Plastic Responses of Magnolia schiedeana Schltdl., a Relict-Endangered Mexican Cloud Forest Tree, to Climatic Events: Evidences from Leaf Venation and Wood Vessel Anatomy

 ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
- (1)
- evaluate the microenvironmental effect (litter depth, temperature and moisture soil, and canopy openness) on leaf vein traits (leaf shape, vein length, primary vein size, leaf base shape, leaf base angle, angle between major secondary veins and primary veins, and vein density);
- (2)
- examine the relationship between tree-ring width and climatic fluctuations, and determine the effects of local climatic factors on the ring-width index of individual trees; and
- (3)
- assess the influence of drought events on the plasticity of annual tree-ring width and vessel traits (density, hydraulic diameter, and percentage of conductive vessel area).
2. Material and Methods
2.1. Study Forests
2.2. Microclimatic Data
2.3. Leaf Sample Collection
Leaf Vein Traits
2.4. Microenvironmental Factors
- Litter depth: was measured by inserting a ruler into the ground until it touched the mineral soil layer.
- Soil moisture: was recorded at a depth of 2–3 cm using a hygrometer (Lincoln Soil Moisture Meters). This factor took values on a scale from 0 to 100%, where 0 = dry and 100 = saturated.
- Soil temperature: was measured with a soil thermometer (Forestry Suppliers, Taylor® Switchable Digital Pocket Thermometer, Jackson, MS, USA).
- Canopy openness: was estimated in each sample plot using a concave mirror forestry densitometer (Forestry Suppliers, Spherical Crown Densitometers, model A, Jackson, MS, USA).
Statistical Analyses
2.5. Dendrochronological Survey
2.6. Climate Data
2.7. Wood Vessel Traits
- (1)
- Vessel density (VD) was directly measured (mm) on each digital image of demarcated wood core. The number of vessels per square (mm·m−2) was calculated within the area between two wood rays per each tree-ring.
- (2)
- (3)
- Percentage of conductive area (PCA) was calculated according to Rodríguez-Ramírez et al. [42]: , where = ratio of cell wall lumen.
3. Results
3.1. Microenvironmental Requirements
3.2. Microenvironmental Effect on Leaf Vein Traits
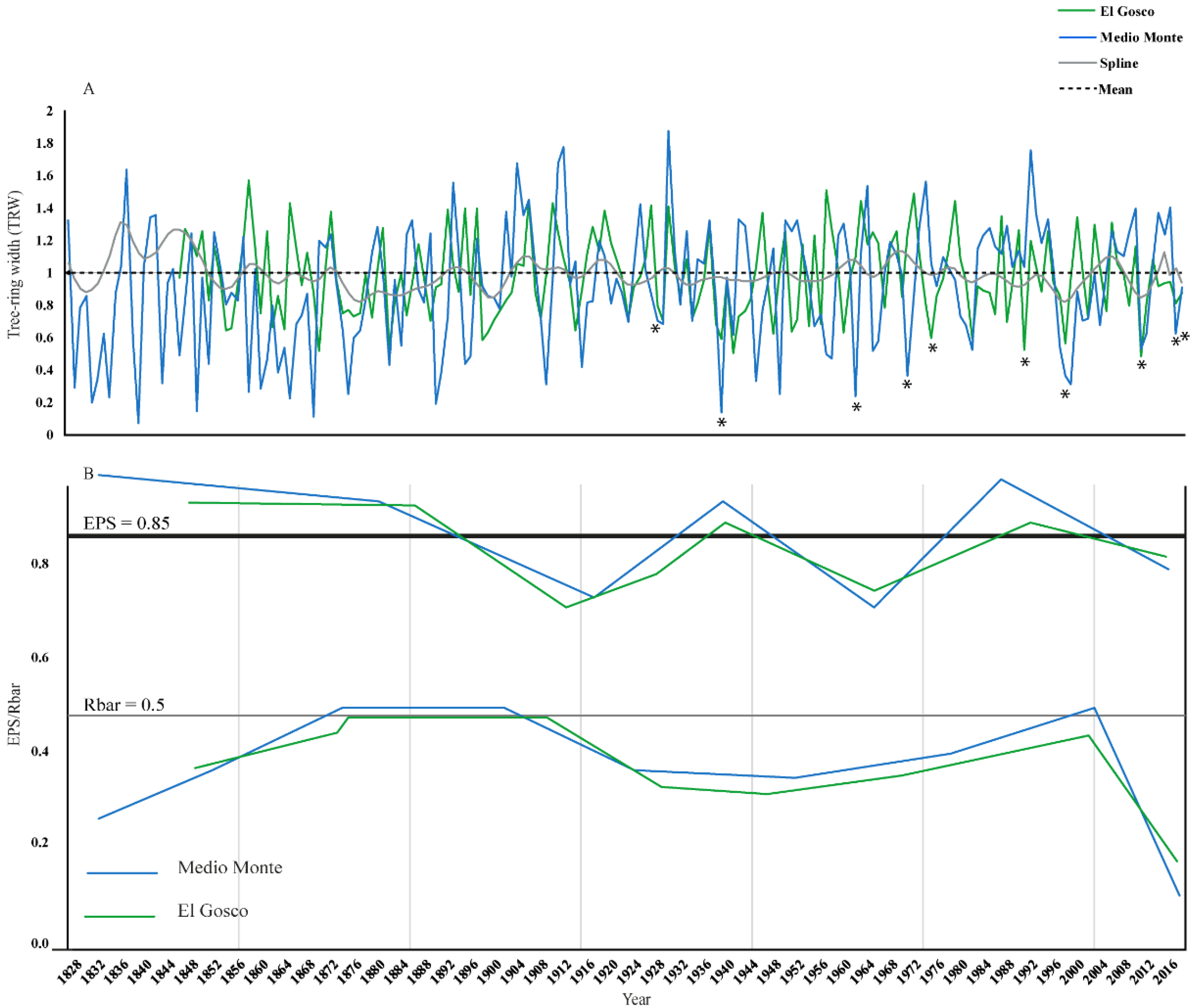
3.3. Tree-Ring Width Chronology
3.4. Tree Growth–Climate Relationship
3.5. The Effect of Drought Events on Vessel Traits
4. Discussion
4.1. Leaf Venation as Evidence to Microenvironmental Adaptation
4.2. Wood Anatomy as Evidence to Climate Adaptation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vázquez-García, J.A. Cloud forest archipelagos: Preservation of fragmented montane ecosystems in tropical America. In Tropical Montane Cloud Forests; Hamilton, L.S., Juvik, J.O., Scatena, F.N., Eds.; Springer: New York, NY, USA, 1995; Volume 110, pp. 296–300. ISBN 9789896540821. [Google Scholar]
- Gauthier, S.; Bernier, P.; Burton, P.J.; Edwards, J.; Isaac, K.; Isabel, N.; Jayen, K.; le Goff, H.; Nelson, E.A. Climate change vulnerability and adaptation in the managed Canadian boreal forest. Environ. Rev. 2014, 22, 256–285. [Google Scholar] [CrossRef]
- Bruijnzeel, L.A.; Mulligan, M.; Scatena, F.N. Hydrometeorology of Tropical Montane Cloud Forests: Emerging Patterns; Cambridge University Press: Cambridge, UK, 2011; Volume 25, ISBN 9780521760355. [Google Scholar]
- Rahbek, C.; Borregaard, M.K.; Antonelli, A.; Colwell, R.K.; Holt, B.G.; Nogues-Bravo, D.; Rasmussen, C.M.Ø.; Richardson, K.; Rosing, M.T.; Whittaker, R.J.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Matzke, N.J.; Dawson, T.E. The incidence and implications of clouds for cloud forest plant water relations. Ecol. Lett. 2013, 16, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, M.; Billington, C.; Edwards, M.; Laidlaw, R. Tropical montane cloud forests: An urgent priority for conservation. WCMC Biodivers. Bull. 1997, 2, 17. [Google Scholar]
- Pouteau, R.; Giambelluca, T.W.; Ah-Peng, C.; Meyer, J.Y. Will climate change shift the lower ecotone of tropical montane cloud forests upwards on islands? J. Biogeogr. 2018, 45, 1326–1333. [Google Scholar] [CrossRef]
- Gupta, A. The physiology of plant responses to drought. Science 2020, 269, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Griggs, D.J.; Noguer, M. Climate change 2001: The scientific basis. Contribution of working group I to the third assessment report of the intergovernmental panel on climate change. Weather 2002, 57, 267–269. [Google Scholar] [CrossRef]
- Ponce-Reyes, R.; Reynoso-Rosales, V.H.; Watson, J.E.M.; Vanderwal, J.; Fuller, R.A.; Pressey, R.L.; Possingham, H.P. Vulnerability of cloud forest reserves in Mexico to climate change. Nat. Clim. Change 2012, 2, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Blackman, C.J.; Brodribb, T.J.; Jordan, G.J. Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol. 2010, 188, 1113–1123. [Google Scholar] [CrossRef]
- Scoffoni, C.; Rawls, M.; Mckown, A.; Cochard, H.; Sack, L. Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture. Plant. Physiol. 2011, 156, 832–843. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramírez, E.C.; Terrazas, T.; Luna-Vega, I. The influence of climate on the masting behavior of Mexican beech: Growth rings and xylem anatomy. Trees Struct. Funct. 2019, 33, 23–35. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gual-Díaz, M.; Rendón-Correa, A. (Eds.) Bosques mesófilos de montaña de México diversidad, ecología y manejo. In Bosques Mesófilos de Montaña de México Diversidad, Ecología y Manejo; CONABIO: Mexico City, Mexico, 2014; pp. 27–67. ISBN 978-607-8328-07-9. [Google Scholar]
- Barradas, V.L.; Luis, J.; Cervantes, J. Implicaciones del cambio climático en la fisiología ecológica de las plantas. In Cambio Climático: Aproximaciones para el Estudio de su Efecto sobre la Biodiversidad; Sánchez-Rojas, G., Barrera, C.B., Pavón, N.P., Eds.; Universidad Autónoma del Estado de Hidalgo: Pachuca de Soto, Mexico, 2011; pp. 71–78. [Google Scholar]
- Rivers, M.; Beech, E.; Murphy, L.; Oldfield, S. (Eds.) The Red List of Magnoliaceae-Revised and Extended, 1st ed.; Botanic Gardens Conservation International: Richmond, UK, 2016; ISBN 9781905164646. [Google Scholar]
- González-Espinosa, M.; Meave, J.A.; Lorea-Hernández, F.G.; Ibarra-Manríquez, G.; Newton, A.C. (Eds.) The Red List of Mexican Cloud Forest Trees, 1st ed.; Fauna & Flora International: Cambridge, UK, 2011; ISBN 9781903703281. [Google Scholar]
- Vásquez-Morales, S.G.; Téllez-Valdés, O.; del Rosario Pineda-López, M.; Sánchez-Velásquez, L.R.; Flores-Estevez, N.; Viveros-Viveros, H. Effect of climate change on the distribution of Magnolia schiedeana: A threatened species. Bot. Sci. 2014, 92, 575. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-García, J.A. Magnolia (Magnoliaceae) in Mexico and Central America: A Synopsis. Brittonia 1994, 46, 1. [Google Scholar] [CrossRef]
- García-González, I.; Fonti, P. Selecting earlywood vessels to maximize their environmental signal. Tree Physiol. 2006, 26, 1289–1296. [Google Scholar] [CrossRef] [Green Version]
- Ariya, U.; Hamano, K.-Y.; Makimoto, T.; Kinoshita, S.; Akaji, Y.; Miyazaki, Y.; Hirobe, M.; Sakamoto, K. Temporal and spatial dynamics of an old-growth beech forest in western Japan. J. For. Res. 2015, 21, 73–83. [Google Scholar] [CrossRef]
- Rodríguez-Ramírez, E.C.; Luna-Vega, I. Dendroecology as a research tool to investigate climate change resilience on Magnolia vovidesii, a threatened Mexican cloud forest tree species of eastern Mexico. In Latin American Dendroecology: Combining Tree-Ring Sciences and Ecology in a Megadiverse Territory; Pompa-García, M., Camarero, J.J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 3–20. [Google Scholar]
- Anderegg, W.R.L. Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation. New Phytol. 2015, 205, 1008–1014. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Meinzer, F.C. Wood anatomy and plant hydraulics in a changing climate. In Functional and Ecological Xylem Anatomy; Springer International Publishing: Cham, Switzerland, 2015; pp. 235–253. ISBN 9783319157832. [Google Scholar]
- Kawai, K.; Okada, N. Leaf vascular architecture in temperate dicotyledons: Correlations and link to functional traits. Planta 2020, 251, 1–12. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, J.; Huang, Y.; Jia, Z.; Fang, Y. Leaf venation variation and phenotypic plasticity in response to environmental heterogeneity in Parrotia subaequalis (H. T. Chang) R. M. Hao et H. T. Wei, an endemic and endangered tree species from China. Forests 2018, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramírez, E.C.; Luna-Vega, I.; Rozas, V. Tree-ring research of Mexican beech (Fagus grandifolia subsp. mexicana) a relict tree endemic to eastern Mexico. Tree-Ring Res. 2018, 74, 94–107. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramírez, E.C.; Sánchez-González, A.; Ángeles-Pérez, G. Relationship between vegetation structure and microenvironment in Fagus grandifolia subsp. mexicana forest relicts in Mexico. J. Plant. Ecol. 2018, 11, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Roberts-Pichette, P.; Gillespie, L. Terrestrial Vegetation Biodiversity Monitoring Protocols; Canada Centre for Inland Waters: Burlington, ON, Canada, 1999. [Google Scholar]
- Shobe, R.W.R.; Lersten, N.R. A technique for clearing and staining gymnosperm leaves. Bot. Gaz. 1967, 128, 150–152. [Google Scholar] [CrossRef]
- Vasco, A.; Thadeo, M.; Conover, M.; Daly, D.C. Preparation of samples for leaf architecture studies, a method for mounting cleared leaves. Appl. Plant. Sci. 2014, 2, 1400038. [Google Scholar] [CrossRef]
- Ash, A.; Ellis, B.; Hickey, L.J.; Johnson, K.; Wilf, P.; Wing, S. (Eds.) Manual of Leaf Architecture-Morphological Description and Categorization of Dicotyledonous and Net-Veined Monocotyledonous Angiosperms, 1st ed.; Smithsonian Institution: Washington, DC, USA, 1999; ISBN 0967755409. [Google Scholar]
- Hickey, L.J. Classification of the architecture of dicotyledonous leaves. Am. J. Bot. 1973, 60, 17. [Google Scholar] [CrossRef]
- Zeileis, A.; Hornik, K. Generalized M-fluctuation tests for parameter instability. Stat. Neerl. 2007, 61, 488–508. [Google Scholar] [CrossRef]
- Package ‘glm2,’. Available online: http://mirror.psu.ac.th/pub/cran/web/packages/glm2/glm2.pdf (accessed on 20 March 2020).
- Polge, H.; Thiercelin, F. Dégâts occasionnés par les sondages à la tarière. Rev. For. Française 1970, 461, 629–636. [Google Scholar] [CrossRef] [Green Version]
- Thiercelin, F.; Arnould, M.-F.; Mangenot, F.; Polge, H. Altérations du bois provoquées par les sondages à la tarière. Leur contrôle. Ann. Des. Sci. For. 1972, 29, 107–133. [Google Scholar] [CrossRef] [Green Version]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Guide for computer program ARSTAN. In The International Tree-Ring Data Bank Program Library Version 2.0 User’s Manual; Grissino-Mayer, H.D., Holmes, R.L., Fritts, H.C., Eds.; Laboratory of Tree-Ring Research: Tucson, AZ, USA, 1996; pp. 75–87. [Google Scholar]
- Briffa, K.R. Interpreting high-resolution proxy climate data—The example of dendroclimatology. In Analysis of Climate Variability; von Storch., H., Navarra, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; Volume 0500, pp. 77–94. [Google Scholar]
- Gareca, E.E.; Fernández, M.; Stanton, S. Dendrochronological investigation of the high Andean tree species Polylepis besseri and implications for management and conservation. Biodivers. Conserv. 2010, 19, 1839–1851. [Google Scholar] [CrossRef]
- Dittmar, C.; Elling, W. Dendroecological investigation of the vitality of common beech (Fagus sylvatica L.) in mixed mountain forests of the Northern Alps (South Bavaria). Dendrochronologia 2007, 25, 37–56. [Google Scholar] [CrossRef]
- Stahle, D.W.; Cook, E.R.; Burnette, D.J.; Villanueva, J.; Cerano, J.; Burns, J.N.; Griffin, D.; Cook, B.I.; Acuña, R.; Torbenson, M.C.A.; et al. The Mexican drought Atlas: Tree-ring reconstructions of the soil moisture balance during the late pre-Hispanic, colonial, and modern eras. Quat. Sci. Rev. 2016, 149, 34–60. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramírez, E.C.; Vázquez-García, J.A.; García-González, I.; Alcántara-Ayala, O.; Luna-Vega, I. Drought effects on the plasticity in vessel traits of two endemic magnolia species in the tropical montane cloud forests of eastern Mexico. J. Plant. Ecol. 2020, 13, 331–340. [Google Scholar] [CrossRef]
- Sperry, J.S.; Saiendra, N.Z. Intra- and inter-plant variation in xylem cavitation in Betula Occident. Plant Cell Environ. 1994, 17, 1233–1241. [Google Scholar] [CrossRef]
- García-González, I.; Souto-Herrero, M.; Campelo, F. Ring-porosity and earlywood vessels: A review on extracting environmental information through time. IAWA J. 2016, 37, 295–314. [Google Scholar] [CrossRef]
- Souto-Herrero, M.; Rozas, V.; García-González, I. A 481-year chronology of oak earlywood vessels as an age-independent climatic proxy in NW Iberia. Glob. Planet. Change 2017, 155, 20–28. [Google Scholar] [CrossRef]
- Camarero, J.J. Linking functional traits and climate-growth relationships in Mediterranean species through wood density. IAWA J. 2019, 40, 215–240. [Google Scholar] [CrossRef] [Green Version]
- Souto-Herrero, M.; Rozas, V.; García-González, I. Earlywood vessels and latewood width explain the role of climate on wood formation of Quercus pyrenaica Willd. across the Atlantic-Mediterranean boundary in NW Iberia. For. Ecol. Manag. 2018, 425, 126–137. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Hintze, J.L.; Nelson, R.D. Violin plots: A box plot-density trace synergism. Am. Stat. 1998, 52, 181–184. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Gentleman, R., Hornik, K., Parmigiani, G., Eds.; Springer: Houston, TX, USA, 2016; Volume 35, ISBN 978-0-387-98140-6. [Google Scholar]
- Rodríguez-Ramírez, E.C.; Martínez-Falcón, A.P.; Luna-Vega, I. Spatial patterns of Mexican beech seedlings (Fagus grandifolia subsp. mexicana (Martínez) A.E. Murray): Influence of canopy openness and conspecific trees on recruitment mechanisms. Ann. For. Sci. 2018. [Google Scholar] [CrossRef] [Green Version]
- Williams-Linera, G.; Tolome, J. Litterfall, temperate and tropical dominant trees, and climate in a Mexican lower montane forest. Biotropica 1996, 28, 649–656. [Google Scholar] [CrossRef]
- Jiménez-García, D.; Peterson, A.T. Climate change impact on endangered cloud forest tree species in Mexico. Rev. Mex. de Biodivers. 2019. [Google Scholar] [CrossRef]
- Givnish, T.J. Leaf and canopy adaptations in tropical forests. In Physiological Ecology of Plants of the Wet Tropics; Medina, E., Mooney, H.A., Vazquez-Yanes, C., Eds.; Springer: Mexico City, Mexico, 1984; pp. 51–84. [Google Scholar]
- Blonder, B.; Kapas, R.E.; Dalton, R.M.; Graae, B.J.; Heiling, J.M.; Opedal, Ø.H. Microenvironment and functional-trait context dependence predict alpine plant community dynamics. J. Ecol. 2018, 106, 1323–1337. [Google Scholar] [CrossRef]
- Vergara-Gómez, D.; Williams-Linera, G.; Casanoves, F. Leaf functional traits vary within and across tree species in tropical cloud forest on rock outcrop versus volcanic soil. J. Veg. Sci. 2020, 31, 129–138. [Google Scholar] [CrossRef]
- Bittencourt, P.R.L.; Barros, F.d.V.; Eller, C.B.; Müller, C.S.; Oliveira, R.S. The fog regime in a tropical montane cloud forest in Brazil and its effects on water, light and microclimate. Agric. For. Meteorol. 2019, 265, 359–369. [Google Scholar] [CrossRef]
- Guerin, G.R.; Wen, H.; Lowe, A.J. Leaf morphology shift linked to climate change. Biol. Lett. 2012, 8, 882–886. [Google Scholar] [CrossRef]
- Williams-Linera, G. Leaf demography and leaf traits of temperate-deciduous and tropical evergreen-broadleaved trees in a Mexican montane cloud forest. Plant. Ecol. 2000, 149, 233–244. [Google Scholar] [CrossRef]
- Sack, L.; Frole, K. Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology 2006, 87, 483–491. [Google Scholar] [CrossRef]
- Rodríguez-Ramírez, E.C.; Sánchez-González, A.; Ángeles-Pérez, G. Current distribution and coverage of Mexican beech forests Fagus grandifolia subsp. mexicana in Mexico. Endanger. Species Res. 2013, 20, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zou, D.; Shrestha, N.; Xu, X.; Wang, Q.; Jia, W.; Wang, Z. Spatiotemporal variation in leaf size and shape in response to climate. J. Plant. Ecol. 2020, 13, 87–96. [Google Scholar] [CrossRef]
- Takahashi, K.; Okuhara, I. Comparison of climatic effects on radial growth of evergreen broad-leaved trees at their northern distribution limit and co-dominating deciduous broad-leaved trees and evergreen conifers. Ecol. Res. 2012, 27, 125–132. [Google Scholar] [CrossRef]
- Pederson, N.; D’Amato, A.W.; Orwig, D.A. Natural history from dendrochronology: Maximum ages and canopy persistence of rarely studied hardwood species. In Proceedings of the 15th Central Hardwood Forest Conference, Knoxville, TN, USA; Buckley, D.S., Clatterbuck, W.K., Eds.; University of Tennessee, Wildlife and Fisheries, Knoxville Department of Agriculture Forest Service, Southern Research Station: Knoxville, TN, USA, 2006; pp. 695–701. [Google Scholar]
- Venegas-González, A.; von Arx, G.; Chagas, M.P.; Filho, M.T. Plasticity in xylem anatomical traits of two tropical species in response to intra-seasonal climate variability. Trees Struct. Funct. 2015, 29, 423–435. [Google Scholar] [CrossRef]
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Vein Traits | Leaf Features | Variable Units | ||
|---|---|---|---|---|
| 1. Leaf shape (LS) | Elliptic | Broadly elliptic | Qualitative | |
| 2. Vein length (VL) | Distance from apex to basal leaf length | cm | ||
| 3. Primary vein size (PVS) | Vein width/leaf width × 100% = 1.25%–2% (moderate) | Vein width/leaf width × 100% = <1.25% (weak) | cm | |
| 4. Leaf base shape (LBS) is taken into consideration from the vertex to 0.25 lm) | (0) Rounded | (1) Concave | Qualitative | |
| 5. Leaf base angle (LBA). Angle from the vertex to the points located 0.25 lm from the lamina | (0) Acute (<90°) | (1) Obtuse (>90°) | Qualitative | |
| 6. Angle between major secondary veins and primary veins (SVPV) | (1) Narrow acute (<45°) | (2) Moderate (45°–65°) | (3) Wide acute (65°–80°) | Qualitative |
| 7. Vein density (VD) | The vein density was expressed as the sum of the length of all its segments (mm) per unit area (mm2) | (mm·mm−2) | ||
| LS | VL | PVS | LBS | LBA | SVPV | VD | |
|---|---|---|---|---|---|---|---|
| Intercept | 5.58 | 13.84 | 1.08 | −0.77 | 60.49 | 36.02 | 0.47 |
| Soil temperature (°C) | 0.14 | −0.99 | 0.24 | 1.88 | 0.04 | 5.85 | 0.07 |
| Canopy openness (%) | 0.25 | 0.85 | −0.35 | 0.3 | −0.63 | −4.02 | −0.06 |
| Soil moisture (%) | −0.95 | 1.17 | 0.06 | 0.63 | 1.79 | 0.95 | −0.002 |
| Litter depth (cm) | −0.65 | 0.12 | −0.12 | −1.65 | −2.06 | −1.35 | −0.05 |
| Akaike information criterion | 29.1 | 95.005 | 39.17 | 24.3 | 127.06 | 124.09 | −33.06 |
| Magnolia Populations | ||
|---|---|---|
| Statistics | Medio Monte | El Gosco |
| Sampled trees | 20 | 20 |
| Cross-dated series a | 32 | 30 |
| Maximum and minimum age sampled | 190/80 | 172/64 |
| Series intercorrelation b | 0.698 | 0.785 |
| Master series (year) | 1829–2019 | 1847–2019 |
| False rings | 3 | 8 |
| Autocorrelation a | 0.632 | 0.587 |
| EPS c | 0.95 | 0.86 |
| Rbar c | 0.46 | 0.44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Ramírez, E.C.; Valdez-Nieto, J.A.; Vázquez-García, J.A.; Dieringer, G.; Luna-Vega, I. Plastic Responses of Magnolia schiedeana Schltdl., a Relict-Endangered Mexican Cloud Forest Tree, to Climatic Events: Evidences from Leaf Venation and Wood Vessel Anatomy. Forests 2020, 11, 737. https://doi.org/10.3390/f11070737
Rodríguez-Ramírez EC, Valdez-Nieto JA, Vázquez-García JA, Dieringer G, Luna-Vega I. Plastic Responses of Magnolia schiedeana Schltdl., a Relict-Endangered Mexican Cloud Forest Tree, to Climatic Events: Evidences from Leaf Venation and Wood Vessel Anatomy. Forests. 2020; 11(7):737. https://doi.org/10.3390/f11070737
Chicago/Turabian StyleRodríguez-Ramírez, Ernesto C., Jeymy Adriana Valdez-Nieto, José Antonio Vázquez-García, Gregg Dieringer, and Isolda Luna-Vega. 2020. "Plastic Responses of Magnolia schiedeana Schltdl., a Relict-Endangered Mexican Cloud Forest Tree, to Climatic Events: Evidences from Leaf Venation and Wood Vessel Anatomy" Forests 11, no. 7: 737. https://doi.org/10.3390/f11070737