
Diversity of Botryosphaeriaceae Species Associated with Grapevine Trunk Diseases in the Czech Republic
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Isolation
2.2. Morphology
2.3. DNA Extraction and Amplification
2.4. Phylogenetic Analyses
3. Results
3.1. Fungal Isolation
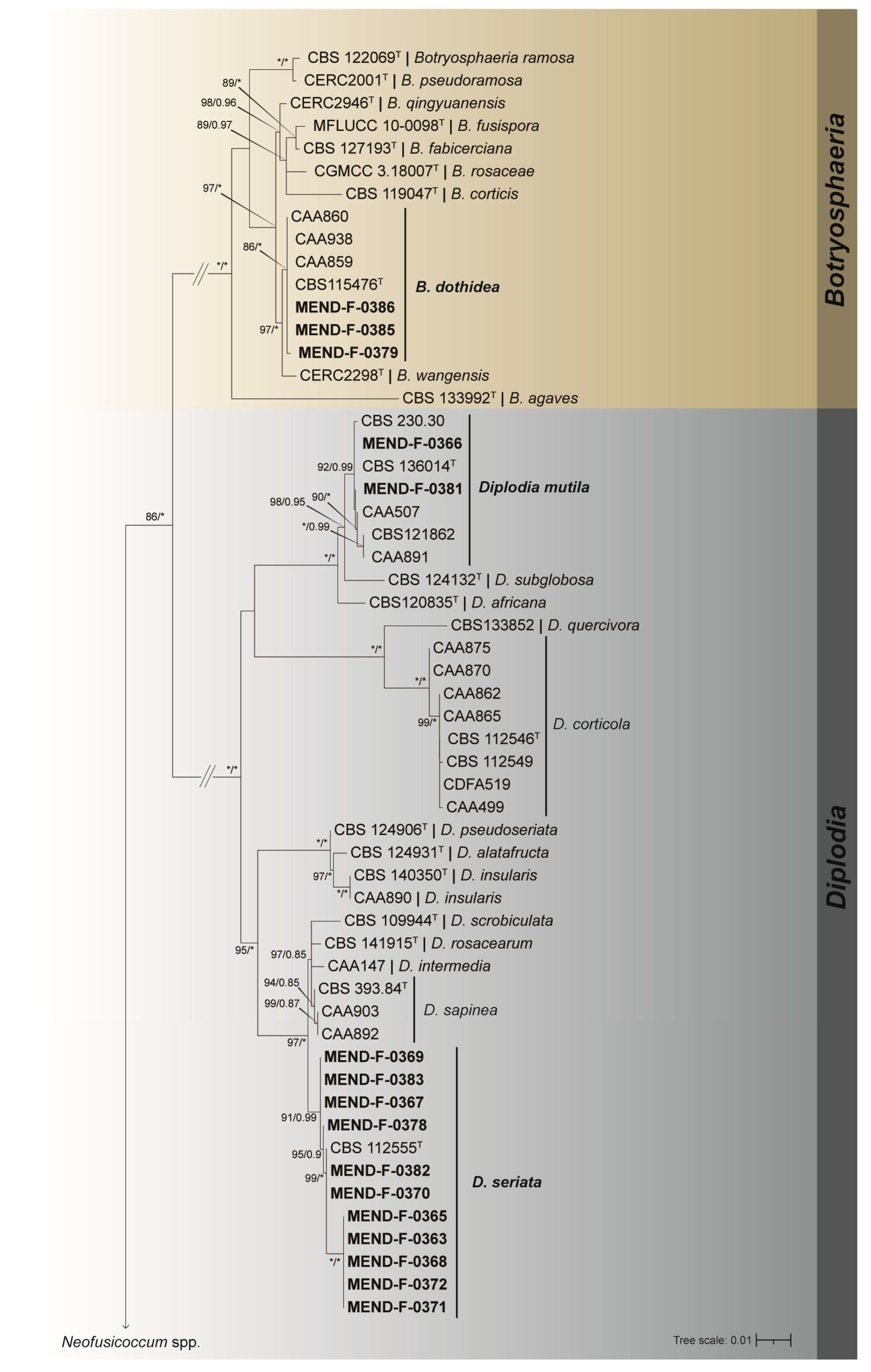
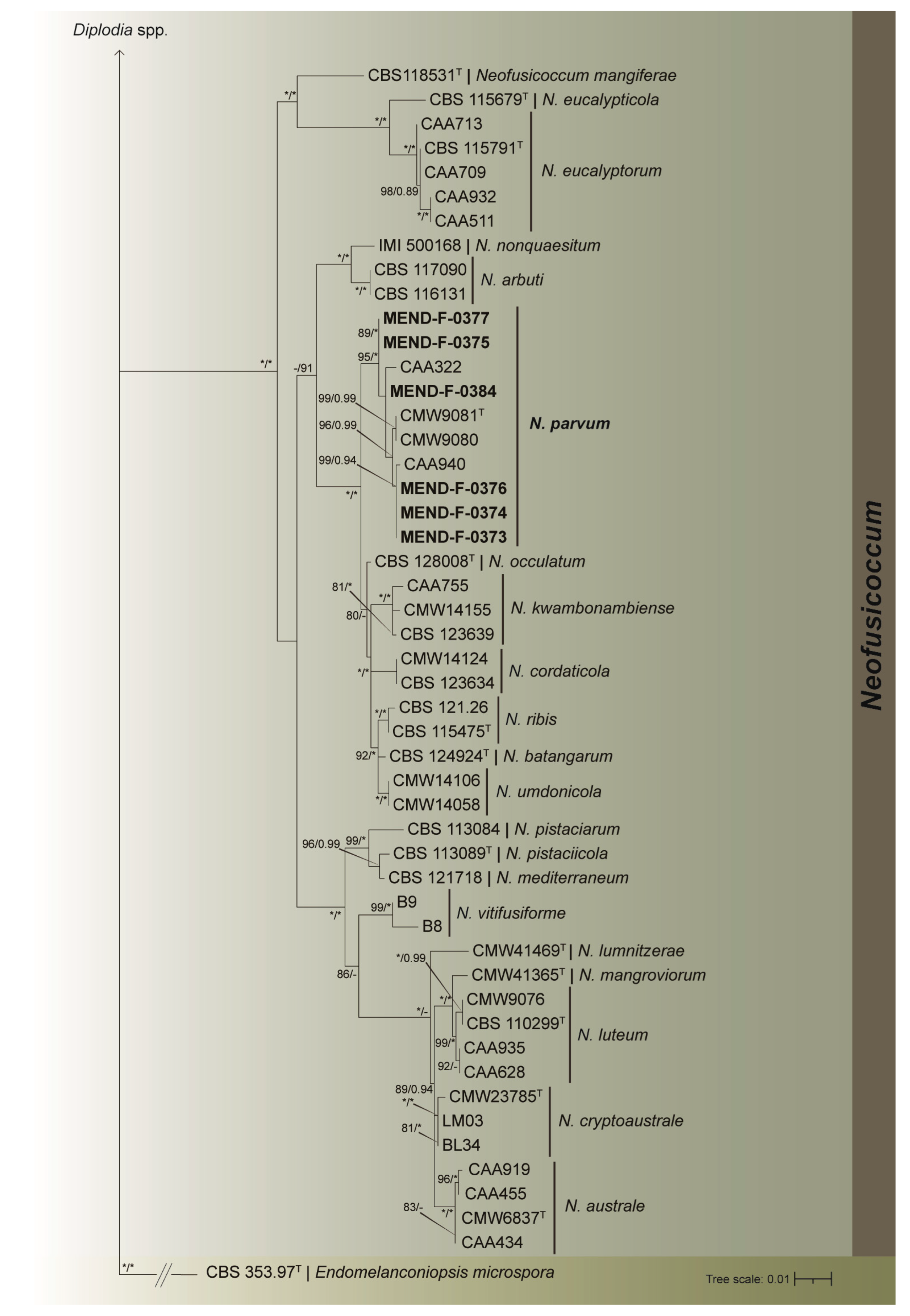
3.2. Phylogenetic Analyses
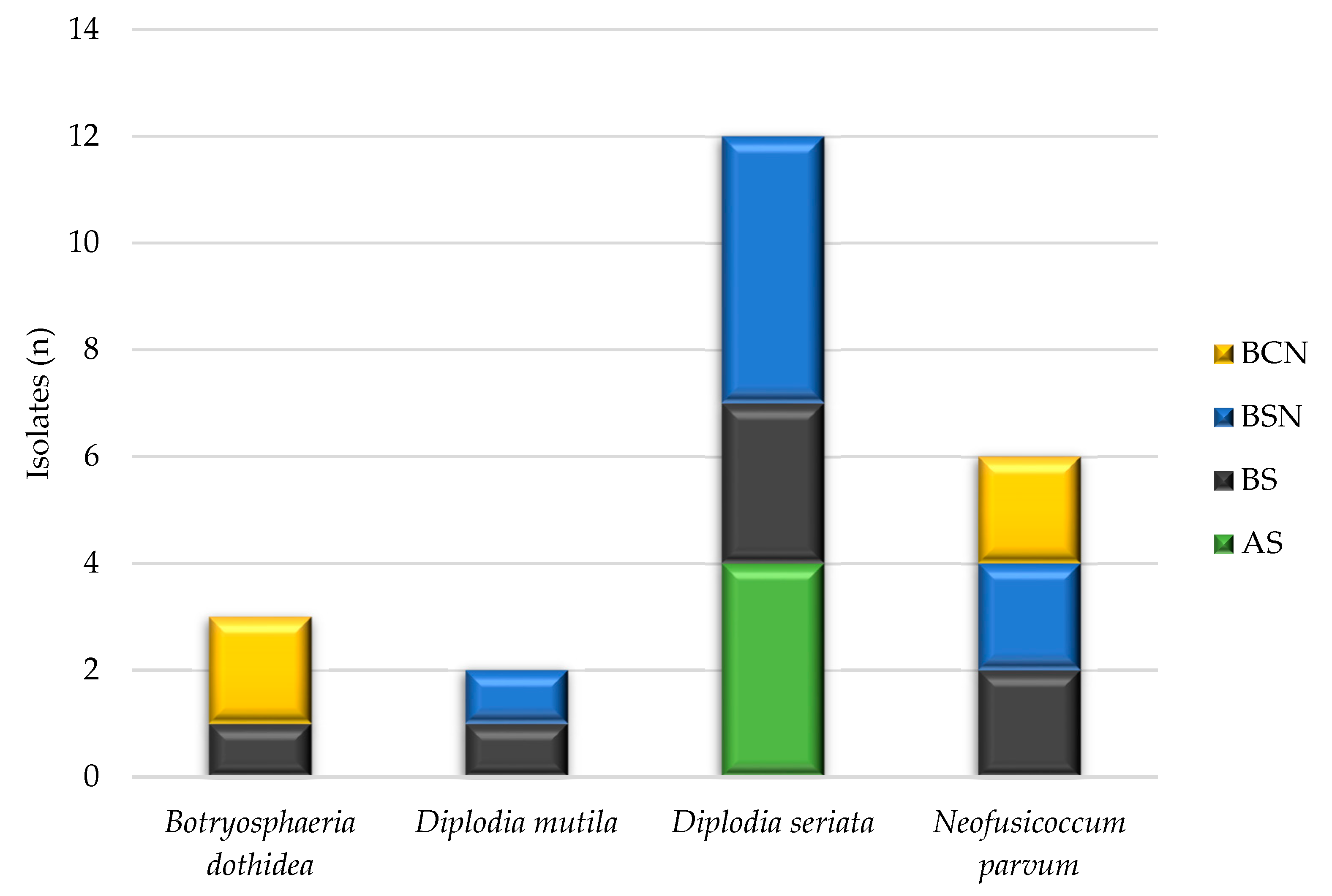
3.3. Species Diversity in Different Grapevine Varieties and Wood necrosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations; FAOSTAT Database: Rome, Italy, 2023. [Google Scholar]
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. Grapevine Trunk Diseases. A Review; OIV Publications: Paris, France, 2016; p. 24. [Google Scholar]
- Bertsch, C.; Ramírez-Suero, M.; Magninrobert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk Diseases With Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Dis. 2017, 102, 12–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbez-Torres, J. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, 5–45. [Google Scholar]
- Aroca, Á.; Gramaje, D.; Armengol, J.; García-Jiménez, J.; Raposo, R. Evaluation of the grapevine nursery propagation process as a source of Phaeoacremonium spp. and Phaeomoniella chlamydospora and occurrence of trunk disease pathogens in rootstock mother vines in Spain. Eur. J. Plant Pathol. 2010, 126, 165–174. [Google Scholar] [CrossRef]
- Pintos, C.; Redondo, V.; Costas, D.; Aguín, O.; Mansilla, P. Fungi associated with grapevine trunk diseases in nursery-produced Vitis vinifera plants. Phytopathol. Mediterr. 2018, 57, 407–424. [Google Scholar] [CrossRef]
- Carbone, M.J.; Gelabert, M.; Moreira, V.; Mondino, P.; Alaniz, S. Grapevine nursery propagation material as source of fungal trunk disease pathogens in Uruguay. Front. Fungal Biol. 2022, 3. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Ridgway, H.J.; Jones, E.E.; Jaspers, M.V. Inoculum sources of Botryosphaeriaceae species in New Zealand grapevine nurseries. Eur. J. Plant Pathol. 2013, 135, 159–174. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Berraf-Tebbal, A.; Mahamedi, A.E.; Aigoun-Mouhous, W.; Spetik, M.; Čechová, J.; Pokluda, R.; Baránek, M.; Eichmeier, A.; Alves, A. Lasiodiplodia mitidjana sp. nov. and other Botryosphaeriaceae species causing branch canker and dieback of Citrus sinensis in Algeria. PLoS ONE 2020, 15, e0232448. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.B.; Aronstein, K.; Flores, J.M.; Vojvodic, S.; Palacio, M.A.; Spivak, M. Standard methods for fungal brood disease research. J. Apic. Res. 2019, 52, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Eichmeier, A.; Pecenka, J.; Spetik, M.; Necas, T.; Ondrasek, I.; Armengol, J.; León, M.; Berlanas, C.; Gramaje, D. Fungal Trunk Pathogens Associated with Juglans regia in the Czech Republic. Plant Dis. 2019, 104, 761–771. [Google Scholar] [CrossRef]
- Spetik, M.; Berraf-Tebbal, A.; Gramaje, D.; Mahamedi, A.E.; Stusková, K.; Burgova, J.; Eichmeier, A. Paecilomyces clematidis (Eurotiales, Thermoascaceae): A new species from Clematis root. Phytotaxa 2022, 559, 238–246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic rDNA ITS2 Types within a Monophyletic Lineage of the Fungus Fusarium Are Nonorthologous. Mol. Phylogenetics Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Cowley, A.; Uludag, M.; Gur, T.; McWilliam, H.; Squizzato, S.; Park, Y.M.; Buso, N.; Lopez, R. The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Res. 2015, 43, 580–584. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R.; Gubler, W.D. Pathogenicity of Botryosphaeriaceae Species Isolated from Grapevine Cankers in California. Plant Dis. 2009, 93, 584–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akgul, D.S.; Savas, N.G.; Eskalen, A. First Report of Wood Canker Caused by Botryosphaeria dothidea, Diplodia seriata, Neofusicoccum parvum, and Lasiodiplodia theobromae on Grapevine in Turkey. Plant Dis. 2013, 98, 568. [Google Scholar] [CrossRef]
- Arkam, M.; Alves, A.; Lopes, A.; Čechová, J.; Pokluda, R.; Eichmeier, A.; Zitouni, A.; Mahamedi, A.E.; Berraf-Tebbal, A. Diversity of Botryosphaeriaceae causing grapevine trunk diseases and their spatial distribution under different climatic conditions in Algeria. Eur. J. Plant Pathol. 2021, 161, 933–952. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Savocchia, S. A review of Botryosphaeriaceae species associated with grapevine trunk diseases in Australia and New Zealand. Australas. Plant Pathol. 2019, 48, 3–18. [Google Scholar] [CrossRef]
- Yan, J.-Y.; Xie, Y.; Zhang, W.; Wang, Y.; Liu, J.-K.; Hyde, K.D.; Seem, R.C.; Zhang, G.-Z.; Wang, Z.-Y.; Yao, S.-W.; et al. Species of Botryosphaeriaceae involved in grapevine dieback in China. Fungal Divers. 2013, 61, 221–236. [Google Scholar] [CrossRef]
- Pitt, W.M.; Huang, R.; Steel, C.C.; Savocchia, S. Identification, distribution and current taxonomy of Botryosphaeriaceae species associated with grapevine decline in New South Wales and South Australia. Aust. J. Grape Wine Res. 2010, 16, 258–271. [Google Scholar] [CrossRef]
- Phillips, A.J.L. Botryosphaeria species associated with diseases of grapevines in Portugal. Phytopathol. Mediterr. 2002, 41, 3–18. [Google Scholar] [CrossRef]
- Mohammadi, H.; Gramaje, D.; Banihashemi, Z.; Armengol, J. Characterization of Diplodia seriata and Neofusicoccum parvum associated with grapevine decline in Iran. J. Agric. Sci. Technol. 2013, 15, 603–616. [Google Scholar]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Guerrero, J.C.; Guevara, J.; Gubler, W.D. Identification and Pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the Causal Agents of Bot Canker Disease of Grapevines in Mexico. Plant Dis. 2008, 92, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Kovács, C.; Balling, P.; Bihari, Z.; Nagy, A.; Sándor, E. Incidence of grapevine trunk diseases is influenced by soil, topology and vineyard age, but not by Diplodia seriata infection rate in the Tokaj Wine Region, Hungary. Phytoparasitica 2017, 45, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Chebil, S.; Fersi, R.; Bouzid, M.; Quaglino, F.; Chenenaoui, S.; Melki, I.; Durante, G.; Zacchi, E.; Bahri, B.A.; Bianco, P.A.; et al. Fungi from the Diaporthaceae and Botryosphaeriaceae families associated with grapevine decline in Tunisia. Cienc. E Investig. Agrar. 2017, 44, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Fourie, P.H.; Halleen, F. Occurrence of grapevine trunk disease pathogens in rootstock mother plants in South Africa. Australas. Plant Pathol. 2004, 33, 313–315. [Google Scholar] [CrossRef]
- Aroca, A.; García-Figueres, F.; Bracamonte, L.; Luque, J.; Raposo, R. A Survey of Trunk Disease Pathogens within Rootstocks of Grapevines in Spain. Eur. J. Plant Pathol. 2006, 115, 195–202. [Google Scholar] [CrossRef]
- Eichmeier, A.; Pečenka, J.; Peňázová, E.; Baránek, M.; Català-García, S.; León, M.; Armengol, J.; Gramaje, D. High-throughput amplicon sequencing-based analysis of active fungal communities inhabiting grapevine after hot-water treatments reveals unexpectedly high fungal diversity. Fungal Ecol. 2018, 36, 26–38. [Google Scholar] [CrossRef]
- Baránek, M.; Armengol, J.; Holleinová, V.; Pečenka, J.; Calzarano, F.; Peňázová, E.; Vachůn, M.; Eichmeier, A. Incidence of symptoms and fungal pathogens associated with grapevine trunk diseases in Czech vineyards: First example from a north-eastern European grape-growing region. Phytopathol. Mediterr. 2018, 57, 449–458. [Google Scholar] [CrossRef]
- Pečenka, J.; Eichmeier, A.; Peňázová, E.; Baránek, M.; León, M.; Armengol, J. First Report of Dactylonectria torresensis Causing Black-Foot Disease on Grapevines in the Czech Republic. Plant Dis. 2018, 102, 2038. [Google Scholar] [CrossRef]
- Mondragón, F.A.; Rodríguez-Alvarado, G.; Gmez-Dorantes, N.; Guerra-Santos, J.J.; Fernández-Pavía, S.P. Botryosphaeriaceae: A complex, diverse and cosmopolitan family of fungi. Rev. Mex. Cienc. Agrícolas 2021, 12, 643–654. [Google Scholar] [CrossRef]
- Spetik, M.; Cechova, J.; Eichmeier, A. First report of Neofusicoccum parvum causing stem blight and dieback of highbush blueberry in the Czech Republic. Plant Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Pečenka, J.; Tekielska, D.A.; Kocanová, M.; Peňázová, E.; Berraf-Tebbal, A.; Eichmeier, A. First Report of Lasiodiplodia theobromae Causing Decline of Blueberry (Vaccinium corymbosum) in the Czech Republic. Plant Dis. 2020, 105, 215. [Google Scholar] [CrossRef] [PubMed]
- Elena, G.; DI Bella, V.; Armengol, J.; Luque, J. Viability of Botryosphaeriaceae species pathogenic to grapevine after hot water treatment. Phytopathol. Mediterr. 2015, 54, 325–334. [Google Scholar] [CrossRef]
- Štůsková, K.; Pečenka, J.; Tekielska, D.A.; Špetík, M.; Bytešníková, Z.; Švec, P.; Ondreáš, F.; Ridošková, A.; Richtera, L.; Adam, V.; et al. The in vitro effects of selected substances and nanomaterials against Diaporthe eres, Diplodia seriata and Eutypa lata. Ann. Appl. Biol. 2023, 182, 226–237. [Google Scholar] [CrossRef]
- Špetík, M.; Balík, J.; Híc, P.; Hakalová, E.; Štůsková, K.; Frejlichová, L.; Tříska, J.; Eichmeier, A. Lignans Extract from Knotwood of Norway Spruce—A Possible New Weapon against GTDs. J. Fungi 2022, 8, 357. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling | Locality | Sampling Year | Age of the Vineyards | Sampled Vines (n) | Cultivar |
|---|---|---|---|---|---|
| 1. | Klentnice (48°51′27.4″ N 16°39′04.9″ E) | 2019 | 30 | 10 | Pálava * |
| 2. | Pavlov (48°51′49.1″ N 16°39′23.0″ E) | 2019 | 30 | 10 | Blaufränkisch ** |
| 3. | Maliny (48°49′36.6″ N 16°37′29.9″ E) | 2019 | 30 | 10 | Pinot Noir ** |
| 4. | Maliny (48°49′34.9″ N 16°37′23.6″ E) | 2019 | 3 | 10 | Welschriesling * |
| Locus | Primer | Primer DNA Sequence (5′-3′) | Reference |
|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG | [18] |
| ITS4 | TCCTCCGCTTATTGATATGC | ||
| tef | EF1-728F | CATCGAGAAGTTCGAGAAGG | [19] |
| EF1-986R | TACTTGAAGGAACCCTTACC | ||
| tub2 | T1 | AACATGCGTGAGATTGTAAGT | [20] |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC | [21] |
| Species | Strain | Host | Geographic Origin | ITS | tub2 | tef |
|---|---|---|---|---|---|---|
| Botryosphaeria agaves | CBS 133992T | Agave sp. | Thailand | JX646791 | JX646841 | JX646856 |
| B. corticis | CBS 119047T | Vaccinium corymbosum | United States | DQ299245 | EU673107 | EU017539 |
| B. dothidea | CBS 115476T | Prunus sp. | Switzerland | AY236949 | AY236927 | AY236898 |
| B. dothidea | CAA859 | Quercus ilex | Portugal | MK940302 | MT309378 | MT309403 |
| B. dothidea | CAA938 | Quercus suber | Portugal | MT237173 | MT309379 | MT309401 |
| B. dothidea | CAA860 | Quercus suber | Portugal | MK940295 | MT309380 | MT309402 |
| B. dothidea | MEND-F-0386 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987974 | OQ994785 | OQ994763 |
| B. dothidea | MEND-F-0385 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987975 | OQ994786 | OQ994764 |
| B. dothidea | MEND-F-0379 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987976 | OQ994787 | OQ994765 |
| B. fabicerciana | CBS 127193T | Eucalyptus sp. | China | HQ332197 | KF779068 | HQ332213 |
| B. fusispora | MFLUCC 10–0098T | Entada sp. | Thailand | JX646789 | JX646839 | JX646854 |
| B. pseudoramosa | CERC2001T | Eucalyptus sp. | China | KX277989 | KX278198 | KX278094 |
| B. qingyuanensis | CERC2946T | Eucalyptus sp. | China | KX278000 | KX278209 | KX278105 |
| B. ramosa | CBS 122069T | Eucalyptus camaldulensis | Australia | EU144055 | KF766132 | EU144070 |
| B. rosaceae | CGMCC 3.18007T | – | China | KX197074 | KX197101 | KX197094 |
| B. wangensis | CERC2298T | Cedrus deodara | China | KX278002 | KX278211 | KX278107 |
| Diplodia africana | CBS 120835T | Prunus persica | South Africa | EF445343 | KF766129 | EF445382 |
| D. alatafructa | CBS 124931T | Pterocarpus angolensis | South Africa | FJ888460 | MG015799 | FJ888444 |
| D. corticola | CBS 112546T | Quercus ilex | Spain | AY259090 | EU673117 | EU673310 |
| D. corticola | CBS 112549 | Quercus suber | Portugal | AY259100 | DQ458853 | AY573227 |
| D. corticola | CAA862 | Eucalyptus globulus | Portugal | MK940298 | MT309381 | MT309410 |
| D. corticola | CAA865 | Pinus pinaster | Portugal | MK940296 | MT309382 | MT309411 |
| D. corticola | CAA870 | Quercus ilex | Portugal | MK940303 | MT309383 | MT309408 |
| D. corticola | CAA875 | Quercus suber | Portugal | MK940297 | MT309384 | MT309409 |
| D. corticola | CAA499 | Eucalyptus globulus | Portugal | MG015741 | MG015800 | MG015723 |
| D. corticola | CDFA519 | Quercus sp. | United States | GU799472 | GU799466 | GU799469 |
| D. insularis | CBS 140350T | Pistacia lentiscus | Italy | KX833072 | MG015809 | KX833073 |
| D. insularis | CAA890T | Eucalyptus globulus | Portugal | MK940299 | MT309385 | MT309406 |
| D. intermedia | CAA147T | Malus pumila | Portugal | GQ923857 | MG015811 | GQ923825 |
| D. mutila | CBS 136014 | Populus alba | Portugal | KJ361837 | MG015815 | KJ361829 |
| D. mutila | CBS 230.30 | Phoenix dactylifera | United States | DQ458886 | DQ458849 | DQ458869 |
| D. mutila | CAA507 | Fraxinus ornus | Portugal | MG015746 | MG015816 | MG015728 |
| D. mutila | CBS 121862 | Pyrus communis | Netherlands | KX464093 | KX464799 | KX464567 |
| D. mutila | CAA891 | Eucalyptus globulus | Portugal | MK940300 | MT309386 | MT309407 |
| D. mutila | MEND-F-0366 | V. vinifera ‘Palava’ | Czechia | OQ987977 | OQ994788 | OQ994766 |
| D. mutila | MEND-F-0381 | V. vinifera ‘Palava’ | Czechia | OQ987978 | OQ994789 | OQ994767 |
| D. pseudoseriata | CBS 124906T | Blepharocalyx salicifolius | Uruguay | EU080927 | MG015820 | EU863181 |
| D. quercivora | CBS 133852 | Quercus canariensis | Tunisia | JX894205 | MG015821 | JX894229 |
| D. rosacearum | CBS 141915T | Eriobotrya japonica | Italy | KT956270 | MG015823 | KU378605 |
| D. sapinea | CBS 393.84T | Pinus nigra | Netherlands | DQ458895 | DQ458863 | DQ458880 |
| D. sapinea | CAA892 | Pinus pinaster | Portugal | MK940292 | MT309387 | MT309404 |
| D. sapinea | CAA903 | Quercus suber | Portugal | MK940312 | MT309388 | MT309405 |
| D. scrobiculata | CBS 109944T | Pinus greggii | Mexico | DQ458899 | DQ458867 | DQ458884 |
| D. seriata | CBS 112555T | Vitis vinifera | Portugal | AY259094 | DQ458856 | AY573220 |
| D. seriata | MEND-F-0367 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987979 | OQ994790 | OQ994768 |
| D. seriata | MEND-F-0370a | V. vinifera ‘Welschriesling’ | Czechia | OQ987980 | OQ994791 | OQ994769 |
| D. seriata | MEND-F-0383 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987981 | OQ994792 | OQ994770 |
| D. seriata | MEND-F-0365a | V. vinifera ‘Welschriesling’ | Czechia | OQ987982 | OQ994793 | OQ994771 |
| D. seriata | MEND-F-0363 | V. vinifera ‘Palava’ | Czechia | OQ987983 | OQ994794 | OQ994772 |
| D. seriata | MEND-F-0368 | V. vinifera ‘Blaufränkisch’ | Czechia | OQ987984 | OQ994795 | OQ994773 |
| D. seriata | MEND-F-0372 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987985 | OQ994796 | OQ994774 |
| D. seriata | MEND-F-0369a | V. vinifera ‘Welschriesling’ | Czechia | OQ987986 | OQ994797 | OQ994775 |
| D. seriata | MEND-F-0382 | V. vinifera ‘Blaufränkisch’ | Czechia | OQ987987 | OQ994798 | OQ994776 |
| D. seriata | MEND-F-0371a | V. vinifera ‘Welschriesling’ | Czechia | OQ987988 | OQ994799 | OQ994777 |
| D. seriata | MEND-F-0378 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987989 | OQ994800 | OQ994778 |
| D. subglobosa | CBS 124132T | Fraxinus excelsior | Spain | DQ458887 | DQ458852 | DQ458871 |
| Endomelanconiopsis microspora | CBS 353.97T | Soil | Papua N. Guinea | EU683655 | KX464893 | EU683636 |
| Neofusicoccum arbuti | CBS 116131 | Arbutus menziesii | United States | AY819720 | KF531793 | KF531792 |
| N. arbuti | CBS 117090 | Arbutus menziesii | United States | AY819724 | KF531794 | KF531791 |
| N. australe | CMW6837T | Acacia sp. | Australia | AY339262 | AY339254 | AY339270 |
| N. australe | CAA919 | Eucalyptus globulus | Portugal | MK940294 | MT309395 | MT309423 |
| N. australe | CAA434 | Eucalyptus globulus | Portugal | KT440913 | KX505927 | KT440973 |
| N. australe | CAA455 | Eucalyptus globulus | Portugal | KT440915 | KX505928 | KT440975 |
| N. batangarum | CBS 124924T | Terminalia catappa | Cameroon | FJ900607 | FJ900634 | FJ900653 |
| N. cordaticola | CMW14124 | – | – | EU821925 | EU821865 | EU821895 |
| N. cordaticola | CBS 123634 | Syzygium cordatum | South Africa | EU821898 | EU821838 | EU821868 |
| N. cryptoaustrale | CMW23785T | Eucalyptus sp. | South Africa | FJ752742 | FJ752756 | FJ752713 |
| N. cryptoaustrale | LM03 | Pistacia lentiscus | – | KX505912 | KX505930 | KX505903 |
| N. cryptoaustrale | BL34 | Vitis vinifera | – | KJ638328 | KX505931 | KX505904 |
| N. eucalypticola | CBS 115679T | Eucalyptus grandis | Australia | AY615141 | AY615125 | AY615133 |
| N. eucalyptorum | CBS 115791T | Eucalyptus grandis | South Africa | AF283686 | AY236920 | AY236891 |
| N. eucalyptorum | CAA932 | Eucalyptus globulus | Portugal | MK940311 | MT309396 | MT309422 |
| N. eucalyptorum | CAA511 | Eucalyptus globulus | Portugal | KX505907 | KX505919 | KX505896 |
| N. eucalyptorum | CAA709 | Eucalyptus globulus | Portugal | KT440941 | KX505920 | KT441001 |
| N. eucalyptorum | CAA713 | Eucalyptus globulus | Portugal | KT440943 | KX505921 | KT441003 |
| N. kwambonambiense | CBS 123639 | Syzygium cordatum | South Africa | EU821900 | EU821840 | EU821870 |
| N. kwambonambiense | CAA755 | Eucalyptus globulus | Portugal | KT440946 | KX505917 | KT441006 |
| N. kwambonambiense | CMW14155 | – | – | EU821923 | EU821863 | EU821893 |
| N. lumnitzerae | CMW41469T | Barringtonia racemosa | South Africa | KP860881 | KP860801 | KP860724 |
| N. luteum | CBS 110299T | Vitis vinifera | Portugal | AY259091 | DQ458848 | KX464688 |
| N. luteum | CAA935 | Eucalyptus globulus | Portugal | MK940305 | MT309397 | MT309418 |
| N. luteum | CAA628 | Fraxinus excelsior | Portugal | KX505911 | KX505929 | KX505902 |
| N. luteum | CMW9076 | – | – | AY236946 | AY236922 | AY236893 |
| N. mangiferae | CBS 118531T | Mangifera indica | Australia | AY615185 | AY615172 | DQ093221 |
| N. mangroviorum | CMW41365T | Avicennia marina | South Africa | KP860859 | KP860779 | KP860702 |
| N. mediterraneum | CBS 121718 | Eucalyptus sp. | Greece | GU251176 | GU251836 | GU251308 |
| N. mediterraneum | CAA002 | Pistacia vera | United States | EU017537 | KX505925 | KX505900 |
| N. mediterraneum | SPA9 | Pistacia lentiscus | – | KX505910 | KX505926 | KX505901 |
| N. nonquaesitum | IMI500168 | Vaccinium corymbosum | – | JX217819 | KX505918 | KX505895 |
| N. occulatum | CBS 128008T | Eucalyptus grandis | Australia | EU301030 | EU339472 | EU339509 |
| N. parvum | CMW9081T | Populus nigra | New Zealand | AY236943 | AY236917 | AY236888 |
| N. parvum | CAA940 | Eucalyptus globulus | Portugal | MK940304 | MT309399 | MT309421 |
| N. parvum | CMW9080 | – | – | AY236942 | AY236916 | AY236887 |
| N. parvum | CAA322 | Malus pumila | Portugal | KX505906 | KX505916 | KX505894 |
| N. parvum | MEND-F-0375 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987990 | OQ994801 | OQ994779 |
| N. parvum | MEND-F-0376 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987991 | OQ994802 | OQ994780 |
| N. parvum | MEND-F-0377 | V. vinifera ‘Blaufränkisch’ | Czechia | OQ987992 | OQ994803 | OQ994781 |
| N. parvum | MEND-F-0374 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987993 | OQ994804 | OQ994782 |
| N. parvum | MEND-F-0373 | V. vinifera ‘Blaufränkisch’ | Czechia | OQ987994 | OQ994805 | OQ994783 |
| N. parvum | MEND-F-0384 | V. vinifera ‘Pinot Noir’ | Czechia | OQ987995 | OQ994806 | OQ994784 |
| N. pistaciarum | CBS 113084 | – | United States | KX464187 | KX464999 | KX464713 |
| N. pistaciicola | CBS 113089T | Pistacia vera | United States | KX464199 | KX465014 | KX464727 |
| N. ribis | CBS 115475T | Ribes sp. | United States | AY236935 | AY236906 | AY236877 |
| N. ribis | CBS 121.26 | Ribes sp. | – | AF241177 | AY236908 | AY236879 |
| N. umdonicola | CMW14106 | – | – | EU821899 | EU821839 | EU821869 |
| N. umdonicola | CMW14058 | – | – | EU821904 | EU821844 | EU821874 |
| N. vitifusiforme | B8 | Vitis vinifera | – | KC469638 | KC884951 | KC884948 |
| N. vitifusiforme | B9 | Vitis vinifera | – | KX505908 | KX505923 | KX505898 |
| Locus | No. of Sequences | No. of Characters | Parsimony-Informative | Constant | Unique | BI Model |
|---|---|---|---|---|---|---|
| ITS | 134 | 503 | 113 | 366 | 24 | GTR + I + G |
| tef | 134 | 336 | 143 | 150 | 43 | HKY + G |
| tub2 | 123 | 420 | 95 | 306 | 19 | GTR + G |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spetik, M.; Tekielska, D.A.; Berraf-Tebbal, A.; Pecenka, J.; Stuskova, K.; Mahamedi, A.E.; Eichmeier, A. Diversity of Botryosphaeriaceae Species Associated with Grapevine Trunk Diseases in the Czech Republic. Diversity 2023, 15, 800. https://doi.org/10.3390/d15070800
Spetik M, Tekielska DA, Berraf-Tebbal A, Pecenka J, Stuskova K, Mahamedi AE, Eichmeier A. Diversity of Botryosphaeriaceae Species Associated with Grapevine Trunk Diseases in the Czech Republic. Diversity. 2023; 15(7):800. https://doi.org/10.3390/d15070800
Chicago/Turabian StyleSpetik, Milan, Dorota Anna Tekielska, Akila Berraf-Tebbal, Jakub Pecenka, Katerina Stuskova, Alla Eddine Mahamedi, and Ales Eichmeier. 2023. "Diversity of Botryosphaeriaceae Species Associated with Grapevine Trunk Diseases in the Czech Republic" Diversity 15, no. 7: 800. https://doi.org/10.3390/d15070800