Morphology, DNA Phylogeny, and Pathogenicity of Wilsonomyces carpophilus Isolate Causing Shot-Hole Disease of Prunus divaricata and Prunus armeniaca in Wild-Fruit Forest of Western Tianshan Mountains, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Shot-Hole Disease Investigation
2.2. Sampling and Isolation
2.3. Morphological Identification
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Phylogenetic Analysis
2.6. Pathogenicity Tests
3. Results
3.1. Field Symptoms and Isolating the Fungi
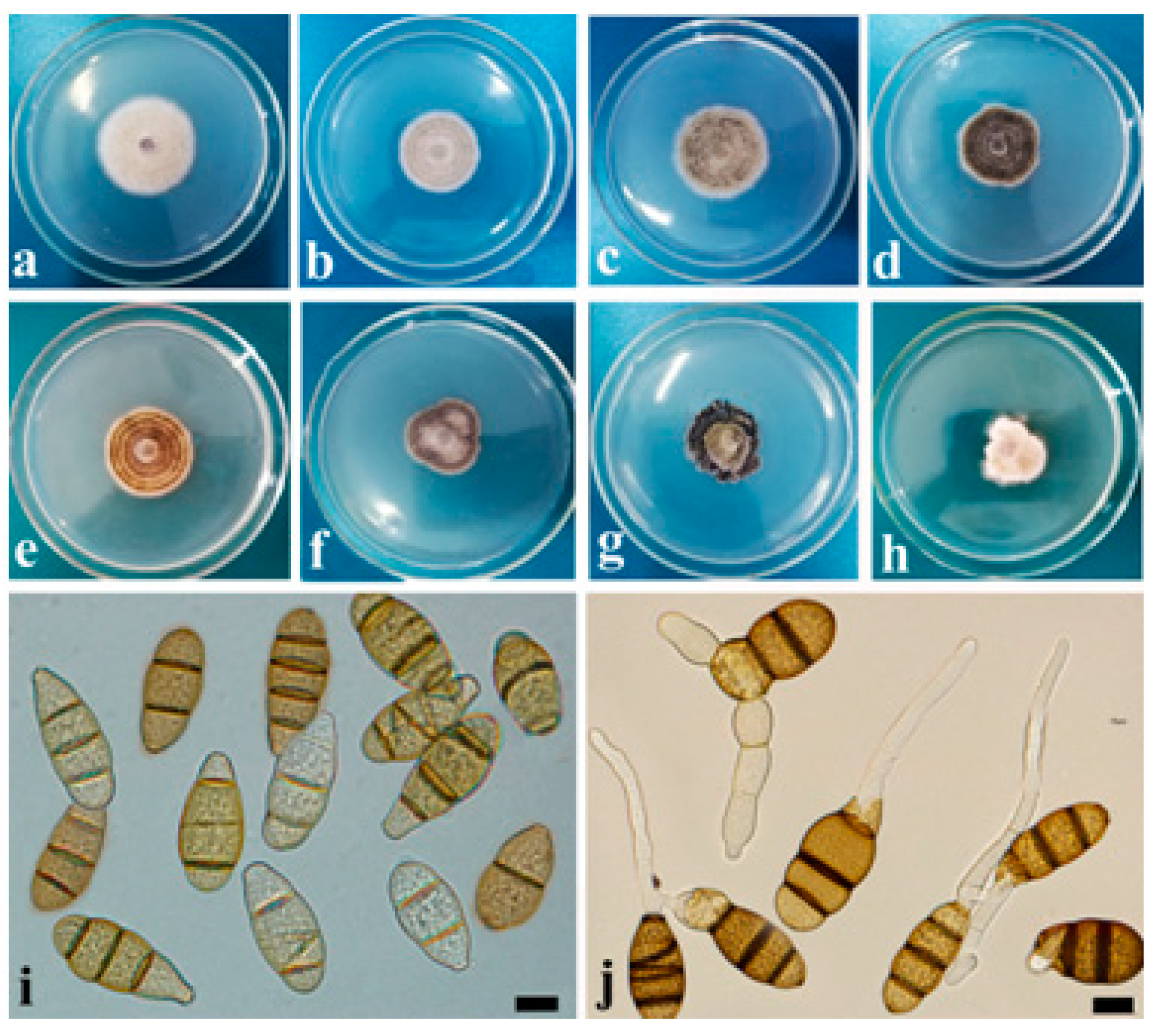
3.2. Cultural and Morphological Characteristics
3.3. Phylogenetic Analysis
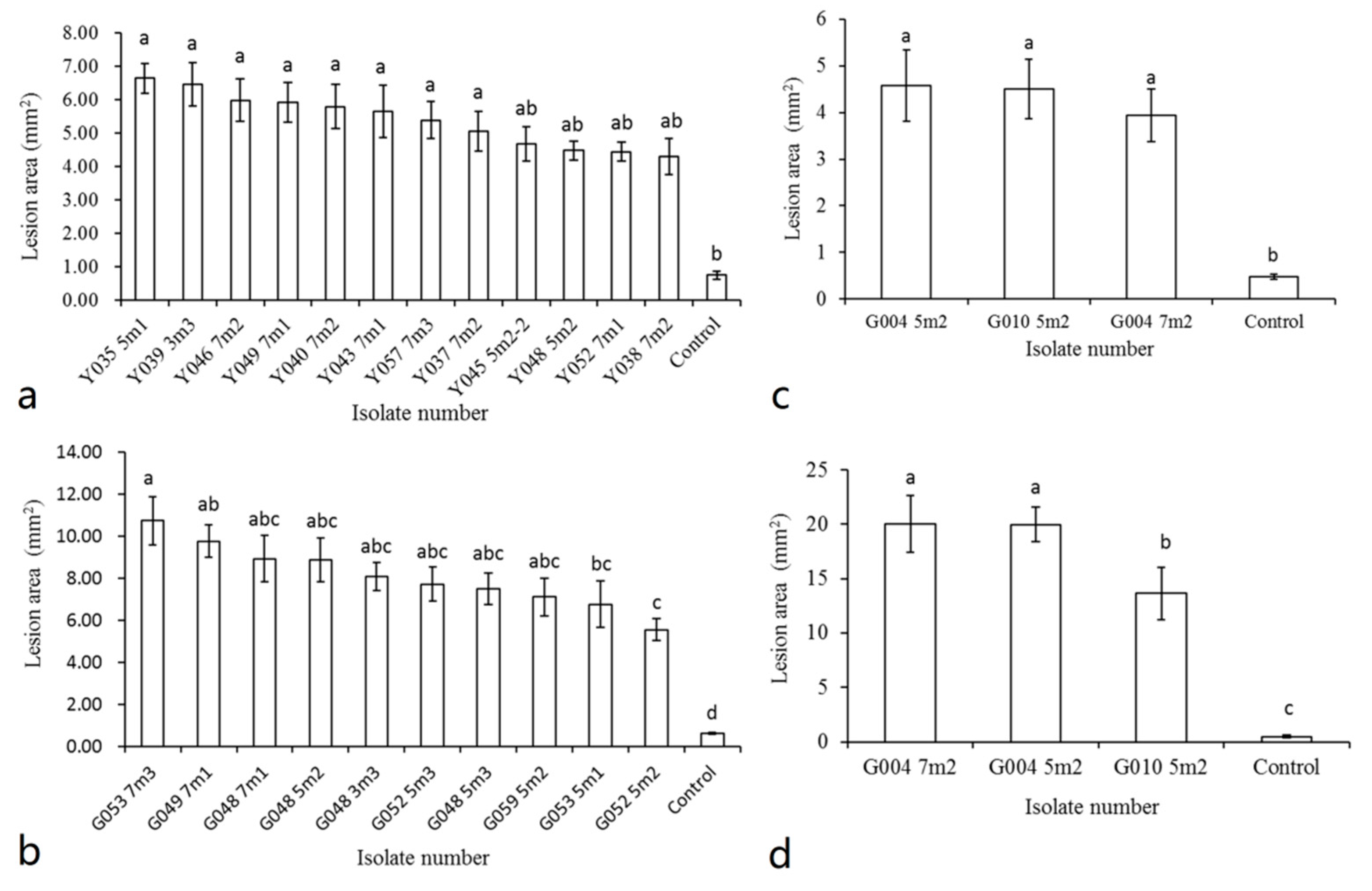
3.4. Pathogenicity Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liao, K. Research on Resources of Wild Fruit Tree in Xinjiang; Xijiang People’s Publishing House: Urumqi, China, 2013. (In Chinese) [Google Scholar]
- Qiu, Y.X.; Lu, Q.X.; Zhang, Y.H.; Cao, Y.N. Phylogeography of East Asia’s Tertiary relict plants: Current progress and future prospects. Biodivers. Sci. 2017, 25, 136–146. (In Chinese) [Google Scholar] [CrossRef] [Green Version]
- Reales, A.; Sargent, D.J.; Tobut, K.R.; Rivera, D. Phylogenetics of Eurasian plums, Prunus L. section Prunus (Rosaceae), according to coding and non-coding chloroplast DNA sequences. Tree Genet. Genomes 2010, 6, 37–45. [Google Scholar] [CrossRef]
- WÖHrmann, T.; Guicking, D.; Khoshbakht, K.; Weising, K. Genetic variability in wild populations of Prunus divaricata Ledeb. in northern Iran evaluated by EST-SSR and genomic SSR marker analysis. Genet. Resour. Crop Evol. 2011, 58, 1157–1167. [Google Scholar] [CrossRef]
- Zhou, L.; Liao, K.; Xu, Z.; Wang, L. Advance in research on germplasm resources in Prunus cerasifera Ehrh. Nonwood For. Res. 2010, 28, 142–145. (In Chinese) [Google Scholar]
- Zhang, S.X. On the Eco-geographical characters and the problems of classification of the Wild Fruit-tree Forest in the Yili Valley of Xinjiang. Acta Bot. Sin. 1973, 36, 39–41. (In Chinese) [Google Scholar]
- Tanwar, B.; Modgil, R.; Goyal, A. Effect of detoxification on biological quality of wild apricot (Prunus armeniaca L.) kernel: Biological quality of detoxified wild apricot kernel. J. Sci. Food Agric. 2019, 99, 517–528. [Google Scholar] [CrossRef]
- Cao, Q. Study on the Genetic Diversity of Daxigou Wild Apricot in Huocheng; Urumqi, Xinjiang Agricultural University: Yili, China, 2016. (In Chinese) [Google Scholar]
- Lu, C.Y. Genetic Diversity Based on SSR and Its Association Analysis with Phenotypic Traits in Prunus armeniaca Var. Ansu; Shenyang Agricultural University: Shenyang, China, 2018. (In Chinese) [Google Scholar]
- Tovar-Pedraza, J.M.; Ayala-Escobar, V.; Segura-León, O.L. Thyrostroma carpophilum causing apricot shot-hole in Mexico. Australas. Plant Dis. Notes 2013, 8, 31–33. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.R.; Zhang, L.Y.; Xu, Z. Damage status and protection of Wild Fruit Forest Ecosystem in Tianshan Mountains. Study Arid Areas 1999, 16, 1–4. (In Chinese) [Google Scholar]
- An, X.L.; Zhou, Y. Resource conservation and rational development and utilization of Wild Fruit Forest in Yili, Xinjiang. J. Jiangsu For. Sci. Technol. 2009, 36, 39–41. (In Chinese) [Google Scholar]
- Bubici, G.; Margherita, D.A.; Cirulli, M. Field reactions of plum cultivars to the shot-hole disease in southern Italy. Crop Prot. 2010, 29, 1396–1400. [Google Scholar] [CrossRef]
- Abolfazl, N.; Teponno, R.B.; Mahdi, A.; Asadollah, B.A.; Marc, S. New secondary metabolites produced by the phytopathogenic fungus, Wilsonomyces carpophilus. Phytochem. Lett. 2018, 26, 212–217. [Google Scholar]
- Grantina-Ievina, L.; Stanke, L. Incidence and severity of leaf and fruit diseases of plums in Latvia. Commun. Agric. Appl. Biol. Sci. 2015, 80, 421–433. [Google Scholar] [PubMed]
- He, S.Q.; Bai, W.; Wen, C.H.; Jin, Z.Q. Identification of the pathogen of the brown spot of peach fruit in Gansu, China. Plant Prot. 2016, 42, 53–57. (In Chinese) [Google Scholar]
- Dar, R.A.; Rai, A.N.; Shiekh, I.A. Stigmina carpophila detected on Prunus armeniaca and Prunus persicain India. Australas. Plant Dis. Notes 2017, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Dai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979; pp. 914–934. (In Chinese) [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China; Mycotaxon, Ltd.: Ithaca, NY, USA, 2005; p. 158. [Google Scholar]
- Zhu, J.L.; Chang, Y.Y.; Chen, D.R.; Kang, Z.Y. Study on Prunus armeniaca fruits spot disease. Gansu Agric. Sci. Technol. 1995, 9, 38–40. [Google Scholar]
- Adaskaveg, J.E.; Ogawa, J.M.; Butler, E.E. Morphology and ontogeny of conidia in Wilsonomyces carpophilus, gen. nov. and comb. nov., causal pathogen of shot hole disease of Prunus species. Mycotaxon 1990, 37, 275–290. [Google Scholar]
- Ahmadpour, A.; Ghosta, Y.; Javan-Nikkhah, M.; Fatahi, R.; Ghazanfari, K. Isolation and pathogenicity tests of Iranian cultures of the shot hole pathogen of Prunus species, Wilsonomyces carpophilus. Australas. Plant Dis. Notes 2009, 4, 133–134. [Google Scholar]
- Jr, I.M.; Tripon, A.; Mitre, I.; Mitre, V. The response of several plum cultivars to natural infection with Monilinia laxa, Polystigma rubrum and Stigmina carpophila. Pediatr. Rehabil. 2015, 7, 219–228. [Google Scholar]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Crous, P.W. Genera of phytopathogenic fungi: GOPGY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [Green Version]
- Sutton, B.C. On Stigmina, Wilsonomyces and Thyrostroma (Hyphomycetes). Arnoldia 1997, 14, 33–35. [Google Scholar]
- Adaskaveg, J.E. Conidial morphology, host colonization, and development of shot hole of almond caused by Wilsonomyces carpophilus. Can. J. Bot. 1995, 73, 432–444. [Google Scholar] [CrossRef]
- Lan, Z.; Scherm, H. Moisture sources in relation to conidial dissemination and infection by Cladosporium carpophilum within peach canopies. Phytopathology 2003, 93, 1581–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalancette, N.; Mcfarland, K.A.; Burnett, A.L. Modeling sporulation of Fusicladium carpophilum on nectarine twig lesions: Relative humidity and temperature effects. Phytopathology 2012, 102, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Sang, J.P.; Zhao, Z.Z.; Guo, H.L. Distributional patterns of forest pathogens fungi in Xiaolongshan forest area. J. Gansu Agric. Univ. 2013, 48, 102–105. (In Chinese) [Google Scholar]
- Rasmus, E.; Berthold, M.; Uli, R.; Gerald, K. Ash Dieback on Sample Points of the National Forest Inventory in South-Western Germany. Forests 2018, 9, 25. [Google Scholar]
- Huang, L.; Zhu, Y.N.; Yang, J.Y.; Li, D.W.; Li, Y.; Bian, L.M.; Ye, J.R. Shoot blight on Chinese fir (Cunninghamia lanceolata) is caused by Bipolaris oryzae. Plant Dis. 2018, 102, 500–506. [Google Scholar] [CrossRef]
- Teviotdale, B.L.; Goodell, N.; Harper, D. Abscission and kernel quality of almond fruit inoculated with the shot hole pathogen Wilsonomyces carpophilus. Plant Dis. 1997, 81, 1454–1458. [Google Scholar] [CrossRef] [Green Version]
- Grove, G.G. Influence of temperature and wetness period on infection of cherry and peach foliage by Wilsonomyces carpophilus. Can. J. Plant Pathol. 2002, 24, 40–45. [Google Scholar] [CrossRef]
- Saito, S.; Margosan, D.; Michailides, T.J.; Xiao, C.L. Botrytis californica, a new cryptic species in the B. cinerea species complex causing gray mold in blueberries and table grapes in California. Mycologia 2016, 108, 330–343. [Google Scholar] [CrossRef]
- Zhang, Q.T.; He, M. Canker on bark of Populus spp. caused by Cytospora tritici, a new disease in China. Plant Dis. 2012, 96, 1578. [Google Scholar] [CrossRef]
- Solarte, F.A.; Muñoz, C.G.; Maharachchikumbura, S.; Álvarez, E. Diversity of Neopestalotiopsis and Pestalotiopsis spp., causal agents of guava scab in Colombia. Plant Dis. 2017, 102, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Ye, H.; Lu, J.; Chen, P.; Yu, Z.F. Isolation and identification of Sydowia polyspora and its pathogenicity on Pinus yunnanensis in Southwestern china. J. Phytopathol. 2018, 166, 386–395. [Google Scholar] [CrossRef]
- Aigoun-Mouhous, W.; Elena, G.; Cabral, A.; León, M.; Sabaou, N.; Armengol, J. Characterization and pathogenicity of Cylindrocarpon-like asexual morphs associated with black foot disease in Algerian grapevine nurseries, with the description of Pleiocarpon algeriense sp. nov. Eur. J. Plant Pathol. 2019, 154, 887–901. [Google Scholar] [CrossRef]
- Ma, R.; Ye, S.H.; Zhao, Y.; Michailides, T.J.; Tian, C.M. New leaf and fruit disease of Juglans regia caused by Juglanconis juglandina in Xinjiang, China. For. Pathol. 2019, 49, e12537. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S. Colletotrichum species associated with anthracnose of pyrus spp. in China. Pers. Mol. Phylogeny Evol. Fungi 2018, 42, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Du, Z.; Liang, Y.M.; Tian, C.M. Melanconis, (Melanconidaceae) associated with Betula spp. Mycol. Prog. 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Ma, R.; Liu, Y.M.; Yin, Y.X.; Tian, C.M. A canker disease of apple caused by Cytospora parasitica recorded in China. For. Pathol. 2018, 48, e12416. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- O’Donnell, K. Fusarium and Its Near Relatives; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A. Morphological and molecular data reveal cryptic species in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Fan, X.L.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Ma, R.; Tian, C.M. Cytospora species associated with canker disease of three anti-desertification plants in northwestern China. Phytotaxa 2015, 197, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony, * and Other Methods. Version 456 4.0b10; Sinauer Associates Inc.: Sunderland, UK, 2003. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.N.; Zhao, Y.; Tian, C.M.; Michailides, T.J.; Ma, R. A new species and a new record of Thyronectria (Nectriaceae, Hypocreales) in china. Phytotaxa 2018, 376, 17–26. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Nouri, M.T.; Lawrence, D.P.; Yaghmour, M.A.; Michailides, T.J.; Trouillas, F.P. Neoscytalidium dimidiatum causing canker, shoot blight and fruit rot of almond in California. Plant Dis. 2018, 102, 1638–1647. [Google Scholar] [CrossRef] [Green Version]
- Ivan, M.; Nenad, K.; Dragan, K.; Zlatan, R.; Justyna, N.; Tomasz, O.; Katarzyna, S.; Tamara, C.; Thomas, J. Isolation and pathogenicity of phytophthora species from poplar plantations in serbia. Forests 2018, 9, 330. [Google Scholar]
- Omar, N.H.; Mohd, M.; Nor, N.M.I.M.; Zakaria, L. Characterization and pathogenicity of Fusarium species associated with leaf spot of mango (Mangifera indica L.). Microb. Pathog. 2018, 114, 362–368. [Google Scholar] [CrossRef]
- Florent, P.T.; Mohamed, T.N.; Daniel, P.L.; Juan, M.; Renaud, T.; Brenna, J.A.; Danielle, L. Identification and Characterization of Neofabraea kienholzii and Phlyctema vagabunda Causing Leaf and Shoot Lesions of Olive in California. Plant Dis. 2019, 103, 3018–3030. [Google Scholar]
- Lu, Y.; Zhang, Y.; Shen, Y.C.; Xu, W.F.; Wang, K.F.; Pan, B.; Gong, W.L.; Wang, Y.H. Several pesticides control effect on peach bacterial shot hole and their residue analysis. Agrochemicals 2016, 55, 287–289. (In Chinese) [Google Scholar]
- Wang, C.A. Isolation and Identification of Pathogen of Peach Shot-Hole Disease; Chinese Academy of Agricultural Sciences Dissertation: Beijing, China, 2018. (In Chinese) [Google Scholar]
- Chen, Y.; Huai, W.X.; Yao, Y.X.; Lin, R.Z.; Liu, Z.J.; Zhao, W.X. The pathogen identification of apricot fruit spots disease in Gongliu County, Xinjiang. For. Res. 2019, 32, 121–126. (In Chinese) [Google Scholar]
- Camele, I.; Lo Cantore, P.; Lasco, R.; Iacobellis, N.S. Xanthomonas arboricola pv. pruni susino in Basilicata. L’informatore Agrar. 2001, 57, 20–689. [Google Scholar]
- Nabi, A.; Shah, M.U.D.; Padder, B.A.; Dar, M.S.; Ahmad, M. Morpho-cultural, pathological and molecular variability in Thyrostroma carpophilum, causing shot hole of stone fruits in India. Eur. J. Plant Pathol. 2017, 151, 613–627. [Google Scholar] [CrossRef]
- Meena, B.S. Morphological and Molecular Variability of Rice Blast Pathogen Pyricularia grisea (Cooke) Sacc. Master’s Thesis, University of Agricultural Sciences, Dharwad, India, 2005. [Google Scholar]
- Torres-Calzada, C.; Tapia-Tussell, R.; Higuera-Ciapara, I.; Perez-Brito, D. Morphological, pathological and genetic diversity of Colletotrichum species responsible for anthracnose in papaya (Carica papaya L.). Eur. J. Plant Pathol. 2013, 135, 67–79. [Google Scholar] [CrossRef]
- Ahmadpour, A.; Ghosta, Y.; Javan-Nikkhah, M.; Ghazanfari, K.; Fatahi, R. Study on morphology, pathogenicity and genetic diversity of Wilsonomyces carpophilus isolates, the causal agent of shot hole of stone fruit trees based on RAPD-PCR in Iran. Arch. Phytopathol. Plant Prot. 2012, 45, 2076–2086. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate no. | Host | Substrate | Location | GenBank Accession Numbers2 | ||
|---|---|---|---|---|---|---|
| ITS | LSU | tef1 | ||||
| XJAU Y035 5m1 | Prunus armeniaca | Leaf | Huocheng | MN817623 | MN817648 | MN817598 |
| XJAU Y037 7m2 | P. armeniaca | Leaf | Xinyuan | MN817624 | MN817649 | MN817599 |
| XJAU Y038 7m2 | P. armeniaca | Leaf | Xinyuan | MN817625 | MN944917 | MN817600 |
| XJAU Y039 3m3 | P. armeniaca | Leaf | Xinyuan | MN817626 | MN817650 | MN817601 |
| XJAU Y040 7m2 | P. armeniaca | Leaf | Xinyuan | MN817627 | MN817651 | MN817602 |
| XJAU Y043 7m1 | P. armeniaca | Leaf | Yining | MN817628 | MN817652 | MN817603 |
| XJAU Y045 5m2-2 | P. armeniaca | Leaf | Yining | MN817629 | MN817653 | MN817604 |
| XJAU Y046 7m2 | P. armeniaca | Leaf | Yining | MN817630 | MN944918 | MN817605 |
| XJAU Y048 5m2 | P. armeniaca | Leaf | Huocheng | MN817631 | MN817654 | MN817606 |
| XJAU Y049 7m1 | P. armeniaca | Leaf | Huocheng | MN817632 | MN817655 | MN817607 |
| XJAU Y052 7m1 | P. armeniaca | Leaf | Huocheng | MN817633 | MN817656 | MN817608 |
| XJAU Y057 7m3 | P. armeniaca | Leaf | Gongliu | MN817634 | MN817657 | MN817609 |
| XJAU G048 3m3 | P. armeniaca | Fruit | Huocheng | MN817613 | MN817638 | MN817588 |
| XJAU G048 5m2 | P. armeniaca | Fruit | Huocheng | MN817614 | MN817639 | MN817589 |
| XJAU G048 5m3 | P. armeniaca | Fruit | Huocheng | MN817615 | MN817640 | MN817590 |
| XJAU G048 7m1 | P. armeniaca | Fruit | Huocheng | MN817616 | MN817641 | MN817591 |
| XJAU G049 7m1 | P. armeniaca | Fruit | Huocheng | MN817617 | MN817642 | MN817592 |
| XJAU G052 5m2 | P. armeniaca | Fruit | Huocheng | MN817618 | MN817643 | MN817593 |
| XJAU G052 5m3 | P. armeniaca | Fruit | Huocheng | MN817619 | MN817644 | MN817594 |
| XJAU G053 5m1 | P. armeniaca | Fruit | Huocheng | MN817620 | MN817645 | MN817595 |
| XJAU G053 7m3 | P. armeniaca | Fruit | Huocheng | MN817621 | MN817646 | MN817596 |
| XJAU G059 5m2 | P. armeniaca | Fruit | Xinyuan | MN817622 | MN817647 | MN817597 |
| XJAU G004 5m2 | Prunus divaricata | Fruit | Huocheng | MN817610 | MN817635 | MN817585 |
| XJAU G004 7m2 | P. divaricata | Fruit | Huocheng | MN817611 | MN817636 | MN817586 |
| XJAU G010 5m2 | P. divaricata | Fruit | Huocheng | MN817612 | MN817637 | MN817587 |
| Species | Isolate no. | Collector | Host | Country | GenBank Accession Numbers | ||
|---|---|---|---|---|---|---|---|
| ITS | LSU | tef1 | |||||
| Dothidotthia symphoricarpi | CBS 119687ET | A. Ramaley | Symphoricarpos rotundifolius | USA | MH863064 | MH874618 | – |
| Mycocentrospora acerina | CBS 113.24 | A. van Luijk | Carum carvi | Netherlands | MH854764 | MH866268 | – |
| Neoascochyta desmazieri | CBS 247.79 | E. Lengauer | Gramineae | Austria | KT389507 | KT389725 | – |
| Neoascochyta desmazieri | CBS 297.69 | U.G. Schlösser | Lolium perenne | Germany | KT389508 | KT389726 | – |
| Neoascochyta europaea | CBS 819.84 | M. Hossfeld | Hordeum vulgare | Germany | KT389510 | KT389728 | – |
| Neoascochyta europaea | CBS 820.84 | M. Hossfeld | Hordeum vulgare | Germany | KT389511 | KT389729 | – |
| Neoascochyta exitialis | CBS 234.52 | E. Muller | Triticum spelta | Switzerland | MH857013 | MH868539 | – |
| Neoascochyta exitialis | CBS 316.81 | H.T. Jachmann | Triticum aestivum | Germany | MH861347 | MH873106 | – |
| Phaeomycocentrospora cantuariensis | CBS 132013 | H.D. Shin | Acalypha australis | South Korea | GU269667 | – | GU384384 |
| Phaeomycocentrospora cantuariensis | CBS 132014 | H.D. Shin | Humulus japonicus | South Korea | GU269668 | – | GU384385 |
| Pleiochaeta carotae | CPC 27452T | M. Truter | carrot | South Africa | KY905669 | KY905663 | – |
| Pleiochaeta setosa | CBS 118.25 | C.M. Doyer | Laburnum | – | KY929373 | KY929376 | – |
| Pleiochaeta setosa | CBS 502.80 | W. Gams | Chamaespartium sagittale | Austria | KY929374 | KY929377 | – |
| Thyrostroma compactum | CBS 335.37 | J.C. Carter | Ulmus pumila | – | KY905670 | KY905664 | KY905681 |
| Thyrostroma cornicola | CBS 141280T | P.W. Crous and H.D. Shin | Cornus officinalis | South Korea | KX228248 | KX228300 | KX228372 |
| Thyrostroma franseriae | CBS 487.71T | F.W. Went | Franseria sp. | USA | KX228249 | KX228301 | KY905680 |
| Thyrostroma franseriae | CBS 700.70 | F.W. Went | Franseria sp. | USA | KX228250 | MH871705 | KY905682 |
| Wilsonomyces carpophilus | CBS 231.89ET | J.W. Veenbaas-Rijks | Prunus subhirtella | – | KY905672 | KY905666 | KY905684 |
| Wilsonomyces carpophilus | CBS 159.51 | G. Goidanich | – | Italy | KY905671 | KY905665 | KY905683 |
| Stigmina platani | CBS 125773 | R. Zare | – | Iran | MH863752 | MH875220 | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, S.; Jia, H.; Cai, G.; Tian, C.; Ma, R. Morphology, DNA Phylogeny, and Pathogenicity of Wilsonomyces carpophilus Isolate Causing Shot-Hole Disease of Prunus divaricata and Prunus armeniaca in Wild-Fruit Forest of Western Tianshan Mountains, China. Forests 2020, 11, 319. https://doi.org/10.3390/f11030319
Ye S, Jia H, Cai G, Tian C, Ma R. Morphology, DNA Phylogeny, and Pathogenicity of Wilsonomyces carpophilus Isolate Causing Shot-Hole Disease of Prunus divaricata and Prunus armeniaca in Wild-Fruit Forest of Western Tianshan Mountains, China. Forests. 2020; 11(3):319. https://doi.org/10.3390/f11030319
Chicago/Turabian StyleYe, Shuanghua, Haiying Jia, Guifang Cai, Chengming Tian, and Rong Ma. 2020. "Morphology, DNA Phylogeny, and Pathogenicity of Wilsonomyces carpophilus Isolate Causing Shot-Hole Disease of Prunus divaricata and Prunus armeniaca in Wild-Fruit Forest of Western Tianshan Mountains, China" Forests 11, no. 3: 319. https://doi.org/10.3390/f11030319