Phytosociological Analysis of Natural and Artificial Pine Forests of the Class Vaccinio-Piceetea Br.-Bl. in Br.-Bl. et al. 1939 in the Sudetes and Their Foreland (Bohemian Massif, Central Europe)




Abstract
:1. Introduction
2. Materials and Methods
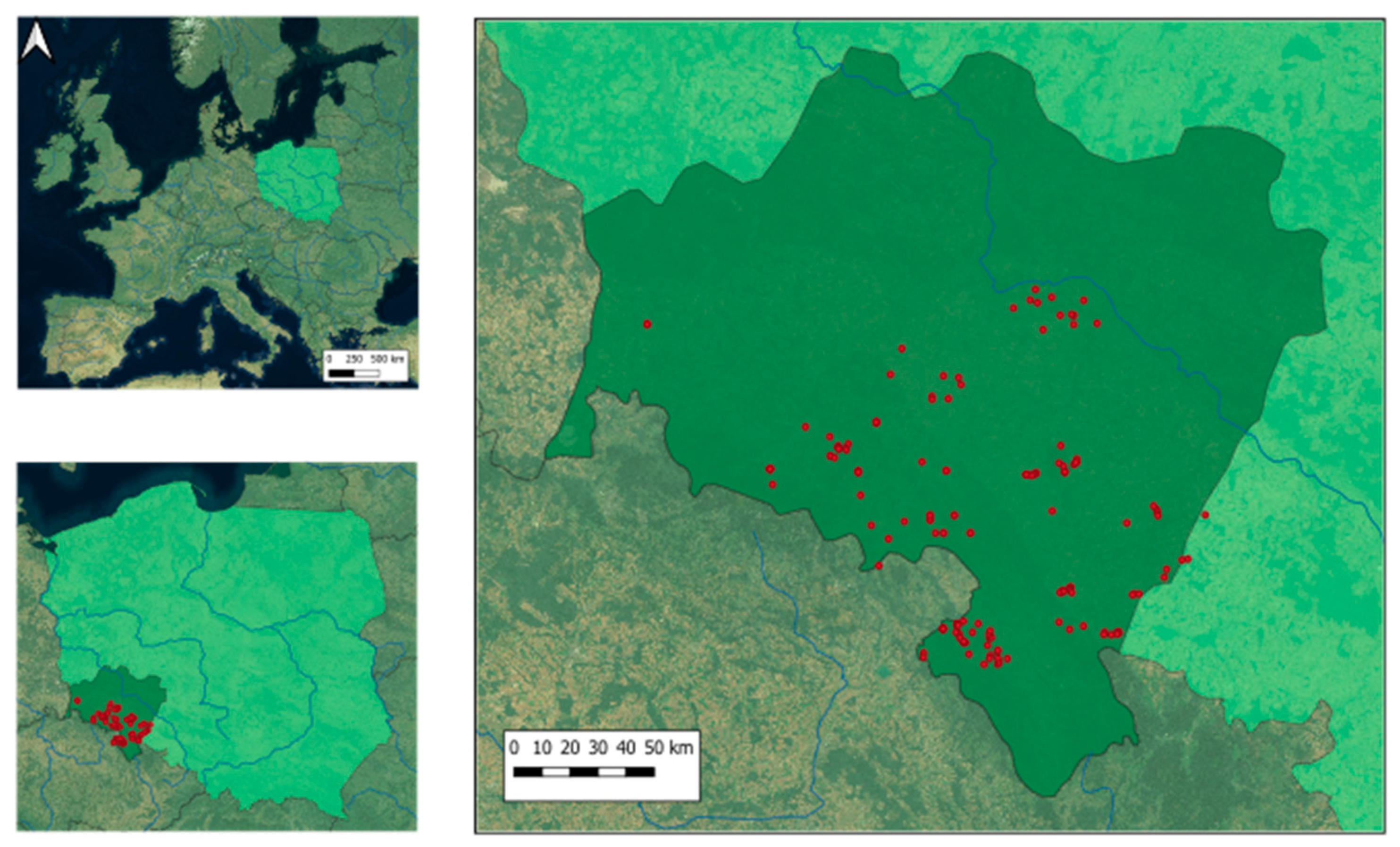
2.1. Study Area
2.2. Field Sampling
2.3. Environmental Variables
2.4. Phytosociological Analysis
2.5. Ecological Analysis
3. Results
3.1. Natural Or Semi-Natural Communities
- Community Brachypodium pinnatum-Pinus sylvestris prov.
- Asplenio cuneifolii-Pinetum sylvestris Pišta ex Husová in Husová et al. 2002.
- Hieracio pallidi-Pinetum sylvestris Stöcker 1965.
- Vaccinio myrtilli-Pinetum sylvestris Juraszek 1928.
- Betulo carpaticae-Pinetum sylvestris Mikyška 1970.
- 6.
- Vaccinio uliginosi-Betuletum pubescentis Libbert 1933.
- 7.
- Comm. Pinus sylvestris-Impatiens parviflora.
- 8.
- Comm. Pinus sylvestris-Prunus serotina.
- 9.
- Comm. Pinus sylvestris-Molinia caerulea.
3.2. Artificial Scots Pine Forests
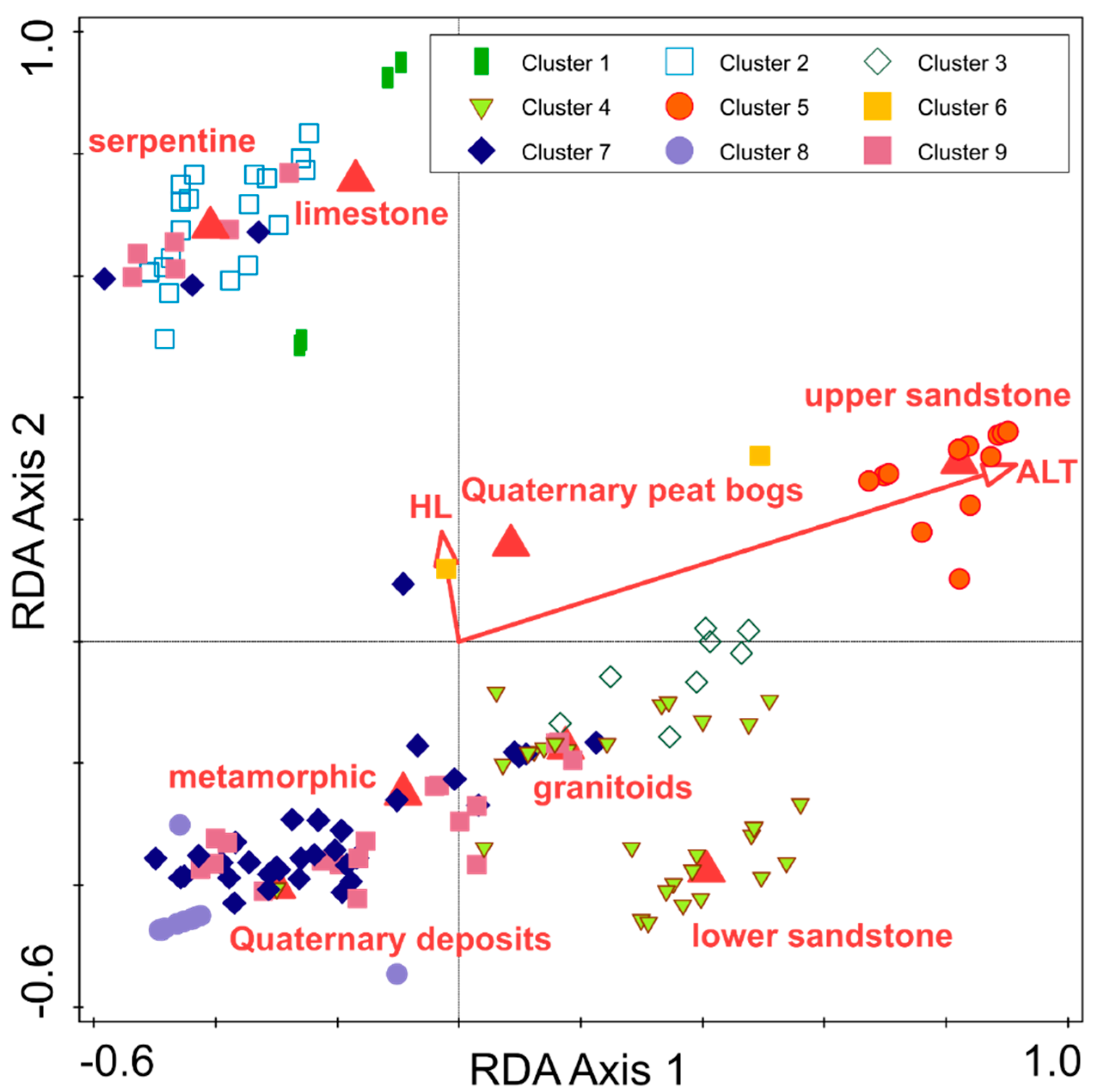
3.3. Ecological Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kobendza, R. Stosunki fitosocjologiczne Puszczy Kampinoskiej. Planta Pol. 1930, 2, 1–200. [Google Scholar]
- Matuszkiewicz, W. Zespoły leśne Białowieskiego Parku Narodowego. Ann. UMCS 1952, Sect. C (Suppl. 6), 1–218. [Google Scholar]
- Matuszkiewicz, W. Zur Systematik der natürlichen Kiefernwälder des mittel- u. osteuropäischen Flachlandes. Mitt. Florist. Soziol. Arbeitsgem. NF 1962, 9, 145–186. [Google Scholar]
- Matuszkiewicz, W. Tymczasowa klasyfikacja zespołów leśnych Polski. Mat. Zakł. Fitosocjol. Stos. UW 1964, 4, 1–5. [Google Scholar]
- Matuszkiewicz, W.; Matuszkiewicz, J.M. Przegląd fitosocjologiczny zbiorowisk leśnych Polski. Vol. 2. Bory sosnowe. Phytocoenosis 1973, 3, 273–356. [Google Scholar]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2001; p. 536. [Google Scholar]
- Matuszkiewicz, J.M. Zespoły Leśne Polski; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2007; p. 357. [Google Scholar]
- Zbiorowiska Roślinne Polski. Lasy i Zarośla; Matuszkiewicz, W.; Sikorski, P.; Szwed, W.; Wierzba, M. (Eds.) Wydawnictwo Naukowe PWN SA: Warszawa, Poland, 2013; p. 517. [Google Scholar]
- Matuszkiewicz, J.M. (Ed.) Geobotanical Identification of the Development Tendencies in Forest Associations in the Regions of Poland; Stanisław Leszczycki Institute of Geography and Spatial Organization Monographies: Warszawa, Poland, 2007; Volume 8, pp. 1–976. [Google Scholar]
- Mikyška, R. Poznamky k některym borum w Čechach a v Kladsku. Preslia 1970, 42, 130–135. [Google Scholar]
- Zelený, D. Vaccinio myrtilli-Pinetum sylvestris Juraszek 1928. In Vegetace České Republiky. 4. Lesní a Křovinná Vegetace; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 394–398. [Google Scholar]
- Matuszkiewicz, A.; Matuszkiewicz, W. 1967. Zespoły roślinne Karkonoskiego Parku Narodowego. Cz. I. Zbiorowiska leśne. Pr. Wrocł. Tow. Nauk. Ser. B 1967, 135, 1–100. [Google Scholar]
- Matuszkiewicz, A.; Matuszkiewicz, W. Mapa zbiorowisk roślinnych Karkonoskiego Parku Narodowego. Ochr. Przyr. 1974, 40, 45–112. [Google Scholar]
- Berdowski, W. Flora mchów i zbiorowisk mszaków Masywu Ślęży. Monogr. Bot. 1974, 45, 125. [Google Scholar]
- Berdowski, W.; Radziuk, M. Szata roślinna Masywu Grochowskiego na Przedgórzu Sudeckim. Acta Univ. Wrat. Prace Bot. 1998, 74, 39–70. [Google Scholar]
- Pender, K. Lasy obszaru Wzgórz Strzelińskich i ich zbiorowiska zastępcze. II. Zbiorowiska zastępcze. Acta Univ. Wrat. Prace Bot 1990, 44, 141–255. [Google Scholar]
- Dengler, J.; Jansen, F.; Glöckler, F.; Peet, R.K.; De Cáceres, M.; Chytrý, M.; Ewald, J.; Oldeland, J.; Lopez-Gonzalez, G.; Finckh, M.; et al. The Global Index of Vegetation-Plot Databases (GIVD): A new resource for vegetation science. J. Veg. Sci. 2011, 22, 582–597. [Google Scholar] [CrossRef]
- Dengler, J.; Oldeland, J.; Jansen, F.; Chytrý, M.; Ewald, J.; Finckh, M.; Glöckler, F.; Lopez-Gonzalez, G.; Peet, R.K.; Schaminée, J.H.J. Vegetation databases for the 21st century. Biodivers. Ecol. 2012, 4, 1–447. [Google Scholar] [CrossRef] [Green Version]
- Willner, W. Syntaxonomische Revision der südmitteleuropäischen Buchenwälder. Phytocoenologia 2002, 32, 337–453. [Google Scholar] [CrossRef]
- Chytrý, M. (Ed.) Vegetace České Republiky 4. Lesní a Křovinná Vegetace. Vegetation of the Czech Republic 4. Forest and Scrub Vegetation; Academia: Praha, Czech Republic, 2013; p. 551. [Google Scholar]
- Douda, J.; Boublík, K.; Slezák, M.; Biurrun, I.; Nociar, J.; Havrdová, A.; Doudová, J.; Aćić, S.; Brisse, H.; Brunet, J.; et al. Vegetation classification and biogeography of European floodplain forests and alder carrs. Appl. Veg. Sci. 2016, 19, 147–163. [Google Scholar] [CrossRef]
- Novák, P.; Willner, W.; Zukal, D.; Kollár, J.; Roleček, K.; Świerkosz, K.; Ewald, J.; Wohlgemuth, T.; Csiky, K.; Onyshchenko, V.; et al. Oak-hornbeam forests of Central Europe: A formalized classification and syntaxonomic revision. Preslia 2020, 92, 1–34. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Dengler, J.; Čarni, A.; Šumberová, K.; Raus, T.; Di Pietro, R.; Tichý, L.; et al. Vegetation of Europe: Hierarchical floristic classification system of plant; lichen; and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 3–264. [Google Scholar] [CrossRef]
- Opracowanie Ekofizjograficzne dla Województwa Dolnośląskiego; Blachowski, J.; Markowicz-Judycka, E.; Zięba, D. (Eds.) Wojewódzkie Biuro Urbanistyczne we Wrocławiu: Wrocław, Poland, 2005; p. 268. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Reprint; The Blackburn Press: Caldwell, NJ, USA, 2002; p. 547. [Google Scholar]
- McCune, B.; Keon, D. Equations for potential annual direct incident radiation and heat load. J. Veg. Sci. 2002, 13, 603–606. [Google Scholar] [CrossRef]
- Bolewski, A.; Parachoniak, W. Petrografia [Petrography]; Wydawnictwo Geologiczne: Warszawa, Poland, 1988; p. 656. [Google Scholar]
- Wojewoda, J. Geology and geological history. In Góry Stolowe. Geology, Landforms, Vegetation Pattern and Human Impacts; Kasprzak, M., Migon, P., Eds.; Instytut Geografii i Rozwoju Regionalnego Uniwersystetu Wrocławskiego: Wrocław, Poland, 2012; pp. 5–11. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, W.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1991, 18, 1–258. [Google Scholar]
- Berg, C.; Welk, E.; Jager, E.J. Revising Ellenberg’s indicator values for continentality based on global vascular plant species distribution. Appl. Veg. Sci. 2017, 20, 482–493. [Google Scholar] [CrossRef]
- Tichý, L.; Holt, J. Juice Program for Management; Analysis and Classification of Ecological Data, 1st ed.; Masaryk University: Brno, Czech Republic, 2006; p. 98. [Google Scholar]
- Fischer, H.S. On the combination of species cover values from different vegetation layers. Appl. Veg. Sci. 2015, 18, 169–170. [Google Scholar] [CrossRef]
- Euro+Med. Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. 2006. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 2 September 2020).
- Ochyra, R.; Żarnowiec, J.; Bednarek-Ochyra, H. Census Catalogue of Polish Mosses. Biodiversity of Poland; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2003; Volume 3, p. 372. [Google Scholar]
- Fałtynowicz, W. The Lichens; Lichenicolous and Allied Fungi of Poland—An Annotated Checklist; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2003; p. 435. [Google Scholar]
- Roleček, J.; Tichý, L.; Zelený, D.; Chytrý, M. Modified TWINSPAN classification in which the hierarchy respects cluster heterogeneity. J. Veg. Sci. 2009, 20, 596–602. [Google Scholar] [CrossRef]
- Tichý, L. JUICE; software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Botta-Dukát, Z.; Chytrý, M.; Hájková, P.; Havlová, M. Vegetation of lowland wet meadows along a climatic continentality gradient in Central Europe. Preslia 2005, 77, 89–111. [Google Scholar]
- Chytrý, M.; Tichý, L.; Holt, J.; Botta-Dukát, Z. Determination of diagnostic species with statistical fidelity measures. J. Veg. Sci. 2002, 13, 79–90. [Google Scholar] [CrossRef]
- Tichý, L.; Chytrý, M. Statistical determination of diagnostic species for site groups of unequal size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Willner, W.; Tichý, L.; Chytrý, M. Effects of different fidelity measures and contexts on the determination of diagnostic species. J. Veg. Sci. 2009, 20, 130–137. [Google Scholar] [CrossRef]
- Dengler, J.; Berg, C.; Jansen, F. New ideas for modern phytosociological monographs. Annali Bot. Nuova Ser. 2005, 5, 193–210. [Google Scholar] [CrossRef]
- Morton, A. DMAP Distribution Mapping Software. 2001. Available online: http://www.dmap.co.uk/ (accessed on 10 October 2020).
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination; Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; p. 496. [Google Scholar]
- Zelený, D.; Schaffers, A.P. Too good to be true: Pitfalls of using mean Ellenberg indicator values in vegetation analyzes. J. Veg. Sci. 2012, 23, 419–431. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; Available online: http://www.R-project.org (accessed on 10 October 2020).
- Gleason, J. Comparative Power of the Anova, Randomization Anova, and Kruskal-Wallis Test. Wayne State University Dissertations 658. 2013. Available online: https://digitalcommons.wayne.edu/oa_dissertations/658 (accessed on 12 December 2020).
- Jupke, J.F.; Schäfer, R.B. Should ecologists prefer model- over distance-based multivariate methods? Ecol. Evol. 2020, 10, 2417–2435. [Google Scholar] [CrossRef]
- Sádlo, J.; Chytrý, M. Festuco-Pinetum sylvestris Kobendza 1930. In Vegetace České Republiky. 4. Lesní a Křovinná Vegetace; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 385–389. [Google Scholar]
- Jarolímek, I.; Šibík, J.; Hegedüšová, K.; Janišová, M.; Kliment, J.; Kučera, P.; Májeková, J.; Michálková, D.; Sadloňová, J.; Zaliberová, M.; et al. A list of vegetation units of Slovakia. In Diagnostic; Constant and Dominant Species of the Higher Vegetation Units of Slovakia; Jarolímek, I., Šibík, J., Eds.; Veda: Bratislava, Slovakia, 2008; pp. 295–329. [Google Scholar]
- Zelený, D. Asplenio cuneifolii-Pinetum sylvestris Pišta ex Husová in Husová et al. 2002. In Vegetace České Republiky. 4. Lesní a Křovinná Vegetace; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 401–404. [Google Scholar]
- Mucina, L.; Grabherr, G.; Wallnöfer, S. Die Pflanzengesellschaften Österreichs. Teil III. Wälder und Gebüsche; Gustav Fischer Verlag: Jena, Germany, 1993; p. 353. [Google Scholar]
- Die Wälder und Gebüsche Österreichs; Willner, W.; Grabherr, G. (Eds.) Spektrum Akademischer Verlag: Heidelberg, Germany, 2007; p. 302. [Google Scholar]
- Świerkosz, K. Zbiorowiska roślinne Góry Chojnik—Eksklawy Karkonoskiego Parku Narodowego. część 1. Zbiorowiska leśne. Parki Nar. Rezerw. Przyr. 1994, 13, 17–36. [Google Scholar]
- Zelený, D. Hieracio pallidi-Pinetum sylvestris Stöcker 1965. In Vegetace České Republiky. 4. Lesní a Křovinná Vegetace; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 398–401. [Google Scholar]
- Pott, R. Die Pflanzengesellschaften Deutschlands, 2nd ed.; Eugen Ulmer Verlag: Stuttgart, Germany, 1995; p. 427. [Google Scholar]
- Seibert, P. Klasse Vaccinio-Piceetea Br.-Bl in Br.-Bl. et al 39. In Süddeutsche Pflanzengesellschaften; Band 4: Wälder und Gebüsche, 2nd ed.; Oberdorfer, E., Ed.; Gustav Fischer Verlag: Jena, Germany, 1992; pp. 53–80. [Google Scholar]
- Passarge, H. Pflanzengesellschaften im Zittauer Bergland. Abh. Ber. Nat. Gorlitz 1981, 54, 1–48. [Google Scholar]
- Härtel, H.; Sadlo, J.; Świerkosz, K.; Markova, I. Phytogeography of the sandstone areas in the Bohemian Cretaceous Basin (Czech Republic/Germany/Poland). In Sandstone Landscapes; Härtel, H., Cilek, V., Hebren, T., Jackson, A., Williams, R., Eds.; Academia: Praha, Czech Republic, 2007; pp. 177–189. [Google Scholar] [CrossRef]
- Świerkosz, K. General characteristics of the vascular flora and geobotanical division of the Góry Stołowe Mts; Sudety Mts (Poland). In Sandstone Landscapes; Härtel, H., Cilek, V., Hebren, T., Jackson, A., Williams, R., Eds.; Academia: Praha, Czech Republic, 2007; pp. 194–200. [Google Scholar] [CrossRef]
- Bobowicz, M.A. Intrapopulational variability of Pinus sylvestris L. from Piekielna Góra in respect to cone and needle traits. Acta Soc. Bot. Pol. 1993, 62, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Krzakowa, M.; Lisowska, D. Genetyczne zróżnicowanie populacji sosny zwyczajnej (Pinus sylvestris L.) z Piekielnej Góry. In Środowisko Przyrodnicze Parku Narodowego Gór Stołowych; Park Narodowy Gór Stołowych: Kudowa Zdrój, Poland, 1996; pp. 125–131. [Google Scholar]
- Boratyński, A.; Danielewicz, W. Chronione i godne ochrony drzewa i krzewy polskiej części Sudetów; Pogórza i Przedgórza Sudeckiego. 5. Betula pubescens Ehrh. ssp. carpatica (Waldst. et Kit) Asch. et Greabn. Arbor. Kórnickie 1989, 34, 71–88. [Google Scholar]
- Świerkosz, K.; Boratyński, A. Chorological and synanthropodynamical analysis of trees and shrubs of the Stołowe Mts (Middle Sudety). Dendrobiology 2002, 48, 75–85. [Google Scholar]
- Białobrzeska, M.; Truchanowicz, J. Badania systematyczno-biometryczne nad rodzajem Betula L. ze Szczelińca w Górach Stołowych. Rocz. Sekc. Dendr. 1991, 39, 51–78. [Google Scholar]
- Boratyński, A. Sosna błotna (Pinus uliginosa Neumann) w rezerwacie “Błędne Skały” w Górach Stołowych. Arbor. Kórnickie 1978, 23, 261–267. [Google Scholar]
- Sobierajska, K.; Wachowiak, W.; Zaborowska, J.; Łabiszak, B.; Wójkiewicz, B.; Sękiewicz, M.; Jasińska, A.K.; Sękiewicz, K.; Boratyńska, K.; Marcysiak, K.; et al. Genetic Consequences of Hybridization in Relict Isolated Trees Pinus sylvestris and the Pinus mugo Complex. Forests 2020, 11, 1086. [Google Scholar] [CrossRef]
- Kącki, Z.; Szymura, M.; Świerkosz, K.; Swacha, G.; Pender, K. Zbiorowiska roślinne Gór Stołowych. In Góry Stołowe—Przyroda i Ludzie; Kabała, C., Ed.; Park Narodowy Gór Stołowych: Kudowa Zdrój, Poland, 2018; pp. 311–328. [Google Scholar]
- Navrátilová, J. Vaccinio uliginosi-Pinetum sylvestris de Kleist 1929. In Vegetace České Republiky. 4. Lesní a Křovinná Vegetace; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 424–426. [Google Scholar]
- Valachovič, M.; Hegedüšová, K. Forest communities of the Jurske jazero (Malé Karpaty Mts). Nat. Tutela 2013, 17, 15–19. [Google Scholar]
- Perring, M.P.; Diekmann, M.; Midolo, G.; Schellenberger Costa, D.; Bernhardt-Römermann, M.; Otto, J.C.J.; Gilliam, S.F.; Hedwall, P.O.; Nordin, A.; Dirnböck, T.; et al. How nitrogen deposition affects forest understorey plant community change: Towards understanding context dependency. Environ. Pollut. 2018, 242, 1787–1799. [Google Scholar] [CrossRef]
- Staude, I.; Waller, D.M.; Bernhardt-Römermann, M.; Bjorkman, A.D.; Brunet, J.; De Frenne, P.; Hédl, R.; Jandt, U.; Lenoir, J.; Máliš, F.; et al. Replacements of small- by larger-ranged species scale to diversity loss in Europe’s temperate forest biome. Nat. Ecol. Evol. 2020, 4, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Pergl, J.; Essl, F.; Lenzner, B.; Dawson, W.; Kreft, H.; Weigelt, P.; Winter, M.; Kartesz, J.; Nishino, M.; et al. Naturalized alien flora of the world: Species diversity; taxonomic and phylogenetic patterns; geographic distribution and global hotspots of plant invasion. Preslia 2017, 89, 203–274. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- De Frenne, P.; Zellweger, F.; Rodríguez-Sánchez, F.; Scheffers, B.R.; Hylander, K.; Luoto, M.; Vellend, M.; Verheyen, K.; Lenoir, J. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, F.; De Frenne, P.; Lenoir, J.; Vangansbeke, P.K.; Bernhardt-Römermann, M.; Baeten, L.; Hédl, R.; Berki, I.; Brunet, J.; Van Calster, H.; et al. Forest microclimate feedbacks drive plant responses to warming. Science 2020, 368, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Miller, J.H.; Wang, H.; Taylor, J.W., Jr. Invasion of tallow tree into southern US forests: Influencing factors and implications for mitigation. Can. J. For. Res. 2009, 39, 1346–1356. [Google Scholar] [CrossRef]
- Lenda, M.; Knops, J.H.; Skórka, P.; Moroń, D.; Woyciechowski, M. Cascading effects of changes in land use on the invasion of the walnut Juglans regia in forest ecosystems. J. Ecol. 2018, 106, 671–686. [Google Scholar] [CrossRef]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef]
- Glasby, T.M.; Connell, S.D.; Holloway, M.G.; Hewitt, C.L. Nonindigenous biota on artificial structures: Could habitat creation facilitate biological invasions? Mar. Biol. 2007, 151, 887–895. [Google Scholar] [CrossRef]
- Padmanaba, M.; Corlett, R.T. Minimizing Risks of Invasive Alien Plant Species in Tropical Production Forest Management. Forests 2014, 5, 1982–1998. [Google Scholar] [CrossRef] [Green Version]
- Charbonneau, N.C.; Fahrig, L. Influence of canopy cover and amount of open habitat in the surrounding landscape on proportion of alien plant species in forest sites. Écoscience 2004, 11, 278–281. [Google Scholar] [CrossRef]
- Langmaier, M.; Lapin, K. A Systematic Review of the Impact of Invasive Alien Plants on Forest Regeneration in European Temperate Forests. Front. Plant Sci. 2020, 11, 524969. [Google Scholar] [CrossRef] [PubMed]
- Foxcroft, L.C.; Spear, D.; van Wilgen, N.J.; McGeoch, M.A. Assessing the association between pathways of alien plant invaders and their impacts in protected areas. NeoBiota 2019, 43, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Eichenberg, D.; Bowler, D.E.; Bonn, A.; Bruelheide, H.; Grescho, V.; Harter, D.; Jandt, U.; May, R.; Winter, M.; Jansen, F. Widespread decline in Central European plant diversity across six decades. Glob. Chang. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster No. | Const. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| No. of relevés | ratio | 5 | 22 | 10 | 33 | 20 | 8 | 39 | 14 | 24 |
| Comm. Brachypodium pinnatum-Pinus sylvestris | ||||||||||
| Sanguisorba minor | 100.0 | 100 100 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Scabiosa ochroleuca | 100.0 | 100 100 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Encalypta streptocarpa | 100.0 | 100 100 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Galium album | 17.6 | 80 85.6 | 5 __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Poa compressa | 100.0 | 80 88.4 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Carlina vulgaris | 100.0 | 80 88.4 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Lotus corniculatus | 2.9 | 80 73.1 | 27 16.2 | . __ | . __ | . __ | . __ | 3 __ | . __ | . __ |
| Cornus sanguinea | 7.8 | 60 70.3 | . __ | . __ | . __ | . __ | . __ | 8 __ | . __ | . __ |
| Solidago virgaurea | 11.7 | 60 72.0 | . __ | . __ | . __ | . __ | . __ | 5 __ | . __ | . __ |
| Salix caprea | 19.8 | 60 73.4 | . __ | . __ | 3 __ | . __ | . __ | . __ | . __ | . __ |
| Origanum vulgare | 100.0 | 60 75.6 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Potentilla tabernaemontani | 13.2 | 60 72.4 | 5 __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Fragaria viridis | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Syntrichia ruralis | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Epipactis atrorubens | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Hieracium vulgatum | 2.8 | 40 39.2 | 5 __ | 10 __ | . __ | . __ | . __ | 5 __ | 14 __ | 4 __ |
| Leucanthemum vulgare | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Anthyllis vulneraria | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Centaurea jacea | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Leontodon autumnalis | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Viola hirta | 100.0 | 40 61.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Asplenio cuneifolii-Pinetum sylvestris Pišta ex Husová in Husová et al. 2002 | ||||||||||
| Galium verum | 19.6 | . __ | 82 86.9 | . __ | . __ | . __ | . __ | . __ | . __ | 4 __ |
| Silene vulgaris | 30.1 | . __ | 77 85.1 | . __ | . __ | . __ | . __ | 3 __ | . __ | . __ |
| Achillea millefolium | 1.9 | 40 __ | 77 67.4 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Festuca ovina | 3.4 | . __ | 73 67.1 | . __ | . __ | . __ | . __ | 3 __ | 21 __ | 8 __ |
| Lychnis viscaria | 100.0 | . __ | 73 83.9 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Rosa canina | 7.1 | . __ | 59 64.8 | . __ | . __ | . __ | . __ | 8 __ | . __ | 8 __ |
| Galium rotundifolium | 4.4 | . __ | 55 60.5 | . __ | . __ | . __ | . __ | 5 __ | . __ | 12 __ |
| Hypericum montanum | 10.9 | . __ | 45 60.0 | . __ | . __ | . __ | . __ | 3 __ | . __ | 4 __ |
| Hypericum perforatum | 2.0 | 20 __ | 41 33.3 | . __ | . __ | . __ | . __ | 13 __ | 14 __ | 12 __ |
| Hieracium sabaudum | 4.9 | . __ | 41 51.7 | . __ | . __ | . __ | . __ | 5 __ | . __ | 8 __ |
| Robinia pseudoacacia | 5.1 | . __ | 36 50.1 | . __ | . __ | . __ | . __ | 3 __ | 7 __ | . __ |
| Hylotelephium maximum | 100.0 | . __ | 36 58.1 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Potentilla alba | 6.2 | . __ | 32 49.4 | . __ | . __ | . __ | . __ | 5 __ | . __ | . __ |
| Melica nutans | 1.6 | 20 __ | 32 38.2 | . __ | . __ | . __ | . __ | 3 __ | . __ | . __ |
| Viola riviniana | 2.5 | . __ | 32 37.1 | . __ | . __ | . __ | . __ | 5 __ | 7 __ | 12 __ |
| Hieracium pilosella | 100.0 | . __ | 27 50.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Asplenium cuneifolium | 100.0 | . __ | 27 50.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Genista tinctoria | 100.0 | . __ | 27 50.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Poa angustifolia | 3.2 | . __ | 23 35.1 | . __ | . __ | . __ | . __ | . __ | 7 __ | 4 __ |
| Viola canina | 100.0 | . __ | 23 45.5 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Danthonia decumbens | 100.0 | . __ | 23 45.5 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Phleum phleoides | 100.0 | . __ | 23 45.5 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Polypodium vulgare | 5.5 | . __ | 23 41.0 | . __ | . __ | . __ | . __ | . __ | . __ | 4 __ |
| Centaurea stoebe | 100.0 | . __ | 23 45.5 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Hieracio pallidi-Pinetum sylvestris Stöcker 1965 | ||||||||||
| Polytrichum piliferum | 23.1 | . __ | . __ | 70 80.1 | 3 __ | . __ | . __ | . __ | . __ | . __ |
| Cladonia uncialis | 100.0 | . __ | . __ | 30 52.5 | . __ | . __ | . __ | . __ | . __ | . __ |
| Grimmia pulvinata | 100.0 | . __ | . __ | 20 42.7 | . __ | . __ | . __ | . __ | . __ | . __ |
| Vaccinio myrtilli-Pinetum sylvestris Juraszek 1928 | ||||||||||
| Dicranum polysetum | 3.3 | . __ | 5 __ | 10 __ | 33 38.1 | . __ | . __ | . __ | 7 __ | 4 __ |
| Leucobryum glaucum | 3.0 | . __ | 5 __ | 10 __ | 30 34.2 | 5 __ | . __ | . __ | . __ | 8 __ |
| Hylocomium splendens | 3.6 | . __ | . __ | . __ | 30 45.3 | . __ | . __ | . __ | . __ | 8 __ |
| Dicranella heteromalla | 1.7 | . __ | . __ | 10 __ | 21 21.7 | . __ | . __ | 13 __ | . __ | 12 __ |
| Betulo carpaticae-Pinetum sylvestris Mikyška 1970 | ||||||||||
| Betula pubescens var. glabrata | 100.0 | . __ | . __ | . __ | . __ | 90 94.3 | . __ | . __ | . __ | . __ |
| Empetrum nigrum subsp. nigrum | 100.0 | . __ | . __ | . __ | . __ | 40 61.0 | . __ | . __ | . __ | . __ |
| Vaccinium uliginosum | 100.0 | . __ | . __ | . __ | . __ | 40 61.0 | . __ | . __ | . __ | . __ |
| Pinus x rhaetica | 100.0 | . __ | . __ | . __ | . __ | 35 56.9 | . __ | . __ | . __ | . __ |
| Cetraria islandica | 3.5 | . __ | . __ | 10 __ | . __ | 35 48.6 | . __ | . __ | . __ | . __ |
| Vaccinio uliginosi-Betuletum pubescentis Libbert 1933 | ||||||||||
| Sphagnum fallax | 100.0 | . __ | . __ | . __ | . __ | . __ | 100 100 | . __ | . __ | . __ |
| Betula pubescens var. pubescens | 2.5 | . __ | . __ | 40 17.5 | 15 __ | . __ | 100 70.3 | . __ | 21 __ | 4 __ |
| Polytrichum commune | 100.0 | . __ | . __ | . __ | . __ | . __ | 88 92.8 | . __ | . __ | . __ |
| Molinia caerulea subsp. arundinacea | 6.4 | . __ | 14 __ | . __ | 9 __ | . __ | 88 80.0 | 3 __ | . __ | . __ |
| Carex rostrata | 100.0 | . __ | . __ | . __ | . __ | . __ | 75 85.3 | . __ | . __ | . __ |
| Juncus effusus | 9.8 | . __ | . __ | . __ | 3 __ | . __ | 75 73.1 | 8 __ | 7 __ | 4 __ |
| Eriophorum vaginatum | 100.0 | . __ | . __ | . __ | . __ | . __ | 62 77.3 | . __ | . __ | . __ |
| Salix aurita | 100.0 | . __ | . __ | . __ | . __ | . __ | 50 68.6 | . __ | . __ | . __ |
| Sphagnum palustre | 100.0 | . __ | . __ | . __ | . __ | . __ | 50 68.6 | . __ | . __ | . __ |
| Carex acutiformis | 100.0 | . __ | . __ | . __ | . __ | . __ | 25 47.8 | . __ | . __ | . __ |
| Trientalis europaea | 3.0 | . __ | . __ | . __ | 3 __ | . __ | 25 37.6 | . __ | . __ | 8 __ |
| Carex leporina | 2.0 | . __ | . __ | . __ | . __ | . __ | 25 35.2 | 3 __ | . __ | 12 13.8 |
| Carex nigra | 100.0 | . __ | . __ | . __ | . __ | . __ | 25 47.8 | . __ | . __ | . __ |
| Carex panicea | 100.0 | . __ | . __ | . __ | . __ | . __ | 25 47.8 | . __ | . __ | . __ |
| Eriophorum angustifolium | 100.0 | . __ | . __ | . __ | . __ | . __ | 25 47.8 | . __ | . __ | . __ |
| Comm. Pinus sylvestris-Impatiens parviflora (Artificial Forest) | ||||||||||
| Impatiens parviflora | 2.2 | . __ | 18 __ | . __ | . __ | . __ | . __ | 79 60.6 | 36 __ | 12 __ |
| Dryopteris filix-mas | 2.8 | . __ | 18 __ | . __ | . __ | . __ | . __ | 59 52.8 | 21 __ | 4 __ |
| Urtica dioica | 11.3 | . __ | 5 __ | . __ | . __ | . __ | . __ | 51 66.1 | . __ | . __ |
| Acer platanoides | 2.4 | 20 __ | 14 __ | . __ | . __ | . __ | . __ | 49 41.6 | 7 __ | 12 __ |
| Sambucus nigra | 3.1 | . __ | . __ | . __ | . __ | . __ | . __ | 38 50.1 | . __ | 12 __ |
| Crataegus monogyna | 1.8 | 20 __ | 5 __ | . __ | . __ | . __ | . __ | 36 33.8 | 14 __ | 4 __ |
| Galeopsis pubescens | 2.2 | . __ | . __ | . __ | . __ | . __ | . __ | 31 41.7 | 14 __ | . __ |
| Brachypodium sylvaticum | 2.8 | . __ | 9 __ | . __ | . __ | . __ | . __ | 26 34.5 | . __ | 8 __ |
| Drymochloa sylvatica | 1.6 | . __ | . __ | . __ | . __ | . __ | . __ | 21 31.6 | . __ | 12 __ |
| Comm. Pinus sylvestris-Prunus serotina (Artificial Forest) | ||||||||||
| Prunus serotina | 8.4 | . __ | . __ | . __ | . __ | . __ | . __ | 10 __ | 86 85.9 | . __ |
| Viscum album | 100.0 | . __ | . __ | . __ | . __ | . __ | . __ | . __ | 71 83.1 | . __ |
| Convallaria majalis | 2.8 | . __ | 18 __ | . __ | 3 __ | . __ | . __ | 10 __ | 50 50.4 | . __ |
| Holcus mollis | 4.3 | . __ | . __ | . __ | . __ | . __ | . __ | 3 __ | 36 48.7 | 8 __ |
| Atrichum undulatum | 2.8 | . __ | . __ | . __ | 3 __ | . __ | . __ | 8 __ | 21 34.0 | . __ |
| Hedera helix | 1.7 | . __ | . __ | . __ | . __ | . __ | . __ | 13 13.4 | 21 27.7 | 8 __ |
| Comm. Pinus sylvestris-Molinia caerulea (Artificial Forest) | ||||||||||
| Maianthemum bifolium | 3.0 | . __ | . __ | . __ | 3 __ | . __ | 12 __ | 18 __ | 14 __ | 54 47.7 |
| Molinia caerulea subsp. caerulea | 2.5 | . __ | 5 __ | . __ | 6 __ | . __ | . __ | 5 __ | 21 __ | 54 51.4 |
| Athyrium filix-femina | 4.5 | . __ | . __ | . __ | . __ | . __ | . __ | 10 __ | . __ | 46 57.9 |
| Rubus hirtus agg. | 2.3 | . __ | . __ | . __ | . __ | . __ | . __ | 18 __ | . __ | 42 49.8 |
| Luzula pilosa | 2.4 | . __ | 5 __ | . __ | 9 __ | . __ | . __ | 10 __ | . __ | 25 30.5 |
| Pteridium aquilinum | 1.7 | . __ | . __ | . __ | 15 __ | . __ | . __ | 10 __ | . __ | 25 29.8 |
| Pseudoscleropodium purum | 1.7 | . __ | 5 __ | . __ | 12 __ | . __ | . __ | 3 __ | 7 __ | 21 24.7 |
| Polytrichastrum formosum | 1.6 | . __ | 18 __ | . __ | 42 __ | 10 __ | 12 __ | 33 __ | 29 __ | 67 35.8 |
| Species diagnostic for at least two clusters | ||||||||||
| Hieracium murorum | 2.8 | 100 69.7 | 36 14.0 | 20 __ | 9 __ | . __ | . __ | 18 __ | . __ | . __ |
| Campanula rotundifolia | 7.6 | 100 71.9 | 64 39.4 | . __ | . __ | . __ | . __ | 3 __ | . __ | 8 __ |
| Brachypodium pinnatum | 16.0 | 80 68.0 | 41 27.8 | . __ | . __ | . __ | . __ | 3 __ | . __ | . __ |
| Thymus pulegioides | 100.0 | 60 58.1 | 32 25.2 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Pimpinella saxifraga | 100.0 | 60 47.4 | 64 51.1 | . __ | . __ | . __ | . __ | . __ | . __ | . __ |
| Euphorbia cyparissias | 21.3 | 60 49.3 | 55 43.5 | . __ | . __ | . __ | . __ | 3 __ | . __ | . __ |
| Ceratodon purpureus | 5.0 | 80 58.2 | 5 __ | 50 30.3 | . __ | 10 __ | . __ | 5 __ | 7 __ | . __ |
| Frangula alnus | 1.5 | 80 29.4 | 36 __ | . __ | 9 __ | . __ | 50 __ | 23 __ | 100 43.9 | 54 10.8 |
| Quercus petraea | 0.9 | . __ | 100 43.3 | 30 __ | 42 __ | . __ | 75 __ | 26 __ | 14 __ | 71 22.3 |
| Calamagrostis epigejos | 5.0 | . __ | 64 43.1 | . __ | . __ | . __ | . __ | 13 __ | 71 50.3 | 8 __ |
| Poa nemoralis | 2.3 | 20 __ | 45 29.0 | . __ | . __ | . __ | . __ | 51 34.6 | 7 __ | 17 __ |
| Fragaria vesca | 100.0 | . __ | 27 32.1 | . __ | . __ | . __ | . __ | 26 29.6 | . __ | . __ |
| Prunus avium | 1.7 | . __ | 64 41.1 | . __ | . __ | . __ | 12 __ | 49 27.5 | 29 __ | 12 __ |
| Luzula luzuloides | 1.9 | . __ | 41 33.8 | 10 __ | 3 __ | . __ | . __ | 15 __ | . __ | 29 20.6 |
| Moehringia trinervia | 2.2 | . __ | 36 21.9 | . __ | . __ | . __ | . __ | 13 __ | 64 50.0 | 17 __ |
| Pyrus communis agg. | 100.0 | . __ | 27 30.8 | . __ | . __ | . __ | . __ | . __ | 29 32.7 | . __ |
| Fallopia dumetorum | 100.0 | . __ | 27 34.1 | . __ | . __ | . __ | . __ | . __ | 21 25.0 | . __ |
| Rumex acetosella | 2.7 | . __ | 27 24.9 | 10 __ | . __ | . __ | . __ | 3 __ | 29 26.6 | 4 __ |
| Calamagrostis arundinacea | 1.4 | . __ | 45 26.1 | 10 __ | 9 __ | . __ | . __ | 33 __ | . __ | 58 38.1 |
| Cladonia macilenta | 6.6 | . __ | . __ | 70 53.2 | 9 __ | 60 43.4 | . __ | . __ | . __ | . __ |
| Calluna vulgaris | 1.7 | . __ | 18 __ | 80 43.8 | 48 18.4 | 80 43.8 | . __ | . __ | . __ | 4 __ |
| Vaccinium vitis-idaea | 4.4 | . __ | 14 __ | 60 26.2 | 61 26.7 | 100 58.0 | . __ | . __ | . __ | 8 __ |
| Dicranum scoparium | 1.5 | . __ | 9 __ | 40 __ | 61 31.5 | 60 31.0 | 12 __ | 8 __ | . __ | 17 __ |
| Fagus sylvatica | 1.4 | 20 __ | 14 __ | 80 34.1 | 48 __ | . __ | 25 __ | 38 __ | 14 __ | 67 24.2 |
| Pleurozium schreberi | 2.5 | . __ | 18 __ | . __ | 45 25.7 | 5 __ | 12 __ | 5 __ | 64 43.1 | 8 __ |
| Hypnum cupressiforme agg. | 1.2 | . __ | 50 __ | 50 __ | 61 24.5 | . __ | . __ | 59 23.2 | . __ | 42 __ |
| Lysimachia vulgaris | 3.7 | . __ | . __ | . __ | . __ | . __ | 38 30.5 | 10 __ | 7 __ | 42 35.2 |
| Corylus avellana | 1.2 | . __ | 32 __ | . __ | . __ | . __ | . __ | 49 33.9 | 14 __ | 38 22.7 |
| Acer pseudoplatanus | 1.9 | . __ | 23 __ | . __ | 6 __ | . __ | . __ | 77 48.5 | 29 __ | 54 28.8 |
| Oxalis acetosella | 100.0 | . __ | . __ | . __ | . __ | . __ | . __ | 33 31.9 | . __ | 42 42.5 |
| Stellaria media | 1.8 | . __ | 18 __ | . __ | . __ | . __ | . __ | 33 22.7 | 50 40.7 | 8 __ |
| Quercus robur | 1.9 | . __ | 5 __ | . __ | 27 __ | . __ | . __ | 64 32.2 | 93 55.7 | 33 __ |
| Quercus rubra | 2.2 | . __ | 9 __ | . __ | . __ | . __ | . __ | 15 __ | 43 35.4 | 33 24.8 |
| Carex pilulifera | 3.8 | . __ | . __ | . __ | . __ | . __ | . __ | 8 __ | 43 42.2 | 29 25.2 |
| Dryopteris carthusiana | 2.8 | . __ | 14 __ | 10 __ | 12 __ | . __ | 25 __ | 69 25.9 | 100 48.7 | 79 33.2 |
| Rubus fruticosus agg. | 3.0 | . __ | 77 25.8 | . __ | 6 __ | . __ | 25 __ | 87 32.9 | 100 42.1 | 75 24.2 |
| The most common accompanying species | ||||||||||
| Pinus sylvestris | 100 __ | 100 __ | 100 __ | 100 __ | 100 __ | 75 __ | 100 __ | 100 __ | 100 __ | |
| Betula pendula | 80 __ | 32 __ | 60 __ | 76 19.3 | 30 __ | 38 __ | 28 __ | 50 __ | 42 __ | |
| Sorbus aucuparia | 1.2 | 60 __ | 95 23.0 | 60 __ | 61 __ | 25 __ | 12 __ | 79 __ | 93 21.1 | 92 20.2 |
| Picea abies | 1.0 | 40 __ | 5 __ | 90 23.5 | 82 17.7 | 65 __ | 88 __ | 36 __ | 29 __ | 79 15.8 |
| Avenella flexuosa | 1.1 | . __ | 73 __ | 80 __ | 94 28.5 | 20 __ | 12 __ | 23 __ | 93 27.7 | 88 23.9 |
| Vaccinium myrtillus | 1.1 | . __ | 64 __ | 90 __ | 97 24.8 | 90 19.8 | 88 __ | 26 __ | 29 __ | 83 14.9 |
| Axis 1 | Axis 2 | |||
|---|---|---|---|---|
| rho1 | p2 | rho | p | |
| ElV_Light | 0.54 | 0.164 | −0.65 | 0.004 ** |
| EIV_Temperature | −0.68 | 0.044 * | −0.35 | 0.112 |
| EIV_Continentality | 0.63 | 0.064 | 0.13 | 0.52 |
| EIV_Moisture | −0.19 | 0.712 | 0.34 | 0.176 |
| EIV_Soil Reaction | −0.86 | 0.004 ** | −0.04 | 0.772 |
| EIV_Nutrients | −0.91 | 0.004 ** | 0.32 | 0.18 |
| Environmental Variable | Simple Term Effects | Conditional Effects | ||||
|---|---|---|---|---|---|---|
| Lambda | Pseudo-F | p | Lambda | Pseudo-F | p | |
| Altitude | 9.2 | 17.4 | 0.002 | 9.2 | 17.4 | 0.002 |
| Upper sandstone | 6.7 | 12.3 | 0.002 | 2.8 | 5.8 | 0.002 |
| Serpentine | 5.3 | 9.6 | 0.002 | 4.6 | 9.0 | 0.002 |
| Lower sandstone | 3.1 | 5.5 | 0.002 | 2.1 | 4.6 | 0.002 |
| Quaternary deposits | 3.0 | 5.3 | 0.002 | 0.7 | 1.5 | 0.022 |
| Limestone | 2.5 | 4.4 | 0.002 | 2.7 | 5.6 | 0.002 |
| Quaternary peat bogs | 2.4 | 4.3 | 0.002 | 2.6 | 5.5 | 0.002 |
| Metamorphic | 1.9 | 3.3 | 0.002 | 1.4 | 3.1 | 0.002 |
| Granitoids | 1.3 | 2.2 | 0.002 | 0.5 | 1.2 | 0.232 |
| Heat load index | 0.9 | 1.6 | 0.014 | 0.7 | 1.5 | 0.008 |
| Cluster No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Impatiens parviflora | - | 18 | - | - | - | - | 79 | 36 | 13 |
| Quercus rubra | - | 9 | - | - | - | - | 15 | 43 | 33 |
| Robinia pseudoacacia | - | 36 | - | - | - | - | 3 | 7 | - |
| Solidago gigantea | - | 5 | - | - | - | - | 3 | 7 | - |
| Prunus serotina | - | - | - | - | - | - | 10 | 86 | - |
| Pseudotsuga menziesii | - | - | 25 | 6 | - | - | - | - | - |
| Amelanchier spicata | - | - | - | - | - | - | 3 | 7 | - |
| Oxalis corniculata | - | - | - | - | - | - | 5 | - | - |
| Symphoricarpos albus | - | - | - | - | - | - | 5 | - | - |
| Pinus strobus | - | - | - | - | - | - | 3 | - | - |
| Parthenocissus inserta | - | - | - | - | - | - | 7 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reczyńska, K.; Pech, P.; Świerkosz, K. Phytosociological Analysis of Natural and Artificial Pine Forests of the Class Vaccinio-Piceetea Br.-Bl. in Br.-Bl. et al. 1939 in the Sudetes and Their Foreland (Bohemian Massif, Central Europe). Forests 2021, 12, 98. https://doi.org/10.3390/f12010098
Reczyńska K, Pech P, Świerkosz K. Phytosociological Analysis of Natural and Artificial Pine Forests of the Class Vaccinio-Piceetea Br.-Bl. in Br.-Bl. et al. 1939 in the Sudetes and Their Foreland (Bohemian Massif, Central Europe). Forests. 2021; 12(1):98. https://doi.org/10.3390/f12010098
Chicago/Turabian StyleReczyńska, Kamila, Paweł Pech, and Krzysztof Świerkosz. 2021. "Phytosociological Analysis of Natural and Artificial Pine Forests of the Class Vaccinio-Piceetea Br.-Bl. in Br.-Bl. et al. 1939 in the Sudetes and Their Foreland (Bohemian Massif, Central Europe)" Forests 12, no. 1: 98. https://doi.org/10.3390/f12010098