A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia


School of Biological Sciences, University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia
Diversity 2019, 11(8), 123; https://doi.org/10.3390/d11080123
Submission received: 25 June 2019
/
Revised: 11 July 2019
/
Accepted: 16 July 2019
/
Published: 26 July 2019
(This article belongs to the Special Issue The Ecology and Diversity of Orchids)
Abstract
:The pollination of 20 common terrestrial orchids was studied in a 60-ha urban banksia and eucalypt dominated woodland in Western Australia. Five years of data (24,000 flowers, 6800 plants) measured fruit set relative to floral areas, capsule volumes, climate, phenology, pollination mechanisms, disturbance tolerance and demography. Pollination varied from 0–95% of flowers, floral displays from 90–3300 mm2 and capsules from 15–1300 mm3 per spike. Pollination traits strongly influenced outcomes, with self-pollination highest (59—95%), followed by sexually deceptive autumn or winter-flowering (18–39%), visual deception (0–48%) and sexually deceptive spring-flowering (13–16%). Pollination was limited by drought in autumn or spring and cool winter temperatures. Some orchids were resilient to drought and one formed seed after the leaves withered. Plant density had the greatest impact on fruit set for orchids forming large groups, especially for sexually deceptive pollination. Consequently, small group average (SGA) pollination was up to 4× greater than overall averages and peak seed production occurred in the best locations for genetic exchange and dispersal. SGA rates and seedpod volumes were strongly linked to clonality, but not to demographic trends. Resource competition limited flowering at higher plant densities and competition within spikes resulted in smaller, later-forming seedpods. Pollination data from co-occurring common orchids identified five evolutionary trade-offs linked to pollination, provided baseline data for rare species and revealed impacts of changing climate.
1. Introduction
Orchid pollination and the subsequent dispersal and germination of seeds are key limiting factors that determine the persistence and spread of orchids in natural habitats [1,2,3,4,5]. Pollination is more complex in orchids than in most other plants, due to highly specialized flower structures which encourage cross pollination and are linked to relatively specific interactions with insects [6,7,8]. It is normally expected that orchids with nectar should have higher rates of pollination than non-rewarding (visually deceptive) species [1,9,10,11]. However, deceptive means of attracting insects without food rewards are more common, occurring in an estimated one-third of all orchid species [10,12,13,14].
Australia is a global center of diversity for plants with deceptive pollination [15,16]. Orchids that attract insects by visual deception, exploit insect color perception biases and often resemble other flowers that contain nectar or pollen rewards and are scented [17,18,19,20]. In these orchids, some insect specificity can occur because the mimicked flowers are preferred by particular pollinators, but these usually include multiple categories of insects (Table A1). Detailed studies of Australian visually deceptive orchids, such as Thelymitra and Diuris species, found them to attract a suite of pollinators associated with co-flowering plants in other families of similar appearance [17,19,21,22,23]. Schiestl, Tremblay et al. and Kagawa and Takimoto discussed the ecology and evolution of plants with deceptive pollination which include the following elements: (i) orchid species should be less common than mimicked species, (ii) polymorphism of floral characters has a selective advantage by confusing pollinators and reducing learned avoidance responses, (iii) orchid floral displays are often larger and more colorful than the mimicked species, and (iv) the impact of orchids on pollinators is expected to be small [12,13,24].
Existing information about pollination of the main orchids included in this study is listed in Table A1. Only Eriochilus dilatatus, Pyrorchis nigricans and two primarily self-pollinating orchids (Microtis media and Disa bracteata) produce small amounts of nectar. Half of the total species have visual deception (9 out of 20) and five have sexually deceptive pollination by pheromone production. The remaining four species are primarily self-pollinated. Despite the fact that pollinators can be very difficult to confirm from observations [25], their identity has been established in most cases by detailed studies extending back over 100 years (see Table A1).
The southwest of Western Australia (WA) is recognized as a global diversity hotspot with exceptionally high plant species richness and turnover [26,27]. This is linked to areas with long-term climate stability, deeply leached and weathered soils and many species with adaptations to grow in these highly infertile soils [27,28,29]. However, hotpots also tend to occur in areas where complex landforms, soils and hydrology support many specialized plants and function as refugia from climate change [27,30]. Western Australia is the most important global center for evolution of new strategies for plant mineral nutrition [30]. This region also has a higher than expected diversity of terrestrial orchids, despite their exclusion from the arid interior of Australia [30,31,32]. This study occurred in banksia woodland, a threatened plant community where it is especially important to monitor population trends for important families of plants and identify processes which impact on their ecological sustainability [33,34]. The overall objectives of this study were to determine typical pollination rates and annual variations in these rates for common orchid species in an urban nature reserve. Specific objectives were to measure the floral area and fruit set of orchids, as well as the chronology of their flowering and fruiting relative to pollination traits (syndromes). The effect of local plant density on pollination and climactic conditions on flowering and capsule production were also investigated and associated insects observed. Finally, consequences of reproductive traits for each orchid were considered, relative to their population trends in an isolated urban vegetation remnant.
2. Materials and Methods
2.1. Study Site
The study was conducted at Warwick Conservation Area (WCA, Warwick Open Space) in the Swan Coastal Plain bioregion of Western Australia (–31.839124° S, 115.816894° E). This 60-ha urban reserve is in the City of Joondalup of the Perth Metropolitan Region. Vegetation consists of banksia and eucalypt woodland on Spearwood Sand Dunes with underlying Tamala limestone [35]. Banksia woodlands on the Swan Coastal Plain are sclerophyllous plant communities with an overstory dominated by Banksia species, especially B. attenuata R.Brown and B. menziesii R.Brown, along with Eucalyptus species, especially E. marginata Sm. [33]. The understory has a high diversity of shrubs, herbs, geophytes and sedges. Substantial areas of these communities have been lost to urban development and they face other threats such as water table drawdown, Phytophthora dieback disease, weeds and increased fire [33,34]. The study area is bound by 4 major roads and is further subdivided by an internal road, three sporting complexes and a school. However, this site is in relatively good condition and serious weeds are the target of ongoing management by the City of Joondalup and hand weeding by volunteers.
2.2. Study Species and Survey Methods
The study site (WCA) has well-studied flora and vegetation with over 200 spp. of flowering plants, including 31 taxa of terrestrial orchids [36]. All orchid species present at WCA were subject to detailed study, except for those that appear to be locally extinct (Caladenia marginata, Thelymitra crinita), or are locally rare (C. falcata, C. hirta, C. reptans, Cyanicula gemmata and Prasophyllum elatum), leaving 20 species in total as the main study species (see Table A1). These are illustrated in Figure 1, with their full names and ecological information in Table A1. Limited data was also available for two other species Pterostylis recurva, which was uncommon, and Leptoceras menziesii, which primarily flowers after fire (Table A2). Observations of flowering and pollination were from 2008 to 2018, with the majority of data collected in 2016, 2017 and 2018. In total 4235 individual flowering plants, 24,332 flowers and 5234 capsules were included in this study (Table S1). All orchids in this study produce 1 spike per flowering plant. Common orchids were only observed in selected areas, while all known flowering plants were assessed for uncommon species. Three species were studied over multiple nature reserves to obtain sufficient data (Table S1). Species that were disturbance opportunists were also studied in adjacent small patches of remnant vegetation (< 100 m from the reserve) including the median strip of adjacent road reserves (Caladenia latifolia, Disa bracteata, Microtis media, and Pterostylis ectypha).
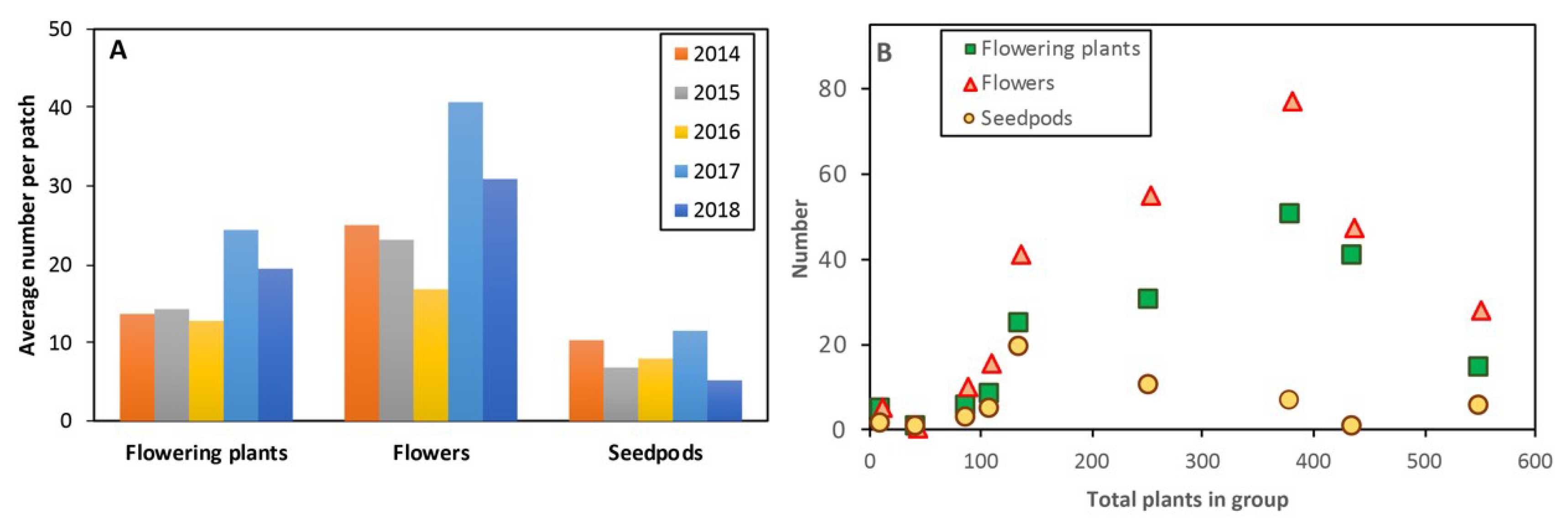
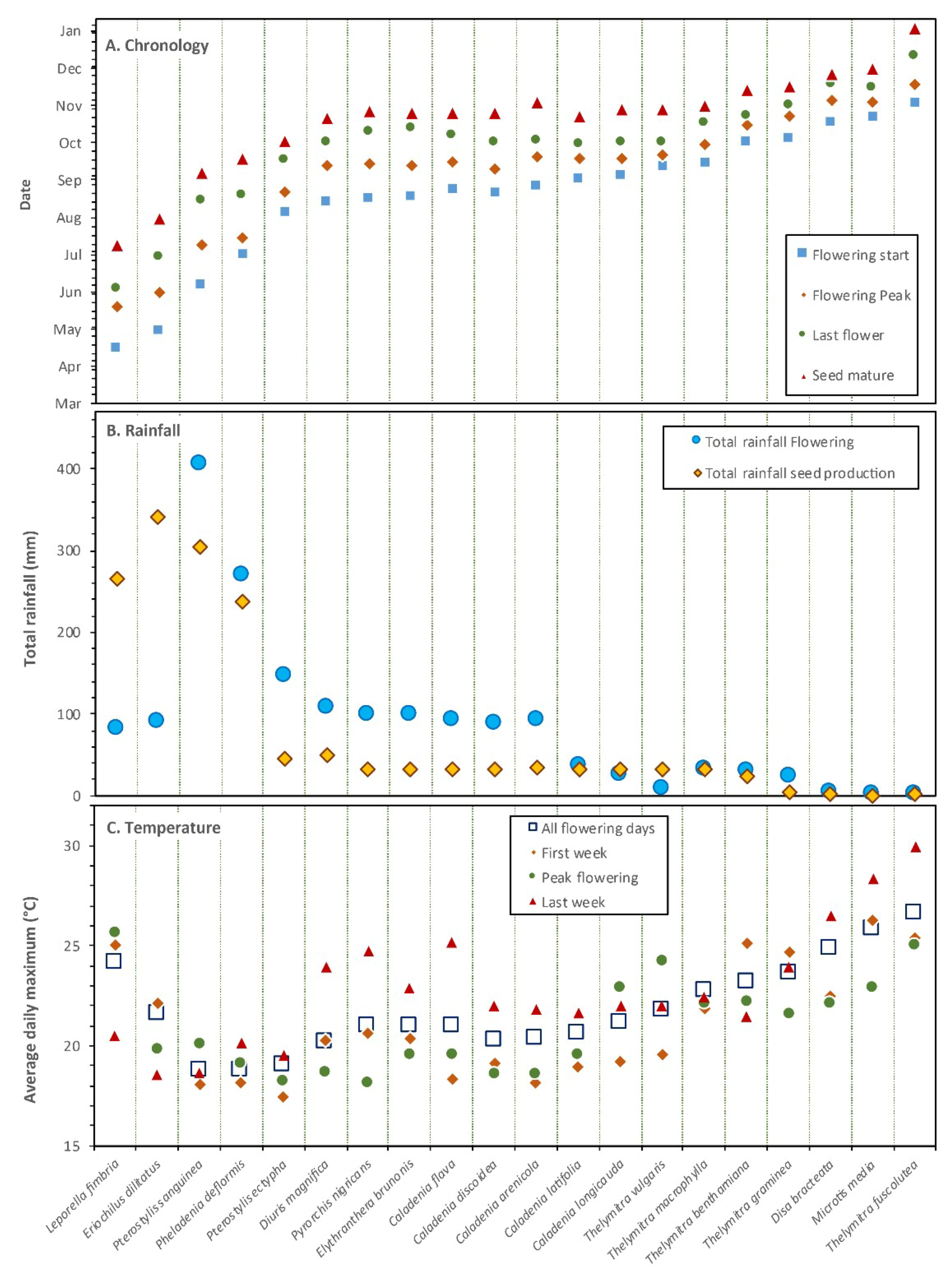
Flowering and pollination data were obtained from flowering plants within an area where orchids were aggregated and separated from other groups by at least 5 m. Areas observed were consistent within species, most were 0.5–5 m in diameter, but T. macrophylla occurred in groups up to 30 m across. Study locations were marked using GPS waypoints (± 5 m) and visited multiple times each year to count flowering plants, non-flowering plants and flowers in groups. Preliminary data (Figure 2) showed that plant density was important, so a wider range of plant numbers in similar-sized groups were sampled when possible. Annual variations in population sizes were determined for three orchids with relatively localized distributions (Figure 3). Most other orchids were too widespread to count, but the proportion of flowering plants of all species were determined in 2018. For each species, dates for the start, peak and end of flowering, as well as seed release were recorded (Figure 4, Table A1), using observations and photographic records, primarily from 2018.
Plant height, flower numbers, as well as flower and leaf areas were determined from 10–30 plants to allow plant size and floral area to be compared (Table A1). These were measured to the nearest 0.5 mm using live plants, photographs with a ruler included, or herbarium specimens from the local area. Floral area was estimated for the front view of flowers as tepal length × average width, summed for all tepals, then added to lip height × average width (if relevant). Fully overlapping flower parts were not measured and the area of Pterostylis sp. flowers was estimated as average width × height since most tepals were fused together. Upper flowers were often smaller than lower flowers in spikes, so both were measured to obtain average sizes. Leaf areas were estimated for flowering plants from the length × average width × leaf number (1 in most cases). Plants were designated as clonal if they normally produce more than one tuber per plant per year. They are unconnected, so were assessed as individuals (orchids forming a group may not all be the same genet).
In this study, fruit set is defined as the proportion of seed capsules per flower that reach full maturity, which may be less than the pollination rate due to seed abortion and can result from insect-pollen transfer or self-pollination. Seed viability was not measured, but is known to be high for many of the species studied [37,38]. This is not always the case elsewhere [39]. Capsules were counted on 4235 spikes (Table A2). Only mature capsules were counted, as some that appeared pollinated later aborted. The length and width of capsules from 22 species were measured with digital callipers to the nearest 0.1 mm including some from the bottom and top of spikes (oldest and youngest). These were primarily from WCA in 2018, with 30–170 capsules per species for most species (Table A2). Capsule volumes are highly correlated with seed numbers, with up to 80,000 seeds in large Caladenia sp. capsules [40]. Capsule volumes were calculated using the formula for a prolate spheroid (=3/4πab2, where a is the short and b the long axis radius) and multiplied by average numbers per plant to determine the reproductive output of species (Table A2). Data are presented relative to numbers of both flowering plants and total plants.
Rainfall data was obtained from a rain gauge at our home < 1 km from the study site and the Wanneroo weather station 11 km away [41]. These were in close overall agreement (± 2%), but both had 2 to 14-day gaps in records so were consolidated. Rainfall was summed for the flowering period and fruit set period for each orchid (Figure 4A,B). Flowering chronology data in Table A1 was used to calculate the average daily maximum temperature during the total flowering period, as well as the first, last and middle flowering week for each orchid (Figure 4C). Monthly differences from long-term averages in rainfall or temperatures were also noted (Figure S1).
Flower baiting for sexually deceptive orchids (Leporella fimbriata, Caladenia discoidea and C. arenicola) followed standard protocols [42,43]. This method involves moving orchid flowers or plants growing in pots to different locations to locate pollinators. Photographic records and movies (see Supplemental data) were used to estimate pollinator numbers when there were multiple responders, since these insects moved rapidly, and visits were often brief. Observations of possible pollinators and threats to orchids were also recorded (Table A1, Figure 5).
2.3. Analysis of Data
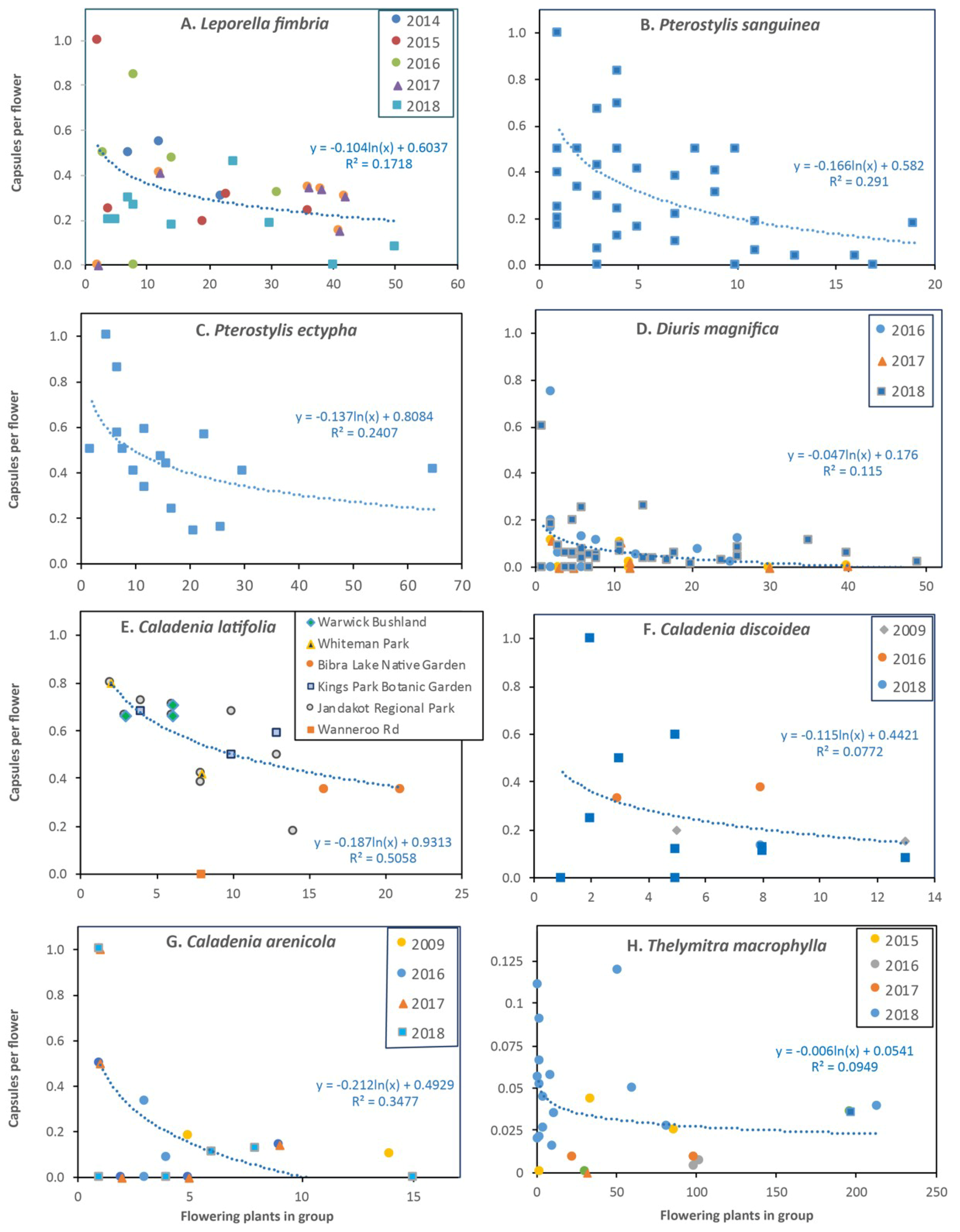
Initial comparisons of data, shown in Figure 2 for L. fimbriata, demonstrated that there were annual variations in reproductive output, as well as a strong impact of high plant density on both flowering and fruit set (Figure 2 and Figure 3). For subsequent data analysis, density dependence was visualized by graphing pollination outcomes against the total number of flowers and flowering plants in similar-sized areas which are called groups (e.g., Figure 6A, Figure S2A). For orchids with adequate sampling, plant density was investigated using Log (ln) regressions of capsules per flower and per plant. The Small Group Average (SGA) fruit set rate for each species was determined from the y intercept from the regression. This represents the pollination rate for single plants well separated from other plants. Overall averages were calculated and Large Group Average (LGA) pollination rates was also estimated from the trend line in graphs relative to the largest group observed for that species.
A metric for the pollination density effects was also calculated (SGA/overall mean fruit set). Differential rates of pollination were only calculated for orchids with aggregated distribution patterns found in 5 or more groups. The relationships between pollination outcomes and total flowering area (average flower area × average flowers per spike) was also calculated. The layout of results below follows the flowering sequence of orchids as shown in Figure 1 and Table A1, except for orchids with limited data. Orchids are referred to using species names only with their full taxonomy presented in Table A1.
3. Results
3.1. Leporella fimbriata (Hare Orchid)
This highly clonal orchid has small brown and green flowers (Figure 1A) that open in autumn, usually before leaves of non-flowering plants emerge (Figure 4). It normally grows in large groups, but over 90% of plants were sterile (Table A1) and some groups lack flowers altogether. Five years of monitoring (Figure 2 and Figure 3A) revealed that most groups were stable or increasing in numbers, but three groups declined substantially. There was a very strong negative effect of high plant density on seed production (Figure 6A, Figure S2A) with 17% of flowers and 27% of spikes setting seed overall and an SGA of 60%. There also was resource competition within large groups (Figure 7A) and within flower spikes where upper seed pods were smaller than earlier-formed seedpods (Figure 8A). Autumn drought also affected fruit set (Section 2.2).
This orchid has a well-studied sexually deceptive pollination system where male bull ants (Myrmecia spp.) are strongly attracted to flowers by a sexual pheromone as shown in Figure 1A and Video S1. Four-years of pollination baiting found there were often rapid responses by 1–10 ants for about 15 min before activity became uncommon. Moving the orchid 10 m caused renewed pollinator activity.
3.2. Eriochilus dilatatus (Bunny Orchid)
This orchid, which produces multiple small white flowers (Figure 1B), is uncommon in WCA so most flowering and fruit set data was obtained from other sites within 10 km with similar vegetation. Flowering occurred in WCA in 2016 and 2017, but not in 2018 when local populations were impacted by severe drought (Figure 4B), and a late autumn fire. Seed production (17%) was higher than most visually deceptive orchids, perhaps because flowers of this orchid contain nectar in a floral tube formed by the base of the lip [32]. Native bees and European honeybees visit its flowers (Table A1). This orchid flowers at the same time as local Ericaceae species that also have white flowers (Consostephium spp. and Astroloma spp.), so floral mimicry may also contribute to its pollination.
3.3. Pterostylis sanguinea (Banded Greenhood Orchid)
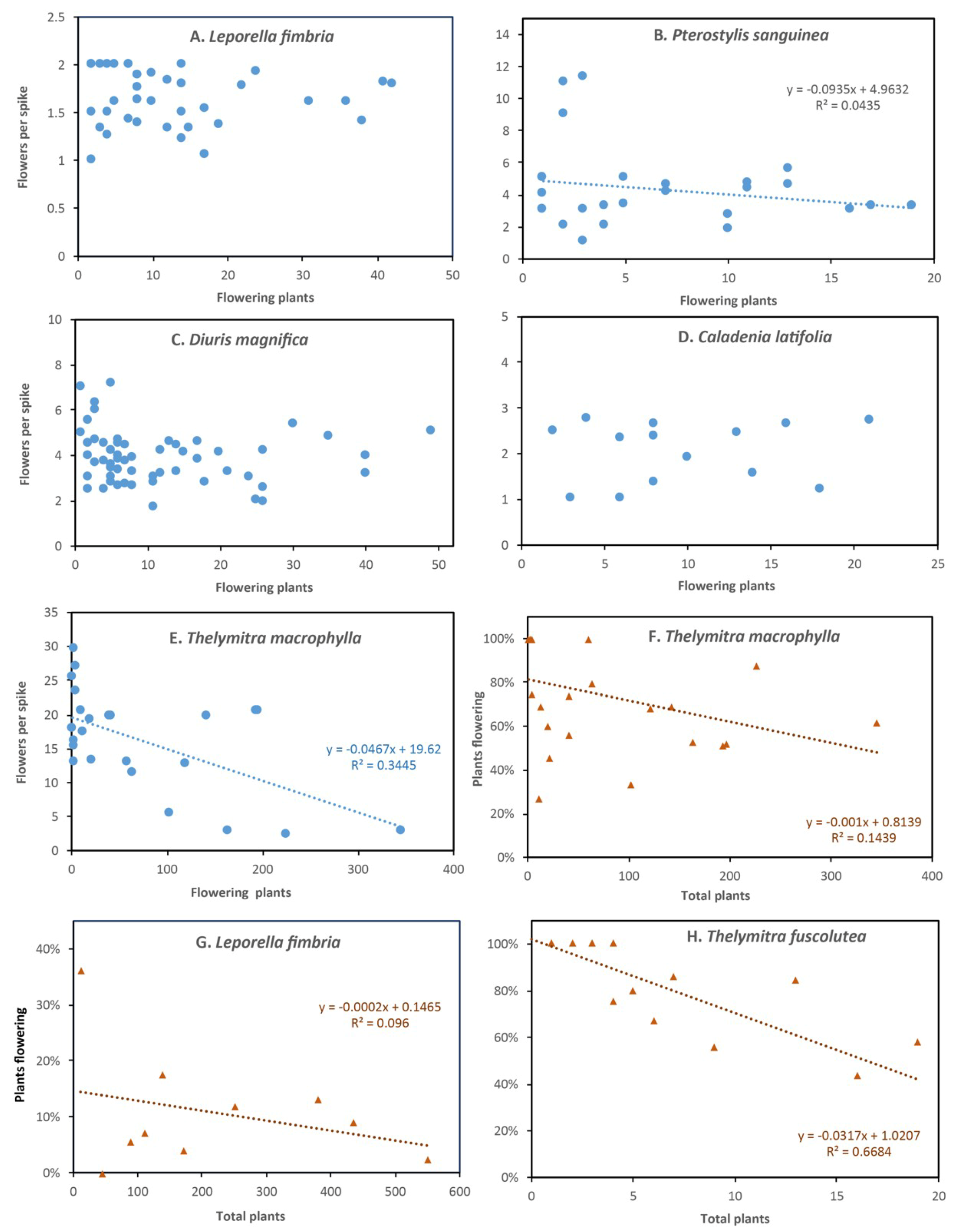
Flowers of the banded greenhood are translucent with darker red, green or brown stipes and have a floral chamber formed of fused tepals above a plate formed by two partly fused sepals (Figure 1C,D). This orchid is very common and widespread in the local area with relatively stable populations. It reproduces by seed and clonally by forming multiple tubers in winter [32]. Tubers sprout in late summer, but the shoot remains below the soil surface until late autumn when it emerges and leaves form rapidly in early winter, followed by flowering, seed formation and seed dispersal [32]. Data from 241 plants (979 flowers) established an overall pollination rate of 23% of flowers and 91% of spikes, but this decreased substantially in larger groups from an SGA value of 58% to an LGA value of about 10% (Table A2, Figure 6B, Figure S2B). Higher plant densities also reduced flowering effort (Figure 7B) providing evidence for resource competition. However, there was little effect of competition with a spike on seed production, as seedpods of this orchid were relatively large, even when 4 or more occurred on one spike (Figure 8A, Figure S3A,B).
Observations and flower baiting trials showed that fungus gnats (Mycomya sp.) were strongly attracted to flowers (Figure 1E). The pollinator is locally common and relatively unaffected by habitat condition, since the pollination rate of plants growing in pots in our backyard (about 200 m from WCA) and the study site (23%).
3.4. Pheladenia deformis (Blue Beard Orchid)
This orchid occurred in low densities at several widely separated locations. Flowers of this small orchid are bright blue in color, lack nectar and are not strongly scented (Figure 1G). They would be expected to be pollinated by native bees, or perhaps beetles (Table A1). This orchid has an exceptionally high flowering effort, where flower area often greatly exceeds leaf area (Figure 1F, Table A1). However, no seed pods formed in the monitored population in 2016 or 2018 (Table A2). Native bees were rarely observed visiting other plants when it flowered in 2018, perhaps due to cool wet winter conditions. Hand pollinated flowers were used to determine the size of seed pods.
3.5. Pterostylis ectypha (Short-Sepalled Snail Orchid)
This orchid is a member of a species complex that includes many similar forms (Pterostylis nana complex). Flowers form a small green chamber this is open at the front (Figure 1H) that is shaped to direct fungus gnats towards the lip and small column at the base of this chamber. A second unnamed member of this species complex with longer sepals also occurs in WCA. The short-sepalled form was locally common, especially in disturbed areas like the margins of bushland or within road reserves where mulch was applied. It produces single flowers from most basal leaf rosettes. The overall pollination rate was relatively high (39%) and only weakly density dependent (Figure 6C), but seedpods were small (Table A2). Pollination was not strongly affected by habitat condition since plants growing in the median strip of a major road and in pots growing in our backyard also formed seedpods. Small fungus gnats and wasps were observed near flowers, but more work is required to identify the pollinator(s) of this species.
3.6. Diuris magnifica (Pansy Orchid)
This donkey orchid is very similar to Diuris corymbosa (they intergrade within a species complex) and flowers in late winter and early spring. It has relatively large and colorful flowers (Figure 1I) in a large floral display (Table A1). This orchid has large clonal groups and most plants flower each year. It is also distributed throughout the area in hundreds of smaller groups. Large colonies contain > 50 flowering plants (Figure 5E). This orchid seems to be relatively shade-intolerant and is especially prolific in semi-disturbed areas at the margins of intact bushland (Figure 5E). Preliminary observations of co-flowering plants suggest pollination of this visually deceptive species is linked to the presence of nectar producing shrubs or climbers, including Hibbertia hypericoides, Hardenbergia comptoniana and Hypocalymma robustum. Native bees, flies and forester moths (Pollanisus sp., Zygaenidae) were the most commonly observed potential pollinators in 2018 (Figure 5A).
The very low average pollination rate (6% of flowers and 21% of spikes) was compensated for by the large numbers of flowers produced, with up to 250 flowers in a group. Pollination outcomes are strongly influenced by group size, with an SGA of 20% and an LGA close to 0% (Figure 6D). There was little effect of group size on flowering effort (Figure 7C). A separate analysis of 198 flowering plants in 12 large groups revealed that most pollination occurred at the margins of these groups (73%), providing further evidence of localized competition for pollinators. Resource competition within plants resulted in smaller flowers at the top of spikes and smaller upper seedpods in spikes (Figure 8A).
3.7. Pyrorchis nigricans (Red Beak Orchid)
This orchid is very common in Warwick bushland forming clonal groups that often include hundreds of leaves. It has dark red flowers that may include some nectar and are reported to be pollinated by native bees (Figure 1J, Table A1). It flowers prolifically after fire, but only very rarely without fire. Pollination rates for this orchid are relatively high (31% in 2018) and seed pods were the largest of any species (Table A2), which would help compensate for very infrequent flowering. The high rate of pollination supports earlier reports that flowers contain nectar (Table A1), but further investigation is required.
3.8. Elythranthera brunonis (Purple Enamel Orchid)
Populations of this orchid are relatively stable and are scattered over large areas at low densities. Most of these non-clonal orchids produce 1–3 shiny purple flowers that lack nectar and are not strongly scented (Figure 1K). Like other visually deceptive orchids, the suspected pollinators include bees and flies (Table A1). This orchid had adequate rates of pollination overall (24%), but there were insufficient plants to investigate density effects (Table A2).
3.9. Caladenia flava (Cowslip Orchid)
These orchids have highly visible yellow flowers in late winter (Figure 1L) that lack nectar and are not strongly scented. This is a relatively common clonal orchid that forms large groups where most plants do not flower. They usually occur in relatively shady, undisturbed areas and flowering is greatly enhanced by fire. Pollination success (48%) was similar to E. brunonis and C. latifolia (Table A2) and weakly related to plant density since most groups contained few flowering plants. Erickson [44] provided detailed observations of pollination by relatively large native bees, but the only potential pollinators I observed were nectar scarab beetles (Figure 5B, Table A1).
3.10. Caladenia latifolia (Pink Fairy Orchid)
This orchid has relatively large brightly colored flowers that, like C. flava, lack nectar and are not strongly scented (Figure 1P). Three color forms were present in one group (pink, white, dark pink). It tends to be most abundant in semi-disturbed habitats with only partial shade, unlike C. flava which prefers undisturbed woodland. These two species of fairy orchids are closely related and hybridize. Additional plants were studied in gardens growing under flowering native shrubs (Table S1). There were strong trends for seed production (Figure 6E) and flowering effort (Figure 7D) to be lower in large groups.
3.11. Caladenia longicauda (White Spider Orchid)
This orchid has white flowers that are similar in shape to C. arenicola, but are pollinated by visual deception (Figure 1Q). They have a large floral areas per spike (Table A1), but low pollination rates per flower (Table A2). Two subspecies of C. longicauda are infrequently present at WCA (Table S1). Fruit set by C. longicauda subsp. calcigena occurred in 2016 but not 2018, so additional data was obtained from a nearby site (Table S1). Plants formed small clumps by tuber multiplication. Hoverflies with pollinia attached have been observed in flowers of this species (Table A1).
3.12. Caladenia discoidea (Dancing Bee Orchid)
This relatively small orchid may be declining in the local area as it is absent from many suitable habitats nearby. It has 1–4 small red and greenish-yellow flowers with a broad lip (Figure 1M). Flower baiting efficiently detected pollinators both at WCA and areas where it is absent (see Video S2). When temperatures were above 20°C a small black male wasp (Thynninae) arrived in groups of up to 12, swarmed around flowers than most departed after 5–10 min. A second swarm sometimes formed after 25 min (Figure 1M). The pollinator is a Phymatothynnus sp. [45]. Despite the relatively high abundance of pollinators detected by baiting, only 16% of flowers were pollinated (Figure 6F). There was strong density dependence (Figure 6F, Figure S2E), even though groups were relatively small (SGA = 44%). A confounding factor was that the two largest groups of this orchid occurred in semi-disturbed habitats near the edge of the reserve where pollinators may be less likely to visit, especially if prevailing winds were blowing away from the bushland.
3.13. Caladenia arenicola (Carousel Spider Orchid)
This orchid produces up to 4 tall flowers with long narrow reddish-colored petals and sepals (Figure 1N). A white flowered form also occurred and one plant with a deformed flower was observed (2 lips). Population sizes have decreased substantially in the past 2 decades in most areas where it occurs at WCA, but some large groups remain. Grazing of leaves and flowers was rare, but some plants appeared to be virus-affected and failed to emerge the following year (Figure 5I). The pollinator, which is a relatively large thynnine wasp (Table A1), was not detected in WCA despite many attempts at pollinator baiting. However, the same flowers successfully attracted pollinators at another location (see Figure 1O, Figure 5C). Overall pollination rates were similar to C. discoidea, a species where pollinators were commonly detected. Low rates of pollination (13%) are partially offset by relatively large seed pods (Table A2). There was a strong effect of plant density on fruit set (SGA 49%), but no evidence of resource competition (Figure 6G, Figure S2F). Much larger seed pods were produced by hand pollinated plants in pots, which also had larger leaves and flowers.
3.14. Thelymitra vulgaris (Common Sun Orchid)
This highly variable orchid is a close relative of the larger blue sun orchids described below, but has much smaller flowers that self-pollinate (Figure 1R, Figure 5M). It has several local populations which are expanding and was first observed 15 years earlier. Seed pods are smaller than most other orchids studied, but set many capsules (94%). Resource competition within plants results in smaller upper flowers and upper seed pods, as well as some that abort (Figure 5M, Figure 8A, Figure S3E). Overall seed production was high due to capsule numbers per spike and their size (Table A2, Figure S3F). This plant flowers before the other sun orchids, so usually avoids drought stress (Figure 4).
3.15. Thelymitra macrophylla (Blue Lady Orchid)
This orchid was first observed in the local area about 10 years earlier and has a rapidly expanding population, due increasing groups sizes and locations (Figure 3B). It is late-spring flowering and is one of a number of similar sun orchids with many bright blue flowers in a spike that lack nectar and are weakly scented (Figure 1S). These flowers only open on warm days and are pollinated by visual mimicry (Table A1). The most likely mimicked species are two blue native irises, that have coinciding flowering periods and are locally common (Orthrosanthus laxus and Patersonia occidentalis). Potential pollinators were rarely observed, presumably because their visitation was rare and/or weather dependent.
Overall pollination rates were low every year (5% of flowers), but an average of 7 flowers per spike results in 24% of flowering plants setting seed and seedpods were relatively large (Table A2). Many flowers with swollen ovaries did not form seed and some may have aborted due to dry conditions. There were many non-flowering leaves, often in clumps due to clonal division. The maximum per flower SGA was very low (5.4%, Figure 6H) due to a strong negative relationship between density and pollination, which also affected the proportion of flowering plants and number of flowers per spike (Figure 7E,F). There also was competition within spikes for seed formation (Figure S3C,D).
3.16. Thelymitra graminea (Slender Blue Sun-Orchid)
This orchid is another member of the T. macrophylla complex with smaller blue unscented flowers (Figure 1U). It has narrower leaves than T. macrophylla (< 10 mm) and flowered several weeks later than its sister species. There were two expanding local populations which have a recent origin. Some hybrids between this species and T. macrophylla were present and had a similar rate of pollination. Flowering when insect activity was observed to be high resulted in 17% of flowers setting seed which is substantially higher than T. macrophylla. Density effects could not be measured. Hoverflies (Melangyna sp., Syrphidae) visited some flowers (Figure 1U).
3.17. Thelymitra benthamiana (Leopard Orchid)
This orchid has attractive yellow-green flowers with brown spots (Figure 1T) that only remain open for a few days. The local form self-pollinates with seedpods from 90–94% of flowers and these are also relatively large (Table A2, Figure 8A). Self-pollination was confirmed by placing spikes with unopened buds were in seedbags. The local population is steadily increasing both in numbers and in area, but all known plants are within 100 m of each other. Population growth trends suggest this orchid also arrived in the area within the last 2 decades. Resource limitations within plants resulted in smaller pods from later opening flowers and some that failed to set seed (Figure 8A). Seedpods opened and closed with changes in humidity so seed dispersal extended for several weeks.
3.18. Disa bracteata (South African Orchid)
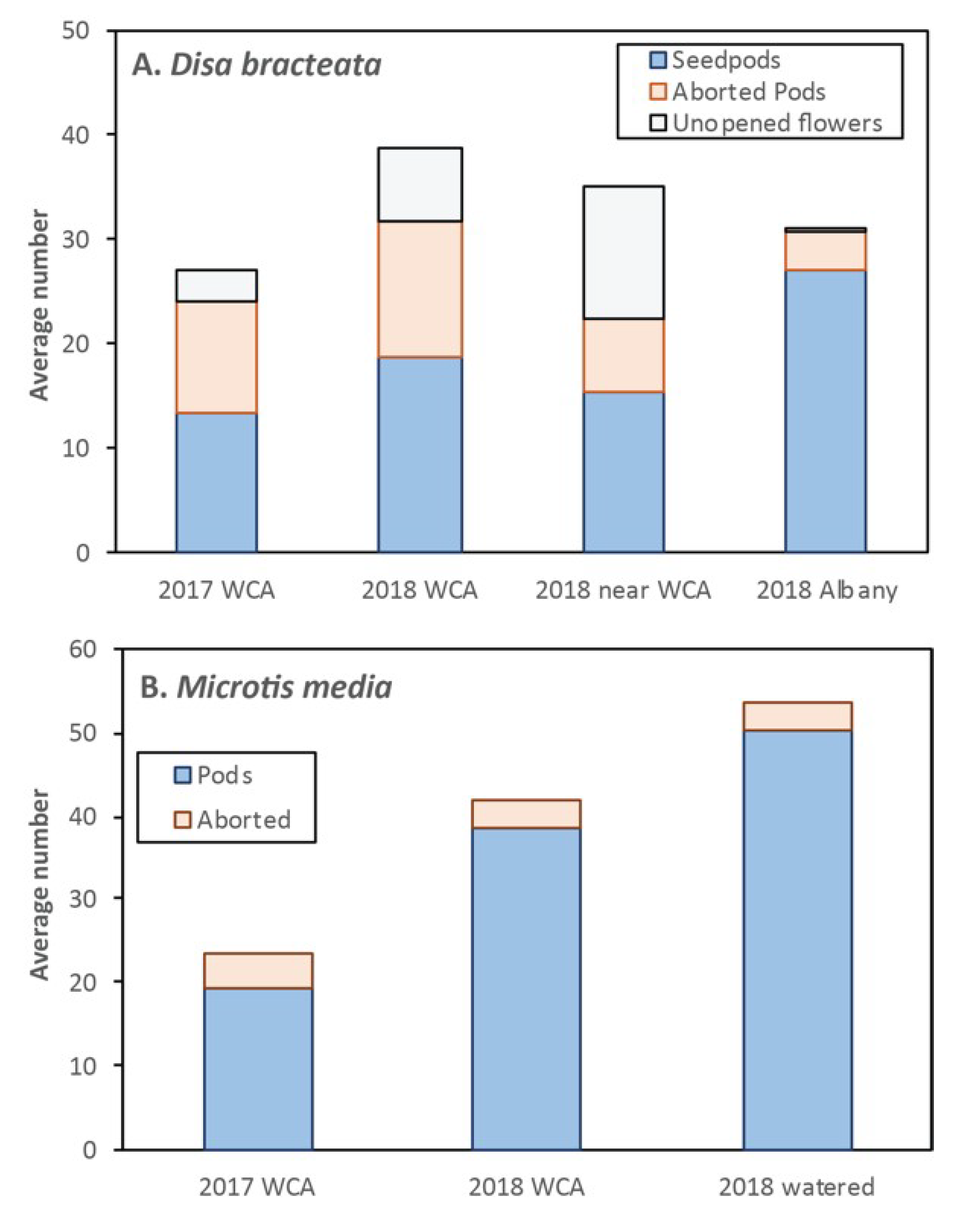
This orchid has dense spikes of very small greenish flowers (Figure 1V) that self-pollinate when pollen becomes crumbly and falls onto the stigma [32]. It occurs in both disturbed habitats and intact bushland, but is rarely common in very good condition areas (some hand removal of this orchid has occurred in the area studied). Flowers also seem to be designed for insect pollination as they have a spur that contains some nectar [46]. Visits by insects were not observed. Larger pods were formed by flowers at the base of spikes that opened first (Figure 8B), while most aborted flowers were at the top of spikes in 2018 (Figure 5N). Only 48% of potential flowers set seed, because 33% of pods aborted and 18% of flowers failed to develop (Figure 9A). The proportion of flowers producing seed was similar at WCA in 2017 (63%), but was much higher in Albany in an area where plants were watered and received more rainfall (Figure 9A). This weedy orchid has semi-indeterminate flower and seed production, since upper flower buds fail to develop in dry conditions (Figure 5N,O). Overall seed volume was about average for the species examined due to their moderate size (Table A2). There was a small plant density effect on seed production (Table A2).
3.19. Microtis media (Mignonette Orchid)
This orchid has up to 100 very small green flowers (Figure 1W) that open gradually over several weeks in late spring like D. bracteata. It was most common in semi-disturbed habitats, such as the margins of bushland areas where weeds were also present. As shown in Figure 8B, most flowers set seed due to self-pollinating (apomixis), but Microtis species have also been reported to be capable of pollination by insects, especially ants and flies (Figure 1W, Table A1). However, visitation of flowers by insects was rare. Fruit set, while potentially 100%, is usually lower due to the failure of some flower buds or seedpods to develop at the top of spikes (Figure 9B). Flowering of this species coincided with drought conditions in 2018 (Figure 4) and most had > 10 undeveloped buds at the top of spikes in natural habitats. In contrast, plants grown in a garden with added water developed seed from all their flowers (Figure 9B). Thus, this orchid has semi-indeterminate flower and seed production like D. bracteata. Seed pods were very small, and even smaller at the top of spikes (Figure 8B), but 30–70 pods in spikes resulted in moderate total volumes (Table A2). Pollination in large or small groups was similar (Table A2).
3.20. Thelymitra fuscolutea (Chestnut Sun Orchid)
This orchid was the last to flower each year and has yellow and brown spotted flowers. These are similar to T. benthamiana but remain open for weeks and are larger and darker in color (Figure 1X). They lack nectar or a clearly discernible scent and are difficult to detect visually because they resemble seedpods of other species from a distance (Burchardia congesta and Orthrosanthus laxus). The study site has one of the largest populations of this orchid in Perth and surveys in 2018 revealed twice as many flowering plants (102), as the previous year (45). These additional plants were found by extending surveys over two months since some plants started flowering after others had finished. However, the area where this plant is known to occur also increased since observations began in 2013. With one exception (rediscovered after 14 years), plants that were not observed for several years never re-emerged. In total, three groups of plants first recorded in 2013 were lost. Figure 3C shows that for areas checked consistently this species has a stable or increasing local population, but trends were different for each group. Ten flower spikes were lost due to trampling in 2018, but should re-emerge in 2019. Symptoms of possible infection by a rust fungus was commonly observed on leaves (Figure 5F inset).
This orchid flowered in very late spring (mid-November to early December) after leaves senesced (Figure 4 and Figure 5F) and seed formation in some plants continued after stems turned brown (see Figure 5K). It is one of the last to flower at a time with high activity of flies and native bees, but overall pollination rates were low (7% of flowers, 43% of plants). In total, 49 pods formed from 632 flowers on 102 flowering plants, but 4 of these were lost to grazing by insects (Figure 5G). Density effects on seed production were small due to small size of groups (< 12 plants), but there was a negative relationship between leaf density and flowering due to clumps of leaves from vegetative division (Figure 7H, Figure S2H). Competition within plants affected seedpod size (Figure 8A). Possible pollinators include hoverflies (Melangyna sp., Syrphidae) that were observed near flowers (Figure 5D).
3.21. Other less Common Orchids
Pterostylis recurva (the jug orchid) is uncommon at WCA, so was studied at another location nearby. It had fewer flowers than P. sanguinea, but had higher rates of pollination and larger seedpods (Table A2). Pollinators of this species are likely to be fungus gnats like other greenhoods (Table A1). Leptoceras menziesii (the rabbit orchid) flowers primarily after fire. Large clonal groups with hundreds of plants occur at the study site, but very rarely flower. One group that flowered in 2008 after severe local soil disturbance and provided preliminary data on seed production and seedpod size (Table A2). Hoverflies were observed visiting flowers of this orchid.
3.22. Pollination Outcomes Relative to Climate and Ecology
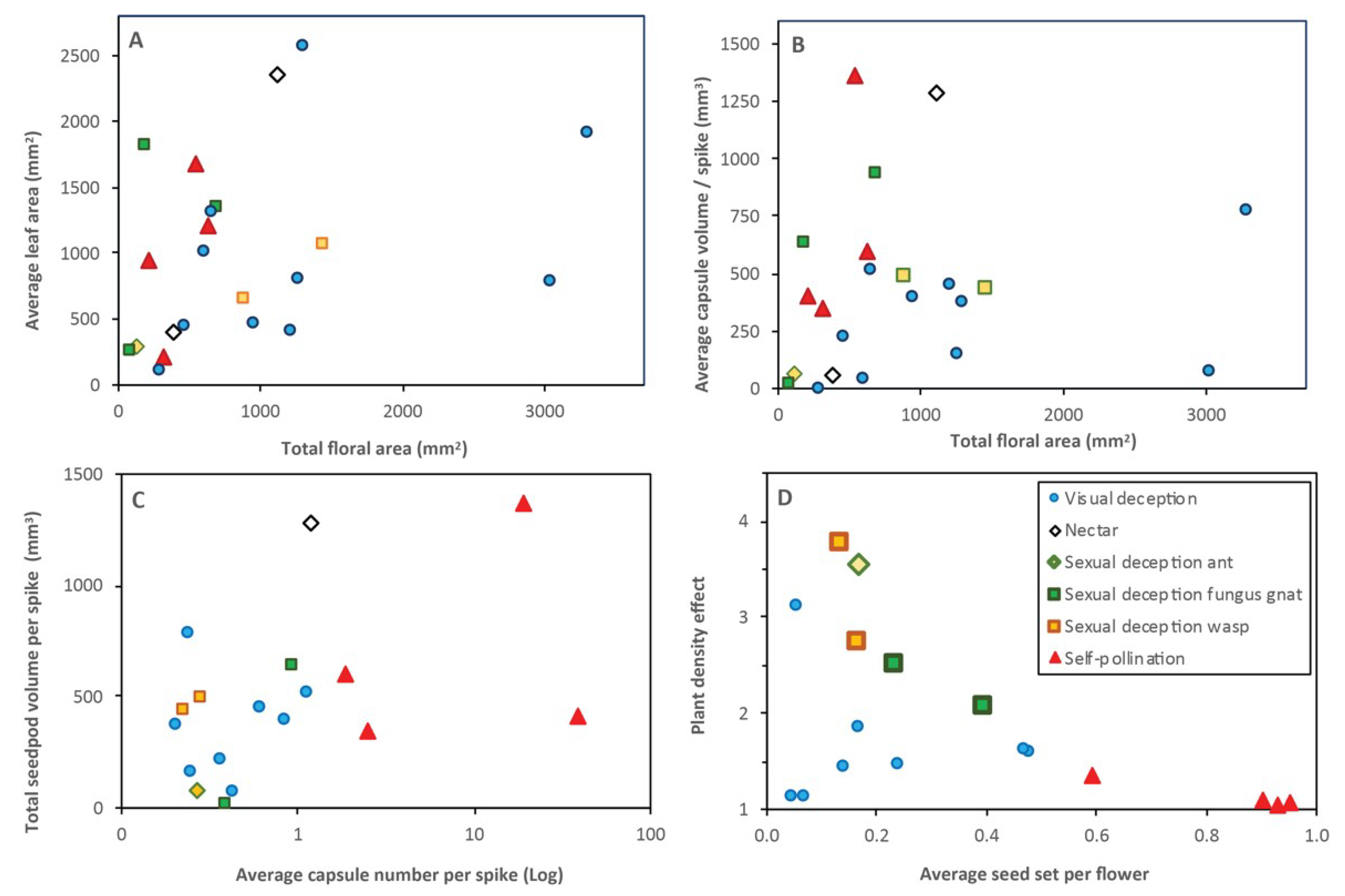
Figure 10 and Figure 11 show relationships between fruit set, leaf area, flowering effort (floral area and production) and plant density relative to pollination traits of orchids. Overall, flowering effort was highest for orchids with visual deception and lowest for self-pollinating orchids, but was not well correlated with leaf area or capsule volume (Figure 11A,B). Visually deceptive orchids had relatively large and brightly colored floral displays (Figure 1, Table A1), but most had low rates of pollination (Figure 10). The number of capsules per spike was also poorly correlated with their total volume due to differences between pollination trait categories (Figure 11C). There were different functional groups for sexual deception due to separate insect vectors and flowering times. As shown in Figure 11D, orchids with sexually deceptive pollination had very high SGA values relative to average fruit set (Caladenia spp. and Leporella sp.), while fungus gnat pollinated orchids (Pterostylis spp.) were less affected by plant density. High plant density had a lower impact on most visually deceptive orchids (Figure 11D). As would be expected, self-pollinating orchids were least affected by plant density and always produced many seedpods, but only one (D. bracteata) had relatively high total seedpod volumes (Figure 11C). When only visually deceptive orchids are compared there were weak relationships between leaf and flower area, or between seed output and flower area (Figure 11A,B).
Effects of plant density on pollination were also apparent in clonal orchids that occur in large groups (Figure 12B). As shown in Figure 12A, 4 out of 9 highly clonal orchids had very low reproductive output (P. nigricans, L. fimbriata, C. flava and Leptoceras menziesii). These flower most commonly (or only) after fire and often occur in large groups. Three clonal orchids also had low seed production due to very small seedpods (L. fimbriata and L. menziesii and P. ectypha).
Overall, there were trade-offs between seedpod sizes, flowering frequencies, flowering efforts, and pollination rates that resulted in seedpod volumes per plant that varied over orders of magnitude from 6 to 1300 mm3. Self-pollinating orchids produced the most flowers and seedpods, but these were relatively small (Figure 11C). Two orchids had much higher seed volumes than the rest. One of these produces very large seedpod volumes, but only after fire (P. nigricans) and the other was a self-pollinating weed with up to 50 seedpods per spike (D. bracteata). Overall, differences in pollination traits between orchids were the most important determinants of pollination outcomes, but plant density was also very important for some species. There also were impacts of competition for resources within groups on flowering (Figure 7) and within plants on seed production (Figure 9), but these were of lesser importance.
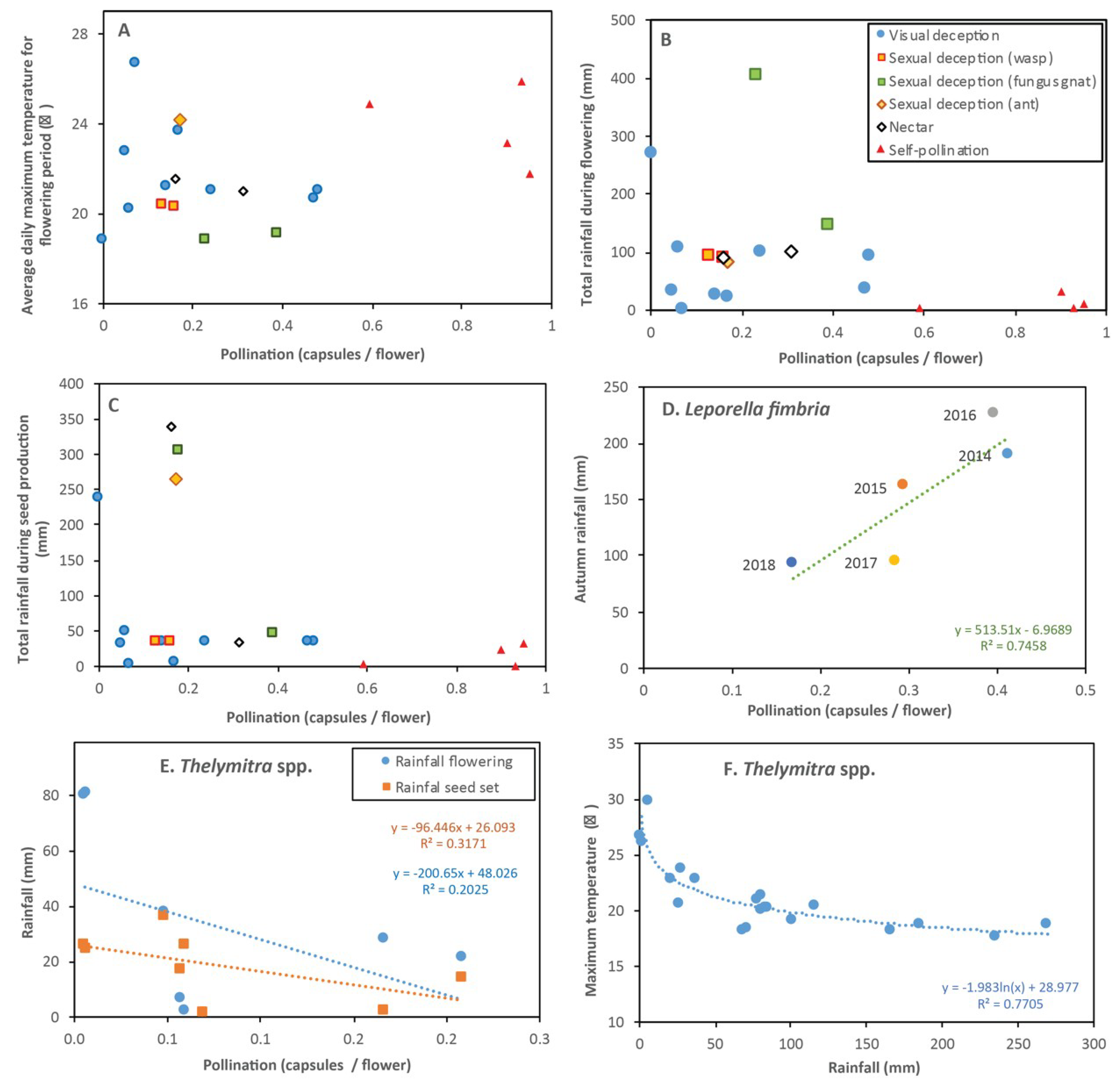
The sequence of dates for flowering, seedpod development and seed release are summarized in Figure 4A, along with prevailing climatic conditions. This sequence starts with five autumn and winter flowering orchids, followed by a peak in late winter to mid-spring when half of the orchids flowered and ending with three orchids where flowering continues in early summer. Overall, there were no clear relationships between climate and fruit set for orchids, because each functional group had a different response (Figure 13A,B,C). Orchids generally developed less seed in winter when temperatures were lowest, especially P. deformis which remained unpollinated. Low fruit set was correlated with autumn drought for the earliest flowering orchid (L. fimbriata - Figure 13D). Some orchids developed seed during severe drought in late spring (Figure 4B, Figure 13C). There seemed to be a negative relationship between rainfall and fruit set for visually deceptive Thelymitra spp. (Figure 13E), but the strong correlation between temperature and rainfall suggest this may be indirect (Figure 13F).
The main year of this study (2018) was drier in autumn and late spring than expected from long-term records (Figure S1). It was observed that native bees were much less common than expected in spring, perhaps because a late start to the growing season caused by autumn drought affected their phenology. Self-pollinating orchids can theoretically form seed with 100% of their flowers, but in reality, their pollination was limited by severe drought in late spring that caused their last flowers to abort. This was observed to a small extent in two sun orchids (T. benthamiana, T. vulgaris) and to a larger extent in two weedy orchids with many small flowers (M. media, D. bracteata - Figure 9).
Grazing of orchid seed pods and disturbance impacts on orchid reproduction were also observed, but was uncommon (Figure 5G,L). Disease symptoms caused by viruses and rust fungi were also noted (Figure 5F,I). A hot fire much later in autumn than normal (mid-May 2018) impacted on orchids with growing buds just below the soil surface. Leporella and Pyrorchis leaves were severely damaged at the edges of the fire (see Figure 5H) and many plants died in the middle of the burnt area, where tubers were smaller and less abundant than in adjacent parts of the same large group.
4. Discussion
In one of the largest orchid pollination studies ever undertaken, seed production and related factors were measured for all common orchids in an urban habitat over multiple years. Clear differences in pollination success rates were linked to plant density, flowering times and pollination traits (syndromes). Reproductive output expressed as capsule volume provided the best understanding of pollination outcomes, due to very large differences in seedpod sizes and numbers between species. When expressed this way, reproductive outputs of orchids with different pollination traits were more similar than expected. However, seed pod numbers provide a more direct reflection of pollinator activity, so are also important. Ecological factors linked to reproductive success in orchids are listed by Tremblay et al. [13]. These include inflorescence size, phenology, plant density, climate and associated vegetation, as discussed below.
4.1. Climate and Habitat Conditions
Many earlier studies have shown that orchid flowering and seed production can be strongly influenced by rainfall and temperature [13]. Temperature regulates orchid flower opening and flowering duration [22,47]. The impact of cool temperatures on pollination of P. deformis by a native bee (Halictus sp.) was also noted by Rogers [48]. In the present study different orchids flowered from autumn to early summer at the same time every year (usually within a week). These flowering times should be attuned to seasonal variations in insect availability, but some pollinators appeared at different times each year and sometimes were several months later than expected. For example, native bees and nectar scarabs were rare in spring in 2018 when a late start to autumn rainfall caused many plants other than orchids to flower late. This seem to be a major point of vulnerability for orchids which often have less flexible phenologies than other flowering plants or insects. In particular, late flowering orchids such as T. fuscolutea, M. media and D. bracteata were severely affected by drought, but still produced some seed. Brown and York also observed that severe drought had major impacts on Diuris maculata seed production [49]. Overall trends suggest a trade-off between flowering when rainfall was adequate, but temperatures were cooler, as opposed to when warmer temperatures favor insect activity, but plants were subject to drought stress during seed formation. However, some orchids were highly resilient to drought and seedpod development in otherwise dormant plants was observed for T. fuscolutea, a rare phenomenon that warrants further investigation. Similar mechanisms occur in several other West Australian plants, most notably Spiculaea ciliata (Orchidaceae) and Isotoma hypocrateriformis (Campanulaceae) that form seed in early summer using only resources stored in stems.
Several visually deceptive orchids had relatively higher overall rates of pollination. These include Caladenia latifolia which can be common in gardens under flowering shrubs. Caladenia flava also had relatively high rates of pollination, but at low flower densities. Another complicating factor that may occur in this case is that the majority of orchid seed falls close to parent plants [53], so areas which have relatively high insect activity may result in higher orchid densities, as seems to be the case for C. latifolia in gardens. With the exception of M. media, other native orchids were less well suited to exploiting artificial situations, presumably due to habitat constraints (such as compatible fungi) or dispersal limitations. It is also possible that orchids were more common in habitats preferred by their pollinators. These include visually deceptive orchids such as D. magnifica and T macrophylla which were more abundant than most other orchids. Diuris magnifica prefers relatively open areas where pollinators (native bees, flies, etc.) can also be relatively abundant [54]. One explanation for this is that orchid flowers are more visible to pollinators in open areas. This is also the case after fire, when many WA orchids flower most prolifically [32]. Fungus gnats prefer damp shady habitats (where mushrooms are also most common) and so do many greenhood orchids (Pterostylis spp.). However, orchids pollinated by fungus gnats seemed to be less sensitive to habitat conditions than sexually deceptive orchids pollinated by thynnine wasps and tend to have higher rates of pollination than other sexually deceptive or visually deceptive orchids.
Results of baiting studies, where orchid flowers are used to attract pollinators, did not agree with rates of seedpod production for the same species. In particular, there was a discrepancy between C. arenicola and C. discoidea where the apparent abundance of insect vectors differed considerably, but both species had similar rates of pollination (13–16%). Orchid pollinators are often very hard to detect because their activity is regulated by specific environmental conditions and varies considerably from year to year [55]. Thus, negative results of baiting do not provide conclusive proof that pollinators were absent, but helped to identify pollinators and provided evidence that habitat preferences of thynnine wasps may differ from the orchids they pollinate.
The iconic Perth carousel spider orchid (C. arenicola) is one of the most studied orchids in Australia, but its population dynamics are still poorly understood. At the study site it had unstable local populations due to loss of some groups of plants and establishment of others over several decades. Most missing orchids do not re-appear in later years, so were not dormant. This differs from more arid habitats in WA where dormancy lasting one or more years is common [56]. Sudden decline of local populations can occur in habitats which are in good condition. Grazing animals such as kangaroos observed in other studies [56,57] were rare at the study site. However, suspected symptoms of virus infection were sometimes observed before plants disappeared. Viruses are common in WA orchids, but usually do not cause visual symptoms [58]. There was some predation of seedpods by insects, but these impacts were relatively minor compared to other studies (e.g., Light and MacConaill [47]).
Australian orchids differ considerably in their responses to disturbance and some rapidly colonize new habitats, while others are absent even after decades [59]. This study included two orchids considered to be weed-like (D. bracteata and M. media). These are self-pollinating with prolific fruit set, despite some losses due to drought. They also have very broad fungal compatibility [37] and relatively high rates of seed germination in natural habitats [60]. However, there was no evidence that these orchids were increasing in numbers despite all of these “weedy” characteristics. Orchids indigenous to the area fall along a continuum from those that are favored by disturbance to those that are highly intolerant of it. Approximately half of them are most abundant in areas with some disturbance, such as margins of the reserve or tracks, where they probably benefit from higher light levels and lower competition. However, these areas need to be in good condition since orchids are generally absent in areas with high weed cover [38].
Orchid fruit set is a key vital statistic for managing rare orchids and outcomes for rare and common orchids relative to pollination traits are presented in Table 1. Because of limited data on the lifespans, dispersal or germination of Australian orchids, it is not possible to establish the values required for sustainable populations. However, some rare orchids have pollination rates that are much lower than expected (Table 1). There are similar numbers of rare WA orchids with generalized and specialized pollination overall and orchid pollination strategies are also not well correlated with their rarity in Europe [3,61].
Many of the orchid species studied had an extended period of seed development due to multiple flowers on spikes and erratic pollination resulting in seed dispersal lasting for weeks or months. The narrow slits on capsules were observed to open then close multiple times on different days in response to moisture or humidity and some seed remained in capsules weeks after opening. This facilitates dispersal by exposing seed to changes in prevailing wind strength and direction. Intensive observations at the study site by the author and others have documented the arrival of five orchid species into the area over two decades. Thus, effective long-range dispersal in isolated urban vegetation remnants is rare, but does occur. Several species also seem to have disappeared locally over the same time period, showing that new orchids may not persist, especially when growing outside their normal habitat types.
4.2. Plant Density Effects and Resource Limitations
Studying a specific urban nature reserve containing a wide range of orchid group sizes revealed that seedpod production for sexually and visually deceptive orchids was substantially reduced at high plant densities. This was especially noticeable for D. magnifica where most pollination occurred at the edges of large groups. Plant density impacts on seed production were stronger than the impact of climatic factors for most orchids. High plant density also reduced flowering within large groups and orchid flowers and seed pods that formed later were consistently smaller than those that formed first at the base of the same spike. This was most evident in self-pollinating orchids with many seedpods and density effects on pollination have also been observed in European and tropical orchids [62,63]. It was also noted that orchids cultivated in pots could produce larger flowers and seedpods than in situ plants. Flowers are less expensive to produce than seeds, especially if nutrients can be reabsorbed during senescence. Soil resources such as mineral nutrients and water, supplied by mycorrhizal fungi or absorbed by the orchids themselves, are normally in limited supply in West Australian soils [28,30]. This results in an access of carbon supply relative to mineral nutrients for plants. This may explain why visually deceptive orchids often produce large floral displays with low pollination rates per flower, but have similar seed volume outputs to other orchids.
Earlier studies found that orchid density is linked to pollination rates in some cases, but not in others [13]. The opposite trend, where pollination was greater in groups of orchids than for isolated plants, was noted for Elythranthera brunonis by Tremblay et al. [64], but cannot be directly be compared because areas occupied by groups of orchids in their study were much larger. It is often difficult to compare studies because population size rather than plant density is reported. A strong inverse relationship between flower density and pollination is expected for sexually deceptive pollination, due to learned avoidance responses by insects [52,65]. While sexually deceptive Caladenia species did have very high impacts of plant density on pollination, I found similar trends for visually deceptive orchids, suggesting that their pollinators also learn to avoid them, and/or are limited in abundance. Strong density-dependent effects on pollination were also observed by Dafni and Calder [66] for Thelymitra antennifera in Eastern Australia, where it was linked to very low rates of pollinator visits (flies and bees). Similar trends occurred for the European orchid Neottia ovata [67,68]. Visits by pollinators tend to be brief and inconsistent so often only a few flowers on spikes set seed [7,13,69]. Visually deceptive orchids are more likely to saturate pollinators at higher flower densities when co-occurring rewarding plants are uncommon [17,21,23,70]. It also seems likely that climatic conditions interact strongly with the other factors in Table 2, due to correlations with temperate or rainfall in some cases. Thus, limited numbers of pollinators, their learned avoidance responses, resource limitations and adverse climatic conditions can all lead to low rates of orchid pollination, and these mechanisms often occur together. Separating the factors in Table 2 requires more detailed data on pollinator numbers and their behavior.
A logical consequence of density dependence is that pollination tends to occur in sparsely occupied areas where it is needed most, so low overall rates of pollination may not be detrimental to orchids. Seed produced in the middle of large groups is most likely to be wasted, because most will fall were plants are already common. In contrast, seed produced at the margins of large groups or by isolated plants is more likely to found new groups or expand existing ones. It is also more likely to lead to genetic exchanges between different genotypes, especially for clonal orchids. Consequently, SGA pollination rates (the intercept from a plot of pollination vs plant density), will be more informative than the overall average as a vital statistic for orchid ecology or conservation. However, overall average pollination rates will be similar if plants are not aggregated into groups. The best approach may be to use several different metrics to compare pollination rates for locations or species (see Table A2). Both SGA and average or LGA values can be calculated relative to the number of flowers, spikes, flowering plants and total plants (where relevant). Comparing these metrics can help to separate the effects of flower production, pollinator abundance and plant density on reproductive outputs. Many rare orchids do occur in dense aggregations [56], so their critical pollination rates should be reconsidered using SGA and LGA values. Annual variability in pollination rates was substantial in some cases, but was found to be less important than plant density. Nevertheless, at least four years of data was needed to measure population sizes for rare Australian orchids [56] and to identify climatic influences on pollination in the present study.
4.3. Pollination Outcomes Relative to Orchid Ecology and Evolution
Orchids included in this study belong to different categories and subcategories based on their reproductive strategies, categories of insects and flowering times (Table 1). These include (i) visual deception with bright colored flowers that mimic native peas (D. magnifica) or irises (Thelymitra spp.), (ii) sexual deception with relatively wispy flowers with neutral colors that attract male wasps (C. arenicola, C. discoidea), (iii) sexual deception attracting winged male ants (L. fimbriata), (iv) sexual deception with complex green-colored flower traps that attract fungus gnats (Pterostylis spp.) and (v) self-pollinating flowers with abundant very small greenish flowers (M. media, D. bracteata), or small colorful flowers (Thelymitra spp.). Visual deception subcategories include flowering in mid-winter—where temperature had strong influence on pollination, late winter to mid spring—where flower densities were low, but pollination tended to be relatively high and spring to early summer—where floral displays were relatively large, but pollination rates were low. These categories and subcategories result in trade-offs due to widely differing reproduction costs and outcomes (Table 2).
Another evolutionary trade-off is the relative investments orchid make in vegetative reproduction (clonal division) relative to seed production. In general, highly clonal plants had lower rates of flowering per plant due to their aggregation and displayed greater impacts of density on pollination than other orchids. However, they would be expected to compensate for this by having much longer lifespans. Populations of large groups of clonal orchids tended to be stable over the period of observation (5 years), with some losses of groups due perhaps to predation of tubers, or disease impacts. Populations of these species are characterized by gradual expansion of colonies and very occasional loss of a colony or establishment of a new one. Presumably they invest resources in flowering and fruit set primarily to allow very occasional short or long-range dispersal events. However, this strategy is less likely to succeed now than in the past, due to extensive land clearing which has greatly reduced the chance of seed landing in a suitable site (most Swan Coastal Plain habitats are 50–90% cleared [71]).
A poor relationship between flowering effort and pollination outcomes was observed in this study, since visually deceptive orchids with the largest displays also tended to grow at high densities which made these displays less effective. Production of many flowers may help offset flower herbivory [63] and often results in longer flowering periods which should increase the probability of coinciding with peaks in pollinator activity. Another likely explanation for large floral displays is that visually deceptive orchids are caught in arms races with other species of co-occurring native plants they mimic. At the study site, the main species that orchids are likely to mimic were members of the pea and iris families (Fabaceae and Iridaceae), with very large floral displays [17,21,22,23,70]. High variability in the colors and shape of flowers occurred in visually deceptive species, especially D. magnifica and C. latifolia, presumably to better match the shape and color of multiple mimicked species, and reduce pollinator avoidance by making it harder to recognize orchid flowers [32,72].
Orchids are expected to cope with low pollination rates better than most other plant families because they produce numerous seeds [1,13,69,73] that are readily dispersed by wind [2,53]. Low rates of pollination are also expected to result in more outcrossing and increased rates of speciation [7,10,13,74,75]. All the orchids in this study belong to the Diurideae, with the exception of Pterostylis pp. (Cranichideae) and Disa bracteata (Orchideae). Phylogenetic studies have shown that visual deception is the ancestral condition of orchids in the Diurideae with 4–7 independent origins of sexual deception and several independent origins of nectar production [8]. Pollination by visual deception occurs in basal Caladenia species and the closely related genera Elythranthera and Pheladenia, as well as Thelymitra and Diuris which diverged from this group much earlier [8,75,76]. In Caladenia and Pterostylis sexual deception by pheromones to attract specific insects has independently led to a situation where high potential pollinator diversity allows rapid orchid speciation [77,78]. It seems likely that switching from visual deceptive to sexually deception would initially result in increased rates of pollination and allow smaller floral displays to be effective, but in the longer-term may also lead to risks associated with reliance on a single, or very few pollen vectors.
5. Conclusions
The use pollination metrics that incorporate plant density, such as SGA, provide powerful new tools for understanding insect abundance and other factors limiting orchid pollination. They should also allow for orchid dispersal, recruitment and demographics to be investigated more efficiently, as well as genetic patterns in orchid populations. They can also be used to set targets to trigger more intensive management of rare species [56]. Orchid pollination integrates multiple ecological processes including (i) the impact of climate and weather on orchid growth and flowering, (ii) the effect of climate and weather on insects, (iii) the reproductive potential of specific insects attracted by pheromones, (iv) the relative performance of orchids with generalist or specific pollination and (v) the relative success of disturbance tolerant vs. intolerant orchids. The first two address climate impacts while last three are surrogates of habitat condition and complexity. The insects observed here have key roles as pollinators (flies, bees, moths and beetles), recyclers (fungus gnats and ants) and biological control agents (thynnine wasps). Thus, orchid pollination data also represents the activity of key insect groups linked to ecosystem complexity and sustainability and can also help identify insect decline. These data are especially important in highly fragmented urban landscapes because many insects have specific food sources, are intolerant of habitat degradation and have a limited dispersibility, so are susceptible to local extinction [79,80]. It is also important to study common orchids to determine important factors that limit population viability for all orchids [81].
Comparison of closely and distantly related orchid species revealed many ecological differences. Closely related orchid species differed in floral displays, flowering times and pollination strategies, especially within the large genera Thelymitra and Caladenia. This study provided evidence for evolutionary trade-offs concerning pollination mechanisms and other ecological factors linked to pollination success rates. The first trade-off involves the relative investment in flowering effort and seed production, resulting in a continuum of inflorescence sizes and seed pod volumes from many small flowers and seedpods, through to very large displays that result in a few large seedpods. Competition between visually deceptive orchids and mimicked plants may have caused a visual display arms race, where inflorescences become larger and more colorful, but pollination rates remained low. The flowering time continuum represents a second trade-off between flowering earlier in early spring when water is more available, but insects may be scarce, or flowering later when higher temperatures promoted insect activity, but drought stress was severe. In this study, the majority of orchids flowered in early spring, but often had low rates of pollination. Plants that flowered later in spring had adequate pollination rates and were remarkably resilient to drought conditions during seed formation. Plants that flower in autumn have faced severe drought stress in recent years. A third reproductive trade-off involves their relative investments in clonal and/or sexual reproduction. The former should be linked to long-term population stability and the latter with acquisition of new territory. Another related attribute concerns the degree of aggregation of plants in groups, due to lower rates of pollination in large groups. A fourth trade-off involves disturbance tolerance which is also linked to shade tolerance. Some disturbance tolerant orchids had higher rates of pollination in more open areas, but were also subject to increased risks from grazing and weeds. The degree to which fire promotes flowering is a related attribute (which varies from an absolute requirement through to a deterrent), since flowers should be much more detectable to pollinators after fire, but fires are normally infrequent. The fifth trade-off involves the relative specificity of insect pollinators, since orchids that require particular insects should be more vulnerable to habitat decline or fragmentation than those with a suite of pollinators. In the current study, visually deceptive orchids attracted different categories of pollinators in different situations (e.g., bees, beetles, or moths), while sexually deceptive orchids pollinated by one thynnine wasp had very low fruit set, especially at high plant density. Pollination needs to be measured relative to plant density to investigate any of these trade-offs.
The relative merits of competing reproductive strategies need to be evaluated over longer timeframes, since there was no clear winner when pollination strategies are compared (Table 1). Self-pollination was most efficient overall, but may eventually lead to reduced fitness due to genetic bottlenecks [13,82]. Extreme specialization due to sexual deception via insect pheromone production with high pollinator specificity works well in some cases but not in others. This may result because orchids with specific pollinators are more susceptible to factors that impact on the phenology and abundance of a particular insect (Table 2). However, there also were substantial impacts of climate on pollination of visually attractive orchids that attract a suite of pollinators and even the performance of self-pollinating orchids was limited by severe drought. Climatic impacts on Australian orchid seed production are not new (e.g., Rogers 1931 [48]), but rainfall deficits and temperature extremes have become mere severe and frequent in recent decades [41].
The South West Australian Floristic Region is a globally important terrestrial diversity hotspot and centre for evolution of plant nutrition traits [30]. This region is equally important as a key centre of evolution for pollination strategies with an exceptionally high diversity of plants with deceptive pollination, highly specific insect interactions, animal pollination, secondary pollen presentation, or other extreme floral modifications [29]). Perhaps due to arms races to attract limited pollinators, this region has evolved one of the world’s most spectacular terrestrial orchid floras. These orchids have very complex and fascinating interactions with insects that will require ongoing research and monitoring to reveal how relationships work, why they evolved and how they will continue to respond to climate change. Impacts of climate extremes and fire on orchids were identified during this study, but require further investigation.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/11/8/123/s1, Figure S1: Monthly average daily maximum temperatures and total rainfall measured in the vicinity of the study site, Figure S2: Relationships between plant density and capsules per plant for 8 orchid species, Figure S3: Average capsule volumes or total volumes relative to the total number of seedpods, Table S1: Summary of sampling locations and years for orchid pollination data, Video S1: Pollination of Leporella fimbria, Video S2 Caladenia discoidea pollination.
Funding
This project was not funded and carried out in my own time.
Acknowledgments
Special thanks to Karen Clarke for her endless patience, joining me on many walks through the study site and comments on the manuscript. I am also very grateful for help locating orchids by Tim Hodgkins, Geoff and Robyn Foley, Jay and Bob Steer, Ken Mulavin, Julie van Oosten, Marina Karyagina, Beth and Ron Kinsey, the Friends of Warwick Bushland and City of Joondalup staff.
Conflicts of Interest
The author declares no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Summary of measured attributes of species in this study for flowering chronology and effort (see methods), along with observed pollinators and key references.
Table A1.
Summary of measured attributes of species in this study for flowering chronology and effort (see methods), along with observed pollinators and key references.
| Full Name | Flowering (fls.) Start (date) | Fls. Peak (date) | Fls. End (date) | Seed Ripe (date) | Spike Height (mm) | Leaf no. | Leaf Length (mm) | Leaf Width (mm) | Leaf Area mm3 | Plants With Fls. | Mean Fls. per Spike | Fl. no. Range | Fl. Area (mm3) | Fl. Area per Spike | Fl. Area per Plant (mm3) | Flower Colors | Pollination Syndrome | Observed and Reported Pollinators | References for Pollinators |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leporella fimbriata (Lindl.) A.S.George | 16–4 | 20–5 | 4–6 | 10–7 | 180 | 1–2 | 16–20 | 4–6 | 300 | 0.09 | 1.6 | 1–3 | 70 | 112 | 9.7 | Green and purple | Sexual deception (SD) | Male ants (Myrmecia spp.) | [55,86] |
| Eriochilus dilatatus Lindl. subsp. dilatatus | 1–5 | 30–5 | 1–7 | 1–8 | 320 | 1 | 10–70 | 4–8 | 400 | 0.77 | 2.4 | 1–8 | 160 | 384 | 295 | White with pink and green markings | Nectar | Native bees, honeybees | [32,44,55] |
| Pterostylis sanguinea D.L.Jones and M.A.Clem. | 7–6 | 11–7 | 16–8 | 9–9 | 210 | 7–13 | 4–14 | 3–6 | 1800 | 0.95 | 3.9 | 1–15 | 50 | 195 | 185.25 | Translucent green, red or brown | SD | Fungus gnats (Mycomya sp.) | [55,88] |
| Pheladenia deformis (R.Br.) D.L.Jones and M.A.Clem. | 2–7 | 17–7 | 7–8 | 20–9 | 130 | 1 | 45–70 | 1–3 | 90 | 0.83 | 1 | 1–2 | 300 | 300 | 249 | Bright blue | Visual deception (VD) | Native bees | [48] |
| Pterostylis ectypha D.L.Jones and C.J.French | 7–8 | 23–8 | 19–9 | 5–10 | 80 | 3–7 | 11–19 | 4–7 | 210 | 0.65 | 1 | 1 | 90 | 90 | 58.5 | Green | SD | Fungus gnats | [55,89] |
| Diuris magnifica D.L.Jones | 15–8 | 15–9 | 4–10 | 24–10 | 480 | 2–5 | 60–280 | 5–10 | 2560 | 0.95 | 3.6 | 2–8 | 360 | 1296 | 1231.2 | Yellow, orange, purple | VD | Moth, bees, flies, beetles | [17,22] |
| Pyrorchis nigricans (R.Br.) D.L.Jones and M.A.Clem. | 19–8 | 16–9 | 12–10 | 30–10 | 180 | 1–2 | 40–100 | 30–80 | 2350 | 0.3 | 3.7 | 2–7 | 300 | 1110 | 333 | Dark red | Some nectar | Bees, flies, etc. | [55] |
| Elythranthera brunonis (Endl.) A.S.George | 20–8 | 15–9 | 16–10 | 24–10 | 330 | 1 | 60–145 | 5–7 | 440 | 0.71 | 1.6 | 1–3 | 290 | 464 | 329.44 | Glossy dark purple | VD | Bees, flies, etc. | [32,64] |
| Caladenia flava R.Br. subsp. flava | 26–8 | 18–9 | 10–10 | 29–10 | 150 | 1 | 50–100 | 5–12 | 450 | 0.24 | 1.8 | 1–4 | 530 | 954 | 228.96 | Bright yellow | VD | Native bees, Beetles | [32,44] |
| Caladenia discoidea Lindl. | 23–8 | 11–9 | 4–10 | 28–10 | 230 | 1 | 75–150 | 8–5 | 650 | 0.62 | 1.8 | 1–4 | 490 | 882 | 546.84 | Orange-green | SD | Thynnine wasp | [45] |
| Caladenia arenicola Hopper and A.P.Br. | 28–8 | 22–9 | 5–10 | 6–11 | 440 | 1 | 100–225 | 8–12 | 1050 | 0.60 | 1.8 | 1–4 | 805 | 1449 | 869.4 | Red and green | SD | Thynnid wasp | [42,77] |
| Caladenia latifolia R.Br. | 4–9 | 20–9 | 24–10 | 7–11 | 300 | 1 | 65–210 | 8–21 | 1300 | 0.64 | 2.5 | 1–5 | 260 | 650 | 416 | Pink | VD | Beetles, bees | See C. flava |
| Caladenia longicauda subsp. calcigena Hopper and A.P.Br. | 7–9 | 20–9 | 4–10 | 1–11 | 320 | 1 | 120–200 | 7–12 | 800 | 0.25 | 1.8 | 1–4 | 700 | 1260 | 315 | White | VD | Flies, beetles, bees | [32,42] |
| Thelymitra vulgaris Jeanes | 14–9 | 23–9 | 4–10 | 1–11 | 315 | 1 | 65–155 | 1–4 | 210 | 0.9 | 2.6 | 1–6 | 120 | 312 | 280.8 | Blue | Self-pollination | none observed | [90] |
| Thelymitra macrophylla Lindl. | 17–9 | 2–10 | 20–10 | 4–11 | 700 | 1 | 140–250 | 7–25 | 1900 | 0.74 | 6.6 | 2–15 | 500 | 3300 | 2442 | Blue | VD | Flies, bees, etc. | [70] |
| Thelymitra benthamiana Rchb.f. | 4–10 | 14–10 | 26–10 | 16–11 | 330 | 1 | 40–210 | 10–40 | 1200 | 0.50 | 2.1 | 1–8 | 300 | 630 | 315 | Yellow and brown | Self-pollination | none observed | [91] |
| Thelymitra graminea Lindl. | 7–10 | 26–10 | 4–11 | 20–11 | 360 | 1 | 55–170 | 8–3 | 400 | 0.97 | 3.8 | 2–8 | 320 | 1216 | 1179.52 | Blue | VD | Hoverflies, etc. | This study |
| Disa bracteata Sw. | 20–10 | 8–11 | 21–11 | 30–11 | 240 | 5–16 | 15–150 | 2–10 | 1670 | 0.95 | 32 | 10–70 | 17 | 544 | 516.8 | Greenish | Self-pollination (some nectar) | none observed | [46] |
| Microtis media R.Br. subsp. media | 25–10 | 7–11 | 18–11 | 25–11 | 250 | 1 | 140–270 | 1.8–2.2 | 950 | 1 | 42 | 10–100 | 5 | 210 | 210 | Green | Self-pollination (some nectar) | ants, micro wasps | [55,92] |
| Thelymitra fuscolutea R.Br. | 6–11 | 21–11 | 14–12 | 7–1 | 380 | 1 | 60–140 | 7–15 | 780 | 0.72 | 6.2 | 2–10 | 490 | 3038 | 2187.36 | Orange and brown | VD | Hover flies, etc. | This study |
Table A2.
Capsule numbers, sizes and volumes for each species. Totals per plant include flowering and non–flowering plants. Shaded columns show reproductive vital statistics.
Table A2.
Capsule numbers, sizes and volumes for each species. Totals per plant include flowering and non–flowering plants. Shaded columns show reproductive vital statistics.
| Orchid | Location | Year | Spikes | Flowers | Capsules/Flower | Capsules/Spike | Capsules/Flower SGA | Capsules/Spike SGA | Capsules/Flower LGA | Capsules Measured | Capsules/Spike Range | Average Capsule Length (mm) | Capsule Length Range (mm) | Average Capsule Width (mm) | Capsule Width Range (mm) | Average Capsule Volume (mm3) | Capsule Volume Range | Total Capsule Volume/Spike (mm3) | Total Capsule Volume/Plant (mm3) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leporella fimbriata | WCA | 2018 | 175 | 278 | 0.17 | 0.27 | 0.60 | 1.2 | 0.2 | 21 | 0–3 | 8.7 | 6–11 | 3 | 2–5 | 45 | 30–90 | 72 | 6 |
| Eriochilus dilatatus | WCA, etc. | 2014–16 | 61 | 149 | 0.16 | 0.59 | ND | ND | ND | 3 | 0–4 | 13 | 11–15 | 4 | 3–4 | 100 | 70–120 | 59 | 45 |
| Pterostylis sanguinea | WCA | 2018 | 173 | 680 | 0.23 | 0.95 | 0.58 | 2.5 | 0.1 | 61 | 0–6 | 13 | 9–16 | 7 | 5–10 | 300 | 100–690 | 630 | 599 |
| Pheladenia deformis | WCA | 2018 | 39 | 39 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 18 | 15–20 | 4.5 | 4–5 | 190 | NA | 0 | 0 |
| Pterostylis ectypha | WCA | 2018 | 265 | 265 | 0.39 | 0.39 | 0.81 | 0.81 | 0.2 | 26 | 0–1 | 11 | 7–20 | 3.5 | 3–5 | 80 | 35–290 | 14 | 9 |
| Pterostylis recurva* | WCA, etc. | 2016–18 | 4 | 8 | 0.5 | 1 | ND | ND | ND | 4 | 0–2 | 16 | 15–19 | 9 | 7.5–10 | 720 | 500–1000 | 936 | 936 |
| Diuris magnifica | WCA | 2018 | 465 | 1690 | 0.058 | 0.21 | 0.18 | 0.76 | 0.00 | 46 | 0–3 | 13 | 12–17 | 7 | 5–8 | 320 | 230–640 | 368 | 350 |
| Pyrorchis nigricans | Marang-aroo | 2018 | 18 | 67 | 0.31 | 1.17 | ND | ND | ND | 9 | 2–3 | 14.5 | 10–19 | 8.2 | 6.8–11.8 | 570 | 220–1400 | 1283 | 385 |
| Elythranthera brunonis | WCA | 2016–18 | 35 | 55 | 0.24 | 0.37 | ND | ND | ND | 6 | 0–2 | 12.2 | 9.1–14.1 | 5.3 | 4.8–6.4 | 180 | 112–246 | 216 | 153 |
| Caladenia flava | WCA | 2018 | 34 | 61 | 0.48 | 0.85 | 0.76 | 1.4 | 0.05 | 10 | 0–2 | 15 | 13–18 | 6 | 4–6.5 | 280 | 130–400 | 392 | 94 |
| Caladenia arenicola | WCA | 2008–18 | 101 | 177 | 0.13 | 0.23 | 0.49 | 0.78 | 0.00 | 12 | 0–2 | 14.9 | 10–20 | 6.7 | 5–9 | 360 | 170–650 | 432 | 259 |
| Caladenia discoidea | WCA | 2015–18 | 52 | 95 | 0.16 | 0.29 | 0.44 | 0.44 | 0.15 | 7 | 0–2 | 15 | 10–21 | 5.8 | 3–9 | 350 | 40–580 | 490 | 304 |
| Caladenia latifolia | WCA | 2018 | 105 | 217 | 0.47 | 1.16 | 0.75 | 2.3 | 0.3 | 16 | 0–2 | 16 | 11–20 | 6 | 4–7.5 | 320 | 80–510 | 512 | 328 |
| Caladenia longicauda | Sorento | 2008–18 | 4 | 7 | 0.14 | 0.25 | ND | ND | ND | 2 | 0–1 | 20 | NA | 7 | NA | 520 | NA | 151 | 38 |
| Leptoceras menziesii* | WCA | 2008 | 6 | 10 | 0.3 | 0.5 | ND | ND | ND | 3 | 0–2 | 9.5 | 9–10 | 4 | 3–5 | 80 | 60–120 | 40 | 12 |
| Thelymitra vulgaris | WCA | 2018 | 29 | 76 | 0.95 | 2.48 | ND | ND | ND | 65 | 0–5 | 10.5 | 8–19 | 4.4 | 3–5.5 | 133 | 30–290 | 346 | 311 |
| Thelymitra macrophylla | WCA | 2018 | 698 | 4574 | 0.048 | 0.24 | 0.054 | 1.2 | 0.04 | 171 | 0–8 | 13 | 21–7 | 5.5 | 3.5–9 | 215 | 40–820 | 774 | 573 |
| Thelymitra benthamiana | WCA | 2018 | 38 | 79 | 0.90 | 1.87 | 0.99 | 2.6 | 0.90 | 36 | 0–5 | 15 | 7–22 | 6 | 4–8 | 300 | 90–660 | 600 | 300 |
| Thelymitra graminea | WCA | 2018 | 64 | 240 | 0.17 | 0.63 | 0.31 | 0.6 | 0.13 | 29 | 0–5 | 10.5 | 17–6 | 4.7 | 7.5–3.5 | 140 | 30–360 | 448 | 435 |
| Disa bracteata | WCA | 2018 | 26 | 821 | 0.59 | 19 | ND | ND | ND | 58 | 3–46 | 11 | 18–7 | 3.4 | 2–4.5 | 72 | 18–138 | 1368 | 1300 |
| Microtis media | WCA | 2018 | 38 | 1583 | 0.93 | 39 | 0.97 | 42 | 0.92 | 96 | 30–67 | 4.1 | 5.7–2.8 | 2.4 | 3.7–1.5 | 14 | 5–40 | 406 | 406 |
| Thelymitra fuscolutea | WCA | 2018 | 102 | 632 | 0.070 | 0.43 | 0.078 | 0.54 | 0.06 | 78 | 0–4 | 11 | 8–14 | 5.2 | 4–7 | 160 | 70–310 | 67 | 48 |
Notes: * Locally uncommon species not included in Table A1. SGA = small group average and LGA = large group average from a graph of pollination vs. plant density. ND = not determined (> 5 groups, or not grouped.)
References
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef]
- Brundrett, M.C. Scientific approaches to Australian temperate terrestrial orchid conservation. Aust. J. Bot. 2007, 55, 293–307. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Phillips, R.D.; Peakall, R.; Hutchinson, M.F.; Linde, C.C.; Xu, T.; Dixon, K.W.; Hopper, S.D. Specialized ecological interactions and plant species rarity: the role of pollinators and mycorrhizal fungi across multiple spatial scales. Biol. Conserv. 2014, 169, 285–295. [Google Scholar] [CrossRef]
- Darwin, C. The Various Contrivances by which British and Foreign Orchids are Fertilised By Insects, 2nd ed.; John Murray: London, UK, 1877; pp. 1–300. [Google Scholar]
- Johnson, S.D.; Nilsson, L.A. Pollen carryover, geitonogamy, and the evolution of deceptive pollination systems in orchids. Ecology 1999, 80, 2607–2619. [Google Scholar] [CrossRef]
- Weston, P.H.; Perkins, A.J.; Indsto, J.O.; Clements, M.A. Phylogeny of Orchidaceae tribe Diurideae and its implications for the evolution of pollination systems. In Darwin’s Orchids: Then and Now; University of Chicago Press: Chicago, IL, USA, 2014; pp. 91–154. [Google Scholar]
- Dafni, A. Mimicry and deception in pollination. Annu. Rev. Ecol. Syst. 1984, 15, 259–278. [Google Scholar] [CrossRef]
- Cozzolino, S.; Widmer, A. Orchid diversity: an evolutionary consequence of deception? Trends Ecol. Evol. 2005, 20, 487–494. [Google Scholar] [CrossRef]
- Vereecken, N.J.; Dafni, A.; Cozzolino, S. Pollination syndromes in Mediterranean orchids—implications for speciation, taxonomy and conservation. Bot. Rev. 2010, 76, 220–240. [Google Scholar] [CrossRef]
- Schiestl, F.P. On the success of a swindle: Pollination by deception in orchids. Naturwissenschaften 2005, 92, 255–264. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification. Biol. J. Linn. Soc. 2005, 84, 1–54. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Dafni, A.; Bernhhardt, P. Pollination of terrestrial orchids of Southern Australia and the Mediterranean region. Systematics, ecological and evolutionary implications. Evol. Biol. 1990, 24, 193–252. [Google Scholar]
- Herberstein, M.E.; Baldwin, H.J.; Gaskett, A.C. Deception down under: is Australia a hot spot for deception? Behav. Ecol. 2013, 25, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Beardsell, D.; Clements, M.; Hutchinson, J.; Williams, E. Pollination of Diuris maculata R Br (Orchidaceae) by floral mimicry of the native legumes Daviesia spp. and Pultenaea scabra R Br. Aust. J. Bot. 1986, 34, 165–173. [Google Scholar] [CrossRef]
- Elliott, C.P.; Ladd, P.G. Pollen limitation of fruit set in Western Australian terrestrial orchids. J. R. Soc. West. Aust. 2002, 85, 165–168. [Google Scholar]
- Edens-Meier, R.; Bernhardt, P. The Sun Orchids (Thelymitra) Then and Now: Large Flowers versus Small Flowers and Their Evolutionary Implications. Darwins Orchids Then Now 2014, 173–200. [Google Scholar]
- Gaskett, A.C.; Endler, J.A.; Phillips, R.D. Convergent evolution of sexual deception via chromatic and achromatic contrast rather than colour mimicry. Evol. Ecol. 2017, 31, 205–227. [Google Scholar] [CrossRef]
- Cropper, S.C.; Calder, D.M. The floral biology of Thelymitra epipactoides (Orchidaceae), and the implications of pollination by deceit on the survival of this rare orchid. Plant Syst. Evol. 1990, 170, 11–27. [Google Scholar] [CrossRef]
- Indsto, J.O.; Weston, P.H.; Clements, M.A.; Dyer, A.G.; Batley, M.; Whelan, R.J. Pollination of Diuris maculata (Orchidaceae) by male Trichocolletes venustus bees. Aust. J. Bot. 2006, 54, 669–679. [Google Scholar] [CrossRef]
- Scaccabarozzi, D.; Cozzolino, S.; Guzzetti, L.; Galimberti, A.; Milne, L.; Dixon, K.W.; Phillips, R.D. Masquerading as pea plants: behavioural and morphological evidence for mimicry of multiple models in an Australian orchid. Ann. Bot. 2018, 122, 1061–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagawa, K.; Takimoto, G. Inaccurate color discrimination by pollinators promotes evolution of discrete color polymorphism in food-deceptive flowers. Am. Nat. 2016, 187, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.B.; Lawson, S.D. Pollination in Australian orchids: a critical–assessment of the literature 1882–1992. Aust. J. Bot. 1993, 41, 553–575. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Hopper, S.D.; Gioia, P. The southwest Australian floristic region: evolution and conservation of a global hot spot of biodiversity. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 623–650. [Google Scholar] [CrossRef]
- Lambers, H.; Brundrett, M.C.; Raven, J.A.; Hopper, S.D. Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant Soil 2011, 348, 7. [Google Scholar] [CrossRef]
- Groom, P.K.; Lamont, B. Plant Life of Southwestern Australia: Adaptations for Survival; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2015; pp. 1–258. [Google Scholar]
- Brundrett, M.C. Distribution and evolution of mycorrhizal types and other specialised roots in Australia. In Biogeography of Mycorrhizal Symbiosis; Springer: Cham, Switzerland, 2017; Volume 230, pp. 361–394. [Google Scholar]
- Phillips, R.D.; Barrett, M.D.; Dixon, K.W.; Hopper, S.D. Do mycorrhizal symbioses cause rarity in orchids? J. Ecol. 2011, 99, 858–869. [Google Scholar] [CrossRef]
- Brundrett, M. Identification and Ecology of Southwest Australian Orchids; Western Australian Naturalists’ Club Inc.: Perth, WA, Australia, 2014; p. 424. [Google Scholar]
- Banksia Woodlands of the Swan Coastal Plain: a nationally-protected ecological community. Available online: https://www.environment.gov.au/biodiversity/threatened/publications/banksia-woodlands-swan-coastal-plain-guide (accessed on 16 September 2016).
- Ramalho, C.E.; Laliberte, E.; Poot, P.; Hobbs, R.J. Complex effects of fragmentation on remnant woodland plant communities of a rapidly urbanizing biodiversity hotspot. Ecology 2014, 95, 2466–2478. [Google Scholar] [CrossRef] [Green Version]
- Government of Western Australia Directory of Bush Forever Sites. In Bush Forever: Keeping the Bush in the City; Department of Environmental Protection: Perth, Western Australia, Australia, 2000; Volume 2, pp. 1–530.
- Friends of Warwick Bushland Warwick Bushland. Available online: Friendsofwarwickbushland.com (accessed on 24 June 2019).
- Bonnardeaux, Y.; Brundrett, M.; Batty, A.; Dixon, K.; Koch, J.; Sivasithamparam, K. Diversity of mycorrhizal fungi of terrestrial orchids: compatibility webs, brief encounters, lasting relationships and alien invasions. Mycol. Res. 2007, 111, 51–61. [Google Scholar] [CrossRef]
- Scade, A.; Brundrett, M.; Batty, A.; Dixon, K.; Sivasithamparam, K. Survival of transplanted terrestrial orchid seedlings in urban bushland habitats with high or low weed cover. Aust. J. Bot. 2006, 54, 383–389. [Google Scholar] [CrossRef]
- Tao, Z.-B.; Ren, Z.-X.; Bernhardt, P.; Wang, W.-J.; Liang, H.; Li, H.-D.; Wang, H. Nocturnal hawkmoth and noctuid moth pollination of Habenaria limprichtii (Orchidaceae) in sub-alpine meadows of the Yulong Snow Mountain (Yunnan, China). Bot. J. Linn. Soc. 2018, 187, 483–498. [Google Scholar] [CrossRef]
- Petit, S.; Jusaitis, M.; Bickerton, D. Effect of pollen load, self-pollination and plant size on seeds and germination in the endangered pink-lipped spider orchid, Caladenia behrii. Aust. J. Bot. 2009, 57, 307–314. [Google Scholar] [CrossRef]
- Bureau of Meteorology Climate data online. Available online: www.bom.gov.au (accessed on 18 January 2019).
- Stoutamire, W.P. Wasp-pollinated species of Caladenia (Orchidaceae) in south-western Australia. Aust. J. Bot. 1983, 31, 383–394. [Google Scholar] [CrossRef]
- Phillips, R.; Peakall, R.; Retter, B.; Montgomery, K.; Menz, M.; Davis, B.; Hayes, C.; Brown, G.; Swarts, N.; Dixon, K. Pollinator rarity as a threat to a plant with a specialized pollination system. Bot. J. Linn. Soc. 2015, 179, 511–525. [Google Scholar] [CrossRef] [Green Version]
- Erickson, R. Orchids of the west; Paterson, Brokensha, Pty. Ltd.: Perth, Western Australia, Australia, 1951; pp. 1–109. [Google Scholar]
- Pollinator Photos by Mark Brundrett & Orchid Photographers Identifications by Graham Brown. Available online: https://www.researchgate.net/publication/269280032_Wasp_Pollinator_Photos_By_Mark_Brundrett_Orchid_Photographers_Identifications_by_Graham_Brown (accessed on 9 December 2014).
- Johnson, S.D.; Linder, H.P.; Steiner, K.E. Phylogeny and radiation of pollination systems in Disa (Orchidaceae). Am. J. Bot. 1998, 85, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Light, M.H.S.; MacConaill, M. Climatic influences on flowering and fruiting of Cypripedium parviflorum var. pubescens. In Trends and Fluctuations and Underlying Mechanisms in Terrestrial Orchid Populations; Backhuys Publishers: Leiden, Netherland, 2002; pp. 85–97. [Google Scholar]
- Rogers, R.S. Pollination of Caladenia deformis, R. Br. Trans. Proc. R. Soc. S. Aust. 1931, 55, 143–146. [Google Scholar]
- Brown, J.; York, A. Fire, food and sexual deception in the neighbourhood of some Australian orchids. Austral Ecol. 2017, 42, 468–478. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PloS One 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Ridsdill Smith, T. The behaviour of Hemithynnus hyalinatus (Hymenoptera: Tiphiidae), with notes on some other Thynninae. Aust. J. Entomol. 1970, 9, 196–208. [Google Scholar]
- Menz, M.H.; Phillips, R.D.; Dixon, K.W.; Peakall, R.; Didham, R.K. Mate-searching behaviour of common and rare wasps and the implications for pollen movement of the sexually deceptive orchids they pollinate. PLoS One 2013, 8, e59111. [Google Scholar] [CrossRef]
- Jersáková, J.; Malinová, T. Spatial aspects of seed dispersal and seedling recruitment in orchids. New Phytol. 2007, 176, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Woodland Restoration Project Annual Report 6 January - December 2017. Available online: https://www.researchgate.net/publication/325965309_Banksia_Woodland_Restoration_Project_Annual_Report_6_January_-_December_2017 (accessed on 25 June 2018).
- Kuiter, R.H. Orchid Pollinators of Victoria, 3rd ed.; Aquatic Photographics: Seaford, Victoria, VIC, Australia, 2015; pp. 252–525. [Google Scholar]
- Brundrett, M.C. Using vital statistics and core-habitat maps to manage critically endangered orchids in the Western Australian wheatbelt. Aust. J. Bot. 2016, 64, 51–64. [Google Scholar] [CrossRef]
- Petit, S.; Dickson, C.R. Grass-tree (Xanthorrhoea semiplana, Liliaceae) facilitation of the endangered pink-lipped spider orchid (Caladenia syn. Arachnorchis behrii, Orchidaceae) varies in South Australia. Aust. J. Bot. 2005, 53, 455–464. [Google Scholar] [CrossRef]
- Ong, J.; Li, H.; Sivasithamparam, K.; Dixon, K.W.; Jones, M.; Wylie, S.J. Novel and divergent viruses associated with Australian orchid-fungus symbioses. Virus Res. 2018, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Brundrett, M. Recovery of terrestrial orchids in natural ecosystems after severe disturbance. In Mining in Ecologically Sensitive Landscapes; CSIRO PUBLISHING: Victoria, Australia, 2015; pp. 141–158. [Google Scholar]
- Brundrett, M.; Scade, A.; Batty, A.; Dixon, K.; Sivasithamparam, K. Development of in situ and ex situ seed baiting techniques to detect mycorrhizal fungi from terrestrial orchid habitats. Mycol. Res. 2003, 107, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J.H. Does nectar reward affect rarity and extinction probabilities of orchid species? An assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 121, 257–263. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Aide, T.M. Patterns of fruit production in a neotropical orchid: Pollinator vs. resource limitation. Am. J. Bot. 1989, 76, 67–73. [Google Scholar] [CrossRef]
- Vallius, E. Position-dependent reproductive success of flowers in Dactylorhiza maculata (Orchidaceae). Funct. Ecol. 2000, 14, 573–579. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Bateman, A.P.; Brown, A.; Hachandourian, M.; Hutchings, M.J.; Kell, S.; Koopowitz, H.; Lehnebach, C.; Whigham, D.F. Density induced rates of pollinaria removal and deposition in the purple enamel orchid Elythranthera brunonis (Endl.). AS George. 2007, 7, 229–239. [Google Scholar] [CrossRef]
- Peakall, R. Responses of male Zaspilothynnus trilobatus Turner wasps to females and the sexually deceptive orchid it pollinates. Funct. Ecol. 1990, 159–167. [Google Scholar] [CrossRef]
- Dafni, A.; Calder, D. Pollination by deceit and floral mimesis in Thelymitra antennifera (Orchidaceae). Plant Syst. Evol. 1987, 158, 11–22. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Honnay, O. Large population sizes mitigate negative effects of variable weather conditions on fruit set in two spring woodland orchids. Biol. Lett. 2009, 5, 495–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brys, R.; Jacquemyn, H.; Hermy, M. Pollination efficiency and reproductive patterns in relation to local plant density, population size, and floral display in the rewarding Listera ovata (Orchidaceae). Bot. J. Linn. Soc. 2008, 157, 713–721. [Google Scholar] [CrossRef]
- Perez-Hernandez, H.; Damon, A.; Valle-Mora, J.; Sanchez-Guillen, D. Orchid pollination: specialization in chance? Bot. J. Linn. Soc. 2011, 165, 251–266. [Google Scholar] [CrossRef]
- Edens-Meier, R.M.; Raguso, R.A.; Westhus, E.; Bernhardt, P. Floral fraudulence: Do blue Thelymitra species (Orchidaceae) mimic Orthrosanthus laxus (Iridaceae)? Telopea 2014, 17, 15–28. [Google Scholar] [CrossRef]
- Western Australian Local Government Association 2013 Native vegetation on the Swan Coastal Plain. Available online: http://pbp.walga.asn.au/Publications.html (accessed on 2 April 2019).
- Ackerman, J.D.; Cuevas, A.A.; Hof, D. Are deception-pollinated species more variable than those offering a reward? Plant Syst. Evol. 2011, 293, 91–99. [Google Scholar] [CrossRef]
- Sonkoly, J.E.; Vojtkó, A.; Tökölyi, J.; Török, P.; Sramkó, G.; Illyés, Z.; Molnár, V.A. Higher seed number compensates for lower fruit set in deceptive orchids. J. Ecol. 2016, 104, 343–351. [Google Scholar] [CrossRef]
- Calvo, R.N. Evolutionary demography of orchids: intensity and frequency of pollination and the cost of fruiting. Ecology 1993, 74, 1033–1042. [Google Scholar] [CrossRef]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.; Clements, M.A.; Arroyo, M.T.; Leebens-Mack, J. Orchid phylogenomics and multiple drivers of their extraordinary diversification. Proc R Soc B 2015, 282, 20151553. [Google Scholar] [CrossRef]
- Nauheimer, L.; Schley, R.J.; Clements, M.A.; Micheneau, C.; Nargar, K. Australasian orchid biogeography at continental scale: molecular phylogenetic insights from the sun orchids (Thelymitra, Orchidaceae). Mol. Phylogenet. Evol. 2018, 127, 304–319. [Google Scholar] [CrossRef]
- Phillips, R.D.; Faast, R.; Bower, C.C.; Brown, G.R.; Peakall, R. Implications of pollination by food and sexual deception for pollinator specificity, fruit set, population genetics and conservation of Caladenia (Orchidaceae). Aust. J. Bot. 2009, 57, 287–306. [Google Scholar] [CrossRef]
- Overview of Pterostylis Pollination (Orchidaceae) in Victoria. Available online: https://www.researchgate.net/publication/318282443_Overview_of_Pterostylis_Pollination_Orchidaceae_in_Victoria (accessed on 8 July 2017).
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Williams, M.R. Habitat resources, remnant vegetation condition and area determine distribution patterns and abundance of butterflies and day-flying moths in a fragmented urban landscape, south-west Western Australia. J. Insect Conserv. 2011, 15, 37–54. [Google Scholar] [CrossRef]
- Light, M.H.; MacConaill, M. The role of common orchids in appreciating the complexity of biodiversity conservation. Lankesteriana Int. J. Orchid. 2011, 11, 293–330. [Google Scholar] [CrossRef]
- Catling, P.M. Auto-pollination in the Orchidaceae. Timber Press 1990, 5, 121–158. [Google Scholar]
- Newman, B.J.; Ladd, P.; Brundrett, M.; Dixon, K.W. Effects of habitat fragmentation on plant reproductive success and population viability at the landscape and habitat scale. Biol. Conserv. 2013, 159, 16–23. [Google Scholar] [CrossRef]
- Orchid Rescue Project: Case Studies of Collaborative Orchid Conservation in Western Australia; University of Western Australia. Available online: https://www.researchgate.net/publication/259500490_Wheatbelt_Orchid_Rescue_Project_Case_Studies_of_Collaborative_Orchid_Conservation_in_Western_Australia (accessed on 2 January 2011).
- Sydes, M.A.; Calder, D.M. Comparative reproductive biology of two sun-orchids; the vulnerable Thelymitra circumsepta and the widespread T. ixioides (Orchidaceae). Aust. J. Bot. 1993, 41, 577–589. [Google Scholar] [CrossRef]
- Peakall, R. The unique pollination of Leporella fimbriata (Orchidaceae): pollination by pseudocopulating male ants (Myrmecia urens, Formicidae). Plant Syst. Evol. 1989, 167, 137–148. [Google Scholar] [CrossRef]
- Batty, A.L.; Dixon, K.W.; Brundrett, M.; Sivasithamparam, K. Constraints to symbiotic germination of terrestrial orchid seed in a mediterranean bushland. New Phytol. 2001, 152, 511–520. [Google Scholar] [CrossRef]
- Phillips, R.D.; Scaccabarozzi, D.; Retter, B.A.; Hayes, C.; Brown, G.R.; Dixon, K.W.; Peakall, R. Caught in the act: pollination of sexually deceptive trap-flowers by fungus gnats in Pterostylis (Orchidaceae). Ann. Bot. 2013, 113, 629–641. [Google Scholar] [CrossRef]
- Hoffman, N.; Brown, A. Orchids of South-West Australia; Noel Hoffman: Perth, Western Australia, Australia, 2011; pp. 1–512. [Google Scholar]
- Jeanes, J.A. A revision of the Thelymitra pauciflora R. Br. (Orchidaceae) complex in Australia. Muelleria 2004, 19, 19–79. [Google Scholar]
- Rogers, R. Mechanism of pollination in certain Australian Orchids. Trans. Proc. R. Soc. S. Aust. 1913, 37, 48–61. [Google Scholar]
- Bates, R. The genus Microtis R. Br. (Orchidaceae): A taxonomic revision with notes on biology. J. Adel. Bot. Gard. 1984, 7, 45–89. [Google Scholar]
Figure 1.
Photographs showing differences in the form and color of flowers of 20 orchid species with some associated insects; (A) Leporella fimbria with male bull ant (Myrmecia sp.); (B) Eriochilus dilatatus. (CDE) Pterostylis sanguinea. (D) red and green color forms of this species co-occur; (E) Fungus gnat with pollinia (Mycomya sp.); (FG) Pheladenia deformis; (H) Pterostylis ectypha; (I) Diuris magnifica; (J) Pyrorchis nigricans; (K) Elythranthera brunonis; (L) Caladenia flava; (M) Caladenia discoidea flower and male thynnine wasp with attached pollen (Phymatothynnus sp.); (NO) Caladenia arenicola; (O) Male thynnine wasps competing for a C. arenicola flower (Zaspilothynnus sp.); (P) Caladenia latifolia; (Q) Caladenia longicauda subsp. calcigena; (R) Thelymitra vulgaris; (S) Thelymitra macrophylla; (T) Thelymitra benthamiana; (U) Thelymitra graminea with hoverfly; (V) Disa bracteata; (W) Microtis media; (X) Thelymitra fuscolutea.
Figure 1.
Photographs showing differences in the form and color of flowers of 20 orchid species with some associated insects; (A) Leporella fimbria with male bull ant (Myrmecia sp.); (B) Eriochilus dilatatus. (CDE) Pterostylis sanguinea. (D) red and green color forms of this species co-occur; (E) Fungus gnat with pollinia (Mycomya sp.); (FG) Pheladenia deformis; (H) Pterostylis ectypha; (I) Diuris magnifica; (J) Pyrorchis nigricans; (K) Elythranthera brunonis; (L) Caladenia flava; (M) Caladenia discoidea flower and male thynnine wasp with attached pollen (Phymatothynnus sp.); (NO) Caladenia arenicola; (O) Male thynnine wasps competing for a C. arenicola flower (Zaspilothynnus sp.); (P) Caladenia latifolia; (Q) Caladenia longicauda subsp. calcigena; (R) Thelymitra vulgaris; (S) Thelymitra macrophylla; (T) Thelymitra benthamiana; (U) Thelymitra graminea with hoverfly; (V) Disa bracteata; (W) Microtis media; (X) Thelymitra fuscolutea.

Figure 2.
Observations of flowering and fruit set trends for Leporella fimbria; (A) Flowering and pollination outcomes per group of plants; (B) Numbers of leaves, flowers and capsules relative to group size.
Figure 2.
Observations of flowering and fruit set trends for Leporella fimbria; (A) Flowering and pollination outcomes per group of plants; (B) Numbers of leaves, flowers and capsules relative to group size.

Figure 3.
Population size trends for populations of orchid species showing annual differences in numbers of (A) Leporella fimbria, (B) Thelymitra fuscolutea and (C) Thelymitra macrophylla.
Figure 3.
Population size trends for populations of orchid species showing annual differences in numbers of (A) Leporella fimbria, (B) Thelymitra fuscolutea and (C) Thelymitra macrophylla.

Figure 4.
Climatic trends for the main orchid species in this study; (A) The timing of flowering and seed production; (B) Total rainfall during periods of flowering and seed production; (C) Temperature trends during flowering periods.
Figure 4.
Climatic trends for the main orchid species in this study; (A) The timing of flowering and seed production; (B) Total rainfall during periods of flowering and seed production; (C) Temperature trends during flowering periods.

Figure 5.
Observations of potential pollinators, habitats, threats and seed production from this study; (A) Diuris magnifica with forester moth (Pollanisus sp., Zygaenidae—arrow); (B) Caladenia flava with nectar scarab beetles (Phyllotocus sp.—arrows); (C) Caladenia arenicola and male thynnine wasp with attached female (arrow, Zaspilothynnus sp.); (D) Thelymitra fuscolutea with hoverfly (Melangyna sp., Syrphidae—arrow); (E) The largest colonies of Diuris magnifica occurred in semi-disturbed areas; (F) Thelymitra fuscolutea leaves dried out before flowering; (F Inset) Rust pustules on leaves; (G) Thelymitra fuscolutea with web of an insect that consumed seedpods (arrow); (H) Leporella fimbria leaves damaged by a fire (arrows); (I) Caladenia arenicola flower showing suspected virus symptoms; (J) Thelymitra macrophylla with multiple seedpods showing size gradation; (K) Thelymitra fuscolutea seedpod maturation while stem is dying (arrow); L. Caladenia latifolia seedpod grazing by an insect (arrow); (M) Thelymitra vulgaris upper seedpods (arrow) are smaller than lower seedpods; (N) Disa bracteata seed abortion in upper flowers (arrow) due to drought; (O) Well-watered Disa bracteata with seed formation from all flowers (arrow).
Figure 5.
Observations of potential pollinators, habitats, threats and seed production from this study; (A) Diuris magnifica with forester moth (Pollanisus sp., Zygaenidae—arrow); (B) Caladenia flava with nectar scarab beetles (Phyllotocus sp.—arrows); (C) Caladenia arenicola and male thynnine wasp with attached female (arrow, Zaspilothynnus sp.); (D) Thelymitra fuscolutea with hoverfly (Melangyna sp., Syrphidae—arrow); (E) The largest colonies of Diuris magnifica occurred in semi-disturbed areas; (F) Thelymitra fuscolutea leaves dried out before flowering; (F Inset) Rust pustules on leaves; (G) Thelymitra fuscolutea with web of an insect that consumed seedpods (arrow); (H) Leporella fimbria leaves damaged by a fire (arrows); (I) Caladenia arenicola flower showing suspected virus symptoms; (J) Thelymitra macrophylla with multiple seedpods showing size gradation; (K) Thelymitra fuscolutea seedpod maturation while stem is dying (arrow); L. Caladenia latifolia seedpod grazing by an insect (arrow); (M) Thelymitra vulgaris upper seedpods (arrow) are smaller than lower seedpods; (N) Disa bracteata seed abortion in upper flowers (arrow) due to drought; (O) Well-watered Disa bracteata with seed formation from all flowers (arrow).

Figure 6.
Relationships between plant density (plants within 5 m) and fruit set per flower for 8 orchid species with regressions to show the effect of plant density. Data are for a single year and location unless separate symbols are used.
Figure 6.
Relationships between plant density (plants within 5 m) and fruit set per flower for 8 orchid species with regressions to show the effect of plant density. Data are for a single year and location unless separate symbols are used.

Figure 7.
Relationships between plant density and flower production, or flowering plants relative to total plants. Figures A–E show number of flowers per spike relative to numbers of flowering plants within a 5 m radius for Leporella fimbria (A), Pterostylis sanguinea (B), Diuris magnifica (C), Caladenia latifolia (D) and Thelymitra macrophylla (E). Figures F,G,H show proportion of flowering plants relative to or total plants in the same areas for T. fuscolutea (F), L. fimbria (G) and Thelymitra macrophylla (H).
Figure 7.
Relationships between plant density and flower production, or flowering plants relative to total plants. Figures A–E show number of flowers per spike relative to numbers of flowering plants within a 5 m radius for Leporella fimbria (A), Pterostylis sanguinea (B), Diuris magnifica (C), Caladenia latifolia (D) and Thelymitra macrophylla (E). Figures F,G,H show proportion of flowering plants relative to or total plants in the same areas for T. fuscolutea (F), L. fimbria (G) and Thelymitra macrophylla (H).

Figure 8.
Comparisons of capsule sizes relative to formation sequence (A) or between upper, middle and lower flowers from self-pollinating orchids with numerous flowers (B).
Figure 8.
Comparisons of capsule sizes relative to formation sequence (A) or between upper, middle and lower flowers from self-pollinating orchids with numerous flowers (B).

Figure 9.
Effect of climate on flowering and capsule production for two self-pollinating orchids; (A) Disa bracteata; (B) Microtis media.
Figure 9.
Effect of climate on flowering and capsule production for two self-pollinating orchids; (A) Disa bracteata; (B) Microtis media.

Figure 10.
Average fruit set rates for species and years sampled separated into pollination trait groups.
Figure 10.
Average fruit set rates for species and years sampled separated into pollination trait groups.

Figure 11.
The influence of leaf, flower and capsule sizes or plant density on seed production for orchids with different pollination traits (one point per species); (A) Comparison of leaf and flower area; (B) Seed output per spike relative to total flower area per spike; (C) Seedpod volume per spike relative to fruit set; (D) The impact of plant density on fruit set (small group average/overall mean) relative to average pollination. Orchids with sexually deceptive pollination have larger symbols.
Figure 11.
The influence of leaf, flower and capsule sizes or plant density on seed production for orchids with different pollination traits (one point per species); (A) Comparison of leaf and flower area; (B) Seed output per spike relative to total flower area per spike; (C) Seedpod volume per spike relative to fruit set; (D) The impact of plant density on fruit set (small group average/overall mean) relative to average pollination. Orchids with sexually deceptive pollination have larger symbols.

Figure 12.
Graphs showing links between clonality and seed production for 22 orchid species (one point each); (A) Seed output relative to the proportion of plants with flowers; (B) The impact of plant density on fruit set (small group average/mean) relative to the average rate relative to clonality (see 11D for comparison).
Figure 12.
Graphs showing links between clonality and seed production for 22 orchid species (one point each); (A) Seed output relative to the proportion of plants with flowers; (B) The impact of plant density on fruit set (small group average/mean) relative to the average rate relative to clonality (see 11D for comparison).

Figure 13.
The influence of climate on fruit set for orchids with different pollination traits (one point for each species); (A) Average daily maximum temperature during flowering and seed production; (B) Total rainfall during flowering relative to fruit set; (C) Rainfall accumulation during fruit set relative to seed production; (D) Autumn rainfall (March to May) relative to fruit set by Leporella fimbria; (E) Rainfall during flowering and fruit set for visually deceptive Thelymitra species (4 species, 3 years); (F) The relationship between rainfall accumulation and average daily maximum temperature during flowering for the samples in Figure 13E. Overall pollination rates were low for many orchids, especially visually deceptive species, but were generally in agreement with earlier studies of the same or similar species (Table 1). However, Erickson observed that most flowers of Diuris sp. and Caladenia flava were visited by native bees and almost all received pollen, suggesting that historic activity levels by native bees were much higher in the 1920’s [44]. This provides some evidence of long-term declines in pollinator numbers (e.g., Hallman et al. [50]), but other local data supporting this was not available. There also was a tendency for orchid pollination to vary considerably between years and sites (e.g., 4–18% for C. arenicola) suggesting that pollinator activity is highly variable in space and time. The data in Table 1 is often lower for Australian orchids than expected from global averages. For example, a threshold for effective pollination was set at 50% by Neiland and Wilcock, which is much higher than most Australian orchids [1]. However, there was no evidence that low rates of pollination were detrimental, since rates as low as 5% for species such as T. macrophylla and D. magnifica where local populations were stable or expanding. These species often occurred in large groups, where competition for insect vectors would have reduced fruit set. Pollination outcomes per flowering plant are more substantial (above 20%) and probably more meaningful than outcomes per flower, due to large floral displays. Two Caladenia species with sexually deceptive pollination by thynnine wasps had fruit set rates that may be of concern (Table 1), but long-term studies are required to confirm this. These wasps are parasites of other insects, especially larvae of Scarabaeoidea beetles, but we know very little about their ecology [51,52].
Figure 13.
The influence of climate on fruit set for orchids with different pollination traits (one point for each species); (A) Average daily maximum temperature during flowering and seed production; (B) Total rainfall during flowering relative to fruit set; (C) Rainfall accumulation during fruit set relative to seed production; (D) Autumn rainfall (March to May) relative to fruit set by Leporella fimbria; (E) Rainfall during flowering and fruit set for visually deceptive Thelymitra species (4 species, 3 years); (F) The relationship between rainfall accumulation and average daily maximum temperature during flowering for the samples in Figure 13E. Overall pollination rates were low for many orchids, especially visually deceptive species, but were generally in agreement with earlier studies of the same or similar species (Table 1). However, Erickson observed that most flowers of Diuris sp. and Caladenia flava were visited by native bees and almost all received pollen, suggesting that historic activity levels by native bees were much higher in the 1920’s [44]. This provides some evidence of long-term declines in pollinator numbers (e.g., Hallman et al. [50]), but other local data supporting this was not available. There also was a tendency for orchid pollination to vary considerably between years and sites (e.g., 4–18% for C. arenicola) suggesting that pollinator activity is highly variable in space and time. The data in Table 1 is often lower for Australian orchids than expected from global averages. For example, a threshold for effective pollination was set at 50% by Neiland and Wilcock, which is much higher than most Australian orchids [1]. However, there was no evidence that low rates of pollination were detrimental, since rates as low as 5% for species such as T. macrophylla and D. magnifica where local populations were stable or expanding. These species often occurred in large groups, where competition for insect vectors would have reduced fruit set. Pollination outcomes per flowering plant are more substantial (above 20%) and probably more meaningful than outcomes per flower, due to large floral displays. Two Caladenia species with sexually deceptive pollination by thynnine wasps had fruit set rates that may be of concern (Table 1), but long-term studies are required to confirm this. These wasps are parasites of other insects, especially larvae of Scarabaeoidea beetles, but we know very little about their ecology [51,52].

Table 1.
Typical fruit set rates for common or rare (R) Australian orchids with different pollination strategies. Data are from studies that include substantial numbers of orchids (n > 100) and are the percentage of flowers where seedpods develop fully from natural pollination (see Table A1 for insect pollinator references).
Table 1.
Typical fruit set rates for common or rare (R) Australian orchids with different pollination strategies. Data are from studies that include substantial numbers of orchids (n > 100) and are the percentage of flowers where seedpods develop fully from natural pollination (see Table A1 for insect pollinator references).
| No | Strategy | Insect Group | Season | Orchid Genus | Average Gruit Set per Flower |
|---|---|---|---|---|---|
| 1A | Visual deception | bees, etc. | winter | Pheladenia Diuris | P. deformis—0%* D. brumalis—2% [23] |
| 1B | Visual deception | bees, beetles, flies, etc. | spring | Caladenia | C. flava—most flowers [44],—25% [18], —18% [83], —32% [77], —48%*; C. latifolia—47%*; C. polychroma—16% [77]; C. melanema R—8% [84]; |
| Diuris | D. corymbosa—most flowers [44]; D. magnifica—11% [83], —6%*; D. maculata—15% [22]; | ||||
| 1C | Visual deception | Flies, bees, etc. | spring - early summer | Thelymitra | T. antennifera—3.2% [66]; T. ixioides—28% [85]; T. macrophylla—4.8%*; T. graminea—17%*; T. fuscolutea (drought stressed)—7%* |
| 2A | Some nectar | bees | autumn | Eriochilus | E. dilatatus—95% pollen deposition [44], —16%* |
| 2B | Some nectar | bees | spring | Pyrorchis | P. nigricans—31%* |
| 3A | Sexual deception | male ant | autumn | Leporella | L. fimbria—18% [86], —17%* |
| 3B | Sexual deception | fungus gnats | winter or spring | Pterostylis | P. recurva—50%, [18]), —50%*; P. sanguinea—44%, [83], —23%*; P. ectypha—39%* |
| 3C | Sexual deception | thynnine wasps | spring | Caladenia | C. arenicola—4% [87], —10% [77], —18% [83], —13%*; C. discoidea—16%*; C. huegelii R—7% [77], —2% (Brundrett unpublished); C. graniticola R—6% [84]; C. williamsiae R—15% [84] |
| Drakaea | D. glyptodon—0 to 59% 9 sites [65]; D. elastica R—5% (Brundrett unpublished) | ||||
| 4A | Self–pollination | none | spring | Thelymitra | T. vulgaris—95%*; T. benthamiana—90%*; T. circumsepta R—96% [85] |
| 4B | Self–pollination (some nectar) | none (usually) | spring – summer | Microtis Disa | M. media—93%*; D. bracteata (drought stressed)—59%* |
Notes: Data with an * are from this study.
Table 2.
Evidence that pollinator availability and behavior or resource limitations restrict orchid pollination. These take the form of four alternative hypotheses that may all be correct (see text for supporting data and references).
Table 2.
Evidence that pollinator availability and behavior or resource limitations restrict orchid pollination. These take the form of four alternative hypotheses that may all be correct (see text for supporting data and references).
1. Pollinator abundance is the key limiting factor to pollination success.
|
2. Pollination is primarily limited by learned avoidance responses by insects in deceptive species.
|
3. Pollinator abundance is strongly limited by seasonal climatic factors.
|
4. Flowering and seedpod development are limited by resources available to the plant.
|
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brundrett, M.C. A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia. Diversity 2019, 11, 123. https://doi.org/10.3390/d11080123
AMA Style
Brundrett MC. A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia. Diversity. 2019; 11(8):123. https://doi.org/10.3390/d11080123
Chicago/Turabian StyleBrundrett, Mark C. 2019. "A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia" Diversity 11, no. 8: 123. https://doi.org/10.3390/d11080123
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.