
Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China
by

Chuli Liu
1,
Hailong Lu
1,†,
Hongjin Chen
1,
Shuikang Chen
1,
Caixia Wang
1,
Zhiyong Zhou
2 and
Rong Ma
1,* 1
College of Forestry and Landscape Architecture, Xinjiang Agricultural University, Urumqi 830052, China
2
Xiaokelan Management and Conservation Center, Altay Branch of State Forestry Administration, Altay 836599, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Diversity 2023, 15(12), 1190; https://doi.org/10.3390/d15121190
Submission received: 23 October 2023
/
Revised: 28 November 2023
/
Accepted: 29 November 2023
/
Published: 30 November 2023
(This article belongs to the Special Issue Recent Advances in Plant-Pathogen Interactions - 2nd Edition)
Abstract
:Populus laurifolia is one of the most valuable tree species in the world and an important silvicultural tree species in the Xinjiang Uygur Autonomous Region, China. In July 2017, an unreported brown leaf spot disease was observed on P. laurifolia in Altay City, Xinjiang. The causal agent of this leaf spot disease was isolated, and Koch’s postulates were performed to confirm its pathogenicity. Based on a morphological characterization and phylogenetic analyses, the causal organism was identified to be a fungal species, Coniolariella gamsii. The optimum mycelial growth conditions for C. gamsii are on PLPDA (Populus leaves potato dextrose agar) medium, at 28 °C, in the dark. The sporulation time when using PLPDA medium (12 days) is much less than that for PDA medium (25 days). Pathogenicity tests revealed that C. gamsii can also infect two other Populus species (P. bolleana and P. tomentosa). This is the first report of C. gamsii causing brown leaf spot disease on P. laurifolia, and the optimum culture and sporulation conditions have also been optimized for the first time. This study provides a theoretical basis for the diagnosis of this disease and the monitoring of the disease’s occurrence and epidemic status.
1. Introduction
Laurel poplar (Populus laurifolia) is native to Kazakhstan, the Mongolian Altai Mountains, and the Xinjiang Uygur Autonomous Region, China [1]. This species hybridizes readily with other Populus species and has high genetic variation [2]. Populus laurifolia is mainly distributed in the Altay and Tacheng areas of Xinjiang and plays an important environmental role, increasing the soil water availability and decreasing flood events [2]. Wood from P. laurifolia trees has traditionally been used for handicrafts and as a construction material for buckets, saddle frames, and the center ring of yurts [1,2].
In July 2017, a previously unreported leaf disease that formed circular, necrotic areas on P. laurifolia and developed into black circular or irregular leaf spots was found in Xiaodonggou Forest Park, Altay City. During severe infections, the black circular spots frequently coalesced, causing the entire leaf to wither and die.
Coniolariella belongs to Ascomycota, Sordariomycetes, Xylariales, Xylariaceae. This genus was recently segregated from Coniochaeta (Coniochaetales) and was introduced due to its affinities to Xylariales; it replaced the previously invalidly published name Coniolaria Seigle-Mur. in 2006 [3,4,5]. The Coniolariella includes five species that have been recorded to cause leaf diseases in Eryngium campestre, Hordeum vulgare, Vitis vinifera, and Pistacia chinensis [4,5]. Prior to the present study, the only reported fungal pathogen on the leaves of P. laurifolia was the rust fugus Melampsora allii-populina [6,7]. Species of Coniolariella can be characterized as having asci and lacking visible apical structures and polyblastic conidiogenesis [5]. Recently, morphological and phylogenetic analyses using the internal transcribed spacer (ITS) and the large subunit (LSU) regions revealed five species in this genus, namely, C. ershadii, C. gamsii, C. hispanica, C. limoniispora, and C. macrothecia [3,4,5].
The objective of this study was to identify the pathogen-causing disease on P. laurifolia in the Xinjiang Uygur Autonomous Region based on morphological identification and molecular analyses. Koch’s postulates were performed to confirm the pathogenicity of the isolated fungus on P. laurifolia leaves. In order to improve sporulation efficiency, we optimized the formula of the medium used for this fungal species. This study will provide a scientific diagnosis of the leaf spots found on P. laurifolia.
2. Materials and Methods
2.1. Sample Survey, Collection, and Fungal Isolation
Surveys of the disease incidence were conducted in Xiaodonggou Forest Park Altay City, Xinjiang Uygur Autonomous Region during the period of 2017–2019. The results indicated that the incidence rates for 2017, 2018, and 2019 were 51.4%, 62.5%, and 79.2%, respectively. During the surveys, a total of 30 P. laurifolia leaf samples with spot symptoms were collected and kept in paper bags for further study.
The leaf samples were first rinsed under running tap water for 30 s and immersed in 75% ethanol for 30 s. Tissue pieces (5 mm2) cut at the junctions of diseased and healthy tissue were disinfected by being placed in 3% sodium hypochlorite (NaClO) solution for 3 min. After rinsing three times in sterile distilled water and drying them naturally on sterilized filter paper [8], these tissue pieces were incubated on potato dextrose agar (PDA) at 28 °C for 3 to 4 d in 12/12 h light/darkness. The growing edges of distinct fungal colonies were subcultured onto fresh PDA, and pure cultures were obtained by subculturing hyphal tips. After obtaining axenic cultures, single-spore cultures were cultured on fresh PDA. Isolates were maintained as a spore suspension in 25% glycerol at −80 °C until they were ready for use in further studies [9,10,11,12]. Living cultures were deposited in the China Forestry Culture Collection Center (CFCC) and the Pathology Laboratory of the Forestry and Horticulture College (XJAU).
2.2. DNA Extraction, PCR Amplification, and Sequencing
Total genomic DNA was extracted from 5-day-old mycelium grown on PDA at 25 °C in the light using the CTAB method. DNA sequences were generated for the internal transcribed spacer (ITS) region, which was amplified using primers ITS1/ITS4 [13], and the large subunit (LSU) of the nuclear rDNA region, which was amplified using primers NL1/NL4 [14]. PCR reactions were performed in a 30 µL reaction volume containing 1.5 µL of DNA template, 1.5 µL of each forward and reverse primer, 15 µL of PCR master mix, and 10.5 µL of double-distilled water (dd H2O). The following PCR conditions were used: an initial denaturation step of 2 min at 94 °C (ITS) or at 95 °C (LSU), followed by 35 cycles of 30 s at 94 °C (ITS) or 25 s at 95 °C (LSU), 45 s primer annealing at 51 °C (ITS) or 55 °C (LSU), 1 min extension at 72 °C, and a final 7 min extension at 72 °C (ITS) or a 10 min extension at 72 °C (LSU) [15]. PCR products were detected using a 1% agarose gel under 120 V stable voltage, and the electrophoresis time was 25 min. PCR bands were observed in a gel imager to determine whether they were clear and to determine the size of the target bands. PCR products were sent to Shanghai Sangon Bioengineering Co., Ltd. (Shanghai, China) for bidirectional sequencing.
2.3. Phylogenetic Analyses
The quality of the obtained chromatograms was checked, and the nucleotide sequences were assembled using SeqMan v.7.1.0 and the DNASTAR lasergene core suite software (DNASTAR Inc., Madison, WI, USA). BLAST searches were performed to identify homologous sequences in GenBank (http://blast.ncbi.nlm.nih.gov; accessed on 21 October 2023). Reference sequences and newly generated nucleotide sequences are listed in Table 1. Sequences were aligned using MAFFT v. 6 and corrected manually using MEGA 7.0.21 [16,17].
The phylogenetic analyses of the combined loci were performed using the Maximum Likelihood (ML) and Bayesian Inference (BI) methods. The ML was implemented on the CIPRES Science Gateway portal using RAxML-HPC BlackBox 8.2.10 [18,19], employing a GTRGAMMA substitution model with 1000 bootstrap replicates. The Bayesian Posterior Probabilities (BPPs) were determined with Markov Chain Monte Carlo (MCMC) sampling in MrBayes v. 3.2.6 [20]. The phylogram was visualized in FigTree v.1.3.1. The newly generated nucleotide sequences were deposited in GenBank (Table 1).
2.4. Pathogenicity Test
The CFCC 55706 strain was used for pathogenicity testing on detached leaves of Populus bolleana Lauche and P. tomentosa Carr. in August 2020 and the leaves of 2-year-old seedlings of P. laurifolia from Altay City in August 2021.
The detached leaves and seedling leaves were washed under running tap water to remove surface dust, surface-washed with sterile distilled water three times, and dried naturally. The leaves were pierced with a No. 3 insect needle to create a single wound of uniform size on the left- and right-hand side of each leaf. Mycelial discs (5 mm in diameter) taken from the growing edge of 3-day-old cultures grown on PDA and incubated at 25 °C were transferred to the right-hand wound of each leaf, with the mycelium facing the exposed wound. The left-hand wound on each leaf was mock-inoculated with a sterile PDA disc, which acted as a negative control. There were five leaf replicates for each poplar species and five leaves per seedling. The detached leaves were placed on a tray with holes in it. Distilled water was placed in the tray holes to maintain humid conditions. The inoculum was removed from the wound after 24 h. The development of leaf lesions around the inoculated wound area was monitored, and lesion diameters were measured every two days. Inoculation experiments were carried out in a greenhouse maintained at 25 °C. Fungal isolates were reisolated from the lesion margins of the inoculated leaf tissue and identified again based on morphological characteristics and DNA sequencing.
2.5. Determination of Colony Morphologies and Growth Rates
Mycelial plugs (5 mm in diameter) excised from the periphery of 3-day-old colonies were used to inoculate the centers of fresh PDA plates. The plates were incubated at 25 °C under light conditions. The colony diameter was measured using the crossing method at the same time each day. The daily growth rate was used to calculate the 5-day average of mean daily growth. Morphological observations of the anamorph features included the stromata color, growth rate, developing colony color, mature colony color, presence and color of exudate droplets, the reverse color of mature colonies, and the morphology of colony margins.
2.6. Analysis of Culture Characteristics
To investigate the effect of temperature on the mycelial growth of strain CFCC 55706, mycelial agar discs measuring 5 mm in diameter were removed from the edge of 3-day-old colonies using a sterilized punch. The discs were transferred to fresh plates of PDA medium (90 mm in diameter) and cultured at 0 °C, 4 °C, 8 °C, 12 °C, 16 °C, 20 °C, 24 °C, 28 °C, 32 °C, 36 °C, 37 °C, 38 °C, or 40 °C in darkness. Each treatment was repeated five times. Colony diameters were measured using the straight-edge crisscrossing method at 24 h, 48 h, 72 h, 96 h, 120 h, and 144 h post-inoculation [21]. The mean average of five replicates was calculated for each temperature treatment.
We also analyzed the effect of different illumination conditions at 24 °C on the mycelial growth of strain CFCC 55706. Three light conditions were investigated: 24 h of light, 24 h of darkness, and 12/12 h light/darkness. Inoculation, culture, and colony measurements were performed as in the method described above.
To determine whether mycelial growth and sporulation time could be enhanced using different media, strain CFCC 55706 was cultured on three different types of modified PDA media: Populus leaf potato dextrose agar (PLPDA), Salix leaf potato dextrose agar (SLPDA), and Malus leaf potato dextrose agar (MLPDA). To select the most suitable formulation, 5 g, 10 g, 15 g, or 20 g of Populus bolleana, Salix babylonica, or Malus pumila leaves was added to PDA medium and boiled for 5 min, 10 min, 15 min, or 20 min. The cross treatment of leaf grams and treatment time was added, and 16 treatments were performed in each group. Colony growth on the different media was measured after incubation at 28 °C in the dark for 7 d. The colonies were observed for a further month to determine the sporulation time.
2.7. Statistical Analysis
The means and standard errors of the data were calculated using Microsoft Excel formulas. Analysis of variance was used to detect differences among treatments using IBM SPSS Statistics 22.0 and Origin 9.5.
3. Results
3.1. Incidence and Fungal Isolation
In Xinjiang, diseased Populus laurifolia were found and collected from Xiaodonggou Forest Park in 2017, 2018 and 2019. Symptomatic leaf samples at different stages of P. laurifolia leaf brown spot disease development were also collected. In total, 25 fungal strains were isolated from diseased poplar samples. These isolates had the same colony morphology and the same sequences of ITS and LSU.
3.2. Phylogenetic Analyses
The combined sequence dataset of ITS and LSU was analyzed to infer the phylogenetic placement of our new isolates within the genus Coniolariella. The dataset consisted of nine sequences, including one outgroup taxon, Xylaria badia (BCC 1190). A total of 1075 characters, including gaps (565 for ITS and 510 for LSU), were included in the phylogenetic analysis. Of these characters, 991 were constant, 83 were variable but parsimony-uninformative, and 1 was parsimony-informative. The best ML tree (lnL = −800.19) revealed by RAxML is shown in Figure 1. The topologies resulting from the ML and BI analyses of the concatenated dataset were congruent (Figure 1). Five species of Coniolariella were classified into two groups. The first group included C. gamsii (CBS 114319), C. ershadii (CBS 119785), C. limoniispora (AH 24323 and 108fa), C. macrothecia (CBS 125772), and two new isolates from the present study (CFCC 55706 and XJAU KY-2-1). The second group only included the species C. hispanica (CBS 124506). The phylogeny based on the combined loci of ITS and LSU cannot distinguish C. ershadii, C. gamsii, C. limoniispora, and C. macrothecia. Hence, it is impossible to identify our new isolates at the species level based on the phylogenetic analyses of ITS and LSU loci (Figure 1).
3.3. Description of Coniolariella gamsii
Coniolariella gamsii (Asgari & Zare) Dania García, Stchigel & Guarro, Mycol. Res. 110(11): 1285 (2006).
The sexual state was observed, and the ascomata were either superficial or buried at the base; were globose, solitary or in small groups; had a rough surface with no bristles; were dark brown to black and 200 to 700 μm in diameter (Figure 2D); and had white projections protruding from the top, as well as slit-like ostioles (Figure 2E). The shell of the ascomata consists of thick-walled polygonal cells in the outer layer and subglobular or angular cells in the inner layer (Figure 2G,H). The asci are cylindrical, tapering slightly to the base, eight-spored, apical without an apical ring, and 83 to 125 μm × 11 to 16 μm (Figure 2I,J). The ascospores are arranged in the asci in a single row, with a straight germ slit, and a protrusion at one or both ends. Initially, they are colorless to olive green, becoming dark-chocolate brown or black and elliptical, and are 30 to 40 × 10 to 20 μm (Figure 2L). Immature ascospores with a hole or a few pores in water are shown in Figure 2K. Paraphyses are numerous, filiform, septate, simple, slightly undulate, and considerably longer than the asci (Figure 2M). Octahedral crystals were observed in the culture (Figure 2O).
Conidiophores were produced laterally or terminally on the hyphae; were 6 to 45 × 2 to 3 μm; erect or ascending; mostly simple, smooth, and septate; had inconspicuous denticles bearing solitary blastoconidia; and had a distinct scar. The conidia were holoblastic, hyaline, smooth, oblong, and cylindrical with a rounded apex and an attenuated and truncated base 5 to 12 × 2 to 3 μm in size (Figure 2N).
The colonies were white, pinnate, and sparse (Figure 2A) at 3 d, turning gray after 10 d on the PDA medium. The mycelial growth rate was 11 mm/d. The ascomata emerged after 12 d and ascospores were observed at 28 d (Figure 2B,C). The ascomata appeared to be white during the early stages, later turning black, with a globose and randomly distributed colony on the PDA medium surface. The conidia were observed after 15 d.
Materials examined: Xinjiang Uygur Autonomous Region, Altay City, 47°56′27.41″ N, 88°08′03.48″ E, alt. 1058 m, on leaf of P. laurifolia, 7 July 2017, R. Ma (BJFC-S1875, cultures CFCC 55706 and XJAU KY-2-1).
Notes: The species Coniolariella gamsii was initially proposed in the genus Coniochaeta as Coniochaeta gamsii based on two collections from the host Hordeum vulgare in Iran [22]. The genus Coniolariella was later separated from Coniochaeta (Coniochaetales) by transferring the species affinities to Xylariales based on molecular phylogeny [7]. Morphologically, specimens collected from P. laurifolia in the present study matched with the type specimen of C. gamsii (500 to 800 μm diam. of perithecia, 110 to 130 × 12 to 14 μm of asci, 16 to 19 × 6 to 11 μm of ascospores in IRAN 12363 F; 2 to 46 × 2 to 3 μm of conidiophores, 6.5 to 11 × 2 to 3 μm of conidia in ex-type culture CBS 114319) [22]. Phylogenetically, the CFCC 55706 and XJAU KY-2-1 isolates from P. laurifolia in the present study formed a supported clade with the ex-type strain of C. gamsii, CBS 114319, in the phylogram based on combined ITS and LSU loci (Figure 1). In addition, there were no differences between CFCC 55706, XJAU KY-2-1, and CBS 114319 in the ITS and LSU sequence data. Hence, the samples collected from P. laurifolia in Xinjiang, China were identified as the types of C. gamsii from the species of Coniochaeta based on both morphology and molecular data.
3.4. Pathogenicity Test
P. laurifolia leaves that were inoculated with the CFCC 55706 isolate developed symptoms of brown leaf spot disease that were the same as those observed in the field under natural conditions. Inoculated leaves initially developed a white, irregularly round, necrotic lesion, becoming brownish black over time, although the center of the lesion remained white (Figure 3F). The negative control leaves did not develop disease symptoms (Figure 3E). The lesion expansion on the leaves of the P. laurifolia seedlings was the same as that observed on detached leaves. The fungus was reisolated from symptomatic leaves and identified as C. gamsii based on morphological characteristics and DNA sequence data, thus fulfilling Koch’s postulates.
Coniolariella gamsii showed different degrees of pathogenicity on P. laurifolia, P. bolleana, and P. tomentosa leaves (Figure 4). The average lesion diameters that developed on inoculated P. bolleana, P. tomentosa, and P. laurifolia leaves were 25.51 mm, 17.71 mm, and 12.18 mm, respectively, 10 days after inoculation (Figure 5).
3.5. Cultural Characteristics of the Pathogen
The colony growth rates of the CFCC 55706 isolate at 0 °C, 4 °C, 37 °C, 38 °C, and 40 °C were significantly slower than those at temperatures ranging from 8 °C to 36 °C (p < 0.05). The optimum temperature for hyphal growth was 28 °C (Figure 6A).
The growth rate of mycelia was significantly different under different light conditions (p < 0.05). The daily growth rates of colonies under 24 h of light, 12/12 h of light/darkness, and 24 h of darkness were 2 mm/d, 12 mm/d, 15 mm/d, respectively. The optimum illumination for hyphal growth was 24 h of darkness (Figure 6B).
There was no significant difference in the growth rate and sporulation of the CFCC 55706 strain in the PLPDA, SLPDA, and MLPDA media (based on 16 treatments in each group). However, there was a significant difference in the growth rate and sporulation time of the CFCC 55706 strain on the three different types of leaf medium (p < 0.05). Mycelia grew the fastest (26 mm/d) on the PLPDA medium (Figure 7A) and only took 12 d to produce spores. The growth rates of the strain on the MLPDA and SLPDA media were 18 mm/d (Figure 7B) and 20 mm/d (Figure 7C), and the sporulation took more than 20 d. The slowest growth rate of the strain was on the PDA medium (12 mm/d), and sporulation took more than 25 d (Figure 7D). The optimum medium formulation was the one where the PLPDA medium contained 5 g/liter of leaves that had been boiled for 5 min, the MLPDA medium contained 15 g/liter of leaves that had been boiled for 20 min, and the SLPDA medium contained 10 g/liter of leaves that had been boiled for 5 min.
4. Discussion
In this study, Coniolariella gamsii (Xylariaceae) was isolated and tested on P. laurifolia following Koch’s postulates, which confirmed C. gamsii as a pathogen affecting P. laurifolia leaves. The vast majority of Xylariaceae species are saprophytes that play a role in wood decomposition, and some species spend part of their lifecycle as endophytes [23,24,25]. Previous studies of Xylariaceae have mainly focused on their distribution and classification status. In contrast, there are few reports of plant diseases caused by members of the Xylariaceae [26]. A few species of Xylariaceae are pathogens that can infect plants, principally causing cankers, needle blights, and root-associated diseases on woody hosts [27]. There are reports of Xylariaceae causing leaf spot disease, viz. Xylaria schweinitzii causing leaf spots on Neosinocalamus affinis and X. cubensis causing leaf spots on Hemerocallis fulva [28]. In this study, we consider the fungus Coniolariella gamsii as a new leaf spot pathogen caused by a member of the Xylariaceae that can infect P. laurifolia leaves. The pathogenicity test indicated that C. gamsii can also infect P. bolleana and P. tomentosa, which play important roles as farmland shelterbelts and windbreaks, and in sand fixation, in Xinjiang, China [29,30]. Analyses of variance revealed that the lesions that developed on the three Populus spp. were significantly different. The largest lesions developed on P. bolleana leaves, suggesting that P. bolleana was the most susceptible to C. gamsii. Further research is needed to determine whether this species is a potential pathogen which can affect other species of poplar. Xylariaceous plant pathogens tend to either infect their hosts via wounded tissue or via root tissue, with the exception of those that seem to transition from an endophytic lifestyle to a pathogenic lifestyle [26]. For example, E. mammata is known to cause cankers on several woody plant genera, infecting the host via open wounds [31,32]. In contrast, Rosellinia necatrix is a soil fungus and does not appear to need wound openings to enter the root tissue of susceptible plants [33,34]. Our pathogenicity tests showed that C. gamsii can infect Populus spp. via leaf wounds. This is consistent with the findings of previous studies, which have shown that the mechanism of pathogenicity of Xylariaceous species is either through direct damage to the vascular system or through the translocation of phytotoxins, or both [26]. C. gamsii can infect three Populus spp.; its mechanism of pathogenicity is still unclear.
The host plant can have an important effect on spore formation. Ushiyam suggested that adding plant leaves to a water–agar medium could induce the sporulation of fungi [35]. For example, Phoma herbarum’s production of pycnidium can be enhanced by adding Commelina communis juice to OMA+ medium [36]. In this study, the addition of Populus leaves to a PDA medium to make a PLPDA medium induced C. gamsii sporulation more rapidly than the MLPDA or SLPDA media. The optimum temperature for the mycelial growth of X. C. gamsii is 28 °C. Furthermore, the hyphae of C. gamsii continued to grow at 0 °C but ceased to grow at 40 °C. Therefore, 40 °C is a lethal temperature for this pathogen. These results indicate that CFCC 55706 grows the fastest under warm conditions but that little or no growth occurs at high or low temperatures, whereas the optimum illumination condition for C. gamsii hyphal growth is 24 h of darkness.
5. Conclusions
Coniolariella gamsii is an emerging pathogen causing P. laurifolia brown leaf spot in Altay City, Xinjiang, China. The pathogenicity test confirmed that C. gamsii is a pathogenic fungus that can also infect P. bolleana and P. tomentosa leaves. The optimum culture conditions for C. gamsii mycelial growth are 28 °C and darkness. The most suitable sporulation medium is a PDA medium with 5 g/liter of Populus leaves that have been boiled for 5 min. The data obtained in this study should provide a theoretical basis for monitoring and preventing this disease. The potential impact of C. gamsii on poplar production in this area of China warrants further investigation to determine potential disease management strategies.
Author Contributions
Conceptualization, R.M.; formal analysis, C.L., R.M. and C.W.; funding acquisition, R.M.; investigation, R.M., H.C. and Z.Z.; software, H.L. and S.C.; visualization, C.L.; writing—original draft preparation, C.L. and R.M.; writing—review and editing, R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 31960316). The project was also supported by the Scientific Research Program of the Forestry Department of the Xinjiang Uygur Autonomous Region 2020LCJPC-1.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wiehle, M.; Vornam, B.; Wesche, K.; Goenster, S.; Buerkert, A. Population structure and genetic diversity of Populus laurifolia in fragmented riparian gallery forests of the Mongolian Altai Mountains. Flora 2016, 224, 112–122. [Google Scholar] [CrossRef]
- Zheng, S.; Zhang, J.G.; He, C.Y.; Bao, J.; Duan, A.G.; Zeng, Y.F.; Sai, L. Genetic diversity of Populus laurifolia and Populus nigra along Erqis River. For. Res. 2014, 27, 295–301. [Google Scholar]
- Checa, J.; Arenal, F.; Blanco, N.; Rogers, J.D. Coniolariella hispanica sp. nov. and other additions to Coniolariella. Mycol. Res. 2008, 112, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Zare, R.; Asgari, B.; Gams, W. The species of Coniolariella. Mycologia 2010, 102, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Stchigel, A.M.; Cano, J.; Calduch, M.; Hawksworth, D.L.; Guarro, J. Molecular phylogeny of Coniochaetales. Mycol. Res. 2006, 110, 1271–1289. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.Y. Rust fungi from the desert of northern Xinjiang. Acta Mycol. Sin. 1989, 8, 259–269. [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China; Mycotaxon, Ltd.: Ithaca, NY, USA, 2005; p. 430. [Google Scholar]
- Jiang, N.; Voglmayr, H.; Xue, H.; Piao, C.G.; Li, Y. Morphology and Phylogeny of Pestalotiopsis (Sporocadaceae, Amphisphaeriales) from Fagaceae Leaves in China. Microbiol Spectr. 2022, 10, e03272-22. [Google Scholar] [CrossRef]
- Zhang, Q.T.; He, M. Canker on bark of Populus spp. caused by Cytospora tritici, a new disease in China. Plant Dis. 2012, 96, 1578. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, Y.N.; Yang, J.Y.; Li, D.W.; Li, Y.; Bian, L.M.; Ye, J.R. Shoot blight on Chinese fir (Cunninghamia lanceolata) is caused by Bipolaris oryzae. Plant Dis. 2017, 102, 500–506. [Google Scholar] [CrossRef]
- Solarte, F.A.; Muñoz, C.G.; Maharachchikumbura, S.; Álvarez, E. Diversity of Neopestalotiopsis and Pestalotiopsis spp., causal agents of guava scab in Colombia. Plant Dis. 2017, 102, 49–59. [Google Scholar] [CrossRef]
- Ma, R.; Ye, S.H.; Zhao, Y.; Michailides, T.J.; Tian, C.M. New leaf and fruit disease of Juglans regia caused by Juglanconis juglandina in Xinjiang, China. For. Pathol. 2019, 49, e12537. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- O'Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus fusariumare nonorthologous. Molecul Phylogenet Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Ma, C.Y.; Xue, H.; Piao, C.G.; Li, Y. A new Arthrinium-like genus of Amphisphaeriales in China. MycoKeys 2022, 92, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees; Institute of Electrical and Electronics Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Jia, X.H.; Wang, W.; Fu, J.F.; Du, Y.M.; Wang, Y.; Zhou, R.J.; Liu, B.L. Biological characteristics and host range of Athelia bombacina causing postharvest fruit rot on pear. Acta Hortic. Sin. 2020, 47, 1253–1263. [Google Scholar]
- Asgari, B.; Zare, R. Two new Coniochaeta species from Iran. Nova Hedwig. 2006, 82, 227–236. [Google Scholar] [CrossRef]
- Davis, E.C.; Franklin, J.B.; Shaw, A.J.; Vilgalys, R. Endophytic Xylaria (Xylariaceae) among liverworts and angiosperms: Phylogenetics, distribution, and symbiosis. Am J Bot. 2003, 90, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.D. The Xylariaceae: Systematic, biological and evolutionary aspects. Mycologia 1979, 71, 1–42. [Google Scholar] [CrossRef]
- González, F.E.S.M. A Mycofloristic and Cultural Study of the Xylariaceae of México; Washington State University: Washington, DC, USA, 1992. [Google Scholar]
- Garcia-Aroca, T.; Price, P.P.; Tomaso-Peterson, M.; Allen, T.W.; Wilkerson, T.H.; Spurlock, T.N.; Faske, T.R.; Bluhm, B.; Conner, K.; Sikora, E.; et al. Xylaria necrophora sp. nov. is an emerging root-associated pathogen responsible for taproot decline of soybean in the southern United States. Mycologia 2021, 113, 326–347. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.L.; Jonglaekha, N.; Kshirsagar, A.; Maitland, D.J.; Mekkamol, S.; Nugent, L.K.; Phosri, C.; Rodtong, S.; Ruchichachorn, N.; Sangvichien, E.; et al. The Xylariaceae as phytopathogens. Recent Res. Dev. Plant Sci. 2003, 1, 1–19. [Google Scholar]
- Shen, J.W.; Zhou, S.J.; Yu, H.Y.; Zhang, B.; Zhou, H. Studies on the biological characteristics of Xylaria pedunculata. J. Henan Agric. Sci. 2008, 12, 75–77. [Google Scholar]
- Li, P. Studies on the Physiological Mechanism of Populus bolleana in Response to the Fungal Canker Diseases; Chinese Academy of Forestry Sciences: Beijing, China, 2019. [Google Scholar]
- Jiang, H.; Zhou, F.; Xia, S.C.; Che, J.H.; Wang, G.F. Providencia rettgeri as the causal agent of the brown slime flux of Populus tomentosa. For. Pathol. 2020, 50, e12576. [Google Scholar]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Introductory Mycology, 4th ed.; Wiley & Sons: New York, NY, USA, 1996. [Google Scholar]
- Balasuriya, A.; Adikaram, N.K.B. Some spatial, temporal and spatio-temporal considerations of wood decay of tea (Camellia sinensis), caused by Nemania diffusa (syn. Hypoxylon vestitum). Crop Prot. 2009, 28, 273–279. [Google Scholar] [CrossRef]
- Pérez-Jiménez, R.M. A review of the biology and pathogenicity of Rosellinia necatrix—The cause of white root rot disease of fruit trees and other plants. J. Phytopathol. 2006, 154, 257–266. [Google Scholar] [CrossRef]
- Eguchi, N.; Kondo, K.I.; Yamagishi, N. Bait twig method for soil detection of Rosellinia necatrix, causal agent of white root rot of Japanese pear and apple, at an early stage of tree infection. J. Gen. Plant Pathol. 2009, 75, 325–330. [Google Scholar] [CrossRef]
- Ushiyam, K. Mass production of conidiophores of Phomopsis spp., Colletotrichum and Pestalotiopsis spp. by agar leaf disk method. Proceeding Kanto-Tosan Plant Prot. Soc. 1995, 42, 101–103. [Google Scholar]
- Liang, D.D.; Zhang, Y.L.; Gu, Z.M. Study on sporulation conditions of Phoma herbarum SYAU-06 Strain. Agrochemicals 2020, 59, 219–222. [Google Scholar]
Figure 1.
Phylogram of Coniolariella resulting from a Maximum Likelihood analysis, based on a combined matrix of ITS and LSU. Numbers above the branches indicate ML bootstraps (left, ML BS ≥ 50%) and Bayesian Posterior Probabilities (right, BPP ≥ 0.90). Xylaria badia (BCC 1190) was selected as an outgroup. New isolates from the present study are marked in blue.
Figure 1.
Phylogram of Coniolariella resulting from a Maximum Likelihood analysis, based on a combined matrix of ITS and LSU. Numbers above the branches indicate ML bootstraps (left, ML BS ≥ 50%) and Bayesian Posterior Probabilities (right, BPP ≥ 0.90). Xylaria badia (BCC 1190) was selected as an outgroup. New isolates from the present study are marked in blue.

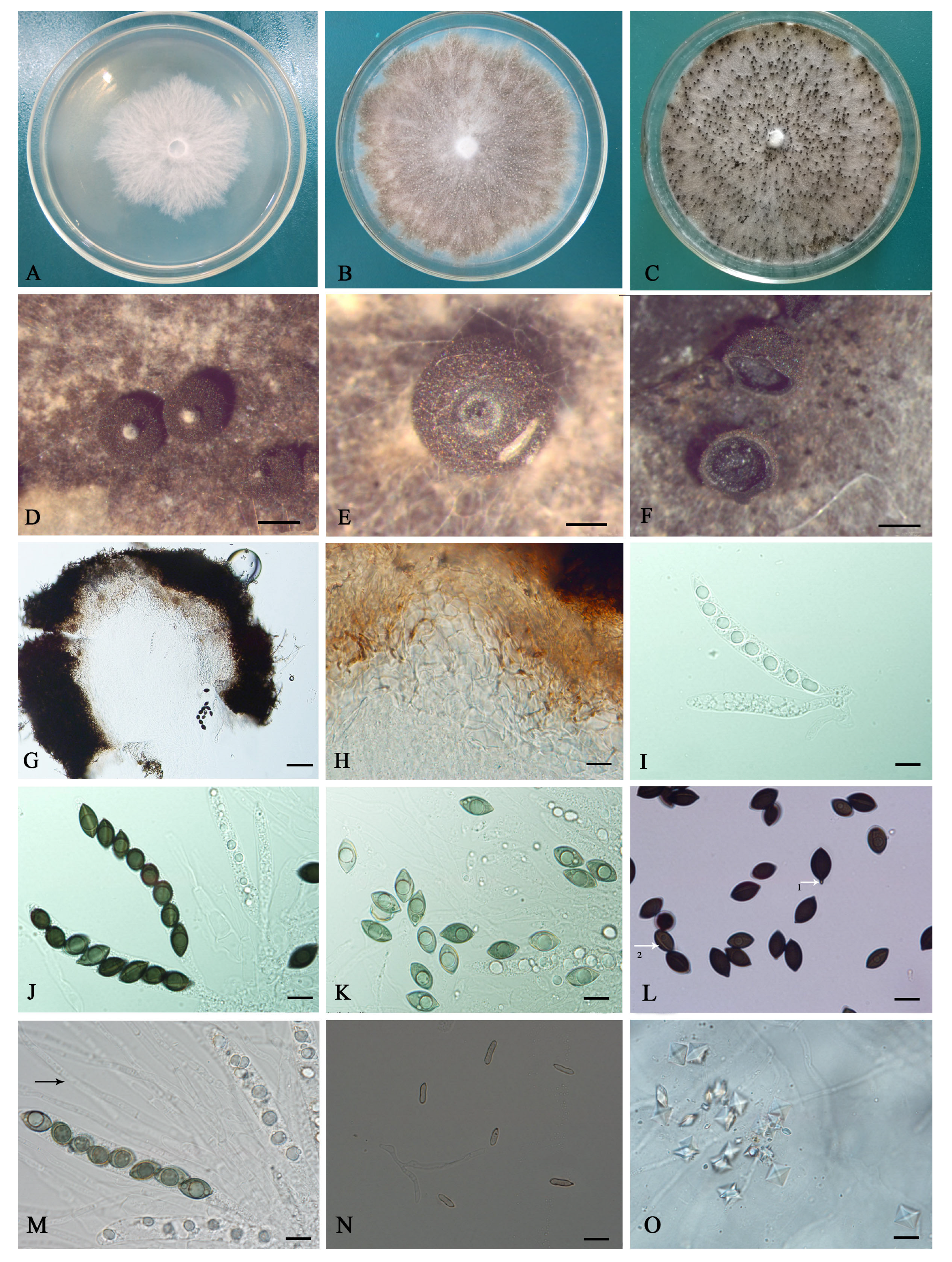
Figure 2.
Cultural characteristics and morphological characteristics of Coniolariella gamsii. (A) colony on PDA after 3 d; (B) colony on PDA after 15 d; (C) colony growth on PDA after 25 d; (D) ascomata formation on PDA; (E) transverse section of the top of the ascomata showing white projections protruding; (F) ascomata slit; (G,H) sections of ascomata showing three-layered peridium. (I) immature asci containing ascospores; (J) mature asci; (K) immature ascospores with an obvious guttule; (L) mature ascospores with protruding ends (arrow 1) and germ slits (arrow 2); (M) paraphyses (arrow); (N) conidiophores and conidia; (O) crystals. (Bars: D, F = 200 μm; E = 100 μm; G = 50 μm; H–O = 10 μm).
Figure 2.
Cultural characteristics and morphological characteristics of Coniolariella gamsii. (A) colony on PDA after 3 d; (B) colony on PDA after 15 d; (C) colony growth on PDA after 25 d; (D) ascomata formation on PDA; (E) transverse section of the top of the ascomata showing white projections protruding; (F) ascomata slit; (G,H) sections of ascomata showing three-layered peridium. (I) immature asci containing ascospores; (J) mature asci; (K) immature ascospores with an obvious guttule; (L) mature ascospores with protruding ends (arrow 1) and germ slits (arrow 2); (M) paraphyses (arrow); (N) conidiophores and conidia; (O) crystals. (Bars: D, F = 200 μm; E = 100 μm; G = 50 μm; H–O = 10 μm).

Figure 3.
Brown leaf spot disease symptoms on Populus laurifolia leaves. (A–C) Natural disease symptoms on P. laurifolia growing in the field. (D) Leaves of a seedling inoculated with Coniolariella gamsii; (E) the control leaf; (F) symptoms of disease on a leaf inoculated with C. gamsii.
Figure 3.
Brown leaf spot disease symptoms on Populus laurifolia leaves. (A–C) Natural disease symptoms on P. laurifolia growing in the field. (D) Leaves of a seedling inoculated with Coniolariella gamsii; (E) the control leaf; (F) symptoms of disease on a leaf inoculated with C. gamsii.

Figure 4.
Brown leaf spot disease symptoms on (A–C) Populus Laurifolia; (D–F) Populus bolleana; and (G–I) Populus tomentosa leaves. The wound on the left-hand side of each leaf was mock-inoculated (control - CK); the wound on the right-hand side of each leaf was inoculated with Coniolariella gamsii. (A,D,G) 2 d after inoculation; (B,E,H) 6 d after inoculation; (C) 10 d after inoculation; (F,I) 12 d after inoculation.
Figure 4.
Brown leaf spot disease symptoms on (A–C) Populus Laurifolia; (D–F) Populus bolleana; and (G–I) Populus tomentosa leaves. The wound on the left-hand side of each leaf was mock-inoculated (control - CK); the wound on the right-hand side of each leaf was inoculated with Coniolariella gamsii. (A,D,G) 2 d after inoculation; (B,E,H) 6 d after inoculation; (C) 10 d after inoculation; (F,I) 12 d after inoculation.

Figure 5.
Analysis of variance of lesions that developed on Populus laurifolia, Populus bolleana, and Populus tomentosa leaves following inoculation with Coniolariella gamsii. Values shown are means ± the standard errors. Different lowercase letters above bars indicate a significant difference in lesion size between poplar species at p < 0.05.
Figure 5.
Analysis of variance of lesions that developed on Populus laurifolia, Populus bolleana, and Populus tomentosa leaves following inoculation with Coniolariella gamsii. Values shown are means ± the standard errors. Different lowercase letters above bars indicate a significant difference in lesion size between poplar species at p < 0.05.

Figure 6.
Cultural characteristics of Coniolariella gamsii strain CFCC 55706. (A) Diameter of 7-day-old colony incubated at three different temperatures. (B) Diameter of 3-day-old colony grown under three different illumination conditions. Values shown are means ± the standard errors. Different lowercase letters above bars indicate a significant difference in colony diameter between treatments at p < 0.05.
Figure 6.
Cultural characteristics of Coniolariella gamsii strain CFCC 55706. (A) Diameter of 7-day-old colony incubated at three different temperatures. (B) Diameter of 3-day-old colony grown under three different illumination conditions. Values shown are means ± the standard errors. Different lowercase letters above bars indicate a significant difference in colony diameter between treatments at p < 0.05.

Figure 7.
The growth rate and sporulation time of Coniolariella gamsii strain CFCC 55706 on different media. (A–C) media: PDA and modified PDA media that included 5 g/liter, 10 g/liter, 15 g/liter, or 20 g/liter of Populus (PLPDA), Malus (MLPDA), or Salix (SLPDA) leaves that had been boiled for 5 min, 10 min, 15 min, or 20 min. (D) Optimal media formulations for mycelial growth and sporulation. Values shown are means ± the standard error. Different lowercase letters above bars indicate a significant difference in colony diameter at p < 0.05.
Figure 7.
The growth rate and sporulation time of Coniolariella gamsii strain CFCC 55706 on different media. (A–C) media: PDA and modified PDA media that included 5 g/liter, 10 g/liter, 15 g/liter, or 20 g/liter of Populus (PLPDA), Malus (MLPDA), or Salix (SLPDA) leaves that had been boiled for 5 min, 10 min, 15 min, or 20 min. (D) Optimal media formulations for mycelial growth and sporulation. Values shown are means ± the standard error. Different lowercase letters above bars indicate a significant difference in colony diameter at p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Isolates and GenBank accession numbers of species of Coniolariella for phylogenetic analysis.
Table 1.
Isolates and GenBank accession numbers of species of Coniolariella for phylogenetic analysis.
| Species of Coniolariella | Isolate | Host | Country | Sequences | |
|---|---|---|---|---|---|
| ITS | LSU | ||||
| C. ershadii | CBS 119785 | Pistacia vera | Iran | GU553328 | GU553331 |
| C. gamsii | CBS 114379 | Hordeum vulgare | Iran | GU553325 | GU553329 |
| C. gamsii | CFCC 55706 | Populus laurifolia | China | MZ423210 | MZ436978 |
| C. gamsii | XJAU KY-2-1 | Populus laurifolia | China | MZ423211 | MZ436979 |
| C. hispanica | CBS 124506 | Eryngium campestre | Spain | MH863381 | MH874902 |
| C. limoniispora | 108fa | Vitis vinifera | USA | NA | EF489469 |
| C. limoniispora | AH24323 | Nicotiana glauca | Spain | AY908997 | EF489464 |
| C. macrothecia | CBS 125772 | Soil | USA | MH863751 | MH875219 |
| Xylaria badia | BCC 1190 | NA | Thailand | AB376711 | NA |
Note: NA, not applicable. Strains used in this study are marked in bold.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, C.; Lu, H.; Chen, H.; Chen, S.; Wang, C.; Zhou, Z.; Ma, R. Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China. Diversity 2023, 15, 1190. https://doi.org/10.3390/d15121190
AMA Style
Liu C, Lu H, Chen H, Chen S, Wang C, Zhou Z, Ma R. Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China. Diversity. 2023; 15(12):1190. https://doi.org/10.3390/d15121190
Chicago/Turabian StyleLiu, Chuli, Hailong Lu, Hongjin Chen, Shuikang Chen, Caixia Wang, Zhiyong Zhou, and Rong Ma. 2023. "Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China" Diversity 15, no. 12: 1190. https://doi.org/10.3390/d15121190
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.