Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus


Department of Forest Ecosystems Protection, University of Agriculture in Krakow, Al. 29 Listopada 46, 31-425 Krakow, Poland
*
Author to whom correspondence should be addressed.
Forests 2021, 12(10), 1412; https://doi.org/10.3390/f12101412
Submission received: 30 August 2021
/
Revised: 9 October 2021
/
Accepted: 11 October 2021
/
Published: 16 October 2021
(This article belongs to the Section Forest Health)
Abstract
:Studies on fungal communities in the previous year’s leaf petioles of Fraxinus excelsior found in litter in five ash stands in southern Poland were made in 2017. Fungi were identified on the basis of isolation from 300 surface sterilized leaf petioles and by in situ inventory of fruit bodies (on 600 petioles, in spring and autumn). Identification was based on morphology of colonies and fruit bodies, and sequencing of ITS region of the rRNA gene cluster. In total, 2832 isolates from 117 taxa (Ascomycota—100; Basidiomycota—15; Mucoromycota—2 taxa) were obtained with the isolation method. The most frequent taxa (with frequency >10%) were: Nemania serpens, Hymenoscyphus fraxineus, Alternaria sp. 1, Boeremia sp., Helotiales sp. 1, Epicoccum nigrum, Venturia fraxini, Fusarium sp., Fusarium lateritium, Nemania diffusa, Typhula sp. 2 (in descending order). In total, 45 taxa were detected with the in situ inventory method. Eleven taxa were classified as dominant: Hymenoscyphus fraxineus, Venturia fraxini, Leptosphaeria sp. 2, Cyathicula fraxinophila, Typhula sp. 2, Hypoderma rubi, Pyrenopeziza petiolaris, Cyathicula coronata, Hymenoscyphus scutula, Leptosphaeria sclerotioides and Hymenoscyphus caudatus. Among 202 leaf petioles colonized by H. fraxineus, 177 petioles also showed fructification of 26 other fungi. All the isolated saprotrophs were tested in dual-culture assay for antagonism to two strains of H. fraxineus. Three interaction types were observed: type A, mutual direct contact, when the two fungi meet along the contact line (occurred with 43.3% of test fungi); type B, with inhibition zone between colonies (with 46.9% of test fungi); type C, when the test fungus overgrows the colony of H. fraxineus (with 9.8% of test fungi). The possible contribution of the fungal saprotrophs in limiting of the expansion of H. fraxineus in ash leaf petioles, which may result in reduction in the inoculum of ash dieback causal agent, is discussed.
1. Introduction
European ash (Fraxinus excelsior L.) started to show symptoms of serious disease in the early 1990s and, currently, dieback can be seen in most of its range in Europe [1,2,3,4,5,6,7,8]. The disease is caused by an alien ascomycete, Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz & Hosoya, which probably came from Eastern Asia where it occurs as an endophyte, extensive leaf colonizer and, locally, as a leaf pathogen of Fraxinus mandshurica Rupr. and Fraxinus chinensis Roxb. ssp. rhynchophylla (Hance) A. E. Murray [9,10,11]. The role of leaves, mainly petioles, as sites of infection in the H. fraxineus life cycle and ash dieback has been emphasized. Each year H. fraxineus forms apothecia, predominantly on the previous year’s leaf petioles and leaflet veins, in the litter on the forest floor [5,7,8]. In Poland, apothecia of H. fraxineus are formed mainly in July and August. However, in some years, they can also be found in September and October [12]. In other European regions, apothecia can be found much earlier, from the middle of May [13].
Wind-disseminated ascospores of H. fraxineus infect leaves of F. excelsior, and both leaf blades and leaf petioles have been confirmed as targets for primary infections [14]. The fungus is able to colonize all types of ash-leaf cells. In ash leaf petioles, its hyphae were observed in xylem vessels, phloem, axial parenchyma cells and also in the pith region [14].
After infection, the first symptoms in the form of local necrotic lesions appear on leaf blades, midribs and petioles [14]. The fungus can spread to woody tissues through the petiole-shoot junction prior to leaf fall [15]. The first symptoms are followed by dieback of shoots, twigs or main stems, resulting in crown dieback, bark lesions and sapwood discoloration.
During colonization of fallen leaves, H. fraxineus forms pseudosclerotial plates on petioles and some leaflet veins. Numerous new apothecia are formed on them in the following year, starting from early summer and completing the pathogen’s one-year life cycle associated with leaves [5,8,16]. On shoots and stems, apothecia of H. fraxineus are formed only sporadically and have no significance in the development of epidemics [5,7,8,17,18].
Options for preventing, inhibiting or mitigating disease and damage caused by H. fraxineus in forest stands are very limited [8,19,20,21]. There are, however, some environmentally friendly alternatives to chemical control.
Considering the significance of petioles as a necessary substrate for the formation of apothecia and production of H. fraxineus inoculum, there is the possibility of removing (burning or composting) fallen leaves. Such methods may be possible in tree nurseries or town greenery but not, however, in forest stands [22]. In the latter, creating an environment suitable for faster decomposition of ash leaves and debris may be considered [19,20].
Another alternative considered increasingly often is microbial biocontrol, similar to that used with some success against diseases of some agriculturally important herbaceous plants and forest trees [23,24,25]. Several common Fraxinus leaf endophytes (e.g., Boeremia exigua, Kretzschmaria deusta, Paraconiothyrium sp., Neofabraea alba and Venturia fraxini) have been shown able to reduce germination of H. fraxineus ascospores [26], and some F. excelsior leaf and stem endophytes (e.g., B. exigua, Epicoccum nigrum, Hypoxylon rubiginosum and Phoma macrostoma) can inhibit growth of H. fraxineus hyphae [27,28,29,30]. Certain forest fungal communities can stimulate decomposition of litter and may be employed in the long term for indirect microbial control. The required quality of litter and its desirable microbial status can be achieved in certain conditions or after certain physical and chemical treatments [20,31,32,33,34,35]. For example, higher concentrations of nitrogen (N) and phosphorus (P) in litter will promote its decomposition, while a higher concentration of lignin (difficult to degrade) will often inhibit its degradation and removal [35,36,37]. Urea generally accelerates the decomposition of leaf debris [19] and additionally prevents the formation of H. fraxineus apothecia (important for dissemination of spores).
Leaves of F. excelsior are easily degraded compared with leaves of other broad-leaved trees in temperate forests [20,38,39] because of their lower C/N ratio and lower lignin content [40,41,42]. However, F. excelsior leaf petioles, which constitute 20% of leaf biomass, are more resistant to decomposition [20,43]. Starch concentration, considerably higher in ash petioles than in leaflets [43], ensures continuously favorable conditions for fungi. Additionally, infected petioles are the substrate for formation of pseudosclerotial plates of H. fraxineus which protect the fungus from biotic and abiotic stresses [16].
Fungi are the key players in litter decomposition and nutrient cycling owing to their ability to produce a wide range of extracellular enzymes which allow them to attack efficiently the recalcitrant lignocellulose matrix [44,45,46,47,48,49,50,51].
There are only few reports from qualitative and quantitative mycological analyses of F. excelsior petioles from litter [52,53]. More studies have been made on endophytes in petioles of living leaves, with and without symptoms, aimed at detecting possible pathogens [54,55,56].
The fungi of most relevance in leaf petioles from the litter are those which are antagonistic and suppressive to H. fraxineus, inhibiting the pathogen’s growth and reducing inoculum production. Such activity depends on colonization strategies and relationships among different members of the fungal community [57,58]. This subject needs careful investigation, as in other studies on endophytes [27,28,54].
The objectives here were to evaluate: (i) fungal species diversity in leaf petioles of F. excelsior from the previous year, found in the litter, using the classical isolation method and an inventory made in situ, based on the occurrence of fruit bodies; and (ii) interactions between H. fraxineus and co-occurring members of fungal communities, in dual cultures, in vitro, and any reduction in growth rate in each fungus. An additional objective was to assess the risk of leaf petioles of F. excelsior being exploited as a substrate for production of inoculum by other pathogens of trees and annual plants.
2. Materials and Methods
2.1. Study Sites and Sampling
The study sites were five stands of F. excelsior, 27–82 years old, in Brody, Dynów, Miechów, Myślenice and Ojców, located in southern Poland (Table 1). Fraxinus excelsior was growing in mixed populations with other trees, mostly broad-leaved, and comprised 10–80% of the total of trees. The ash trees were showing symptoms of dieback similar to those observed over many years in other F. excelsior stands [59]. Leaf petioles from the previous year were collected from the litter on the forest floor between 10 May and 8 June 2017 (Table 1). A total of 6 petioles were collected beneath each of 30 trees located in various parts of each stand (180 petioles per stand). On the next day, in the laboratory, each sample was randomly divided into three subsamples, each of 60 petioles. Petioles were stored temporarily in sterile plastic boxes at 5 °C. Subsamples (I, II, III) were treated differently.
2.2. Fungal Isolation and Identification
Subsample I was used for isolation of fungi and identification based on colony morphology (Table 1). In total, isolations were made from 300 petioles (3600 pieces). After removing the remains of leaflets, petioles were surface sterilized by soaking first for 1 min in 96% ethanol, then for 5 min in sodium hypochlorite solution (NaOCl, approx. 4% available chlorine) and finally for 30 s in 96% ethanol. After drying in layers of blotting paper, each petiole was divided into 12 small, equally sized pieces which were placed on the surface of malt extract agar (MEA; 20 g L−1 malt extract (Difco; Sparks, MD, USA), 15 g L−1 Difco agar) supplemented with 100 mg L−1 streptomycin sulphate to suppress bacterial growth, in two Petri dishes. The sequence of petiole pieces (1–12, from base to apex) was marked on the Petri dish cover, 1–6 and 7–12. The plates were incubated in darkness at 20 °C for 42 d and examined every 1–2 weeks. Emerging individual mycelia were regularly transferred to sterile Petri dishes with MEA and incubated for 3–4 weeks. Initial identification of fungi was based on morphology, which included color and structure of the colony, presence of stromatic structures and the type of anamorphic conidiation and conidia. Isolates were grouped in morphotypes [60]. Morphology of H. fraxineus was compared with descriptions in previous studies [2,61,62].
2.3. In Situ Inventory of Fungi from Fruit Bodies on Petioles
This was based on the presence of fruit bodies and survival structures formed on leaf petioles, either in May–June or from the beginning of September to the end of November, in subsamples II and III, respectively [63]. Petioles from subsample II were stored at 5 °C and analyzed as quickly as possible.
Petioles from subsample III were laid out, avoiding contact between petioles from different stands, on the surface of a fenced meadow which was moderately shaded by trees and bushes (with no Fraxinus) and moderately humid, with extensive patches of mosses, particularly of Atrichum undulatum Web. et. Mohr. and Pleurozium schreberi (Willd. ex Brid.) Mitt., among grasses. They were examined with a magnifying lens every 10–14 days. Petioles with fructification were placed on moist filter paper in plastic boxes and transferred to the laboratory for subsequent mycological analysis. Representative, strongly colonized petioles were deposited in an herbarium. Most petioles, however, were marked with a number and returned to the meadow for further fructification. In results, assessments in May–June (subsample II) and September–November (subsample III) are referred to as “spring” and “autumn”, respectively.
Identification of H. fraxineus was based on the presence of pseudosclerotial plates or the pathogen’s anamorph (in subsample II) [16,62,64] or of apothecia formed on pseudosclerotial plates, naturally, in July–August (in subsample III). Identification of the genus Typhula was based on the presence of basidiocarps or ungerminated sclerotia [65,66].
Observations and photomicrographs were made with a Zeiss V12 Discovery stereo microscope and with a Zeiss Axiophot light microscope using differential interference contrast (DIC) illumination and an AxioCam MRc5 and HR3 cameras (Zeiss, Göttingen, Germany). For microscopic analysis, hand-cut sections and 1–3 slides were prepared from each fruit body being studied. Fungi were classified and identified to the lowest taxonomic rank using mycological keys and monographs.
Isolation as pure cultures was attempted on each differentiated taxon, from ascospores formed in perithecia/pseudothecia, basidiospores formed in basidiocarps (for Marasmius, Typhula), conidia formed in conidiomata or sclerotia, which were spread or placed on MEA, either in a Petri dish or inside a Petri dish cover, and incubated at 20 °C [67].
Cultures representative of taxa distinguished on the basis of fruit bodies or survival structures were analyzed with the nucleotide sequences of ITS region of the rRNA gene cluster.
The term “petiole” is used for petiole and rachis. The petiole was considered as colonized by a certain taxon if it was isolated from at least 1 of 12 petiole pieces. Representative fungal isolates are stored on MEA slants at 5 °C in the Culture Collection, and exsiccates of fungi in Mycotheca of the Department of Forest Ecosystems Protection, University of Agriculture, in Kraków. The nomenclature of taxa follows Index Fungorum [68].
2.4. DNA Extraction, PCR and Sequencing
To verify the morphology-based identification and to identify non-sporulating morphotypes, the nucleotide sequences of the ITS region of the rRNA gene cluster of representative cultures were determined.
DNA was extracted from 3-week-old cultures using the Genomic Mini AX Plant Kit (A&A Biotechnology, Gdynia, Poland) according to the manufacturer’s protocol. The ITS region of the rRNA gene cluster was amplified for sequencing. Primers used were ITS1-F [69] and ITS4 [70] or ITS5 and ITS4 [70]. Gene fragments were amplified in 25 µL reaction mixture containing 0.25 µL Phusion High-Fidelity DNA Polymerase (Thermo Fisher Scientific Baltics UAB, Vilnius, Lithuania), 5 µL Phusion HF Buffer (5x), 0.5 µL dNTPs (10 mM), 0.75 µL DMSO (100%), 0.5 µL of each primer (25 µM) and 20–100 ng μL−1 of template DNA. PCR amplification was performed in a Labcycler thermocycler (SensoQuest Biomedical Electronics GmbH, Göttingen, Germany) under the following conditions: an initial denaturation step at 98 °C for 30 s followed by 35 cycles of 5 s at 98 °C, 10 s at 52–64 °C (depending on the optimal Tm of the primers and fungal species) and 30 s at 72 °C, and a final elongation at 72 °C for 8 min. The PCR products were visualized on 2% agarose gel stained with Midori Green DNA Stain (Nippon Genetics Europe) under UV light. Amplified products were sequenced bi-directionally with the BigDye® Terminator v 3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) and ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA), at the DNA Research Centre (Poznań, Poland) using the same primers as for the PCR.
Searches using the BLASTn algorithm were performed to retrieve similar sequences from NCBI GenBank (http://www.ncbi.nlm.nih.gov accessed on 2 August 2021) [71]. Newly obtained sequences were deposited in GenBank with accession numbers presented in Supplementary Material Table S1.
Sequence matches of ≥98% were used to define species boundaries and matches ≥90% were used to define order boundaries [72]. If sequencing of the ITS region of the rRNA gene cluster was ambiguous, the taxonomy was determined by phylogenetic analysis with reference sequences from NCBI GenBank.
The ITS region of the rRNA gene cluster sequences were aligned using MAFFT v.6 [73] with the E-INS-i option, a gap-opening penalty of 1.53 and an offset value of 0.00. The alignments were checked manually with BioEdit v.2.7.5 [74].
Phylogenetic analyses were performed individually, for each dataset, using maximum likelihood (ML) and Bayesian inference (BI). The best-fit substitution models for dataset were established for ML and BI using the corrected Akaike Information Criterion (AICc) in jModelTest v.2.1.10 [75,76]. Maximum likelihood (ML) analysis was run in PhyML 3.0 [77] via the Montpelier online server (http://www.atgc-montpellier.fr/phyml/ (accessed on 2 August 2021)) with 1000 bootstrap replicates. The best evolutionary substitution model for the ITS region of the rRNA gene cluster was GTR+I+G. The BI analysis was carried out in MrBayes v.3.1.2 [78]. The MCMC chains were run for 10 million generations using the best-fit model. Trees were sampled every 100 generations, resulting in 100,000 trees from both runs. The default burnin, first 25% of samples, was used. The resulting phylogenetic trees (Supplementary Material Figure S1) contain all the ITS region of the rRNA gene cluster sequences generated in this study.
2.5. Dual-Culture Assays
Representative fungi of each morphotype from subsample I were screened for their ability to suppress mycelial growth of two isolates of H. fraxineus by in vitro dual-culture assays on malt extract agar (MEA).
The H. fraxineus isolates were obtained in 2017 from the previous year’s leaf petioles with pseudosclerotia found in the litter, in two different F. excelsior stands in southern Poland. Both isolates (Hf1 and Hf2) were characterized by moderately quick and even growth (35 and 39 mm on MEA after 3 weeks at 20 °C).
Plugs (8 mm diameter.) of young, actively growing mycelium of H. fraxineus and the test fungus were placed simultaneously on MEA in a Petri dish, opposite each other and 25 mm from the Petri dish edge. Plates inoculated only with H. fraxineus served as negative controls. Fungal growth was measured after incubation for 3 weeks in darkness at 20 °C. The measurements were: (i) radial mycelial growth of H. fraxineus (Ri) and of the saprotrophic test fungus (Rs) taken along an axis joining the centers of the plugs of the two fungi; (ii) radial mycelial growth of H. fraxineus on the control plate (Rc); (iii) the average of the maximum radial mycelial growth in the upper and lower parts of the test-fungus colony (Rm). Percentage inhibition of radial growth of H. fraxineus and of the test fungus was calculated according to the formulae (Rc − Ri)/Rc × 100 and (Rm − Rs)/Rm × 100, respectively [79,80]. The width of the inhibition zone (mm) was measured along the axis joining the plugs of the two fungi. Growth was measured with a ruler to an accuracy of 1 mm.
Previous observations showed that four types of interactions between H. fraxineus and a test fungus can be expected: type A, mutual direct contact, when both colonies meet along the contact line, with no inhibition zone; type B, with an inhibition zone between colonies; type C, the test fungus overgrows and covers the colony of H. fraxineus; type D, the H. fraxineus colony overgrows the colony of the test fungus [81]. For type B, the width of inhibition zone was scored as: Bs, <3 mm; Bm, 4–5 mm; Bw, 6–8 mm; Bv, >8 mm. Inhibition of mycelial growth was scored according to the amount of decreased growth as: f, no reduction; a, <25%; b, 26–50%; c, 51–75%; d, >75%.
When a test fungus overgrew and covered the H. fraxineus colony, its efficacy against of H. fraxineus was checked using re-isolations of the fungus (from 6–12 inocula taken from each combination of pathogen and test fungus).
2.6. Data Analyses
Frequency of an individual taxon was defined as percentage (%) of petioles colonized by that taxon. Diversity in microfungal communities from the five stands was analyzed with Shannon diversity index (H’) and Simpson’s diversity index (SDI) [82,83]. Species dominance in microfungal communities was determined with the Camargo method [84].
The association between frequency of fungi and their origin was analyzed using principal component analysis (PCA). PCA was performed for species with at least 10% frequency in subsample I (communities isolated from the previous year’s leaf petioles) and 5% frequency in subsamples II and III (detected as fruit bodies on petioles).
The same dataset was used to determine whether the differences in the occurrence of fungi on particular stands were statistically significant using the Marascuilo procedure. Similarities in fungal communities from the five stands were analyzed using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA), clustering based on the Bray–Curtis similarity index.
Co-occurrence of taxa in two fungal communities isolated from leaf petioles (subsample I) or detected as fruit bodies (subsamples II and III) was analyzed using a co-occurrence network created with Gephi visualization software v.0.9.2 combined with the Fruchterman–Reingold layout or the Yifan Hu proportional layout graph algorithm [85], after previous calculation of Spearman’s rank correlation coefficient and statistically significant difference (p < 0.05). All calculations were made using Statistica v.12 [86], PAST v.3.11 [87] and Marascuilo procedure in R software v.3.5.1 [88].
3. Results
3.1. Fungi Isolated from the Previous Year’s Leaf Petioles
All 300 leaf petioles of F. excelsior from the previous year and collected from the litter yielded fungi. A total of 2832 cultures was successfully isolated from 2501 petiole pieces (69.5% of the 3600 pieces) on MEA. Cultures were obtained slightly more frequently from basal petiole pieces (72.2%) than from pieces nearer the apex (66.7%). A total of 164 morphotypes was distinguished (Supplementary Material Table S1). Since a single taxon could be represented by 1–5 visually different morphotypes, sequencing of ITS region of the rRNA gene cluster of representative isolates resulted in detection of 117 fungal taxa (Supplementary Material Tables S1 and S2). The number of taxa detected in a single stand ranged from 53 (Brody) to 70 (Dynów) (Table 2).
The fungi isolated belonged to three phyla: Mucoromycota (1.7%), Ascomycota (85.5%) and Basidiomycota (12.8%) (Supplementary Material Table S2). Mucoromycota was represented by two taxa: Mortierella and Umbelopsis (Table 2). Ascomycota was represented by 100 taxa belonging to 19 orders. Pleosporales, Helotiales and Xylariales were represented by 32, 15 and 8 species, respectively, and were dominant in the fungal community. Eight orders were each represented only by one species (Figure 1 and Supplementary Material Table S2). The highest species diversity was in Leptosphaeria and Phoma (five species), and Alternaria, Coniochaeta and Hymenoscyphus (four species each) (Table 2). Basidiomycota was represented by 15 taxa belonging to six orders. Typhula sp. 2 (11.0%), Peniophora incarnata (5.0%), as well as Agaricales and Russulales, were the most frequent (Figure 1 and Table 2). Yeast-like basidiomycetes were represented by two species from the genera Fellozyma (Pucciniomycotina) and Tilletiopsis (Ustilagomycotina). Ten taxa were detected only in 1–2 petioles.
Among 117 taxa identified, 27 taxa were classified as dominants (Table 2). The most frequent taxa (>10%) were: Nemania serpens, Hymenoscyphus fraxineus, Alternaria sp. 1, Boeremia sp., Helotiales sp. 1, Epicoccum nigrum, Venturia fraxini, Fusarium sp., Fusarium lateritium, Nemania diffusa and Typhula sp. 2 (Table 2).
A total of 18 of the detected taxa occurred in all five stands, while 35 taxa occurred only in one stand (Table 2). The Shannon diversity index (H’) and Simpson’s diversity index (SDI), indicating diversity in all five fungal communities, were H’ = 3.86 and SDI = 0.96. There most diversity was at Dynów (H’ = 3.67, SDI = 0.96) and least at Ojców (SDI = 0.93) and Brody (H′ = 3.31).
Principal component analysis (PCA) separated the localities along the first axis (explaining 67.97% of the variation), mainly on the basis of prevalence of Nemania serpens, H. fraxineus, Helotiales sp. 1 and other dominant species (Figure 2). The second PCA axis explained 13.87% of the variation and separated Myślenice from other locations. This axis separated locations mainly on the basis of prevalence of Epicoccum nigrum and the other dominant species, e.g., Alternaria sp. 1, Boeremia sp. and Nemania diffusa (Figure 2). The difference in the prevalence of E. nigrum at Myślenice and at Ojców and Dynów was statistically supported by the Marascuilo procedure. Similarly, N. serpens was significantly more frequent at Ojców than at the other locations, whereas H. fraxineus was significantly more frequent at Dynów and Miechów than at other locations (Table 2).
The dendrogram based on the Bray–Curtis index showed similarity between fungal communities in stands at Dynów and Miechów and dissimilarity between them and communities at Ojców, Brody and Myślenice (Figure 3).
Analysis of co-occurrence of fungi in communities isolated from the previous year’s leaf petioles (subsample I) made with Spearman’s rank correlation coefficient showed 233 relationships that were statistically significant, including 129 positive and 104 negative correlations. There were five modules of co-occurrence. The first consisted of 16 taxa and included mostly F. lateritium, N. diffusa and Typhula sp. 2. The second consisted of 21 taxa and included mostly Epicoccum nigrum. The third consisted of 14 taxa and included mostly Boeremia sp. The fourth consisted of 16 taxa and included mostly Alternaria sp. 1. The fifth consisted of 14 taxa and included mostly Helotiales sp. 1, H. fraxineus and N. serpens (Figure 4).
Taxa with the most frequent relationships with others were Cladosporium sp. 1, Helotiales sp. 1, Microsphaeropsis sp. and Xylaria polymorpha. Co-occurrences of Cladosporium sp. 1 with other taxa were equally positively or negatively correlated (with slightly higher values for the positive correlations). Microsphaeropsis sp. had similar numbers of positive and negative correlations. Helotiales sp. 1 and X. polymorpha had a positive correlation, but occurrences of Helotiales sp. 1 or X. polymorpha with other fungi were often negatively correlated. A negative correlation suggests that increased frequency of one taxon causes decreased frequency of another. A good example is the co-occurrence of Helotiales sp. 1 and H. fraxineus, the latter decreasing in the presence of the former; this is the only negative statistically significant correlation for co-occurrence of H. fraxineus with other fungi (Figure 4).
3.2. In Situ Inventory of Fungi Based on Fruit Bodies on Petioles
A total of 45 taxa was detected from fruit bodies present on the 600 previous year’s leaf petioles (subsamples II and III) (Figure 5 and Table 3). All 45 taxa were cultured and subject to molecular analysis. Twenty-three of them had been isolated previously from petioles (subsample I). Twenty-two taxa were additional and extended the list of fungi from petioles (Table 2 and Table 3). The fungi detected were Ascomycota and Basidiomycota (Figure 5). Ascomycota was represented by 41 taxa (91.1%) from nine orders. Those in the order Helotiales were most frequent (51.0%) and had most diversity (17 species) (Figure 5 and Supplementary Material Table S2).
The high level of taxonomic diversity was demonstrated by the diversity of fruit bodies on the dead petioles (Figure 6a–t). Colonization of petioles by certain fungi was associated with discoloration of the surface of the colonized petiole pieces, e.g., Hymenoscyphus scutula, Helotiales sp. 1, Hypoderma rubi and L. sclerotioides caused, respectively, grey-brown, rusty-brown, light-cream or dark-grey discoloration (Figure 6d,e,g,j).
Basidiomycota were represented by four taxa, all from the order Agaricales (Figure 5, Table 3 and Supplementary Material Table S2).
The formation of fruit bodies and sclerotia of some fungi, including H. scutula, Pyrenochaeta sp. or Typhula sp. 2, may contribute to loosening and desquamation of the peripheral tissue of petioles (Figure 6d,o,r,s). Additionally, the petiole’s surface tissues may be eaten by small animals living in the litter, exposing the inner tissues to colonization but also causing loss of taxa that fruit on the eaten tissue edges (Figure 6n).
Among 45 taxa identified, 11 were classified as dominants (Table 3). This group included Hymenoscyphus fraxineus, Venturia fraxini, Leptosphaeria sp. 2, Cyathicula fraxinophila, Typhula sp. 2, Hypoderma rubi and Pyrenopeziza petiolaris (in descending order, with frequency 33.7%–5.2%).
A total of 8 of the detected taxa occurred in all five stands, while 13 occurred only in one stand (Table 3). The numbers of taxa detected in a single stand ranged from 17 (Miechów) to 31 (Ojców) (Table 3).
The Shannon diversity index (H′) and Simpson’s diversity index (SDI), indicating the diversity over all five stands, were H′ = 2.73 and SDI = 0.89. There was most diversity at Ojców (H′ = 2.82, SDI = 0.91) and least at Miechów (H′ = 1.99, SDI = 0.79) (Table 3).
PCA separated the localities along the first axis (explaining 73.98% of the variation), mainly on the basis of the prevalence of H. fraxineus. The second PCA axis explained 16.69% of the variation and separated Ojców from the other locations. This axis separated locations mainly on the basis of the prevalence of Leptosphaeria sp. 2 as well as the other dominant species, H. fraxineus, C. fraxinophila and V. fraxini (Supplementary Material Figure S2). The difference in prevalence of H. fraxineus at Miechów from that at Brody, Ojców and Dynów was statistically supported by the Marascuilo procedure. Similarly, Leptosphaeria sp. 2 was statistically more frequent at Myślenice than at Ojców (Table 3).
The dendrogram based on the Bray–Curtis index showed similarity between fungal communities in stands at Dynów and Miechów and dissimilarity between them and communities in stands at Ojców, Brody and Myślenice (Supplementary Material Figure S3). These results support those of analysis based on isolation of fungi from petioles (subsample I).
The fungi detected varied depending on their time of fructification on F. excelsior petioles (Figure 7). Six taxa, including P. petiolaris, fruited only in spring (Figure 7). Thirteen taxa fruited in both spring and autumn, with variable frequency observed in some of them, particularly V. fraxini (Figure 7). Only H. rubi fruited in spring and autumn with similar frequency, although in spring the fungus produced only immature hysterothecia (as well as the anamorph) (Figure 6e). Most fungi, including C. coronata, C. fraxinophila, H. caudatus, H. scutula and Pyrenochaeta sp., fruited in autumn (Figure 7). The autumn-fruiting group also includes basidiomycetous Typhula spp. It should be emphasized, however, that, in spring, Typhula formed only sclerotia with no sporophores (Figure 7).
In spring, identification of H. fraxineus was based on the presence of pseudosclerotial plates with phialides and endoconidia (Figure 8a–c). Apothecia appeared on pseudosclerotial plates at the beginning of July (Figure 8d). Careful morphological analysis excluded the presence of the related species, H. albidus.
Fruit bodies of 26 other fungal species were present on 177 out of 202 leaf petioles colonized by H. fraxineus (Table 3). One, two and three additional taxa co-occurred on 124, 42 and 11 petioles, respectively. Usually, the petiole pieces colonized by them were visually different, being distinctly brighter than those with H. fraxineus pseudosclerotia (Figure 8a–o). On one petiole, Cyathicula coronata produced its apothecium on the H. fraxineus pseudosclerotium (Figure 8o).
Observation showed that H. fraxineus may co-exist spatially in ash petioles with V. fraxini in a way that is different from its co-existence with some other fungi. Pseudosclerotia of H. fraxineus were formed subepidermally and appeared in late spring, after desquamation of the epidermis (Figure 8a,b), which led to the removal of pseudothecia of V. fraxini formed in the epidermis earlier. Desquamation of the epidermis could be caused by H. fraxineus, but also by other physical and biological factors (Figure 6m,n).
Co-occurrence of fungi was positive or negative (data not shown) and statistically significant if strongly positively correlated. Increased frequency of H. fraxineus was associated with increased frequency of Leptosphaeria sclerotioides, Leptospora rubella and Paraophiobolus arundinis (Supplementary Material Figure S4 and Table 3).
3.3. Dual-Culture Assays
Three types of interaction (A, B, C) were observed on MEA in dual-culture assays with either of two isolates of H. fraxineus, originating from two stands of F. excelsior, and 117 fungal species representing 164 morphotypes, isolated from the previous year’s leaf petioles (Figure 9a–t and Table 4, Supplementary Material Table S1). The fourth type (D) listed in Methods did not occur (Table 4, Supplementary Material Table S1). The types occurred with different frequencies. Types A and B were significantly more frequent than type C (Table 4). The difference in frequency between type A and type B was statistically non-significant. The frequency of any type did not depend on the H. fraxineus isolate used (Table 4). One species of test fungus was represented by 1–5 morphotypes (Supplementary Material Table S1). Differences in interaction type between different morphotypes of one species were sporadic and often temporary, e.g., D. acicola FeF42 had an inhibition zone only for the first 3 weeks, then covered the zone and finally had an effect similar to that observed with D. acicola FeF119 (Supplementary Material Table S1).
In type A interactions, the size of the H. fraxineus colony and the shape of the contact line between two colonies often depended on the rate of growth of the test fungus (Figure 9b–d).
Type B interactions, the formation of the inhibition zone, were the most common. The most frequent width of the inhibition zone was Bs, and the least frequent was Bv. The Bs width (Figure 9e,f) occurred significantly more frequently than Bw or Bv (Table 4).
Very wide inhibition zones (>8 mm) were observed in 6.4% of cases, in pairings of H. fraxineus with Coniochaeta angustispora, Cytospora pruinosa, Fusarium sp., Lanzia sp. FeF26, Malbranchea sp. (inhibition zone 16 mm), Peziza ninguis, Plenodomus sp. 1, Pseudocoleophoma polygonicola (inhibition zone 27 mm) or Pseudoophiobolus italicus (Figure 9j–l and Supplementary Material Table S1). Visible morphological and physiological changes in colonies were often observed: Hypoderma rubi produced intensive, brownish-red pigment at the border with H. fraxineus (Figure 9g); Lanzia sp. FeF26 caused the formation of blackish areas in H. fraxineus colonies, possibly from melanization of the pathogen’s hyphae (Figure 9j); Plenodomus sp. 1 or Pleospora sp. FeF80 caused the production of a black, melanin-like substance which stained the agar (Figure 9h).
Type C interactions, in which the test fungus overgrows the colony of H. fraxineus, occurred in assays with morphotypes of 12 taxa: Clonostachys rosea, Desmazierella acicola FeF119 (thin cover), Hypholoma fasciculare, Lasiosphaeriaceae sp. (thin cover), Nemania diffusa, N. serpens, Peniophora incarnata, Rosellinia corticium, R. nectrioides, Trichoderma viride, Xylaria polymorpha and Xylaria sp. 1 (Figure 9m–t and Supplementary Material Table S1). This type of interaction was often complex and preceded by: (i) earlier, considerable inhibition of the H. fraxineus colony; (ii) formation of an inhibition zone caused by accumulation of non-volatile metabolites produced by H. fraxineus (Figure 9p); and (iii) overgrowth of the inhibition zone (Figure 9p). Hypholoma fasciculare and T. viride entirely covered the inhibition zone and the H. fraxineus colony after 3 weeks (Figure 9t), while Lasiosphaeriaceae sp. and D. acicola FeF119 covered them only slightly. Rosellinia abscondita started to cover the narrow inhibition zone (width Bs < 3 mm) and the H. fraxineus colony after 4 weeks, and so could not be assessed.
Only the test fungi were re-isolated from colonies of H. fraxineus overgrown by the test fungi.
The reduction in growth rate of H. fraxineus mycelium, in the region growing towards the test fungus, was determined in all dual-culture combinations by comparison with the control. Amounts of reduction differed. Most often it ranged between 51 and 75% and only sporadically was >75% (Table 5). The origin of H. fraxineus did not affect the amount of inhibition (Table 4 and Table 5). Two statistically different groups of taxa were obtained according to the amount of growth reduction they caused in H. fraxineus. The first group included taxa causing 26–50 or 51%–75% reduction; the second group included other taxa (Table 5). A reduction in the growth of the saprotrophic partners towards H. fraxineus was observed in 90.0% of dual cultures. Most often (61.3%), the reduction rate was between 26 and 50% (Table 5).
4. Discussion
4.1. Fungal Diversity
Analyses of fungal communities from dead leaf petioles of F. excelsior from five locations show that they have high diversity of both common and rare species and are comparable with fungal communities from other tree species. Taxonomically, the fungi detected belonged mostly to the Ascomycota, less often to the Basidiomycota and occasionally to the Mucoromycota. The results are consistent with earlier findings on fungal communities from litter [47,48,50,51,89,90], although the relative occurrences of Ascomycota and Basidiomycota depend on the stage of decomposition of the substrate. Ascomycota are usually relatively more abundant in the early stages of decomposition while Basidiomycota increase over time, becoming most frequent at the advanced decomposition stage [48,51,89,90,91]. Ascomycetous fungi can even decrease the rate of decomposition by exclusion of secondary saprotrophic Basidiomycota, so affecting the microbiological status of woody tissues [34].
The classical method of isolation of fungi on agar medium led to detection of more taxa than the in situ inventory of fungi based on the presence of fruit bodies. The latter method supplemented the former. There were, however, a few taxa, such as Cyathicula fraxinophila and Leptosphaeria sp. 2, which were detected more often in situ. This could have resulted from the possible domination of the successful fungus or from the method of disinfection used. The fungus detected might be faster growing in vitro, especially from the surface epidermal and subepidermal layers, and resistant to the disinfectants used [92].
A total of 117 taxa was detected by isolation, and 45 taxa by in situ inventory based on the presence of fruit bodies. Among the 45 taxa producing fruit bodies on petioles, 23 were also isolated on agar medium. Therefore, a total of 139 taxa was detected on the dead leaf petioles of F. excelsior in litter.
The difference between the two methods in number of taxa detected may be expected. Among the reasons for fewer taxa being detected in situ are that: (i) some fungi do not produce fruit bodies in their life cycle, (ii) some fungi produce filamenous anamorphs rather than teleomorphs, and (iii) yeast forms may dominate in situ [93]. Some taxa detected produce fruit bodies on ash leaves less often than on other substrates, such as tree logs, branches and stumps (e.g., Coniochaeta, Nemania, Peniophora, Trametes, Xylaria) or on tree stems and twigs (e.g., Cytospora pruinosa, Diplodia fraxini) [94,95,96,97,98].
The ascomyceous taxa detected belong to 20 orders, indicating high diversity among the Ascomycota. Most diversity was observed in the Pleosporales, which were comparably abundant and diverse as endophytes in living ash leaves [29,81,99]. The findings support those of Hyde et al. [100], Promputtha et al. [101] and De Silva et al. [102], which showed that some fungal species associated with living leaves were also found in leaf litter. These are the host-specific fungal endophytes that most often switch to become saprotrophs [103]. Certain taxa found in the dead F. excelsior petioles, namely Boeremia exigua, Colletotrichum acutatum, Cytospora pruinosa, Diaporthe eres, Fusarium lateritium, Nemania diffusa, N. serpens, Venturia. fraxini and X. polymorpha, have been found previously in living, symptomless leaflets and petioles of F. excelsior [54,81,99].
Ascomycetous fungi detected in this study differed in their trophic character, host range, tissue specificity, lifestyle and significance in forests. Most frequent among the potential pathogens was Venturia fraxini which, in favorable conditions, can cause withering and premature leaf fall in F. excelsior stands [104,105], although in some regions it is a dominant species in symptomless leaflets or petioles of F. excelsior [81,99,106]. It is one of the rare groups of fungi that easily produce fruit bodies on ash petioles, with mature ascospores appearing very early, in May-June, before infectious ascospores of H. fraxineus. This predestines the fungus as a very early endophyte colonizing living leaves of F. excelsior. Both H. fraxineus and V. fraxini can survive in ash petioles in the litter and the early colonization of leaves by V. fraxini may contribute to temporary protection against H. fraxineus. This is not supported, however, by the assays in vitro and observations in situ. Venturia fraxini was not among the fungi most antagonistic to H. fraxineus: it colonizes the surface tissues of petioles which does not prevent H. fraxineus from colonizing other, empty spatial niches and formation of pseudosclerotia [16,106]. Other frequent potential pathogens were Boeremia exigua, Cytospora pruinosa and Diplodia fraxini. The first two were found previously on F. excelsior with symptoms of decline [55,97,107]. In dual cultures, both fungi were strongly antagonistic towards H. fraxineus which may suggest their effectiveness against H. fraxineus in nature. Diplodia fraxini (with pycnidia on the sampled petioles) was found on ash leaves for the first time. It has previously been associated with cankers and dieback of F. excelsior [98].
Most of the taxa detected in leaf petioles of F. excelsior are, however, known exclusively or generally to have a saprotrophic lifestyle on leaf residues of forest trees, dead wood or the remains of herbaceous plants. The group of the most frequent saprotrophs included representatives of the genera Cyathicula, Hypoderma, Hymenoscyphus and Pyrenopeziza, and the family Xylariaceae. Cyathicula species (Helotiales) are known from their common occurrence in ash leaf petioles [108,109]. These fungi were shown here not to have been eliminated by H. fraxineus from the petiole tissues. Cyathicula fraxinophila occurred most frequently. This fungus was often observed previously on ash petioles in the Czech Republic [53] but not in Ukraine [52]. It is host specific to F. excelsior and the epidemics of ash dieback in Europe may be associated with a decrease in its population. Another member of the genus, C. coronata, occurs on a wide range of herbaceous plants [108]. Hypoderma rubi has been reported on leaves, petioles and twigs of a wide range of woody plants, mostly dicotyledons; its main, most colonized host is Rubus [110]. It has been found on F. excelsior and other species of ash [111,112]. Its high frequency in dead leaf petioles of ash is shown and emphasized here. Unexpectedly frequent was also Pyrenopeziza petiolaris (Helotiales), which has been found previously on numerous broad-leaved trees (Acer, Aesculus, Ailanthus, Betula, Fagus, Populus, Sorbus and Tilia), particularly on Acer pseudoplatanus [109,113]. It is possible that A. pseudoplatanus occurred frequently in the sampled stands and so may have contributed to the frequent occurrence of P. petiolaris on F. excelsior. Apothecia of P. petiolaris mature in late spring, and the involvement of ascospores in early development of an endophytic phase in living leaves, followed by a saprotrophic life cycle and survival in dead leaf petioles until the following year, cannot be excluded.
Two members of the genus Hymenoscyphus found in this study, H. caudatus and H scutula, can colonize various species of plants. Hymenoscyphus caudatus occurs mostly in leaf debris of broad-leaved trees, including Aesculus, Corylus, Populus and Salix. Hymenoscyphus scutula occurs mostly on herbaceous stems of dicotyledons. Both fungi have been found previously on F. excelsior but with much lower frequency [52,109,114,115]. We did not detect H. albidus. Studies in other countries, such as the Czech Republic and Norway, suggest it was absent or occurred at low frequency [53,116].
Representatives of the Xylariales on F. excelsior petioles only sporadically produced stromata with conidia and were detected only on the basis of isolation, although numerous species from this order form fruit bodies on wood of many species of broad-leaved trees [95,96]. They are common endophytes in young leaflets and petioles of F. excelsior and other species of broad-leaved trees [81,95,99,117]. This endophytic phase can help them in further colonization of leaves after their fall.
Some species found unexpectedly on F. excelsior leaf petioles include Phacidium lacerum, Leptosphaeria conoidea, Leptosphaeria urticae and Pseudoophiobolus italicus. The first is known in Europe from its occurrence on needles of Pinus sylvestris. It invades the older generations of needles in the crown and persists in the fallen needles [118]. So far, L. conoidea has been found on dead stems of various herbaceous plants, especially Asteraceae (Compositae) or Apiaceae (Umbelliferae) [119], L. urticae on dead stems of Urtica dioica [120] and P. italicus on dead stems of Onobrychis viciifolia [121].
The frequent occurrence on petioles of species in the genera Cyathicula, Hymenoscyphus (other than H. fraxineus), Hypoderma or Typhula may have resulted from their time of fructification, during the leaf-fall season. This would give them priority of access among the other endophytes and pathogens that had colonized leaves in the tree crown before autumn leaf fall.
A similar interpretation may explain the differences in frequency of Peziza varia (9.7%) and P. ninguis (2.0%). The first one fruits in the leaf-fall season and the second one much later, at the spring thaw [122]. Ascospores of P. varia colonize the, as yet, uninhabited (in autumn) petioles of the fallen leaves, while P. ninguis has to compete with fungi that had colonized petioles earlier (during 3–4 months, in late autumn and winter).
Basidomycota detected in leaf petioles of F. excelsior from the previous year perform various ecological functions. It seems that some Typhula species are saprotrophs that contribute to decomposition of plant debris. There are, however, phytopathogenic Typhula species, T. ishikariensis and T. incarnata, which cause important diseases of cereals and grasses [65,66,123,124]. According to Yang et al. [66], sporophores of most Typhula species are usually found in autumn. Our results support this; only sclerotia were found in spring. Peniophora incarnata and Trametes versicolor are known decomposers of wood. Their degradation of all cell-wall constituents causes intensive white rot in logs, branches and stumps. Species of Sistotrema cause brown rot when carbohydrates are extensively removed and lignin is degraded only to a limited extent. Coprinus species, Hypholoma fasciculare and Psathyrella piluliformis form basidiocarps on stumps, often continuously, even from spring to autumn [94,125,126,127]. According to Dix and Webster [128], litter-inhabiting Basidiomycota in the genera Marasmius, Mycena, Clitocybe or Collybia are associated with the final stages of litter decay. Their preferences for pre-decayed substrate could limit their frequency in the previous year’s leaf petioles. Fellozyma sp. and Tilletiopsis washingtonensis are representatives of the basidiomycetous yeasts. Neither taxon had preference for any dominant tree type [129,130]. Despite the significant proportion of yeasts in fungal communities in forest soil and litter, they were found sporadically at the decomposition stage of the ash petioles with the methods being used.
Examples of the relationships between fungi in ash leaf petioles demonstrate their complexity and that of the habitat, as is also indicated by different values of Shannon’s and Simpson’s diversity indices.
4.2. Competition between H. fraxineus and Saprotrophs
In addition to observations on simultaneous occurrence of H. fraxineus and other fungi in dead leaf petioles of F. excelsior in situ, further information on the competitive abilities of saprotrophs against H. fraxineus was obtained from dual-culture assays in vitro. The results show that some of the saprotrophs found can be considered as very effective antagonists of H. fraxineus.
It is well-known that H. fraxineus can produce numerous metabolites, including those with mycotoxic of phytotoxic properties [131,132,133,134,135]. The inhibition of mycelial growth of colonizers of ash stems and branches by H. fraxineus has already been observed in vitro at the beginning of studies on ash dieback [136]. The present results showed much variation among interactions between H. fraxineus and individual saprotrophs. Generally, H. fraxineus was able to reduce mycelial growth of most saprotrophs. However, all tested saprotrophs also reduced the growth of H. fraxineus. It may be assumed that the extent of inhibition depended mostly on the growth rate of the saprotroph and the type of mutual relationship. The present results on H. fraxineus–saprotroph interactions are broadly similar to those of others [27,28,29,137].
The dual-culture assay is expected to demonstrate the different strategies used by various groups to gain occupation of their niche and obtain the nutrition required for survival [57,128,138].
One of these groups includes the fast growing fungi, Alternaria spp., Chromelosporium sp., Colletotrichum acutatum, Coniochaeta sp. 2, Epicoccum nigrum, Phacidium lacerum, Phoma spp. and Trametes versicolor, which strongly inhibited growth of H. fraxineus in vitro, in agreement with results of Haňáčková [28] or Becker [139]. They may dominate during early substrate colonization. In nature, the overall effect may depend on the frequency of particular taxa. The frequent detection of the fast growing fungi in laboratory studies is often due to their affiliation to “pioneer communities of fungi” which may include numerous “weed” species [128]. Their fast growth rate compensates for low metabolic (e.g., antibiotic) activity, in contrast to “later dominants”, which may be highly antagonistic because of production of metabolic inhibitors and mycoparasitism [57,128,140].
The formation of an inhibition zone, regardless of its main producer (H. fraxineus and/or saprotroph), was the type of interaction seen most often. The most potentially valuable saprotrophs were those that inhibited growth of H. fraxineus from the greatest distance and produced wide inhibition zones. Such results were obtained with Malbranchea sp. (Onygenales) and Pseudocoleophoma polygonicola (Pleosporales). The former colonizes decomposed plants or cellulose-containing substrate, is thermophilic and is an important source of lignocellulolytic enzymes [141,142]. Pseudocoleophoma polygonicola is a recently described species with unknown ecology, previously found only in Japan on dead polygonaceous plants [143]. The related species, Coleophoma empetri, produces an antifungal echinocandin-like compound [144], suggesting promise for P. polygonicola which may have similar properties.
Other saprotrophs that produced inhibition zones are also known producers of antifungal metabolites or degrading enzymes. This group includes, among others, Aureobasidium pullulans, Coniochaeta spp., Epicoccum nigrum and Fusarium lateritium. Aureobasidium pullulans produces a group of antifungal metabolites described as aureobasidins, which are derivatives of cyclic deosipeptides, and secretes cell-wall degrading enzymes, including chitinase and β-1,3 glucanase, which contribute to lysis of pathogen hyphae [145,146]. Coniochaeta species produce antibacterial and antifungal coniosetin and coniochaetone-a and -b [147]. The frequently found E. nigrum produces flavipin, epicorazine and epipyrone A [145,148]. The antifungal activities of some of them has led to development of biological control products based on E. nigrum mycelium, spores and metabolites [148]. Moreover, this species secretes cell-wall degrading enzymes which contribute to lysis of pathogen hyphae. Fusarium lateritium produces five antifungal sesquiterpenes, including microsphaeropsisins D and E [149].
Melanins, important natural pigments, may also have an important role in establishing relationships among microorganisms. It is belived that they contribute to survival of fungi in diverse hostile environments [150]. In the present study, the production of dark-colored pigmentation was observed in tests with H. fraxineus and some members of the Pleosporales. Pukalski et al. [151] recently showed that Plenodomus biglobosus (Pleosporales) isolated from living leaf petioles of F. excelsior can produce pheomelanin, which is rarely produced by fungi.
Only a few fungi overgrew and covered the H. fraxineus colony in dual cultures, an effect that may indicate their potential for antagonism in nature. The strongest effects occurred with ascomycetous Clonostachys rosea, Trichoderma viride (Hypocreales), Nemania, Rosellinia and Xylaria (Xylariales), and basidiomycetous Hypholoma fasciculare (Agaricales) and Peniophora incarnata (Russulales). Clonostachys rosea and T. viride are well-known antagonists of many species of fungi [152,153,154,155] and their suppressive effects were expected.
Re-isolations of fungi from the covered colonies seemed to indicate successful replacement of H. fraxineus by the saprotrophs. Replacement of H. fraxineus by some endophyes from ash shoots in vitro has also been observed [28]. Boddy [156] pointed out that similar interactions in dead wood, in situ, may also lead to replacement of the weaker competitor in woody tissues. Successful and effective replacement of H. fraxineus in leaf petioles, however, when the pathogen is often surrounded by outer and inner pseudosclerotial plates, may be difficult [16]. Successful replacement could be demonstrated by the formation of the saprotroph’s fruit bodies on pseudosclerotia of H. fraxineus which is rare. The protective value of pseudosclerotial plates can be compared with that of similar structures formed by saprotrophic Xylaria hypoxylon, which were so firm that they were able to prevent colonization of beech timber blocks by other saprotrophic fungi [157].
Interpretation of dual-culture assays generally supports results on colonization of leaf petioles. On 87.6% of petioles colonized by H. fraxineus, there were also other fungi which colonized other parts of the petiole, possibly making those parts unavailable to H. fraxineus. In situ, growth of H. fraxineus was apparently limited by the same fungi that produced inhibition zones or made physical contact by overgrowth and replacement of H. fraxineus. These results show that the spectrum of fungi able to limit growth of H. fraxineus in situ is wide and includes more than 50% of species fruiting on leaf petioles. Significantly, most of these fungi are not host specific for F. excelsior. They occur on many species of trees, bushes and even herbaceous plants. Among this group, detected in the dead leaf petioles, are phytopathogenic Discohainesia oenotherae, Leptosphaeria sclerotioides and Verticillium dahliae. Discohainesia oenotherae is generally a pathogen or saprotroph on many woody species and cultivated plants, including strawberries, and occurs worldwide [158,159]. Leptosphaeria sclerotioides generally does not occur on trees. It is a pathogen on herbaceous legumes, particularly sweet clover (Melilotus alba) and lucerne (Medicago sativa). It may also be pathogenic on grasses and cereals exposed to low temperature [119,160]. Verticillium dahliae is a dangerous, vascular pathogen. It survives easily and efficiently in the form of microsclerotia in soil and plant debris. Its occurrence additionally in ash stands suggests a risk for economically important agricultural or horticultural production and the health of forests [161].
Ash dieback has caused a reduction in F. excelsior stands in many regions of Europe. This is being followed by a decreased frequency of fungal colonizers of ash [162]. Current studies suggest, however, that they may be replaced by other fungi which are colonizers of other species of trees and shrubs and even of herbaceous plants.
Further studies on antagonism of mycobiota against H. fraxineus in situ are needed. Assessment of the chance of fungi being able to compete with H. fraxineus must take into account the important fact that H. fraxineus has the advantage of having early, pre-competitive infection and colonization of living leaves while still in the crown.
5. Conclusions
An important stage in the life cycle of H. fraxineus on F. excelsior is its successful survival until the following year in dead leaves, particularly in leaf petioles which, in summer, will be the substrate for fruit-body development. This stage can be affected by co-colonizers. Co-colonizers of 139 fungal taxa, including Ascomycota (120 species), Basidiomycota (17 species) and Mucoromycota (2 species), were identified in dead leaf petioles of F. excelsior from the previous year. Many of these fungi showed antagonism to H. fraxineus. Few of these taxa were host specific to F. excelsior. Fungal taxa that occur in a wide range of trees, shrubs and herbaceous plants were more frequent, which is important considering that F. excelsior has a decreased share in European stands owing to ash dieback. The structure of fungal communities in leaf petioles of F. excelsior emphasizes the significance of plant species diversity in forests and the necessity for accumulation of wood debris, which serves as substrate for fruit-body and spore production by potential antagonists of H. fraxineus, including species of Coniochaeta, Nemania, Peniophora, Rosellinia, Trametes and Xylaria.
Supplementary Materials
The following are available online at: https://www.mdpi.com/article/10.3390/f12101412/s1, Table S1: List of morphotypes from the previous year’s leaf petioles of Fraxinus excelsior in five forest stands, in southern Poland in 2017. Sequences deposited in GenBank. Reference sequences from GenBank. Results of interactions of test fungi with two isolates of Hymenoscyphus fraxineus (Hf 1/Hf 2) in vitro; Table S2: Fungi isolated from the previous year’s leaf petioles of Fraxinus excelsior (subsample I) and fungi detected in situ from fruit bodies on the previous year’s leaf petioles of Fraxinus excelsior (subsample II and III); Figure S1: Phylogram obtained from maximum likelihood (ML) analyses of the ITS1/2 data for the isolated fungal taxa. Sequences obtained during this study are presented in bold type. The Bootstrap values ≥75% for ML and Bayesian inference (BI) analyses are presented at branch as follows: ML/BI. * Bootstrap values <75%. The tree is drawn to scale (see bar) with branch length measured in the number of substitutions per site. Fellozyma inositophila and Fellozyma sp. represent the outgroup; Figure S2: Principal component analysis (PCA) with frequencies of fungal variables represented by arrows and sites represented by black circles, for mycobiota detected in situ from fruit bodies on the previous year’s leaf petioles of F. excelsior; Figure S3: Similarity of fungal communities detected in situ from fruit bodies on the previous year’s leaf petioles of F. excelsior in five stands shown by dendrogram based on Bray–Curtis similarity index; Figure S4: Network of co-occurrence of fungi detected in situ from fruit bodies on the previous year’s leaf petioles of Fraxinus excelsior in five stands, in southern Poland, determined on the basis of data grouped for the study plots. Each connection shows a strong (Spearman’s p > 0.6) and significant (p < 0.05) correlation. Blue lines indicate positive correlation. The same color represents fungal taxa with potential co-occurrence in the same module. The frequency of the species was indicated by the size of each node.
Author Contributions
Conceptualization, T.K. and P.B.; methodology, T.K. (mycological aspects) and P.B. (molecular and statistical aspects); investigation, T.K. and P.B.; formal analysis, T.K. and P.B.; data curation, T.K. and P.B.; writing—original draft preparation, T.K. and P.B.; writing—review and editing, T.K. and P.B.; software, P.B.; supervision, T.K. and P.B.; visualization, P.B.; project administration and funding acquisition, T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was conducted in a project no. 2016/21/B/NZ9/01226, financed by the National Science Centre, Poland. Article processing charge was financed by a subvention from the Polish Ministry of Science and Higher Education for the University of Agriculture in Krakow for 2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Material.
Acknowledgments
The authors wish to thank H. Kwaśna for valuable comments, G. L. Bateman for his linguistic correction of the manuscript as well as J. Michalik and D. Ledwich for carrying out numerous laboratory works.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kowalski, T. O zamieraniu jesionów. Tryb. Leśnika 2001, 4, 6–7. [Google Scholar]
- Kowalski, T. Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- Przybył, K. Fungi associated with necrotic apical parts of Fraxinus excelsior shoots. For. Pathol. 2002, 32, 387–394. [Google Scholar] [CrossRef]
- Halmschlager, E.; Kirisits, T. First report of the ash dieback pathogen Chalara fraxinea on Fraxinus excelsior in Austria. Plant Pathol. 2008, 57, 1177. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef]
- Timmermann, V.; Nagy, N.E.; Hietala, A.M.; Børja, I.; Solheim, H. Progression of ash dieback in Norway related to tree age, disease history and regional aspects. Balt. For. 2017, 23, 150–158. [Google Scholar]
- Hietala, A.M.; Børja, I.; Cross, H.; Nagy, N.E.; Solheim, H.; Timmermann, V.; Vivian-Smith, A. Dieback of European Ash: What Can We Learn from the Microbial Community and Species-Specific Traits of Endophytic Fungi Associated with Ash? In Endophytes of Forest Trees. Biology and Applications; Pirttilä, A.M., Frank, A.C., Eds.; Forestry Sciences; Springer International Publishing: Cham, Switzerland, 2018; Volume 86, pp. 229–258. ISBN 978-3-319-89832-2. [Google Scholar]
- Enderle, R. An overview of ash (Fraxinus spp.) and the ash dieback disease in Europe. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Baral, H.-O.; Queloz, V.; Hosoya, T. Hymenoscyphus fraxineus, the correct scientific name for the fungus causing ash dieback in Europe. IMA Fungus 2014, 5, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Cleary, M.; Nguyen, D.; Marčiulynienė, D.; Berlin, A.; Vasaitis, R.; Stenlid, J. Friend or foe? Biological and ecological traits of the European ash dieback pathogen Hymenoscyphus fraxineus in its native environment. Sci. Rep. 2016, 6, 21895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, T.; Okane, I.; Ishiga, Y.; Degawa, Y.; Hosoya, T.; Yamaoka, Y. The life cycle of Hymenoscyphus fraxineus on Manchurian ash, Fraxinus mandshurica, in Japan. Mycoscience 2019, 60, 89–94. [Google Scholar] [CrossRef]
- Kowalski, T. Zamieranie jesionu—Aspekty taksonomiczne sprawcy choroby. Sylwan 2012, 156, 262–269. [Google Scholar]
- Hauptman, T.; Piškur, B.; de Groot, M.; Ogris, N.; Ferlan, M.; Jurc, D. Temperature effect on Chalara fraxinea: Heat treatment of saplings as a possible disease control method. For. Pathol. 2013, 43, 360–370. [Google Scholar] [CrossRef]
- Cleary, M.R.; Daniel, G.; Stenlid, J. Light and scanning electron microscopy studies of the early infection stages of Hymenoscyphus pseudoalbidus on Fraxinus excelsior. Plant Pathol. 2013, 62, 1294–1301. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Koukol, O.; Čmoková, A.; Zahradník, D.; Havrdová, L. Direct evidence of Hymenoscyphus fraxineus infection pathway through the petiole-shoot junction. For. Pathol. 2017, 47, e12370. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O. On the longevity of Hymenoscyphus pseudoalbidus in petioles of Fraxinus excelsior. For. Pathol. 2013, 43, 168–170. [Google Scholar] [CrossRef]
- Kirisits, T. Ascocarp formation of Hymenoscyphus fraxineus on several-year-old pseudosclerotial leaf rachises of Fraxinus excelsior. For. Pathol. 2015, 45, 254–257. [Google Scholar] [CrossRef]
- Kowalski, T.; Kehr, R. Stand des Eschentriebsterbens und zunehmende Schäden an Buche und Berg-Ahorn. In Jahrbuch der Baumpflege; Dujesiefken, D., Ed.; Haymarket Media: Braunschweig, Germany, 2016; pp. 63–83. [Google Scholar]
- Hauptman, T.; Celar, F.; de Groot, M.; Jurc, D. Application of fungicides and urea for control of ash dieback. iFor. Biogeosci. For. 2015, 8, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Bartha, B.; Mayer, A.; Lenz, H.D. Acceleration of ash petiole decomposition to reduce Hymenoscyphus fraxineus apothecia growth—A feasible method for the deprivation of fungal substrate. Balt. For. 2017, 23, 82–88. [Google Scholar]
- Marzano, M.; Woodcock, P.; Quine, C.P. Dealing with dieback: Forest manager attitudes towards developing resistant ash trees in the United Kingdom. Forestry 2019, 92, 554–567. [Google Scholar] [CrossRef]
- Noble, R.; Woodhall, J.W.; Dobrovin-Pennington, A.; Perkins, K.; Somoza-Valdeolmillos, E.; Gómez, H.L.; Lu, Y.; Macarthur, R.; Henry, C.M. Control of Hymenoscyphus fraxineus, the causal agent of ash dieback, using composting. For. Pathol. 2019, 49, e12568. [Google Scholar] [CrossRef]
- Berg, G.; Köberl, M.; Rybakova, D.; Müller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93, 50. [Google Scholar] [CrossRef]
- Terhonen, E.; Kovalchuk, A.; Zarsav, A.; Asiegbu, F.O. Biocontrol Potential of Forest Tree Endophytes. In Endophytes of Forest Trees. Biology and Applications; Pirttilä, A.M., Frank, A.C., Eds.; Forestry Sciences; Springer International Publishing: Cham, Switzerland, 2018; Volume 86, pp. 283–318. ISBN 978-3-319-89832-2. [Google Scholar]
- Collinge, D.B.; Jørgensen, H.J.L.; Latz, M.A.C.; Manzotti, A.; Ntana, F.; Rojas, E.C.; Jensen, B. Searching for novel fungal biological control agents for plant disease control among endophytes. In Endophytes for a Growing World; Cambridge University Press: Cambridge, UK, 2019; pp. 25–51. ISBN 9781108607667. [Google Scholar]
- Schlegel, M.; Dubach, V.; von Buol, L.; Sieber, T.N. Effects of endophytic fungi on the ash dieback pathogen. FEMS Microbiol. Ecol. 2016, 92, fiw142. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Haas, S.; Junker, C.; Andrée, N.; Schobert, M. Fungal endophytes are involved in multiple balanced antagonisms. Curr. Sci. 2015, 109, 39–45. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Havrdová, L.; Černý, K.; Zahradník, D.; Koukol, O. Fungal endophytes in ash shoots—Diversity and inhibition of Hymenoscyphus fraxineus. Balt. For. 2017, 23, 89–106. [Google Scholar]
- Kosawang, C.; Amby, D.B.; Bussaban, B.; McKinney, L.V.; Xu, J.; Kjær, E.D.; Collinge, D.B.; Nielsen, L.R. Fungal communities associated with species of Fraxinus tolerant to ash dieback, and their potential for biological control. Fungal Biol. 2018, 122, 110–120. [Google Scholar] [CrossRef]
- Halecker, S.; Wennrich, J.-P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Stadler, M.; Surup, F.; Schulz, B. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M.; Luis Bonilla, J.; Potvin, C. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 2007, 116, 2108–2124. [Google Scholar] [CrossRef]
- Jonard, M.; Andre, F.; Ponette, Q. Tree species mediated effects on leaf litter dynamics in pure and mixed stands of oak and beech. Can. J. For. Res. 2008, 38, 528–538. [Google Scholar] [CrossRef]
- Purahong, W.; Hyde, K.D. Effects of fungal endophytes on grass and non-grass litter decomposition rates. Fungal Divers. 2011, 47, 1–7. [Google Scholar] [CrossRef]
- Hoeber, S.; Fransson, P.; Weih, M.; Manzoni, S. Leaf litter quality coupled to Salix variety drives litter decomposition more than stand diversity or climate. Plant Soil 2020, 453, 313–328. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Osono, T.; Takeda, H. Decomposition of organic chemical components in relation to nitrogen dynamics in leaf litter of 14 tree species in a cool temperate forest. Ecol. Res. 2005, 20, 41–49. [Google Scholar] [CrossRef]
- Nykvist, N. Leaching and decomposition of litter I. Experiments on leaf litter of Fraxinus excelsior. Oikos 1959, 10, 190. [Google Scholar] [CrossRef]
- Błońska, E.; Piaszczyk, W.; Staszel, K.; Lasota, J. Enzymatic activity of soils and soil organic matter stabilization as an effect of components released from the decomposition of litter. Appl. Soil Ecol. 2021, 157, 103723. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, O.; Tenhunen, J.D. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiol. 1998, 18, 681–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotrufo, M.F.; Briones, M.J.I.; Ineson, P. Elevated CO2 affects field decomposition rate and palatability of tree leaf litter: Importance of changes in substrate quality. Soil Biol. Biochem. 1998, 30, 1565–1571. [Google Scholar] [CrossRef]
- Jacob, M.; Viedenz, K.; Polle, A.; Thomas, F.M. Leaf litter decomposition in temperate deciduous forest stands with a decreasing fraction of beech (Fagus sylvatica). Oecologia 2010, 164, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü. Differences in chemical composition relative to functional differentiation between petioles and laminas of Fraxinus excelsior. Tree Physiol. 1999, 19, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Conway, D.R.; Frankland, J.C.; Saunders, V.A.; Wilson, D.R. Effects of elevated atmospheric CO2 on fungal competition and decomposition of Fraxinus excelsior litter in laboratory microcosms. Mycol. Res. 2000, 104, 187–197. [Google Scholar] [CrossRef]
- Dilly, O.; Bartsch, S.; Rosenbrock, P.; Buscot, F.; Munch, J.C. Shifts in physiological capabilities of the microbiota during the decomposition of leaf litter in a black alder (Alnus glutinosa (Gaertn.) L.) forest. Soil Biol. Biochem. 2001, 33, 921–930. [Google Scholar] [CrossRef]
- Osono, T.; Takeda, H. Comparison of litter decomposing ability among diverse fungi in a cool temperate deciduous forest in Japan. Mycologia 2002, 94, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Shirouzu, T.; Hirose, D.; Fukasawa, Y.; Tokumasu, S. Fungal succession associated with the decay of leaves of an evergreen oak, Quercus myrsinaefolia. Fungal Divers. 2009, 34, 87–109. [Google Scholar]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef]
- Osono, T. Leaf litter decomposition of 12 tree species in a subtropical forest in Japan. Ecol. Res. 2017, 32, 413–422. [Google Scholar] [CrossRef]
- Rosales-Castillo, J.A.; Oyama, K.; Vázquez-Garcidueñas, M.S.; Aguilar-Romero, R.; García-Oliva, F.; Vázquez-Marrufo, G. Fungal community and ligninolytic enzyme activities in Quercus deserticola Trel. litter from forest fragments with increasing levels of disturbance. Forests 2017, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Tennakoon, D.S.; Gentekaki, E.; Jeewon, R.; Kuo, C.H.; Promputtha, I.; Hyde, K.D. Life in leaf litter: Fungal community succession during decomposition. Mycosphere 2021, 12, 406–429. [Google Scholar] [CrossRef]
- Davydenko, K.; Vasaitis, R.; Stenlid, J.; Menkis, A. Fungi in foliage and shoots of Fraxinus excelsior in eastern Ukraine: A first report on Hymenoscyphus pseudoalbidus. For. Pathol. 2013, 43, 462–467. [Google Scholar] [CrossRef]
- Koukol, O.; Haňáčková, Z.; Dvořák, M.; Havrdová, L. Unseen, but still present in Czechia: Hymenoscyphus albidus detected by real-time PCR, but not by intensive sampling. Mycol. Prog. 2016, 15, 6. [Google Scholar] [CrossRef]
- Cross, H.; Sønstebø, J.H.; Nagy, N.E.; Timmermann, V.; Solheim, H.; Børja, I.; Kauserud, H.; Carlsen, T.; Rzepka, B.; Wasak, K.; et al. Fungal diversity and seasonal succession in ash leaves infected by the invasive ascomycete Hymenoscyphus fraxineus. New Phytol. 2017, 213, 1405–1417. [Google Scholar] [CrossRef]
- Trapiello, E.; Schoebel, C.N.; Rigling, D. Fungal community in symptomatic ash leaves in Spain. Balt. For. 2017, 23, 68–73. [Google Scholar]
- Kowalski, T.; Kraj, W.; Bednarz, B.; Rossa, R. The association of Boeremia lilacis with necrotic lesions on shoots and leaf petioles and its pathogenicity towards Fraxinus excelsior. Eur. J. Plant Pathol. 2019, 154, 961–974. [Google Scholar] [CrossRef] [Green Version]
- Wicklow, D.T. Interference competition. In The Fungal Community: Its Organization and Role in the Ecosystem; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 265–274. [Google Scholar]
- Zhang, L.; Andersen, K.H.; Dieckmann, U.; Brännström, Å. Four types of interference competition and their impacts on the ecology and evolution of size-structured populations and communities. J. Theor. Biol. 2015, 380, 280–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, T.; Czekaj, A. Symptomy chorobowe i grzyby na zamierających jesionach (Fraxinus excelsior L.) w drzewostanach Nadleśnictwa Staszów. Disease symptoms and fungi on dying ash trees (Fraxinus excelsior L.) in Staszów Forest District stands. Leśne Pr. Badaw. 2010, 71, 357–368. [Google Scholar]
- Lacap, D.C.; Hyde, K.D.; Liew, E.C.Y. An evaluation of the fungal “morphotype” concept based on ribosomal DNA sequences. Fungal Divers. 2003, 12, 53–66. [Google Scholar]
- Kowalski, T.; Bartnik, C. Morphologial variation in colonies of Chalara fraxinea isolated from ash (Fraxinus excelsior L.) stems with symptoms of dieback and effects of temperature on colony growth and structure. Acta Agrobot. 2010, 63, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Kirisits, T.; Dämpfle, L.; Kräutler, K. Hymenoscyphus albidus is not associated with an anamorphic stage and displays slower growth than Hymenoscyphus pseudoalbidus on agar media. For. Pathol. 2013, 43, 386–389. [Google Scholar] [CrossRef]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Ainthworth and Bisby’s Dictionary of the Fungi, 10th ed.; Kirk, P., Cannon, P., Minter, D., Stalpers, J., Eds.; CABI: Wallingford, UK, 2008; ISBN 978-0-85199-826-8. [Google Scholar]
- Baral, H.-O.; Bemmann, M. Hymenoscyphus fraxineus vs. Hymenoscyphus albidus—A comparative light microscopic study on the causal agent of European ash dieback and related foliicolous, stroma-forming species. Mycology 2014, 5, 228–290. [Google Scholar] [CrossRef] [Green Version]
- Koske, R.E. Typhula erythropus: II. Sclerotial germination and basidiocarp production. Mycologia 1975, 67, 128–146. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, F.; Hsiang, T. Fertile sporophore production of Typhula phacorrhiza in the field is related to temperatures near freezing. Can. J. Microbiol. 2006, 52, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Kraj, W.; Kowalski, T. Genetic variability of Hymenoscyphus pseudoalbidus on ash leaf rachises in leaf litter of forest stands in Poland. J. Phytopathol. 2014, 162, 218–227. [Google Scholar] [CrossRef]
- Index Fungorum—Search Page. Available online: http://www.indexfungorum.org/names/Names.asp (accessed on 29 August 2021).
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, J.; Platas, G.; Paulus, B.; Bills, G.F. High-throughput culturing of fungi from plant litter by a dilution-to-extinction technique. FEMS Microbiol. Ecol. 2007, 60, 521–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BIOEDIT: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Lahlali, R.; Hijri, M. Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 2010, 311, 152–159. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Fernández-González, R.A.; Sanz-Ros, A.V.; Pando, V.; Diez, J.J. Two fungal endophytes reduce the severity of pitch canker disease in Pinus radiata seedlings. Biol. Control 2016, 94, 1–10. [Google Scholar] [CrossRef]
- Bilański, P.; Kowalski, T. Fungal endophytes in Fraxinus excelsior petioles and their in vitro antagonistic potential against the ash dieback pathogen Hymenoscyphus fraxineus. Microbiol. Res. 2021. accepted. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication, 1st ed.; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Camargo, J.A. Must dominance increase with the number of subordinate species in competitive interactions? J. Theor. Biol. 1993, 161, 537–542. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009; pp. 361–362. [Google Scholar]
- STATISTICA (Data Analysis Software System); StatSoft Inc.: Tulsa, OK, USA, 2014.
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core: Vienna, Austria, 2018. [Google Scholar]
- Osono, T. Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Kubartová, A.; Ranger, J.; Berthelin, J.; Beguiristain, T. Diversity and decomposing ability of saprophytic fungi from temperate forest litter. Microb. Ecol. 2009, 58, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Frankland, J.C. Fungal succession—Unravelling the unpredictable. Mycol. Res. 1998, 102, 1–15. [Google Scholar] [CrossRef]
- Paulus, B.; Gadek, P.; Hyde, K. Successional patterns of microfungi in fallen leaves of Ficus pleurocarpa (Moraceae) in an Australian tropical rain forest. Biotropica 2006, 38, 051207072004001. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef] [Green Version]
- Jahn, H.T. Pilze die an Holz Wachsen, 1st ed.; Verlag Busse: Herford, Germany, 1979; ISBN 978-3871208539. [Google Scholar]
- Petrini, L.; Petrini, O. Xylariaceous fungi as endophytes. Sydowia 1985, 38, 216–234. [Google Scholar]
- Granmo, A.; Læssøe, T.; Schumacher, T. The genus Nemania s.l. (Xylariaceae) in Norden. Sommerfeltia 1999, 27, 1. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on stems and twigs in initial and advanced stages of dieback of European ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Linaldeddu, B.T.; Bottecchia, F.; Bregant, C.; Maddau, L.; Montecchio, L. Diplodia fraxini and Diplodia subglobosa: The main species associated with cankers and dieback of Fraxinus excelsior in north-eastern Italy. Forests 2020, 11, 883. [Google Scholar] [CrossRef]
- Schlegel, M.; Queloz, V.; Sieber, T.N. The Endophytic Mycobiome of European Ash and Sycamore Maple Leaves—Geographic Patterns, Host Specificity and Influence of Ash Dieback. Front. Microbiol. 2018, 9, 2345. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Bussaban, B.; Paulus, B.; Crous, P.W.; Lee, S.; Mckenzie, E.H.C.; Photita, W.; Lumyong, S. Diversity of saprobic microfungi. Biodivers. Conserv. 2007, 16, 7–35. [Google Scholar] [CrossRef]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef]
- De Silva, N.I.; Phillips, A.J.L.; Liu, J.-K.; Lumyong, S.; Hyde, K.D. Phylogeny and morphology of Lasiodiplodia species associated with Magnolia forest plants. Sci. Rep. 2019, 9, 14355. [Google Scholar] [CrossRef]
- Osono, T. Role of phyllosphere fungi of forest trees in the development of decomposer fungal communities and decomposition processes of leaf litter. Can. J. Microbiol. 2006, 52, 701–716. [Google Scholar] [CrossRef]
- Sivanesan, A. The Bitunicate Ascomycetes and Their Anamorphs, 1st ed.; Lubrecht & Cramer: Schaan, Liechtenstein, 1984; ISBN 978-3-76821-329-5. [Google Scholar]
- Schubert, K.; Rischel, A.; Braun, D.U. A monograph of Fusicladium s.lat. (Hyphomycetes). Schlechtendalia 2013, 9, 1–132. [Google Scholar]
- Agan, A.; Drenkhan, R.; Adamson, K.; Tedersoo, L.; Solheim, H.; Børja, I.; Matsiakh, I.; Timmermann, V.; Nagy, N.E.; Hietala, A.M. The relationship between fungal diversity and invasibility of a foliar niche—The case of ash dieback. J. Fungi 2020, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Ihrmark, K.; Stenlid, J. Investigations concerning the role of Chalara fraxinea in declining Fraxinus excelsior. Plant Pathol. 2009, 58, 284–292. [Google Scholar] [CrossRef]
- Carpenter, S.E. Monograph of Crocicreas (Ascomycetes, Helotiales, Leotiaceae. Mem. N. Y. Bot. Gard. 1981, 33, 1–290. [Google Scholar]
- Baral, H.-O.; Krieglsteriner, G.J. Bausteine zu einer Askomyzeten-Flora der Bundesrepublik Deutschland: In Süddeutschland gefundene inoperculate Discomyzeten mit taxonomische, okologischen, chorologischen Hinweisen und einer Farbtafel. Beih. Z. Mykol. 1985, 6, 1–160. [Google Scholar]
- Minter, D.W. Hypoderma rubi. In CMI Descriptions of Pathogenic Fungi and Bacteria; Commonwealth Mycological Institute: Kew, UK, 1984; pp. 1–2. [Google Scholar]
- Thomas, P.A. Biological Flora of the British Isles: Fraxinus excelsior. J. Ecol. 2016, 104, 1158–1209. [Google Scholar] [CrossRef]
- Pastirčák, M.; Pastirčáková, K. Hypoderma rubi on two new hosts in Slovakia. Mycotaxon 2017, 132, 849–855. [Google Scholar] [CrossRef]
- Hütter, R. Untersuchungen über die Gattung Pyrenopeziza Fuck. J. Phytopathol. 1958, 33, 1–54. [Google Scholar] [CrossRef]
- Pastirčáková, K.; Ivanová, H.; Pastirčák, M. Species diversity of fungi on damaged branches and leaves of ashes (Fraxinus spp.) in different types of stands in Slovakia. Cent. Eur. For. J. 2018, 64, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Pastircáková, K.; Adamcíková, K.; Barta, M.; Pažitnỳ, J.; Hot’ka, P.; Sarvašová, I.; Horáková, M.K. Host range of Hymenoscyphus fraxineus in Slovak Arboreta. Forests 2020, 11, 596. [Google Scholar] [CrossRef]
- Hietala, A.M.; Børja, I.; Solheim, H.; Nagy, N.E.; Timmermann, V. Propagule pressure build-up by the invasive Hymenoscyphus fraxineus following its introduction to an ash forest inhabited by the native Hymenoscyphus albidus. Front. Plant Sci. 2018, 9, 1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Kowalski, T. Zur Pilzflora toter Kiefernnadeln. Z. Mykol. 1988, 54, 159–173. [Google Scholar]
- Boerema, G.H.; Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual. Differentiation of Specific and Infra-Specific Taxa in Culture; Boerema, G.H., de Gruyter, J., Noordeloos, M.E., Hamers, M.E.C., Eds.; CABI: Wallingford, UK, 2004; ISBN 978-0-85199-743-8. [Google Scholar]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Phookamsak, R.; Wanasinghe, D.N.; Hongsanan, S.; Phukhamsakda, C.; Huang, S.-K.; Tennakoon, D.S.; Norphanphoun, C.; Camporesi, E.; Bulgakov, T.S.; Promputtha, I.; et al. Towards a natural classification of Ophiobolus and ophiobolus-like taxa; introducing three novel genera Ophiobolopsis, Paraophiobolus and Pseudoophiobolus in Phaeosphaeriaceae (Pleosporales). Fungal Divers. 2017, 87, 299–339. [Google Scholar] [CrossRef]
- Hohmeyer, H. Ein Schlüssel zu den europaischen Arten der Gattung Peziza L. Z. Mykol. 1986, 52, 161–188. [Google Scholar]
- Remsberg, R.E. Studies in the genus Typhula. Mycologia 1940, 32, 52–96. [Google Scholar] [CrossRef]
- Wu, C.; Hsiang, T. Pathogenicity and formulation of Typhula phacorrhiza, a biocontrol agent of gray snow mold. Plant Dis. 1998, 82, 1003–1006. [Google Scholar] [CrossRef]
- Zabel, R.; Morrell, J. Wood Microbiology: Decay and Its Prevention, 1st ed.; Academic Press: San Diego, CA, USA, 1992; ISBN 978-0-32313-946-5. [Google Scholar]
- Worrall, J.J.; Anagnost, S.E.; Zabel, R.A. Comparison of wood decay among diverse lignicolous fungi. Mycologia 1997, 89, 199–219. [Google Scholar] [CrossRef]
- Boddy, L. Fungal community ecology and wood decomposition process in angiosperms: From standing tree to complete decay of coarse woody debris. Ecol. Bull. 2001, 49, 43–56. [Google Scholar]
- Dix, N.J.; Webster, J. Fungal Ecology, 1st ed.; Springer: Dordrecht, The Netherlands, 1995; ISBN 978-94-010-4299-4. [Google Scholar]
- Mašínová, T.; Bahnmann, B.D.; Větrovský, T.; Tomšovský, M.; Merunková, K.; Baldrian, P. Drivers of yeast community composition in the litter and soil of a temperate forest. FEMS Microbiol. Ecol. 2017, 93, fiw223. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Yurkov, A.M.; Boekhout, T.; Stadler, M. Diversity of Tilletiopsis-Like Fungi in Exobasidiomycetes (Ustilaginomycotina) and Description of Six Novel Species. Front. Microbiol. 2019, 10, 2544. [Google Scholar] [CrossRef] [PubMed]
- Grad, B.; Kowalski, T.; Kraj, W. Studies on secondary metabolite produced by Chalara fraxinea and its phytotoxic influence on Fraxinus excelsior. Phytopathologia 2009, 54, 61–69. [Google Scholar]
- Andersson, P.F.; Bengtsson, S.; Stenlid, J.; Broberg, A. B-norsteroids from Hymenoscyphus pseudoalbidus. Molecules 2012, 17, 7769–7781. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.L.H.; Zaspel, I.; Schuemann, M.; Stephanowitz, H.; Krause, E. Rapid In-Vitro and In-Vivo Detection of Chalara fraxinea by Means of Mass Spectrometric Techniques. Am. J. Plant Sci. 2013, 4, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Andersson, P.F.; Johansson, S.B.K.; Stenlid, J.; Broberg, A. Isolation, identification and necrotic activity of viridiol from Chalara fraxinea, the fungus responsible for dieback of ash. For. Pathol. 2010, 40, 43–46. [Google Scholar] [CrossRef]
- Citron, C.A.; Junker, C.; Schulz, B.; Dickschat, J.S. A volatile lactone of Hymenoscyphus pseudoalbidus, pathogen of European ash dieback, inhibits host germination. Angew. Chem. Int. Ed. 2014, 53, 4346–4349. [Google Scholar] [CrossRef]
- Kowalski, T.; Holdenrieder, O. Pathogenicity of Chalara fraxinea. For. Pathol. 2009, 39, 1–7. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [Green Version]
- Frankland, J.C. Mechanisms in fungal succession. In The Fungal Community: Its Organization and Role in the Ecosystem; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 383–401. ISBN 978-0824786052. [Google Scholar]
- Becker, R.; Ulrich, K.; Behrendt, U.; Kube, M.; Ulrich, A. Analyzing ash leaf-colonizing fungal communities for their biological control of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 590944. [Google Scholar] [CrossRef]
- Timpe, D. Introduction to the theory of science. Philos. Hist. 1981, 14, 37–39. [Google Scholar] [CrossRef]
- Mahajan, C.; Basotra, N.; Singh, S.; Di Falco, M.; Tsang, A.; Chadha, B.S. Malbranchea cinnamomea: A thermophilic fungal source of catalytically efficient lignocellulolytic glycosyl hydrolases and metal dependent enzymes. Bioresour. Technol. 2016, 200, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.M.O.; Vici, A.C.; Pinheiro, M.P.; Heinen, P.R.; de Oliveira, A.H.C.; Ward, R.J.; Prade, R.A.; Buckeridge, M.S.; de Lourdes Teixeira de Moraes Polizeli, M. A highly glucose tolerant ß-glucosidase from Malbranchea pulchella (MpBg3) enables cellulose saccharification. Sci. Rep. 2020, 10, 6998. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Yawata, K.; Orino, Y.; Ueda, S.; Isogai, Y.; Taguchi, G.; Shimosaka, M.; Hashimoto, S. Agrobacterium tumefaciens-mediated transformation of antifungal lipopeptide producing fungus Coleophoma empetri F-11899. Curr. Genet. 2009, 55, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Finlay, R.; Ward, J.S. Antifungal compounds produced by Epicoccum purpurascens against soil-borne plant pathogenic fungi. Soil Biol. Biochem. 1987, 19, 657–664. [Google Scholar] [CrossRef]
- Bozoudi, D.; Tsaltas, D. The multiple and versatile roles of Aureobasidium pullulans in the vitivinicultural sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Asgari, B.; Zare, R.; Gams, W. Coniochaeta ershadii, a new species from Iran, and a key to well-documented Coniochaeta species. Nov. Hedwig. 2007, 84, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.J.; Cadelis, M.M.; Kim, S.H.; Swift, S.; Copp, B.R.; Villas-Boas, S.G. Epipyrone a, a broad-spectrum antifungal compound produced by Epicoccum nigrum ICMP 19927. Molecules 2020, 25, 5997. [Google Scholar] [CrossRef]
- Zhu, J.; Xiong, P.; Li, Z.; Li, J.; Lin, L.; Fu, X.; Huang, Y.; Xiong, Y.; Li, C. Antifungal sesquiterpenes with post-harvest anthracnose control effect on bananas from the fungus Fusarium lateritium. Nat. Prod. Res. 2021, 15, 1–8. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal Melanin: What do we know about structure? Front. Microbiol. 2015, 6, 1463. [Google Scholar] [CrossRef] [Green Version]
- Pukalski, J.; Marcol, N.; Wolan, N.; Płonka, P.M.; Ryszka, P.; Kowalski, T.; Latowski, D. Detection of a pheomelanin-like pigment by EPR spectroscopy in the mycelium of Plenodomus biglobosus. Acta Biochim. Pol. 2020, 67, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Barnett, H.L.; Lilly, V.G. A destructive mycoparasite, Gliocladium roseum. Mycologia 1962, 54, 72–77. [Google Scholar] [CrossRef]
- Pachenari, A.; Dix, N.J. Production of toxins and wall degrading enzymes by Gliocladium roseum. Trans. Br. Mycol. Soc. 1980, 74, 561–566. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Crozier, J.; Thomas, S.E.; Samuels, G.J.; Vinyard, B.T.; Holmes, K.A. Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol. Control 2008, 46, 24–35. [Google Scholar] [CrossRef]
- John, R.P.; Tyagi, R.D.; Prévost, D.; Brar, S.K.; Pouleur, S.; Surampalli, R.Y. Mycoparasitic Trichoderma viride as a biocontrol agent against Fusarium oxysporum f. sp. adzuki and Pythium arrhenomanes and as a growth promoter of soybean. Crop Prot. 2010, 29, 1452–1459. [Google Scholar] [CrossRef]
- Boddy, L. Interspecific combative interactions between wood-decaying basidiomycetes. FEMS Microbiol. Ecol. 2000, 31, 185–194. [Google Scholar] [CrossRef]
- Rayner, A.D.M.; Boddy, L. Fungal Decomposition of Wood: Its Biology and Ecology, 1st ed.; Wiley: Chichester, UK; New York, NY, USA, 1988; ISBN 0471103101. [Google Scholar]
- Sutton, B.C.; Gibson, I.A. Pezizella oenotherae (conidial state Hainesia lythri). In CMI Descriptions of Pathogenic Fungi and Bacteria; Commonwealth Mycological Institute: Kew, UK, 1977; pp. 1–2. [Google Scholar]
- Debode, J.; Van Hemelrijck, W.; Heungens, K.; Maes, M.; Creemers, P. First report of Pilidium concavum causing tan-brown rot on strawberry fruit in Belgium. Plant Dis. 2011, 95, 1029. [Google Scholar] [CrossRef]
- Wunsch, M.J.; Bergstrom, G.C. Genetic and morphological evidence that Phoma sclerotioides, causal agent of brown root rot of alfalfa, is composed of a species complex. Phytopathology 2011, 101, 594–610. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, C.; Heitmann, B.; Vogel, C. Morphology, vegetative compatibility and pathogenicity of Verticillium dahliae isolates from woody ornamentals in Germany. J. Plant Dis. Prot. 2009, 116, 109–114. [Google Scholar] [CrossRef]
- Pautasso, M.; Aas, G.; Queloz, V.; Holdenrieder, O. European ash (Fraxinus excelsior) dieback—A conservation biology challenge. Biol. Conserv. 2013, 158, 37–49. [Google Scholar] [CrossRef]
Figure 1.
Frequency of fungi detected in situ on the previous year’s leaf petioles of F. excelsior. Relative species frequency calculated as the ratio of the number of specimens in the order to all specimens n = 1251. Percent of species calculated as the ratio of the number of species in the order to all species n = 117.
Figure 1.
Frequency of fungi detected in situ on the previous year’s leaf petioles of F. excelsior. Relative species frequency calculated as the ratio of the number of specimens in the order to all specimens n = 1251. Percent of species calculated as the ratio of the number of species in the order to all species n = 117.

Figure 2.
Principal component analysis (PCA) with frequencies of fungal variables represented by arrows and sites represented by black circles, for mycobiota isolated from the previous year’s leaf petioles of F. excelsior.
Figure 2.
Principal component analysis (PCA) with frequencies of fungal variables represented by arrows and sites represented by black circles, for mycobiota isolated from the previous year’s leaf petioles of F. excelsior.

Figure 3.
Similarity of fungal communities isolated from the previous year’s leaf petioles of F. excelsior in five stands shown by dendrogram based on Bray–Curtis similarity index.
Figure 3.
Similarity of fungal communities isolated from the previous year’s leaf petioles of F. excelsior in five stands shown by dendrogram based on Bray–Curtis similarity index.

Figure 4.
Network of co-occurrence of fungi isolated from the previous year’s leaf petioles of F. excelsior in five stands, in southern Poland, determined on the basis of data grouped for the study plots. Each connection shows a strong (Spearman’s p > 0.6) and significant (p < 0.05) correlation. Blue lines indicate positive and red lines negative correlation. Five modules were generated, and the same color represents fungal taxa with potential co-occurrence in the same module. The frequency of the species was indicated by the size of each node.
Figure 4.
Network of co-occurrence of fungi isolated from the previous year’s leaf petioles of F. excelsior in five stands, in southern Poland, determined on the basis of data grouped for the study plots. Each connection shows a strong (Spearman’s p > 0.6) and significant (p < 0.05) correlation. Blue lines indicate positive and red lines negative correlation. Five modules were generated, and the same color represents fungal taxa with potential co-occurrence in the same module. The frequency of the species was indicated by the size of each node.

Figure 5.
Frequency of fungi detected in situ from fruit bodies on the previous year’s leaf petioles of F. excelsior. Relative species frequency calculated as the ratio of the number of specimens in the order to all specimens n = 882. Percent of species calculated as the ratio of the number of species in the order to all species n = 45.
Figure 5.
Frequency of fungi detected in situ from fruit bodies on the previous year’s leaf petioles of F. excelsior. Relative species frequency calculated as the ratio of the number of specimens in the order to all specimens n = 882. Percent of species calculated as the ratio of the number of species in the order to all species n = 45.

Figure 6.
Fungal fruit bodies on the previous year’s leaf petioles of F. excelsior in the litter, in 2017: (a–f) apothecia: a—Cyathicula fraxinophila, (b) Cyathicula sp. 2, (c) Hymenoscyphus caudatus, (d) Hymenoscyphus scutula, (e) Helotiales sp. 1, (f) Pyrenopeziza petiolaris, (g) Hypoderma rubi, hysterothecia and pycnidia, (h) Paraophiobolus arundinis pseudothecia, (i) Leptosphaeria sp. 2, pseudothecia, (j) Leptosphaeria sclerotioides, pycnidia, (k) Neonectria sp. 2, perithecia, (l–n) Venturia fraxini, with setae lining the ostiolum of the pseudothecium (l), pseudothecia on the exfoliated periderm (m), pseudothecia on petioles with damage made by small fauna (n), (o) setaceous pycnidia of Pyrenochaeta sp., (p) anamorph of Xylaria sp. 1, (q) anamorph of Xylaria sp. 3, (r,s)—Typhula sp. 2, sclerotia (r) and carpogenic sclerotium (s), (t) Typhula erythropus, carpogenic sclerotium.
Figure 6.
Fungal fruit bodies on the previous year’s leaf petioles of F. excelsior in the litter, in 2017: (a–f) apothecia: a—Cyathicula fraxinophila, (b) Cyathicula sp. 2, (c) Hymenoscyphus caudatus, (d) Hymenoscyphus scutula, (e) Helotiales sp. 1, (f) Pyrenopeziza petiolaris, (g) Hypoderma rubi, hysterothecia and pycnidia, (h) Paraophiobolus arundinis pseudothecia, (i) Leptosphaeria sp. 2, pseudothecia, (j) Leptosphaeria sclerotioides, pycnidia, (k) Neonectria sp. 2, perithecia, (l–n) Venturia fraxini, with setae lining the ostiolum of the pseudothecium (l), pseudothecia on the exfoliated periderm (m), pseudothecia on petioles with damage made by small fauna (n), (o) setaceous pycnidia of Pyrenochaeta sp., (p) anamorph of Xylaria sp. 1, (q) anamorph of Xylaria sp. 3, (r,s)—Typhula sp. 2, sclerotia (r) and carpogenic sclerotium (s), (t) Typhula erythropus, carpogenic sclerotium.

Figure 7.
Fruit bodies of fungi (with frequency >2%) from the previous year’s leaf petioles of F. excelsior according to season. In spring, Typhula spp. produced only sclerotia with no sporophores.
Figure 7.
Fruit bodies of fungi (with frequency >2%) from the previous year’s leaf petioles of F. excelsior according to season. In spring, Typhula spp. produced only sclerotia with no sporophores.

Figure 8.
Hymenoscyphus fraxineus and other fungi on the previous year’s leaf petioles of F. excelsior in the litter: (a,b) H. fraxineus pseudosclerotial plate revealed by the exfoliating epidermis, with pseudothecia of Venturia fraxini (arrows), (c) phialide and endoconidium of Chalara fraxinea on pseudosclerotial plate, (d) apothecia of H. fraxineus (July 2017), (e–n) pseudosclerotial plate of H. fraxineus and: (e,f) Cyathicula coronata, (g) Cyathicula fraxinophila, (h) Trichopeziza sp. 1, (i) Pyrenopeziza petiolaris, (j) Leptosphaeria sp. 2, (k) Lophiostoma corticola, (l) Xylaria sp. 2, (m,n) Typhula sp. 2, sclerotium at the base of petiole (m), carpogenic sclerotium (n), (o) apothecia of C. coronata on pseudosclerotial plate of H. fraxineus.
Figure 8.
Hymenoscyphus fraxineus and other fungi on the previous year’s leaf petioles of F. excelsior in the litter: (a,b) H. fraxineus pseudosclerotial plate revealed by the exfoliating epidermis, with pseudothecia of Venturia fraxini (arrows), (c) phialide and endoconidium of Chalara fraxinea on pseudosclerotial plate, (d) apothecia of H. fraxineus (July 2017), (e–n) pseudosclerotial plate of H. fraxineus and: (e,f) Cyathicula coronata, (g) Cyathicula fraxinophila, (h) Trichopeziza sp. 1, (i) Pyrenopeziza petiolaris, (j) Leptosphaeria sp. 2, (k) Lophiostoma corticola, (l) Xylaria sp. 2, (m,n) Typhula sp. 2, sclerotium at the base of petiole (m), carpogenic sclerotium (n), (o) apothecia of C. coronata on pseudosclerotial plate of H. fraxineus.

Figure 9.
Various types of interactions observed in dual cultures (MEA, 3 weeks, 20 °C), Hymenoscyphus fraxineus (Hf1 or Hf2) from the left, test fungus on the right: (a–d) type A interaction, (a) Hymenoscyphus fraxineus (FeF164), (b) Alternaria sp. 1 (FeF152), (c) Microsphaeropsis sp. (FeF92b), (d) Pyrenochaeta sp. (FeF13), (e–l) type B interaction, (e) Phacidium lacerum (FeF246), (f) Plectosphaerella cucumerina (FeF161b), (g) Hypoderma rubi (FeF203), (h) Pleospora sp. (FeF80), (i) Paraphaeosphaeria neglecta FeF109), (j) Lanzia sp. (FeF26), (k) Malbranchea sp. (FeF243), (l) Pseudocoleophoma polygonicola (FeF201), (m–t) type C interaction, (m) Nemania diffusa (FeF104), (n) Nemania serpens (FeF245), (o) Rosellinia corticium (FeF167), (p) Rosellinia nectrioides (FeF187) (with inhibition zone only partly covered, see text), (q) Xylaria sp. 1 (FeF97a), (r) Xylaria polymorpha (FeF30), (s) Desmazierella acicola FeF119, (t) Hypholoma fasciculare (FeF241).
Figure 9.
Various types of interactions observed in dual cultures (MEA, 3 weeks, 20 °C), Hymenoscyphus fraxineus (Hf1 or Hf2) from the left, test fungus on the right: (a–d) type A interaction, (a) Hymenoscyphus fraxineus (FeF164), (b) Alternaria sp. 1 (FeF152), (c) Microsphaeropsis sp. (FeF92b), (d) Pyrenochaeta sp. (FeF13), (e–l) type B interaction, (e) Phacidium lacerum (FeF246), (f) Plectosphaerella cucumerina (FeF161b), (g) Hypoderma rubi (FeF203), (h) Pleospora sp. (FeF80), (i) Paraphaeosphaeria neglecta FeF109), (j) Lanzia sp. (FeF26), (k) Malbranchea sp. (FeF243), (l) Pseudocoleophoma polygonicola (FeF201), (m–t) type C interaction, (m) Nemania diffusa (FeF104), (n) Nemania serpens (FeF245), (o) Rosellinia corticium (FeF167), (p) Rosellinia nectrioides (FeF187) (with inhibition zone only partly covered, see text), (q) Xylaria sp. 1 (FeF97a), (r) Xylaria polymorpha (FeF30), (s) Desmazierella acicola FeF119, (t) Hypholoma fasciculare (FeF241).


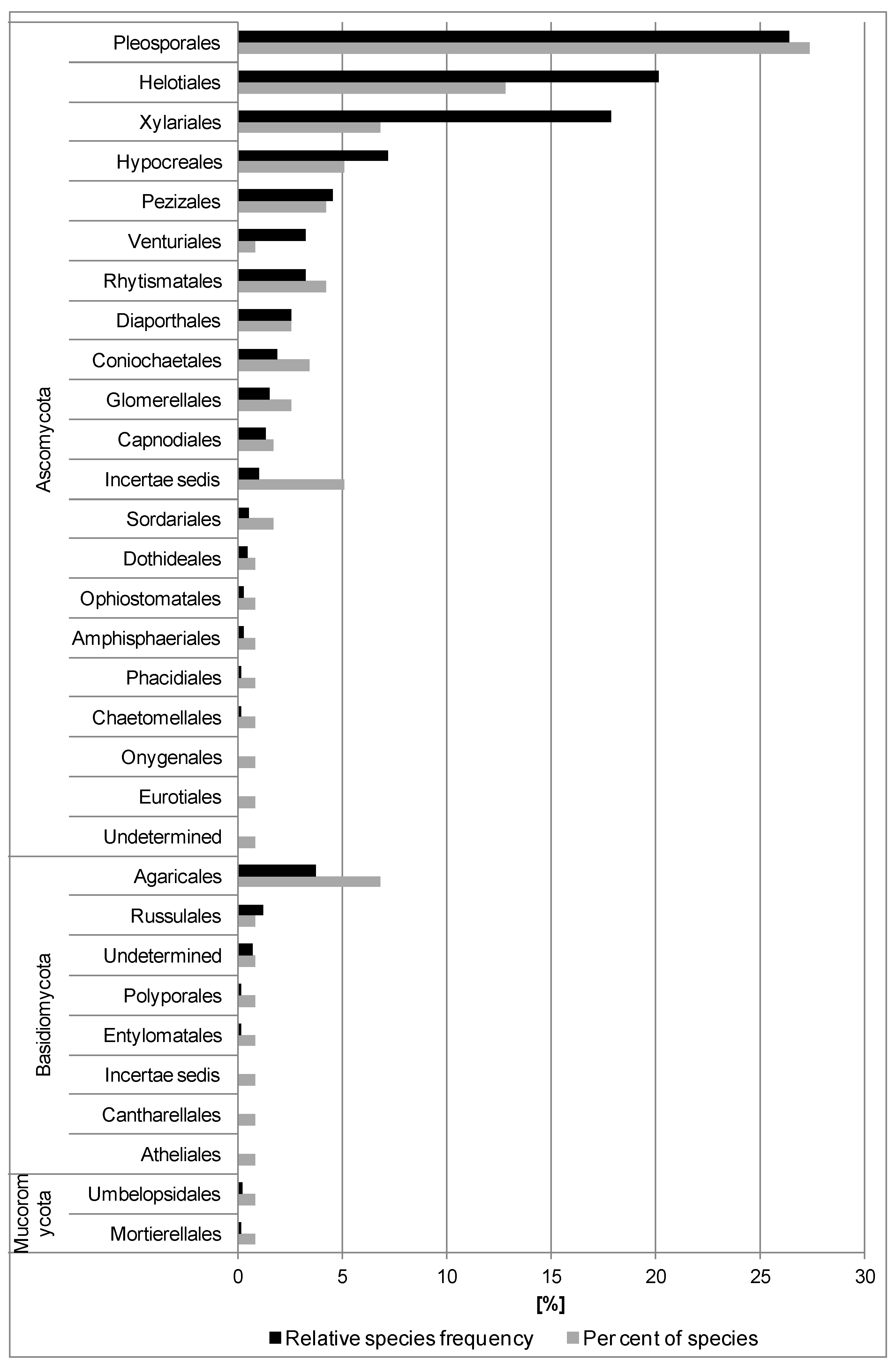{kind=link}
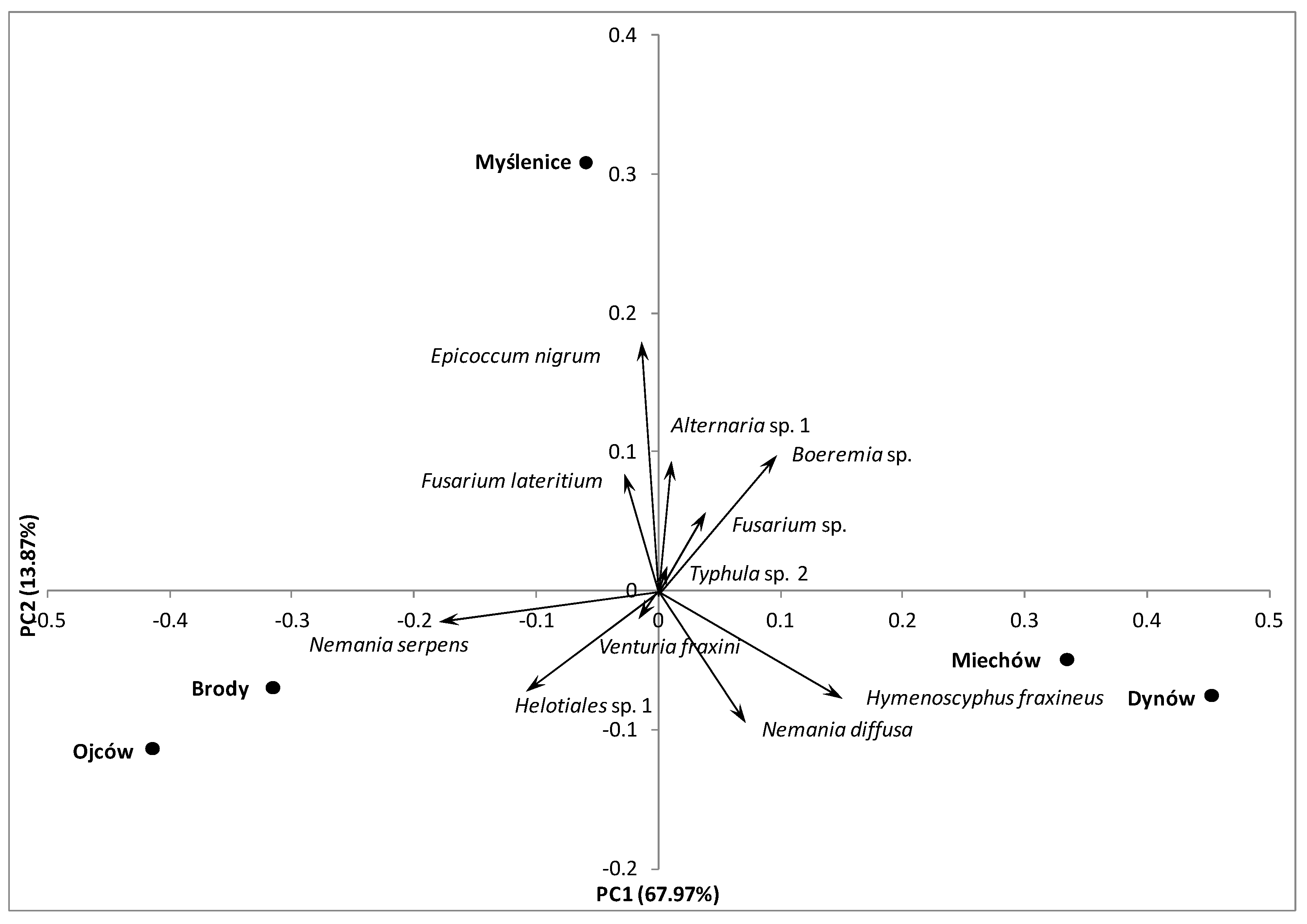{kind=link}
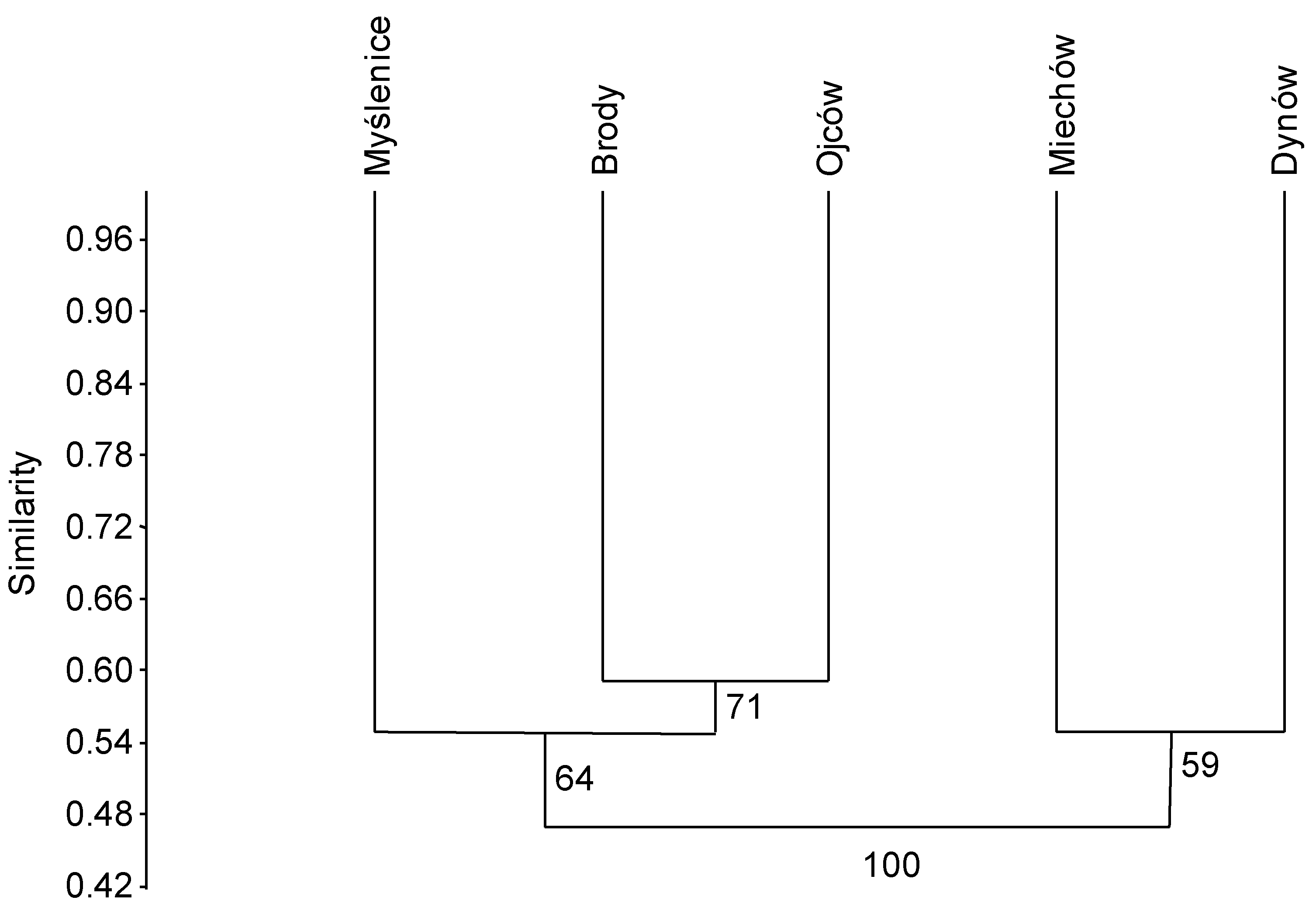{kind=link}
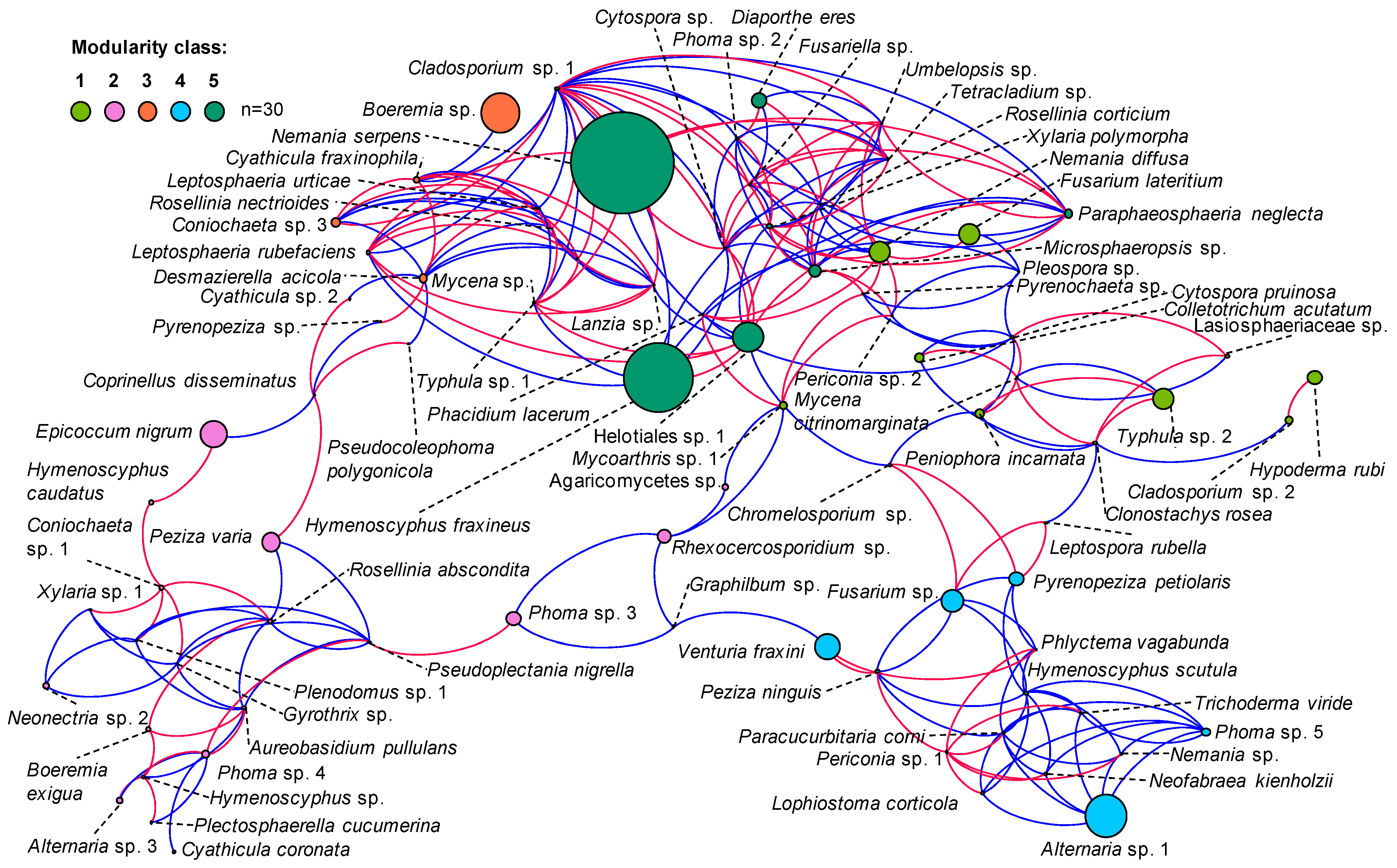{kind=link}
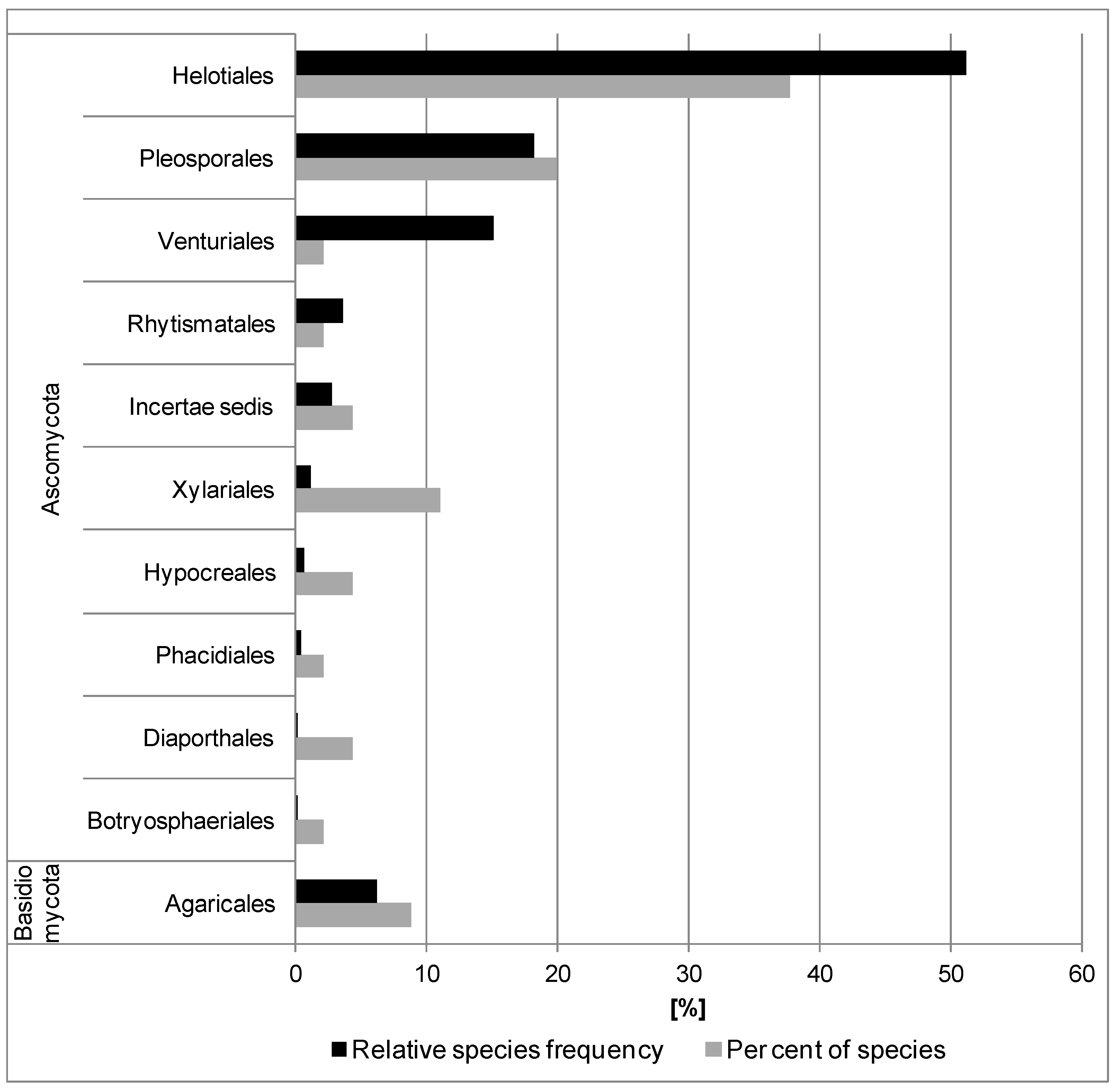{kind=link}
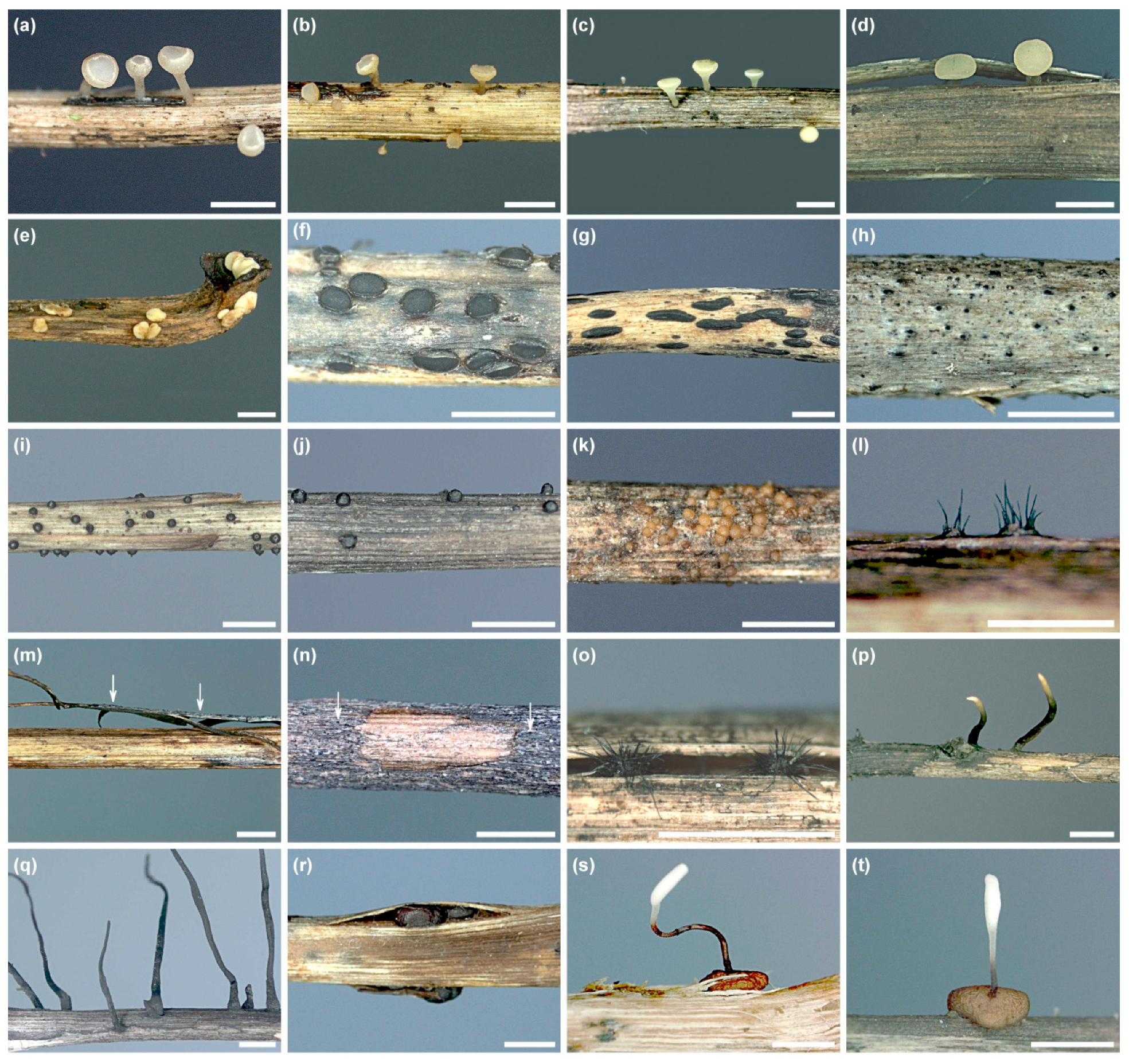{kind=link}
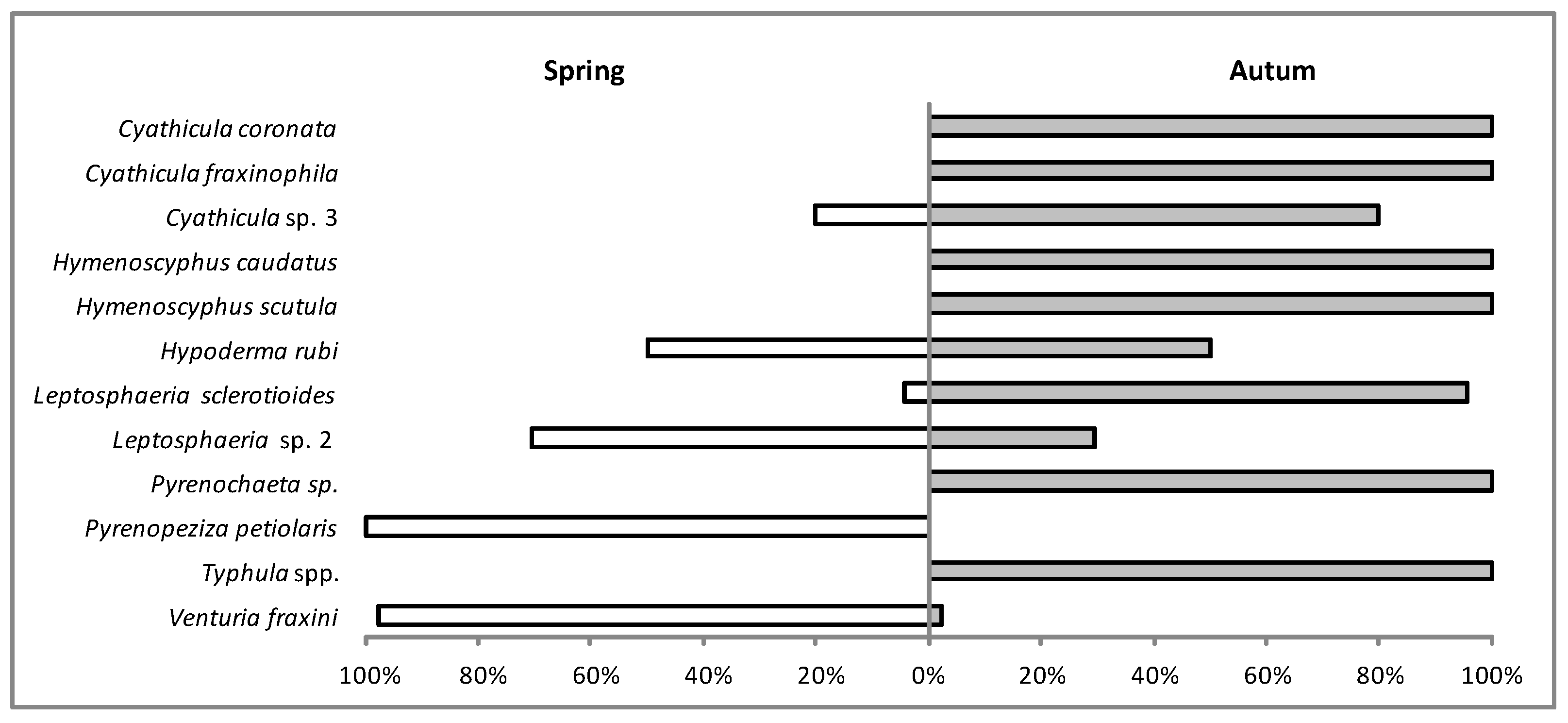{kind=link}
{kind=link}
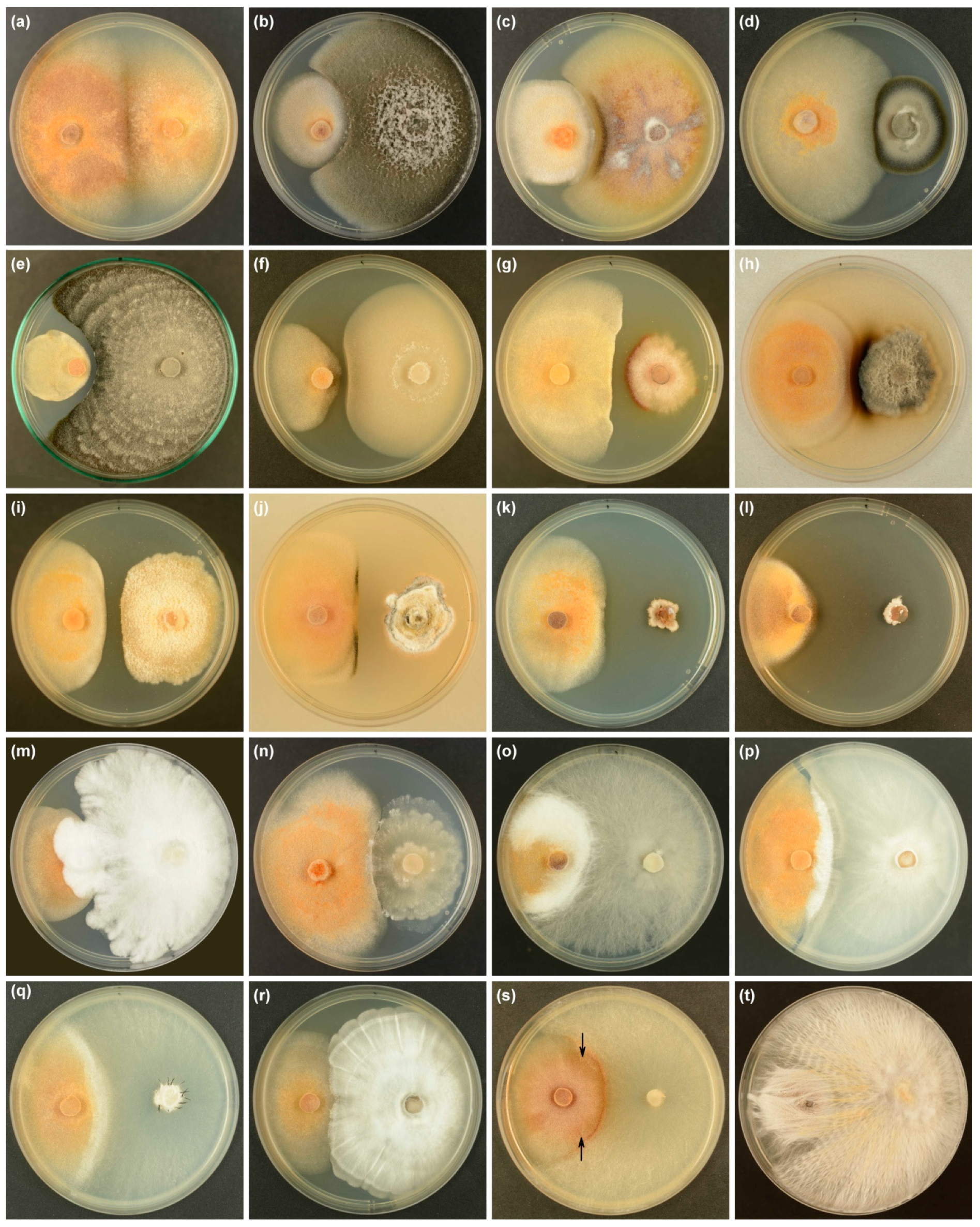{kind=link}
Table 1.
Sampling sites and number of leaf petioles of F. excelsior from the previous year analyzed in the study.
Table 1.
Sampling sites and number of leaf petioles of F. excelsior from the previous year analyzed in the study.
| Research Plots | Brody | Dynów | Miechów | Myślenice | Ojców |
|---|---|---|---|---|---|
| Coordinates | 49°51′34″ N | 49°47′17″ N | 50°13′02″ N | 49°56′45″ N | 50°11′59″ N |
| 19°40′32″ E | 22°15′20″ E | 20°02′09″ E | 19°52′48″ E | 19°48′43″ E | |
| Altitude [m a.s.l.] | 369 | 417 | 281 | 356 | 446 |
| Fraxinus excelsior: | |||||
| age | 54 | 60 | 27 | 82 | 63 |
| Share (%) | 30 | 10 | 10 | 50 | 80 |
| Other tree species present 1 | Ag, Ap, Cb, Qr, Tc | Bp, Cb, Fs, Ps, Qr | Ap, Fs, Ld, Pt, Qr | Ap, Ag, Ld, Qr, Tc | Ap, Bp, Fs, Ps, Qr |
| Habitat type | Fresh upland broadleaved forest | Fresh upland broadleaved forest | Fresh upland broadleaved forest | Fresh upland broadleaved forest | Fresh upland broadleaved forest |
| Sampling date | 10 May 2017 | 16 May 2017 | 08 June 2017 | 23 May 2017 | 30 May 2017 |
| Number of petioles (petiole sections) used in fungal isolations | 60 (720) | 60 (720) | 60 (720) | 60 (720) | 60 (720) |
| Number of petioles (petiole sections) yielding colonies of fungi | 60 (419) | 60 (582) | 60 (465) | 60 (487) | 60 (548) |
1 Ag—Alnus glutinosa, Ap—Acer pseudoplatanus, Bp—Betula pendula, Cb—Carpinus betulus, Fs—Fagus sylvatica, Ld—Larix decidua, Ps—Pinus sylvestris, Pt—Populus tremula, Qr—Quercus robur, Tc—Tilia cordata.
Table 2.
Fungi isolated from the previous year’s leaf petioles of F. excelsior in the litter in five forest stands in southern Poland in spring 2017.
Table 2.
Fungi isolated from the previous year’s leaf petioles of F. excelsior in the litter in five forest stands in southern Poland in spring 2017.
| Taxon | Phylum 1 | Order | Number of Colonized Ash Petioles in Study Plots 2 | |||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | Total Number (%) | |||
| Agaricomycetes sp. | B | - | 4 | 5 | 9 (3.0) | |||
| Alternaria sp. 1 | A | Pleosporales | 11 a 3 | 16 a | 9 a | 20 a | 11 a | 67 (22.3) 4 |
| Alternaria sp. 2 | A | Pleosporales | 1 | 1 (0.3) | ||||
| Alternaria sp. 3 | A | Pleosporales | 2 | 7 | 9 (3.0) | |||
| Alternaria sp. 4 | A | Pleosporales | 1 | 1 (0.3) | ||||
| Athelia sp. | B | Atheliales | 1 | 1 (0.3) | ||||
| Aureobasidium pullulans | A | Dothideales | 3 | 2 | 1 | 6 (2.0) | ||
| Boeremia exigua | A | Pleosporales | 1 | 1 | 3 | 2 | 7 (2.3) | |
| Boeremia sp. | A | Pleosporales | 4 bc | 13 ab | 26 a | 17 a | 2 c | 62 (20.7) 4 |
| Chaetomium globosum | A | Sordariales | 2 | 2 (0.7) | ||||
| Chromelosporium sp. | A | Pezizales | 2 | 1 | 1 | 1 | 5 (1.7) | |
| Cladosporium sp. 1 | A | Capnodiales | 2 | 2 | 1 | 5 (1.7) | ||
| Cladosporium sp. 2 | A | Capnodiales | 3 | 1 | 6 | 1 | 1 | 12 (4.0) 4 |
| Clonostachys rosea | A | Hypocreales | 2 | 2 | 1 | 5 (1.7) | ||
| Colletotrichum acutatum | A | Glomerellales | 4 | 2 | 4 | 4 | 14 (4.7) 4 | |
| Coniochaeta angustispora | A | Coniochaetales | 1 | 1 (0.3) | ||||
| Coniochaeta sp. 1 | A | Coniochaetales | 1 | 2 | 3 | 2 | 8 (2.7) | |
| Coniochaeta sp. 2 | A | Coniochaetales | 2 | 2 (0.7) | ||||
| Coniochaeta sp. 3 | A | Coniochaetales | 6 | 2 | 2 | 3 | 13 (4.3) 4 | |
| Coniothyrium cf. dispersellum | A | Pleosporales | 7 | 7 (2.3) | ||||
| Coprinellus disseminatus | B | Agaricales | 1 | 2 | 3 (1.0) | |||
| Coprinellus micaceus | B | Agaricales | 1 | 1 (0.3) | ||||
| Cyathicula coronata | A | Helotiales | 1 | 1 | 1 | 2 | 5 (1.7) | |
| Cyathicula fraxinophila | A | Helotiales | 1 | 2 | 3 | 2 | 1 | 9 (3.0) |
| Cyathicula sp. 2 | A | Helotiales | 1 | 1 | 1 | 3 (1.0) | ||
| Cytospora pruinosa | A | Diaporthales | 2 | 1 | 2 | 5 (1.7) | ||
| Cytospora sp. | A | Diaporthales | 2 | 1 | 3 (1.0) | |||
| Dactylaria sp. | A | Rhytismatales | 1 | 1 (0.3) | ||||
| Desmazierella acicola | A | Pezizales | 7 | 2 | 1 | 1 | 3 | 14 (4.7) 4 |
| Diaporthe eres | A | Diaporthales | 2 | 1 | 1 | 16 | 4 | 24 (8.0) 4 |
| Discohainesia oenotherae | A | Chaetomellales | 2 | 2 (0.7) | ||||
| Epicoccum nigrum | A | Pleosporales | 8 ab | 1 b | 9 ab | 20 a | 4 b | 42 (14.0) 4 |
| Eupenicillium sp. | A | Eurotiales | 1 | 1 (0.3) | ||||
| Fellozyma sp. | B | Incertae sedis | 1 | 1 (0.3) | ||||
| Fusariella sp. | A | Incertae sedis | 1 | 1 | 1 | 3 (1.0) | ||
| Fusarium lateritium | A | Hypocreales | 10 a | 4 a | 2 a | 12 a | 5 a | 33 (11.0) 4 |
| Fusarium sp. | A | Hypocreales | 2 b | 14 a | 3 ab | 11 ab | 5 ab | 35 (11.7) 4 |
| Graphilbum sp. | A | Ophiostomatales | 3 | 1 | 4 (1.3) | |||
| Gyrothrix sp. | A | Incertae sedis | 1 | 1 | 2 (0.7) | |||
| Helotiales sp. 1 | A | Helotiales | 26 a | 0 c | 3 bc | 6 bc | 14 ab | 49 (16.3) 4 |
| Herpotrichia sp. | A | Pleosporales | 2 | 2 (0.7) | ||||
| Hymenoscyphus caudatus | A | Helotiales | 1 | 5 | 1 | 1 | 8 (3.0) | |
| Hymenoscyphus fraxineus | A | Helotiales | 14 b | 37 a | 34 a | 15 b | 10 b | 110 (36.7) 4 |
| Hymenoscyphus scutula | A | Helotiales | 2 | 1 | 2 | 1 | 6 (2.0) | |
| Hymenoscyphus sp. | A | Helotiales | 2 | 1 | 1 | 4 (1.3) | ||
| Hypholoma fasciculare | B | Agaricales | 1 | 1 (0.3) | ||||
| Hypoderma rubi | A | Rhytismatales | 2 | 5 | 1 | 9 | 6 | 23 (7.7) 4 |
| Lanzia sp. | A | Helotiales | 2 | 1 | 3 (1.0) | |||
| Lasiosphaeriaceae sp. | A | Sordariales | 2 | 3 | 5 (1.7) | |||
| Lemonniera sp. | A | Helotiales | 2 | 2 (0.7) | ||||
| Leptosphaeria conoidea | A | Pleosporales | 1 | 1 (0.3) | ||||
| Leptosphaeria rubefaciens | A | Pleosporales | 4 | 1 | 1 | 6 (2.0) | ||
| Leptosphaeria urticae | A | Pleosporales | 1 | 1 | 1 | 3 (1.0) | ||
| Leptosphaeria sp. 1 | A | Pleosporales | 1 | 1 (0.3) | ||||
| Leptosphaeria sp. 2 | A | Pleosporales | 1 | 1 | 1 | 1 | 4 (1.3) | |
| Leptospora rubella | A | Incertae sedis | 3 | 1 | 4 (1.3) | |||
| Leptospora sp. | A | Incertae sedis | 1 | 1 (0.3) | ||||
| Lophiostoma corticola | A | Pleosporales | 1 | 3 | 4 (1.3) | |||
| Malbranchea sp. | A | Onygenales | 1 | 1 (0.3) | ||||
| Microsphaeropsis sp. | A | Pleosporales | 12 | 4 | 3 | 19 (6.3) 4 | ||
| Mortierella sp. | M | Mortierellales | 2 | 2 (0.7) | ||||
| Mycena citrinomarginata | B | Agaricales | 1 | 1 | 2 (0.7) | |||
| Mycena sp. | B | Agaricales | 1 | 1 | 2 (0.7) | |||
| Mycoarthris sp. 1 | A | Rhytismatales | 1 | 5 | 6 | 12 (4.0) 4 | ||
| Mycoarthris sp. 2 | A | Rhytismatales | 2 | 2 (0.7) | ||||
| Nemania diffusa | A | Xylariales | 0 b | 12 a | 16 a | 0 b | 5 ab | 33 (11.0) 4 |
| Nemania serpens | A | Xylariales | 38 b | 13 c | 25 bc | 34 b | 54 a | 164 (54.7) 4 |
| Nemania sp. | A | Xylariales | 1 | 3 | 4 (1.3) | |||
| Neofabraea kienholzii | A | Helotiales | 2 | 3 | 5 (1.7) | |||
| Neonectria sp. 1 | A | Hypocreales | 4 | 4 (1.3) | ||||
| Neonectria sp. 2 | A | Hypocreales | 2 | 6 | 1 | 9 (3.0) | ||
| Paracucurbitaria corni | A | Pleosporales | 2 | 2 | 4 (1.3) | |||
| Paraophiobolus arundinis | A | Pleosporales | 1 | 1 (0.3) | ||||
| Paraphaeosphaeria michotii | A | Pleosporales | 1 | 1 (0.3) | ||||
| Paraphaeosphaeria neglecta | A | Pleosporales | 9 | 4 | 13 (4.3) 4 | |||
| Peniophora incarnata | B | Russulales | 6 | 1 | 3 | 3 | 2 | 15 (5.0) 4 |
| Periconia sp. 1 | A | Pleosporales | 1 | 1 | 2 | 4 (1.3) | ||
| Periconia sp. 2 | A | Pleosporales | 1 | 1 | 2 (0.7) | |||
| Peziza ninguis | A | Pezizales | 5 | 1 | 6 (2.0) | |||
| Peziza varia | A | Pezizales | 8 | 7 | 5 | 3 | 6 | 29 (9.7) 4 |
| Pezizomycetes sp. | A | - | 1 | 1 (0.3) | ||||
| Phacidium lacerum | A | Phacidiales | 1 | 1 | 2 (0.7) | |||
| Phlyctema vagabunda | A | Helotiales | 2 | 1 | 3 (1.0) | |||
| Phoma sp. 1 | A | Pleosporales | 3 | 3 (1.0) | ||||
| Phoma sp. 2 | A | Pleosporales | 2 | 2 | 4 (1.3) | |||
| Phoma sp. 3 | A | Pleosporales | 1 | 17 | 2 | 3 | 23 (7.7) 4 | |
| Phoma sp. 4 | A | Pleosporales | 1 | 2 | 3 | 5 | 11 (3.7) 4 | |
| Phoma sp. 5 | A | Pleosporales | 5 | 6 | 2 | 13 (4.3) 4 | ||
| Plectosphaerella cucumerina | A | Glomerellales | 1 | 3 | 4 (1.3) | |||
| Plenodomus sp. 1 | A | Pleosporales | 1 | 1 | 2 (0.7) | |||
| Pleospora sp. | A | Pleosporales | 1 | 3 | 4 (1.3) | |||
| Psathyrella piluliformis | B | Agaricales | 1 | 1 (0.3) | ||||
| Pseudocoleophoma polygonicola | A | Pleosporales | 1 | 1 | 1 | 3 (1.0) | ||
| Pseudoophiobolus italicus | A | Pleosporales | 4 | 4 (1.3) | ||||
| Pseudoplectania nigrella | A | Pezizales | 2 | 1 | 3 (1.0) | |||
| Pyrenochaeta sp. | A | Incertae sedis | 1 | 1 | 2 (0.7) | |||
| Pyrenochaetopsis leptospora | A | Pleosporales | 2 | 2 (0.7) | ||||
| Pyrenopeziza petiolaris | A | Helotiales | 1 | 10 | 2 | 6 | 3 | 22 (7.3) 4 |
| Pyrenopeziza sp. | A | Helotiales | 1 | 1 | 2 (0.7) | |||
| Rhexocercosporidium sp. | A | Helotiales | 3 | 4 | 5 | 4 | 5 | 21 (7.0) 4 |
| Rosellinia abscondita | A | Xylariales | 3 | 2 | 1 | 1 | 7 (2.3) | |
| Rosellinia corticium | A | Xylariales | 1 | 1 | 2 (0.7) | |||
| Rosellinia nectrioides | A | Xylariales | 1 | 1 | 2 (0.7) | |||
| Sistotrema sp. | B | Cantharellales | 1 | 1 (0.3) | ||||
| Tetracladium sp. | A | Rhytismatales | 2 | 1 | 3 (1.0) | |||
| Tilletiopsis washingtonensis | B | Entylomatales | 2 | 2 (0.7) | ||||
| Trametes versicolor | B | Polyporales | 2 | 2 (0.7) | ||||
| Trichoderma viride | A | Hypocreales | 1 | 3 | 4 (1.3) | |||
| Truncatella sp. | A | Amphisphaeriales | 4 | 4 (1.3) | ||||
| Typhula sp. 1 | B | Agaricales | 2 | 1 | 1 | 4 (1.3) | ||
| Typhula sp. 2 | B | Agaricales | 3 a | 10 a | 3 a | 8 a | 9 a | 33 (11.0) 4 |
| Umbelopsis sp. | M | Umbelopsidales | 1 | 1 | 1 | 3 (1.0) | ||
| Vargamyces aquaticus | A | Incertae sedis | 1 | 1 (0.3) | ||||
| Venturia fraxini | A | Venturiales | 9 a | 4 a | 11 a | 7 a | 10 a | 41 (13.7) 4 |
| Verticillium dahliae | A | Glomerellales | 1 | 1 (0.3) | ||||
| Xylaria polymorpha | A | Xylariales | 6 | 1 | 1 | 8 (2.7) | ||
| Xylaria sp. 1 | A | Xylariales | 1 | 3 | 4 (1.3) | |||
| Number of petioles analyzed | - | - | 60 | 60 | 60 | 60 | 60 | 300 |
| Number of colonies obtained | - | - | 446 | 696 | 519 | 576 | 595 | 2832 |
| Species richness (S) | - | - | 53 | 70 | 58 | 59 | 58 | 117 |
| Sum of frequency of taxa | - | - | 218 | 272 | 249 | 274 | 238 | 1251 |
| Simpson’s index of diversity (SID, SID = 1 − D) | - | - | 0.94 | 0.96 | 0.94 | 0.95 | 0.93 | 0.96 |
| Shannon index of diversity (H′) | - | - | 3.31 | 3.67 | 3.36 | 3.49 | 3.40 | 3.86 |
1 A—Ascomycota, B—Basidiomycota, M—Mucoromycota. 2 1—Brody, 2—Dynów, 3—Miechów, 4—Myślenice, 5—Ojców. 3 Within row, values with different subscript letters are statistically different (p < 0.05) according to χ2 multiple comparisons test using the Marascuilo procedure. 4 Indicates dominant species (taxa) of fungi determined according to Camargo’s index [84] (1/S). A species was defined as dominant if Pi > 1/S, where Pi is the relative frequency of a species i and S represents species richness.
Table 3.
Fungi detected in situ by inventory of fruit bodies on the previous year’s leaf petioles of F. excelsior in five forest stands, in southern Poland in 2017.
Table 3.
Fungi detected in situ by inventory of fruit bodies on the previous year’s leaf petioles of F. excelsior in five forest stands, in southern Poland in 2017.
| Taxon | Order 1 | Phylum 2 | Taxa No. | Number of Petioles Colonized in Study Plots 3 | Total Number (%) | Hf 6 | Accession | BLASTn | Identities % | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||||||||
| Amniculicola sp. | P | A | FeF402 | 2 | 2 (0.3) | MZ492963 | MK353143 | 92.42 | |||||
| Anthostoma turgidum | X | A | FeF425 | 2 | 2 (0.3) | MZ493125 | KC774617 | 99.84 | |||||
| Apiognomonia hystrix | D | A | FeF401 | 1 | 1 (0.2) | MZ493111 | KX776442 | 100 | |||||
| Boeremia exigua | P | A | FeF403 | 2 | 6 | 8 (1.3) | 1 | MZ493112 | MN540289 | 100 | |||
| Calycina sp. | H | A | FeF404 | 1 | 1 | 2 (0.3) | MZ493113 | JF908571 | 100 | ||||
| Cyathicula coronata | H | A | FeF405 | 1 | 16 | 7 | 3 | 27 (4.5) 5 | 16 | MZ492976 | MH858141 | 100 | |
| Cyathicula fraxinophila | H | A | FeF406 | 21 a 4 | 11 a | 18 a | 12 a | 20 a | 82 (13.7) 5 | 31 | MZ492980 | MK584998 | 99.09 |
| Cyathicula sp. 1 | H | A | FeF407 | 3 | 3 (0.5) | MZ492983 | MK584998 | 98.7 | |||||
| Cyathicula sp. 2 | H | A | FeF408 | 2 | 1 | 2 | 5 (0.8) | MZ492978 | MK584998 | 98.7 | |||
| Cyathicula sp. 3 | H | A | FeF409 | 6 | 1 | 6 | 2 | 15 (2.5) | 5 | MZ493114 | MK584998 | 98.35 | |
| Cyathicula sp. 4 | H | A | FeF445 | 1 | 2 | 3 (0.5) | MZ492982 | MK584998 | 99.09 | ||||
| Diaporthe sp. 1 | D | A | FeF446 | 1 | 1 (0.2) | 1 | MZ493057 | EU552122 | 99.67 | ||||
| Diplodia fraxini | B | A | FeF410 | 1 | 1 | 2 (0.3) | 2 | MZ493115 | MT587347 | 100 | |||
| Helotiales sp. 1 | H | A | FeF413 | 2 | 1 | 4 | 7 (1.2) | MZ492996 | GU174285 | 98.97 | |||
| Helotiales sp. 2 | H | A | FeF414 | 1 | 1 (0.2) | MZ492997 | MH578458 | 98.41 | |||||
| Helotiales sp. 3 | H | A | FeF415 | 2 | 2 | 1 | 2 | 7 (1.2) | 1 | MZ493124 | KY430513 | 99.44 | |
| Hymenoscyphus caudatus | H | A | FeF416 | 3 | 4 | 5 | 4 | 5 | 21 (3.5) 5 | 7 | MZ492985 | KM114539 | 99.38 |
| Hymenoscyphus fraxineus | H | A | FeF417 | 30 bc | 59 ab | 66 a | 21 c | 26 c | 202 (33.7) 5 | MZ492992 | LLCC00000000 | 100 | |
| Hymenoscyphus scutula | H | A | FeF418 | 8 | 2 | 4 | 6 | 7 | 27 (4.5) | 12 | MZ492989 | MK674606 | 99.83 |
| Hymenoscyphus sp. | H | A | FeF419 | 1 | 1 | 2 | 4 (0.7) | MZ492987 | MK674606 | 97.03 | |||
| Hypoderma rubi | R | A | FeF420 | 8 | 4 | 3 | 13 | 4 | 32 (5.3) 5 | 13 | MZ493025 | GU138741 | 99.64 |
| Lasionectria sp. | Hy | A | FeF421 | 1 | 1 (0.2) | MZ493040 | KY607542 | 96.47 | |||||
| Leptosphaeria sclerotioides | P | A | FeF422 | 7 | 14 | 2 | 23 (3.8) 5 | 5 | MZ492959 | FJ179155 | 100 | ||
| Leptosphaeria sp. 2 | P | A | FeF423 | 29 ab | 17 ab | 17 ab | 38 a | 11 b | 112 (18.7) 5 | 39 | MZ492957 | KC965764 | 95.33 |
| Leptospora rubella | Is | A | FeF424 | 2 | 2 | 1 | 1 | 6 (1.0) | 5 | MZ493116 | MH857603 | 99.83 | |
| Lophiostoma corticola | P | A | FeF426 | 2 | 1 | 1 | 4 (0.7) | 2 | MZ493117 | KU712227 | 100 | ||
| Marasmius epiphyllus | A | B | FeF427 | 1 | 1 (0.2) | MZ493079 | FN293008 | 99.66 | |||||
| Neonectria sp. 2 | Hy | A | FeF411 | 2 | 1 | 2 | 5 (0.8) | 1 | MZ493037 | JF735313 | 99.82 | ||
| Paraophiobolus arundinis | P | A | FeF428 | 2 | 2 | 5 | 9 (1.5) | 6 | MZ492953 | MG520945 | 100 | ||
| Phacidium lacerum | Ph | A | FeF429 | 2 | 1 | 1 | 4 (0.7) | 3 | MZ493005 | KU942438 | 100 | ||
| Plenodomus sp. 2 | P | A | FeF430 | 1 | 1 (0.2) | MZ492947 | KU973715 | 100 | |||||
| Pleosporales sp. | P | A | FeF431 | 1 | 1 (0.2) | MZ492961 | KC965786 | 96.51 | |||||
| Pseudocoleophoma polygonicola | P | A | FeF432 | 1 | 1 (0.2) | MZ492973 | NR_154274 | 99.62 | |||||
| Pyrenochaeta sp. | Is | A | FeF433 | 11 | 2 | 4 | 2 | 19 (3.2) | 2 | MZ492948 | MT236853 | 98.98 | |
| Pyrenopeziza petiolaris | H | A | FeF434 | 7 | 10 | 3 | 11 | 31 (5.2) 5 | 12 | MZ493015 | MH857804 | 99.29 | |
| Trichopeziza sp. 1 | H | A | FeF435 | 8 | 1 | 1 | 1 | 11 (1.8) | 1 | MZ493009 | AB481285 | 97.93 | |
| Trichopeziza sp. 2 | H | A | FeF436 | 1 | 1 | 1 | 3 (0.5) | MZ493008 | JN033398 | 99.23 | |||
| Typhula erythropus | A | B | FeF439 | 1 | 1 | 1 | 3 (0.5) | 2 | MZ493118 | MT232359 | 100 | ||
| Typhula sp. 1 | A | B | FeF437 | 1 | 3 | 1 | 6 | 4 | 15 (2.5) | 3 | MZ493119 | KR673716 | 99.86 |
| Typhula sp. 2 | A | B | FeF438 | 10 | 2 | 15 | 10 | 37 (6.7) 5 | 16 | MZ493081 | NR_132792 | 88.65 | |
| Venturia fraxini | V | A | FeF440 | 26 a | 24 a | 27 a | 27 a | 29 a | 133 (22.2) 5 | 53 | MZ493120 | EU035457 | 99.83 |
| Xylaria sp. 1 | X | A | FeF441 | 2 | 2 (0.3) | MZ493122 | MF774330 | 99.83 | |||||
| Xylaria sp. 2 | X | A | FeF442 | 1 | 1 | 2 (0.3) | 1 | MZ493121 | MF774330 | 99.48 | |||
| Xylaria sp. 3 | X | A | FeF443 | 1 | 2 | 3 (0.3) | 1 | MZ493059 | MH578518 | 97.86 | |||
| Xylaria sp. 4 | X | A | FeF444 | 1 | 1 (0.2) | MZ493123 | MH578518 | 97.1 | |||||
| Number of petioles analyzed | - | - | - | 120 | 120 | 120 | 120 | 120 | 600 | - | - | - | - |
| Species richness (S) | - | - | - | 25 | 24 | 17 | 30 | 31 | 45 | - | - | - | - |
| Sum of frequency of taxa | - | - | - | 180 | 171 | 171 | 194 | 166 | 882 | 241 | - | - | - |
| Simpson’s index of diversity (SID, SID=1-D) | - | - | - | 0.9 | 0.84 | 0.79 | 0.91 | 0.91 | 0.89 | - | - | - | - |
| Shannon index of diversity (H′) | - | - | - | 2.61 | 2.36 | 1.99 | 2.72 | 2.82 | 2.73 | - | - | - | - |
1 Order: P—Pleosporales, X—Xylariales, D—Diaporthales, H—Helotiales, B—Botryosphaeriales, R—Rhytismatales, Hy—Hypocreales, Is—Incertae sedis, A—Agaricales, Ph—Phacidiales, V—Venturiales. 2 A—Ascomycota, B—Basidiomycota. 3 Study plots: 1—Brody, 2—Dynów, 3—Miechów, 4—Myślenice, 5—Ojców. 4 Within row, values with different subscript letters are statistically different (p < 0.05) according to χ2 multiple comparisons test using the Marascuilo procedure. 5 Indicates dominant species (taxa) of fungi determined according to Camargo’s index (1/S) [84]. A species was defined as dominant if Pi > 1/S, where Pi is the relative abundance of a species i and S represents species richness. 6 Number of petioles with H. fraxineus and co-occurring fungus.
Table 4.
Interaction types in dual cultures of H. fraxineus and test fungus from the previous year’s leaf petioles of F. excelsior.
Table 4.
Interaction types in dual cultures of H. fraxineus and test fungus from the previous year’s leaf petioles of F. excelsior.
| Interaction Types | Hymenoscyphus fraxineus (Strain) Number (%) | ||
|---|---|---|---|
| Hf 1 2 | Hf 2 2 | Total 3 | |
| A (mutual direct contact of mycelia) | 73 (44.5) a | 69 (42.1) a | 142 (43.3) a |
| B (inhibition zone) 1 | 75 (45.7) a | 79 (48.2) a | 154 (46.9) a |
| C (overgrowth of H. fraxineus colony by a saprotroph) | 16 (9.8) a | 16 (9.8) a | 32 (9.8) b |
| D (overgrowth of saprotroph’s colony by H. fraxineus) | 0 | 0 | 0 c |
| Number of pathogen/co-partner combinations in dual cultures | 164 | 164 | 328 |
| 1 inhibition zone width | |||
| Bs (up to 3 mm) | 29 a | 31 a | 60 (18.3) a |
| Bm (4–5 mm) | 25 a | 26 a | 51 (15.5) ab |
| Bw (6–8 mm) | 10 a | 12 a | 22 (6.7) b |
| Bv (>8 mm) | 11 a | 10 a | 21 (6.4) b |
2 Within rows in two columns (Hf 1 and Hf 2), values with the same letter are not significantly different (p > 0.05) according to χ2 test. 3 Within column, values with different subscript letters are statistically different (p < 0.05) according to χ2 multiple comparisons test using the Marascuilo procedure separately for “interaction types” and “inhibition zone width” datasets.
Table 5.
Reduction in H. fraxineus and the test fungi colonies in dual cultures.
| Reduction Rate | Hymenoscyphus fraxineus | Fungal Co-Partners | ||||
|---|---|---|---|---|---|---|
| Hf 1 1 | Hf 2 1 | Total Number (%) 2 | Hf 1 1 | Hf 2 1 | Total Number (%) 2 | |
| a (<25%) | 5 a | 5 a | 10 (3.0) b | 37 a | 46 a | 83 (25.3) b |
| b (26–50%) | 68 a | 57 a | 125 (38.1) a | 104 a | 97 a | 201 (61.3) a |
| c (51–75%) | 85 a | 93 a | 178 (54.3) a | 5 a | 6 a | 11 (3.4) c |
| d (>75%) | 6 a | 9 a | 15 (4.6) b | 0 | 0 | 0 c |
| f (0%) | 0 | 0 | 0 (0.0) b | 18 a | 15 a | 33 (10.0) bc |
| Total | 164 | 164 | 328 (100.0) | 164 | 164 | 328 (100.0) |
1 Within rows in two columns (Hf 1 and Hf 2), values with the same letter are not significantly different (p > 0.05) according to χ2 test. 2 Within columns, values with different subscript letters are statistically different (p < 0.05) according to χ2 multiple comparisons test using the Marascuilo procedure.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kowalski, T.; Bilański, P. Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus. Forests 2021, 12, 1412. https://doi.org/10.3390/f12101412
AMA Style
Kowalski T, Bilański P. Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus. Forests. 2021; 12(10):1412. https://doi.org/10.3390/f12101412
Chicago/Turabian StyleKowalski, Tadeusz, and Piotr Bilański. 2021. "Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus" Forests 12, no. 10: 1412. https://doi.org/10.3390/f12101412
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.