

Alien Species in the Pioneer and Ruderal Vegetation of Ukraine
by
 , , ,
, , ,
Dmytro V. Dubyna
1,
Tetiana P. Dziuba
1,*,
Svitlana M. Iemelianova
1,2,
Vira V. Protopopova
1,3 and
Myroslav V. Shevera
1,3 1
Department of Geobotany and Ecology, M.G. Kholodny Institute of Botany, National Academy of Sciences of Ukraine, 01004 Kyiv, Ukraine
2
Department of Botany and Zoology, Faculty of Science, Masaryk University, 602 00 Brno, Czech Republic
3
Ferenc Rákóczi II Transcarpathian Hungarian College of Higher Education, 90200 Beregove, Ukraine
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(12), 1085; https://doi.org/10.3390/d14121085
Submission received: 8 November 2022
/
Revised: 27 November 2022
/
Accepted: 6 December 2022
/
Published: 8 December 2022
(This article belongs to the Special Issue Changes and Evolution of Flora and Vegetation under Human Impacts)
Abstract
:Invasions of nonnative plants are widely recognized as one of the major threats to the biodiversity of natural ecosystems on a global scale. Pioneer and ruderal habitats are the primary locations for the penetration of alien plants. Both pioneer and ruderal vegetation are very close in their genesis and beginning of development; therefore, a comparative analysis of their alien components and historical trends would contribute to clarifying the direction of successional changes and the possible management of destructive processes caused by anthropogenic influences in different types of habitats. The results of a structural and comparative analysis of the alien fractions of the coenofloras of the pioneer and ruderal vegetation of Ukraine indicated that the systematic, biomorphological, ecological, and geographical structures of these species show a high similarity, according to many of the main indicators, which allows them to successfully implement a strategy of invasion, particularly in communities characterized by instability and weak coenotic connections. It was established that the ecotopes of both types of vegetation are very favorable to the penetration and establishment of alien species; however, disturbed habitats of the ruderal type are more prone to invasions. In the communities of both pioneer and ruderal vegetation, alien species can become successfully established at the coenotic level, forming phytocoenoses of different hierarchical ranks. The results of this study will contribute to the identification of general patterns of invasions and the optimization (management) of disturbed and unstable natural ecosystems.
1. Introduction
Especially high rates of nonnative species invasions were recorded in the 20th century, which is probably a consequence of the intensification of world trade, cultivation of plants in agriculture, and introductions from botanical and private gardens [1]. These nonnative species pose a real threat to the biodiversity of natural ecosystems.
Pioneer and ruderal habitats are places for the primary penetration of alien species. Newly formed natural or permanently disturbed anthropogenic habitats have a significant percentage of free ecological niches and weak coenotic connections. Alien species with a high adaptive potential and wide ecological amplitude can easily penetrate such unstable phytocoenoses, change their structure by displacing the native component, and dominate the community. This ecological and coenotic strategy is characteristic of alien species with high invasive potential and of transformers. They are able to form a series of replacement communities with different successional statuses [2].
There is special interest in the comparative analysis of the alien fractions of the coenofloras of pioneer and ruderal vegetation. Although exhibiting some degree of similarity, they differ, first of all, in the history of their formation and development [3]. In pioneer communities, these processes take place with a predominance of elements of an autochthonous group of species; in ruderal communities, allochthonous ones predominate [4,5,6]. Pioneer and ruderal vegetation differ in the floristic and phytocoenotic “closedness” of their communities. The size of the phytocoenotic barrier, which mostly depends on the environment-creating capacity of the main edificators in pioneer vegetation, despite its generally low degree, is higher in pioneer communities than in ruderal ones. At the same time, the value of the phytocoenotic barrier increases in the community as it approaches others, e.g., zonal types of vegetation [7], which is one of the strategies of the development of pioneer communities. Therefore, the high specific weight of alien species in the composition of pioneer vegetation is due to the greater availability of free ecological niches favorable for the development of representatives of this fraction [4]. The anthropic factor also has an increasing influence. In ruderal vegetation, the role of free ecological niches is combined with an almost complete absence of a phytocoenotic barrier in the community. Therefore, in the formation of phytodiversity of the alien fraction of ruderal vegetation, the soil and microclimatic mosaic play a greater role than in the pioneer mosaic. This was confirmed by the results of a comparative structural analysis of the coenofloras of pioneer and ruderal vegetation. Such an analysis of their alien components and historical trends appears to be informative for clarifying the direction of successional changes and the possible management of destructive processes caused by anthropogenic influences in different types of biotope. Identifying factors leading to successful invasion provides an opportunity to predict and mediate future anthropogenic invasions [8].
The high degree of adventization of Ukrainian flora has been defined in a wide range of studies. Publications on this topic from 1787 to 2022 were summarized in nine issues of bibliographic references [9], which included 2905 works. These indicators increased this year. The directions of research are as follows: floristics, chorology, biology (primarily of weeds), ecology, distribution control and control means, taxonomy, population, vegetation, molecular genetics, economic evaluation, paleobotany, etc. The species of the listed genera that have been studied most completely include Ambrosia, Amaranthus, Impatiens, Heracleum, Solidago, Xanthium etc. The study of the alien flora has over 220 years of history and is connected with the general research of the flora of Ukraine and its regions. Initial and fragmentary information about alien plants was recorded in the works of naturalists of the 18th–19th centuries; several publications began to indicate the origin of some alien species. Taliev was the first who noted that human activity had become a botanical and geographical factor. In the 1920s, a study of alien plants as a special group began, e.g., Kotov investigated the mass resettlement of alien plants associated with human activity. In the second half of the 20th century, a comprehensive study of the regional alien flora of the Forest Steppe, Steppe, and Crimea was started with the works by Protopopova and Kozhevnikova. The special study of the synanthropic fraction flora of Ukraine was started with the works by Protopopova. In this study, particular attention was paid to the correlations between the distribution of alien plants and changes in the plant cover, determined by human activity. One of the interesting directions in the studies of that time was the research of the anthropogenic transformation of ecosystems; principally, their phytodiversity (Burda). Another trend in the study of alien plants of the flora of Ukraine is the especially comprehensive research of the urban flora of Ukraine, initiated by Burda. The adventization of the plant cover of the protected areas of Ukraine began to be studied in the 1980s. The study of phytoinvasion was focused on the different directions of these phenome, e.g., ecology, participation and role of invasive species in different vegetation types and habitats, dynamics trends of its distribution, transformers in different regions, and its regional features, etc. [10]. In Ukraine, studies on this topic are combined into general investigations, in accordance with the classic and modern approaches used, methods, techniques, and the classifications and terminology of national and foreign scientists.
A Short History of the Development of the Territory of Ukraine
The most noticeable changes in the plant cover occurred in the forest steppe zone of Ukraine, where there was agriculture by the third–second millennia BC; and in the 12th–14th centuries AD, in some areas, the agricultural landscape became a stable phenomenon. The first cities also arose in this area to meet the needs of agriculture and protection from enemies, and forests were cut down over large areas. With each subsequent century, the processes of plant cover transformation accelerated and contributed to the emergence of various transformations, mostly ruderal ecotopes. In the forest zone, radical changes in vegetation occurred in the 17th–19th centuries, when many forests were destroyed due to the development of industry. As a result, conditions were created for the spread of open landscape species, especially alien species. In the 20th century, melioration contributed to increasing the adventization of the flora of this zone. Radical changes in the natural terrain of the steppe zone began at the end of the 18th century, after the annexation of Crimea. In the initial period of economic development of the steppe zone, animal husbandry developed intensively. Overgrazing contributed to the disappearance of turf grasses and bushes and their replacement by annuals, as well as the destruction of the surface horizon of the soil, the emergence of numerous failed areas, and the transformation of steppe areas into a semidesert. The secondary associations were more xerophytic than the original ones and were typical of more southern and southeastern zones. Thus, the development of animal husbandry played a significant role in the introduction and spread of alien species. The next significant changes in the environment took place at the end of the 18th century, as a result of the development of grain farming. Almost all the land suitable for sowing was ploughed, which contributed to the spread of weeds in agricultural fields and the increase in ruderal habitats. At the same time, industry was progressing, the establishment and development of new cities occurred, and the population increased. Significant areas of the steppe were turned into semideserts and pastures, connected by a network of roads and railways. New ecotopes, urban wastelands, landfills, railway embankments, and human-made environments (dumps of various strata, quarries, and construction debris) appeared. They were mainly inhabited by eurytopian species, forming unstable communities penetrated by alien and local species from more southerly regions. The formation of the alien fraction flora in Ukraine was partially influenced by the location of the country. Since ancient times, important trade routes have passed through here. Trade and wars with nomads, Crimean Tatars, and Turks contributed to the introduction of diasporas of species from the Eastern Mediterranean, Iran–Turanian, and Asia Minor origin. The majority of archaeophytes of the country belong to these groups by origin. The largest number of such species is in the forest steppe zone. In the 11th century, trade with the countries to the east and Byzantium occupied a significant place in the economy. Trade was also conducted with the countries of Central Europe.
The development of railway transport caused an increase in the introduction of alien plants from Europe. Significant changes in the species composition of the alien flora of Ukraine occurred with the development of transcontinental connections between European countries. Plants were mainly introduced to Ukraine from southwestern and western European countries, as well as to the Azov and Black Sea ports from other continents. Erigeron canadensis and Amaranthus retroflexus probably entered Ukraine during this time. The number of species of American origin and their role in the plant cover of the transformed habitats grew rapidly, significantly changing the character of the plant community. In the first half of the 19th century, the alien flora of the country was still at the stage of forming its stable core. In the 20th century, the expansion of American species (e.g., Ambrosia artemisiifolia, Iva xanthiifolia, Matricaria discoidea, Xanthium orientale subsp. riparium, etc.) in some regions completely changed the character of the alien flora. At the same time, some species that were quite widespread, mainly in agricultural crops (e.g., Agrostemma githago, Persicaria lapathifolia subsp. leptoclada), began to noticeably decrease in area. Perhaps this was due to the cessation of cultivation of the crops they associated with. The dynamics of the alien flora also changed: the processes of colonization of anthropogenic ecotopes by new species slowed down. Presently, there is mainly an expansion of the range of habitats, an increase in the degree of naturalization, an exchange of species between anthropogenic floristic complexes, and an increase in their coenotic role. Recently, the process of plants escaping from botanic gardens, private parks, and introduction centers has been highly prominent. A significant reduction in the lag phase of some of these has been observed, which is probably explained by the fact that most of these species are already adapted to local conditions in European countries.
In the formation of the alien fraction of the flora of Ukraine, the main role is played by flora from the Ancient Mediterranean, as well as the North American prairies, areas of the Rocky Mountains, the Mandrean Region, local areas within the central and western parts of the North American continent, and the arid Chilean–Patagonian region of South America. Some of the indicators of the biological and ecological structures of the alien flora of Ukraine testify to its Mediterranean character. At present, the main trends of its development are determined by the xerophytization of the flora under the influence of climatic changes. This is facilitated by the strengthening of the xerothermic regime of anthropogenic ecotopes compared to natural ones. Despite the significant advantage of species that originated from different regions of the Ancient Mediterranean region and their extended period of existence in the conditions of Ukraine, North American species have the widest amplitude of adaptation. Species of Asian origin (especially East and Central Asian) also adapt quickly [11].
At present, in this period of macroscale reduction of biodiversity, excessive land use, urbanization, and climate change, it is relevant to review the current state of pioneer and ruderal vegetation and the presence and distribution of alien species.
The hypothesis that forms the basis of this work is as follows: we assumed that the alien fractions of pioneer and ruderal vegetation should be largely similar.
Our study focused on the analysis of the (a) composition, (b) structure, (c) peculiarity, and (d) historical trends of alien species of the coenofloras of the pioneer and ruderal vegetation of Ukraine.
2. Materials and Methods
2.1. Study Area
Ukraine is located within forest, forest steppe, and steppe zones and has two mountain systems (Ukrainian Carpathians and Crimean Mountains), while the southern coast of Crimea is located in the ecoregion of the Crimea submediterranean forest complex. All these zones contain smaller classification units. Each one of these is characterized by a distinct vegetation cover type, with a distinct species composition, including invasive plants, and by a certain state of preservation and transformation.
2.2. The Dataset
Phytosociological data from the databases “Anthropogenic vegetation of Ukraine” (registration number in GIVD [12] EU-UA-011) and “Pioneer vegetation of Ukraine” [13] were the basis for establishing the syntaxonomic diversity and compiling lists of alien species of the pioneer and ruderal vegetation of Ukraine. A total of 10,423 phytosociological relevés were analyzed. The syntaxonomy of the specified types of vegetation was developed based on the method of floristic classification, which made it possible to establish the composition of their coenoflora. By coenoflora, we understand the floral composition of phytocoenoses of certain classes of vegetation.
The objects of the study were the alien fractions of the coenofloras of the pioneer and ruderal vegetation of Ukraine.
2.3. Analysis
The analysis of the structures of these fractions has been carried out according to various approaches: systematic by Tolmachev [14], life form by Raunkiaer [15], geographical by Takhtajan [16], and historical-geographical classification of alien species by Kornaś [17]. The indices proposed by Jackowiak [18] were used to assess the degree of anthropogenic transformation of coenofloras and for demonstration of the main trends in the process of invasion. We used indices of:
- anthropogenization (IAn), as the proportion between alien plant species and all species in the studied flora (coenofloras);
- archaeophytization (IArch), as the proportion of archaeophytes in all species pools of each coenoflora;
- kenophytization (IKen), as the proportion of kenophytes in all species pools of each coenoflora;
- modernization (IM), as the proportion between kenophytes and all species in the studied coenofloras;
- fluctuation (instability) (IF), as the proportion of diaphytes (ergasiophytes and ephemerophytes) in all species pools of each coenoflora.
Calculation of the environmental parameters for a set of alien species participating in communities (in fact, phytosociological relevés) of certain classes of vegetation was performed using the standardized point scales of synphytoindication ecological amplitudes developed by Didukh [19]. These scales allow one to conduct an ordination analysis using 12 factors, including seven edaphic factors: soil moisture, moisture variability, soil aeration, available nitrogen content, soil acidity, salt regime, and carbonate content; and five climatic factors: thermo-regime, ombroregime, continentality, cryoregime, and light intensity. Phytoindication analysis of the interactions between alien species of plant communities and environmental factors and the range of their distribution was carried out using the method of synphytoindication [19], the JUICE program [20], and basic statistical analysis in the Statistica 7.0 package (StatSoft Inc., Tulsa, OK, USA). The set of alien species ranges and optima for each of the 12 environmental factors were calculated with the Statistica 7.0 package. In the ecological assessment of alien species, average scores of all types of phytocenosis were used, which together represent the average value. Data were represented in “box-and-whisker” form. The “boxes” represent a interquartile range (25–75% of the values observed) and correspond to the ecological optimum of the set of alien species, and the “whiskers” represent minimum and maximum values, while the middle point represents the median.
2.4. Nomenclature
3. Results and Discussion
The syntaxonomic structure of the ruderal vegetation in Ukraine includes 210 associations belonging to 36 alliances, 16 orders, and 8 classes, e.g., Stellarietea mediae, Artemisietea vulgaris, Galio-Urticetea, Bidentetea, Robinietea, Epilobietea angustifolii, Polygono-Poetea annuae, and Plantaginetea majoris [23]. The pioneer vegetation of Ukraine is represented by 84 associations, which are united into 21 alliances, 10 orders, and 10 classes, e.g., Cakiletea maritimae, Ammophiletea, Crithmo-Staticetea, Crypsietea aculeatae, Therosalicornietea, Helichryso-Crucianelletea maritimae, Festucetea vaginatae, Koelerio-Corynephoretea canescentis, Isoëto-Nanojuncetea, and Bidentetea, and representing psammophytic, halophytic, littoral, and hygrophilous plant communities [24,25,26].
The coenoflora of the pioneer vegetation in Ukraine, in general, consists of 844 species of vascular plants from 338 genera and 80 families [27], including 113 species of alien plants. The ruderal vegetation includes 1637 species, of which 325 are aliens. The total index of anthropogenization was 13.4% for pioneer vegetation and 19.8% for ruderal vegetation. The index of ruderal vegetation exceeds this indicator for the alien flora of Ukraine (14%) [28], and that of pioneer vegetation approaches it. This indicates that anthropogenic habitats are more favourable for the introduction of alien species.
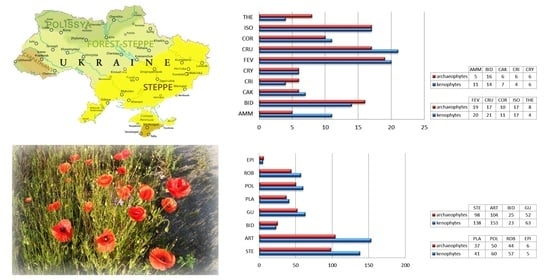
There were 103 common alien species in both vegetation types. There was an overlap of the allochthonous component in the composition of pioneer vegetation by 91.2% (Figure 1). A significant percentage of common species indicates the similarity of the conditions of their habitats and that both pioneer and ruderal phytocoenoses are subject to invasions of a certain group of alien species. These species are mostly eurytopic, with wide ecological plasticity to all abiotic factors, and are characterized by rapid adaptation to rapidly changing environmental conditions. Among them the highest frequency across the two vegetation types included Amaranthus blitoides, Ambrosia artemisiifolia, Anisantha tectorum, Artemisia absinthium, Atriplex prostrata, A. tatarica, Ballota nigra, Bidens connata, B. frondosa, Capsella bursa-pastoris, Lepidium draba, Centaurea diffusa, Cichorium intybus, Descurainia sophia, Echinochloa crus-galli, Echinocystis lobata, Erigeron annuus, E. canadensis, Hordeum murinum, Iva xanthiifolia, Juncus tenuis, Lactuca serriola, Oenothera biennis, Oxybasis rubra, Portulaca oleracea, Sisymbrium loeselii, S. orientale, S. polymorphum, Tribulus terrestris, Tripleurospermum inodorum, Xanthium orientale subsp. riparium, and X. strumarium.
Some common species are those that occur in the coenoflora of the Bidentetea class, because this class combines both the pioneer vegetation of seasonally flooded and nutrient-rich river alluvial sediments and lake shores, and the vegetation of nitrified anthropogenic habitats [29]. It should be noted that in the analysis of ruderal vegetation, we included relevés from the Bidentetea class in anthropogenically disturbed areas, while the pioneer vegetation of these classes included communities that periodically flooded bank river habitats [26]. Therefore, we considered this class to be part of both pioneer and ruderal vegetation, but with different compositions of actual phytosociological material.
Among the plant communities of pioneer vegetation, the highest degree of anthropogenization was observed in the coenoses of overmoistened nitrified ecotopes, e.g., Bidentetea, Cakiletea maritimae, and Isoëto-Nanojuncetea classes (Figure 2a). These habitats are favourable for the appearance of alien species and the successful implementation of their ecological and coenotic strategies. It is believed that river banks are particularly vulnerable to the invasion of nonnative elements [30]. The coenofloras of the Ammophiletea and Helichryso-Crucianelletea maritimae classes are close to the general index. The smallest number of alien species was noted in the Koelerio-Corynephoretea canescentis and Therosalicornietea plant communities. The small percentage of the allochthonous component in the coenofloras of these classes can be explained by the limiting influence of the leading factors of ecological differentiation of their groups. A high concentration of mineral salts in the soil is a leading factor for Therosalicornietea, and the increased acidity of the soil solution is a leading factor for Koelerio-Corynephoretea canescentis.
In the coenoflora of ruderal vegetation, the largest number of alien species was recorded in the coenoses of the Polygono-Poetea annuae, Stellarietea mediae, and Plantaginetea majoris classes (Figure 2b). A significant number of alien species was noted in the plant community of the Artemisietea vulgaris and Bidentetea classes. The high rates of adventization in these classes are due to the location of these coenoses in the initial stages of ecological and coenotic series and due to permanent disturbance of their ecotopes with the transfer of diaspores of alien species through trampling and compacting the soil. As in pioneer vegetation, alien species of ruderal coenoflora gravitate towards nutrient-rich ecotopes. Galio-Urticetea coenoses are less affected by alien species, as tall herbaceous thickets have high competitiveness and stability [31]. The lowest percentage of the allochthonous component was recorded in the coenoflora of the Epilobietea angustifolii class (5.8%), which contains the vegetation of cuttings. This is explained by the preservation of their close connection with the previous natural forest vegetation, which is one of the most sustainable ecosystems [32,33,34,35], as well as their isolation from other habitats, which can be a source of propagules of alien species [31]. The quantitative distribution of alien species in the coenoflora is consistent with the data of Mollot et al. [36], where species richness losses from alien invaders differed by habitat type.
The taxonomical spectrum of the alien fraction of the coenofloras of the pioneer and ruderal vegetation of Ukraine reveals a great similarity, which indicates the unidirectionality of their developmental processes. Asteraceae, Brassicaceae, Poaceae, Chenopodiaceae, and Fabaceae occupy the first places among the ten leading families in both vegetation types (Table 1). In general, this distribution corresponds to a similar distribution of the alien flora of Ukraine as a whole [11] and indicates the Mediterranean character of the spectra and the essential role of arid floristic centers in the formation of the nonnative core of the studied coenofloras. Some differences in the distribution of the leading families were observed, starting from the sixth place (Table 1). The presence of Polygonaceae and Amaranthaceae among the leading families indicates a recent strengthening of the role of North American floristic centers in the ruderal community.
The taxonomical spectra of invasive species at the global scale demonstrate a somewhat different distribution. According to a meta-analysis in 2017, the first places in taxonomical classification of invaders belonged to Poaceae, Asteraceae, Fabaceae, and Polygonaceae [36]. The subsequent places, occupied by Myrtaceae, Balsaminaceae, Pinaceae, Caprifoliaceae, etc. are atypical for our latitudes and less common.
In the systematic spectra at the genus level in both pioneer and ruderal vegetation, genera represented by one species were prevailing. The largest polymorphism in the alien fraction of pioneer coenoflora was characteristic of the typical arid genera Sisymbrium, Lepidium, Anisantha, and Atriplex. For ruderal ones, the leading genera in the spectra were Xanthium, Amaranthus, Chenopodium, Sisymbrium, and Veronica (Table 2).
Raunkiær’s life-form spectra of species of the alien fractions of both pioneer and ruderal vegetation are similar to the corresponding spectrum of nonnative species of Ukraine [11]. Therophytes prevail in the pioneer (58.4%) and ruderal (57.2%) phytocoenoses (Figure 3). The predominance of therophytes, which are usually the first to occupy newly formed and disturbed ecotopes, is characteristic of pioneer and anthropogenic habitats and reflects the mechanisms of plant adaptation to conditions of regular disturbance [37,38,39]. Significant participation of therophytes is also characteristic of unfavorable, sometimes extreme, ecological conditions, in which groups of the studied types of vegetation were formed.
Hemicryptophytes and cryptophytes are much less represented. However, it is among these that species with vegetative reproduction, having an advantage in occupying new territories are found [40,41], e.g., Solidago canadensis, Helianthus tuberosus, Heracleum mantegazzianum, H. pubescens, Reynoutria japonica, Oenothera biennis, Onopordum acanthium, Cenchrus longispinus, etc.
An insignificant number of phanerophytes and chamaephytes are characteristic only for coenoses of the Robinietea and Epilobietea angustifolii classes, e.g., Acer negundo, A. saccharinum, Amorpha fruticosa, Elaeagnus angustifolia, Ailanthus altissima, Robinia pseudoacacia, Caragana arborescens, Celtis occidentalis, Cotinus coggygria, Fraxinus pennsylvanica, Gleditsia triacanthos, Morus alba, M. nigra, Quercus rubra, etc. They occur mainly in tree and shrub plantations. Juvenile individuals of Acer negundo, Amorpha fruticose, and Elaeagnus angustifolia were also recorded in herbaceous communities.
The ecological structure of the alien fractions of the studied coenofloras, on the one hand, differs in the conditions of habitats of some groups of phytocoenoses and, on the other side, reveals the similarity of those coenoflora that develop in similar ecological conditions. The ecological characteristics and conditions of habitats are the determining factors in plant invasions [42]. It is believed that the characteristics and habitat requirements of alien species need to be similar to native ones [43,44]. The success of a species in a habitat depends on its ecophysiological or morphological characteristics, which can be interpreted as determinants of distribution along ecological gradients. The functional characteristics of the values of ecological indicators help to clarify the mechanisms and processes that determine the ecological niche of a plant [45]. To establish these ecological niches, most European researchers use the Ellenberg bioindication system [46,47,48,49,50]; however, for many kenophytes, the values of ecological indicators according to Ellenberg are currently missing. To establish the peculiarities of their ecological structure, we used the national system of ecomorphs followed by Didukh [19], which is comparable to Ellenberg’s scale.
Submesophytes, mesophytes, and subxerophytes prevail in the general spectrum of hydromorphs of the alien fractions of the pioneer and ruderal vegetation communities, which indicates their adaptability to ecotopes with moderate and insignificant soil moisture (Figure 4). Species of the hygrophytic group prevail in the Bidentetea, Isoëto-Nanojuncetea, Galio-Urticetea, and Plantaginetea majoris classes, and species of the xerophytic group prevail in the Festucetea vaginatae, Helichryso-Crucianelletea maritimae, and Koelerio-Corynephoretea canescentis classes, which are of natural origin.
Regarding the variability of soil moisture during the vegetative period, in the coenosis of both pioneer and ruderal vegetation, hemihydrocontrastophyles and hydrocontrastophyles prevail (Figure 5). This emphasizes their ability to adapt to rapidly changing environmental conditions.
With respect to soil acidity, the alien components of the pioneer and ruderal vegetation of Ukraine prefer soils with slightly acidic or neutral pH values. They were distributed mainly between the subacidophilic and neutrophilic ecological groups (Figure 6). The analysis of the flora of different syntaxonomic units of the highest rank, according to these indices, revealed that the majority of plant communities prefer subacidophilic conditions, and the phytocoenoses of the Stellarietea mediae, Artemisietea vulgaris, and Ammophiletea classes are more frequently found in neutral soils. Alien species of the Cakiletea maritimae and Crypsietea aculeatae classes gravitate toward alkaline soils.
Regarding the salt regime of edaphotopes, in both pioneer and ruderal vegetation, the alien species tolerate a lack of mineral salts well and grow in mesotrophic ecological conditions (Figure 7). The representatives of the Bidentetea, Galio-Urticetea, and Isoëto-Nanojuncetea classes belong to the semieutrophic group.
By nitrogen content, alien species of both pioneer and ruderal vegetation gravitate toward heminitrophilic and nitrophilic edaphic conditions (Figure 8). Nitrophilic and eunitrophilic conditions are more characteristic of the Isoëto-Nanojuncetea, Bidentetea, Galio-Urticetea, Plantaginetea majoris, Polygono-Poetea annuae, and Robinietea classes.
Regarding the carbonate content in the substrate, the majority of alien species of both pioneer and ruderal vegetation are represented by the acarbonatophilic group; they prefer neutral ecotopes with an insignificant content of carbonates in the soil (Figure 9). Species of the Isoëto-Nanojuncetea, Koelerio-Corynephoretea canescentis, and Plantaginetea majoris classes belong to the hemicarbonatophobes. Hemicarbonatophilous species growing on soils enriched with carbonates belong to the community of Cakiletea maritimae, Crypsietea aculeatae, and Therosalicornietea, and partially to the Koelerio-Corynephoretea canescentis, Festucetea vaginatae, Plantaginetea majoris, and Polygono-Poetea annuae classes.
The species that prefer open and well-lit habitats and belong to the heliohyte and hemiheliophyte groups are dominant in the spectra of the heliomorphs of the alien fractions of both the pioneer and ruderal vegetation of Ukraine (Figure 10). Shade-tolerant plants, hemisciophytes, occur mainly in artificial tree plantations. Heliophytes dominate in the Bidentetea, Stellarietea mediae, Artemisietea vulgaris, Polygono-Poetea annuae, and Plantaginetea majoris classes. Coenofloras of Isoëto-Nanojuncetea, Cakiletea maritimae, Crypsietea aculeatae, and Festucetea vaginatae classes are characterized by a wide ecological amplitude regarding this factor.
Regarding their arrival time, the proportions of kenophytes (51.5%) and archaeophytes (49.5%) were almost the same in the coenofloras of pioneer vegetation (Figure 11a).
The general ratio between these groups is 1.0:1.02, and this is significantly different from the alien fraction of the flora of Ukraine (1.0:4.5) [28]. Such a proportion between migrochronological elements represents the specificity of the process of penetration of alien species into pioneer coenoses.
The newly formed habitats are a fairly favourable recipient environment and are characterized by a free ecological space for the emergence of new species and their distribution, including alien species. The rapidly changing and sometimes extreme abiotic conditions in which communities of pioneer vegetation are formed is a limiting factor for the free penetration and rapid adaptation of nonnative plants.
In ruderal vegetation, the proportion of archaeophytes and kenophytes differs in the other direction from the ratio for the alien flora of Ukraine and is 1.0:1.73. That is, kenophytes penetrate more intensively into ruderal coenoses. This has been facilitated by the development of transport communications, the expansion of trade relations, and environmental changes [51]. Active urbanization processes [2,52,53], agricultural intensification, and global climate changes [54] also contribute to the dispersal of kenophytes in plant communities on a global scale.
The share of archaeophytes in the anthropogenic vegetation of Ukraine is 7.3%, which is comparable to their percentage in Italy (from 11.6% in artificial surfaces to 18.3% in agricultural land) [55], Switzerland (9.7%) [54], Great Britain (6.7%) [56], and the Balkans region (4.3%) [57]; however, it is much lower than that in the ruderal vegetation of the Czech Republic (35.5%) [56].
The share of archaeophytes in the pioneer vegetation of Ukraine is 6.5%. This value is comparable to the average in pioneer coenoses of Central Europe, with values of 6.0% in dry grassland and 8.1% in inland saline grass and herb-dominated habitats in the Czech Republic [58].
Kenophytes represent the majority of the alien fractions of pioneer and ruderal coenofloras, which indicates the active processes of the addition of an allochthonous component to the floristic composition at the current historical stage. Archaeophytes prevail only in the communities of the Epilobietea angustifolii, Bidentetea, Therosalicornietea, and Crithmo-Staticetea classes (Figure 11b). This is explained by the fact that the Epilobietea angustifolii coenoses of forest clearings still have a connection with the previous forest vegetation, which is quite resistant to the introduction of new alien species. In coenoses of the Bidentetea class, the introduction, adaptation, and further distribution of kenophytes are limited by rapidly changing environmental conditions, while in the Therosalicornietea and Crithmo-Staticetea classes, they are limited by the extreme conditions of excessive salinity and wind–wave action.
In total, 6.8% of kenophytes participate in the coenoses of pioneer vegetation in Ukraine. The share of kenophytes in separate types of biotopes, e.g., coastal habitats, is 4.0%, which is twice as high as in the Mediterranean islands of Italy (2.0%) [59] but closer to the values in Catalonia and Great Britain (3.3% each). In the saline habitats in the Czech Republic, kenophytes account for 1.6%, while in Catalonia, the value is 2.2%, and in Great Britain, the value is 0.7% [56]. In Ukraine, pioneer saline biotopes contain 2.4% kenophytes.
The percentage share of kenophytes in the ruderal vegetation of Ukraine is 12.6%, and this indicator exceeds the Central European average. In the anthropogenic habitats of the Northwestern Balkans, it is 8.4% [57]; in Central Europe, it is 6.9%; in Great Britain, it is 4.5%; and in Catalonia, it is 5.3% [56].
Regarding the degree of naturalization, among the alien plants in pioneer and ruderal vegetation of Ukraine, the majority are epoecophytes (Figure 12a,b). In the coenoses of ruderal vegetation, there are twice as many ergasiophytes and half as many agrio-epoecophytes and agriophytes. The number of unstable elements, ephemerophytes, is approximately the same in both types of vegetation.
The proportions of these groups differ slightly in the Bidentetea and Galio-Urticetea classes, which are seminatural and have a higher proportion of agriophytes. In the coenoses of Therosalicornetea, Crypsietea aculeatae, and Crithmo-Staticetea, an absence of diaphytes is observed, which is due to the extreme conditions of the development of these coenoses (in particular, excessive moisture and/or salinity of the substrate), which significantly limits the free penetration and further naturalization of alien species.
A specific feature of the ecological structure of the alien fraction of the coenofloras of the pioneer and ruderal vegetation of Ukraine, both as a whole and within individual groups, is the predominance of eurytopic species with a very wide ecological plasticity for all abiotic factors. This explains their rapid adaptation to rapidly changing environmental conditions in pioneer vegetation coenoses and to the constant and varying intensity of human influence in ruderal coenoses.
In the geographical structure spectrum of the alien fractions of the coenofloras of pioneer and ruderal vegetation, species with Ancient Mediterranean origins dominate. In the plant communities of both types of vegetation, the majority of representatives are characterized by Mediterranean–Iranian–Turanian and Mediterranean origins (Figure 13). The proportions of species with North American and Asian origins are similar. Species with origins in Europe and other regions are much less represented. Such ratios are preserved in almost all coenofloras of the pioneer and ruderal vegetation classes. The species of North American origin, which are adapted to more mesophytic conditions, are dominant in Bidentetea, Galio-Urticetea, and Plantaginetea majoris.
Indices of archaeophytization, kenophytization, modernization, and instability of flora were calculated to determine the degree of anthropogenic transformation of coenofloras of the pioneer and ruderal vegetation classes (Table 3). High indices of archaeophytization and kenophytization of coenofloras of the Bidentetea and Plantaginetea majoris classes indicate the ancient and stable naturalization of nonnative species in the floristic composition of these communities and testify to active processes of the introduction of alien species in the current period. There is a deepening of the processes of transformation of the flora of these communities under the influence of modern anthropogenic factors.
The Ammophiletea, Artemisietea vulgaris, and Stellarietea mediae classes are characterized by the highest indices of modernization of coenofloras, which indicates significant qualitative changes in the species composition of coastal sand dune communities and the most common ruderal phytocoenoses due to eukenophytes, especially those with high invasiveness. Instead, the specific ecological conditions of the coenoses of abrasive beaches and cliffs, periodically flooded salt marshes, and forest clearings are the limiting factor of intensive invasions in the Crithmo-Staticetea, Therosalicornietea, and Epilobietea angustifolii classes. The indices of modernization are the lowest here.
The general index of instability of the coenoflora of the pioneer vegetation classes of Ukraine is low (1.5%). Thus, it can be argued that the more the habitat is disturbed, the more it undergoes invasions and experiences subsequent species loss [36,60].
The distributions of alien species in the main natural regions of Ukraine differ in composition, number of species, character of their distribution, and role in plant cover. Only 15% of species are common to all regions. The most specific alien fraction flora of Crimea, Polissia (forest zone), and the forest steppe zone are most similar to each other. The Carpathians and the Crimea differ the most.
Certain regularities in the distribution of alien species were found, which indicates that their distribution is not chaotic. In the anthropogenic ecotopes of all regions, alien species predominate in the synanthropic fraction flora; their number gradually increases in the southern direction, and their percentage with respect to the total sum of synanthropic species is higher in the south than in the north. The majority of them are in the forest steppe zone and Crimea, in the regions that were most anciently settled by man. Similar trends are characteristic of some other groups of alien flora; for example, the number of archaeophytes increases in the northern direction, and the number of kenophytes increases in the southern direction. This distribution is determined, not only by natural and climatic conditions, but also by the character of changes in the plant cover as a result of its transformation [11].
Along the latitudinal gradient, the number of highly active invasive species decreases in the southern direction (from 61 to 39), and the composition of stable elements, including agriophytes and agrio-epoecophytes, as well as epoecophytes, increases. In addition, towards the south, the degree of naturalization of these species decreases, and the amplitude of distribution narrows. The latter is explained by climatic conditions, since most of the species of this group belong to the xeromesophytes and mesophytes, and in the warmer conditions of the southern regions, they achieve only a low degree of naturalization. The specificity of the species composition of highly active invasive species at both regional and zonal levels is small. Only five out of 60 species were recorded in the forest zone, two out of 44 species were recorded in the steppe zone, and four out of 18 species were recorded in Crimea [61].
At the zonal level, a balance of the species composition of the stable alien component in the synanthropic flora is observed. This is evidenced by the following: The ratio of the sum of ephemerophytes and ergasiophygophytes and the sum of all species of zonal synanthropic floras in all regions of Ukraine indicates the existence within these floras of a relative equilibrium of an unstable component limited by the threshold of species diversity. This component in regional synanthropic floras of Ukraine is 1/5 (e.g., Carpathians—0.175, plain forest territories—0.197, forest steppe—0.200, steppe—0.200, Crimea—0.195). Thus, in all regions, it is approximately 0.2. Minor fluctuations of this indicator occur with the degree of anthropogenic transformation of the plant cover in different regions, increasing in the most transformed areas [11].
To a large extent, zonation also determines the contribution of mainly Mediterranean species to the autochthonous or allochthonous fractions of the flora. Some of the species of Mediterranean origin, which in Crimea and in the south of the steppe zone are within the natural range, are alien in the northern regions of Ukraine.
Nonnative species have already become not only a stable component of the floristic structure of pioneer and ruderal coenoses of Ukraine but also have high fidelity and are characteristic of many syntaxa of the highest and middle hierarchical levels. Therefore, mostly archaeophytes (47 species) were noted as being part of the diagnostic blocks of alliances, orders, or even classes. The participation of kenophytes (34 sp.) was also quite significant (Table 4).
At the level of associations, alien species also participate in the community as diagnostic, constant, and dominant species. According to the results of the analysis, the number of alien species in the ruderal vegetation is significantly higher than that in the pioneer vegetation. For example, in the classes of pioneer vegetation Ammophiletea and Therosalicornietea, they are completely absent in the diagnostic blocks. The coenoses of the Stellarietea mediae and Artemisietea vulgaris classes are characterized by the largest number of alien species that are diagnostic (Table 5).
Alien species present over a long time become the stable components of the community of subsequent associations, e.g., Amarantho retroflexi-Echinochloetum cruris-galli, Setario pumilae-Echinochloetum cruris-galli, Digitarietum ischaemii, Portulacetum oleracei (Figure 14a), Ambrosietum artemisiifoliae, Brometum tectorum, Hordeetum murini, Galio aparines-Papaveretum rhoeadis (Figure 14b), Artemisietum annuae, Chamaeplietum officinalis, Ivaetum xanthiifoliae (Figure 14c), Conyzo canadensis-Lactucetum serriolae, Asclepiadetum syriacae, Achilleo millefolii-Grindelietum squarrosae, Erigeretum canadensi-acris (Figure 14d), Carduo acanthoidis-Onopordetum acanthii, Xanthietum spinosi (Figure 14e), Urtico dioicae-Heracleetum mantegazziani, Polygonetum cuspidati (Figure 14f), Myosoto aquatici-Bidentetum frondosae, etc.
Analysis of the alien species frequency of the studied coenofloras shows that the highest values of constancy belong to Anisantha tectorum and Descurainia sophia. They occur in the coenoses of a few vegetation classes.
Diagnostic types of associations are usually dominant among alien species. Often, their coverage is 80–100%. As a result of the dominance of invasive species in the habitats invaded by them, native species are suppressed, and the species composition of communities as a whole decreases [62]. It remains to be seen, in the future, how dominant kenophytes behave at local, regional, and temporal scales, but some studies have indicated that their participation in assemblages declines in the later stages of restoration succession. These data support spontaneous succession as a positive recovery option and emphasize the early detection of potentially problematic species [2].
The most stable component of the coenofloras of pioneer and ruderal vegetation is the constant alien species in the syntaxa of these types. This group includes 60 species, including 29 archaeophytes and 31 kenophytes. In pioneer coenoflora, archaeophytes and kenophytes are almost equal. In ruderal coenoflora, there are only seven species of archaeophytes, and almost twice as many kenophytes. This is explained by the higher competitiveness of the latter. By origin, Ancient Mediterranean species predominate among archaeophytes (including 6 Mediterranean, 14 Mediterranean-Iran-Turanian and 7 Iranian-Turanian), and there are also one South European, one Southeast Asian, and two of unknown origin. These data reflect the ancient and various migration routes of alien species due to the various relations of Ukraine with the countries to the east. Among the kenophytes (introduced in 1809–1986), North American species predominate (12), and this group also includes eight Ancient Mediterranean species (6 Mediterranean–Iranian–Turanian, 1 Mediterranean, 1 Mediterranean–Turanian), one Mediterranean-Asian, one Mediterranean–South Asian; one Balkan–Asia Minor, and five European species (1 European, 4 South European), with one species of unknown origin. Archaeophytes and kenophytes differ both in their life forms and in relation to the moisture regime. Therophytes predominate (23) among the archaeophytes; this group also includes seven hemicryptophytes and four therophytes/hemicryptophytes. Approximately half the kenophytes are therophytes; with eight hemicryptophytes, eight therophytes/hemicryptophytes, and two phanerophytes.
A significant change in the character of the alien flora, associated with the appearance of new sources of species introduction, has occurred as a result of the expansion of international trade relations; the diversification of transport routes; and the faster adaptation, dynamism, breadth of distribution, and higher coenotic activity of species with transcontinental and transzonal types of area. During this period, the introduction of new species mainly occurred from Europe by rail and from other continents by sea. The adaptation of species was facilitated by the diversification of ecotopes and agents of plant spread and preliminary adaptation to the natural conditions of Europe. The rate of naturalization of alien species and the number of invasive species have increased (from 3 archaeophytes to 7 kenophytes). Thus, the results of the analysis of constant species showed that the first aliens from adjacent geographical zones, which were less competitive, spread to ecotopes with reduced competition and more homogeneous conditions, mainly in weedy places. Primarily Mediterranean species, they were adapted to habitats with similar conditions. The mass and speed of introduction of alien species with transcontinental and transzonal types of area, often preadapted to new conditions, was more successful and significantly changed the character of the plant communities. The kenophytes introduced at that time had a wider ecological amplitude and polyvariance of their adaptation complex, which provided them with a greater competitive ability and led to a wide distribution. There is a tendency to accelerate the naturalization of introduced species and the formation of ruderal phytocoenoses.
Thus, despite the general tendency toward xerophytization of anthropogenically transformed habitats, the increasing of role of transcontinental and transzonal species of the mesophytic group, mostly North American, was observed in the studied coenofloras. In the group of archaeophytes, there is an increase in the cenotic activity of some species, which is manifested in the formation of the plant community; for example, with participation of Anisantha tectorum and Apera spica-venti and native species. The expansion of the adaptive capabilities of other species occurred due to the formation of new microspecies/morphotypes; for example, with the participation of Portulaca oleracea, due to which their area and spectrum of habitats have expanded.
The group of highly active invasive species poses a particular threat to biodiversity. They are able to impair the vitality of populations of native species, suppressing their growth, development, reproduction, and stability, and reducing density. The potential threat of hybridization with native species is dangerous, especially for those that are diagnostic in syntaxa of different ranks, which leads to the local disappearance of representatives of the local flora and irreversible changes in the composition of plant communities [63]. The community of the Bidentetea class, in which 14 species of invasive plants were recorded (53.8% of all found in the pioneer coenoses), and the Isoëto-Nanojuncetea class, in which 13 taxa (50%) of this group were noted, were the most vulnerable to invasions. The coenoses of these classes, which are formed on disturbed ruderalized, well-moistened, and nitrified ecotopes, are extremely favourable for the introduction, growth, and active distribution of plants with high invasiveness. Under such conditions, these communities are a potential source of threat, not only for ecologically close coenoses, in particular river bank habitats, but also for other more mesophytic ones. For the extremely vulnerable and unique nanoephemeretum (cl. Isoëto-Nanojuncetea), invasions are particularly dangerous. They cause changes in the floristic composition and coenotic structure of these communities and are also able to transform environmental conditions, by enriching the soil with nitrogen compounds or consuming a significant amount of water resources.
In the ruderal vegetation of Ukraine, 52 invasive species were found. The most common of these are Erigeron canadensis, E. annuus, Ambrosia artemisiifolia, Anisantha tectorum, Helianthus tuberosus, Iva xanthiifolia, Impatiens parviflora, I. glandulifera, Solidago canadensis, Amorpha fruticosa, Apera spica-venti, Bidens frondosa, Centaurea diffusa, Echinocystis lobata, Heracleum mantegazzianum, Robinia pseudoasacia, Portulaca oleracea, Quercus rubra, Xanthium orientale subsp. riparium, and Asclepias syriaca. They often form monodominant communities, transforming the ecosystem and reducing natural biodiversity. In the last 20 years, in connection with global warming and the xerophytization of soil and hydrological conditions, there has been a trend toward a spatial displacement of some taxa in the southern to northern regions. This trend is especially noticeable for phytocoenoses with the participation of thermophilic species [54]. The community of Brometum tectorum, Hordeetum murini, Descurainietum sophiae, Cardarietum drabae, Anisantho-Artemisietum austriacae, and Setario viridis-Erigeronetum canadensis associations expanded their coenotic range beyond the borders of the steppe zone. The active development of infrastructure facilities, communication systems, transport routes (especially railways), and urbanization processes will contribute to these and other synchronological changes in the future.
Biodiversity positively influences ecosystem functions and subsequently services [64], and biological invasions impair the reliability and quality of these services. As human activities also depend on the well-being of ecosystems [65], it is important to understand and identify the environmental conditions that favor species invasion. The results of our research have shown that although pioneer and ruderal vegetation are very close in their genesis and beginning of development, disturbed habitats are more prone to invasions.
The results of our research correspond to the hypothesis of increased competitiveness of invasive species when disturbed, fragmented, and heterogeneous areas, without almost any concurrent competitors, serve as ecological niches for them [66,67]. According to our data, the habitats of both pioneer and ruderal vegetation are very favourable for the penetration and stabilization of the invasion of alien species at the coenotic level. Ruderal vegetation is particularly vulnerable.
Biological invasions are recognized as a factor in global changes as well as species richness and functioning of ecosystems [36,66], and are used to define biogeography in an era of global change [68]. Most scientific studies on biological invasions have focused on the evaluation of hypotheses at the taxonomic level [69]. We focused our investigation on the analysis of alien species participation at the coenotic level in plant communities, which are the basis of functioning of ecosystems. Number of species, projective cover, fidelity, and frequency of occurrence of alien species are more informative than presence/absence data in floristic studies [57].
The results of our research indicate that alien species successfully form coenotic connections, particularly in communities of pioneer and ruderal vegetation, forming new associations and displacing native species, and thereby having a transformative impact on biodiversity.
Of special scientific interest are the coenoflora involved in the formation of a community in which the leading role is played by alien species with wide ecological amplitudes, e.g., Bidens tripartita and B. frondosa. Some phytocoenologists attribute their communities (Bidentetea class) to pioneer ephemeral or swamp vegetation [70,71,72,73,74,75], and others include synanthropic vegetation [29,76] or azonal communities of partly native or partly anthropogenic origin [77]. We believe that class coenoses are formed both in pioneer alluvial ecotopes and in disturbed ruderal ecotopes [23,26]. Such a dualistic approach, in our opinion, allows us to identify the alien component and its distinguishing features in the community, the formation of which is driven by the same edificators but can be separated into temporal, edaphic, microclimatic, and ultimately evolutionary measurements. This approach is aimed at clarifying the role of pioneer vegetation as a basic component of the formation of communities of other types of vegetation, particularly zonal vegetation. The next task is to determine the degree of influence of the alien fraction on the nature of the identified processes and their vectorability. Given the modern anthropogenic transformations of vegetation and the excessive vulnerability of the pioneer vegetation, this will allow us to validate methods of minimizing these negative impacts, in particular, of the alien fraction on the state of pioneer communities unique in the history of their evolution and integral to the optimization of ruderal communities.
These and other questions are of particular importance in the current historical period of Ukraine, when as a result of military operations, significant areas occupied by natural vegetation have undergone changes due to military activity. In the territories affected by explosions and other impacts (destruction of plant cover by machinery, fires, etc.), processes of regrowth are taking place. As shown by previous studies (e.g., in the Kyiv region), the character of the regrowth depends on the magnitude of the military transformation of ecotopes, the primary vegetation, and the degree of coenotic organization, as well as the presence of diaspores, in particular alien species.
The general trends found in our study are consistent with other studies that reported that anthropogenic vegetation is the most invaded habitat type, due to strong disturbances and management [42,56,57,78]; pioneer vegetation, especially riparian freshwater and coastal sand dune systems, is very vulnerable to invasion [30,79,80], and the negative impacts of biological invasion on the species richness of phytosystems as a whole [62,81,82].
The formation of the general character of spontaneous vegetation with the participation of alien species in Central European countries depends on the height above sea level, climate, substrate, and level of urbanization [2]. The important role of macroecological conditions in the distribution of alien species was emphasized in the work of Šilc et al. [57]. Other studies have shown that the success of invasions is also determined by the following factors: plant strategies [44,83], local abundance, geographic range size, habitat breadth in naturalized distributions [60], fluctuation in resource availability in plant communities [84,85], phylogenetic and functional similarities to natives [86,87,88,89], and high propagule pressure [43]. Undoubtedly, all these factors collectively affect the position and role of alien species in phytocoenoses, especially in unstable ones such as pioneer and anthropogenic habitats. Studying the impact of alien species in these types of vegetation at the coenotic level will help identify the potential consequences of alien species invasion and provide valuable information for landscape management and nature conservation [90].
Effective global plant diversity management and plant invasion management policies require the implementation of integrated frameworks for observing, modelling, and forecasting various forms of anthropogenic biodiversity change processes at the scale of regional landscapes, aiming at preserving biodiversity in new anthropogenic plant communities [91]. A set of measures aimed at minimizing the impact of invasions is required. The organization of monitoring of the emergence and spread of alien species, the direction of dynamic processes, the creation of relevant databases, and the implementation of preventive measures aimed at preventing invasions of nonnative species in phytocoenoses are the main tasks. Management of pioneer and ruderal communities should also include methods of limiting the acclimatization of invasive species and their replacement with native plants [92].
4. Conclusions
The hypothesis about the similarity of the alien fraction of the coenofloras of the pioneer and ruderal vegetation of Ukraine was confirmed, and the reasons for this natural phenomenon were clarified. Their essence lies in the closeness of ecogenesis of these two types of vegetation. The naturally higher rate of anthropogenization of ruderal vegetation is due to a larger spectrum of existing ecological niches than that of pioneer ones. It has been proven that alien species successfully naturalize in the composition of pioneer and ruderal vegetation at the coenotic level and form phytocoenoses of different hierarchical ranks. A tendency to accelerate the increase in the degree of naturalization and the coenotic role of alien species within the transcontinental and transzonal area, as well as the occurrence of local cases of their active dispersal in the studied coenofloras were observed. In recent years, there has been an expansion of the coenotic range of communities of the studied types of vegetation, with the participation of thermophilic species.
The results of our research can contribute to the understanding of the role of alien species in the processes of synanthropization of vegetation, they will be useful for understanding the behavior of alien species in coenotically unstable communities, will help to reveal general patterns of invasion, and will contribute to the optimization of disturbed ecosystems. Completing the mentioned tasks is especially relevant in connection with the military transformation of large areas in Ukraine and the restoration of natural vegetation cover, which during the stages of overgrowth will take place with the participation of pioneer vegetation. As previous studies have shown, alien species will play a major role in the composition, in the trends, and directions of dynamic successions.
Author Contributions
Conceptualization, D.V.D.; methodology, D.V.D., T.P.D., M.V.S., V.V.P. and S.M.I.; software, T.P.D. and S.M.I.; validation, D.V.D., T.P.D., S.M.I. and M.V.S.; formal analysis, T.P.D. and S.M.I.; investigation, D.V.D., T.P.D., V.V.P., M.V.S. and S.M.I.; data curation, D.V.D. and V.V.P.; writing—original draft preparation, T.P.D., S.M.I., V.V.P. and M.V.S.; writing—review and editing, D.V.D., T.P.D., S.M.I., V.V.P. and M.V.S.; visualization, T.P.D.; supervision, D.V.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to Bogdan Jackowiak (Adam Mickiewicz Uniwersity, Poznan) and the Editors of Diversity Journal for valuable and difference help and advice.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, M.; Vítovcová, K.; Řehounková, K.; Müllerová, A.; Janečková, P.; Pospíšilová, P.; Prach, K. Alien species in vegetation succession: Participation, temporal trends and determining factors in various central European series. Biol. Invasions 2021, 23, 3435–3445. [Google Scholar] [CrossRef]
- Kuzmichev, A.I. Hyhrophyllous Flora of South-West of Russian Lowland and Its Genesis; Parfyonov, V.I., Ed.; Hydrometizdat: Sankt-Petersburg, Russia, 1992. [Google Scholar]
- Ilyin, M.M. Flora of littoral and desert in their connections. Sov. Bot. 1947, 5, 249–256. [Google Scholar]
- Zosimovich, V.P. Evolution of Goosefoot and the Genus Beta. Scientific Papers on Selection, Agricultural Technology, Mechanization and Crop Protection; Ahrarna Nauka: Kyiv, Ukraine, 1958; Volume 38, pp. 59–73. [Google Scholar]
- Zozulin, G.M. Historical suites of vegetation of the European part of the USSR. Bot. J. 1973, 58, 1081–1092. [Google Scholar]
- Zlobin, Y.A. Phytocoenotic Barrier. Materials on the Dynamics of the Vegetation Cover; Reports at the Interuniversity Conference in September 1968; Vladimir Printing House of the Glavpoligrafprom: Vladimir, Russia, 1968; pp. 9–10. [Google Scholar]
- Sax, D.F.; Stachowicz, J.J.; Brown, J.H.; Bruno, J.F.; Dawson, M.N.; Gaines, S.D.; Grosberg, R.K.; Hastings, A.; Holt, R.D.; Mayfield, M.M.; et al. Ecological and evolutionary insights from species invasions. Trends Ecol. Evol. 2007, 22, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Burda, R.I.; Protopopova, V.V.; Shevera, M.V.; Kucher, O.O.; Koniakin, S.M. Alien Species in the Flora of Ukraine: Years and Authors. Bibliographic List/Kyiv. 2022. Available online: http://www.botany.kiev.ua/doc/bibliograf9.pdf (accessed on 1 November 2022).
- Shevera, M.V.; Protopopova, V.V.; Burda, R.I.; Zavialova, L.V.; Kucher, O.O.; Korniyenko, O.M. Historical overview of the studies of alien flora of Ukraine. Acta Horti Bot. Bucurest 2018, 45, 5–32. [Google Scholar]
- Protopopova, V.V. Synanthropic Flora of Ukraine and Ways of Its Development; Naukova Dumka: Kyiv, Ukraine, 1991. [Google Scholar]
- Dengler, J.; Jansen, F.; Glöckler, F.; Peet, R.K.; De Cáceres, M.; Chytrý, M.; Ewald, J.; Oldeland, J.; Lopez-Gonzalez, G.; Finckh, M.; et al. The Global Index of Vegetation-Plot Databases (GIVD): A new resource for vegetation science. J. Veget. Sci. 2012, 22, 582–597. [Google Scholar] [CrossRef]
- Dubyna, D.; Dziuba, T.; Iemelianova, S. Database of pioneer vegetation of Ukraine. In Proceedings of the 15th meeting of the German Working Group on Vegetation Databases, Potsdam, Germany, 2–4 March 2016; p. 42. [Google Scholar]
- Tolmachev, A.I. Introduction to Plant Geography; Leningrad Univ. Press: Sankt-Petersburg, Russia, 1974. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography, Being the Collected Papers of C. Raunkiær; Gilbert-Carter, H.; Fausbøll, A.; Tansley, A.G., Translators; Oxford University Press: Oxford, UK, 1934. [Google Scholar]
- Takhtajan, A.L. The Floristic Regions of the World; Nauka: Sankt-Petersburg, Russia, 1978. [Google Scholar]
- Kornaś, J. Geograficzno-historyczna klasyfikacja roślin synantropijnych. Mater. Zakładu Fitosocjologii Stosow. Uniw. Warsz. 1968, 25, 33–41. [Google Scholar]
- Jackowiak, B. Antropogeniczne Przemiany Flory Roślin Naczyniowych Poznania; Seria Biologia; Wydawnictwo Naukowe Uniwersytetu Adama Mickiewicza: Poznań, Poland, 1990; Volume 42. [Google Scholar]
- Didukh, Y.P. The Ecological Scales for the Species of Ukrainian Flora and Their Use in Synphytoindication; Phytosociocentre: Kyiv, Ukraine, 2011. [Google Scholar]
- Tichy, L. JUICE, software for vegetation classification. J. Veget. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://www.europlusmed.org (accessed on 22 November 2022).
- Dubyna, D.V.; Dziuba, T.P.; Iemelianova, S.M.; Bagrikova, N.O.; Borysova, O.V.; Borsukevych, L.M.; Vynokurov, D.S.; Gapon, S.V.; Gapon, Y.V.; Davydov, D.A.; et al. Prodrome of the Vegetation of Ukraine; Naukova Dumka: Kyiv, Ukraine, 2019. [Google Scholar]
- Dubyna, D.; Iemelianova, S.; Dziuba, T.; Ustymenko, P.; Felbaba-Klushyna, L.; Davydova, A.; Davydov, D.; Tymoshenko, P.; Baranovski, B.; Borsukevych, L.; et al. Ruderal vegetation of Ukraine: Syntaxonomic diversity and territorial differentiation. Chornomors’k. Bot. Z. 2021, 17, 253–275. [Google Scholar] [CrossRef]
- Dubyna, D.V.; Dziuba, T.P.; Iemelianova, S.M.; Makhynia, L.M. Syntaxonomy and ecological differentiation of the pioneer vegetation of Ukraine. 1. Classes: Cakiletea maritimae, Ammophiletea, Crithmo-Staticetea, Crypsietea aculeatae, Therosalicornietea. Biosyst. Divers. 2020, 28, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Dubyna, D.V.; Dziuba, T.P.; Iemelianova, S.M.; Tymoshenko, P.A. Syntaxonomy and ecological differentiation of the pioneer vegetation of Ukraine. 2. Helichryso-Crucianelletea maritimae, Festucetea vaginatae, Koelerio-Corynephoretea canescentis classes. Biosyst. Divers. 2020, 28, 298–319. [Google Scholar] [CrossRef]
- Dubyna, D.V.; Dziuba, T.P.; Iemelianova, S.M.; Felbaba-Klushyna, L.M. Syntaxonomy and ecological differentiation of the pioneer vegetation of Ukraine. Classes: Isoëto-Nanojuncetea, Bidentetea. Environ. Soc.-Econ. Stud. 2021, 9, 32–52. [Google Scholar] [CrossRef]
- Dubyna, D.V.; Dvoretskyi, T.V.; Iemelyanova, S.M.; Dzyuba, T.P.; Tymoshenko, P.A. Taxonomic structure of coenofloras of the classes of pioneer vegetation of Ukraine. Ukr. Bot. J. 2017, 74, 421–430. [Google Scholar] [CrossRef]
- Protopopova, V.V.; Mosyakin, S.L.; Shevera, M.V. Plant Invasions in Ukraine as a Threat to Biodiversity: The Present Situation and Tasks for the Future; M.G. Kholodny Institute of Botany, NAS of Ukraine: Kyiv, Ukraine, 2002. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar]
- Tickner, D.P.; Angold, P.G.; Gurnell, A.M.; Mountford, J.O. Riparian plant invasions: Hydrogeomorphological control and ecological impacts. Prog. Phys. Geogr. 2001, 25, 22–52. [Google Scholar] [CrossRef]
- Medvecká, J.; Jarolímek, I.; Zaliberová, M. Dynamics and distribution of neophytes in ruderal vegetation of the Horná Orava Region (Northern Slovakia). Hacquetia 2010, 8, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Verheyen, K.; Baeten, L.; De Frenne, P.; Bernhardt-Römermann, M.; Brunet, J.; Cornelis, J.; Decocq, G.; Dierschke, H.; Eriksson, O.; Hédl, R.; et al. Driving factors behind the eutrophication signal in understorey plant communities of deciduous temperate forests. J. Ecol. 2012, 100, 352–365. [Google Scholar] [CrossRef]
- Förster, A.; Becker, T.; Gerlach, A.; Meesenburg, H.; Leuschner, C. Long-term change in understorey plant communities of conventionally managed temperate deciduous forests: Effects of nitrogen deposition and forest management. J. Veg. Sci. 2017, 28, 747–761. [Google Scholar] [CrossRef]
- Vild, O.; Šipoš, J.; Szabó, P.; Macek, M.; Chudomelová, M.; Kopecký, M.; Suchánková, S.; Houška, J.; Kotačka, M.; Hédl, R. Legacy of historical litter raking in temperate forest plant communities. J. Veg. Sci. 2018, 29, 596–606. [Google Scholar] [CrossRef]
- Adolf, C.; Tovar, C.; Kühn, N.; Behling, H.; Berrío, J.C.; Dominguez-Vázquez, G.; Figueroa-Rangel, B.; Gonzalez-Carranza, Z.; Islebe, G.A.; Hooghiemstra, H.; et al. Identifying drivers of forest resilience in long-term records from the Neotropics. Biol. Lett. 2020, 16, 20200005. [Google Scholar] [CrossRef] [PubMed]
- Mollot, G.; Pantel, J.H.; Romanuk, T.N. The effects of invasive species on the decline in species richness: A global meta-analysis. Networks of Invasion: A Synthesis of Concepts. Adv. Ecol. Res. 2017, 56, 61–83. [Google Scholar] [CrossRef]
- Pyšek, P.; Prach, K.; Šmilauer, P.; Rejmánek, M.; Wade, M. Relating Invasion Success to Plant Traits: An Analysis of the Czech Alien Plora. Plant Invasions: Gerneral Aspects and Special Problems; Pyšek, P., Prach, K., Rejmanek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 39–60. [Google Scholar]
- Pfadenhauer, J.S.; Klötzli, F.A. Die Vegetation der Erde. Grundlagen, Ökologie, Verbreitung; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Axmanová, I.; Kalusová, V.; Danihelka, J.; Dengler, J.; Pergl, J.; Pyšek, P.; Večeřa, M.; Attorre, F.; Biurrun, I.; Boch, S.; et al. Neophyte invasions in European grasslands. J. Veget. Sci. 2021, 32, e12994. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Levin, D.A. Ecological Speciation: Lessons from Invasive Species. Syst. Bot. 2003, 28, 643–650. Available online: https://www.jstor.org/stable/25063912 (accessed on 22 November 2022).
- Simonová, D.; Lososová, Z. Which factors determine plant invasions in man-made habitats in the Czech Republic? Perspect. Plants Ecol. 2008, 10, 89–100. [Google Scholar] [CrossRef]
- Vedder, D.; Leidinger, L.; Cabral, J.S. Propagule pressure and an invasion syndrome determine invasion success in a plant community model. Ecol. Evol. 2021, 11, 17106–17116. [Google Scholar] [CrossRef]
- Dalle Fratte, M.; Bolpagni, R.; Brusa, G.; Caccianiga, M.; Pierce, S.; Zanzottera, M.; Cerabolini, B.E.L. Alien plant species invade by occupying similar functional spaces to native species. Flora 2019, 257, 151419. [Google Scholar] [CrossRef]
- Bartelheimer, M.; Poschlod, P. Functional characterizations of Ellenberg indicatorvalues—A review on ecophysiological determinants. Funct. Ecol. 2016, 30, 506–516. [Google Scholar] [CrossRef]
- Berg, C.; Welk, E.; Jäger, E.J. Revising Ellenberg’s indicator values for continentality based on global vascular plant species distribution. App. Veg. Sci. 2017, 20, 482–493. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Dřevojan, P.; Sádlo, J.; Zelený, D. Ellenberg-type indicator values for the Czech flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef] [Green Version]
- Rion, V.; Gallandat, J.D.; Gobat, J.M.; Vittoz, P. Recent changes in the plant composition of wetlands in the Jura Mountains. Appl. Veg. Sci. 2018, 21, 121–131. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Brooker, R.W.; O’Brien, D.; Genney, D. Using species records and ecological attributes of bryophytes to develop an ecosystem health indicator. Ecol. Ind. 2019, 104, 127–136. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Carlón, L.; Fernández-Pascual, E.; Acedo, C.; Alfaro-Saiz, E.; Alonso Redondo, R.; Cires, E.; del Egido Mazuelas, F.; del Río, S.; Díaz-González, T.E.; et al. Checklist of the vascular plants of the Cantabrian Mountains. Mediterr. Bot. 2021, 42, e74570. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; van Kleunen, M.; Winter, M.; et al. Global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. USA 2018, 115, E2264–E2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Protopopova, V.V.; Shevera, M.V. Participation of alien species in urban floras in different botanical and geographical zones of Ukraine: A preliminart assessment. Biodiv. Res. Conserv. 2008, 11–12, 9–16. [Google Scholar]
- Protopopova, V.V.; Shevera, M.V.; Anischenko, I.M.; Teren’teva, N.G. Analysis of the species composition of kenophytes in urban floras of different phytogeographical zones of Ukraine using mathematical statistics methods. Ukr. Bot. J. 2010, 67, 536–546. [Google Scholar]
- Scherrer, D.; Bürgi, M.; Gessler, A.; Kessler, M.; Nobis, M.P.; Wohlgemuth, T. Abundance changes of neophytes and native species indicate a thermophilisation and eutrophisation of the Swiss flora during the 20th century. Ecol. Indic. 2022, 135, 108558. [Google Scholar] [CrossRef]
- Celesti-Grapow, L.; Alessandrini, A.; Arrigoni, P.V.; Assini, S.; Banfi, E.; Barni, E.; Bovio, M.; Brundu, G.; Cagiotti, M.R.; Camarda, I.; et al. Non-native flora of Italy: Species distribution and threats. Plant Biosyst. 2010, 144, 12–28. [Google Scholar] [CrossRef]
- Chytrý, M.; Maskell, L.C.; Pino, J.; Pyšek, P.; Vilà, M.; Font, X.; Smart, S.M. Habitat invasions by alien plants: A quantitative comparison among Mediterranean, subcontinental and oceanic regions of Europe. J. Appl. Ecol. 2008, 45, 448–458. [Google Scholar] [CrossRef]
- Šilc, U.; Vrbničanin, S.; Božić, D.; Čarni, A.; Dajić Stevanović, Z. Alien plant species and factors of invasiveness of anthropogenic vegetation in the Northwestern Balkans—A phytosociological approach. Cent. Eur. J. Biol. 2012, 7, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Chytrý, M.; Pyšek, P.; Tichý, L.; Knollová, I.; Danihelka, J. Invasions by alien plants in the Czech Republic: A quantitative assessment across habitats. Preslia 2005, 77, 339–354. [Google Scholar]
- Pretto, F.; Celesti-Grapow, L.; Carli, E.; Brundu, G.; Blasi, C. Determinants of non-native plant species richness and composition across small Mediterranean islands. Biol. Invasions 2012, 14, 2559–2572. [Google Scholar] [CrossRef]
- Fristoe, T.S.; Chytrý, M.; Dawson, W.; Essl, F.; Heleno, R.; Kreft, H.; Maurel, N.; Pergl, J.; Pyšek, P.; Seebens, H.; et al. Dimensions of invasiveness: Links between local abundance, geographic range size, and habitat breadth in Europe’s alien and native floras. Proc. Natl. Acad. Sci. USA 2021, 118, e2021173118. [Google Scholar] [CrossRef] [PubMed]
- Protopopova, V.V.; Shevera, M.V. Invasive species in the flora of Ukraine. I. The group of highly active species. GEO&BIO 2019, 17, 116–135. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Vilà, M.; Weber, E.; Antonio, C.M. Conservation Implications of Invasion by Plant Hybridization. Biol. Invasions 2000, 2, 207–217. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.-S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Pejchar, L.; Liba, P.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef]
- Murphy, G.E.P.; Romanuk, T.N. A meta-analysis of declines in local species richness from human disturbances. Ecol. Evol. 2014, 4, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Chabrerie, O.; Massol, F.; Facon, B.; Thevenoux, R.; Hess, M.; Ulmer, R.; Pantel, J.H.; Braschi, J.; Amsellem, L.; Baltora-Rosset, S.; et al. Biological Invasion Theories: Merging Perspectives from Population, Community and Ecosystem Scales. Preprints 2019, 2019100327. Available online: https://hal-amu.archives-ouvertes.fr/hal-02345539 (accessed on 22 November 2022).
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Lowry, E.; Rollinson, E.J.; Laybourn, A.J.; Scott, T.; Aiello-Lammens, M.; Gray, S.M.; Mickley, J.; Gurevitch, J. Biological invasions: A field synopsis, systematic review and database of the literature. Ecol. Evol. 2013, 3, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Martínez, S.; Fernández-González, F.; Loidi, J.; Lousã, M.; Penas, A. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobot. 2001, 14, 5–341. [Google Scholar]
- Bardat, J.; Bioret, F.; Botineau, M.; Boullet, V.; Delpech, R.; Genu, J.-M.; Haury, J.; Lacoste, A.; Rameau, J.-C.; Royer, J.-M.; et al. The Prodrome of French Vegetation; Muséum National d’Histoire Naturelle: Paris, France, 2004. [Google Scholar]
- Tzonev, R.T.; Dimitrov, M.A.; Roussakova, V.H. Syntaxa according to the Braun-Blanquet approach in Bulgaria. Phytol. Balc. 2009, 15, 209–233. [Google Scholar]
- Chytry, M. (Ed.) Vegetation of the Czech Republic. 3. Aquatic and Wetland Vegetation; Academia: Prague, Czech Republic, 2011. [Google Scholar]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef] [Green Version]
- Stępień, E.; Rosadziński, S. Communities of the Bidentetea class of small coastal river valleys of the Western Pomerania (Poland). Ecol. Montenegrina 2020, 28, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Abramova, L.M.; Golovanov, Y.M. Review of higher units of synanthropic vegetation in the European part of Russia. Work. State Nikitsk. Bot. Gard. 2016, 143, 7–15. [Google Scholar]
- Borhidi, A. Plant Associations in Hungary; Akadémiai Kiadó: Budapest, Hungary, 2003. [Google Scholar]
- Šilc, U. Synanthropic Vegetation: Pattern of Various Disturbances on Life History Traits. Acta Bot. Croat. 2010, 69, 215–227. Available online: https://hrcak.srce.hr/59732 (accessed on 22 November 2022).
- Felzines, J.-C.; Loiseau, J.-E. Les groupements fluviatiles des Bidentetea de la Loire moyenne, du bas Allier et de la Dordogne moyenne. Modifications apportées à la synsystématique de la classe des Bidentetea. Bull. Société Bot. Cent. Ouest 2005, 36, 159–204. [Google Scholar]
- Marignani, M.; Lussu, M.; Murru, V.; Bacaro, G.; Cogoni, A. Effect of Invasive Alien Species on the Co-Occurrence Patterns of Bryophytes and Vascular Plant Species—The Case of a Mediterranean Disturbed Sandy Coast. Diversity 2020, 12, 160. [Google Scholar] [CrossRef] [Green Version]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Tordoni, E.; Bacaro, G.; Weigelt, P.; Cameletti, M.; Janssen, J.A.M.; Acosta, A.T.R.; Bagella, S.; Filigheddu, R.; Bergmeier, E.; Buckley, H.L.; et al. Disentangling native and alien plant diversity in coastal sand dune ecosystems worldwide. J. Veget. Sci. 2021, 32, e12861. [Google Scholar] [CrossRef]
- Gallien, L.; Mazel, F.; Lavergne, S.; Renaud, J.; Douzet, R.; Thuiller, W. Contrasting the effects of environment, dispersal and biotic interactions to explain the distribution of invasive plants in alpine communities. Biol. Invasions 2015, 17, 1407–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Dostál, P.; Dawson, W.; van Kleunen, M.; Keser, L.H.; Fischer, M. Central European plant species from more productive habitats are more invasive at a global scale. Glob. Ecol. Biogeogr. 2013, 22, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Carboni, M.; Münkemüller, T.; Lavergne, S.; Choler, P.; Borgy, B.; Violle, C.; Essl, F.; Roquet, C.; Munoz, F.; Thuiller, W. What it takes to invade grassland ecosystems: Traits, introduction history and filtering processes. Ecol. Lett. 2016, 19, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Henn, J.J.; Yelenik, S.G.; Damschen, E.I. Environmental gradients influence differences in leaf functional traits between native and non-native plants. Oecologia 2019, 191, 397–409. [Google Scholar] [CrossRef]
- Tordoni, E.; Petruzzellis, F.; Nardini, A.; Bacaro, G. Functional divergence drives invasibility of plant communities at the edges of a resource availability gradient. Diversity 2020, 12, 148. [Google Scholar] [CrossRef] [Green Version]
- Kalusová, V.; Cubino, J.P.; Fristoe, T.S.; Chytrý, M.; van Kleunen, M.; Dawson, W.; Essl, F.; Kreft, H.; Mucina, L.; Pergl, J.; et al. Phylogenetic structure of alien plant species pools from European donor habitats. Glob. Ecol. Biogeogr. 2021, 30, 2354–2367. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Yu, F.; Barreiro, R. EDITORIAL: Plant invasions: Mechanisms, impacts and management. Flora 2020, 267, 151603. [Google Scholar] [CrossRef]
- Ellis, E.C.; Antill, E.C.; Kreft, H. All is not loss: Plant biodiversity in the anthropocene. PLoS ONE 2012, 7, e30535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rentch, J.S.; Fortney, R.H.; Stephenson, S.L.; Adams, H.S.; Grafton, W.N.; Anderson, J.T. Vegetation-site relationships of roadside plant communities in West Virginia, USA. J. Appl. Ecol. 2005, 42, 129–138. [Google Scholar] [CrossRef]
Figure 1.
The number of alien species in the coenofloras of pioneer and ruderal vegetation.

Figure 2.
Anthropogenization index values of coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine (in %). Here and below: AMM—Ammophiletea; CAK—Cakiletea maritimae; CRI—Crithmo-Staticetea; CRY—Crypsietea aculeatae; THE—Therosalicornietea; BID—Bidentetea; ISO—Isoëto-Nanojuncetea; FEV—Festucetea vaginatae; CRU—Helichryso-Crucianelletea maritimae; COR—Koelerio-Corynephoretea canescentis; STE—Stellarietea mediae; ART—Artemisietea vulgaris; GU—Galio-Urticetea; PLA—Plantaginetea majoris; POL—Polygono-Poetea annuae; ROB—Robinietea; EPI—Epilobietea angustifolii.
Figure 2.
Anthropogenization index values of coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine (in %). Here and below: AMM—Ammophiletea; CAK—Cakiletea maritimae; CRI—Crithmo-Staticetea; CRY—Crypsietea aculeatae; THE—Therosalicornietea; BID—Bidentetea; ISO—Isoëto-Nanojuncetea; FEV—Festucetea vaginatae; CRU—Helichryso-Crucianelletea maritimae; COR—Koelerio-Corynephoretea canescentis; STE—Stellarietea mediae; ART—Artemisietea vulgaris; GU—Galio-Urticetea; PLA—Plantaginetea majoris; POL—Polygono-Poetea annuae; ROB—Robinietea; EPI—Epilobietea angustifolii.

Figure 3.
Distribution of alien fraction species of coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine, according Raunkiær’s system of life forms.
Figure 3.
Distribution of alien fraction species of coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine, according Raunkiær’s system of life forms.


Figure 4.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by soil water regime. Here and below, the classes are in the following order: Stellarietea mediae; Artemisietea vulgaris; Galio-Urticetea; Bidentetea (ruderal); Plantaginetea majoris; Polygono-Poetea annuae; Epilobietea angustifolii; Robinietea; Bidentetea (pioneer); Isoëto-Nanojuncetea; Ammophiletea; Cakiletea maritimae; Crithmo-Staticetea; Crypsietea aculeatae; Therosalicornietea; Koelerio-Corynephoretea canescentis; Festucetea vaginatae; Helichryso-Crucianelletea maritimae.
Figure 4.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by soil water regime. Here and below, the classes are in the following order: Stellarietea mediae; Artemisietea vulgaris; Galio-Urticetea; Bidentetea (ruderal); Plantaginetea majoris; Polygono-Poetea annuae; Epilobietea angustifolii; Robinietea; Bidentetea (pioneer); Isoëto-Nanojuncetea; Ammophiletea; Cakiletea maritimae; Crithmo-Staticetea; Crypsietea aculeatae; Therosalicornietea; Koelerio-Corynephoretea canescentis; Festucetea vaginatae; Helichryso-Crucianelletea maritimae.

Figure 5.
Distribution of alien species of coenofloras of the pioneer and ruderal vegetation in Ukraine by soil moisture variability.
Figure 5.
Distribution of alien species of coenofloras of the pioneer and ruderal vegetation in Ukraine by soil moisture variability.

Figure 6.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by acidity.
Figure 6.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by acidity.

Figure 7.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by salt regime.
Figure 7.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by salt regime.

Figure 8.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by nitrogen content.
Figure 8.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by nitrogen content.

Figure 9.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by carbonate content.
Figure 9.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by carbonate content.

Figure 10.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by light intensity.
Figure 10.
Distribution of alien species of the coenofloras of the pioneer and ruderal vegetation in Ukraine by light intensity.

Figure 11.
Distribution of alien species of the coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine by arrival time.
Figure 11.
Distribution of alien species of the coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine by arrival time.

Figure 12.
Distribution of alien species of coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine by the degree of naturalization.
Figure 12.
Distribution of alien species of coenofloras of the classes of pioneer (a) and ruderal (b) vegetation in Ukraine by the degree of naturalization.

Figure 13.
Distribution of alien species in pioneer (a) and ruderal (b) vegetation in Ukraine by origin.
Figure 13.
Distribution of alien species in pioneer (a) and ruderal (b) vegetation in Ukraine by origin.


Figure 14.
Alien species in communities of association in Ukraine: (a) Portulacetum oleraceae on the railways in Fastiv (Kyiv region); (b) Galio aparines-Papaveretum rhoeadis in the vicinity of the village Myrotske (Kyiv region); (c) Ivaetum xanthiifoliae in the rice fields of the Odessa region; (d) Erigeretum canadensi-acris in the vicinity of the village Blystavytsa (Kyiv region); (e) Xanthietum spinosi in the vicinity of the village Zoria (Odesa region); (f) Polygonetum cuspidati in Beregove (Transcarpathian Region) (photo by T. Dziuba).
Figure 14.
Alien species in communities of association in Ukraine: (a) Portulacetum oleraceae on the railways in Fastiv (Kyiv region); (b) Galio aparines-Papaveretum rhoeadis in the vicinity of the village Myrotske (Kyiv region); (c) Ivaetum xanthiifoliae in the rice fields of the Odessa region; (d) Erigeretum canadensi-acris in the vicinity of the village Blystavytsa (Kyiv region); (e) Xanthietum spinosi in the vicinity of the village Zoria (Odesa region); (f) Polygonetum cuspidati in Beregove (Transcarpathian Region) (photo by T. Dziuba).



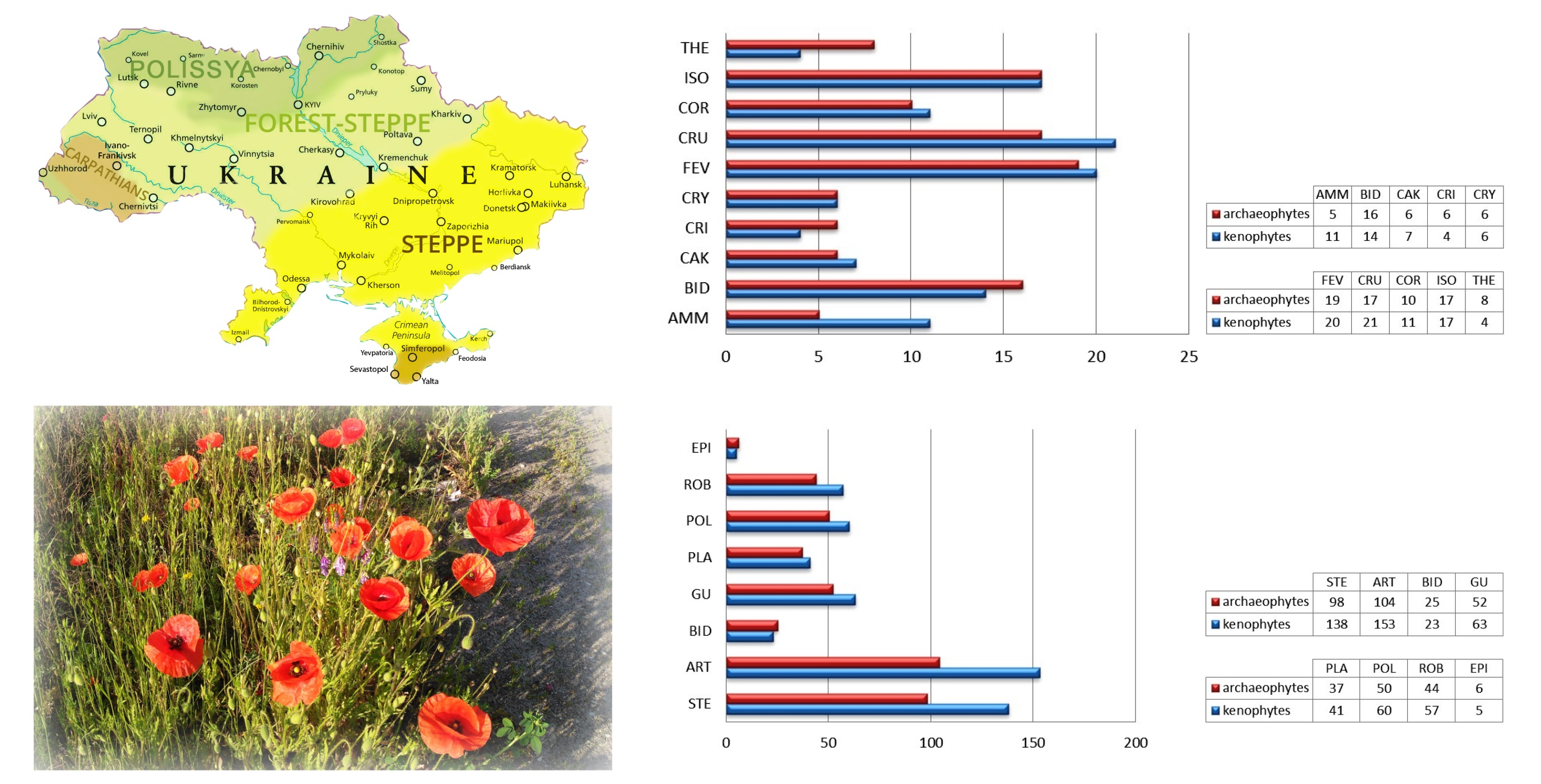{kind=link}
{kind=link}
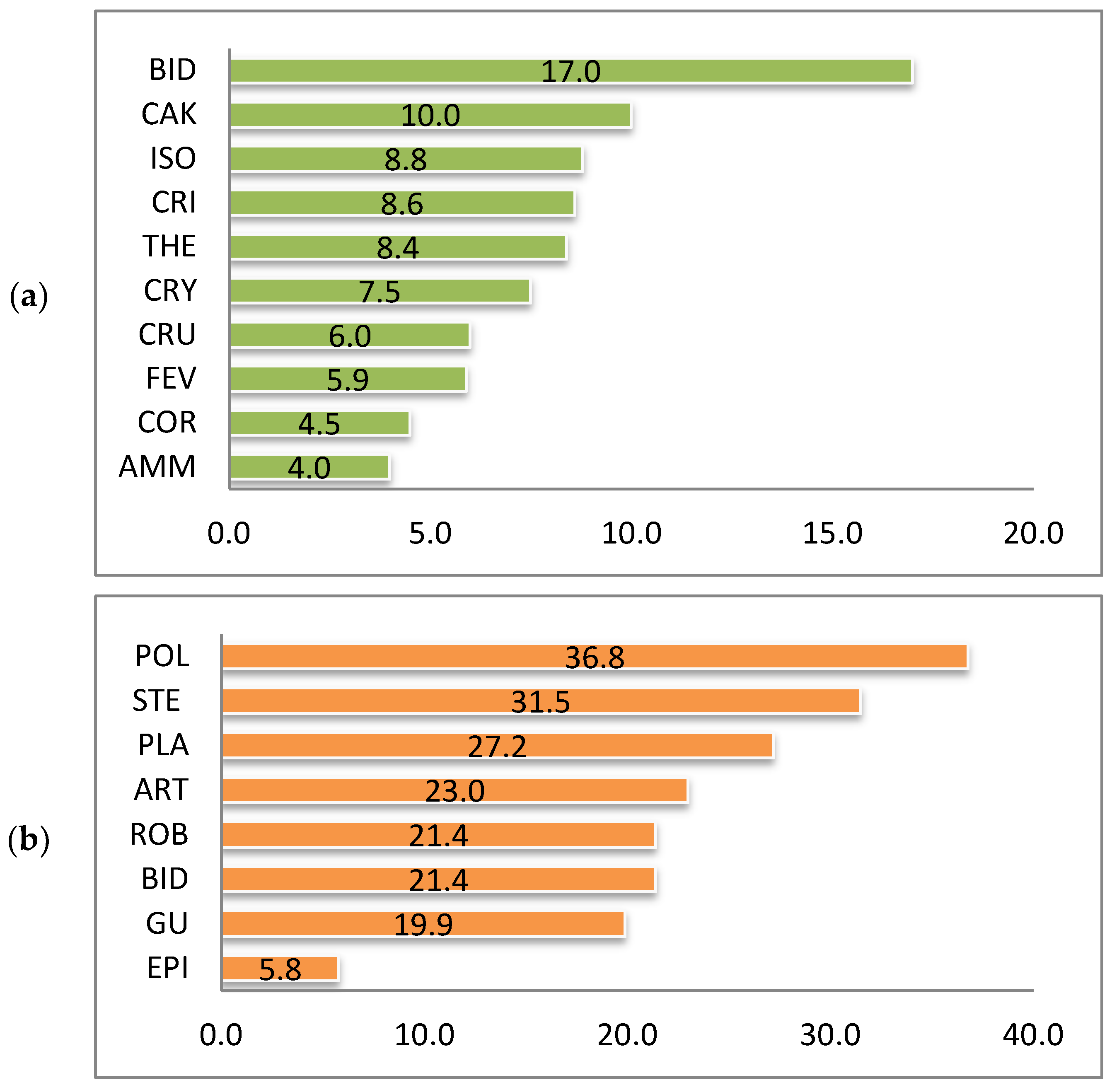{kind=link}
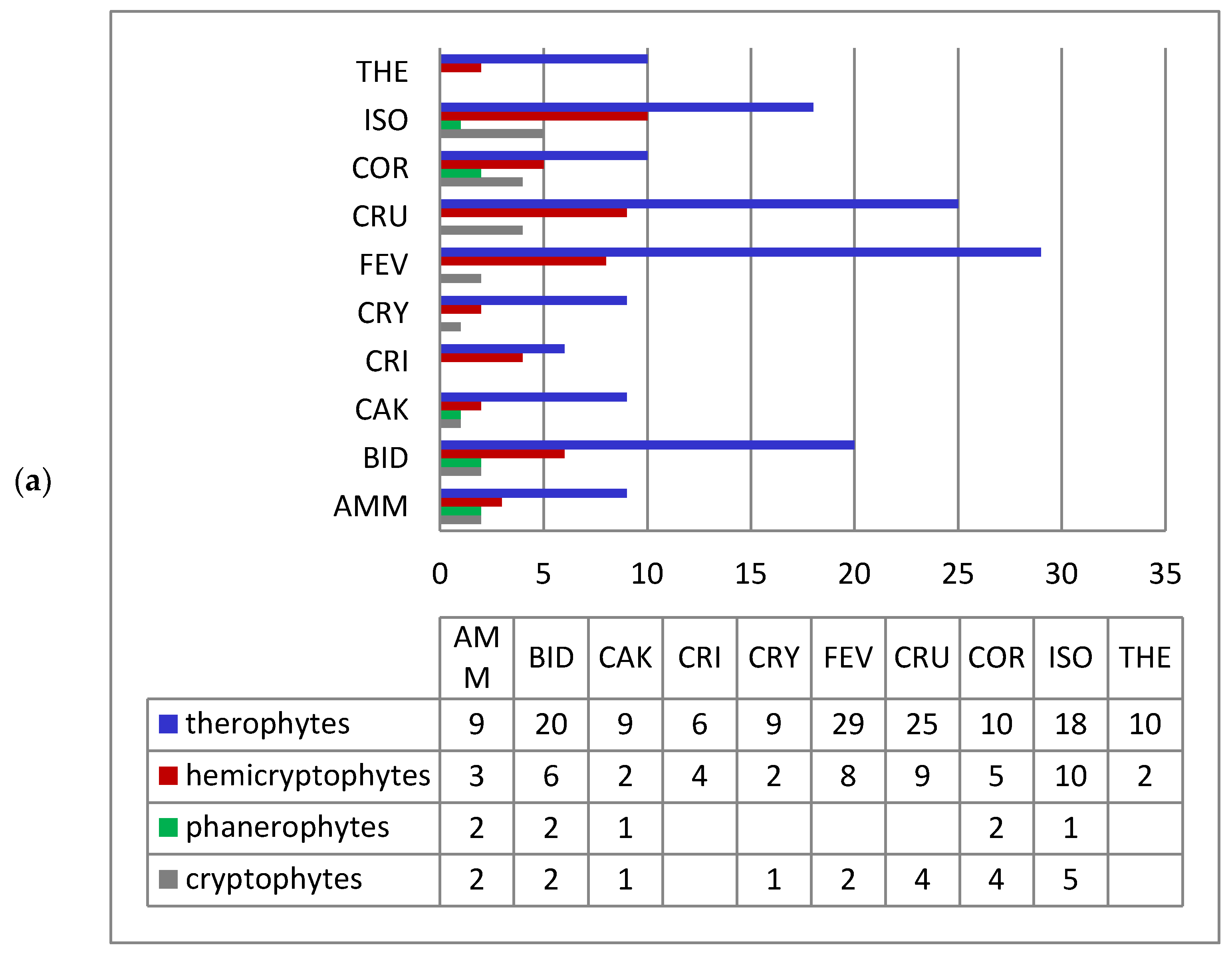{kind=link}
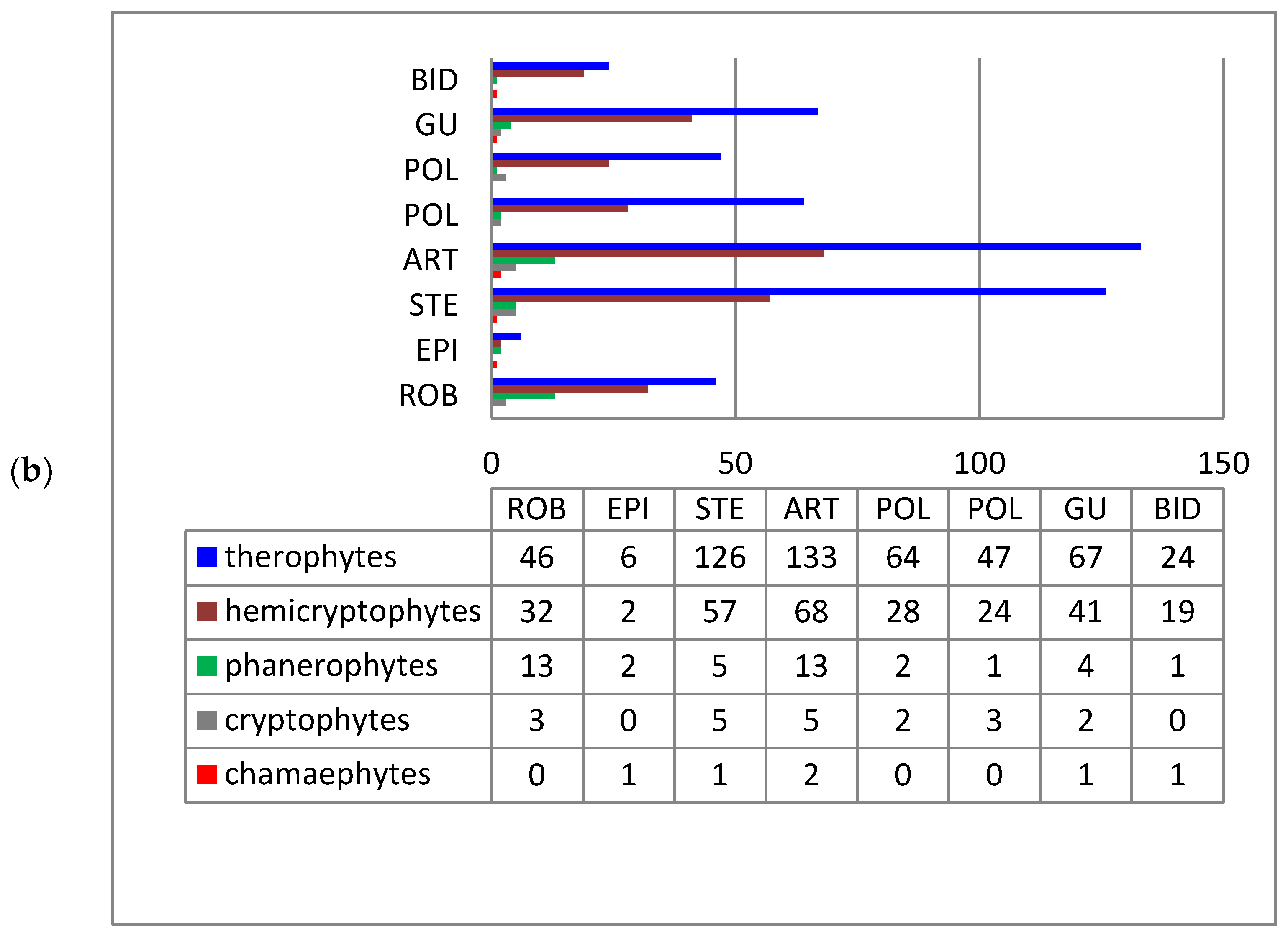{kind=link}
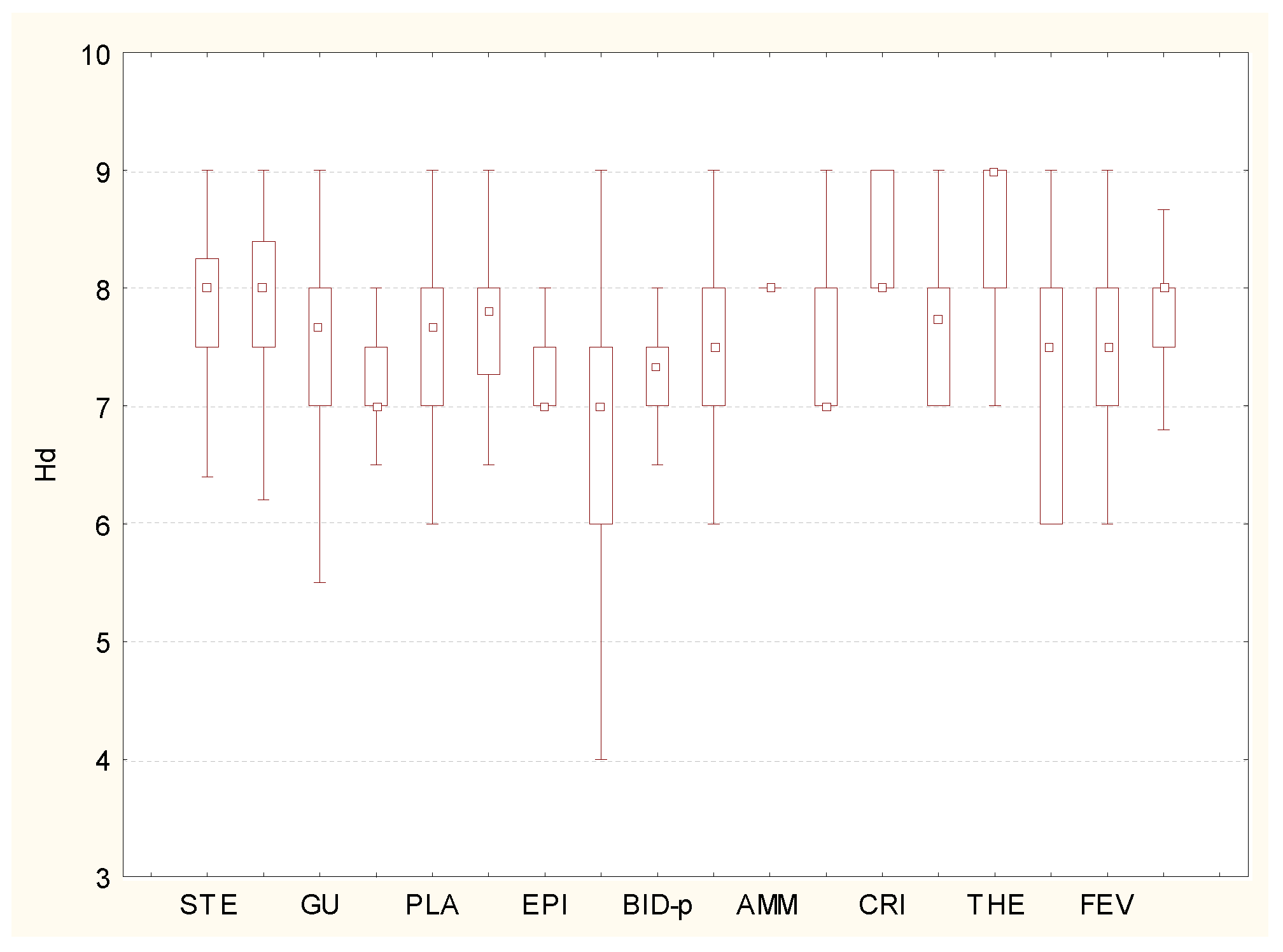{kind=link}
{kind=link}
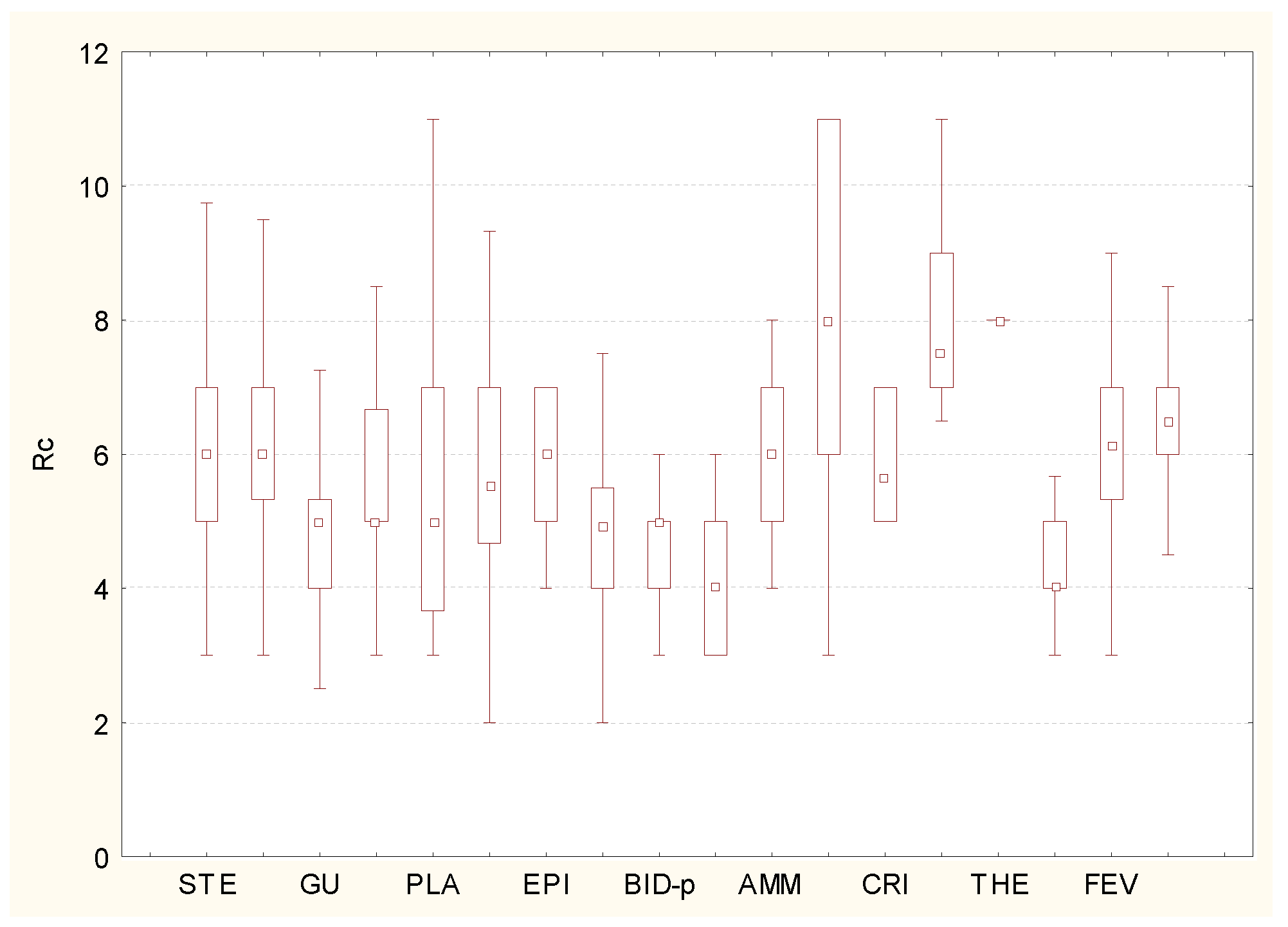{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
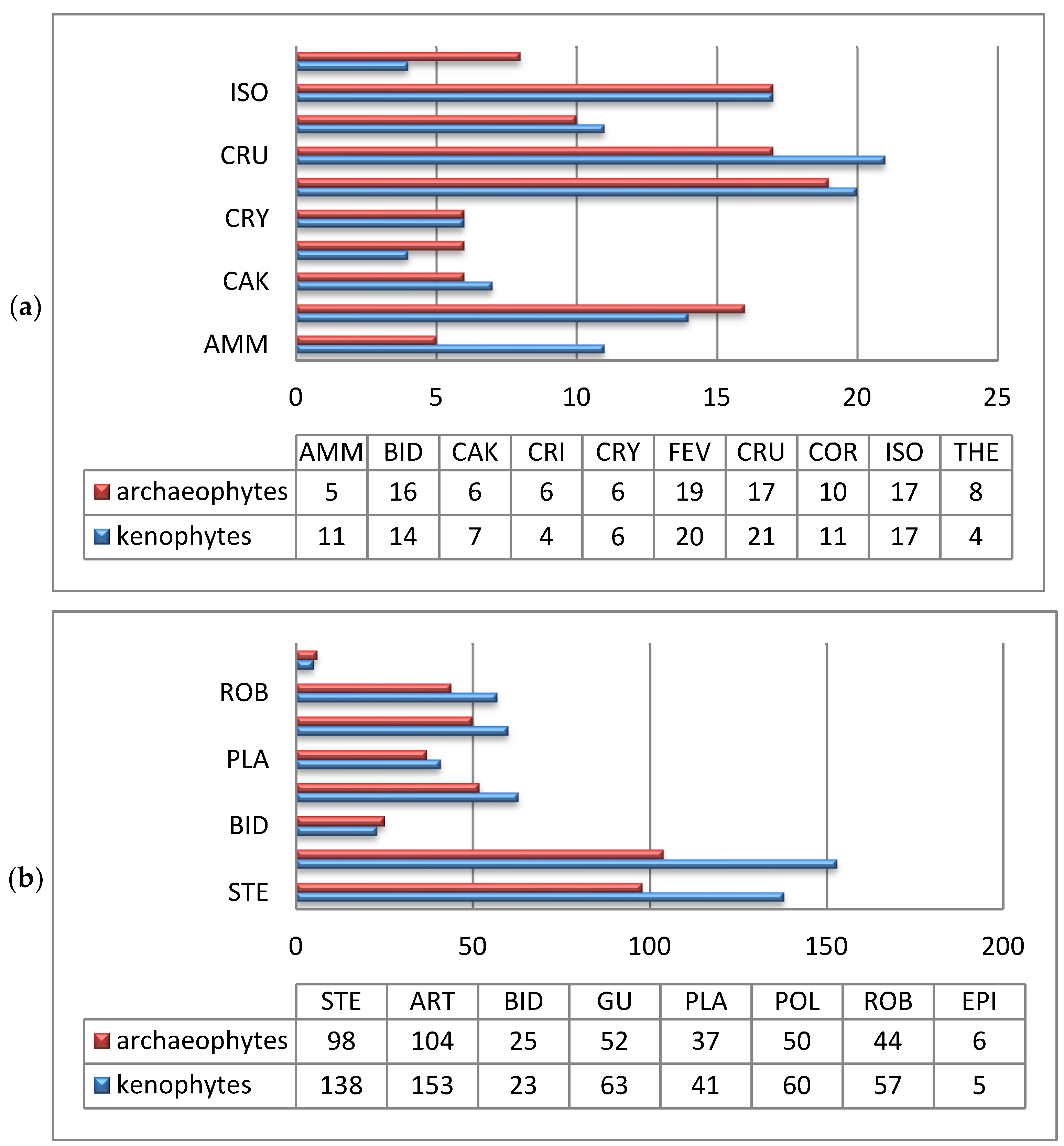{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Spectrum of the leading families of the alien fraction of the coenofloras of the pioneer and ruderal vegetation in Ukraine.
Table 1.
Spectrum of the leading families of the alien fraction of the coenofloras of the pioneer and ruderal vegetation in Ukraine.
| Pioneer Vegetation | Ruderal Vegetation | ||||||
|---|---|---|---|---|---|---|---|
| Rank | Families | Number | Rank | Families | Number | ||
| Species | % | Species | % | ||||
| 1 | Asteraceae | 26 | 23.01 | 1 | Asteraceae | 65 | 20.00 |
| 2 | Brassicaceae | 22 | 19.47 | 2 | Brassicaceae | 38 | 11.69 |
| 3 | Poaceae | 15 | 13.27 | 3 | Poaceae | 29 | 8.92 |
| 4 | Chenopodiaceae | 6 | 5.31 | 4 | Fabaceae | 16 | 4.92 |
| 5 | Fabaceae | 5 | 4.42 | 5 | Chenopodiaceae | 15 | 4.62 |
| 6 | Lamiaceae | 4 | 3.54 | 6 | Apiaceae | 13 | 4.00 |
| 7 | Caryophillaceae | 3 | 2.65 | 7 | Lamiaceae | 10 | 3.08 |
| Cucurbitaceae | 3 | 2.65 | Boraginaceae | 10 | 3.08 | ||
| 8 | Malvaceae | 2 | 1.77 | 8 | Polygonaceae | 9 | 2.77 |
| Onagraceae | 2 | 1.77 | 9 | Amaranthaceae | 8 | 2.46 | |
| Ranunculaceae | 2 | 1.77 | Scrophulariaceae | 8 | 2.46 | ||
| Verbenaceae | 2 | 1.77 | 10 | Malvaceae | 7 | 2.15 | |
Table 2.
Spectrum of the leading genera of the alien fraction of the coenofloras of the pioneer and ruderal vegetation in Ukraine.
Table 2.
Spectrum of the leading genera of the alien fraction of the coenofloras of the pioneer and ruderal vegetation in Ukraine.
| Pioneer Vegetation | Ruderal Vegetation | ||||
|---|---|---|---|---|---|
| Rank | Generas | Number of Species | Rank | Generas | Number of Species |
| 1 | Sisymbrium | 5 | 1 | Xanthium | 9 |
| 2 | Lepidium | 4 | 2 | Amaranthus | 8 |
| 3 | Anisantha | 3 | 3 | Chenopodium | 6 |
| Atriplex | 3 | Geranium | 6 | ||
| 4 | Artemisia | 2 | Sisymbrium | 6 | |
| Bidens | 2 | Veronica | 6 | ||
| Bromus | 2 | 4 | Atriplex | 5 | |
| Camelina | 2 | Vicia | 5 | ||
| Centaurea | 2 | 5 | Anthemis | 4 | |
| Digitaria | 2 | Bromus | 4 | ||
| Eragrostis | 2 | Centaurea | 4 | ||
| Mentha | 2 | Lepidium | 4 | ||
| Oenothera | 2 | Malva | 4 | ||
| Papaver | 2 | 6 | Anchusa | 3 | |
| Senecio | 2 | Carduus | 3 | ||
| Setaria | 2 | Cuscuta | 3 | ||
| Sonchus | 2 | Euphorbia | 3 | ||
| Thlaspi | 2 | Fumaria | 3 | ||
| Verbena | 2 | Helianthus | 3 | ||
| Vicia | 2 | Hordeum | 3 | ||
| Xanthium | 2 | Lamium | 3 | ||
Table 3.
Indices of archaeophytization, kenophytization, modernization, and fluctuation of pioneer (a) and ruderal (b) phytocoenoses of Ukraine.
Table 3.
Indices of archaeophytization, kenophytization, modernization, and fluctuation of pioneer (a) and ruderal (b) phytocoenoses of Ukraine.
| IArch | IKen | IM | IJ | ||
|---|---|---|---|---|---|
| (a) | AMM | 4.0 | 9.0 | 68.0 | 3.0 |
| CAK | 10.0 | 11.7 | 53.8 | – | |
| CRI | 8.6 | 5.7 | 40.0 | – | |
| CRY | 7.5 | 7.5 | 50.0 | 8.3 | |
| THE | 8.4 | 4.2 | 33.3 | – | |
| BID | 17.0 | 15.3 | 46.6 | 4.3 | |
| ISO | 8.8 | 8.8 | 50.0 | 2.1 | |
| CRU | 6.0 | 7.5 | 55.3 | 5.2 | |
| FEV | 5.9 | 6.2 | 51.3 | 1.2 | |
| COR | 4.5 | 4.9 | 52.4 | 0.4 | |
| All | 6.6 | 6.8 | 50.4 | 1.5 | |
| (b) | STE | 13.1 | 18.3 | 58.3 | 5.0 |
| ART | 9.4 | 13.6 | 59.2 | 3.9 | |
| BID | 11.4 | 10.0 | 46.9 | 2.2 | |
| GU | 9.2 | 10.7 | 53.8 | 2.2 | |
| PLA | 12.6 | 14.6 | 53.8 | 2.4 | |
| POL | 16.6 | 20.3 | 55.0 | 3.4 | |
| ROB | 9.1 | 12.3 | 57.4 | 3.4 | |
| EPI | 3.2 | 2.6 | 45.5 | 1.1 | |
| All | 7.3 | 12.6 | 63.4 | 3.8 |
Table 4.
Alien species that are diagnostic for syntaxa of higher and middle ranks.
| Species | Syntaxa |
|---|---|
| Acer negundo (ken.) | Chelidonio-Acerion negundi |
| Acer platanoides (ken.) | Geo-Acerion platanoidis |
| Anisantha tectorum (arch.) | Festucion beckeri |
| Apera spica-venti (arch.) | Scleranthion annui |
| Artemisia absinthium (arch.) | Onopordion acanthii |
| Atriplex prostrata (arch.) | Crypsietea aculeatae, Crypsietalia aculeatae, Cypero-Spergularion salinae, Heleochloion schoenoidis, Crithmo-Staticetea |
| Bassia laniflora (ken.) | Koelerion glaucae |
| Ballota nigra (arch.) | Artemisietea vulgaris, Onopordetalia acanthii, Arction lappae |
| Bidens connata (ken.) | Bidentetea |
| Bidens frondosa (ken.) | |
| Capsella bursa-pastoris (arch.) | Stellarietea mediae |
| Carduus acanthoides (arch.) | Artemisietea vulgaris, Onopordetalia acanthii, Arction lappae |
| Carduus nutans (arch.) | Onopordion acanthii |
| Centaurea diffusa (ken.) | Koelerio-Corynephoretea |
| Conium maculatum (arch.) | Arction lappae |
| Consolida regalis (arch.) | Aperetalia spicae-venti |
| Cyanus segetum (arch.) | Scleranthion annui |
| Descurainia sophia (arch.) | Stellarietea mediae, Sisymbrietalia sophiae, Chenopodio albi-Descurainion sophiae |
| Diplotaxis tenuifolia (ken.) | Medicagini falcatae-Diplotaxion tenuifoliae |
| Erigeron annuus (ken.) | Robinietea, Chelidonio-Robinietalia pseudoacaciae, Chelidonio majoris-Robinion pseudoacaciae |
| Erigeron canadensis (ken.) | Stellarietea mediae |
| Fallopia convolvulus (arch.) | Stellarietea mediae |
| Fumaria officinalis (arch.) | |
| Fumaria schleicheri (arch.) | Chenopodio albi-Descurainion sophiae |
| Heracleum mantegazzianum (ken.) | Galio-Urticetea |
| Impatiens glandulifera (ken.) | |
| Impatiens parviflora (ken.) | |
| Iva xanthiifolia (ken.) | Sisymbrietalia sophiae |
| Lamium album (arch.) | Artemisietea vulgaris |
| Lappula squarrosa (arch.) | Onopordion acanthii |
| Leonurus cardiaca (arch.) | Arction lappae |
| Lepidium campestre (arch.) | Scleranthion annui |
| Lepidium draba (ken.) | Agropyretalia intermedio-repentis, Convolvulo arvensis-Agropyrion repentis |
| Lepidium ruderale (arch.) | Stellarietea mediae, Polygono-Poetea annuae, Polygono-Coronopodion |
| Matricaria chamomilla (arch.) | Anthemido ruthenicae-Sisymbrion orientalis |
| Matricaria discoidea (ken.) | Polygono-Poetea annuae, Polygono-Coronopodion |
| Myosotis arvensis (arch.) | Aperetalia spicae-venti |
| Nigella arvensis (arch.) | Caucalidion |
| Oenothera biennis (ken.) | Koelerio-Corynephoretea |
| Oenothera rubricaulis (ken.) | Koelerion glaucae |
| Onopordum acanthium (arch.) | Onopordetalia acanthi, Onopordion acanthii |
| Oxalis stricta (ken.) | Oxalidion europaeae |
| Papaver rhoeas (arch.) | Papaveretalia rhoeadis, Galeopsion bifidae |
| Portulaca oleracea (arch.) | Eragrostietalia, Eragrostion, Saginion procumbentis |
| Raphanus raphanistrum (arch.) | Panico-Setarion |
| Rapistrum rugosum (ken.) | Sisymbrietalia sophiae |
| Reseda lutea (arch.) | Onopordion acanthii |
| Reynoutria japonica (ken.) | Galio-Urticetea |
| Reynoutria sachalinensis (ken.) | |
| Robinia pseudoacacia (ken.) | Robinietea, Chelidonio-Robinietalia pseudoacaciae, Chelidonio majoris-Robinion pseudoacaciae, Balloto nigrae-Robinion pseudoacaciae, Galio-Urticetea |
| Rubus idaeus (ken.) | Sambucetalia racemosae |
| Rudbeckia laciniata (ken.) | Galio-Urticetea |
| Scleranthus annuus (arch.) | Koelerio-Corynephoretea |
| Sclerochloa dura (arch.) | Polygono-Poetea annuae, Polygono arenastri-Poetalia annuae |
| Senecio viscosus (ken.) | Epilobion augustifolii |
| Senecio vulgaris (arch.) | Veronico-Euphorbion |
| Setaria pumila (arch.) | Atriplici-Chenopodietalia albi, Panico-Setarion |
| Setaria viridis (arch.) | |
| Sinapis arvensis (arch.) | Stellarietea mediae, Papaveretalia rhoeadis, Panico-Setarion |
| Sisymbrium altissimum (ken.) | Sisymbrion officinalis |
| Sisymbrium officinale (arch.) | |
| Sisymbrium orientale (ken.) | Anthemido ruthenicae-Sisymbrion orientalis, Sisymbrion officinalis |
| Sisymbrium loeselii (ken.) | Sisymbrion officinalis |
| Solidago canadensis (ken.) | Galio-Urticetea |
| Sonchus arvensis (arch.) | Stellarietea mediae, Papaveretalia rhoeadis |
| Sonchus asper (arch.) | Stellarietea mediae |
| Sonchus oleraceus (arch.) | |
| Stachys annua (arch.) | Panico-Setarion |
| Thlaspi arvense (arch.) | Stellarietea mediae, Atriplici-Chenopodietalia albi |
| Tribulus terrestris (ken.) | Eragrostietalia, Saginion procumbentis |
| Tripleurospermum inodorum (arch.) | Stellarietea mediae, Cakiletea maritimae, Aperetalia spicae-venti, Verbenion supinae, Polygono-Chenopodion, Sisymbrion officinalis |
| Urtica urens (arch.) | Malvion neglectae |
| Verbena supina (ken.) | Isoëto-Nanojuncetea |
| Veronica arvensis (arch.) | Koelerio-Corynephoretea |
| Veronica persica (ken.) | Veronico-Euphorbion |
| Vicia pannonica (arch.) | Caucalidion |
| Vicia villosa (arch.) | Anthemido ruthenicae-Sisymbrion orientalis |
| Viola arvensis (arch.) | Aperetalia spicae-venti |
| Xanthium orientale subsp. riparium (ken.) | Bidentetea, Bidentetalia, Chenopodion rubri |
| Xanthium pungens (ken.) | Sisymbrion officinalis |
| Xanthium strumarium (arch.) | Bidentetea, Bidentetalia, Chenopodion rubri, Thero-Atriplicetalia, Sisymbrietalia sophiae, Sisymbrion officinalis, Amarantho blitoidis-Echinochloion cruris-galli |
Explanations: arch.—archaeophyte, ken.—kenophyte.
Table 5.
Distribution of alien species in the syntaxa of pioneer and ruderal vegetation as diagnostic, constant, and dominant.
Table 5.
Distribution of alien species in the syntaxa of pioneer and ruderal vegetation as diagnostic, constant, and dominant.
| Syntaxon of Classes | Diagnostic | Constant | Dominant |
|---|---|---|---|
| Stellarietea mediae | Adonis aestivalis, Amaranthus albus, A. blitoides, A. retroflexus, Ambrosia artemisiifolia, Anagallis arvensis, Anisantha sterilis, A. tectorum, Apera spica-venti, Artemisia annua, Atriplex prostrata, A. sagittata, A. tatarica, Bassia scoparia, Bromus squarrosus, Camelina microcarpa, Capsella bursa-pastoris, Lepidium draba, Carduus acanthoides, Centaurea diffusa, Chenopodiastrum hybridum, Conium maculatum, Consolida regalis, Crepis micrantha, Dasypyrum villosum, Descurainia sophia, Digitaria sanguinalis, D. ischaemum, Diplotaxis muralis, Echinochloa crus-galli, Eragrostis minor, Erigeron canadensis, Erysimum repandum, Euphorbia helioscopia, E. peplus, Fallopia convolvulus, Galinsoga parviflora, Hibiscus trionum, Hordeum murinum, H. murinum subsp. leporinum, Hyoscyamus niger, Iva xanthiifolia, Lactuca serriola, Lamium amplexicaule, L. purpureum, Lappula squarrosa, Lepidium campestre, L. perfoliatum, Malva neglecta, Matricaria chamomilla, Myosotis arvensis, Neslia paniculata, Oenothera biennis, Oxalis dillenii, O. stricta, Oxybasis rubra, Papaver rhoeas, P. dubium, Peganum harmala, Portulaca oleracea, Raphanus raphanistrum, Rapistrum rugosum, Reseda lutea, R. luteola, Scleranthus annuus, Senecio vulgaris, Setaria pumila, S. viridis, Sinapis arvensis, Sisymbrium altissimum, S. loeselii, S. officinale, S. orientale, Solanum nigrum, Sonchus arvensis, S. asper, S. oleraceus, Spergula arvensis, Stachys annua, Thlaspi arvense, Noccaea perfoliata, Tribulus terrestris, Tripleurospermum inodorum, Urtica urens, Veronica persica, Vicia sativa subsp. nigra, V. tetrasperma, V. pannonica, Viola arvensis, Xanthium strumarium, X. orientale subsp. riparium, X. spinosum | Amaranthus retroflexus, Ambrosia artemisiifolia, Anisantha sterilis, A. tectorum, Apera spica-venti, Artemisia absinthium, Atriplex prostrata, Ballota nigra, Capsella bursa-pastoris, Lepidium draba, Carduus acanthoides, Centaurea diffusa, Cichorium intybus, Consolida regalis, Cyanus segetum, Descurainia sophia, Diplotaxis muralis, Echinochloa crus-galli, Eragrostis minor, Erigeron canadensis, Erysimum repandum, Fallopia convolvulus, Hordeum murinum, Iva xanthiifolia, Lactuca serriola, Lepidium perfoliatum, L. ruderale, Oenothera biennis, Onopordum tauricum, Papaver dubium, P. rhoeas, Portulaca oleracea, Setaria pumila, S. viridis, Sisymbrium loeselii, S. officinale, Sonchus arvensis, S. oleraceus | Amaranthus albus, A. retroflexus, Ambrosia artemisiifolia, Anisantha sterilis, A. tectorum, Artemisia annua, Atriplex prostrata, A. tatarica, Bromus squarrosus, Erigeron canadensis, Descurainia sophia, Digitaria sanguinalis, D. ischaemum, Echinochloa crus-galli, Eragrostis minor, Fallopia convolvulus, Galinsoga parviflora, Geranium pusillum, Hordeum murinum, Iva xanthiifolia, Bassia scoparia, Lamium purpureum, Lepidium draba, L. perfoliatum, L. ruderale, Malva neglecta, M. pusilla, Peganum harmala, Portulaca oleracea, Reseda lutea, Scleranthus annuus, Setaria viridis, S. pumila, Sisymbrium altissimum, S. officinale, S. loeselii, Sonchus oleraceus, Tribulus terrestris |
| Artemisietea vulgaris | Anchusa officinalis, Anisantha tectorum, Artemisia absinthium, Asclepias syriaca, Atriplex prostrata, A. sagittata, A. tatarica, Ballota nigra, Bupleurum rotundifolium, Carduus acanthoides, C. nutans, Centaurea diffusa, Conium maculatum, Erigeron canadensis, Crepis micrantha, Diplotaxis tenuifolia, D. muralis, Ecballium elaterium, Galium spurium, Grindelia squarrosa, Lamium album, L. purpureum, Lathyrus tuberosus, Leonurus cardiaca, Lepidium draba, L. graminifolium, Lycium barbarum, Malva pusilla, M. sylvestris, Onopordum acanthium, Papaver rhoeas, Prunus cerasifera, Ranunculus arvensis, Reseda lutea, Sinapis arvensis, Solanum nigrum, Sonchus oleraceus, Verbena officinalis, Veronica arvensis, V. triphyllos, Vulpia ciliata, Xanthium spinosum, X. strumarium | Amaranthus retroflexus, Ambrosia artemisiifolia, Anisantha tectorum, Apera spica-venti, Artemisia absinthium, Atriplex prostrata, Ballota nigra, Bromus squarrosus, Capsella bursa-pastoris, Lepidium draba, Carduus acanthoides, Centaurea diffusa, Cichorium intybus, Conium maculatum, Consolida regalis, Erigeron canadensis, Crepis micrantha, Descurainia sophia, Diplotaxis tenuifolia, Erigeron annuus, Grindelia squarrosa, Lactuca serriola, Lamium purpureum, Leonurus cardiaca, Onopordum acanthium, Sisymbrium loeselii | Ambrosia artemisiifolia, Anisantha tectorum, Artemisia absinthium, Asclepias syriaca, Atriplex sagittata, Ballota nigra, Bromus squarrosus, Lepidium draba, Carduus acanthoides, Conium maculatum, Erigeron canadensis, Grindelia squarrosa, Lathyrus tuberosus, Leonurus cardiaca, Lepidium perfoliatum, Lycium barbarum, Onopordum acanthium, Rhamnus alaternus, Xanthium orientale subsp. riparium, X. strumarium, X. spinosum |
| Galio-Urticetea | Asclepias syriaca, Calepina irregularis, Chrozophora tinctoria, Echinocystis lobata, Erysimum cheiranthoides, Geranium rotundifolium, Helianthus tuberosus, Heracleum mantegazzianum, H. pubescens, Impatiens glandulifera, I. parviflora, Lamium album, Lepidium graminifolium, Medicago arabica, Ranunculus arvensis, Reynoutria japonica, R. sachalinensis, Robinia pseudoacacia, Rudbeckia laciniata, Solidago canadensis, S. gigantea, Tripleurospermum inodorum, Verbena officinalis | Ballota nigra, Erigeron annuus | Bryonia alba, Helianthus tuberosus, Heracleum mantegazzianum, H. pubescens, Impatiens glandulifera, I. parviflora, Solidago canadensis |
| Bidentetea (ruderal) | Atriplex prostrata, Bidens connata, B. frondosa, Echinochloa crus-galli, Xanthium orientale subsp. riparium, X. strumarium | Bidens connata, B. frondosa | Bidens connata, B. frondosa |
| Robinietea | Acer negundo, A. platanoides, Ailanthus altissima, Amorpha fruticosa, Ballota nigra, Caragana arborescens, Celtis occidentalis, Cotinus coggygria, Erigeron annuus, E. canadensis, Fraxinus pennsylvanica, Galinsoga parviflora, Gleditsia triacanthos, Impatiens parviflora, Lactuca serriola, Parthenocissus quinquefolia, Quercus rubra, Robinia pseudoacacia, Rubus idaeus, Solanum nigrum | Acer negundo, A. platanoides, Ballota nigra, Robinia pseudoacacia | Acer negundo, A. platanoides, Ailanthus altissima, Anisantha sterilis, A. tectorum, Ballota nigra, Celtis occidentalis, Cotinus coggygria, Gleditsia triacanthos, Impatiens parviflora, Parthenocissus quinquefolia, Robinia pseudoacacia |
| Epilobietea angustifolii | Erigeron canadensis, Oenothera biennis, Rubus idaeus, Senecio viscosus | Rubus idaeus | |
| Polygono-Poetea annuae | Anagallis arvensis, Erigeron canadensis, Eragrostis minor, Lappula squarrosa, Lepidium ruderale, Portulaca oleracea, Sclerochloa dura, Sisymbrium orientale, Tragus racemosus, Tribulus terrestris | Capsella bursa-pastoris | Eragrostis minor, Sclerochloa dura |
| Plantaginetea majoris | Juncus tenuis, Lepidium ruderale, Mentha pulegium, Ranunculus arvensis, Verbena officinalis | Erigeron canadensis, Juncus tenuis | Juncus tenuis |
| Cakiletea maritimae | Atriplex prostrata, Tripleurospermum inodorum, Xanthium strumarium s. l. | Atriplex prostrata | – |
| Festucetea vaginatae | Anisantha tectorum, Bromus squarrosus, Centaurea diffusa, Tragus racemosus, Tribulus terrestris | Ambrosia artemisiifolia, Anisantha sterilis, A. tectorum, Apera spica-venti, Bromus squarrosus, Camelina rumelica, Centaurea diffusa, Cichorium intybus, Descurainia sophia, Eragrostis minor, Erysimum repandum | Anisantha sterilis, Apera spica-venti |
| Crithmo-Staticetea | Atriplex prostrata | Anisantha tectorum, Atriplex prostrata | – |
| Crypsietea aculeatae | Atriplex prostrata | Atriplex tatarica | – |
| Therosalicornietea | – | Atriplex tatarica, A. prostrata, Bromus squarrosus | – |
| Helichryso-Crucianelletea maritimae | Anisantha tectorum | Anisantha tectorum, Bromus squarrosus, Carduus acanthoides, Bassia laniflora | – |
| Koelerio-Corynephoretea canescentis | Centaurea diffusa, Bassia laniflora, Oenothera biennis, O. rubricaulis, Veronica arvensis | Artemisia absinthium, Erigeron annuus | – |
| Isoëto-Nanojuncetea | Amaranthus retroflexus, Digitaria ischaemum, Mentha pulegium, Tripleurospermum inodorum, Verbena supina | Oxybasis rubra | – |
| Bidentetea (pion.) | Atriplex prostrata, Bidens connata, B.frondosa, Oxybasis glauca, Xanthium strumarium | Bidens connata, B. frondosa | Bidens connata, B. frondosa, Oxybasis glauca |
| Ammophiletea | – | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dubyna, D.V.; Dziuba, T.P.; Iemelianova, S.M.; Protopopova, V.V.; Shevera, M.V. Alien Species in the Pioneer and Ruderal Vegetation of Ukraine. Diversity 2022, 14, 1085. https://doi.org/10.3390/d14121085
AMA Style
Dubyna DV, Dziuba TP, Iemelianova SM, Protopopova VV, Shevera MV. Alien Species in the Pioneer and Ruderal Vegetation of Ukraine. Diversity. 2022; 14(12):1085. https://doi.org/10.3390/d14121085
Chicago/Turabian StyleDubyna, Dmytro V., Tetiana P. Dziuba, Svitlana M. Iemelianova, Vira V. Protopopova, and Myroslav V. Shevera. 2022. "Alien Species in the Pioneer and Ruderal Vegetation of Ukraine" Diversity 14, no. 12: 1085. https://doi.org/10.3390/d14121085
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.