
Schiedeella bajaverapacensis (Orchidaceae, Spiranthinae), a New Orchid Species from Guatemala
 ,
,  , and
, and
Abstract
:1. Introduction
2. The History of Classification of Schiedeella Complex
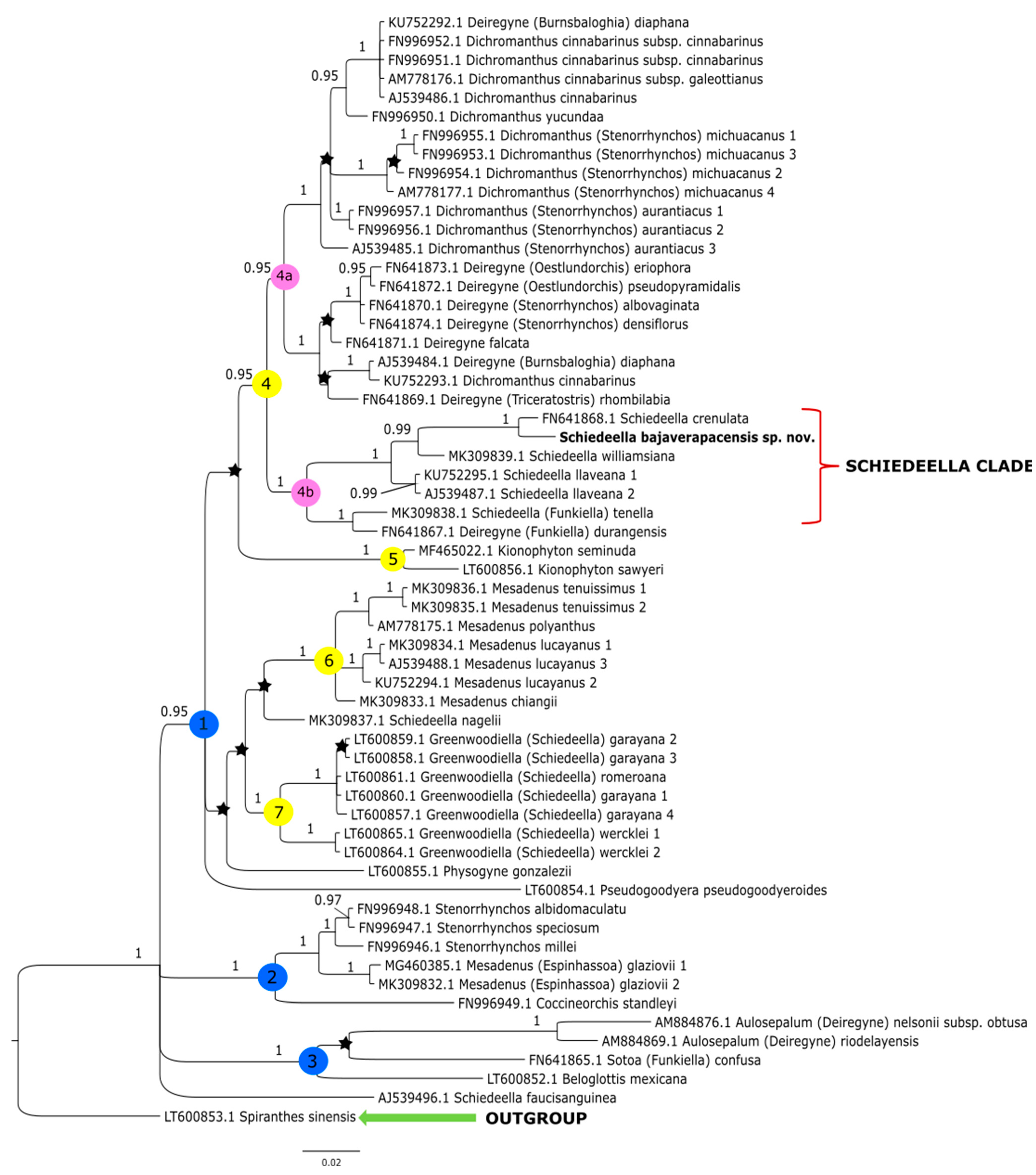
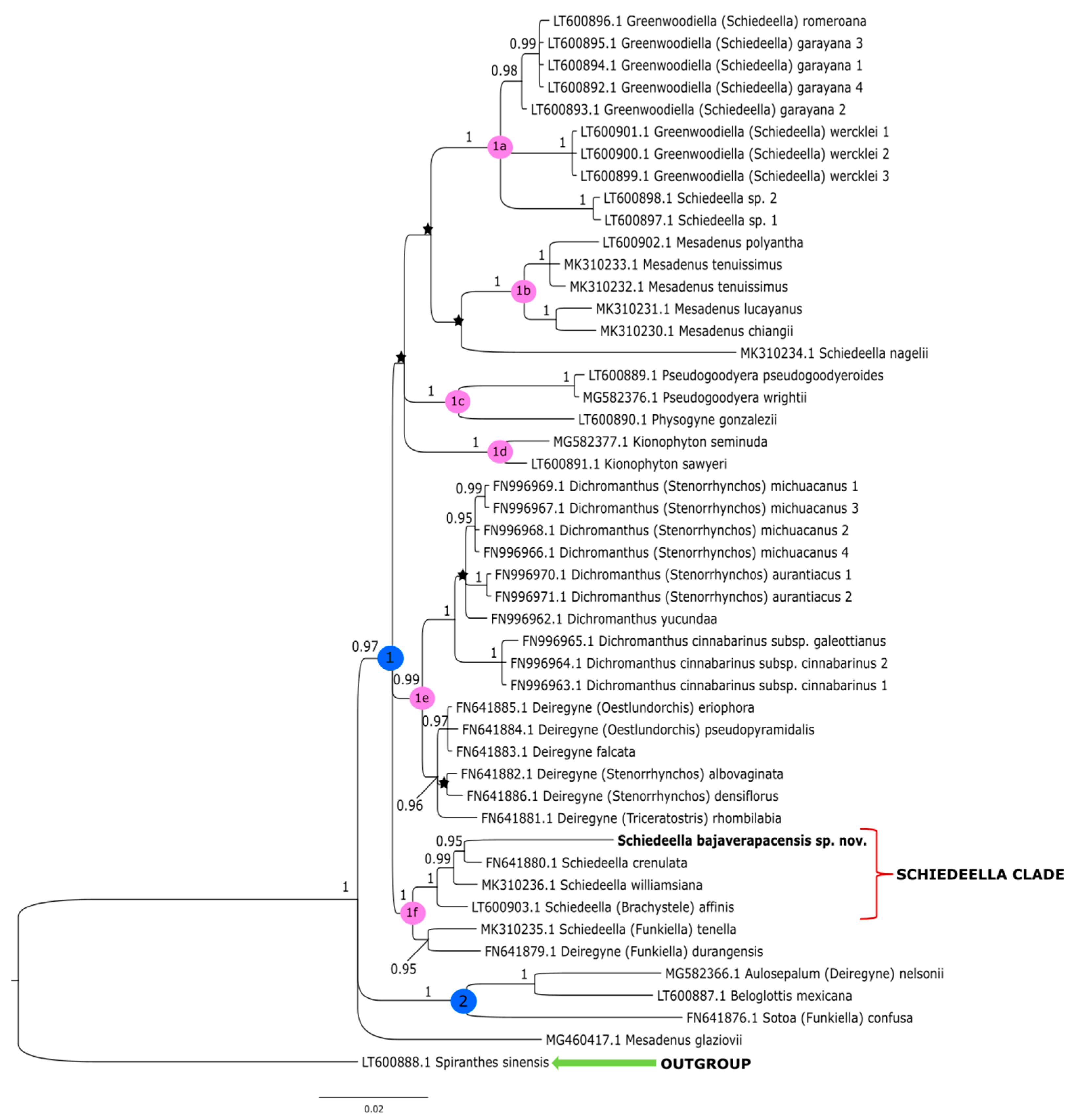
3. Results
3.1. ITS Matrix
3.2. Plastid Matrix
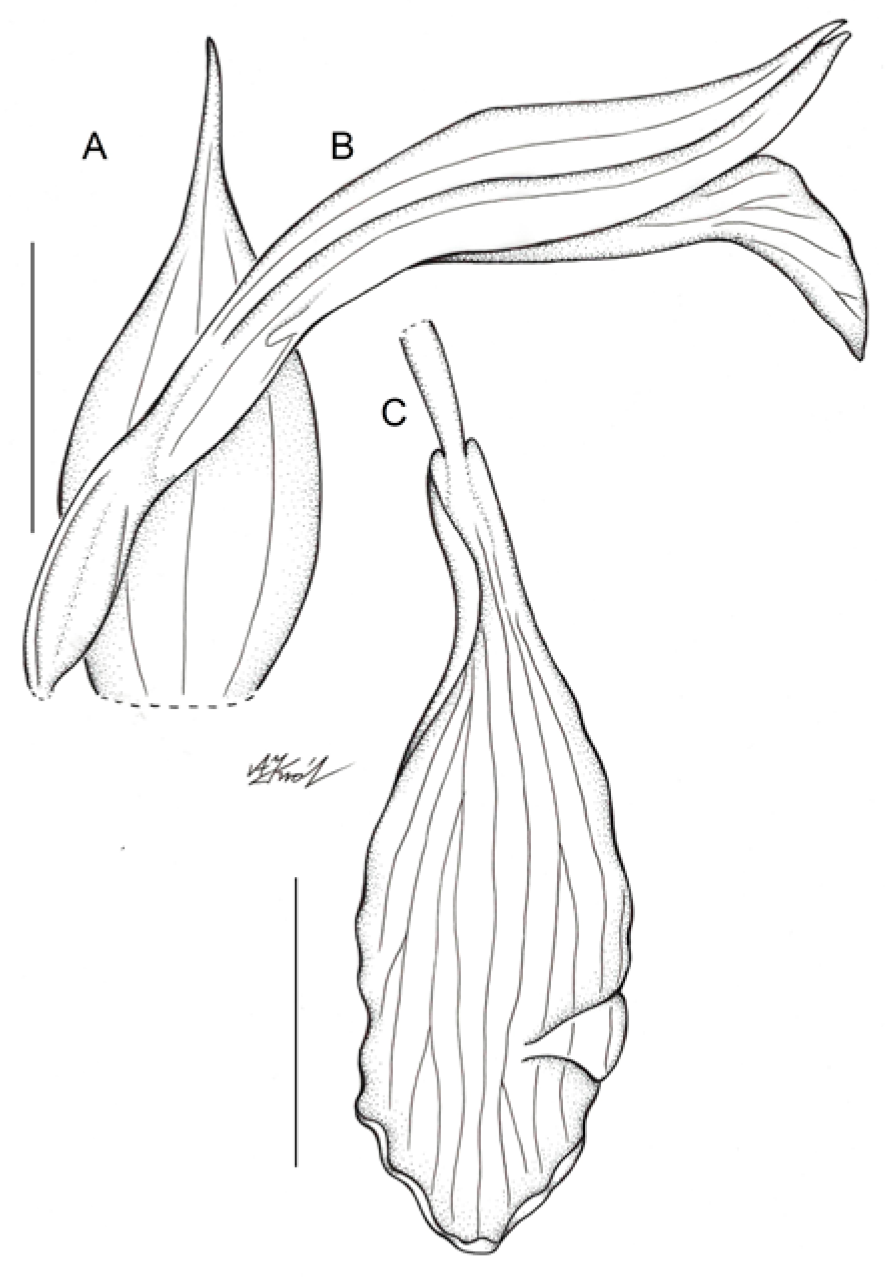
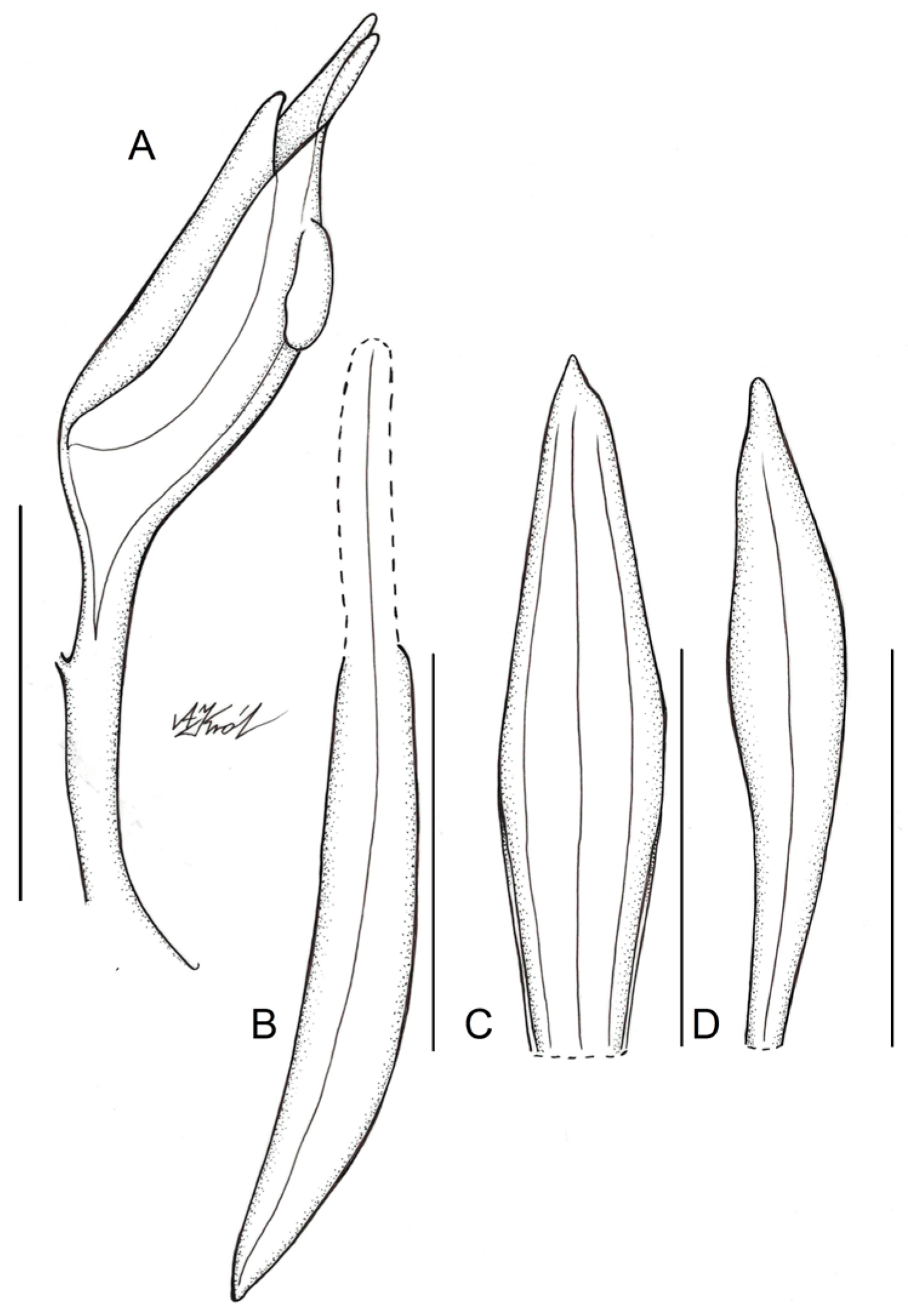
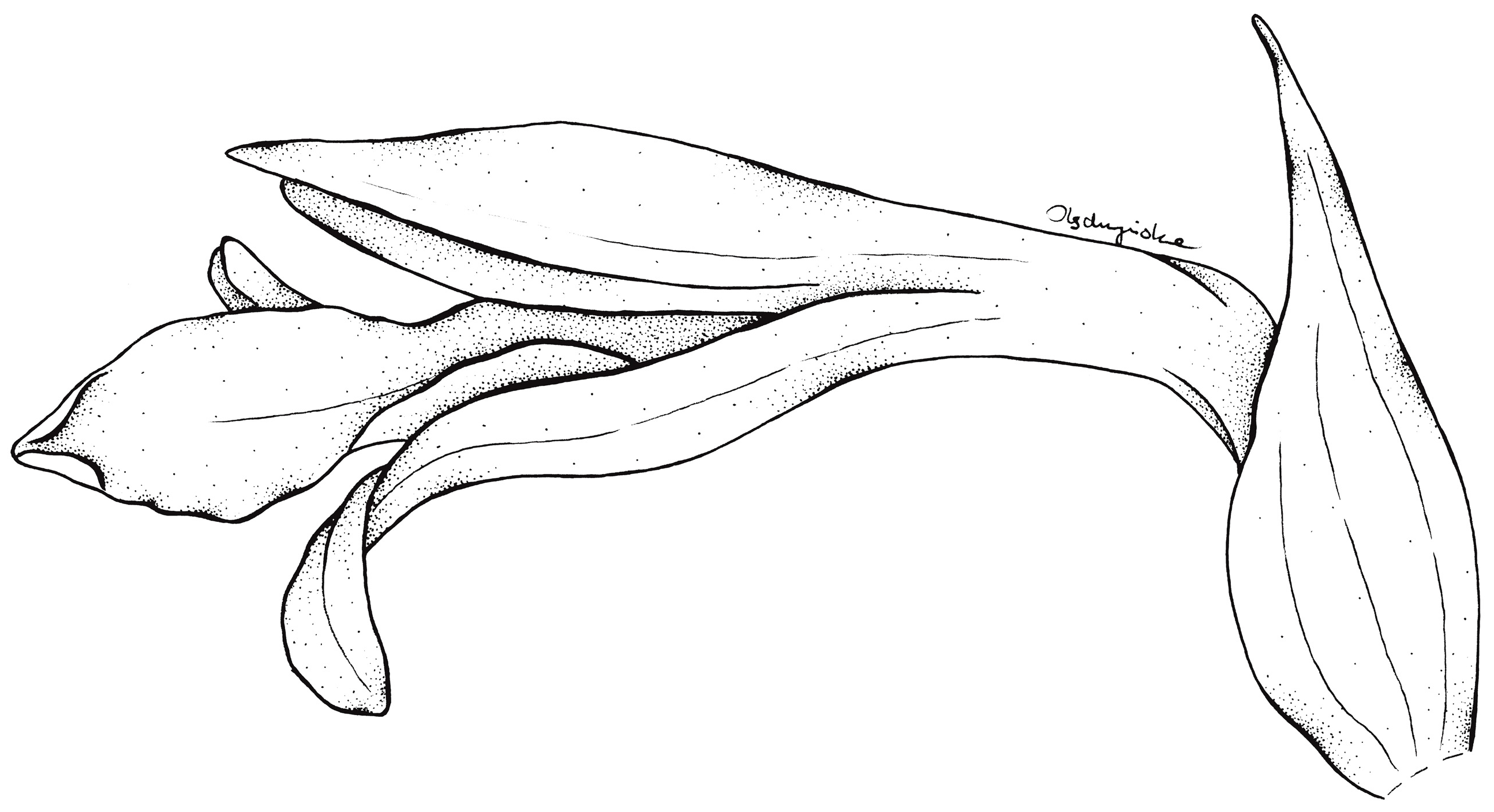
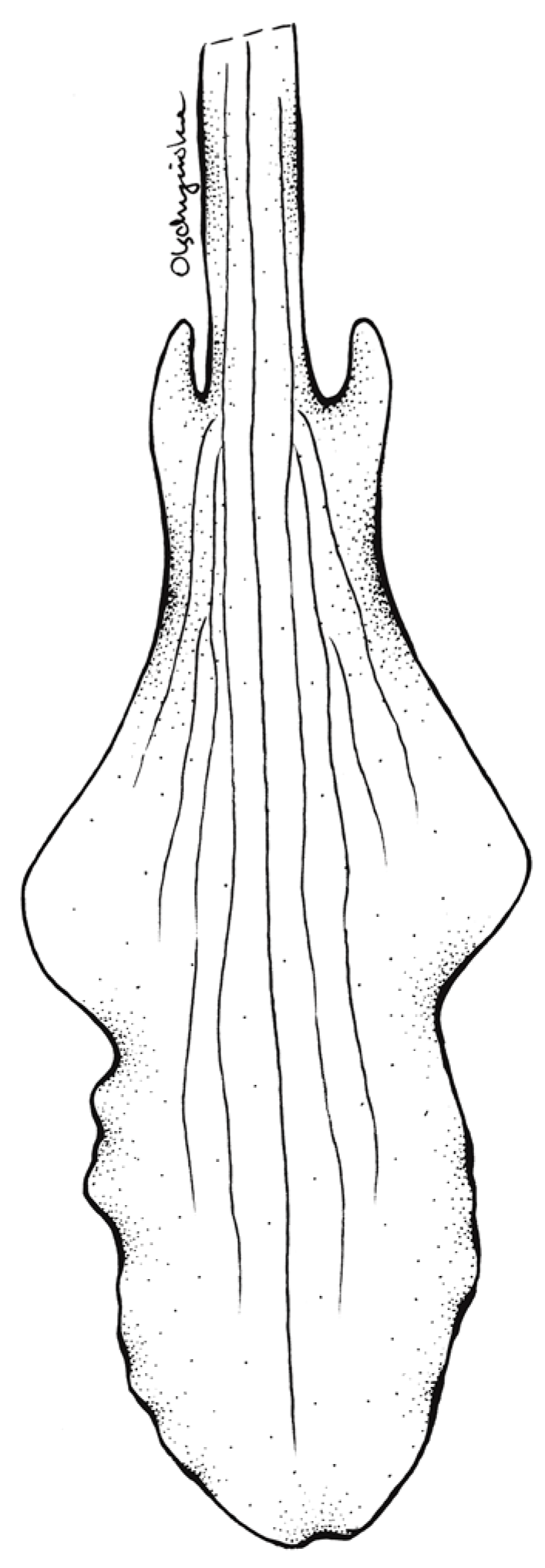
3.3. Taxonomic Treatment
4. Discussion
5. Materials and Methods
5.1. Morphological Study
5.2. Molecular Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Accession Number of ITS | Accession Number of trnL-trnF |
|---|---|---|
| Aulosepalum nelsonii | ------- | MG582366.1 |
| Aulosepalum nelsonii subsp. obtusa | AM884876.1 | ------- |
| Aulosepalum riodelayensis | AM884869.1 | ------- |
| Beloglottis mexicana | LT600852.1 | LT600887.1 |
| Coccineorchis standleyi | FN996949.1 | ------- |
| Deiregyne albovaginata | FN641870.1 | FN641882.1 |
| Deiregyne densiflorus | FN641874.1 | FN641886.1 |
| Deiregyne diaphana 1 | KU752292.1 | ------- |
| Deiregyne diaphana 2 | AJ539484.1 | ------- |
| Deiregyne durangensis | FN641867.1 | FN641879.1 |
| Deiregyne eriophora | FN641873.1 | FN641885.1 |
| Deiregyne falcata | FN641871.1 | FN641883.1 |
| Deiregyne pseudopyramidalis | FN641872.1 | FN641884.1 |
| Deiregyne rhombilabia | FN641869.1 | FN641881.1 |
| Dichromanthus aurantiacus 1 | FN996957.1 | FN996970.1 |
| Dichromanthus aurantiacus 2 | FN996956.1 | FN996971.1 |
| Dichromanthus aurantiacus 3 | AJ539485.1 | ------- |
| Dichromanthus cinnabarinus 1 | KU752293.1 | ------- |
| Dichromanthus cinnabarinus 2 | AJ539486.1 | ------- |
| Dichromanthus cinnabarinus subsp. cinnabarinus 1 | FN996952.1 | FN996964.1 |
| Dichromanthus cinnabarinus subsp. cinnabarinus 2 | FN996951.1 | FN996963.1 |
| Dichromanthus cinnabarinus subsp. galeottianus | AM778176.1 | FN996965.1 |
| Dichromanthus michuacanus 1 | FN996955.1 | FN996969.1 |
| Dichromanthus michuacanus 2 | FN996954.1 | FN996968.1 |
| Dichromanthus michuacanus 3 | FN996953.1 | FN996967.1 |
| Dichromanthus michuacanus 4 | AM778177.1 | FN996966.1 |
| Dichromanthus yucundaa | FN996950.1 | FN996962.1 |
| Greenwoodiella garayana 1 | LT600860.1 | LT600894.1 |
| Greenwoodiella garayana 2 | LT600859.1 | LT600893.1 |
| Greenwoodiella garayana 3 | LT600858.1 | LT600895.1 |
| Greenwoodiella garayana 4 | LT600857.1 | LT600892.1 |
| Greenwoodiella romeroana | LT600861.1 | LT600896.1 |
| Greenwoodiella wercklei 1 | LT600865.1 | LT600901.1 |
| Greenwoodiella wercklei 2 | LT600864.1 | LT600900.1 |
| Greenwoodiella wercklei 3 | ------- | LT600899.1 |
| Kionophyton sawyeri | LT600856.1 | LT600891.1 |
| Kionophyton seminuda | MF465022.1 | MG582377.1 |
| Mesadenus chiangii | MK309833.1 | MK310230.1 |
| Mesadenus glaziovii 1 | MG460385.1 | MG460417.1 |
| Mesadenus glaziovii 2 | MK309832.1 | ------- |
| Mesadenus lucayanus 1 | MK309834.1 | MK310231.1 |
| Mesadenus lucayanus 2 | KU752294.1 | ------- |
| Mesadenus lucayanus 3 | AJ539488.1 | ------- |
| Mesadenus polyanthus | AM778175.1 | LT600902.1 |
| Mesadenus tenuissimus 1 | MK309836.1 | MK310233.1 |
| Mesadenus tenuissimus 2 | MK309835.1 | MK310232.1 |
| Physogyne gonzalezii | LT600855.1 | LT600890.1 |
| Pseudogoodyera pseudogoodyeroides | LT600854.1 | LT600889.1 |
| Pseudogoodyera wrightii | ------- | MG582376.1 |
| Schiedeella affinis | ------- | LT600903.1 |
| Schiedeella crenulata | FN641868.1 | FN641880.1 |
| Schiedeella faucisanguinea | AJ539496.1 | ------- |
| Schiedeella llaveana 1 | KU752295.1 | ------- |
| Schiedeella llaveana 2 | AJ539487.1 | ------- |
| Schiedeella nagelii | MK309837.1 | MK310234.1 |
| Schiedeella sp. 1 | ------- | LT600897.1 |
| Schiedeella sp. 2 | ------- | LT600898.1 |
| Schiedeella tenella | MK309838.1 | MK310235.1 |
| Schiedeella williamsiana | MK309839.1 | MK310236.1 |
| Sotoa confusa | FN641865.1 | FN641876.1 |
| Spiranthes sinensis | LT600853.1 | LT600888.1 |
| Stenorrhynchos albidomaculatu | FN996948.1 | ------- |
| Stenorrhynchos millei | FN996946.1 | ------- |
| Stenorrhynchos speciosum | FN996947.1 | ------- |
References
- Archila, F.; Szlachetko, D.L.; Chiron, G.; Lipińska, M.M.; Mystkowska, K.; Bertolini, V. Orchid Genera and Species in Guatemala; Koeltz Botanical Books: Oberreifenberg, Germany, 2018; pp. 1–724. [Google Scholar]
- Bateman, J. Orchidaceae of Mexico and Guatemala; J. Ridgway & Sons: London, UK, 1843; pp. 1–264. [Google Scholar]
- Ames, O.; Correll, D.S. Orchids of Guatemala; Chicago Natural History Museum: Chicago, IL, USA, 1952; pp. 1–779. [Google Scholar]
- Archila, F. Recolección, Cultivo y Reproducción de Orquídeas. Escuela Nacional Central de Agricultura (ENCA). Practica Profesional Agrícola y Forestal Supervisada (PPAFS). Informe Técnico; Barcena: Villa Nueva, Guatemala, 1992. [Google Scholar]
- Archila, F. Listado de Orquídeas de Guatemala. Revista Guatemal. 2014, 17, 32–71. [Google Scholar]
- Schlechter, R. Versuch einer systematischen Neuordnung der Spiranthinae. Beih. Bot. Centralbl. 1920, 37, 317–454. [Google Scholar]
- Szlachetko, D.L. Genera and species of the subtribe Spiranthinae (Orchidaceae). 2. A revision of Schiedeella. Fragm. Flor. Geobot. Pol. 1992, 37, 157–204. [Google Scholar]
- Burns-Balogh, P. Generic redefinition in the subtribe Spiranthinae (Orchidaceae). Am. J. Bot. 1982, 69, 1119–1132. [Google Scholar] [CrossRef]
- Garay, L.A. A generic revision of the Spiranthinae. Bot. Mus. Leafl. 1982, 28, 278–425. [Google Scholar] [CrossRef]
- Szlachetko, D.L. Systema Orchidalium. Fragm. Flor. Geobot. Pol. 1995, (Suppl. 3), 1–152. [Google Scholar]
- Burns-Balogh, P. A synopsis of Mexican Spiranthinae. Orquídea 1986, 10, 47–96. [Google Scholar]
- Szlachetko, D.L.; Rutkowski, P.; Mytnik, J. Contributions to the taxonomic revision of the subtribes Spiranthinae, Stenorrhynchidinae and Cyclopogoninae (Orchidaceae) in Mesoamerica and the Antilles. Pol. Bot. Stud. 2005, 20, 3–387. [Google Scholar]
- Salazar, G.A.; Hernández-López, T.J.; Sharma, J.; Jiménez-Machorro, R.; Cabrer, L.I.; Treviño-Carreón, J. Greenwoodiella, a new genus of Spiranthinae (Orchidaceae) from North and Central America and the Greater Antilles, with a new species from the Chihuahuan Desert. Syst. Bot. 2016, 41, 823–838. [Google Scholar] [CrossRef]
- Salazar, G.A.; Batista, J.A.N.; Cabrera, L.I.; Van den Berg, C.; Whitten, W.M.; Smidt, E.C.; Buzatto, C.R.; Singer, R.B.; Gerlach, G.; Jimenez-Machorro, R.; et al. Phylogenetic systematics of subtribe Spiranthinae (Orchidaceae: Orchidoideae: Cranichideae) based on nuclear and plastid DNA sequences of a nearly complete generic sample. Bot. J. Linn. Soc. 2018, 186, 273–303. [Google Scholar] [CrossRef]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria. Version 14. 2019. Available online: https://www.iucnredlist.org/resources/redlistguidelines (accessed on 29 June 2021).
- Holmgren, P.K.; Holmgren, N.H. Index Herbariorum. Taxon 1993, 42, 489–505. [Google Scholar] [CrossRef]
- Chase, M.W.; Hills, H.H. Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon 1991, 40, 215–220. [Google Scholar] [CrossRef]
- Martin, C.M.; Morales, A.M.; Dudek, M.; Szlachetko, D.L. Pachygenium laurense (Orchidaceae, Spiranthinae), a new orchid species from Argentina—Morphological evidence and phylogenetic reconstruction. PeerJ 2022, 10, 13433. [Google Scholar] [CrossRef] [PubMed]
- Douzery, E.J.P.; Pridgeon, M.A.; Kores, P.; Linder, H.P.; Kurzweil, H.; Chase, M.W. Molecular phylogenetics of Disae (Orchidaeae): A contribution from nuclear ribosomal ITS sequences. Am. J. Bot. 1999, 86, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Galtier, N.; Gouy, M.; Gautier, C. SeaView and Phylo_win, two graphic tools for sequence alignment and molecular phylogeny. Bioinformatics 1996, 12, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author. Evolutionary Biology Centre, Uppsala University. Available online: http://www.abc.se/~nylander/ (accessed on 20 January 2023).
- Ronquist, F.; Teslenko, M.; Pvander, M.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Bot. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]








| S. esquintlensis | S. jeanmulleri | S. llaveana | S. parasitica | S. romeroana | S. schlechteriana | S. trilineata | S. trilineata var. thelymitra | S. valerioi | |
|---|---|---|---|---|---|---|---|---|---|
| Name’s authors | Szlach., Rutk. and Mytnik | Szlach., Rutk. and Mytnik | (Lindl ex. Benth) Schltr. | (A. Rich. and Galeotti) Schltr. | Szlach. | Szlach. and Shevia | (Lindl.) Burns-Bal. | (Rchb. f.) Szlach. | (Ames and C. Schweinf.) Szlach. and Sheviak |
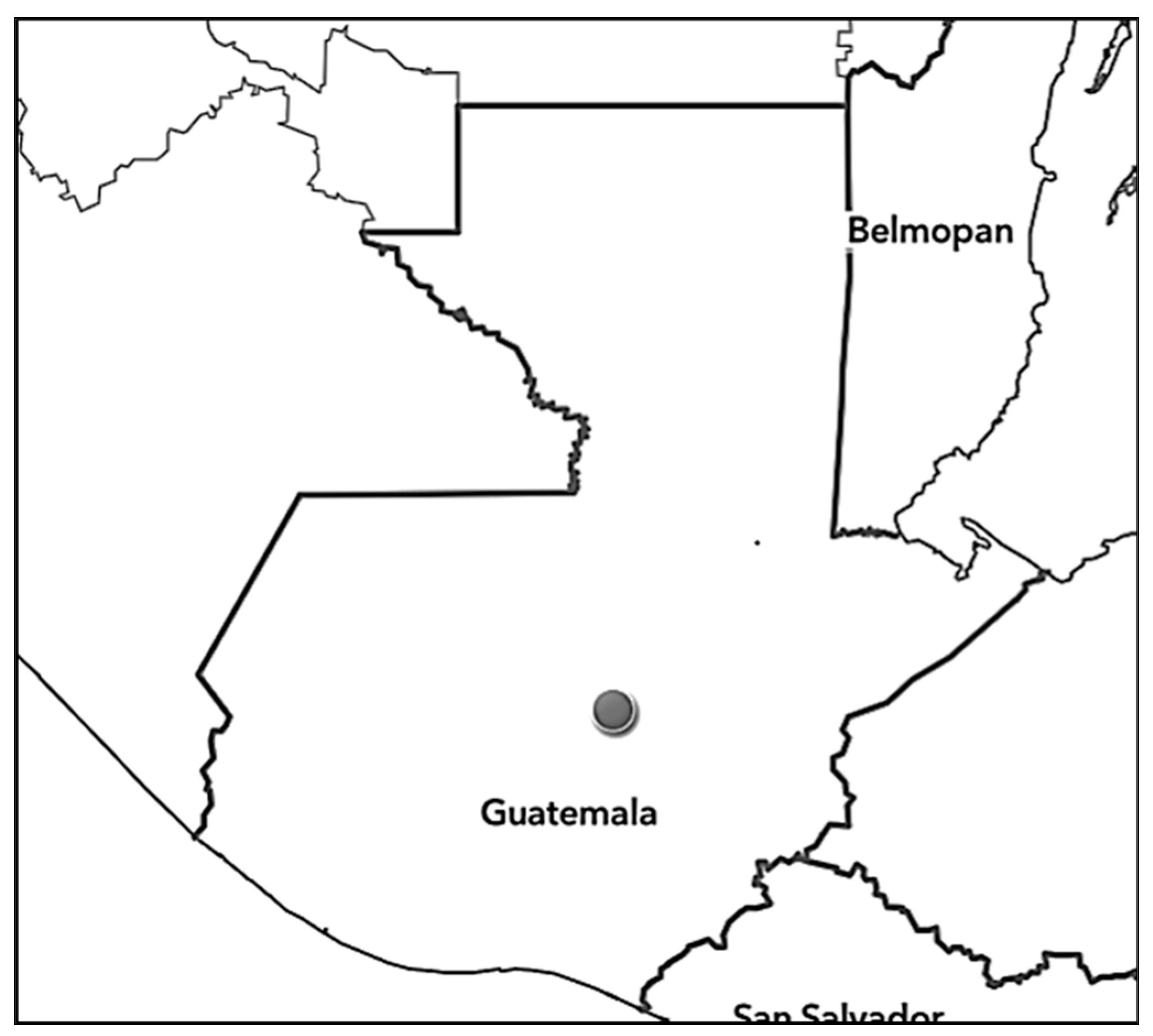
| Distribution | Esquintla | Esquintla | Baja Verapaz, San Marcos, El Progresso | Chimaltenango, San Marcos | Huehuetenango | Totonicapán | Chimaltenango, Chiquimula, Guatemala, Huehuetenango, Zacapa | Chiquimula | Chimaltenango, Quetzaltenango |
| Stem | 10 cm high, delicate, erect, glabrous | Ca. 20 cm high, very delicate, erect, glabrous, 6–7 sheaths | 19–16 (66) cm high, usually delicate, erect, glabrous, 5–7 sheaths | 10–25 (38) cm high, delicate, erect, usually glabrous, rarely sparsely glandular along spike, 5–7 sheaths | 35 cm high, delicate, erect, tinged reddish, sparsely glandular under and within the spike, 9 sheaths | 6.5–8 cm tall, erect, glabrous below, glandular above the uppermost bracts | 6–20.5 cm high, erect, delicate, glabrous, whitish or brownish, 4–8 sheaths | 15–43 cm high, rather delicate, glabrous, only in the upper part glandular, 5–7 sheaths | |
| Sheaths | Imbricate, semiscarious, semitransparent cauline | Cauline, herbaceous, thin, delicate, the lower ones imbricate, the upper ones shorter than internodes, glabrous | Cauline, more or less adnate to the stem, acute, herbaceous, the lower ones longer than internodes, the others shorter | Cauline, adnate to the stem, acuminate, herbaceous, sometimes with hyaline margins, shorter than internodes, except the lowermost, glabrous | Cauline, tubular, acuminate, tightly adnate to the stem, herbaceous, with hyaline margins, glabrous | Cauline, longer than the internodes (the uppermost reaching the inflorescence), sharply pointed, herbaceous, with parallel veins | Cauline, more or less adnate to the stem, sharply pointed, herbaceous with hyaline margins, green, pink or brownish | Cauline, herbaceous, very thin, acute, more or less adnate to the stem | |
| Leaves | Unknown | Basal, 2–3, petiole to 10 cm long, narrow; blade to 8 cm long and1 cm wide, narrowly lanceolate, acute | Basal, petiolate; petiole up to 9 cm long, narrow; blade ca 5 cm long and 1 cm wide, lanceolate, broadly lanceolate to oblong-elliptic, acute | Basal, petiolate; petiole 4.5 cm long, narrow; blade up to 6 cm long, probably lanceolate, acute | Single, basal, petiolate; petiole about 4 cm long, narrow; blade 5 cm long, 2.5 cm wide, ovate-lanceolate, acute, attenuate towards the base | Basal, fugaceous, otherwise unknown | Basal, petiolate; petiole 1.5–2 cm long | Basal, 1–2, petiolate; petiole 0.8–4 cm long, narrow; blade 1–4 cm long and 0.7–1 cm wide, ovate-lanceolate, acute | |
| Inflorescence | 4–5 cm long, laxly 6–8-flowered | 3–4 cm long, laxly 6–8-flowered | 5–23 cm long, 8–19-flowered, lax, secund | 2–10 (13) cm long, (3) 4–10 (16) flowered, rather lax to relatively dense, subsecund | 9 cm long, ca 20-flowered, lax | 1.5–2 cm long, laxly 2- or 3-flowered, the axis glandular | 1–7 cm long, (1) 2–16-flowered, dense, sometimes capitate, to lax | 2–10 (13) cm long, 3–9-flowered, lax | |
| Flowers | Middle-sized, lip white, sepals dull pinkish-white with prominent brownish nerves | Rather small, lip white with dull yellowish throat, sepals dull pinkish or brownish with prominent nerves | Small, tubular, perianth brownish-red or grey-green, lip white, sometimes with a green base | Small, tubular, with slightly divergent apices, perianth green, pink or reddish, lip white with 5 olive-brown nerves or greenish-yellow front | Horizontal, tiny, tubular | Small, tubular, white, perianth segments longer than ovary | Tubular, apical ones erect, tubular, divergent at the apex only, glabrous; usually white with green nerves on petals, rarely dull white or white with pink deposit, or yellow | Tubular, divergent at the apex, perianth dull, greenish-white to pale pink or pinkish red-brown | |
| Floral bracts | Slightly longer than ovary, glabrous, herbaceous with hyaline margins, with 3 prominent nerves, dull pinkish-white | Distinctly longer than ovary, glabrous, herbaceous with hyaline margins, with 3 prominent nerves, dull pinkish or brown-ish | 9–20 mm long, broadly lanceolate, acute, herbaceous | 5–16 mm long, ovate-lanceolate, acuminate, herbaceous, glabrous | 5–7 mm long, ovate, apex obliquely lanceolate, acute, herbaceous, glabrous | 10 mm long, about twice as long as the ovary, thin, herbaceous, semitranslucent, acuminate, with 3 branched nerves | 5–21 mm long, as long as or 1.5 times longer than ovary, ovate to broad lanceolate, acute, semi-transparent, thin, herbaceous with hyaline margins | 7–22 mm long, about 1.5 times longer than ovary, ovate-lanceolate, acute, thin, semi-transparent, pale green, usually 3-nerved | |
| Ovary | 6 mm long, twisted, glabrous, dull pinkish-white | 5 mm long, twisted, sparsely glandular, dull pinkish to brownish | 5–10 mm long, cylindrical, glabrous, or sparsely glandular at the apex only | 3–9 mm long, narrowly cylindrical, apically bent forwards, sparsely glandular at the apex | 4–6 mm long, cylindrical, glabrous | 5 mm long, sparsely glandular | 4–7 mm long, slender, glabrous | 4.2–11 mm long, cylindrical, narrow, glabrous or almost glabrous | |
| Dorsal sepal | 15 mm long,1.6 mm wide, linear with lanceolate, acute apex, 1-nerved | 9 mm long, 1.4 mm wide, linear with lanceolate, acute apex, 1-nerved | (4.2) 7–10.4 mm long, 1–2 mm wide, lanceolate to linear-lanceolate, concave above the base, broadest near the middle, glabrous or sparsely glandular at the base, 1 or obscurely 3-nerved | 5–8 mm long, 1–1.9 mm wide, linear-lanceolate, concave above the base, 1-nerved | 5 mm long, 1 mm wide, oblong-lanceolate, obtuse, thin, semi-transparent, 1-nerved | 6 mm long, 1.5 mm wide, lanceolate, acute, glabrous, indistinctly 3-nerved | 9.5–14 mm long, 1.6–3.2 mm wide, usually lanceolate to oblong-lanceolate, broadest in the middle or beneath, acute, near the apex slightly concave, 1- or rarely 3-nerved | 4.3–7.2 mm long, 1.1–2 mm wide, usually broad- or oblong-lanceolate, acute, rarely obtuse, 1-nerved | |
| Petals | 11 mm long, 0.3–0.4 mm wide, almost filiform, acute,1-nerved, adnate to the dorsal sepal forming a prominent galea | 7.2 mm long, 0.5 mm wide, linear, acute, 1-nerved, adnate to the dorsal one forming a prominent galea. | (4.1) 6.7–9 mm long, (0.5) 0.8–1.2 mm wide, linear to narrow-spathulate, widest in the upper part, rounded or obtuse at the apex, sometimes on the outer margin near the apex minutely dentate | 4.5–7.3 mm long, 0.5–1 mm wide, linear-oblanceolate, widest in the upper part, subacute, 1-nerved | 6 mm long, 1.3 mm wide, oblong, subobtuse, slightly asymmetric, 1-nerved | 5 mm long, 1.2 mm wide, indistinctly falcate-banded, very thin, relatively broad, the apical margin erose, 3-nerved | 9.5–14 mm long, 0.8–1.2 mm wide, usually linear to linear-lanceolate, sigmoid or falcate, acute or obtuse, 1-nerved | 4–6.1 mm long, 0.8–1.5 mm wide, linear-oblanceolate or linear- spathulate, slightly sigmoid or falcate, acute or obtuse, at the apex irregular minutely indented, 1-nerved | |
| Lateral sepals | 17 mm long, 2 mm wide, oblique at the base, linear with oblanceolate apical half, acute to sub-obtuse at apex,1-nerved | 8 mm long, 1.2 mm wide, slightly oblique, linear with lanceolate, acute apex, 1-nerved | (3.8)7–9.5 mm long, (0.8)1–1.9 mm wide, linear-lanceolate, slightly falcate, acute, glabrous or sparsely glandular at the base, 1-nerved | 4.6–9 mm long, 0.9–1.2 mm wide, linear-lanceolate, acute, falcate, 1-nerved | 4.9 mm long, 0.6 mm wide, oblong-spathulate, obtuse, very thin, 1-nerved | 5.2 mm long, 1.3 mm wide, narrowly triangular, acute, glabrous, 1-nerved | 10–14.5 mm long, 1–2.9 mm wide, linear to linear-oblanceolate, acute or obtuse, broadest in the upper part, slightly sigmoid or falcate, 1-nerved | 5–6.4 mm long, 0.8–1.6 mm wide, oblong to narrowly lanceolate, acute, erect or slightly sigmoid, 1-nerved | |
| Lip | On prominent claw, constricted distinctly in the apical fifth; hypochile 13 mm long, 2.9 mm wide, widest below the apex, with prominent digitate auricles at the base, glandularin the center on the upper surface; isthmus shallow; epichile 2.8 mm long, 3 mm wide, almost rounded, sub-triangular at apex, with slightly undulate margins | On prominent claw, constricted distinctly in the apical third; hypochile 7 mm long, 3.2 mm wide, widest at the apex, with prominent digitate auricles at the base, glandular on the upper sur-face; isthmus acute; epichile 3.2 mm long, 4.1 mm wide, almost rounded to slightly transversely elliptic, rounded at the apex, with prominently crispate margins | On claw, divided into hypo- and epichile; in natural position canaliculate with epichile erect or bent down; hypochile 3.5–6 (7.5) mm long, 1.9–4 (4.5) mm wide, more or less rectangular in outline, usually as wide at the base as near the apex, with two prominent, thickened basal auricles; epichile 2–4 mm long, 2–4.1 mm wide, elliptic to ligulate, obtuse | Shortly clawed; lamina distinctly constricted above the middle—hypochile 3.5–7 mm long, 1.8–4 mm wide, shallowly constricted above the basal auricles, than oblong-obovate, widest above the middle, with rounded lateral margins; epichile 1.7–3.7 mm long, 1.9–3.4 mm wide, elliptic, rounded in the apex, irregularly denticulate | Shortly clawed; claw 0.6 mm long, narrow; lamina constricted near apical third—hypochile 4 mm long, 2.2 mm wide, rectangular slightly thickened in the middle, the basal auricles rounded; epichile 2 mm long, 1.9 mm wide, rounded, minutely dentate, papillate, subobtuse | Rather narrow, oblong, with distinctly dendritic proliferation of nerves, shortly clawed, constricted near the middle—hypochile 3 mm long, 2.1 mm wide, at the base the upper surface glandular with two fleshy, very small auricles bent toward the apex of the lip, hypochile gradually tapered to epichile, more or less rhombic, the side lobes triangular; epichile 2 mm long and wide, oval, obtuse, the margin very minutely but distinctly erose | Usually distinctly constricted below the middle, on long claw agglutinate to the lateral sepals—hypochile 5.5–12.5 mm long, 1.8–5.5 mm wide, greatly varied in shape -oblong-triangular, obovate lanceolate, ovate; broadest below apex at the base two, fleshy, pubescent auricles, in the center slightly thickened and pubescent, side lobes rounded, rarely triangular, sharply pointed agglutinate to the clinandrium; epichile 2–5 m long, 2–4.6 mm wide, reniform or cordate, obtuse or sometimes acute, broadest at the base, papillate usually irregularly indented, sometimes folded; if the lip undivided—usually oblanceolate, obtuse below the center with small lobes | With distinct constriction in the lower part dividing into hypo- and epichile. Epichile more or less folded, distinctly smaller than hypochile | Shortly clawed; claw narrow, linear; lamina indistinctly divided, in natural position canaliculate, erect with epichile strongly bent down- hypochile 2.9–4.5 mm long, 1.5–2.1 mm wide, 5-nerved, rectangular, at least 2 times longer than wide, truncate at the base, with slightly thickened auricles, in the center slightly thickened; epichile 1–2 mm long, 1–1.9 mm wide, 3-nerved, lingulate, ovate to transversely ovate, acute or obtuse, minutely irregular indented, papillate |
| Gynostemium | 13 mm | 7 mm | 7–8 mm | 3.5–5 mm | 3.5 mm | 2 mm | 7–13.5 mm | 4–5 mm | |
| Viscidium | 1 mm | 0.5 mm | 0.4 mm | 0.4 mm | Not found | 1 mm | 0.8–1 mm | 0.5 mm | |
| Dataset | No. of Taxa | Total Characters | Constant Characters | Informative Characters |
|---|---|---|---|---|
| ITS1-5.8S-ITS2 | 59 | 703 | 466 | 183 |
| trnL-trnF region | 48 | 1582 | 1228 | 150 |
| S. bajaverapacensis sp. nov. | S. crenulata | |
|---|---|---|
| Stem | 10 cm high, erect, delicate, completely glabrous, 3 sheaths | 6–20.5 cm high, erect, delicate, glabrous, whitish or brownish, 4–8 sheaths |
| Sheaths | Cauline, ovate-lanceolate, acuminate, glabrous, semi-transparent | Cauline, more or less adnate to the stem, sharply pointed, herbaceous with hyaline margins, green, pink or brownish |
| Leaves | Ca. 6.2 cm long and 0.8 cm wide, elliptic, microscopically obliquely acute | Basal, petiolate; petiole 1.5–2 cm long, blade unknown |
| Inflorescence | Ca. 2.5 cm long, laxly 5-flowered | 1–7 cm long, (1)2–16-flowered, dense, sometimes capitate, to lax |
| Flowers | More or less widely opened at the apex, white, sepals and petals with prominent, grayish central vein, lip pure white with pinkish suffusion in the center and grayish central vein | Tubular, apical ones erect, divergent at the apex only, glabrous; usually white with green nerves on petals, rarely dull white or white with pink deposit, or yellow |
| Floral bracts | 10 mm long, obliquely ovate, long-acuminate, with 3 prominent veins, glabrous, semi-transparent, whitish | 5–21 mm long, as long as or 1.5 times longer than ovary, ovate to broad lanceolate, acute, semi-transparent, thin, herbaceous with hyaline margins |
| Ovary | 4–5 mm long, glabrous, white | 4–7 mm long, slender, glabrous |
| Dorsal sepal | Free part 9 mm long, 2 mm wide, lanceolate, acute, basally connate with the gynostemium, somewhat concave in the center, 3-veined, glabrous | 9.5–14 mm long, 1.6–3.2 mm wide, usually lanceolate to oblong-lanceolate, broadest in the middle or beneath, acute, near the apex slightly concave, 1- or rarely 3-nerved |
| Petals | 8.5 mm long, 1.5 mm wide, linear-oblanceolate, acute, subfalcate, adnate to the dorsal sepal forming a kind of galea, 1-veined | 9.5–14 mm long, 0.8–1.2 mm wide, usually linear to linear-lanceolate, sigmoid or falcate, acute or obtuse, 1-nerved |
| Lateral sepals | Free part 8 mm long, 1.5 mm wide, linear, acute, subfalcate, 1-veined, glabrous | 10–14.5 mm long, 1–2.9 mm wide, linear to linear-oblanceolate, acute or obtuse, broadest in the upper part, slightly sigmoid or falcate, 1-nerved |
| Lip | Clawed; claw 3 mm long, narrow, adnate to the sepaline tube; blade 11 mm long in total, 3.2 mm wide when spread, elliptic-ovate in general outline, basally shortly auriculated, auricles somewhat thickened, apical third of lamina somewhat undulate along margins, apex obtuse, multiveined, thin, glabrous | Clawed; claw 3 mm long, narrow, adnate to the sepaline tube; blade 9 mm long in total, 4.2 mm wide when spread, multiveined; hypochile deltoid, 5 × 4.2 mm, basally auriculated; epichile 4 × 2.9 mm, logulate, obtuse, margins crenulate. |
| Gynostemium | 11.5 mm | 9 mm |
| Viscidium | 1.1 mm long | 0.9 mm |
| Name of Sample | No. GenBank of nrITS | No. GenBank of trnL-trnF Region |
|---|---|---|
| Schiedeella bajaverapacensis | ON854135 | ON862914 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Archila Morales, F.L.; Lipińska, M.M.; Dudek, M.; Szlachetko, D.L. Schiedeella bajaverapacensis (Orchidaceae, Spiranthinae), a New Orchid Species from Guatemala. Int. J. Mol. Sci. 2023, 24, 5362. https://doi.org/10.3390/ijms24065362
Archila Morales FL, Lipińska MM, Dudek M, Szlachetko DL. Schiedeella bajaverapacensis (Orchidaceae, Spiranthinae), a New Orchid Species from Guatemala. International Journal of Molecular Sciences. 2023; 24(6):5362. https://doi.org/10.3390/ijms24065362
Chicago/Turabian StyleArchila Morales, Fredy L., Monika M. Lipińska, Magdalena Dudek, and Dariusz L. Szlachetko. 2023. "Schiedeella bajaverapacensis (Orchidaceae, Spiranthinae), a New Orchid Species from Guatemala" International Journal of Molecular Sciences 24, no. 6: 5362. https://doi.org/10.3390/ijms24065362