrossRef]
- Boevink, P.C.; Wang, X.; McLellan, H.; He, Q.; Naqvi, S.; Armstrong, M.R.; Zhang, W.; Hein, I.; Gilroy, E.M.; Tian, Z.; et al. A Phytophthora infestans RXLR Effector Targets Plant PP1c Isoforms That Promote Late Blight Disease. Nat. Commun. 2016, 7, 10311. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xu, J.-R. Effectors and Effector Delivery in Magnaporthe oryzae. PLoS Pathog. 2014, 10, e1003826. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, M.C.; Dagdas, Y.F.; Gupta, Y.K.; Mentlak, T.A.; Yi, M.; Martinez-Rocha, A.L.; Saitoh, H.; Terauchi, R.; Talbot, N.J.; Valent, B. Two Distinct Secretion Systems Facilitate Tissue Invasion by the Rice Blast Fungus Magnaporthe oryzae. Nat. Commun. 2013, 4, 1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, S.D.; Gu, B.; Capelluto, D.G.S.; Dou, D.; Feldman, E.; Rumore, A.; Arredondo, F.D.; Hanlon, R.; Fudal, I.; Rouxel, T.; et al. External Lipid PI3P Mediates Entry of Eukaryotic Pathogen Effectors into Plant and Animal Host Cells. Cell 2010, 142, 284–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonah, H.; Deshmukh, R.K.; Bélanger, R.R. Computational Prediction of Effector Proteins in Fungi: Opportunities and Challenges. Front. Plant Sci. 2016, 7, 00126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petre, B.; Lorrain, C.; Saunders, D.G.O.; Win, J.; Sklenar, J.; Duplessis, S.; Kamoun, S. Rust Fungal Effectors Mimic Host Transit Peptides to Translocate into Chloroplasts. Cell. Microbiol. 2016, 18, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Robin, G.P.; Kleemann, J.; Neumann, U.; Cabre, L.; Dallery, J.-F.; Lapalu, N.; O’Connell, R.J. Subcellular Localization Screening of Colletotrichum higginsianum Effector Candidates Identifies Fungal Proteins Targeted to Plant Peroxisomes, Golgi Bodies, and Microtubules. Front. Plant Sci. 2018, 9, 00562. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lan, X.; Song, S.; Yin, L.; Dry, I.B.; Qu, J.; Xiang, J.; Lu, J. In Planta Functional Analysis and Subcellular Localization of the Oomycete Pathogen Plasmopara viticola Candidate RXLR Effector Repertoire. Front. Plant Sci. 2018, 9, 00286. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Seto, D.; Subramaniam, R.; Desveaux, D. Oh, the Places They’ll Go! A Survey of Phytopathogen Effectors and Their Host Targets. Plant J. 2018, 93, 651–663. [Google Scholar] [CrossRef]
- Scheibner, F.; Marillonnet, S.; Büttner, D. The Tal Effector Avrbs3 from Xanthomonas campestris pv. vesicatoria Contains Multiple Export Signals and Can Enter Plant Cells in the Absence of the Type III Secretion Translocon. Front. Microbiol. 2017, 8, 02180. [Google Scholar] [CrossRef]
- Rodríguez-Puerto, C.; Chakraborty, R.; Singh, R.; Rocha-Loyola, P.; Rojas, C.M. The Pseudomonas syringae Type III Effector HopG1 Triggers Necrotic Cell Death That Is Attenuated by AtNHR2B. Sci. Rep. 2022, 12, 5388. [Google Scholar] [CrossRef]
- Block, A.; Guo, M.; Li, G.; Elowsky, C.; Clemente, T.E.; Alfano, J.R. The Pseudomonas syringae Type III Effector HopG1 Targets Mitochondria, Alters Plant Development, and Suppresses Plant Innate Immunity. Cell. Microbiol. 2010, 12, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Halane, M.K.; Kim, S.H.; Spears, B.J.; Garner, C.M.; Rogan, C.J.; Okafor, E.C.; Su, J.; Bhattacharjee, S.; Gassmann, W. The Bacterial Type III-Secreted Protein AvrRps4 Is a Bipartite Effector. PLoS Pathog. 2018, 14, e1006984. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Toruño, T.Y.; Elowsky, C.G.; Zhang, C.; Steinbrenner, J.; Beynon, J.; Alfano, J.R. The Pseudomonas syringae Type III Effector HopD1 Suppresses Effector-Triggered Immunity, Localizes to the Endoplasmic Reticulum, and Targets the Arabidopsis Transcription Factor NTL9. New Phytol. 2014, 201, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, A.; Rutter, W.; Wang, S.; Akhunova, A.; Bolus, S.; Chao, S.; Anderson, N.; De Soto, M.F.; Rouse, M.; Szabo, L.; et al. Variation in the AvrSr35 Gene Determines Sr35 Resistance against Wheat Stem Rust Race Ug99. Science 2017, 358, 1604–1606. [Google Scholar] [CrossRef] [Green Version]
- Manning, V.A.; Hardison, L.K.; Ciuffetti, L.M. Ptr ToxA Interacts with a Chloroplast-Localized Protein. Mol. Plant-Microbe Interact. 2007, 20, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ni, H.; Du, X.; Wang, S.; Ma, X.-W.; Nürnberger, T.; Guo, H.-S.; Hua, C. The Verticillium-Specific Protein VdSCP7 Localizes to the Plant Nucleus and Modulates Immunity to Fungal Infections. New Phytol. 2017, 215, 368–381. [Google Scholar] [CrossRef] [Green Version]
- Ning, N.; Xie, X.; Yu, H.; Mei, J.; Li, Q.; Zuo, S.; Wu, H.; Liu, W.; Li, Z. Plant Peroxisome-Targeting Effector MoPtep1 Is Required for the Virulence of Magnaporthe oryzae. Int. J. Mol. Sci. 2022, 23, 2515. [Google Scholar] [CrossRef]
- Liu, T.; Ye, W.; Ru, Y.; Yang, X.; Gu, B.; Tao, K.; Lu, S.; Dong, S.; Zheng, X.; Shan, W.; et al. Two Host Cytoplasmic Effectors Are Required for Pathogenesis of Phytophthora sojae by Suppression of Host Defenses. Plant Physiol. 2011, 155, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, M.; Shen, D.; Liu, T.; Chen, Y.; Zhou, J.-M.; Dou, D. A Phytophthora sojae Effector PsCRN63 Forms Homo-/Hetero-Dimers to Suppress Plant Immunity via an Inverted Association Manner. Sci. Rep. 2016, 6, 26951. [Google Scholar] [CrossRef] [Green Version]
- Jing, M.; Guo, B.; Li, H.; Yang, B.; Wang, H.; Kong, G.; Zhao, Y.; Xu, H.; Wang, Y.; Ye, W.; et al. A Phytophthora sojae Effector Suppresses Endoplasmic Reticulum Stress-Mediated Immunity by Stabilizing Plant Binding Immunoglobulin Proteins. Nat. Commun. 2016, 7, 11685. [Google Scholar] [CrossRef] [Green Version]
- Caillaud, M.-C.; Piquerez, S.J.M.; Fabro, G.; Steinbrenner, J.; Ishaque, N.; Beynon, J.; Jones, J.D.G. Subcellular Localization of the Hpa RxLR Effector Repertoire Identifies a Tonoplast-Associated Protein HaRxL17 That Confers Enhanced Plant Susceptibility. Plant J. 2012, 69, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Zhong, X.; Shi, Y.; Liu, Z.; Jiang, N.; Liu, J.; Ding, B.; Li, Z.; Kang, H.; Ning, Y.; et al. A Fungal Effector Targets a Heat Shock-Dynamin Protein Complex to Modulate Mitochondrial Dynamics and Reduce Plant Immunity. Sci. Adv. 2020, 6, eabb7719. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Leary, A.Y.; Tumtas, Y.; Savage, Z.; Dagvadorj, B.; Duggan, C.; Yuen, E.L.; Sanguankiattichai, N.; Tan, E.; Khandare, V.; et al. An Oomycete Effector Subverts Host Vesicle Trafficking to Channel Starvation-Induced Autophagy to the Pathogen Interface. eLife 2021, 10, e65285. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yang, J.; Zhang, Q.; Wang, W.; Feng, L.; Zhao, L.; An, B.; Wang, Q.; He, C.; Luo, H. The Effector Protein CgNLP1 of Colletotrichum gloeosporioides Affects Invasion and Disrupts Nuclear Localization of Necrosis-Induced Transcription Factor HbMYB8-Like to Suppress Plant Defense Signaling. Front. Microbiol. 2022, 13, 911479. [Google Scholar] [CrossRef]
- Dagvadorj, B.; Outram, M.A.; Williams, S.J.; Solomon, P.S. The Necrotrophic Effector ToxA from Parastagonospora nodorum Interacts with Wheat NHL Proteins to Facilitate Tsn1-Mediated Necrosis. Plant J. 2022, 110, 407–418. [Google Scholar] [CrossRef]
- Tang, L.; Yang, G.; Ma, M.; Liu, X.; Li, B.; Xie, J.; Fu, Y.; Chen, T.; Yu, Y.; Chen, W.; et al. An Effector of a Necrotrophic Fungal Pathogen Targets the Calcium-Sensing Receptor in Chloroplasts to Inhibit Host Resistance. Mol. Plant Pathol. 2020, 21, 686–701. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shen, M.; Yang, J.; Xing, Y.; Chen, D.; Li, Z.; Zhao, W.; Zhang, Y. Peroxisomal Fission Is Induced during Appressorium Formation and Is Required for Full Virulence of the Rice Blast Fungus. Mol. Plant Pathol. 2016, 18, 222–237. [Google Scholar] [CrossRef]
- Launay, A.; Jolivet, S.; Clément, G.; Zarattini, M.; Dellero, Y.; Le Hir, R.; Jossier, M.; Hodges, M.; Expert, D.; Fagard, M. DspA/E-Triggered Non-Host Resistance against E. amylovora Depends on the Arabidopsis GLYCOLATE OXIDASE 2 Gene. Int. J. Mol. Sci. 2022, 23, 4224. [Google Scholar] [CrossRef]
- Tzelepis, G.; Dölfors, F.; Holmquist, L.; Dixelius, C. Plant Mitochondria and Chloroplasts Are Targeted by the Rhizoctonia solani RsCRP1 Effector. Biochem. Biophys. Res. Commun. 2021, 544, 86–90. [Google Scholar] [CrossRef]
- Nakano, M.; Mukaihara, T. The Type III Effector RipB from Ralstonia solanacearum RS1000 Acts as a Major Avirulence Factor in Nicotiana benthamiana and Other Nicotiana Species. Mol. Plant Pathol. 2019, 20, 1237–1251. [Google Scholar] [CrossRef] [Green Version]
- Washington, E.J.; Mukhtar, M.S.; Finkel, O.M.; Wan, L.; Banfield, M.J.; Kieber, J.J.; Dangl, J.L. Pseudomonas syringae Type III Effector HopAF1 Suppresses Plant Immunity by Targeting Methionine Recycling to Block Ethylene Induction. Proc. Natl. Acad. Sci. USA 2016, 113, E3577–E3586. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, Y.; Wang, J.; Zou, F.; Jia, Y.; Shen, D.; Zhang, Q.; Jing, M.; Dou, D.; Zhang, M. A Phytophthora capsici Virulence Effector Associates with NPR1 and Suppresses Plant Immune Responses. Phytopathol. Res. 2019, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, H.; Jing, M.; Zhu, J.; Guo, B.; Wang, Y.; Lin, Y.; Chen, H.; Kong, L.; Ma, Z.; et al. A Phytophthora Effector Recruits a Host Cytoplasmic Transacetylase into Nuclear Speckles to Enhance Plant Susceptibility. eLife 2018, 7, e40039. [Google Scholar] [CrossRef]
- Ma, L.; Djavaheri, M.; Wang, H.; Larkan, N.J.; Haddadi, P.; Beynon, E.; Gropp, G.; Borhan, M.H. Leptosphaeria maculans Effector Protein AvrLm1 Modulates Plant Immunity by Enhancing MAP Kinase 9 Phosphorylation. iScience 2018, 3, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wang, Y.; Wang, Y.; Du, Y.; Song, C.; Song, P.; Yang, Q.; He, F.; Bai, X.; Huang, L.; et al. Glycine-Serine-Rich Effector PstGSRE4 in Puccinia striiformis f. sp. tritici Inhibits the Activity of Copper Zinc Superoxide Dismutase to Modulate Immunity in Wheat. PLoS Pathog. 2022, 18, e1010702. [Google Scholar] [CrossRef] [PubMed]
- Plett, J.M.; Daguerre, Y.; Wittulsky, S.; Vayssières, A.; Deveau, A.; Melton, S.J.; Kohler, A.; Morrell-Falvey, J.L.; Brun, A.; Veneault-Fourrey, C.; et al. Effector MiSSP7 of the Mutualistic Fungus Laccaria bicolor Stabilizes the Populus JAZ6 Protein and Represses Jasmonic Acid (JA) Responsive Genes. Proc. Natl. Acad. Sci. USA 2014, 111, 8299–8304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondeau, K.; Blaise, F.; Graille, M.; Kale, S.D.; Linglin, J.; Ollivier, B.; Labarde, A.; Lazar, N.; Daverdin, G.; Balesdent, M.-H.; et al. Crystal Structure of the Effector AvrLm4–7 of Leptosphaeria maculans Reveals Insights into Its Translocation into Plant Cells and Recognition by Resistance Proteins. Plant J. 2015, 83, 610–624. [Google Scholar] [CrossRef]
- Song, J.; Win, J.; Tian, M.; Schornack, S.; Kaschani, F.; Ilyas, M.; van der Hoorn, R.A.L.; Kamoun, S. Apoplastic Effectors Secreted by Two Unrelated Eukaryotic Plant Pathogens Target the Tomato Defense Protease Rcr3. Proc. Natl. Acad. Sci. USA 2009, 106, 1654–1659. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, M.S.; Carvunis, A.-R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently Evolved Virulence Effectors Converge onto Hubs in a Plant Immune System Network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Ceulemans, E.; Ibrahim, H.M.M.; De Coninck, B.; Goossens, A. Pathogen Effectors: Exploiting the Promiscuity of Plant Signaling Hubs. Trends Plant Sci. 2021, 26, 780–795. [Google Scholar] [CrossRef]
- Kataria, R.; Kaundal, R. WeCoNET: A Host–Pathogen Interactome Database for Deciphering Crucial Molecular Networks of Wheat-Common Bunt Cross-Talk Mechanisms. Plant Methods 2022, 18, 73. [Google Scholar] [CrossRef] [PubMed]
- Kataria, R.; Kaundal, R. Deciphering the Host–Pathogen Interactome of the Wheat–Common Bunt System: A Step towards Enhanced Resilience in Next Generation Wheat. Int. J. Mol. Sci. 2022, 23, 2589. [Google Scholar] [CrossRef] [PubMed]
- González-Fuente, M.; Carrère, S.; Monachello, D.; Marsella, B.G.; Cazalé, A.-C.; Zischek, C.; Mitra, R.M.; Rezé, N.; Cottret, L.; Mukhtar, M.S.; et al. EffectorK, a Comprehensive Resource to Mine for Ralstonia, Xanthomonas, and Other Published Effector Interactors in the Arabidopsis Proteome. Mol. Plant Pathol. 2020, 21, 1257–1270. [Google Scholar] [CrossRef]
- Crua Asensio, N.; Muñoz Giner, E.; de Groot, N.S.; Torrent Burgas, M. Centrality in the Host-Pathogen Interactome Is Associated with Pathogen Fitness during Infection. Nat. Commun. 2017, 8, 14092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, N.S.; Torrent Burgas, M. Bacteria Use Structural Imperfect Mimicry to Hijack the Host Interactome. PLoS Comput. Biol. 2020, 16, e1008395. [Google Scholar] [CrossRef]
- Mondal, S.I.; Mahmud, Z.; Elahi, M.; Akter, A.; Jewel, N.A.; Muzahidul Islam, M.; Ferdous, S.; Kikuchi, T. Study of Intra-Inter Species Protein-Protein Interactions for Potential Drug Targets Identification and Subsequent Drug Design for Escherichia coli O104:H4 C277-11. Silico Pharmacol. 2016, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Bauters, L.; Stojilković, B.; Gheysen, G. Pathogens Pulling the Strings: Effectors Manipulating Salicylic Acid and Phenylpropanoid Biosynthesis in Plants. Mol. Plant Pathol. 2021, 22, 1436–1448. [Google Scholar] [CrossRef]
- Chen, H.; Chen, J.; Li, M.; Chang, M.; Xu, K.; Shang, Z.; Zhao, Y.; Palmer, I.; Zhang, Y.; McGill, J.; et al. A Bacterial Type III Effector Targets the Master Regulator of Salicylic Acid Signaling, NPR1, to Subvert Plant Immunity. Cell Host Microbe 2017, 22, 777–788.e7. [Google Scholar] [CrossRef] [Green Version]
- Lal, N.K.; Thanasuwat, B.; Huang, P.-J.; Cavanaugh, K.A.; Carter, A.; Michelmore, R.W.; Dinesh-Kumar, S.P. Phytopathogen Effectors Use Multiple Mechanisms to Manipulate Plant Autophagy. Cell Host Microbe 2020, 28, 558–571.e6. [Google Scholar] [CrossRef]
- Kettles, G.J.; Bayon, C.; Sparks, C.A.; Canning, G.; Kanyuka, K.; Rudd, J.J. Characterization of an Antimicrobial and Phytotoxic Ribonuclease Secreted by the Fungal Wheat Pathogen Zymoseptoria tritici. New Phytol. 2018, 217, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, C.; Ramachandran, S.R.; Zhai, Y.; Bu, C.; Pappu, H.R.; Hulbert, S.H. A Novel Fungal Effector from Puccinia graminis Suppressing RNA Silencing and Plant Defense Responses. New Phytol. 2019, 222, 1561–1572. [Google Scholar]
- Cui, C.; Wang, Y.; Liu, J.; Zhao, J.; Sun, P.; Wang, S. A Fungal Pathogen Deploys a Small Silencing RNA That Attenuates Mosquito Immunity and Facilitates Infection. Nat. Commun. 2019, 10, 4298. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, Y.; Song, N.; Zhao, M.; Liu, R.; Feng, H.; Wang, X.; Kang, Z. Puccinia striiformis f. sp. tritici MicroRNA-like RNA 1 (Pst-MilR1), an Important Pathogenicity Factor of Pst, Impairs Wheat Resistance to Pst by Suppressing the Wheat Pathogenesis-Related 2 Gene. New Phytol. 2017, 215, 338–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.-M.; Thomma, B.; Huang, H.-D.; Jin, H. Bidirectional Cross-Kingdom RNAi and Fungal Uptake of External RNAs Confer Plant Protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Gupta, V.K.; Jiang, Y.; Yang, B.; Gong, L.; Zhu, H. Cross-Kingdom Small RNAs among Animals, Plants and Microbes. Cells 2019, 8, 371. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhao, Y.-L.; Zhao, J.-H.; Wang, S.; Jin, Y.; Chen, Z.-Q.; Fang, Y.-Y.; Hua, C.-L.; Ding, S.-W.; Guo, H.-S. Cotton Plants Export MicroRNAs to Inhibit Virulence Gene Expression in a Fungal Pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.-M.; Palmquist, J.; Huang, S.-D.; Jin, H. Plants Send Small RNAs in Extracellular Vesicles to Fungal Pathogen to Silence Virulence Genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zhai, Y.; Feng, L.; Karimi, H.Z.; Rutter, B.D.; Zeng, L.; Choi, D.S.; Zhang, B.; Gu, W.; Chen, X.; et al. A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility. Cell Host Microbe 2019, 25, 153–165.e5. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cui, C.; Wang, G.; Li, Y.; Wang, S. Insects Defend against Fungal Infection by Employing MicroRNAs to Silence Virulence-Related Genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2023802118. [Google Scholar] [CrossRef]
- Zanini, S.; Šečić, E.; Jelonek, L.; Kogel, K.-H. A Bioinformatics Pipeline for the Analysis and Target Prediction of RNA Effectors in Bidirectional Communication During Plant-Microbe Interactions. Front. Plant Sci. 2018, 9, 1212. [Google Scholar] [CrossRef] [Green Version]
- Derbyshire, M.; Mbengue, M.; Barascud, M.; Navaud, O.; Raffaele, S. Small RNAs from the Plant Pathogenic Fungus Sclerotinia sclerotiorum Highlight Host Candidate Genes Associated with Quantitative Disease Resistance. Mol. Plant Pathol. 2019, 20, 1279–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyman, M.C.; Solofoharivelo, M.-C.; Souza-Richards, R.; Stephan, D.; Murray, S.; Burger, J.T. The Use of High-Throughput Small RNA Sequencing Reveals Differentially Expressed MicroRNAs in Response to Aster Yellows Phytoplasma-Infection in Vitis vinifera cv. ‘Chardonnay’. PLoS ONE 2017, 12, e0182629. [Google Scholar] [CrossRef] [PubMed]
- Dunker, F.; Trutzenberg, A.; Rothenpieler, J.S.; Kuhn, S.; Pröls, R.; Schreiber, T.; Tissier, A.; Kemen, A.; Kemen, E.; Hückelhoven, R.; et al. Oomycete Small RNAs Bind to the Plant RNA-Induced Silencing Complex for Virulence. eLife 2020, 9, e56096. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Banerjee, S.; Surana, P.; Liu, M.; Fuerst, G.; Mathioni, S.; Meyers, B.C.; Nettleton, D.; Wise, R.P. Small RNA Discovery in the Interaction between Barley and the Powdery Mildew Pathogen. BMC Genom. 2019, 20, 610. [Google Scholar] [CrossRef] [Green Version]
- Scharf, D.H.; Heinekamp, T.; Brakhage, A.A. Human and Plant Fungal Pathogens: The Role of Secondary Metabolites. PLoS Pathog. 2014, 10, e1003859. [Google Scholar] [CrossRef] [Green Version]
- Devi, R.; Kaur, T.; Guleria, G.; Rana, K.L.; Kour, D.; Yadav, N.; Yadav, A.N.; Saxena, A.K. Chapter 9—Fungal Secondary Metabolites and Their Biotechnological Applications for Human Health. In New and Future Developments in Microbial Biotechnology and Bioengineering; Rastegari, A.A., Yadav, A.N., Yadav, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 147–161. ISBN 978-0-12-820528-0. [Google Scholar]
- Boruta, T. Uncovering the Repertoire of Fungal Secondary Metabolites: From Fleming’s Laboratory to the International Space Station. Bioengineered 2017, 9, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Hassani, M.A.; Oppong-Danquah, E.; Feurtey, A.; Tasdemir, D.; Stukenbrock, E.H. Differential Regulation and Production of Secondary Metabolites among Isolates of the Fungal Wheat Pathogen Zymoseptoria tritici. Appl. Environ. Microbiol. 2022, 88, e02296-21. [Google Scholar] [CrossRef]
- Graham-Taylor, C.; Kamphuis, L.G.; Derbyshire, M.C. A Detailed in Silico Analysis of Secondary Metabolite Biosynthesis Clusters in the Genome of the Broad Host Range Plant Pathogenic Fungus Sclerotinia sclerotiorum. BMC Genom. 2020, 21, 7. [Google Scholar] [CrossRef] [Green Version]
- Muria-Gonzalez, M.J.; Chooi, Y.-H.; Breen, S.; Solomon, P.S. The Past, Present and Future of Secondary Metabolite Research in the Dothideomycetes. Mol. Plant Pathol. 2015, 16, 92–107. [Google Scholar] [CrossRef]
- Rangel, L.I.; Bolton, M.D. The Unsung Roles of Microbial Secondary Metabolite Effectors in the Plant Disease Cacophony. Curr. Opin. Plant Biol. 2022, 68, 102233. [Google Scholar] [CrossRef]
- Collemare, J.; Griffiths, S.; Iida, Y.; Karimi Jashni, M.; Battaglia, E.; Cox, R.J.; de Wit, P.J.G.M. Secondary Metabolism and Biotrophic Lifestyle in the Tomato Pathogen Cladosporium fulvum. PLoS ONE 2014, 9, e85877. [Google Scholar] [CrossRef]
- Baker, S.E.; Kroken, S.; Inderbitzin, P.; Asvarak, T.; Li, B.-Y.; Shi, L.; Yoder, O.C.; Turgeon, B.G. Two Polyketide Synthase-Encoding Genes Are Required for Biosynthesis of the Polyketide Virulence Factor, T-Toxin, by Cochliobolus heterostrophus. Mol. Plant-Microbe Interact. 2006, 19, 139–149. [Google Scholar] [CrossRef]
- Möbius, N.; Hertweck, C. Fungal Phytotoxins as Mediators of Virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.M.; Bender, C.L.; Kunkel, B.N. The Pseudomonas syringae Phytotoxin Coronatine Promotes Virulence by Overcoming Salicylic Acid-Dependent Defences in Arabidopsis thaliana. Mol. Plant Pathol. 2005, 6, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Vergne, E.; Ballini, E.; Marques, S.; Sidi Mammar, B.; Droc, G.; Gaillard, S.; Bourot, S.; DeRose, R.; Tharreau, D.; Nottéghem, J.-L.; et al. Early and Specific Gene Expression Triggered by Rice Resistance Gene Pi33 in Response to Infection by ACE1 Avirulent Blast Fungus. New Phytol. 2007, 174, 159–171. [Google Scholar] [CrossRef]
- Thynne, E.; Mead, O.L.; Chooi, Y.-H.; McDonald, M.C.; Solomon, P.S. Acquisition and Loss of Secondary Metabolites Shaped the Evolutionary Path of Three Emerging Phytopathogens of Wheat. Genome Biol. Evol. 2019, 11, 890–905. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, X.; He, C.; Zhang, Q.-Y.; Zou, X.; Duan, K.; Gao, Q. Novel Fungal Pathogenicity and Leaf Defense Strategies Are Revealed by Simultaneous Transcriptome Analysis of Colletotrichum fructicola and Strawberry Infected by This Fungus. Front. Plant Sci. 2018, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Chaleckis, R.; Meister, I.; Zhang, P.; Wheelock, C.E. Challenges, Progress and Promises of Metabolite Annotation for LC-MS-Based Metabolomics. Curr. Opin. Biotechnol. 2019, 55, 44–50. [Google Scholar] [CrossRef]
- Pinu, F.R.; Goldansaz, S.A.; Jaine, J. Translational Metabolomics: Current Challenges and Future Opportunities. Metabolites 2019, 9, 108. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.E.; Bhatnagar, D.; Ganley, R.J.; Gillman, C.J.; Monahan, B.J.; Seconi, J.M. Dothistroma pini, a Forest Pathogen, Contains Homologs of Aflatoxin Biosynthetic Pathway Genes. Appl. Environ. Microbiol. 2002, 68, 2885–2892. [Google Scholar] [CrossRef] [Green Version]
- Rokas, A.; Mead, M.E.; Steenwyk, J.L.; Raja, H.A.; Oberlies, N.H. Biosynthetic Gene Clusters and the Evolution of Fungal Chemodiversity. Nat. Prod. Rep. 2020, 37, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Schwelm, A.; Bradshaw, R.E. Genetics of Dothistromin Biosynthesis of Dothistroma septosporum: An Update. Toxins 2010, 2, 2680–2698. [Google Scholar] [CrossRef] [PubMed]
- Joosten, M.; de Wit, P. The Tomato-Cladosporium fulvum Interaction: A Versatile Experimental System to Study Plant-Pathogen Interactions. Annu. Rev. Phytopathol. 1999, 37, 335–367. [Google Scholar] [CrossRef] [PubMed]
- De Wit, P.J.G.M. Cladosporium fulvum Effectors: Weapons in the Arms Race with Tomato. Annu. Rev. Phytopathol. 2016, 54, 1–23. [Google Scholar] [CrossRef] [Green Version]
- West, P.V.; Vleeshouwers, V.G. The Phytophthora infestans-Potato Interaction. In Plant Pathogen Interactions; Talbot, N.J., Ed.; Blackwell Scientific Publishers: Hoboken, NJ, USA, 2004. [Google Scholar]
- Yin, J.; Gu, B.; Huang, G.; Tian, Y.; Quan, J.; Lindqvist-Kreuze, H.; Shan, W. Conserved RXLR Effector Genes of Phytophthora infestans Expressed at the Early Stage of Potato Infection Are Suppressive to Host Defense. Front. Plant Sci. 2017, 8, 2155. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.-F.; He, S.Y. Pseudomonas Syringae Pv. Tomato DC3000: A Model Pathogen for Probing Disease Susceptibility and Hormone Signaling in Plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef]
- Morris, C.E.; Lamichhane, J.R.; Nikolić, I.; Stanković, S.; Moury, B. The Overlapping Continuum of Host Range among Strains in the Pseudomonas Syringae Complex. Phytopathol. Res. 2019, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Chen, X.; Kemppainen, M.; Pardo, A.G.; Veneault-Fourrey, C.; Kohler, A.; Martin, F.M. The Small Secreted Effector Protein MiSSP7.6 of Laccaria bicolor Is Required for the Establishment of Ectomycorrhizal Symbiosis. Environ. Microbiol. 2020, 22, 1435–1446. [Google Scholar] [CrossRef]
- Constantin, M.E.; Fokkens, L.; de Sain, M.; Takken, F.L.W.; Rep, M. Number of Candidate Effector Genes in Accessory Genomes Differentiates Pathogenic From Endophytic Fusarium oxysporum Strains. Front. Plant Sci. 2021, 12, 761740. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jensen, B.; Jørgensen, H.J. Fungal Endophytes in Plants and Their Relationship to Plant Disease. Curr. Opin. Microbiol. 2022, 69, 102177. [Google Scholar] [CrossRef]
- Rovenich, H.; Boshoven, J.C.; Thomma, B.P.H.J. Filamentous Pathogen Effector Functions: Of Pathogens, Hosts and Microbiomes. Curr. Opin. Plant Biol. 2014, 20, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, C.-M. Against Friend and Foe: Type 6 Effectors in Plant-Associated Bacteria. J. Microbiol. Seoul Korea 2015, 53, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Trunk, K.; Peltier, J.; Liu, Y.-C.; Dill, B.D.; Walker, L.; Gow, N.A.R.; Stark, M.J.R.; Quinn, J.; Strahl, H.; Trost, M.; et al. The Type VI Secretion System Deploys Antifungal Effectors against Microbial Competitors. Nat. Microbiol. 2018, 3, 920–931. [Google Scholar] [CrossRef]
- Hernandez, R.E.; Gallegos-Monterrosa, R.; Coulthurst, S.J. Type VI Secretion System Effector Proteins: Effective Weapons for Bacterial Competitiveness. Cell. Microbiol. 2020, 22, e13241. [Google Scholar] [CrossRef]
- Le, N.-H.; Pinedo, V.; Lopez, J.; Cava, F.; Feldman, M.F. Killing of Gram-Negative and Gram-Positive Bacteria by a Bifunctional Cell Wall-Targeting T6SS Effector. Proc. Natl. Acad. Sci. USA 2021, 118, e2106555118. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of Effector-like Proteins in Trichoderma Spp. and Role of a Hydrophobin in the Plant-Fungus Interaction and Mycoparasitism. BMC Genet. 2017, 18, 16. [Google Scholar] [CrossRef] [Green Version]
- Horner, N.R.; Grenville-Briggs, L.J.; van West, P. The Oomycete Pythium Oligandrum Expresses Putative Effectors during Mycoparasitism of Phytophthora infestans and Is Amenable to Transformation. Fungal Biol. 2012, 116, 24–41. [Google Scholar] [CrossRef] [Green Version]
- Romero-Contreras, Y.J.; Ramírez-Valdespino, C.A.; Guzmán-Guzmán, P.; Macías-Segoviano, J.I.; Villagómez-Castro, J.C.; Olmedo-Monfil, V. Tal6 From Trichoderma atroviride Is a LysM Effector Involved in Mycoparasitism and Plant Association. Front. Microbiol. 2019, 10, 02231. [Google Scholar] [CrossRef]
- Snelders, N.C.; Rovenich, H.; Petti, G.C.; Rocafort, M.; van den Berg, G.C.M.; Vorholt, J.A.; Mesters, J.R.; Seidl, M.F.; Nijland, R.; Thomma, B.P.H.J. Microbiome Manipulation by a Soil-Borne Fungal Plant Pathogen Using Effector Proteins. Nat. Plants 2020, 6, 1365–1374. [Google Scholar] [CrossRef]
- Snelders, N.C.; Petti, G.C.; van den Berg, G.C.M.; Seidl, M.F.; Thomma, B.P.H.J. An Ancient Antimicrobial Protein Co-Opted by a Fungal Plant Pathogen for in Planta Mycobiome Manipulation. Proc. Natl. Acad. Sci. USA 2021, 118, e2110968118. [Google Scholar] [CrossRef]
- Sarkar, D.; Rovenich, H.; Jeena, G.; Nizam, S.; Tissier, A.; Balcke, G.U.; Mahdi, L.K.; Bonkowski, M.; Langen, G.; Zuccaro, A. The Inconspicuous Gatekeeper: Endophytic Serendipita vermifera Acts as Extended Plant Protection Barrier in the Rhizosphere. New Phytol. 2019, 224, 886–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma harzianum Interacts with Roots and Triggers Induced Systemic Resistance to Foliar Disease in Maize. Sci. Rep. 2016, 6, 35543. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Ji, S.; Wang, Z.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma asperellum Xylanases Promote Growth and Induce Resistance in Poplar. Microbiol. Res. 2021, 248, 126767. [Google Scholar] [CrossRef] [PubMed]
- Vleeshouwers, V.G.A.A.; Raffaele, S.; Vossen, J.H.; Champouret, N.; Oliva, R.; Segretin, M.E.; Rietman, H.; Cano, L.M.; Lokossou, A.; Kessel, G.; et al. Understanding and Exploiting Late Blight Resistance in the Age of Effectors. Annu. Rev. Phytopathol. 2011, 49, 507–531. [Google Scholar] [CrossRef] [Green Version]
- Bart, R.; Cohn, M.; Kassen, A.; McCallum, E.J.; Shybut, M.; Petriello, A.; Krasileva, K.; Dahlbeck, D.; Medina, C.; Alicai, T.; et al. High-Throughput Genomic Sequencing of Cassava Bacterial Blight Strains Identifies Conserved Effectors to Target for Durable Resistance. Proc. Natl. Acad. Sci. USA 2012, 109, E1972–E1979. [Google Scholar] [CrossRef] [Green Version]
- Giannakopoulou, A.; Steele, J.F.C.; Segretin, M.E.; Bozkurt, T.O.; Zhou, J.; Robatzek, S.; Banfield, M.J.; Pais, M.; Kamoun, S. Tomato I2 Immune Receptor Can Be Engineered to Confer Partial Resistance to the Oomycete Phytophthora infestans in Addition to the Fungus Fusarium oxysporum. Mol. Plant-Microbe Interact. 2015, 28, 1316–1329. [Google Scholar] [CrossRef] [Green Version]
- Van de Wouw, A.P.; Idnurm, A. Biotechnological Potential of Engineering Pathogen Effector Proteins for Use in Plant Disease Management. Biotechnol. Adv. 2019, 37, 107387. [Google Scholar] [CrossRef]
Figure 1.
A timeline of some noteworthy milestones that shaped Effector Biology.

Figure 2.
Examples of effectors and their intracellular targets. Secreted effectors enter the cell apoplast before they reach the interior of the cell; some are retained in the apoplast if their targets are apoplastic while others traverse the cell membrane to reach their targets inside the cell. In the intracellular bacterial effector examples given, the effectors AvrBs3 [159], HopG1 [160,161], AvrRps4 [86,162], and HopD1 [163] target the nucleus, mitochondria, chloroplast, and endoplasmic reticulum (ER), respectively. The fungal effectors displayed are AvrSr35 [164], ChEC21 [156], ToxA [165], VdSCP7 [166], and MoPtep1 [167] which target the ER, Golgi, chloroplast, nucleus, and peroxisomes, respectively. The oomycetes effectors Pi04313 [150] and PsCRN63 [168,169] target the nucleus while PsAvh262 [170] targets the mitochondria and HaRxL17 [171] associates with the plant tonoplast (vacuolar membrane).
Figure 2.
Examples of effectors and their intracellular targets. Secreted effectors enter the cell apoplast before they reach the interior of the cell; some are retained in the apoplast if their targets are apoplastic while others traverse the cell membrane to reach their targets inside the cell. In the intracellular bacterial effector examples given, the effectors AvrBs3 [159], HopG1 [160,161], AvrRps4 [86,162], and HopD1 [163] target the nucleus, mitochondria, chloroplast, and endoplasmic reticulum (ER), respectively. The fungal effectors displayed are AvrSr35 [164], ChEC21 [156], ToxA [165], VdSCP7 [166], and MoPtep1 [167] which target the ER, Golgi, chloroplast, nucleus, and peroxisomes, respectively. The oomycetes effectors Pi04313 [150] and PsCRN63 [168,169] target the nucleus while PsAvh262 [170] targets the mitochondria and HaRxL17 [171] associates with the plant tonoplast (vacuolar membrane).

Figure 3.
Effectors showing different host-range preferences based on the effector classification proposed by Snelders et al. (2018) [43]. The red effectors represent broad-host range effectors that are used by the mutualistic fungus against the plant host (far left) and against a bacterial competitor (center). It is unknown whether the red effector’s target in the bacterial competitor is the same target in the plant cell (large green icons) and the bacterium in response secretes a microbe-specific effector against the fungus (yellow effector). The purple effector represents a plant host-specific effector used by the bacterium (far right) that has an inhibitor function on the target (dark green star) in the plant cell.
Figure 3.
Effectors showing different host-range preferences based on the effector classification proposed by Snelders et al. (2018) [43]. The red effectors represent broad-host range effectors that are used by the mutualistic fungus against the plant host (far left) and against a bacterial competitor (center). It is unknown whether the red effector’s target in the bacterial competitor is the same target in the plant cell (large green icons) and the bacterium in response secretes a microbe-specific effector against the fungus (yellow effector). The purple effector represents a plant host-specific effector used by the bacterium (far right) that has an inhibitor function on the target (dark green star) in the plant cell.


{kind=link}
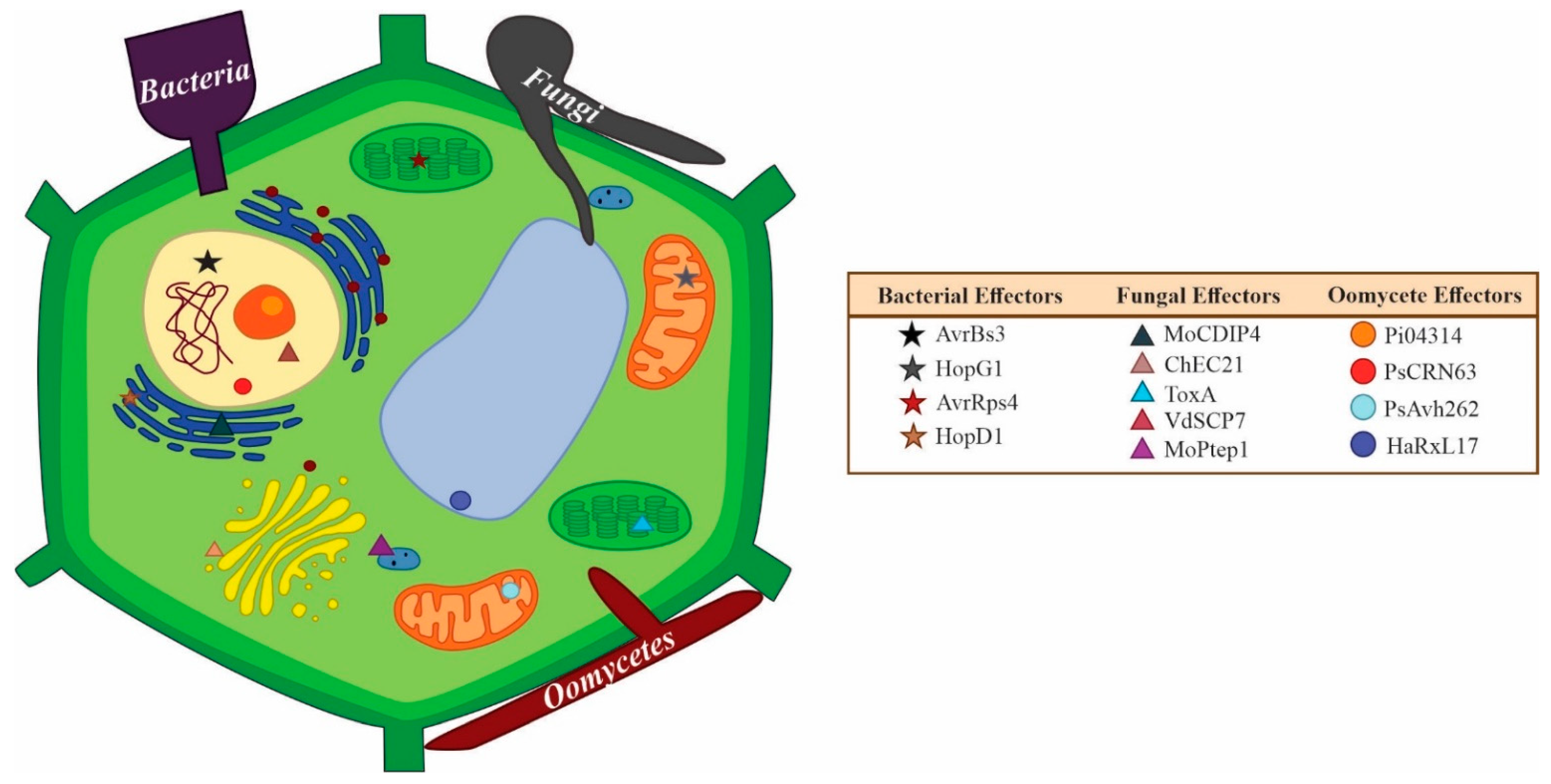{kind=link}
{kind=link}
Table 1.
Effectors can be species-specific, genus-specific or conserved across various genera.
| Distribution * | Effector Type | Organism | Effector Name and Uniprot ID | Function | Reference |
|---|---|---|---|---|---|
| Conserved: Different classes in Ascomycetes | Ceratoplatanin family | Botrytis cinerea | BcSpl1 (A0A384JBC5) | Phytotoxin, elicits systemic resistance (SAR) in tobacco. Necrosis (HR) in tobacco, tomato and Arabidopsis | [125,126] |
| Conserved: Wide distribution in Dothideomycetes class (Ascomycetes) | LysM domain | Cladosporium fulvum | Ecp6 (B3VBK9) | Virulence factor; binds to fungal chitin to prevent chitin triggered-immunity in host | [118] |
| Conserved: Colletotrichum genus (Glomerellales order), Fusarium, Trichoderma, (Hypocreales order) in Sordariomycetes class. Few species in Dothideomycetes class (Ascomycetes) | Necrosis inducing protein | Colletotrichum orbiculare | NIS1 (H7CE97) | Suppresses PAMP-triggered immunity, targets kinases BAK1 and BIK1 Recognition is followed by a plant cell death response (necrosis), potential avirulence effects | [41] |
| Conserved: Few genera in Sclerotiniaceae family (Ascomycetes phylum) | Necrosis and ethylene-inducing proteins (NLPs) | Botrytis cinerea | BcNep2 (Q079H3) | Induces cell death in dicotyledonous plants | [127] |
| Conserved: Fusarium genus (Hypocreales order), Bipolaris genus (Pleosporales order), Verticillium genus (Glomerellales order), in Sordariomycetes class (Ascomycetes phylum) | Necrosis and ethylene-inducing proteins (NLPs) | Fusarium oxysporum f. sp. erythroxyli | NEP1 (O42737) | Induces necrosis and ethylene emission in Erythroxylum coca leaves | [128,129] |
| Conserved: Many species in Colletotrichum genus (Glomerellales order) and Fusarium genus (Hypocreales order) | Necrosis inducing protein | Colletotrichum higginsianum | ChNLP1 (K7N7F9) | Induces necrosis in N. benthamiana | [130] |
| Conserved: Many species in Mycosphaerellaceae family, Dothideomycetes class | Chitin-binding type-2 | Cladosporium fulvum | Avr4 (Q00363) | Triggers a Cf-4-mediated hypersensitive response (HR) in tomato; protects fungal cell walls against hydrolysis by plant chitinases | [55,113,131] |
| Conserved: Species in Ustilaginaceae family | Pep1 | Ustilago maydis | Pep1 (G0X7E8) | Inhibitor of plant peroxidases; targets maize peroxidase POX12 to suppress plant immunity | [106,132] |
| Conserved: Few species in Magnaporthales, Sordariales, Xylariales, Hypocreales, Glomerellales orders | MC69 | Magnaporthe oryzae | MC69 (L7JHY1) | Development of invasive hyphae affected in mc69 mutant; reduced pathogenicity on host | [133] |
| Specific to genus | Avr | Phytophthora sojae | PsAvh163 (G1FRR2) | Suppresses PTI and ETI responses in Arabidopsis Activates immunity (HR response) in the Nicotiana genus. | [134] |
| Specific to genus | Hop | Pseudomonas syringae | HopAl1 (Q888W0) | Phosphothreonine lyase; inactivates MAPKs in Arabidopsis to overcome PTI | [135] |
| Specific to genus | Suppressor of necrosis 1 SNE1 | Phytophthora infestans | SNE1 (A2CLL0) | Suppresses cell death induced by NLP and Avr effectors in N. benthamiana and Solanum lycopersicum | [136] |
| Specific to genus | Avr | Melamspora lini | AvrM (Q2MV46) | Induces ETI in host; targets resistance protein M | [137,138] |
| Specific to genus | Avr | Magnaporthe oryzae | AvrPia (B9WZW9) | Induces ETI in host; targets resistance protein RGA5 | [139] |
| Species-specific | Avr | Cladosporium fulvum | Avr9 (P22287) | Induces ETI in host; Targets resistance protein of Cf-9 | [140] |
| Species-specific | Avr | Magnaporthe oryzae | AVR-Pik (C4B8B8) | Induces ETI in host; targets resistance protein Pik | [56,141] |
| Species-specific | Biotrophy-associated secreted (BAS) proteins | Magnaporthe oryzae | BAS1 (G5EHI7) | Triggers defense response in host; overexpression increases virulence, sporulation and reduces expression of host defense-related genes | [142,143] |
| Species-specific | Ribonuclease family | Blumeria graminis | BEC1054 (N1JJ94) | Ribonuclease like; binds to host ribosomes and inhibits the action of plant ribosome-inactivating proteins (RIPs) | [144] |
* Distribution was determined by Blastp at NCBI for fungi (taxid: 4751) and oomycetes (taxid: 4762).
Table 2.
Examples of intracellular effectors with experimentally proven locations.
| Effector | Cell Localization | Microorganism | Kingdom | Lifestyle | Reference |
|---|---|---|---|---|---|
| CgEP1 | Nucleus | Colletotrichum graminicola | Fungi | Hemibiotrophic | [109] |
| ChECs (nine) | Nucleus, peroxisomes, and microtubules | Colletotrichum higginsianum | Fungi | Hemibiotrophic | [156] |
| MoCDIP4 | Mitochondria | Magnaporthe oryzae | Fungi | Hemibiotrophic | [172] |
| Rab8a | Golgi apparatus | Phytophthora infestans | Oomycete | Hemibiotrophic | [173] |
| CSEP0064/ BEC1054 | Cytoplasmic | Blumeria graminis | Fungi | Biotrophic | [144] |
| Sntf2 | Chloroplast | Colletotrichum gloeosporioides | Fungi | Hemibiotrophic | [111] |
| CgNLP1 | Nucleus | Colletotrichum gloeosporioides | Fungi | Hemibiotrophic | [174] |
| ToxA | Cytoplasm | Parastagonospora nodorum | Fungi | Necrotrophic | [175] |
| SsITL | Chloroplast | Sclerotinia sclerotiorum | Fungi | Necrotrophic | [176] |
| PEF1 | Peroxisomes | Magnaporthe oryzae | Fungi | Hemibiotrophic | [177] |
| DspA/E | Peroxisomes | Erwinia amylovora | Bacteria | Necrotrophic | [178] |
| RsCRP1 | Mitochondria and chloroplasts | Rhizoctonia solani | Fungi | Necrotrophic | [179] |
| RipAA | Chloroplasts | Ralstonia solanacearum | Bacteria | Necrotrophic | [180] |
| HopAF1 | Plasma membrane and cytoplasm | Pseudomonas syringae | Bacteria | Hemibiotrophic | [181] |
| PcRxLR48 | Nucleus | Phytophthora infestans | Oomycete | Hemibiotrophic | [182] |
| PsAvh52 | Cytoplasm and nucleus | Phytophthora infestans | Oomycete | Hemibiotrophic | [183] |
| AvrLm1 | Plasma membrane | Leptosphaeria maculans | Fungi | Hemibiotrophic | [184] |
| PstGSRE4 | Nucleus | Puccinia striiformis f. sp. tritici | Fungi | Biotrophic | [185] |
| MiSSP7 | Nucleus | Laccaria bicolor | Fungi | Ectomycorrhizal symbiont | [186] |
| RsCRP1 | Mitochondria and chloroplasts | Rhizoctonia solani | Fungi | Necrotrophic | [179] |
| PexRD54 | Golgi apparatus | Phytophthora infestans | Oomycete | Hemibiotrophic | [173] |
Table 3.
Examples of hubs (core proteins commonly involved in plant-pathogen interactions) identified in hosts and pathogens.
Table 3.
Examples of hubs (core proteins commonly involved in plant-pathogen interactions) identified in hosts and pathogens.
| Plants | ||||
|---|---|---|---|---|
| Host Target (Hub) (Targets in Different Hosts Targeted by Multiple Effectors) | Function in the Host | Targeted by (Selected Examples) | Microorganisms | References |
| Chorismate mutase (CM) | Salicylic acid (SA) biosynthesis | Cmu1, Mi-CM-3, AvrPtoB | Ustilago maydis (fungus), Meloidogyne javanica (nematode), Pseudomonas syringae (bacterium) | [197,198] |
| Isochorismatase (ICM) | Salicylic acid (SA) biosynthesis | VdIsc1, PsIsc1, HopI1 | Verticillium dahliae (fungus), Phytophthora sojae (oomycete), Pseudomonas syringae (bacterium) | [197] |
| JAZ6, negative regulator of JA induced transcription | Jasmonic acid (JA) biosynthesis | MiSSP7, HopZ1a, HopX1 | Laccaria bicolor and Golovinomyces orontii (fungi), Hyaloperonospora arabidopsidis (oomycete), Pseudomonas syringae (bacterium) | [197] |
| AuTophaGy (ATG) proteins | Autophagy | HrpZ1, HopF3, AvrPtoB, HrpZ1, HopO1-2, | Bremia lactucae, Hyaloperonospora arabidopsidis (oomycetes), Uromyces fabae (fungus), Globodera pallida (nematode), Pseudomonas syringae (bacterium) | [199] |
| Pathogen | ||||
| Pathogen hub (Multiple targets to one effector) | Functions | Targets in the host | Microorganisms where that effector has been identified | References |
| AvrPtoB | Interferes with salicylic acid (SA) biosynthesis and autophagy in plants | CM, ATG1 | Pseudomonas syringae | [198,199] |
| Zt6 | Phytotoxic and antimicrobial ribonuclease effector | Plant and microbial rRNA species | Zymoseptoria tritici | [200,201] |
| VdAMP2 | Soil colonization (antibacterial activity), and induction of necrosis in Nicotiana benthamiana | Unknown | Verticillium dahliae | [17] |
| VdAMP3 | Soil colonization (antifungal activity) | Unknown, mycobiome manipulation | Verticillium dahliae | [17] |
| VirD5 | Stabilizes VirF for Agrobacterium infection | Arabidopsis thaliana AT1G09270.1 AT1G43700.1 AT3G06720.1 AT5G59710.1 VIP2 | Agrobacterium tumefaciens | [193] |
| Avr2 | Inhibits tomato Rcr3 protease | Arabidopsis thaliana AT1G47128.1 AT3G19400.1 AT3G45310.1 AT4G35350.1 AT5G60360.1 | Cladosporium fulvum | [193] |
| HARXL106 | Suppresses transcriptional activation of salicylic acid (SA)-induced defense genes | Arabidopsis thaliana AT1G32230.1 AT2G35510.1 | Hyaloperonospora arabidopsidis | [193] |
| RXLR24 | Inhibitor of RABA GTPase-mediated vesicular secretion of antimicrobial PR-1 and PDF1.2 | Arabidopsis thaliana AT1G06400.1 AT1G09630.1 AT1G16920.1 AT2G30950.1 AT3G18820.1 AT3G46830.1 AT3G56940.1 AT4G18800.1 AT4G39990.1 AT5G45750.1 AT5G47960.1 AT5G59150.1 AT5G60860.1 AT5G65270.1 | Phytophthora brassicae | [193] |
Table 4.
A summary of changes in concepts in the field of effectoromics.
| PAST | PRESENT |
|---|---|
| Effectors show no or limited sequence homology or conservation but may share structural properties | Some effectors are homologs of effectors in other microorganisms. Some orthologs display a high level of sequence conservation |
| Homologs are distributed in close phylogenetic relatives | Some effectors are distributed in phylogenetically related and distant organisms (core effectors) |
| Effectors are small proteins (<300 or 400 amino acids) | Limit in length of known current effectors is ~ 850 amino acids |
| Effectors are secreted proteins with signal Peptides | Many lack signal peptides and are secreted by little-understood non-conventional processes |
| Effector proteins lack TMDs | Some true effectors have one or two TMDs; the current limit for TMDs in a true effector is 6 |
| The majority of effectors initially discovered were extracellular, apoplastic proteins | Many effectors also target cytoplasmic and organellar host proteins. Some effectors may even target both apoplast and cytoplasm |
| Effector interactions follow the gene-for-gene model | Effector interactions encompass the gene-for-gene model, guard model, the decoy model, integrated decoy model etc. |
| Effectors are proteins (Enzymatic or non-enzymatic) | Effectors can be proteins, RNA or secondary metabolites |
| Effectors induce the hypersensitive response (local, visible lesion) on the host leaves | Not all effectors induce HR. Biotrophic effectors can interfere with downstream reactions triggered by effector-R cognate recognition and prevent development of visible lesions |
| MTI and ETI are independent immunity Pathways | MTI and ETI are interconnected |
| Effectors are produced by pathogens to target plant hosts | Produced by pathogens and non-pathogens with targets in a wide spectrum of organisms |
| Effector roles are associated with conquering the plant host | Effector roles documented in host and niche conquest; roles in shaping microbiomes and roles in reproduction and development of producer organism |
| Each effector has one target or cognate | Some effectors are multifunctional or have many different targets in the same host or different hosts; and vice versa, some host targets are targeted by multiple effectors from the same or different microorganisms |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Todd, J.N.A.; Carreón-Anguiano, K.G.; Islas-Flores, I.; Canto-Canché, B. Fungal Effectoromics: A World in Constant Evolution. Int. J. Mol. Sci. 2022, 23, 13433. https://doi.org/10.3390/ijms232113433
AMA Style
Todd JNA, Carreón-Anguiano KG, Islas-Flores I, Canto-Canché B. Fungal Effectoromics: A World in Constant Evolution. International Journal of Molecular Sciences. 2022; 23(21):13433. https://doi.org/10.3390/ijms232113433
Chicago/Turabian StyleTodd, Jewel Nicole Anna, Karla Gisel Carreón-Anguiano, Ignacio Islas-Flores, and Blondy Canto-Canché. 2022. "Fungal Effectoromics: A World in Constant Evolution" International Journal of Molecular Sciences 23, no. 21: 13433. https://doi.org/10.3390/ijms232113433
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.