
Early Field Performance and Genetic Variation of Dalbergia tonkinensis, a Valuable Rosewood in Vietnam
by
 , and
, and
Bernard Dell
1,2,*,
Pham Quang Thu
1,
Nguyen Hoang Nghia
3,
Phi Hong Hai
4,
Nong Phuong Nhung
5 and
Nguyen Minh Chi
1
1
Forest Protection Research Centre, Vietnamese Academy of Forest Sciences, Duc Thang, Bac Tu Liem, Hanoi 11910, Vietnam
2
Agriculture and Forest Sciences, Murdoch University, Murdoch 6150, Australia
3
Research Institute for Sustainable Forest Management and Forest Certification, Hanoi 11910, Vietnam
4
Vietnamese Academy of Forest Sciences, Hanoi 11910, Vietnam
5
Post Graduate Training and International Cooperation Department, Vietnamese Academy of Forest Sciences, Hanoi 11910, Vietnam
*
Author to whom correspondence should be addressed.
Forests 2022, 13(12), 1977; https://doi.org/10.3390/f13121977
Submission received: 26 October 2022
/
Revised: 18 November 2022
/
Accepted: 21 November 2022
/
Published: 23 November 2022
(This article belongs to the Special Issue Advances in Woody Oil Species: Past, Present and Future)
Abstract
:Dalbergia tonkinensis is being promoted in Vietnam for the future supply of prized wood for furniture and chemical extraction. Expansion of the plantation area requires a reliable source of quality seed. This study evaluates the field performance of progeny from mature mother trees in natural forests and urban environments. Trials were established in Tan Son and Doan Hung districts of Phu Tho province and assessed 3 years later. There were significant differences in growth and survival between provenances. The best families reached heights of >3.5 m and came from mother trees in a natural forest. The number of leaflets per leaf on mother trees was positively correlated with height (Ht) and diameter at breast height (Dbh) of the trial trees. The individual heritabilities (ĥ2) for Ht and Dbh were 0.39 and 0.49 for Tan Son, and 0.33 and 0.48 for Doan Hung, respectively. The coefficient of additive variation (CVA) of Ht and Dbh were 36% and 58% for Tan Son, and 23% and 30% for Doan Hung, respectively. Interactions between the growth traits in the two trials were strongly correlated (R2 = 0.88 for Ht, 0.90 for Dbh). These results identify D. tonkinensis gene sources with advantages for early growth performance in plantations. These superior genetics can be used for seed orchards, clonal propagation and for implementing a breeding program.
1. Introduction
Dalbergia is a large pantropical genus containing important timber trees many of which are endangered from overexploitation of wild populations for their valuable heartwood [1,2,3]. Dalbergia tonkinensis Prain occurs naturally in forests of Vietnam, and mature trees can obtain heights of 15–25 m and diameters of 50–90 cm after 60 years [4]. The wood is highly sought after for furniture production [5] and for the extraction of pharmaceutical and other compounds [6,7,8,9]. A closely related taxon, D. odorifera, occurs in south China, and there is a long history of its use in Chinese culture [10]. Recent analysis of the plastid genome has confirmed that D. tonkinensis and D. odorifera are closely related; however, they come from two independent lineages [11].
In 2006, D. tonkinensis was added to annex I in decree 32/2006/ND-CP of the Vietnamese government to prohibit exploitation from the wild due to the extent of habitat destruction and illegal harvesting [12]. Later, the Ministry of Agriculture and Rural Development (MARD) included D. tonkinensis as a major planting species in Vietnam and promoted the development of D. tonkinensis as a plantation tree for the future supply of valuable wood [13]. In 2019, D. tonkinensis was transferred to annex II in decree 06/2019/ND-CP [5,13], allowing farmers and households to cut trees that they have grown after obtaining the relevant permits [5,14].
Over the last decade, small-holder plantings of D. tonkinensis have increased to about 2000 ha [7], mainly using seedlings sourced from parent trees of low quality [15,16]. So far, domestication of the species has taken place without a strong forest science input. The mating system of open-pollinated D. tonkinensis trees has not been investigated. Research has addressed nursery production of seedlings [17], seed germination [18], vegetative propagation [19], and diseases causing stem canker [20,21]. No research on tree selection, sylviculture or field performance has been undertaken. In contrast, there has been considerable effort in genetic improvement, plantation sylviculture and heartwood induction for D. odorifera in China [22,23,24].
As mature D. tonkinensis trees continue to be lost due to illegal harvesting, there is an urgent need to quantify the performance of planting stock obtained from a range of mother trees. Therefore, two field trials were undertaken to identify superior mother trees that could be cloned for preservation of the genotypes, as well as providing a source of material for establishing clonal hedges, and to identify families with superior early growth that could be cloned and used to establish seed orchards. We sought out superior D. tonkinensis trees in a range of habitats from conservation forests to temple grounds and collected seeds for undertaking field trials. The trials were established throughout a short duration of 3 years in order to provide early input for future clonal propagation and breeding programs.
2. Materials and Methods
2.1. Seed Source
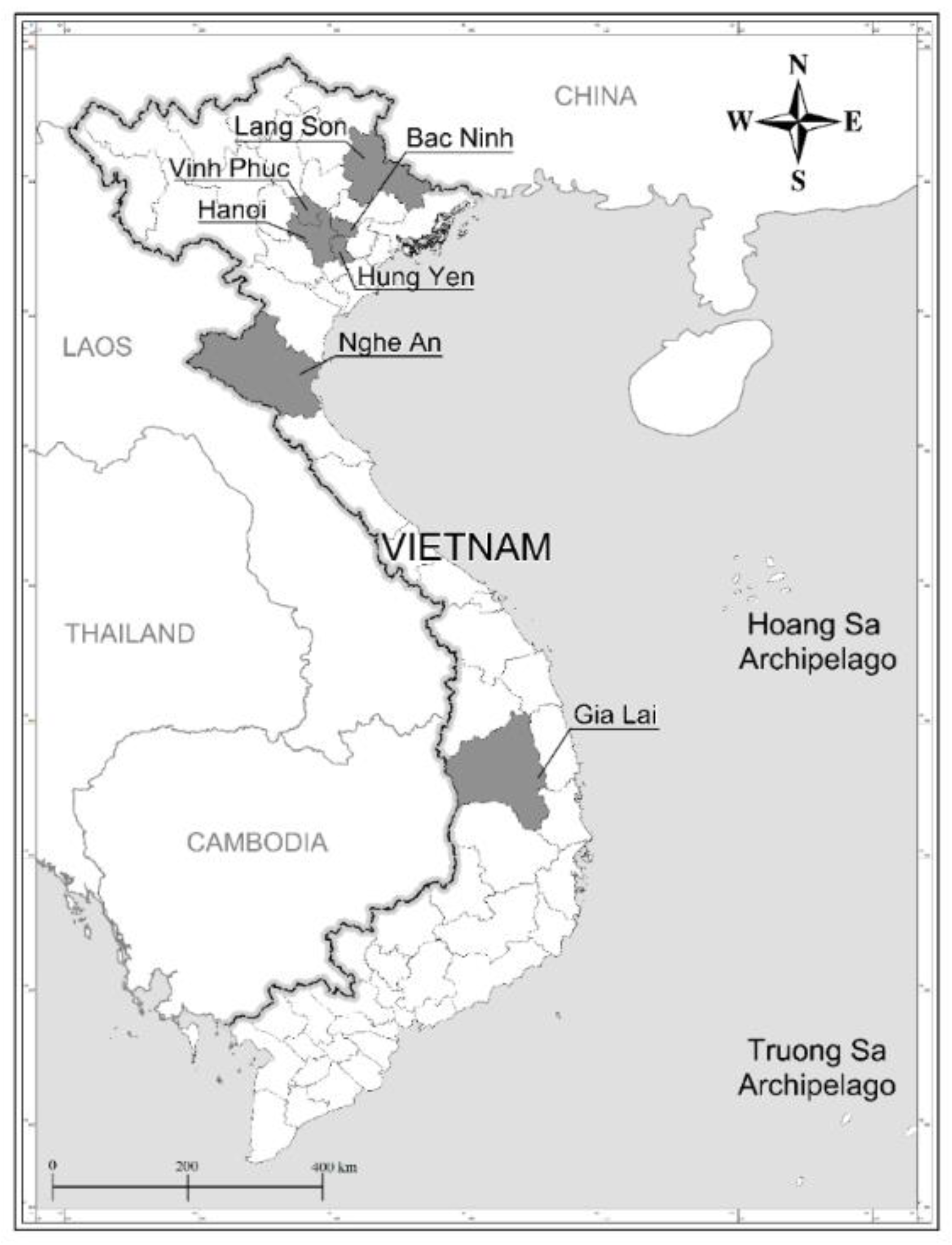
Fruit was collected from 69 mature mother trees, in seven provinces of Vietnam (Figure 1), when the colour of the legumes changed from green to yellow or brown in early winter (December). About 200–300 seeds were obtained from each mother tree.
The criteria used in selecting mother trees were: diameter at breast height (Dbh, 1.3 m) >40 cm; boles straight, round and length >50% of tree height; and trees healthy without visible damage from pests and diseases. The location of suitable trees was sourced through extensive forest and local government networks. The chosen mother trees were located as follows: 26 trees in temple grounds, 16 along streets, 12 in grounds of government offices, 8 in city parks, and 7 in natural forests (Supplementary Table S1). Examples are shown in Figure 2. From local knowledge, the trees were estimated to be more than 60 years old. Height was measured with a height pole, and Dbh was obtained using a diameter tape. We used the number of leaves as a measure of morphological variation between mother trees. The number of leaves was determined for 32 compound leaves that were collected from branches in the middle of the canopy in the four cardinal directions. The seed were manually separated from the fruit. The seeds were dried in a laboratory in Hanoi using a dehumidifier for 5–7 days. Each family’s seeds were vacuum-sealed in separate plastic bags and stored in the dark at 5 °C until use.
2.2. Seedling Production
The seed were placed in cloth bags in water at 70 °C for 12 hours, then drained and kept moist at 35 °C. After 2–3 days the germinated seeds were planted into nursery sand beds that had been solar pasteurized. When 2 pairs of true leaves were present, the seedlings were transplanted into plastic bags (diameter 6 cm, heigh 12 cm) and grown in a research nursery in Hanoi. The container substrate was a red-yellow oxisol taken from the B horizon under a pine forest in Ngoc Thanh award, Phuc Yen district, Vinh Phuc province. A dilute liquid NPK fertilizer was applied when the plants were 2 months old. The seedlings were outplanted at 6 months of age when they were 70–80 cm in height.
2.3. Description of The Progeny Trials
Two sites were selected for evaluating the early field performance of the families. The first was a 2 ha area at Tan Son in Phu Tho province, in north Vietnam; 21.161952 N, 105.089013 E, and 168 m a.s.l. The mean annual temperature is 22 °C, sunshine hours 2517 y−1, and the average rainfall is 1565 mm y−1. The terrain is undulating hills. The soil is a red-yellow oxisol, >50 cm in depth overlying gravel. The site had previously been used for 2 rotations of tea (Camellia sinensis) over 18 years. The second site was a 2 ha area at Doan Hung in Phu Tho province; 21.535924 N, 105.198891 E, and 225 m a.s.l. The mean annual temperature is 23 °C, sunshine hours 2529 y−1, and the average rainfall is 1973 mm y−1. The terrain is flat and has a similar soil type. The site had previously had one rotation of Acacia mangium. The soils are acidic and contain low levels of organic matter, total N, total P2O5 total and total K2O. However, the N, P2O5 and K2O levels in Doan Hung are a little higher than in Tan Son (Supplementary Table S2). Vegetation was dug up by hand and taken off-site. The land was then disc plowed with a tractor. The planting holes (30 × 30 × 30 cm) were dug manually using a shovel and 300 g of NPK (N:P2O5:K2O, 5:10:3) compound fertilizer was added to each hole, mixed with the soil and backfilled.
2.4. Experimental Design
There were two experiments, one at each field site. The experimental design was a randomized block design comprising 4 blocks and 8 replicate plots for each family. There were 59 families common to the two experiments plus an additional 10 families in the Tan Son trial. The unequal numbers were due to the limited availability of planting stock. Each row plot had four trees. A computer program CycDesigN [25] was used to assist the randomization of the families within blocks. The plots and blocks were contiguous. Both experiments were planted in July 2015. Tree spacing was 2 m within rows and the row spacing was 3 m.
2.5. Silviculture
All weeds in the experiments were cut by hand in October 2015 and then in March, August and November for the next 3 years. The soil was lightly cultivated around the base of all trees using a hand hoe and top-dressed with 300 g/tree of NPK (16:16:8) compound fertilizer in March 2016, 2017, 2018. The NPK was banded at the edge of the canopy. For the first year, the trees were supported with bamboo poles. The experimental fields were fenced to exclude water buffalo that were damaging forestry plots in the area.
2.6. Data Collection
At three years, the survival, height (Ht), Dbh and disease level of all trees of each family were assessed. Height was measured using a height pole, and the circumference was measured with a tape and converted to Dbh. The level of stem canker disease was assessed as follows [15]: 0 = no lesions on stem and branches, healthy and green leaves; 1 = length of lesion <10 cm, healthy and green leaves; 2 = length of lesion 10 to <20 cm, some leaf yellowing; 3 = length of lesion 20 to < 30 cm, all leaves yellow; 4 = length of lesion >30 cm, leaves wilted, dried, fallen, and/or dead.
The mean disease index (DI) of each clone was determined using Equation (1):
where: ni is the number of diseased trees at disease level i; vi is the value (range 0 to 4) of the ith level of the disease; and N is total number of plants assessed.
DI = (Ʃnivi)/N
2.7. Data Analysis
Data were analysed using ASReml 4.0 software package. Analysis of variance (ANOVA) was used to test for significant effect of blocks and treatments, followed by Duncan’s Multiple Range Test for comparisons of means of different seed lots. ASReml 4.0 was used to test GxE correlation on the growth of families between the two trials, and individual heritabilities and coefficients of additive genetic variation for the studied traits of families [26].
Family variance (σf2), phenotypic variance (), plot variance (), and environmental variance () for different traits were estimated using ASReml. The estimated variance components were used to calculate the narrow-sense heritabilities for the characters under consideration. Since families in the progeny test came from open-pollinated parent trees, the additive genetic variance () was estimated as three times the family variance component. The additive genetic variance (), total phenotypic variance (), and individual-tree heritability (ĥ2) estimates were calculated using Equations (2)–(4):
σA2 = 3σf2
σP2 = σf2 + σt2 + σe2
Coefficient of additive variation (CVA) was estimated using Equation (5):
where: is the phenotypic mean; σA is additive genetic variance.
Theoretical genetic gain (RY) was estimated using Equation (6):
where: in,N is the intensity of selection based on selection n families from N families participating in the trial; h2Y is the heritability of the trait Y; is the phenotypic variance of trait Y.
Correlation analysis was used to denote the pairwise interaction among quantitative variables obtained from mother trees (MotherH, MotherD, Leaflets and Seed Weight), and the field trials (Ht, Dbh, DI, StemNumber and Survival) using the ggpubr package in R statistical software [27]. Based on the correlation matrix presenting the correlation coefficients, where the correlation was significant, linear regression analysis of the relation between the dependent variables (Ht, Dbh and Survival) and the explanatory variables (Leaves) was explored. The coefficient of determination (R2) was defined in our linear regression models and measures the proportion of variation in the dependent variable explained by the predictors included in the model. To extend it for generalized linear models, we used the variance function to define the total variation of the dependent variable, as well as the remaining variation of the dependent variable after modeling the predictive effects of the independent variables.
3. Results
3.1. Mother Tree Data
The heights of mother trees (range and mean) were 15.5–(21.1)–28.5 m, and the diameters (1.3 m) were 41.4–(62.4)–84.6 cm. A noticeable morphological difference between trees was the number of leaves which ranged from 11 to 17 (mean 15.2). Additionally, there was considerable variation in 1000 seed weight between mother trees: 64.3–(92.1)–155.7 g.
3.2. Field Performance
3.2.1. Tan Son
There were significant differences in growth and survival after 3 years between the seven provenances. On average, the tallest progenies originated from Bac Ninh and Hung Yen provenances (Table 1, Figure 3). However, Hanoi and Gia Lai provenances had the largest diameters, and Hung Yen and Hanoi provenances showed the best survival (Table 1). There was no difference in the number of stems or disease index between the provenances. There was a significant effect of family on growth and survival (p < 0.001). Twelve families achieved heights >3.5 m, more than 29% higher than the trial average (Supplementary Table S3). As for provenances, there was no significant effect of family on the disease index or the number of stems.
The correlation matrix for the characteristics of mother trees (Ht, Dbh, leaves, 1000 seed-weight) and the average of their progenies in the field trial (Height, Dbh, disease index, number of stems, survival) in Tan Son is given in Supplementary Figure S1. The number of leaves on mother trees was positively correlated with tree height, Dbh and survival (p < 0.05). The best linear regression equations between leaves of mother trees and trial factors were for height (y = −1.89 + 0.34x; R2 = 0.74) and Dbh (y = −1.20 + 0.21x; R2 = 0.53).
3.2.2. Doan Hung
There were significant effects of provenance on growth and survival at 3 years of age. Overall, the best provenance was Hanoi, with superior height, Dbh and survival (Table 2, Figure 4b). In addition, the Vinh Phuc provenance showed superior height (Figure 4a).
The height performance of the best families was similar to the trial in Tan Son and 11 of the families were also in the top 12 in Tan Son (Supplementary Table S4). Analysis of the pairwise interaction between characteristics of mother trees and trial factors showed a much weaker correlation between leaflet number and tree growth than in the Tan Son trial (Supplementary Figure S2).
3.3. Heritability
For the Tan Son trial, the individual heritability (ĥ2) of height and Dbh at age 3 years were 0.39 and 0.49, respectively. Coefficients of additive variation (CVA) of height and DBH were 36 and 58%, respectively. Theoretical genetic gain (RY) of height and Dbh were 40.0 and 71.2%, respectively (Table 3a).
The individual heritabilities (ĥ2) of height and diameter at breast height were 0.33 and 0.48, respectively, for the Doan Hung trial. The coefficients of additive variation (CVA) for height and Dbh were 25 and 30%, respectively. Theoretical genetic gain (RY) of height and Dbh were 25.7 and 36.5%, respectively (Table 3b).
3.4. Genotype by Environment Interaction
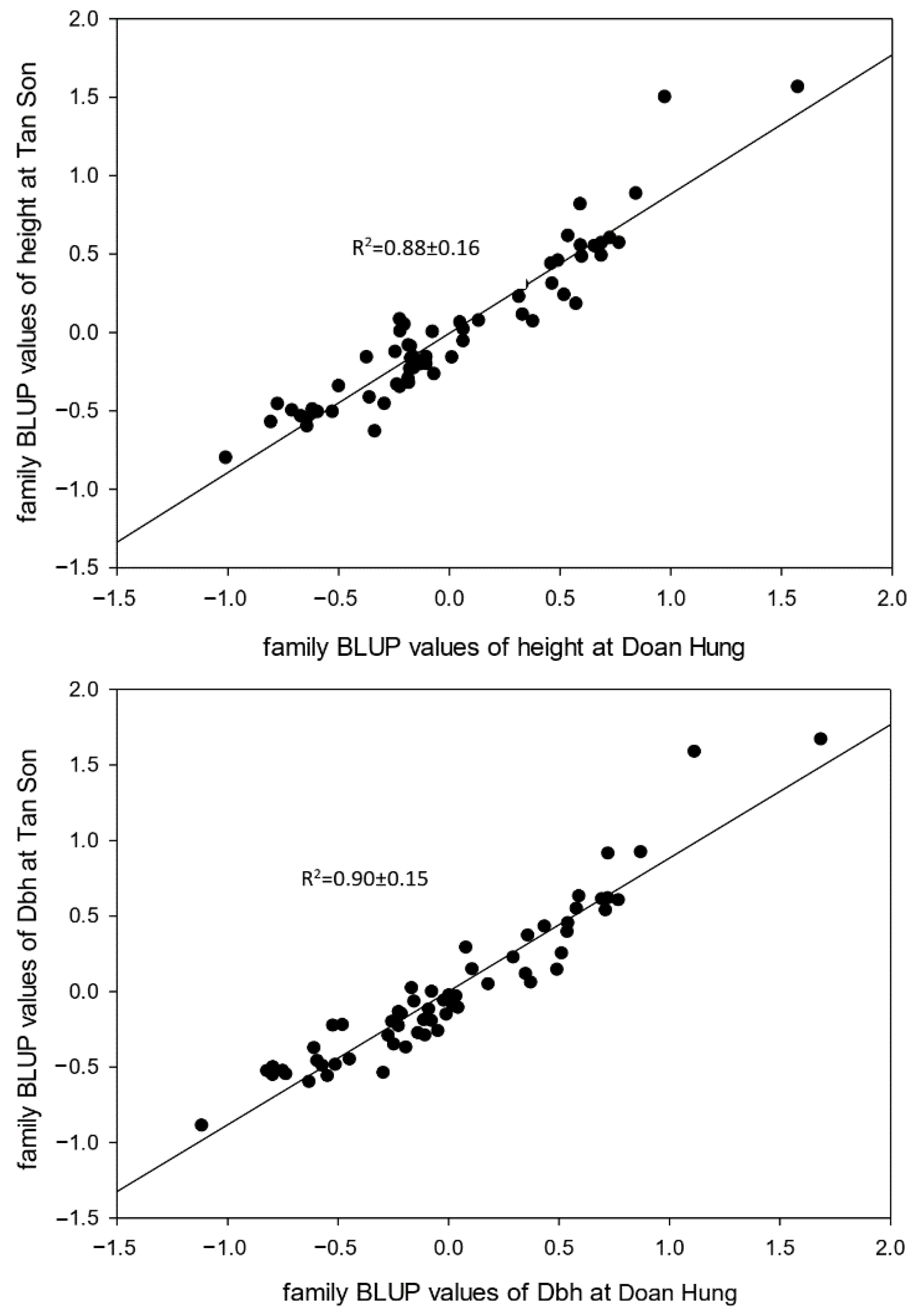
The interaction between the growth traits in the Tan Son and Doan Hung trials was strongly correlated (0.88–0.90), which proves that families with good growth in one site will perform well at the second site—in this case, Doan Hung (Figure 5).
3.5. Location of Mother Trees
Location of mother trees had a significant effect on the height and Dbh of their progenies in the Tan Son trial, and on the height in the Doan Hung trial (Table 4). The highest and thickest progenies all came from mother trees located in natural forest. The poorest performing families were from mother trees in the courtyards of government office buildings or in city parks.
4. Discussion
This is the first study to evaluate the field performance of progenies from mature Dalbergia tonkinensis mother trees growing in natural forests and urban environments. There were significant differences in growth between provenances in the two field trials, with heights ranging from 1.3 to 1.7 m/year and diameters from 1.2 to 1.6 cm/year. Previous investigations have explored the growth of D. tonkinensis in home gardens or in forest restoration using seed from unknown parents. In these studies, D. tonkinensis performed well in moist fertile soil [15,17], reaching maximum heights of 1.5 m/year, and diameters of 1.6 cm/year [15,28]. The poorer growth of trees in Tan Son district may have been due to soil constraints arising from the previous production of tea crops. The Doan Hung soil had higher organic C, total N, soluble P and K2O than in Tan Son (Supplementary Table S2). In the closely related D. odorifera, growth of 2-year-old trees was strongly influenced by site condition. They grew well in the lower parts of hills, achieving a mean height of 2.88 m and mean Dbh of 2.29 cm [29]. Likewise, there was a strong correlation between growth attributes of D. sisso and soil quality [30].
There are no records on the origin of the mother trees in urban settings. It is likely that the mother trees were planted as seedlings; however, we cannot rule out self-regeneration from seed or regeneration from roots. In the future it would be interesting to determine the extent of any inbreeding in progenies from these mother trees as observed in studies on Acacia [31].
Although the soil fertility differed between the two sites, growth traits of D. tonkinensis were strongly correlated between the trials. Nevertheless, progeny from city park trees performed better on the fertile site at Doan Hung than in poor soil at Tan Son. To manage genotype × environment interaction (G × E interaction), usually the best families need to be selected for specific sites to maximize deployment gains [32]. For example, G × E effects were of practical importance for breeding of A. mangium [31]. Significant G × E interactions have been reported in D. sissoo in India [33]. The results of our study are only preliminary. Further studies need to be undertaken to clone the plus trees and test their performance on a wider range of sites.
In our trials, 12 D. tonkinensis families in Tan Son and 8 families in Doan Hung had height growth higher than the trial average. At present, D. tonkinensis seeds are being collected mainly from trees grown in home gardens in Vinh Phuc [5,15]. The Vinh Phuc provenance tended to grow poorly in the Tan Son soil (Table 2). However, family DT66 from Vinh Phuc had good growth in both field trials.
Canker disease caused by Fusarium lateritium and F. decemcellulare can kill young trees [20]. We found that families with the best growth had disease indices that were lower than the trial average. These families should be assessed over the next decade to establish whether disease resistance is maintained. In addition, the timing and extent of heartwood development should be determined. In the meantime, these are promising families for plus tree selection and clonal propagation [19]. In D. sisso, the best clones had superior growth compared to the original mother trees [34]. There are no MARD-recognized sources of D. tonkinensis genetics in Vietnam [16,19]. Therefore, these two trials also have the potential to become seed production areas. Some trees were already producing seeds at 3 years of age.
The theoretical genetic gain (RY) of height and diameter at breast height were 25.7%–40.0% and 36.5%–71.2%, respectively, indicating good potential for the selection of D. tonkinensis clones from these trials. In D. sissoo, five clones with good growth performance had heritability estimates of 87%, 83% and 80% for height, crown width and crown length, respectively [35]. In this study, the individual heritability (ĥ2) of height and diameter at breast height of D. tonkinensis at age 3 years were 0.33–0.39 and 0.48–0.49, respectively. These values are higher than or similar to the individual heritability of fast-growing species such as Acacia auriculiformis, ĥ2 = 0.36–0.39 [36], A. mangium, ĥ2 = 0.11–0.30 [31], and Eucalyptus urophylla, ĥ2 = 0.10–0.31 [37]. From this result, and with a high (25.35%–58.06%) coefficient of additive variation (CVA), we suggest that the improvement of D. tonkinensis in terms of growth traits is completely possible.
Although D. tonkinensis has been over-exploited in Vietnam [5,15,28], we were able to locate 7 healthy mature trees with good form in natural forests that were bearing seed. Greater efforts should be made to locate other plus trees in the wild and to protect them. We can-not disclose the location of the mother trees due to the risk of illegal harvesting. Since the study began, already some of the mother trees in temple grounds have been lost. There is an urgent need to ensure the natural gene pool is preserved not only for biodiversity purposes but also for the future of the rosewood industry in Vietnam. So far, no population study has been undertaken to determine the genetic diversity of D. tonkinensis in the wild or in cultivation. In China, the core germplasm collection of D. odorifera contains 6 wild and 25 cultivated individuals [38]. There are about 100 mature mother trees in parks and pagodas in Vietnam [16] that could be used in conjunction with the genetics in our two field trials to help build a germplasm collection.
5. Conclusions
We collected seeds from plus trees in the wild and in urban settings and set up provenance and progeny trials to quantify early growth, form and disease resistance in north Vietnam. We found that seed from mother trees in natural forests generally outperformed those from urban trees. For the first time for this valuable rosewood, there is a reliable germplasm collection that can be used as seed orchards, to develop clonal plantations and as genetic resources for implementing a breeding program.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/f13121977/s1, Table S1: Number of mother trees in each province according to location. Table S2: Physical and chemical properties of soil in the trials. Table S3: Growth, number of stems and disease index of the 12 best performing Dalbergia tonkinensis families in Tan Son. Table S4: Growth, number of stems and disease index of the 12 best performing D. tonkinensis families in Doan Hung. Figure S1: Correlation analysis of the pairwise interaction between characteristics of mother trees (Ht, Dbh, leaves, 1000 seed-weight) and trial factors (Ht, Dbh, DI, number of stems, survival) in Tan Son. Figure S2: Correlation analysis of the pairwise interaction between characteristics of mother trees (Ht, Dbh, leaves, 1000 seed-weight) and trial factors (Ht, Dbh, DI, number of stems, survival) in Doan Hung.
Author Contributions
Conceptualization, B.D.; Setting trial, N.M.C., N.H.N. and P.Q.T.; Rearing and implementation, N.M.C., N.P.N., P.H.H. and P.Q.T.; Analysis, P.H.H., N.P.N. and N.M.C.; Methodology, B.D., N.M.C. and N.H.N.; Validation, B.D. and N.M.C.; Visualization, B.D., N.H.N. and N.M.C.; Writing—original draft, N.M.C. and B.D.; Writing—review and editing, B.D. and N.M.C. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by an ASEAN-Korea Environmental Cooperation (AKECOP) Project titled “Ex situ Conservation of Dalbergia tonkinensis Prain, a very valuable tree species of Vietnam” for the fiscal year 2014–2015, and by Gondess Pty Ltd.
Data Availability Statement
Datasets presented in this study are available to the user on request.
Acknowledgments
We thank the Tan Son Forest Company and the Forest Science Centre for Central North Vietnam for their cooperation and assistance in trial maintenance; and Duy Long Pham, Nguyen Van Thanh and Tran Xuan Hinh for help with data collection.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Vatanparast, M.; Klitgård, B.B.; Adema, F.A.C.B.; Pennington, R.T.; Yahara, T.; Kajita, T. First molecular phylogeny of the pantropical genus Dalbergia: Implications for infrageneric circumscription and biogeography. S. Afr. J. Bot. 2013, 89, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Crameri, S.; Fior, S.; Zoller, S.; Widmer, A. A target capture approach for phylogenomic analyses at multiple evolutionary timescales in rosewoods (Dalbergia spp.) and the legume family (Fabaceae). Mol. Ecol. Resour. 2022, 22, 3087–3105. [Google Scholar] [CrossRef]
- Rahaingoson, F.R.; Oyebanji, O.; Stull, G.W.; Zhang, R.; Yi, T.S. A dated phylogeny of the pantropical genus Dalbergia L.f. (Leguminosae: Papilionoideae) and its implications for historical biogeography. Agronomy 2022, 12, 1612. [Google Scholar] [CrossRef]
- Nguyen, H.N. Atlas of Vietnam’s Forest Tree Species; Cartographic Publishing House: Hanoi, Vietnam, 2008; Volume 2, p. 249. [Google Scholar]
- Nhung, N.P.; Chi, N.M.; Thu, P.Q.; Thuong, B.H.; Ban, D.V.; Dell, B. Market and policy setting for the trade in Dalbergia tonkinensis, a rare and valuable rosewood, in Vietnam. Trees For. People 2020, 1, 100002. [Google Scholar] [CrossRef]
- Vo, V.C. Dictionary of Vietnamese Medicinal Plants; Medical Publishing House: Hanoi, Vietnam, 1999. [Google Scholar]
- Thu, P.Q.; Quang, D.N.; Chi, N.M.; Hung, T.X.; Binh, L.V.; Dell, B. New and emerging insect pest and disease threats to forest plantations in Vietnam. Forests 2021, 12, 1301. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.-L.; Nhan, N.T.; Nguyen, T.H.; Nguyen, N.P.D.; Nghi, D.H.; Cuong, N.M. New records of potent in-vitro antidiabetic properties of Dalbergia tonkinensis heartwood and the bioactivity-guided isolation of active compounds. Molecules 2018, 23, 1589–1601. [Google Scholar] [CrossRef] [Green Version]
- Cuong, N.M.; Son, N.T.; Nhan, N.T.; Fukuyama, Y.; Ahmed, A.; Saponara, S.; Trezza, A.; Gianibbi, B.; Vigni, G.; Spiga, O.; et al. Vietnamese Dalbergia tonkinensis: A promising source of mono- and bifunctional vasodilators. Molecules 2022, 27, 4505. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, C.; Meng, H.; Yu, Z.; Yang, M.; Wei, J. Dalbergia odorifera: A review of its traditional uses, phytochemistry, pharmacology, and quality control. J. Ethnopharmacol. 2020, 248, 112328. [Google Scholar] [CrossRef]
- Qin, M.; Zhu, C.J.; Yang, J.B.; Vatanparast, M.; Schley, R.; Lai, Q.; Zhang, D.Y.; Tu, T.Y.; Klitgård, B.B.; Li, S.J.; et al. Comparative analysis of complete plastid genome reveals powerful barcode regions for identifying wood of Dalbergia odorifera and D. tonkinensis (Leguminosae). J. Syst. Evol. 2022, 60, 73–84. [Google Scholar] [CrossRef]
- The Government of Vietnam. Vietnam Decree No. 32/2006/ND-CP dated 30/3/2006 of the Government of Vietnam on Management of Endangered and Rare Forest Fauna and Flora; The Government of Vietnam: Hanoi, Vietnam, 2006.
- The Government of Vietnam. MARD Decision No. 4961/QD-BNN-TCLN Dated 17/11/2014 of the Ministry of Agriculture and Rural Development of Vietnam on Promulgating a List of Main Tree Species for Planting Production Forests and a List of Main Tree Species for Afforestation According to Forest Ecological Regions; The Ministry of Agriculture and Rural Development of Vietnam: Hanoi, Vietnam, 2014; p. 11.
- The Government of Vietnam. Vietnam Decree No. 06/2019/ND-CP dated 22/01/2019 of the Government of Vietnam on Management of Endangered, Rare Forest Fauna and FLORA and Implementation of the Convention on International Trade in Endangered Wild Animal and Plant Species; The Government of Vietnam: Hanoi, Vietnam, 2019.
- Nhung, N.P.; Thu, P.Q.; Dell, B.; Chi, N.M. Cultivation of Dalbergia tonkinensis in north Vietnam. Vietnam J. For. Sci. 2019, 2, 64–77. [Google Scholar]
- Thu, P.Q.; Chi, N.M.; Quang, D.N.; Dell, B. Study on phenology and morphological characteristics of some provenances of Dalbergia tonkinensis Prain in Vietnam. Sci. Technol. J. Agric. Rural. Dev. 2014, 1, 247–253. [Google Scholar]
- Ha, V.T. Research techniques for planting Garcinia fagraeoides A. Chev, Cinamomum balanseae Lec and Dalbergia tonkinensis Prain for restoration of degraded natural forest in the Northwest region of Vietnam-result of research. In Forest Science and Technology period 2006–2010; Forest Science Insitute of Vietnam: Hanoi, Vietnam, 2011; pp. 181–186. [Google Scholar]
- Liu, X.J.; Xu, D.P.; Yang, Z.J.; Zhang, N.N.; Chen, M.H. Effects of temperature on seed germination of Dalbergia tonkinensis. For. Res. 2014, 27, 707–709. [Google Scholar]
- Nhung, N.P.; Thu, P.Q.; Chi, N.M.; Dell, B. Vegetative propagation of Dalbergia tonkinensis, a threatened, high-value tree species in South-east Asia. South. For. 2019, 81, 195–200. [Google Scholar] [CrossRef]
- Nhung, N.P.; Thu, P.Q.; Dell, B.; Chi, N.M. First report of canker disease in Dalbergia tonkinensis caused by Fusarium lateritium and Fusarium decemcellulare. Australas. Plant Pathol. 2018, 47, 317–323. [Google Scholar] [CrossRef]
- Chi, N.M.; Nhung, N.P.; Trang, T.T.; Thu, P.Q.; Hinh, T.X.; Nam, N.V.; Quang, D.N.; Dell, B. First report of wilt disease in Dalbergia tonkinensis caused by Ceratocystis manginecans. Australas. Plant Pathol. 2019, 48, 439–445. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.; Xue, S.; Cui, Z.; Xu, D.; Yang, Z. Effects of weeding and fertilization on soil biology and biochemical processes and tree growth in a mixed stand of Dalbergia odorifera and Santalum album. J. For. Res. 2021, 32, 2633–2644. [Google Scholar] [CrossRef]
- Yao, X.; Lan, Y.; Liao, L.; Huang, Y.; Yu, S.; Ye, S.; Yang, M. Effects of nitrogen supply rate on photosynthesis, nitrogen uptake and growth of seedlings in a Eucalyptus/Dalbergia odorifera intercropping system. Plant Biol. 2022, 24, 192–204. [Google Scholar] [CrossRef]
- Cui, Z.; Li, X.; Xu, D.; Yang, Z.; Zhang, N.; Liu, X.; Hong, Z. Physiological changes during heartwood formation induced by plant growth regulators in Dalbergia odorifera (Leguminosae). IAWA J. 2021, 42, 217–234. [Google Scholar] [CrossRef]
- Williams, E.R.; Matheson, A.C.; Harwood, C.E. Experimental Design and Analysis for Tree Improvement; CSIRO Publishing: Melbourne, Australia, 2002; p. 174. [Google Scholar]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Welham, S.J.; Thompson, R. ASReml User Guide Release 1.0. VSN international Ltd.: Hemel Hempstead, UK, 2002; 287p. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Tran, M.D.; Le, T.H. Investigatory result of Dalbergia tonkinensis Prain and propagation status of this tree species in Thua Thien Hue province. Hue Univ. J. Sci. 2012, 75, 19–28. [Google Scholar]
- Lu, S.; Liang, C. Early growth performance of Dalbergia odorifera T. Chen introduced in Zhenhai forest farm. Guangdong For. Sci. Technol. 2009, 4. [Google Scholar]
- Edrisi, S.A.; Tripathi, V.; Abhilash, P.C. Performance analysis and soil quality indexing for Dalbergia sissoo Roxb. grown in marginal and degraded land of Eastern Uttar Pradesh, India. Land 2019, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Hai, P.H.; Duong, L.A.; Toan, N.Q.; Ha, T.T.T. Genetic variation in growth, stem straightness, pilodyn and dynamic modulus of elasticity in second-generation progeny tests of Acacia mangium at three sites in Vietnam. New For. 2015, 46, 577–591. [Google Scholar] [CrossRef]
- Libby, W.J.; Rauter, R.M. Advantages of clonal forestry. For. Chron. 1984, 60, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Bhatt, A.; Ravichandran, S.; Kumar, V.; Dobhal, S. Genotype × environmental interactions for analyzing adaptability and stability in different clones of Dalbergia sissoo Roxb. J. For. Res. 2012, 23, 65–74. [Google Scholar] [CrossRef]
- Liao, H.; Deng, J.; Shi, L.; Zhou, C. Superior plant and clone selection and evaluation of introduced Dalbergia sissoo provenances. For. Res. 2017, 30, 916–920. [Google Scholar]
- Sharma, A.; Bakshi, M. Growth and heritability estimates among clones of Dalbergia sissoo Roxb. in a clonal seed orchard. For. Stud. China 2011, 13, 211–217. [Google Scholar] [CrossRef]
- Hai, P.H.; Jannson, G.; Harwood, C.E.; Hannrup, B.; Thinh, H.H.; Pinyopusarerk, K. Genetic variation in wood basic density and knot index and their relationship with growth traits for Acacia auriculiformis in northern Vietnam. N. Z. J. For. Sci. 2008, 38, 176–193. [Google Scholar]
- Kien, N.D.; Jansson, G.; Harwood, C.; Thinh, H.H. Genetic control of growth and form in Eucalyptus urophylla in Northern Vietnam. J. Trop. For. Sci. 2009, 21, 50–65. [Google Scholar]
- Liu, F.M.; Zhang, N.N.; Liu, X.J.; Yang, Z.J.; Jia, H.Y.; Xu, D.P. Genetic diversity and population structure analysis of Dalbergia odorifera germplasm and development of a core collection using microsatellite markers. Genes 2019, 10, 281. [Google Scholar] [CrossRef]
Figure 1.
Map of Vietnam with the provinces where Dalbergia tonkinensis mother trees were located and seeds were collected.
Figure 1.
Map of Vietnam with the provinces where Dalbergia tonkinensis mother trees were located and seeds were collected.

Figure 2.
Examples of Dalbergia tonkinensis mother trees used for seed collection: (a) in the grounds of a Buddhist temple in Hanoi (Dbh = 70.2 cm, Ht = 25 m), (b) on the side of a roadway in Hung Yen (Dbh = 58.6 cm, Ht = 18 m).
Figure 2.
Examples of Dalbergia tonkinensis mother trees used for seed collection: (a) in the grounds of a Buddhist temple in Hanoi (Dbh = 70.2 cm, Ht = 25 m), (b) on the side of a roadway in Hung Yen (Dbh = 58.6 cm, Ht = 18 m).

Figure 3.
Three-year-old D. tonkinensis planted in Tan Son: (a) superior family from Hung Yen; (b) superior family from Bac Ninh. Note the smaller trial trees surrounding the emergent trees.
Figure 3.
Three-year-old D. tonkinensis planted in Tan Son: (a) superior family from Hung Yen; (b) superior family from Bac Ninh. Note the smaller trial trees surrounding the emergent trees.

Figure 4.
Three-year-old D. tonkinensis planted in Doan Hung: (a) superior family from Vinh Phuc; (b) superior family from Hanoi.
Figure 4.
Three-year-old D. tonkinensis planted in Doan Hung: (a) superior family from Vinh Phuc; (b) superior family from Hanoi.

Figure 5.
Genetic × Environmental correlation on the growth of 60 D. tonkinensis families between the trials at Tan Son and Doan Hung.
Figure 5.
Genetic × Environmental correlation on the growth of 60 D. tonkinensis families between the trials at Tan Son and Doan Hung.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Average height, diameter, number of stems, survival, and disease index of Dalbergia tonkinensis provenances at 3 years in Tan Son. Different lowercase letters in a column indicate a significant difference according to DMRT at p < 0.05.
Table 1.
Average height, diameter, number of stems, survival, and disease index of Dalbergia tonkinensis provenances at 3 years in Tan Son. Different lowercase letters in a column indicate a significant difference according to DMRT at p < 0.05.
| Provenance | No of Families | Height (m) | Diameter (cm) | No of Stems | Survival (%) | Disease Index |
|---|---|---|---|---|---|---|
| Bac Ninh | 8 | 2.98 ± 0.78 a | 1.80 ± 0.66 ab | 1.41 ± 0.08 | 91.8 b | 0.82 ± 0.02 |
| Hanoi | 19 | 2.86 ± 0.79 ab | 1.94 ± 0.67 a | 1.41 ± 0.10 | 94.1 ab | 0.96 ± 0.03 |
| Hung Yen | 7 | 2.94 ± 0.65 a | 1.59 ± 0.49 b | 1.51 ± 0.11 | 96.4 a | 1.07 ± 0.04 |
| Gia Lai | 7 | 2.84 ± 0.74 ab | 1.85 ± 0.61 a | 1.48 ± 0.10 | 94.6 ab | 0.99 ± 0.02 |
| Lang Son | 9 | 2.20 ± 0.54 c | 1.33 ± 0.36 c | 1.30 ± 0.07 | 94.4 ab | 1.07 ± 0.04 |
| Nghe An | 7 | 2.69 ± 0.56 b | 1.62 ± 0.43 b | 1.42 ± 0.11 | 92.0 b | 1.10 ± 0.05 |
| Vinh Phuc | 12 | 2.55 ± 0.69 b | 1.60 ± 0.54 b | 1.43 ± 0.11 | 84.4 c | 0.96 ± 0.03 |
| Mean | 2.72 | 1.71 | 1.42 | 92.37 | 0.99 | |
| F value | <0.001 | <0.001 | 0.146 | <0.001 | 0.210 |
Table 2.
Average height, diameter, number of stems, survival, and disease index of Dalbergia tonkinensis provenances at 3 years in Doan Hung. Different lowercase letters in a column indicate a significant difference according to DMRT at p < 0.05.
Table 2.
Average height, diameter, number of stems, survival, and disease index of Dalbergia tonkinensis provenances at 3 years in Doan Hung. Different lowercase letters in a column indicate a significant difference according to DMRT at p < 0.05.
| Provenance | No of Families | Height (m) | Diameter (cm) | No of Stems | Survival (%) | Disease Index |
|---|---|---|---|---|---|---|
| Bac Ninh | 7 | 3.72 ± 0.76 a | 3.08 ± 0.62 b | 1.43 ± 0.10 | 92.4 c | 0.70 ± 0.01 |
| Hanoi | 10 | 3.81 ± 0.87 a | 3.34 ± 0.63 a | 1.41 ± 0.09 | 97.2 a | 0.86 ± 0.01 |
| Hung Yen | 7 | 3.64 ± 0.66 a | 2.83 ± 0.43 c | 1.51 ± 0.12 | 96.9 a | 0.94 ± 0.02 |
| Gia Lai | 7 | 3.55 ± 0.75 ab | 3.05 ± 0.52 b | 1.49 ± 0.11 | 94.6 b | 0.86 ± 0.03 |
| Lang Son | 9 | 2.93 ± 0.64 c | 2.59 ± 0.31 d | 1.31 ± 0.06 | 95.1 b | 0.93 ± 0.03 |
| Nghe an | 7 | 3.37 ± 0.62 b | 2.84 ± 0.37 c | 1.41 ± 0.09 | 93.3 c | 1.05 ± 0.04 |
| Vinh Phuc | 12 | 3.76 ± 0.87 a | 3.17 ± 0.57 ab | 1.46 ± 0.11 | 95.1 b | 0.98 ± 0.03 |
| Mean | 3.55 | 3.01 | 1.43 | 95.16 | 0.91 | |
| F value | <0.001 | <0.001 | 0.115 | <0.001 | 0.056 |
Table 3.
Trial means, individual heritability, and coefficients of additive genetic variation for the studied traits of D. tonkinensis provenances.
Table 3.
Trial means, individual heritability, and coefficients of additive genetic variation for the studied traits of D. tonkinensis provenances.
| Trait | Unit | Mean | ĥ2 | CVA (%) | Ry (%) |
|---|---|---|---|---|---|
| a. Trial in Tan Son | |||||
| Height | m | 2.72 | 0.39 ± 0.08 | 36 | 40.0 |
| Diameter | cm | 1.71 | 0.49 ± 0.09 | 58 | 71.2 |
| Disease index | score | 0.99 | 0.03 ± 0.03 | ||
| Number of stems | stem | 1.42 | 0.02 ± 0.03 | ||
| b. Trial in Doan Hung | |||||
| Height | m | 3.55 | 0.33 ± 0.08 | 25 | 25.7 |
| Diameter | cm | 3.01 | 0.48 ± 0.09 | 30 | 36.5 |
| Disease index | score | 0.91 | 0.02 ± 0.01 | ||
| Number of stems | stem | 1.43 | 0.03 ± 0.02 | ||
Table 4.
Effect of location of mother trees on growth of Dalbergia tonkinensis at 3 years in field trials in Tan Son and Doan Hung. Different lowercase letters in a column indicate a significant difference according to DMRT at p < 0.05.
Table 4.
Effect of location of mother trees on growth of Dalbergia tonkinensis at 3 years in field trials in Tan Son and Doan Hung. Different lowercase letters in a column indicate a significant difference according to DMRT at p < 0.05.
| Location of Mother Trees | Number of Families | Tan Son | Doan Hung | ||
|---|---|---|---|---|---|
| Height (m) | Diameter (cm) | Height (m) | Diameter (cm) | ||
| Natural forest | 7 | 3.17 ± 0.68 a | 2.03 ± 0.41 a | 3.92 ± 0.74 a | 3.27 ± 0.55 |
| City park | 8 | 2.62 ± 0.59 bc | 1.57 ± 0.38 c | 3.85 ± 0.71 ab | 3.28 ± 0.48 |
| Temple garden | 26 | 2.94 ± 0.63 b | 1.83 ± 0.42 b | 3.69 ± 0.65 ab | 3.07 ± 0.53 |
| Streetscape | 16 | 2.72 ± 0.71 bc | 1.80 ± 0.54 b | 3.53 ± 0.57 ab | 3.05 ± 0.46 |
| Office courtyard | 12 | 2.32 ± 0.60 c | 1.41 ± 0.36 c | 3.22 ± 0.59 b | 2.77 ± 0.42 |
| Mean | 2.72 | 1.71 | 3.55 | 3.01 | |
| F value | 0.006 | 0.040 | 0.029 | 0.069 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dell, B.; Thu, P.Q.; Nghia, N.H.; Hai, P.H.; Nhung, N.P.; Chi, N.M. Early Field Performance and Genetic Variation of Dalbergia tonkinensis, a Valuable Rosewood in Vietnam. Forests 2022, 13, 1977. https://doi.org/10.3390/f13121977
AMA Style
Dell B, Thu PQ, Nghia NH, Hai PH, Nhung NP, Chi NM. Early Field Performance and Genetic Variation of Dalbergia tonkinensis, a Valuable Rosewood in Vietnam. Forests. 2022; 13(12):1977. https://doi.org/10.3390/f13121977
Chicago/Turabian StyleDell, Bernard, Pham Quang Thu, Nguyen Hoang Nghia, Phi Hong Hai, Nong Phuong Nhung, and Nguyen Minh Chi. 2022. "Early Field Performance and Genetic Variation of Dalbergia tonkinensis, a Valuable Rosewood in Vietnam" Forests 13, no. 12: 1977. https://doi.org/10.3390/f13121977
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.