
A Contribution to the Knowledge of Hydnum (Hydnaceae, Cantharellales) in China, Introducing a New Taxon and Amending Descriptions of Five Known Species

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. Molecular Procedures
2.3. Dataset Assembly
2.4. Phylogenetic Analyses
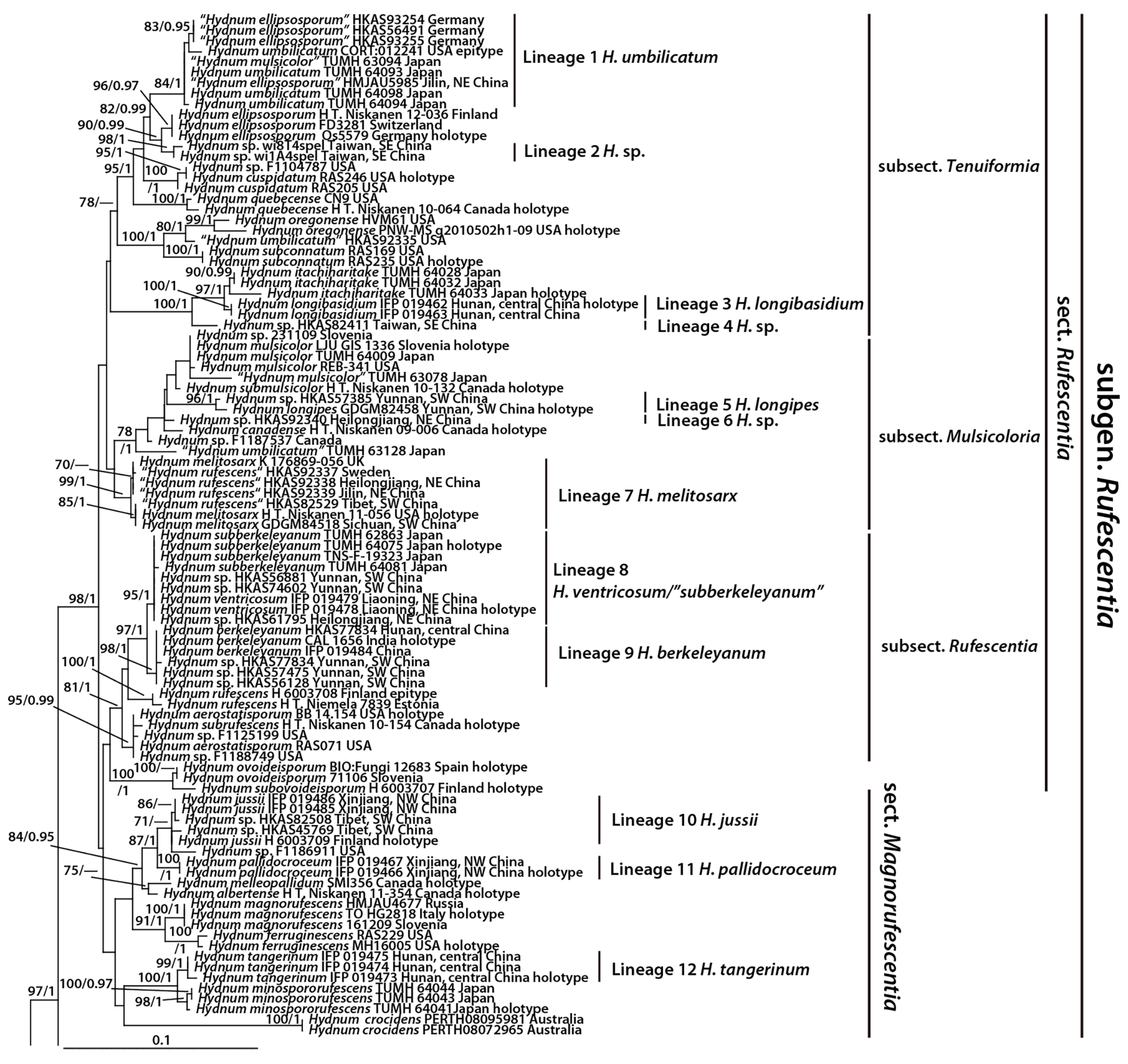
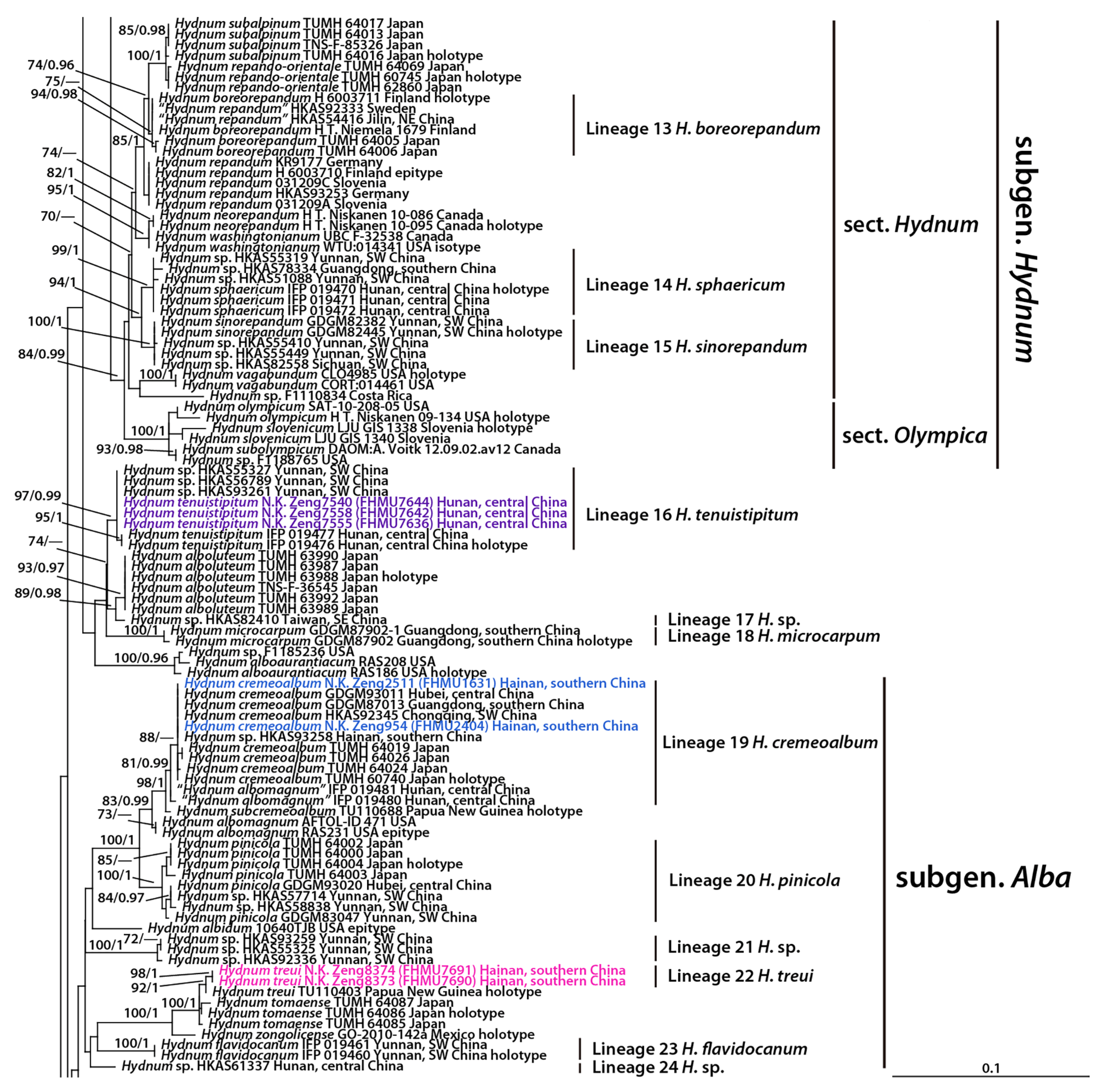
3. Results
3.1. Molecular Data
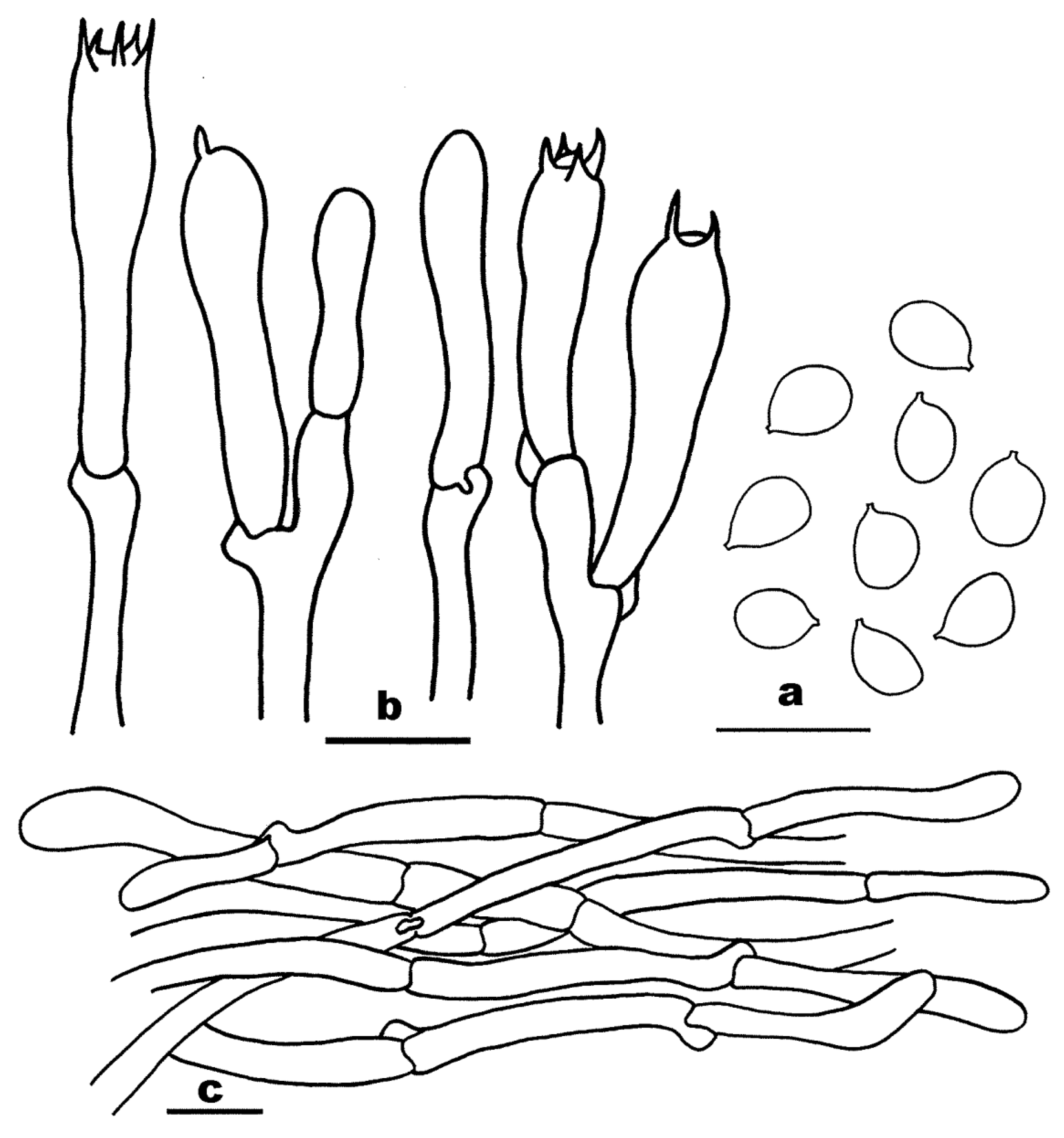
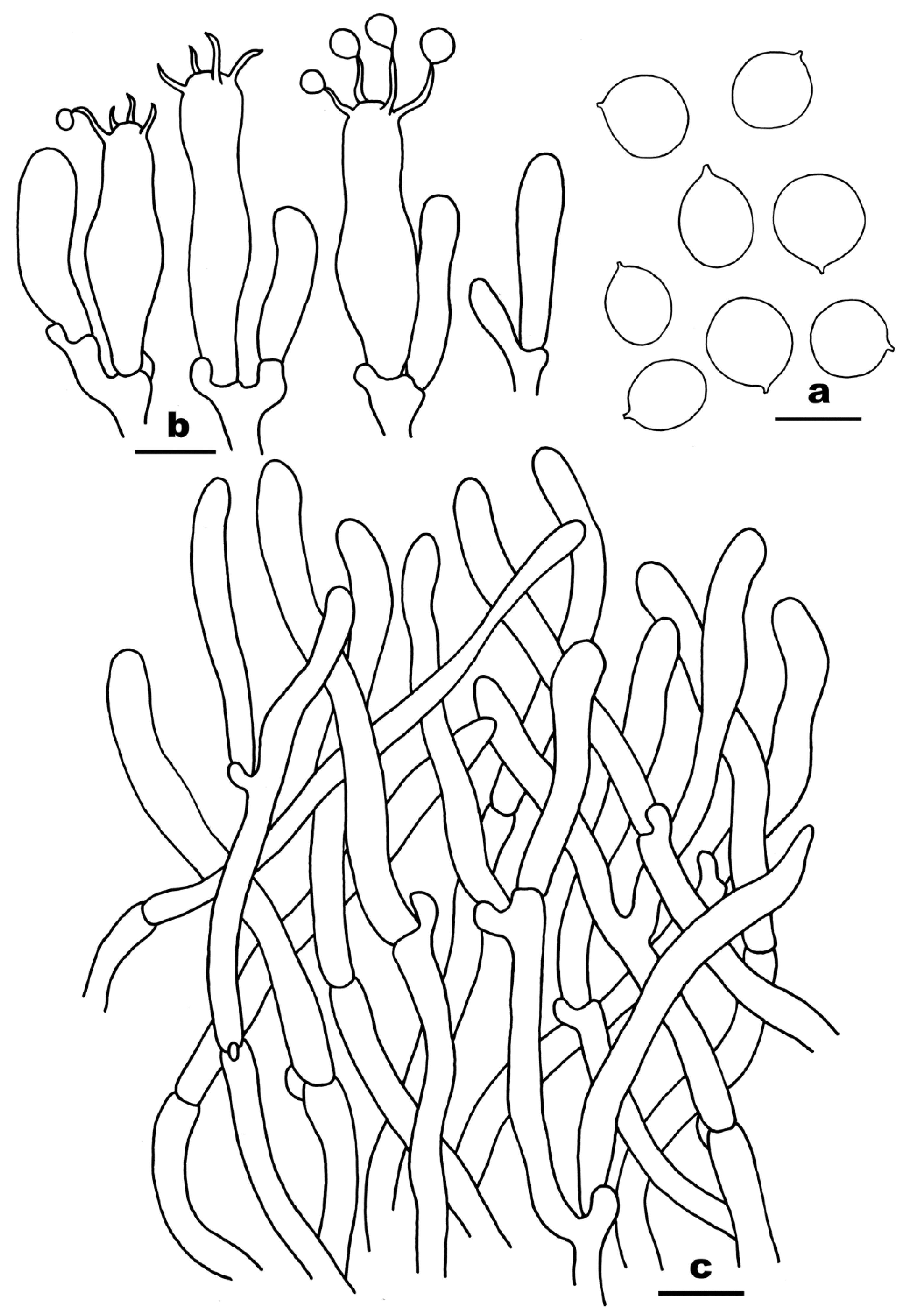
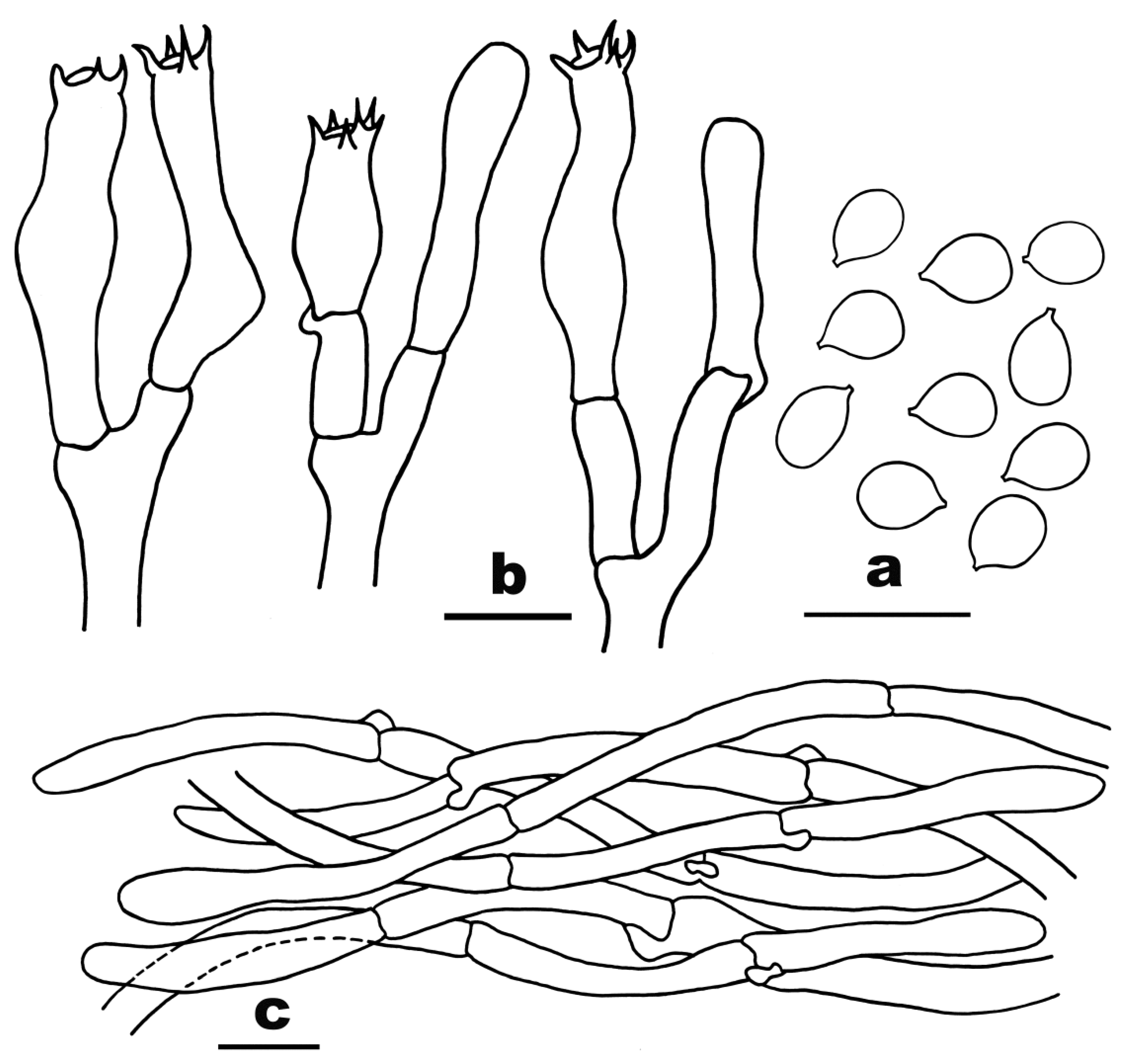
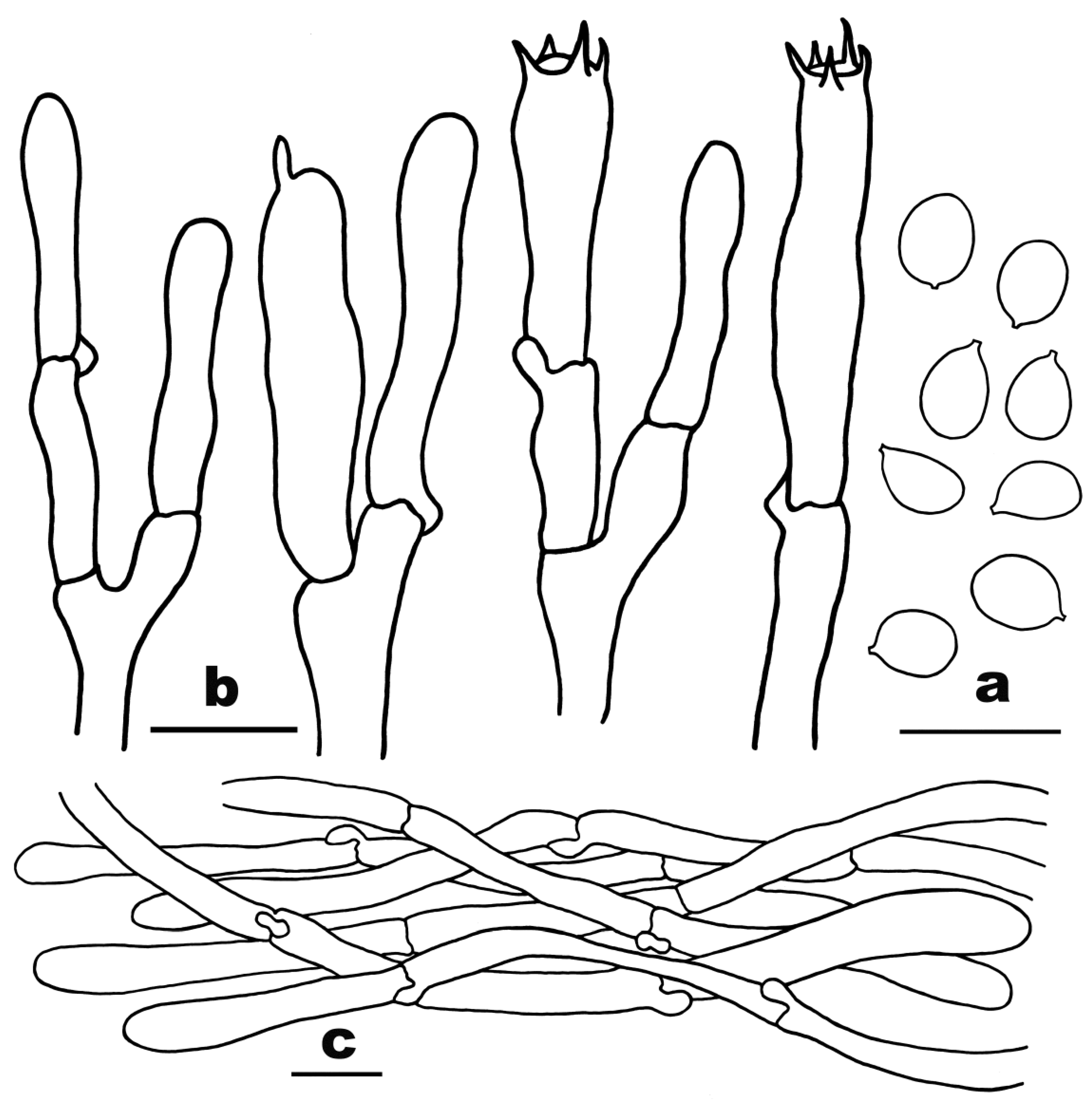
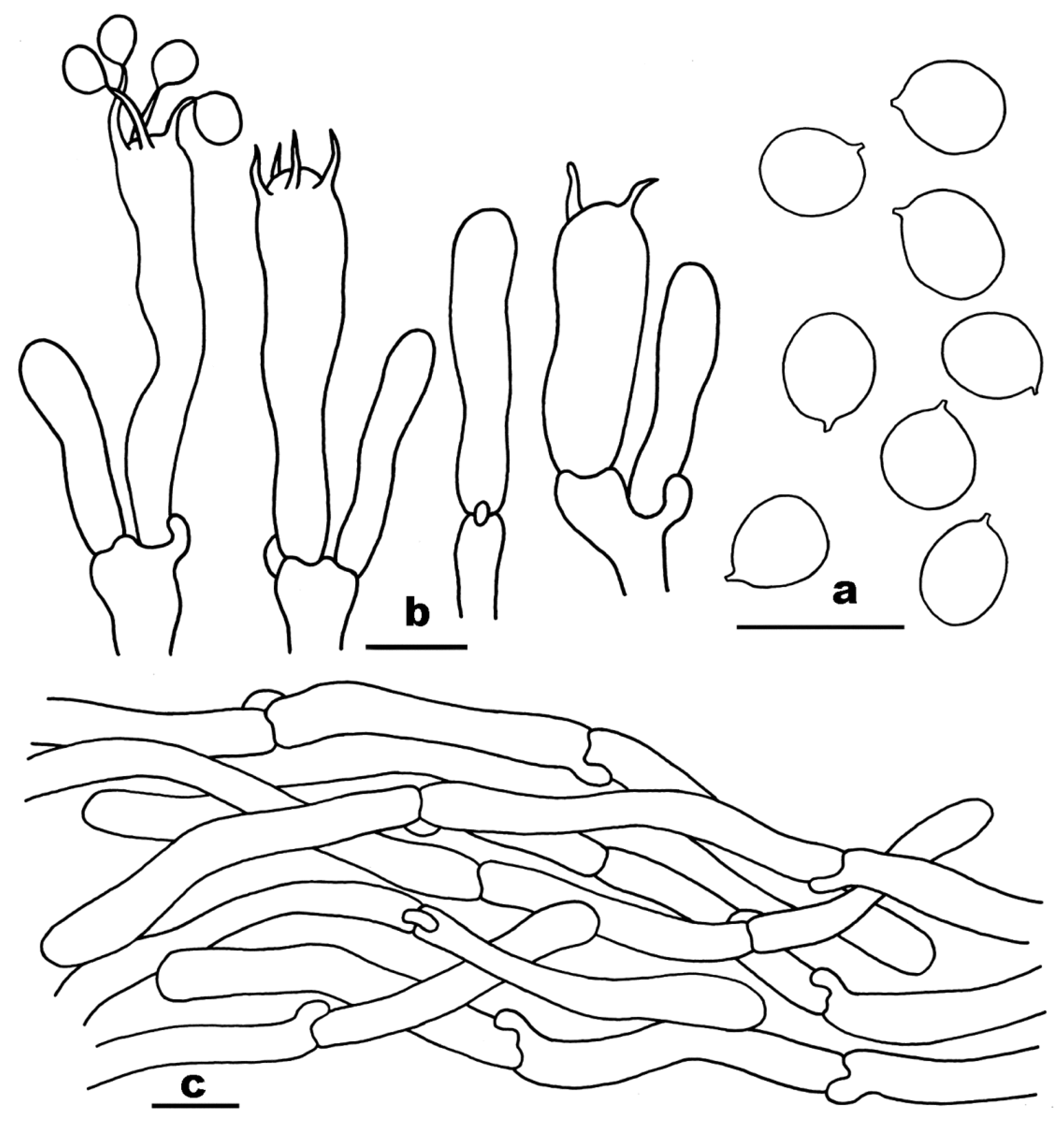
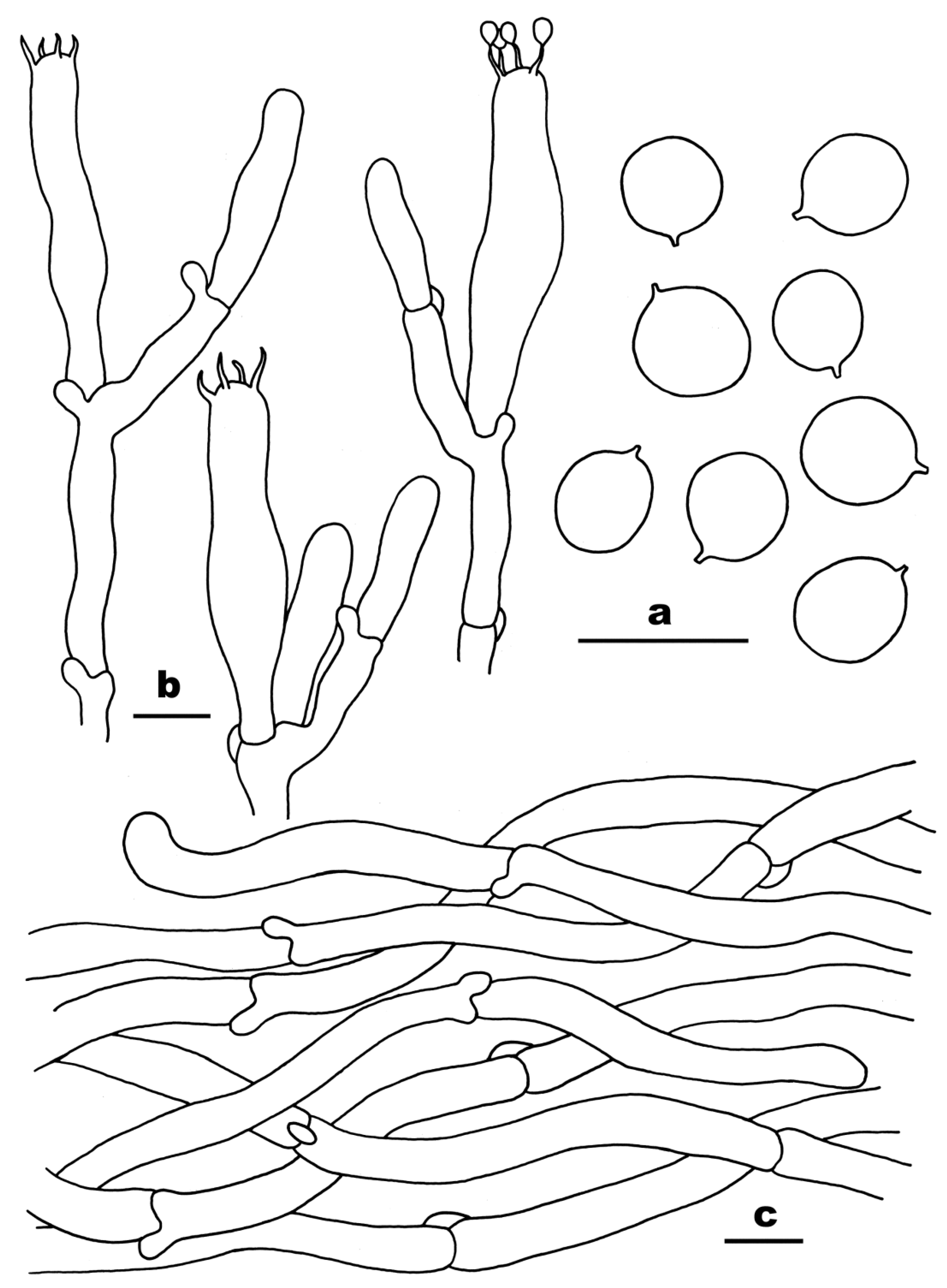
3.2. Taxonomy
- Other known species from China
4. Discussion
5. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Key to the Accepted Species of Hydnum in China | |
|---|---|
| 1. Pileus white, pale cream, cream, yellowish-white, yellowish-grey | 2 |
| 1. Pileus creamy yellow, pale orange, greyish orange, orange | 11 |
| 2. Basidiomata medium to large, pileus > 5 cm diameter | 3 |
| 2. Basidiomata small, pileus < 5 cm diameter | 6 |
| 3. Distributed in sub-alpine regions, elevation > 3000 m, pileal context changing orange-white to pale orange when injured, basidiospores larger (8–9 × 6.5–7.5 μm) | H. sinorepandum |
| 3. Distributed in subtropical/tropical regions, elevation < 2000 m, pileal context unchanging in color when injured, basidiospores smaller (up to 7 μm long and 6.5 μm wide) | 4 |
| 4. Basidiospores larger (6–7 × 5–6.5 μm), basidia longer (21–59 μm), odor sweetish and fragrant | H. treui |
| 4. Basidiospores smaller (up to 5.5 μm long and 4.5 μm wide), basidia shorter (up to 40 μm), odor indistinctive | 5 |
| 5. Stipe cream white, branched, longer (4–6 cm), spines longer (>3 cm long) | H. cremeoalbum |
| 5. Stipe white, not branched, shorter (2–3 cm), spines shorter (<3 cm long) | H. pinicola |
| 6. Context unchanging in color when injured | 7 |
| 6. Context turning red to reddish-brown when injured | H.minus |
| 7. Pileus flabelliform to semicircular, with appressed brownish-orange scales, a distribution in temperate China | H. flabellatum |
| 7. Pileus round, without scales, a distribution in subtropical China | 8 |
| 8. Pileal surface turning brownish-orange to brownish red when injured | H. microcarpum |
| 8. Pileal surface unchanging in color when injured | 9 |
| 9. Basidiospores subglobose, pileipellis composed of frequently branched hyphae | H. tenuistipitum |
| 9. Basidiospores broadly ellipsoid, pileipellis composed of occasionally to rarely branched hyphae | 10 |
| 10. Pileal surface velutinate, pure white, spines shorter (0.2–0.8 mm), basidiospores smaller (5–5.8 × 3.8–4.8 μm) | H. brevispinum |
| 10. Pileal surface glabrous, yellowish-white to yellowish-grey, spines longer (0.5–2 mm), basidiospores larger (7.2–8.8 × 5.5–6.5 μm) | H. flavidocanum |
| 11. Pileipellis a cutis, composed of interwoven to subparallel hyphae | 12 |
| 11. Pileipellis a trichoderm, composed of mostly erect hyphae | H. erectum |
| 12. Basidiomata medium to large, pileus > 5 cm diameter | 13 |
| 12. Basidiomata small, pileus < 5 cm diameter | 14 |
| 13. Basidiospores subglobose, smaller (4.5–6 × 4–5 μm), spines clearly decurrent, pileal surface cream yellow, glabrous, stipe cream white, staining reddish when handled, a distribution in tropical China | H. orientalbidum |
| 13. Basidiospores broadly ellipsoid, larger (7.2–8.8 × 5.8–7 μm), spines non-decurrent, pileal surface orange to brownish-orange, velutinate, stipe white, staining pale orange when handled, a distribution in subtropical China | H. tangerinum |
| 14. Pileal surface changing to orange-white when injured, stipe longer (>5 cm long) | H. longipes |
| 14. Pileal surface unchanging in color when injured, stipe shorter (<5 cm long) | 15 |
| 15. Spines non-decurrent, a distribution in the gymnosperm forest | 16 |
| 15. Spines subdecurrent to decurrent, a distribution in the angiosperm forest | 18 |
| 16. Pileal context white, changing pale orange when injured, basidiospores larger (8–11 × 7–10 μm) | H. melitosarx |
| 16. Pileal context yellowish-white, unchanging in color when injured, basidiospores smaller (up to 9.5 μm long and 8.5 μm) | 17 |
| 17. Pileal surface orange-white to pale orange, stipe longer (3–5.5 cm), light yellow, unchanging in color when injured, basidiospores broadly ellipsoid (7.8–9.5 × 6–7.5 μm), basidia subcylindric or subclavate | H. pallidocroceum |
| 17. Pileal surface orange, stipe shorter (3–3.5 cm), orange-white, staining brownish when handled, basidiospores subglobose (8.2–9 × 7.5–8.5 μm), basidia fusiform to subcylindric, ventricose | H. ventricosum |
| 18. Pileal surface glabrous, basidia shorter (up to 65 μm), stipe surface unchanging in color when handled, basidiospores broadly ellipsoid (Q = 1.2–1.28) | 19 |
| 18. Pileal surface subglabrous to velutinate, basidia longer (up to 82 μm), stipe surface staining pale yellow when handled, basidiospores subglobose (Q = 1.09–1.13) | H. longibasidium |
| 19. Pileus subglobose, pileal surface orange-white, stipe white, spines white, non-decurrent to subdecurrent | H. sphaericum |
| 19. Pileus irregularly round to semicircular, or infundibuliform, pileal surface orange-white to pale orange, but pileal margin whitish (obviously lighter), stipe orange white to pale orange, spines orange white to pale orange, decurrent | H. pallidomarginatum |
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niskanen, T.; Liimatainen, K.; Nuytinck, J.; Kirk, P.; Ibarguren, I.O.; Garibay-Orijel, R.; Norvell, L.; Huhtinen, S.; Kytövuori, I.; Ruotsalainen, J.; et al. Identifying and naming the currently known diversity of the genus Hydnum, with an emphasis on European and North American taxa. Mycologia 2018, 110, 890–918. [Google Scholar] [CrossRef]
- Agerer, R.; Kraigher, H.; Javornik, B. Identification of ectomycorrhizae of Hydnum rufescens on Norway spruce and the variability of the ITS region of H. rufescens and H. repandum (Basidiomycetes). Nova Hedwig. 1996, 63, 183–194. [Google Scholar]
- Moncalvo, J.M.; Nilsson, R.H.; Koster, B.; Dunham, S.M.; Bernauer, T.; Matheny, P.B.; Porter, T.M.; Margaritescu, S.; Weiss, M.; Garnica, S.; et al. The cantharelloid clade: Dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 2006, 98, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Swenie, R.A.; Baroni, T.J.; Matheny, P.B. Six new species and reports of Hydnum (Cantharellales) from eastern North America. MycoKeys 2018, 30, 35–72. [Google Scholar] [CrossRef] [PubMed]
- Yanaga, K.; Sotome, K.; Ushijima, S.; Maekawa, N. Hydnum species producing whitish basidiomata in Japan. Mycoscience 2015, 56, 434–442. [Google Scholar] [CrossRef]
- Hall, D.; Stuntz, D.E. Pileate Hydnaceae of the Puget Sound area. I. White-spored genera: Auriscalpium, Hericium, Dentinum and Phellodon. Mycologia 1971, 63, 937–1098. [Google Scholar] [CrossRef]
- Maas Geesteranus, R.A. Hydnaceous Fungi of the Eastern Old World; North-Holland Publishing Company: Amsterdam, The Netherlands, 1971; pp. 63–77. [Google Scholar]
- Pine, E.M.; Hibbett, D.S.; Donoghue, M.J. Phylogenetic relationships of cantharelloid and clavaroid Homobasidiomycetes based on mitochondrial and nuclear rDNA sequences. Mycologia 1999, 91, 944–963. [Google Scholar] [CrossRef]
- Feng, B.; Wang, X.H.; Ratkowsky, D.; Gates, G.; Lee, S.S.; Grebenc, T.; Yang, Z.L. Multilocus phylogenetic analyses reveal unexpected abundant diversity and significant disjunct distribution pattern of the Hedgehog Mushrooms (Hydnum L.). Sci. Rep. 2016, 6, 25586. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Watling, R.; Noraini-Sikin, Y. Ectomycorrhizal basidiomata fruiting in lowland forests of Peninsular Malaysia. Bois Forêts Trop. 2002, 274, 33–43. [Google Scholar]
- Ostrow, H.; Beenken, L. Hydnum ellipsosporum spec. nov. (Basidiomycetes, Cantharellales)–ein Doppelgänger von Hydnum rufescens Fr. Zeitschrift Für Mykologie 2004, 70, 137–156. [Google Scholar]
- Grebenc, T.; Martín, M.P.; Kraigher, H. Ribosomal ITS diversity among the European species of the genus Hydnum (Hydnaceae). An. Jard. Bot. Madr. 2009, 66, 121–132. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Pine, E.M.; Langer, E.; Langer, G.; Donoghue, M.J. Evolution of gilled mushrooms and puffballs inferred from ribosomal DNA sequences. Proc. Natl. Acad. Sci. USA 1997, 94, 12002–12006. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, R.; Maekawa, N.; Sotome, K.; Nakagiri, A.; Endo, N. Systematic revision of Hydnum species in Japan. Mycologia 2022, 114, 413–452. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Hu, Y.P.; Yu, J.R.; Wei, T.Z.; Yuan, H.S. A phylogenetic overview of the Hydnaceae (Cantharellales, Basidiomycota) with new taxa from China. Stud. Mycol. 2021, 99, 100121. [Google Scholar] [CrossRef]
- Vizzini, A.; Picillo, B.; Ercole, E.; Voyron, S.; Contu, M. Detecting the variability of Hydnum ovoideisporum (Agaricomycetes, Cantharellales) on the basis of Italian collections, and H. magnorufescens sp. nov. Mycosphere 2012, 4, 32–44. [Google Scholar] [CrossRef]
- Olariaga, I.; Grebenc, T.; Salcedo, I.; Martín, M.P. Two new species of Hydnum with ovoid basidiospores: H. ovoideisporum and H. vesterholtii. Mycologia 2012, 104, 1443–1455. [Google Scholar] [CrossRef]
- Buyck, B.; Duhem, B.; Das, K.; Jayawardena, R.S.; Niveiro, N.; Pereira, O.L.; Prasher, I.B.; Adhikari, S.; Albertó, E.O.; Bulgakov, T.S.; et al. Fungal Biodiversity Profiles 21–30. Cryptogam. Myco. 2017, 38, 101–146. [Google Scholar] [CrossRef]
- Sugawara, R.; Yamada, A.; Kawai, M.; Sotome, K.; Maekawa, N.; Nakagiri, A.; Endo, N. Establishment of monokaryotic and dikaryotic isolates of Hedgehog mushrooms (Hydnum repandum and related species) from basidiospores. Mycoscience 2019, 60, 201–209. [Google Scholar] [CrossRef]
- Bi, Z.S.; Zheng, G.Y.; Li, T.H. Macrofungus flora of Guangdong Province; Guangdong Science and Technology Press: Guangzhou, China, 1994. [Google Scholar]
- Zang, M.; Li, B.; Xi, J.X. Fungi of Hengduan Mountains; Science Press: Beijing, China, 1996. [Google Scholar]
- Zhang, M.; Wang, C.; Bai, H.; Deng, W. A Contribution to the Phylogeny and Taxonomy of Hydnum (Cantharellales, Basidiomycota) from China. J. Fungi 2024, 10, 98. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Some new and revised taxa of New Zealand Basidiomycetes (Fungi). N. Z. J. Bot. 1971, 9, 355–370. [Google Scholar] [CrossRef]
- Cooke, M.C. Australian fungi. Grevillea 1890, 19, 44–47. [Google Scholar]
- Massee, G.E. Fungi exotici, XIX. Bull. Misc. Inf. Kew. 1914, 1914, 357–359. [Google Scholar]
- Banker, H.J. A preliminary contribution to a knowledge of the Hydnaceae. Bull. Torrey Bot. Club 1901, 28, 199–222. [Google Scholar] [CrossRef]
- Peck, C.H. Descriptions of new species of New York fungi. Bull. N. Y. St. Mus. Nat. Hist. 1887, 1, 5–24. [Google Scholar]
- Peck, C.H. Report of the State Botanist (1901). Ann. Rep. Reg. N. Y. St. Mus. 1902, 54, 929–984. [Google Scholar]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Hongo, T.; Ueda, T. Yamakei Field Books Kinoko; Yama-Kei Publishers: Tokyo, Japan, 2006. (In Japanese) [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben. 3. Aufl.; Muster-Schmidt Verlag: Göttingen, Germany, 1981; p. 242. [Google Scholar]
- Zhang, Y.Z.; Liang, Z.Q.; Xie, H.J.; Wu, L.L.; Xue, R.; Zeng, N.K. Cantharellus macrocarpus (Cantharellaceae, Cantharellales), a new species from tropical China. Phytotaxa 2021, 484, 170–180. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Lin, W.F.; Buyck, B.; Liang, Z.Q.; Su, M.S.; Chen, Z.H.; Zhang, P.; Jiang, S.; An, D.Y.; Zeng, N.K. Morphological and phylogenetic evidences reveal four new species of Cantharellus subgenus Cantharellus (Hydnaceae, Cantharellales) from China. Front. Microbiol. 2022, 13, 900329. [Google Scholar] [CrossRef]
- Xie, H.J.; Tang, L.P.; Mu, M.; Fan, Y.G.; Jiang, S.; Su, M.S.; Liang, Z.Q.; Zeng, N.K. A contribution to knowledge of Gyroporus (Gyroporaceae, Boletales) in China: Three new taxa, two previous species, and one ambiguous taxon. Mycol. Prog. 2022, 21, 71–92. [Google Scholar] [CrossRef]
- Bas, C. Morphology and subdivision of Amanita and a monograph of its section Lepidella. Persoonia 1969, 5, 285–573. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Kauff, F.; Schoch, C.; Matheny, P.B.; Hofstetter, V.; Cox, C.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of the fungi using a six gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenies. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analyses program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.A.; Dunn, C.W. Phyutility: A phyloinformatics tool for trees, alignments andmolecular data. Bioinformatics 2008, 24, 715–716. [Google Scholar] [CrossRef] [PubMed]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenet. Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Das, K.; Horman, J.; Antonin, V.; Baghela, A.; Chakraborty, D.; Hembrom, M.E.; Nakasone, K.; Ortiz-Santana, B.; Vizzini, A.; et al. Fungal biodiversity profiles 51–60. Cryptogam. Myco. 2018, 39, 211–257. [Google Scholar] [CrossRef]
- Sugawara, R.; Aoki, W.; Yamada, A.; Nakagiri, A.; Endo, N. Ecological speciation of Japanese hedgehog mushroom: Hydnum subalpinum sp. nov. is distinguished from its sister species H. repando-orientale by means of integrative taxonomy. Mycol. Prog. 2022, 21, 94. [Google Scholar] [CrossRef]
- An, D.Y.; Liang, Z.Q.; Jiang, S.; Su, M.S.; Zeng, N.K. Cantharellus hainanensis, a new species with a smooth hymenophore from tropical China. Mycoscience 2017, 58, 438–444. [Google Scholar] [CrossRef]
- Baird, R.; Wallace, L.E.; Baker, G.; Scruggs, M. Stipitate hydnoid fungi of the temperate southeastern United States. Fungal Divers. 2013, 62, 41–114. [Google Scholar] [CrossRef]
- Moeller, H.V.; Peay, K.G.; Fukami, T. Ectomycorrhizal fungal traits reflect environmental conditions along a coastal California edaphic gradient. FEMS Microbiol. Ecol. 2014, 87, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Larsson, E.; Kõljalg, U. Fruiting body-guided molecular identification of root-tip mantle mycelia provides strong indications of ectomycorrhizal associations in two species of Sistotrema (Basidiomycota). Mycol. Res. 2006, 110, 1426–1432. [Google Scholar] [CrossRef] [PubMed]










| Taxon | Voucher | Locality | GenBank/UNITE Accession Nos. | References | |
|---|---|---|---|---|---|
| 28S | ITS | ||||
| Hydnum aerostatisporum | BB 14.154 | USA | — | KY800343 | [18] |
| H. aerostatisporum | RAS071 | USA | — | MH379885 | [4] |
| H. albertense | H T. Niskanen 11-354 | Canada | — | KX388664 | [1] |
| H. albidum | 10640TJB | USA | — | MH379883 | [4] |
| H. alboaurantiacum | RAS186 | USA | — | MH379955 | [4] |
| H. alboaurantiacum | RAS208 | USA | — | MH379937 | [4] |
| H. alboluteum | TNS-F-36545 | Japan | — | LC621797 | [14] |
| H. alboluteum | TUMH 63987 | Japan | — | LC621801 | [14] |
| H. alboluteum | TUMH 63988 | Japan | — | LC621802 | [14] |
| H. alboluteum | TUMH 63989 | Japan | — | LC621803 | [14] |
| H. alboluteum | TUMH 63990 | Japan | — | LC621804 | [14] |
| H. alboluteum | TUMH 63992 | Japan | — | LC621805 | [14] |
| H. albomagnum | AFTOL-ID 471 | USA | AY700199 | DQ218305 | [42] |
| H. albomagnum | RAS231 | USA | — | MH379943 | [4] |
| “H. albomagnum” | IFP 019480 | Hunan, central China | MW979536 | MW980550 | [15] |
| “H. albomagnum” | IFP 019481 | Hunan, central China | MW979537 | MW980551 | [15] |
| H. albopallidum | TUMH 63997 | Japan | LC717904 | LC621807 | [14] |
| H. albopallidum | TUMH 63998 | Japan | — | LC621808 | [14] |
| H. albopallidum | TUMH 63999 | Japan | — | LC621809 | [14] |
| H. berkeleyanum | CAL 1656 | India | NG_070500 | NR_158533 | [43] |
| H. berkeleyanum | HKAS77834 | Hunan, SC China | KU612667 | KU612525 | [9] |
| H. berkeleyanum | IFP 019484 | China | MW979538 | MW980552 | [15] |
| H. boreorepandum | H T. Niemela 1679 | Finland | — | KX388658 | [1] |
| H. boreorepandum | H 6003711 | Finland | — | KX388657 | [1] |
| H. boreorepandum | TUMH 64005 | Japan | LC717880 | LC621814 | [14,44] |
| H. boreorepandum | TUMH 64006 | Japan | LC717881 | LC621815 | [14,44] |
| H. brevispinum | IFP 019464 | Hunan, central China | MW979559 | MW980578 | [15] |
| H. brevispinum | IFP 019465 | Hunan, central China | MW979560 | MW980579 | [15] |
| H. canadense | H T. Niskanen 09-006 | Canada | — | KX388681 | [1] |
| H. cremeoalbum | GDGM87013 | Guangdong, southern China | OR947135 | OR947126 | [22] |
| H. cremeoalbum | GDGM93011 | Hubei, central China | OR947129 | OR947110 | [22] |
| H. cremeoalbum | TUMH60740 | Japan | — | AB906678 | [5] |
| H. cremeoalbum | HKAS92345 | Chongqing, SW China | KU612676 | KU612619 | [9] |
| H. cremeoalbum | TUMH 64024 | Japan | LC717912 | LC621823 | [14,44] |
| H. cremeoalbum | TUMH 64019 | Japan | — | LC621820 | [14] |
| H. cremeoalbum | TUMH 64026 | Japan | — | LC621825 | [14] |
| H. cremeoalbum | N.K. Zeng954 (FHMU2404) | Hainan, southern China | OQ656791 | — | Present study |
| H. cremeoalbum | N.K. Zeng2511 (FHMU1631) | Hainan, southern China | OQ656792 | OQ656784 | Present study |
| H. crocidens | PERTH08072965 | Australia | KU612685 | KU612630 | [9] |
| H. crocidens | PERTH08095981 | Australia | KU612684 | KU612631 | [9] |
| H. cuspidatum | RAS246 | USA | — | MH379944 | [4] |
| H. cuspidatum | RAS205 | USA | — | MH379936 | [4] |
| H. erectum | gui0024 (FHMU7689) | Zhejiang, eastern China | OR722669 | OR722666 | Present study |
| “H. ellipsosporum” | HMJAU5985 | Jilin, NE China | — | KU612602 | [9] |
| H. ellipsosporum | HKAS93254 | Germany | — | KU612604 | [9] |
| “H. ellipsosporum” | HKAS93255 | Germany | — | KU612605 | [9] |
| “H. ellipsosporum” | HKAS56491 | Germany | — | KU612603 | [9] |
| “H. ellipsosporum” | Os5579 | Germany | — | AY817138 | [11] |
| H. ellipsosporum | H T. Niskanen 12-036 | Finland | — | KX388671 | [1] |
| H. ellipsosporum | FD3281 | Switzerland | KX086217 | KX086215 | Unpublished |
| H. ferruginescens | MH16005 | USA | — | MH379905 | [4] |
| H. ferruginescens | RAS229 | USA | — | MH379942 | [4] |
| H. flabellatum | IFP 019459 | Liaoning, NE China | MW979556 | MW980575 | [15] |
| H. flavidocanum | IFP 019460 | Yunnan, SW China | MW979545 | MW980559 | [15] |
| H. flavidocanum | IFP 019461 | Yunnan, SW China | MW979546 | MW980560 | [15] |
| H. ibericum | BIO:Fungi:12330 | Spain | — | HE611086 | [17] |
| H. ibericum | MA-fungi 3457 | Spain | — | AJ547879 | [12] |
| H. itachiharitake | TUMH 64032 | Japan | LC717905 | LC621829 | [14,44] |
| H. itachiharitake | TUMH 64028 | Japan | — | LC621827 | [14] |
| H. itachiharitake | TUMH 64033 | Japan | — | LC621830 | [14] |
| H. jussii | H 6003709 | Finland | — | KX388665 | [1] |
| H. jussii | IFP 019485 | Xinjiang, NW China | MW979539 | MW980553 | [15] |
| H. jussii | IFP 019486 | Xinjiang, NW China | MW979540 | MW980554 | [15] |
| H. longibasidium | IFP 019462 | Hunan, central China | MW979541 | MW980556 | [15] |
| H. longibasidium | IFP 019463 | Hunan, central China | MW979542 | MW980555 | [15] |
| H. longipes | GDGM82458 | Yunnan, SW China | — | OR947121 | [22] |
| H. magnorufescens | HMJAU4677 | Russia | — | KU612550 | [9] |
| H. magnorufescens | 161209 | Slovenia | KU612669 | KU612549 | [9] |
| H. magnorufescens | TO HG2818 | Italy | — | KC293545 | [16] |
| H. melitosarx | GDGM84518 | Sichuan, SW China | OR947136 | OR947117 | [22] |
| H. melitosarx | H T. Niskanen 11-056 | USA | — | KX388683 | [1] |
| H. melitosarx | K 176869 | UK | — | KX388685 | [1] |
| H. melleopallidum | SMI356 | Canada | — | FJ845406 | [14] |
| H. microcarpum | GDGM87902 | Guangdong, southern China | OR947134 | OR947116 | [22] |
| H. microcarpum | GDGM87902-1 | Guangdong, southern China | — | OR947115 | [22] |
| H. minospororufescens | TUMH 64041 | Japan | — | LC621837 | [14] |
| H. minospororufescens | TUMH 64043 | Japan | — | LC621838 | [14] |
| H. minospororufescens | TUMH 64044 | Japan | — | LC621839 | [14] |
| H. minus | TUMH60737 | Japan | AB906675 | [5] | |
| H. minus | IFP 019482 | Hunan, central China | MW979543 | MW980557 | [15] |
| H. minus | IFP 019483 | Hunan, central China | MW979544 | MW980558 | [15] |
| H. minus | TUMH 64050 | Japan | LC717910 | LC621842 | [14,44] |
| H. minus | TUMH 63123 | Japan | — | LC377881 | [14] |
| H. minus | TUMH 64046 | Japan | — | LC621840 | [14] |
| H. minus | N.K. Zeng2819 (FHMU1798) | Yunnan, SW China | KY407528 | KY407533 | [45] |
| H. minus | N.K. Zeng1062 (FHMU2408) | Hainan, southern China | OQ656793 | OQ656785 | Present study |
| H. minus | N.K. Zeng1067 (FHMU7603) | Hainan, southern China | — | OQ656786 | Present study |
| H. mulsicolor | LJU GIS 1336 | Slovenia | — | AJ547885 | [12] |
| H. mulsicolor | REB-341 | USA | — | JX093560 | [46] |
| “H. mulsicolor” | TUMH 63094 | Japan | LC717911 | LC377892 | [19,44] |
| “H. mulsicolor” | TUMH 63078 | Japan | — | LC377890 | [19] |
| H. mulsicolor | TUMH 64009 | Japan | — | LC621843 | [14] |
| H. neorepandum | H T. Niskanen 10-095 | Canada | — | KX388659 | [1] |
| H. neorepandum | H T. Niskanen 10-086 | Canada | — | KX388660 | [1] |
| H. olympicum | H T. Niskanen 09-134 | USA | — | KX388661 | [1] |
| H. olympicum | SAT-10-208-05 | USA | — | MT955159 | Unpublished |
| H. oregonense | HVM61 | USA | — | KF879509 | [47] |
| H. oregonense | PNW-MS g2010502h1-09 | USA | — | AJ534972 | [12] |
| H. orientalbidum | GDGM93480 | Chongqing, SW China | OR947127 | OR947108 | [22] |
| H. orientalbidum | GDGM91301 | Zhejiang, eastern China | OR947130 | OR947111 | [22] |
| H. orientalbidum | TUMH 62998 | Japan | LC717908 | LC377875 | [19,44] |
| H. orientalbidum | TNS-F-35220 | Japan | — | LC621845 | [14] |
| H. orientalbidum | TNS-F-3783 | Japan | — | LC621846 | [14] |
| H. orientalbidum | TUMH 62997 | Japan | — | LC377874 | [14] |
| H. orientalbidum | N.K. Zeng5307 (FHMU6327) | Hainan, southern China | OQ656794 | OQ656787 | Present study |
| H. orientalbidum | N.K. Zeng5308 (FHMU6361) | Hainan, southern China | OQ656795 | OQ656788 | Present study |
| H. orientalbidum | N.K. Zeng5309 (FHMU3153) | Hainan, southern China | OQ656796 | OQ656789 | Present study |
| H. ovoideisporum | BIO:Fungi 12683 | Spain | — | NR_119818 | [48] |
| H. ovoideisporum | 71106 | Slovenia | — | KU612536 | [9] |
| H. pallidocroceum | IFP 019466 | Xinjiang, NW China | MW979554 | MW980568 | [15] |
| H. pallidocroceum | IFP 019467 | Xinjiang, NW China | MW979555 | MW980569 | [15] |
| H. pallidomarginatum | IFP 019468 | Yunnan, SW China | MW979552 | MW980566 | [15] |
| H. pallidomarginatum | IFP 019469 | Yunnan, SW China | MW979553 | MW980567 | [15] |
| H. pinicola | GDGM93020 | Hubei, central China | OR947128 | OR947109 | [22] |
| H. pinicola | GDGM83047 | Yunnan, SW China | OR947137 | OR947119 | [22] |
| H. pinicola | TUMH 64000 | Japan | — | LC621810 | [14] |
| H. pinicola | TUMH 64002 | Japan | — | LC621811 | [14] |
| H. pinicola | TUMH 64003 | Japan | — | LC621812 | [14] |
| H. pinicola | TUMH 64004 | Japan | — | LC621813 | [14] |
| H. quebecense | H T. Niskanen 10-064 | Canada | — | KX388662 | [1] |
| H. quebecense | CN9 | USA | — | MH379881 | [4] |
| H. repando-orientale | TUMH60745 | Japan | — | AB906683 | [5] |
| H. repando-orientale | TUMH 62860 | Japan | LC717900 | LC377883 | [19,44] |
| H. repando-orientale | TUMH 64069 | Japan | LC717903 | LC621873 | [14,44] |
| H. repandum | KR9177 | Germany | — | KU612575 | [9] |
| H. repandum | HKAS93253 | Germany | — | KU612581 | [9] |
| H. repandum | 031209C | Slovenia | — | KU612576 | [9] |
| H. repandum | 031209A | Slovenia | KU612655 | KU612574 | [9] |
| “H. repandum” | HKAS54416 | Jilin, NE China | — | KU612583 | [9] |
| “H. repandum” | HKAS92333 | Sweden | — | KU612582 | [9] |
| “H. repandum” | 420526MF0925 | Hubei, central China | MH141377 | — | Unpublished |
| H. repandum | H 6003710 | Finland | — | NR_164553 | [1] |
| “H. rufescens” | HKAS92339 | Jilin, NE China | KU612658 | KU612537 | [9] |
| H. rufescens | HKAS92338 | Heilongjiang, NE China | KU612659 | KU612538 | [9] |
| “H. rufescens” | HKAS82529 | Tibet, SW China | KU612657 | KU612541 | [9] |
| “H. rufescens” | HKAS92337 | Sweden | KU612656 | KU612539 | [9] |
| H. rufescens | H 6003708 | Finland | — | KX388688 | [1] |
| H. rufescens | H T. Niemela 7839 | Estonia | — | KX388656 | [1] |
| H. sinorepandum | GDGM82445 | Yunnan, SW China | OR947139 | OR947122 | [22] |
| H. sinorepandum | GDGM82382 | Yunnan, SW China | OR947141 | OR947124 | [22] |
| H. slovenicum | LJU GIS 1338 | Slovenia | — | AJ547870 | [12] |
| H. slovenicum | LJU GIS 1340 | Slovenia | — | AJ547884 | [12] |
| H. sp. | F1110834 | Costa Rica | — | KU612598 | [9] |
| H. sp. | F1104787 | USA | — | KU612606 | [9] |
| H. sp. | FRI62832 | Malaysia | — | KU612625 | [9] |
| H. sp. | HKAS57385 | Yunnan, SW China | — | KU612601 | [9] |
| H. sp. | HKAS61337 | Hunan, central China | KU612644 | KU612597 | [9] |
| H. sp. | HKAS82411 | Taiwan, SE China | KU612668 | KU612607 | [9] |
| H. sp. | TM070 | Canada | DQ898744 | — | [3] |
| H. sp. | wi1A4spel | Taiwan, SE China | — | KC679833 | Unpublished |
| H. sp. | wi8T4spel | Taiwan, SE China | — | KC679834 | Unpublished |
| H. sp. | 231109 | Slovenia | KU612660 | KU612545 | [9] |
| H. sp. | F1187537 | Canada | KU612544 | [9] | |
| H. sp. | HKAS92340 | Heilongjiang, NE China | KU612661 | KU612543 | [9] |
| H. sp. | HKAS56128 | Yunnan, SW China | — | KU612526 | [9] |
| H. sp. | HKAS56881 | Yunnan, SW China | — | KU612530 | [9] |
| H. sp. | HKAS57475 | Yunnan, SW China | KU612666 | KU612528 | [9] |
| H. sp. | HKAS61795 | Heilongjiang, NE China | KU612665 | KU612531 | [9] |
| H. sp. | HKAS77834 | Yunnan, SW China | KU612667 | KU612525 | [9] |
| H. sp. | HKAS74602 | Yunnan, SW China | — | KU612532 | [9] |
| H. sp. | F1125199 | USA | — | KU612534 | [9] |
| H. sp. | F1188749 | USA | KU612663 | KU612535 | [9] |
| H. sp. | F1186911 | USA | KU612662 | KU612548 | [9] |
| H. sp. | HKAS82508 | Tibet, SW China | KU612670 | KU612546 | [9] |
| H. sp. | HKAS45769 | Tibet, SW China | — | KU612547 | [9] |
| H. sp. | HKAS55319 | Yunnan, SW China | — | KU612590 | [9] |
| H. sp. | HKAS51088 | Yunnan, SW China | — | KU612587 | [9] |
| H. sp. | HKAS78334 | Guangdong, southern China | — | KU612589 | [9] |
| H. sp. | HKAS55410 | Yunnan, SW China | KU612654 | KU612596 | [9] |
| H. sp. | HKAS55449 | Yunnan, SW China | — | KU612594 | [9] |
| H. sp. | HKAS82558 | Sichuan, SW China | — | KU612595 | [9] |
| H. sp. | F1188765 | USA | KU612653 | KU612599 | [9] |
| H. sp. | HKAS55327 | Yunnan, SW China | KU612682 | KU612568 | [9] |
| H. sp. | HKAS56789 | Yunnan, SW China | KU612680 | KU612571 | [9] |
| H. sp. | HKAS82410 | Taiwan, SE China | KU612679 | KU612573 | [9] |
| H. sp. | HKAS93261 | Yunnan, SW China | KU612681 | KU612567 | [9] |
| H. sp. | F1185236 | USA | — | KU612600 | [9] |
| H. sp. | HKAS93258 | Hainan, southern China | KU612677 | KU612618 | [9] |
| H. sp. | HKAS92345 | Chongqing, SW China | KU612676 | KU612619 | [9] |
| H. sp. | HKAS57714 | Yunnan, SW China | KU612673 | KU612617 | [9] |
| H. sp. | HKAS58838 | Yunnan, SW China | KU612675 | KU612616 | [9] |
| H. sp. | HKAS55325 | Yunnan, SW China | — | KU612613 | [9] |
| H. sp. | HKAS93259 | Yunnan, SW China | KU612683 | KU612612 | [9] |
| H. sp. | HKAS92336 | Yunnan, SW China | — | KU612614 | [9] |
| H. sp. | HKAS52807 | Yunnan, SW China | — | KU612609 | [9] |
| H. sp. | HKAS92350 | Guizhou, SW China | KU612672 | KU612610 | [9] |
| H. sp. | HKAS92347 | Chongqing, SW China | KU612652 | — | [9] |
| H. sp. | HKAS92349 | Guizhou, SW China | KU612671 | — | [9] |
| H. sp. | TUMH 64099 | Japan | — | LC621899 | [14] |
| H. sp. | TUMH 64100 | Japan | — | LC621900 | [14] |
| H. sphaericum | IFP 019470 | Hunan, central China | MW979549 | MW980563 | [15] |
| H. sphaericum | IFP 019471 | Hunan, central China | MW979551 | MW980565 | [15] |
| H. sphaericum | IFP 019472 | Hunan, central China | MW979550 | MW980564 | [15] |
| H. subalpinum | TUMH 64013 | Japan | LC717888 | LC717913 | [44] |
| H. subalpinum | TUMH 64016 | Japan | LC717891 | LC621871 | [14,44] |
| H. subalpinum | TUMH 64017 | Japan | LC717892 | LC621872 | [14,44] |
| H. subalpinum | TNS-F-85326 | Japan | LC717885 | LC621866 | [14,44] |
| H. subcremeoalbum | TU110688 | Papua New Guinea | — | UDB013289 * | [1] |
| “H. subberkeleyanum” | TNS-F-19323 | Japan | — | LC621879 | [14] |
| “H. subberkeleyanum” | TUMH 62863 | Japan | — | LC377889 | [14] |
| “H. subberkeleyanum” | TUMH 64075 | Japan | — | LC621881 | [14] |
| “H. subberkeleyanum” | TUMH 64081 | Japan | — | LC621882 | [14] |
| H. subconnatum | RAS235 | USA | — | MH379930 | [4] |
| H. subconnatum | RAS169 | USA | — | MH379916 | [4] |
| H. submulsicolor | H T. Niskanen 10-132 | Canada | — | KX388682 | [1] |
| H. subolympicum | DAOM:A. Voitk 12.09.02.av12 | Canada | — | MH174257 | [1] |
| H. subovoideisporum | H 6003707 | Finland | — | NR_158494 | [1] |
| H. subrufescens | H T. Niskanen 10-154 | Canada | — | KX388649 | [1] |
| H. subtilior | RAS180 | USA | — | MH379918 | [4] |
| H. subtilior | TENN 073034 | USA | — | NR_164029 | [4] |
| H. tangerinum | IFP 019473 | Hunan, central China | MW979561 | MW980580 | [15] |
| H. tangerinum | IFP 019474 | Hunan, central China | MW979562 | MW980581 | [15] |
| H. tangerinum | IFP 019475 | Hunan, central China | MW979563 | MW980582 | [15] |
| H. tenuistipitum | IFP 019476 | Hunan, central China | MW979557 | MW980576 | [15] |
| H. tenuistipitum | IFP 019477 | Hunan, central China | MW979558 | MW980577 | [15] |
| H. tenuistipitum | N.K. Zeng7540 (FHMU7644) | Hunan, central China | OQ913756 | OQ913759 | Present study |
| H. tenuistipitum | N.K. Zeng7555 (FHMU7636) | Hunan, central China | OQ913757 | OQ913760 | Present study |
| H. tenuistipitum | N.K. Zeng7558 (FHMU7642) | Hunan, central China | OQ913758 | OQ913761 | Present study |
| H. tomaense | TUMH 64086 | Japan | LC717907 | LC621885 | [14,44] |
| H. tomaense | TUMH 64085 | Japan | — | LC621884 | [14] |
| H. tomaense | TUMH 64087 | Japan | — | LC621886 | [14] |
| H. tottoriense | TUMH 64090 | Japan | — | LC621889 | [14] |
| H. tottoriense | TUMH 64091 | Japan | — | LC621890 | [14] |
| H. tottoriense | TUMH 64092 | Japan | — | LC621891 | [14] |
| H. treui | TU110403 | Papua New Guinea | — | UDB013043 * | [1] |
| H. treui | N.K. Zeng8373 (FHMU7690) | Hainan, southern China | OR722670 | OR722667 | Present study |
| H. treui | N.K. Zeng8374 (FHMU7691) | Hainan, southern China | OR722671 | OR722668 | Present study |
| H. umbilicatum | CORT:012241 | USA | — | MH379890 | [4] |
| “H. umbilicatum” | HKAS92335 | USA | KU612678 | KU612608 | [9] |
| “H. umbilicatum” | TUMH 63128 | Japan | LC717909 | LC377891 | [19,44] |
| H. umbilicatum | TUMH 64093 | Japan | — | LC621893 | [14] |
| H. umbilicatum | TUMH 64094 | Japan | — | LC621894 | [14] |
| H. umbilicatum | TUMH 64098 | Japan | — | LC621898 | [14] |
| H. vagabundum | CLO4985 | USA | — | MH379909 | [4] |
| H. vagabundum | CORT:014461 | USA | — | MH379949 | [4] |
| H. ventricosum | IFP 019478 | Liaoning, NE China | MW979547 | MW980561 | [9] |
| H. ventricosum | IFP 019479 | Liaoning, NE China | MW979548 | MW980562 | [9] |
| H. vesterholtii | BIO:Fungi:10429 | Spain | — | HE611084 | [9] |
| H. vesterholtii | BIO:Fungi:10452 | Spain | — | HE611085 | [9] |
| H. vesterholtii | BIO:Fungi:12904 | France | — | HE611087 | [9] |
| “H. vesterholtii” | HKAS56213 | Yunnan, SW China | — | KU612554 | [9] |
| “H. vesterholtii” | HKAS55399 | Yunnan, SW China | — | KU612555 | [9] |
| “H. vesterholtii” | HKAS57047 | Yunnan, SW China | — | KU612558 | [9] |
| “H. vesterholtii” | HKAS58734 | Yunnan, SW China | — | KU612559 | [9] |
| “H. vesterholtii” | HKAS51086 | Yunnan, SW China | KU612650 | KU612557 | [9] |
| “H. vesterholtii” | HKAS92341 | Shaanxi, NW China | KU612647 | KU612562 | [9] |
| “H. vesterholtii” | HKAS92342 | Yunnan, SW China | KU612646 | KU612564 | [9] |
| “H. vesterholtii” | HKAS92343 | Sichuan, SW China | KU612648 | KU612563 | [9] |
| “H. vesterholtii” | HKAS92344 | Heilongjiang, NE China | KU612649 | KU612556 | [9] |
| “H. vesterholtii” | HKAS77884 | Hunan, central China | KU612645 | KU612565 | [9] |
| “H. vesterholtii” | HKAS56489 | Germany | — | KU612561 | [9] |
| “H. vesterholtii” | HKAS92351 | Guizhou, SW China | KU612651 | — | [9] |
| H. washingtonianum | UBC F-32538 | Canada | — | MF954990 | Unpublished |
| H. washingtonianum | WTU:014341 | USA | — | MH379846 | [4] |
| H. zongolicense | GO-2010-142a | Mexico | — | KC152121 | [1] |
| Sistotrema muscicola | taxon:154757 | Finland | AJ606041 | AJ606041 | [49] |
| S. muscicola | KHL 11721 | Finland | AJ606040 | AJ606040 | [49] |
| Subgenus | Section | Subsection | Species | Locality | References |
|---|---|---|---|---|---|
| Alba Niskanen & Liimat. | — | — | H. cremeoalbum | Japan | [5] |
| H. flavidocanum | Yunnan, SW China | [15] | |||
| H. pinicola | Japan | [14] | |||
| H. treui | Papua New Guinea | [1] | |||
| Brevispina T. Cao & H. S. Yuan | — | — | H. brevispinum | Hunan, central China | [15] |
| H. minus | Japan | [5] | |||
| Hydnum L. | Hydnum L. | — | H. sinorepandum | Yunnan Province, SW China | [22] |
| H. sphaericum | Hunan, central China | [15] | |||
| Pallida Niskanen & Liimat. | — | — | H. erectum | Zhejiang, eastern China | Present study |
| H. flabellatum | Liaoning, NE China | [15] | |||
| H. pallidomarginatum | Yunnan, SW China | [15] | |||
| Rufescentia Niskanen & Liimat. | Magnorufescentia Niskanen & Liimat. | — | H. pallidocroceum | Xinjiang, NW China | [15] |
| H. tangerinum | Hunan, central China | [15] | |||
| Mulsicoloria Niskanen & Liimat. | H. longipes | Yunnan Province, SW China | [22] | ||
| H. melitosarx | USA | [1] | |||
| Rufescentia Niskanen & Liimat. | Rufescentia Niskanen & Liimat. | H. ventricosum | Liaoning, NE China | [15] | |
| Tenuiformia Niskanen & Liimat. | H. longibasidium | Hunan, central China | [15] | ||
| — | — | — | H. microcarpum | Guangdong Province, southern China | [22] |
| H. orientalbidum | Japan | [14] | |||
| H. tenuistipitum | Hunan, central China | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, H.-Z.; Liao, Y.-T.; Zhang, Y.-Z.; Lin, W.-F.; Yang, X.-Q.; Zeng, N.-K. A Contribution to the Knowledge of Hydnum (Hydnaceae, Cantharellales) in China, Introducing a New Taxon and Amending Descriptions of Five Known Species. Diversity 2024, 16, 166. https://doi.org/10.3390/d16030166
Qin H-Z, Liao Y-T, Zhang Y-Z, Lin W-F, Yang X-Q, Zeng N-K. A Contribution to the Knowledge of Hydnum (Hydnaceae, Cantharellales) in China, Introducing a New Taxon and Amending Descriptions of Five Known Species. Diversity. 2024; 16(3):166. https://doi.org/10.3390/d16030166
Chicago/Turabian StyleQin, Hua-Zhi, Yu-Ting Liao, Yu-Zhuo Zhang, Wen-Fei Lin, Xin-Quan Yang, and Nian-Kai Zeng. 2024. "A Contribution to the Knowledge of Hydnum (Hydnaceae, Cantharellales) in China, Introducing a New Taxon and Amending Descriptions of Five Known Species" Diversity 16, no. 3: 166. https://doi.org/10.3390/d16030166