
High Variability of Fungal Communities Associated with the Functional Tissues and Rhizosphere Soil of Picea abies in the Southern Baltics

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Sites and Sampling
2.2. DNA Isolation, Amplification, and Sequencing
2.3. Bioinformatics
2.4. Statistical Analyses
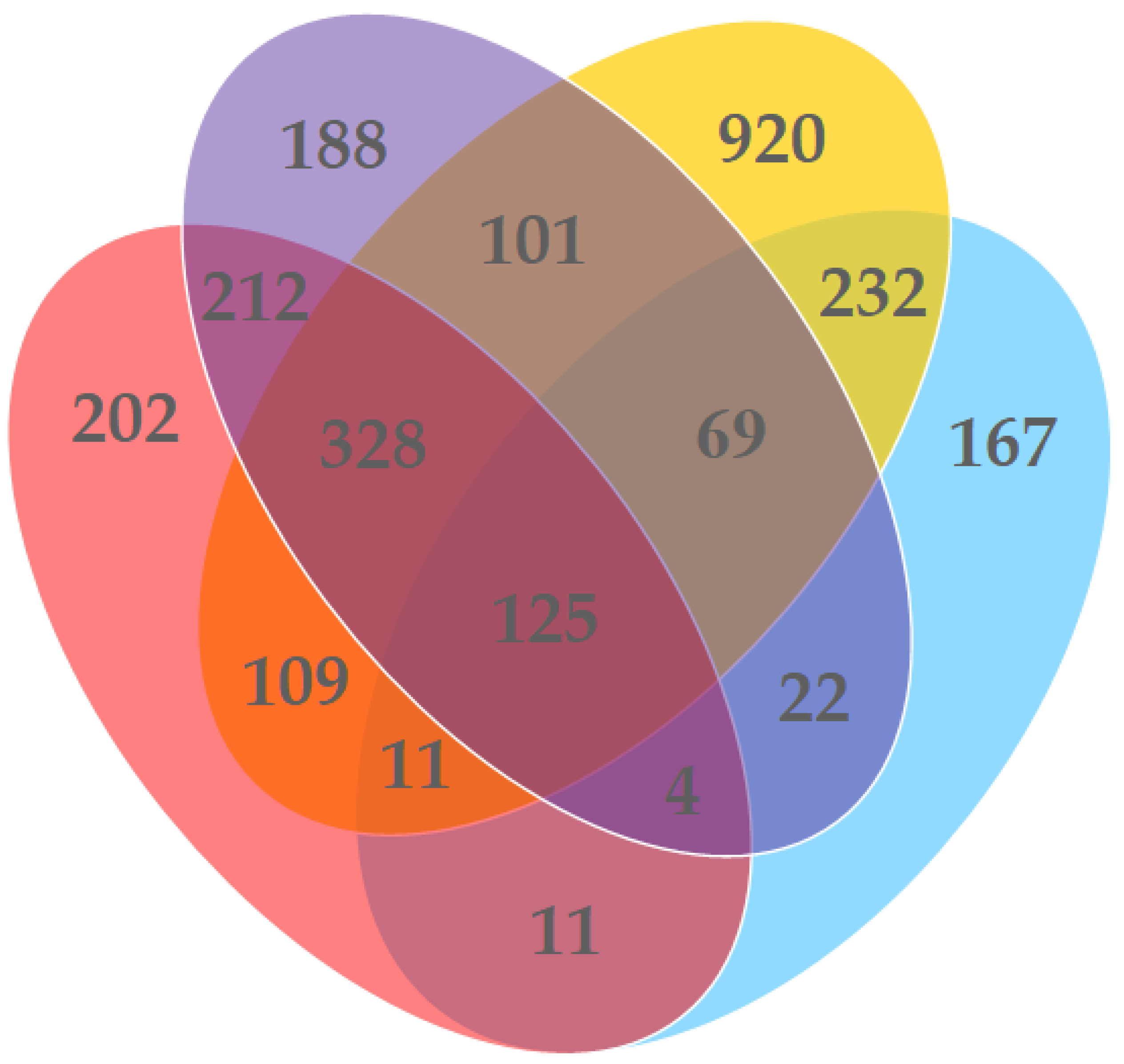
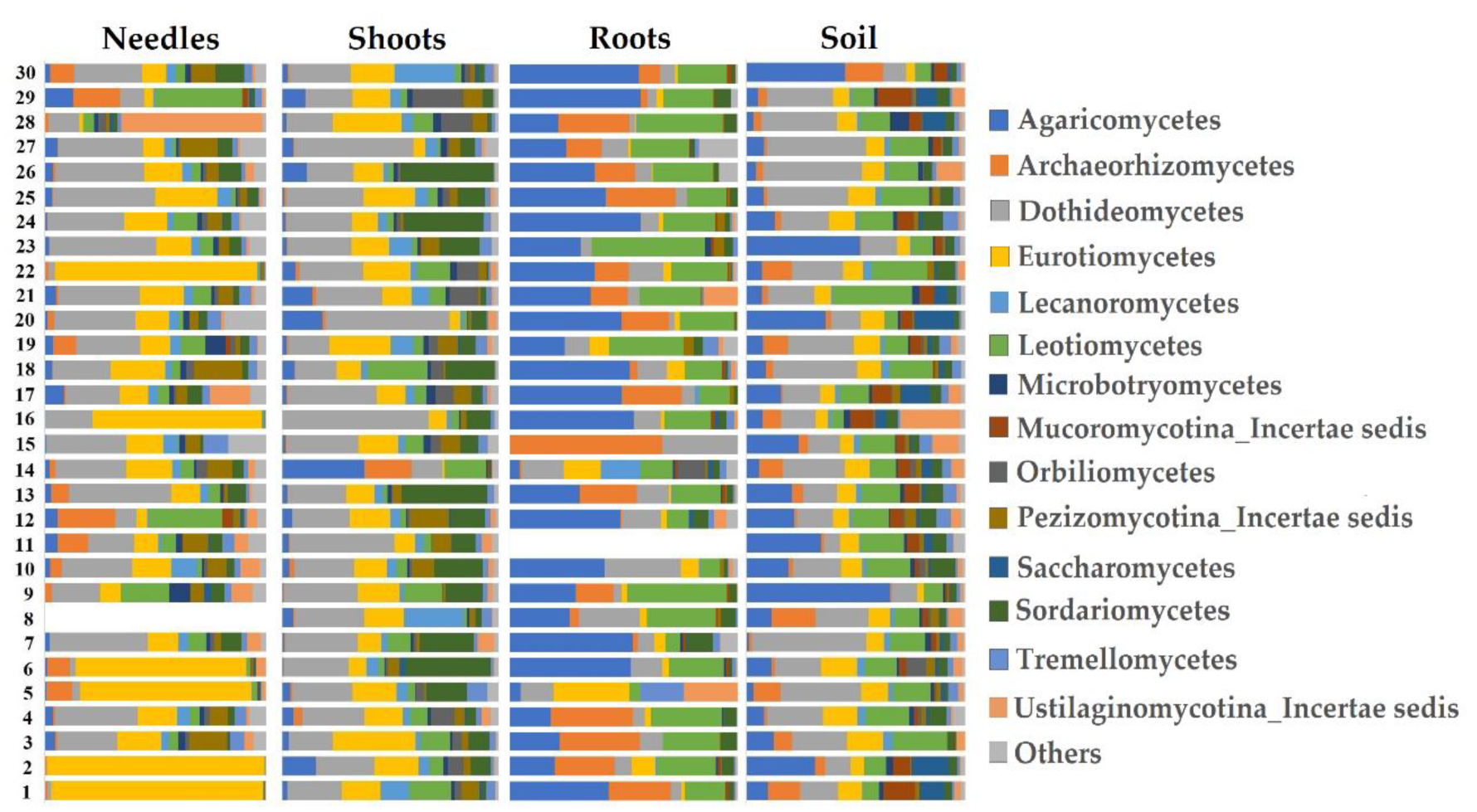
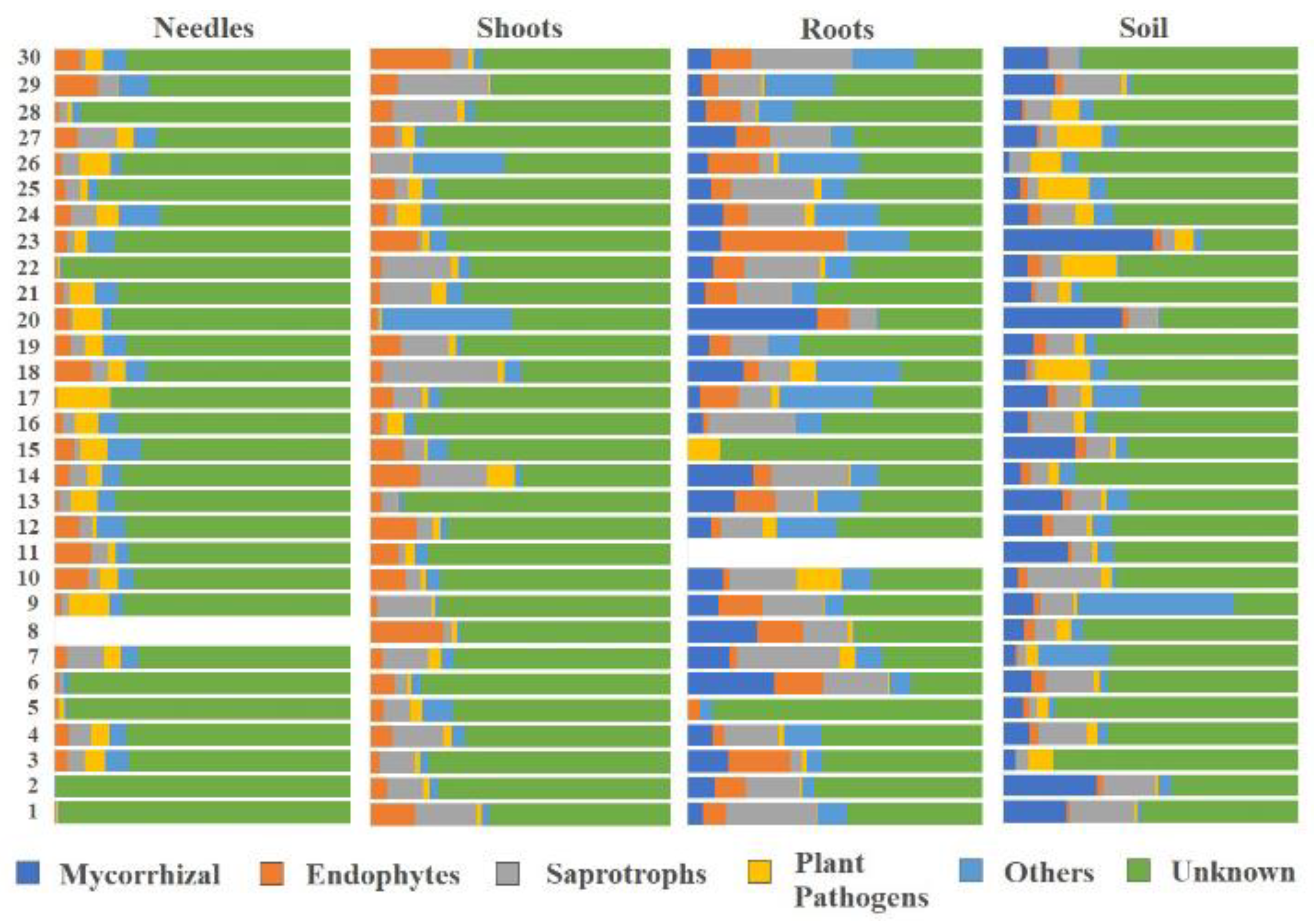
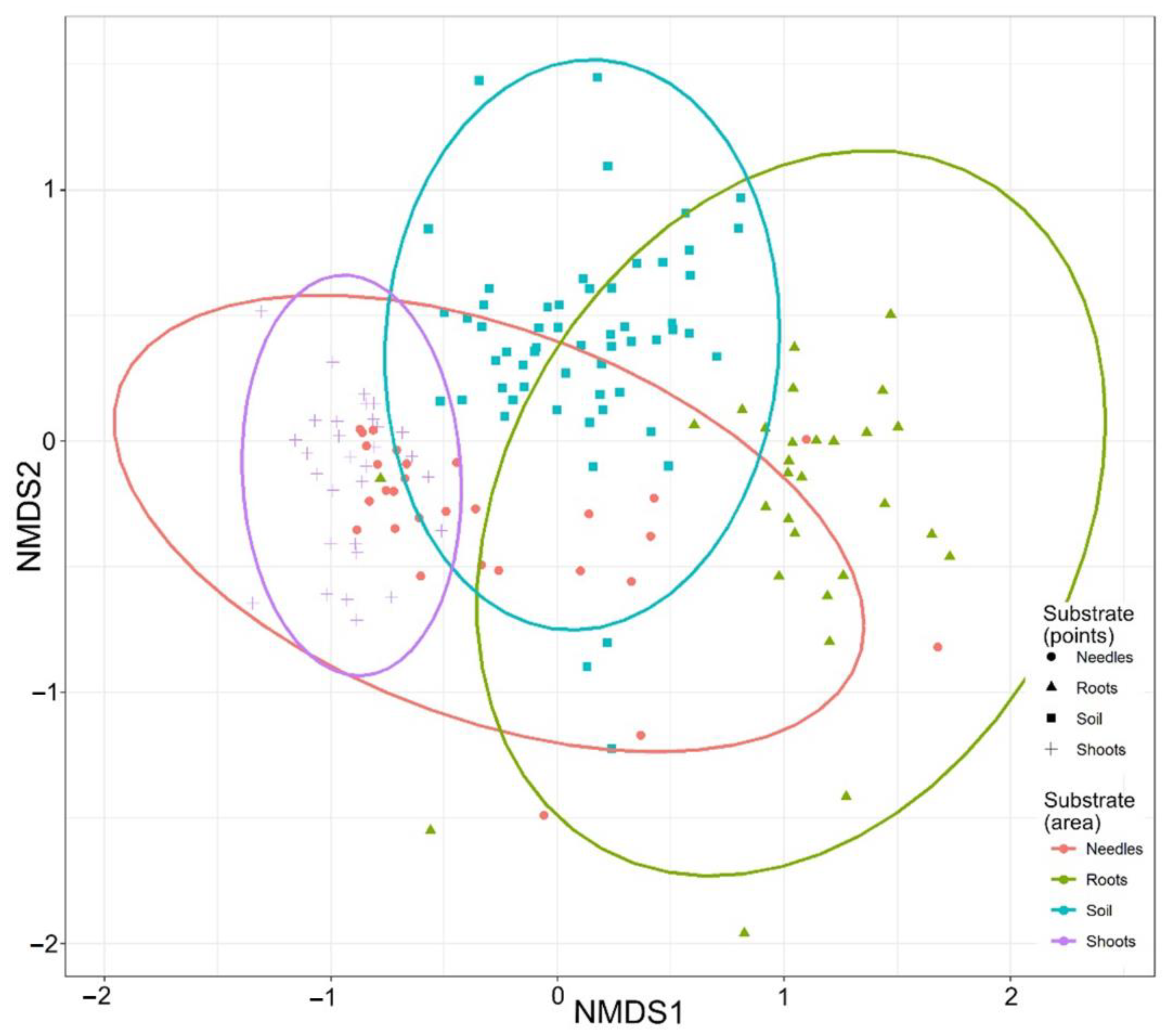
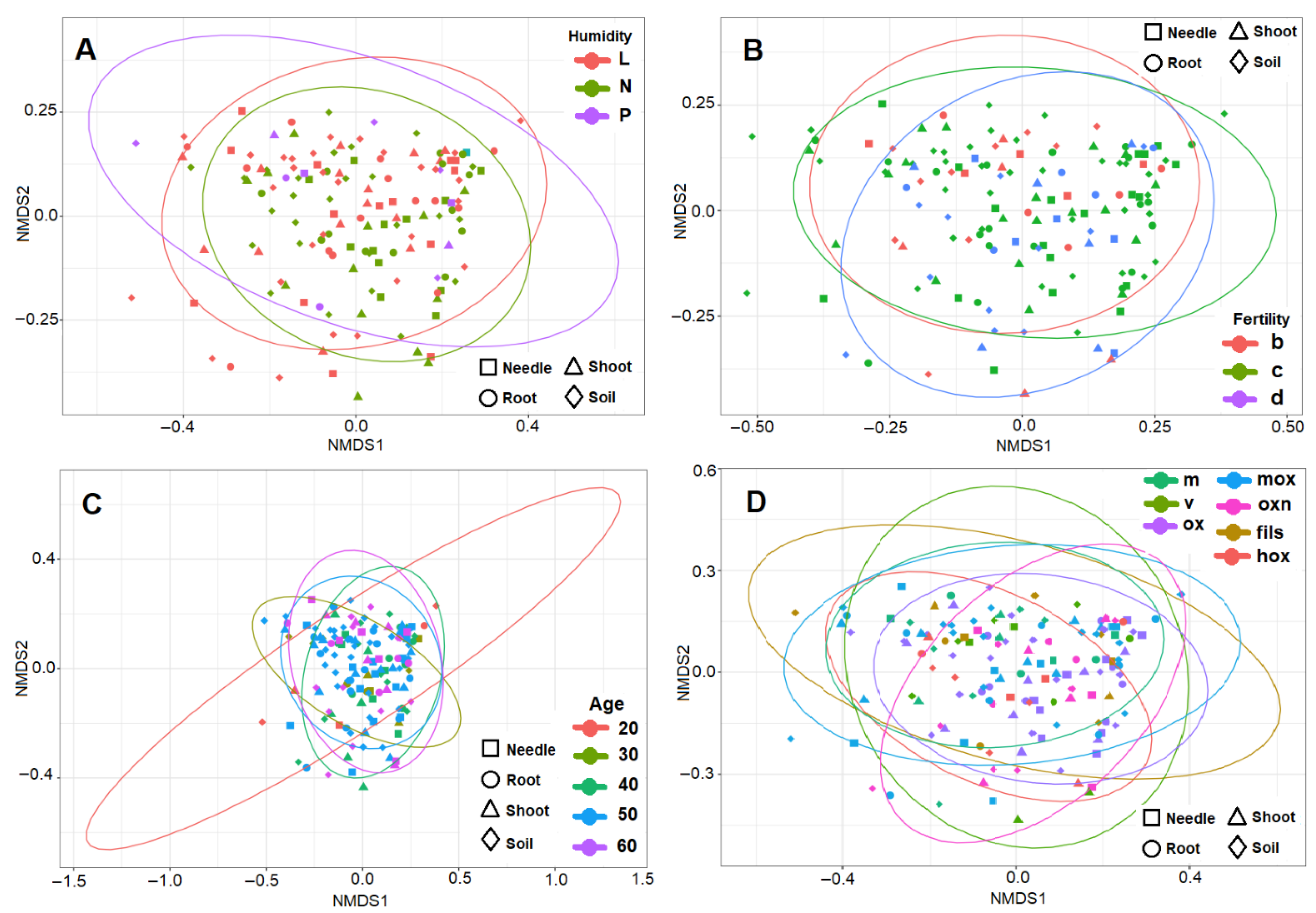
3. Results
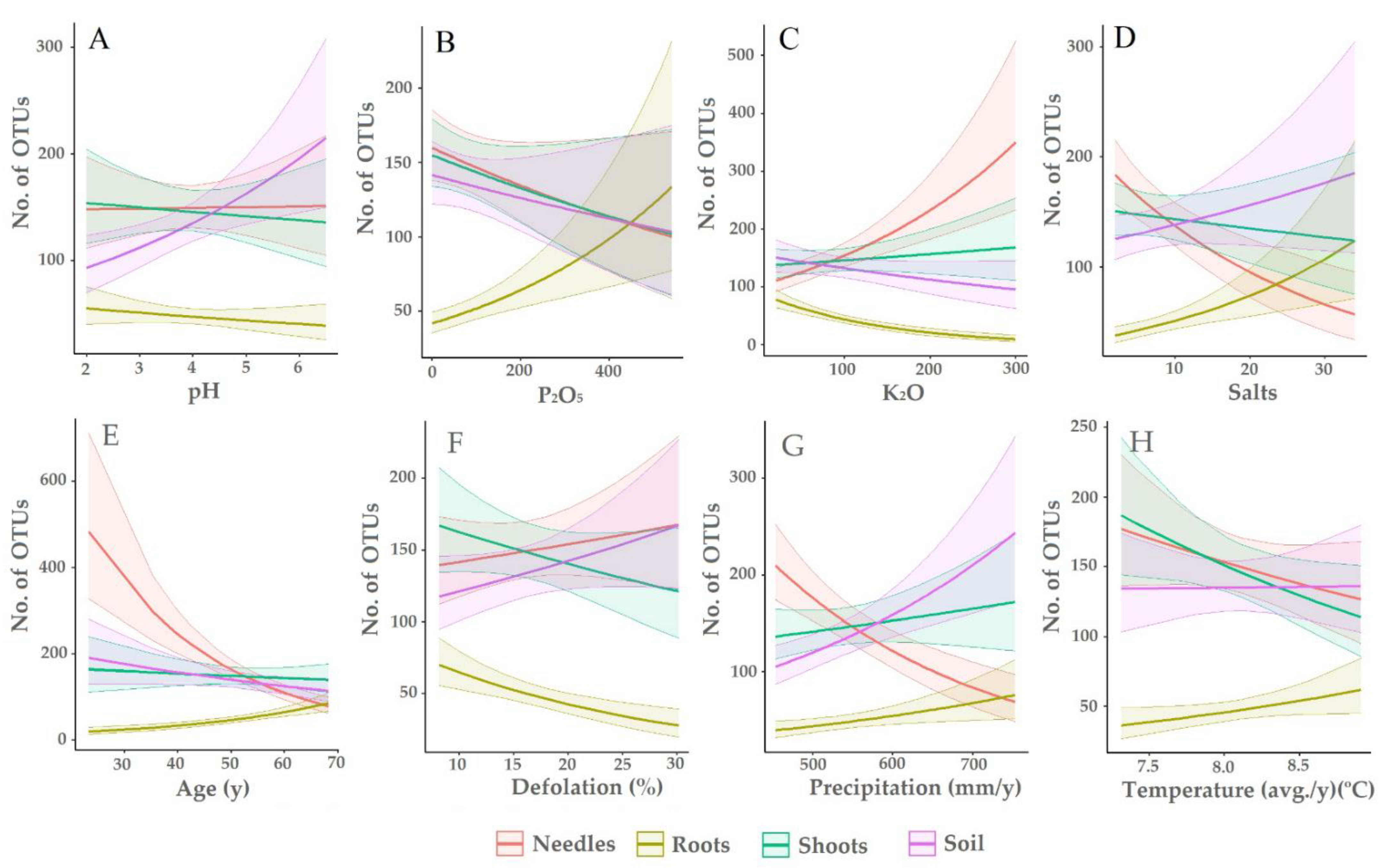
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schlyter, P.; Stjernquist, I.; Barring, L.; Jonsson, A.M.; Nilsson, C. Assessment of the impacts of climate change and weather extremes on boreal forests in northern Europe, focusing on Norway spruce. Clim. Res. 2006, 31, 75–84. Available online: http://www.jstor.org/stable/24869264 (accessed on 29 January 2022). [CrossRef]
- Seppä, H.; Alenius, T.; Bradshaw, R.H.; Giesecke, T.; Heikkilä, M.; Muukkonen, P. Invasion of Norway spruce (Picea abies) and the rise of the boreal ecosystem in Fennoscandia. J. Ecol. 2009, 97, 629–640. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea abies in Europe: Distribution, Habitat, Usage and Threats; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant Mauri, A., Eds.; European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016; pp. 114–116. [Google Scholar]
- Horgan, T.; Keane, M.; McCarthy, R.; Lally, M.; Thompson, D.; O’Carroll, J. A Guide to Forest Tree Species Selection and Silviculture in Ireland; National Council for Forest Research and Development (COFORD): Dublin, Ireland, 2003. [Google Scholar]
- Farjon, A. Handbook of the World’s Conifers: Revised and Updated Edition; Brill: Leiden, The Netherlands, 2017; Volume 1, pp. 21–24. [Google Scholar]
- Bradshaw, R.H.W.; Holmqvist, B.H.; Cowling, S.A.; Sykes, M.T. The effects of climate change on the distribution and management of Picea abies in southern Scandinavia. Can. J. For. Res. 2000, 30, 1992–1998. [Google Scholar] [CrossRef]
- Eckstein, D.; Krause, C. Dendroecologlcal Studies on Spruce Trefs to Monitor Environmental Changes Around Hamburg. IAWA J. 1989, 10, 175–182. [Google Scholar] [CrossRef]
- Lovelius, N.V. Dendroindication of Natural Processes and Anthropogenic Influenses; World and Family: St.-Peterburg, Russia, 1997; 320p. [Google Scholar]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Normand, S.; Pearman, P.B.; Psomas, A. Chapter III: Future ranges in European tree species. In Adapting to Climate Change in European Forests–Results of the MOTIVE Project; Pensoft Publishers: Sofia, Bulgaria, 2013; 108p. [Google Scholar]
- Jandl, R. Climate-induced challenges of Norway spruce in Northern Austria. Trees For. People 2020, 1, 100008. [Google Scholar] [CrossRef]
- Vitasse, Y.; Hoch, G.; Randin, C.F.; Lenz, A.; Kollas, C.; Körner, C. Tree recruitment of European tree species at their current upper elevational limits in the Swiss Alps. J. Biogeogr. 2012, 39, 1439–1449. [Google Scholar] [CrossRef]
- Grigor’ev, A.A.; Moiseev, P.A.; Nagimov, Z.Y. Dynamics of the timberline in high mountain areas of the nether-polar Urals under the influence of current climate change. Russ. J. Ecol. 2013, 44, 312–323. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Woodward, S.; Bodles, W.J.; Oszako, T. Occurence of phytophthora species in European forests. Folia For. Pol. Ser. A For. 2005, 47, 5–12. [Google Scholar]
- Sherwood, P.; Villari, C.; Capretti, P.; Bonello, P. Mechanisms of induced susceptibility to Diplodia tip blight in drought-stressed Austrian pine. Tree Physiol. 2015, 35, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barklund, P.; Rowe, J. Gremmeniella abietina (Scleroderris lagerbergii), a primary parasite in a Norway spruce dieback. Eur. J. For. Pathol. 1981, 11, 97–108. [Google Scholar] [CrossRef]
- Garrett, K.A.; Dendy, S.P.; Frank, E.E.; Rouse, M.N.; Travers, S.E. Climate change effects on plant disease: Genomes to ecosystems. Annu. Rev. Phytopathol. 2006, 44, 489–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Henk, D.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Luchi, N.; Capretti, P.; Feducci, M.; Vannini, A.; Ceccarelli, B.; Vettraino, A.M. Latent infection of Biscogniauxia nummularia in Fagus sylvatica: A possible bioindicator of beech health conditions. iForest Biogeosci. For. 2015, 9, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Carroll, G. Fungal endophytes in stems and leaves: From latent pathogen to mutualistic symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- LeBlanc, N.; Kinkel, L.L.; Kistler, H.C. Soil fungal communities respond to grassland plant community richness and soil edaphics. Microb. Ecol. 2015, 70, 188–195. [Google Scholar] [CrossRef]
- Peay, K.G.; Kennedy, P.G.; Talbot, J.M. Dimensions of biodiversity in the Earth mycobiome. Nat. Rev. Microbiol. 2016, 14, 434–447. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, X.; Tian, L.; Ma, L.; Luo, S.; Zhang, J.; Li, X.; Tian, C. Fungal communities in ancient peatlands developed from different periods in the Sanjiang Plain, China. PLoS ONE 2017, 12, e0187575. [Google Scholar] [CrossRef] [Green Version]
- Von Weissenberg, K. Effect of temperature on dormancy, activation and long-term storage of teliospores of Melampsora pinitorqua. Folia For. 1980, 422, 32–36. [Google Scholar]
- Desprez-Loustau, M.L.; Capron, G.; Dupuis, F. Relating germination dynamics of Melampsora pinitorqua teliospores to temperature and rainfall during overwintering. Eur. J. For. Pathol. 1998, 28, 335–347. [Google Scholar] [CrossRef]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Drenkhan, R.; Pritsch, K.; Buegger, F.; Padari, A. Regional scale in depth analysis of soil fungal diversity reveals strong pH and plant species effects in Northern Europe. Front. Microb. 2020, 11, 1953. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, A.; Johri, B.N. Climate Change is Real: Fungal Perspective. Kavaka 2019, 53, 12–21. [Google Scholar] [CrossRef]
- Huse, K.J. The distribution of fungi in sound-looking stems of Picea abies in Norway. Eur. J. Plant Pathol. 1981, 11, 1–6. [Google Scholar] [CrossRef]
- Müller, M.M.; Hallaksela, A.M. Diversity of Norway spruce needle endophytes in various mixed and pure Norway spruce stands. Mycol. Res. 1998, 102, 1183–1189. [Google Scholar] [CrossRef]
- Müller, M.M.; Hallaksela, A.M. Fungal diversity in Norway spruce: A case study. Mycol. Res. 2000, 104, 1139–1145. [Google Scholar] [CrossRef]
- Terhonen, E.; Blumenstein, K.; Kovalchuk, A.; Asiegbu, F.O. Forest tree microbiomes and associated fungal endophytes: Functional roles and impact on forest health. Forests 2019, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Petrini, O. Fungal endophytes of tree leaves. In Microbial Ecology of Leaves; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Rosling, A. Responses of Ectomycorrhizal Fungi to Mineral Substrates. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2003. [Google Scholar]
- Baldrian, P.; Kolařík, M.; Štursová, M.; Kopecký, J.; Valášková, V.; Větrovský, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, Č.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: London, UK, 1997. [Google Scholar]
- Menkis, A.; Marčiulynas, A.; Gedminas, A.; Lynikienė, J.; Povilaitienė, A. High-throughput sequencing reveals drastic changes in fungal communities in the phyllosphere of Norway spruce (Picea abies) following invasion of the spruce bud scale (Physokermes piceae). Microb. Ecol. 2015, 70, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Rajala, T.; Velmala, S.M.; Tuomivirta, T.; Haapanen, M.; Müller, M.; Pennanen, T. Endophyte communities vary in the needles of Norway spruce clones. Fungal Biol. 2013, 117, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Oger, P.; Tisserand, E.; Cébron, A.; Turpault, M.-P.; Buée, M.; de Boer, W.; Leveau, J.H.J.; Frey-Klett, P. Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities. Sci. Rep. 2016, 6, 27756. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.E.; Nieselt, K.; Garnica, S. Belowground fungal community diversity and composition associated with Norway spruce along an altitudinal gradient. PLoS ONE 2018, 13, e0208493. [Google Scholar] [CrossRef]
- Seidling, W.; Hansen, K.; Strich, S.; Lorenz, M. Part I: Objectives, Strategy and Implementation of ICP Forests. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-Ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2017; pp. 1–12, + Annex; Available online: http://www.icp-forests.net/page/icp-forests-manual (accessed on 23 February 2022).
- Ozolinčius, R.; Stakėnas, V. Tree condition assessment. In Monitoring of Ecosystem in Lithuania; Ozolinčius, R., Armolaitis, K., Augustaitis, A., Kairiūkštis, L., Stakėnas, V., Vaičys, M., Eds.; Lututė: Kaunas, Lithuania, 1999; p. 308. (In Lithuanian) [Google Scholar]
- Vaičys, M. Miško dirvožemių klasifikacija. In Lietuvos Dirvožemiai; Mokslas: Vilnius, Lithuania, 2001; pp. 1040–1043. (In Lithuanian) [Google Scholar]
- Karazija, S. Lietuvos Miško Tipai; Mokslas: Vilnius, Lithuania, 1988; pp. 46–48. [Google Scholar]
- Marčiulynienė, D.; Marčiulynas, A.; Lynikienė, J.; Vaičiukynė, M.; Gedminas, A.; Menkis, A. DNA-metabarcoding of belowground fungal communities in bare-root forest nurseries: Focus on different tree species. Microorganisms 2021, 9, 150. [Google Scholar] [CrossRef] [PubMed]
- Ihrmark, K.; Bodeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fung. Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; 192p. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. GglmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. Available online: https://journal.r-project.org/archive/2017/RJ-2017-066/index.html (accessed on 23 February 2022). [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Lüdecke, D. ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 23 February 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Community Ecology Package. R Package Version 2. 2013. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 24 February 2022).
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponge, J.F. The soil as an ecosystem. Biol. Fertil. Soils 2015, 51, 645–648. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Wagg, C. Soil microbial diversity and agro-ecosystem functioning. Plant Soil 2013, 363, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kivlin, S.N.; Smith, A.P.; Sulman, B.N.; Buscardo, E. Forest Rhizosphere Interactions: Cascading Consequences for Ecosystem-Level Carbon and Nutrient Cycling. Front. For. Glob. Chang. 2021, 4, 33. [Google Scholar] [CrossRef]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [Green Version]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Lagomarsino, A.; Grego, S.; Kandeler, E. Soil organic carbon distribution drives microbial activity and functional diversity in particle and aggregate-size fractions. Pedobiologia 2012, 55, 101–110. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J. Soil moisture as a factor affecting the microbiological and biochemical activity of soil. Plant Soil Environ. 2016, 62, 250–255. [Google Scholar] [CrossRef]
- Bååth, E.; Anderson, T.H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Bioch. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.; Lauber, C.; Lozupone, C.; Caporaso, G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Guo, D. Impacts of environmental factors on fine root lifespan. Front. Plant Sci. 2014, 5, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, D.L.; Griffin, K.P.; Danner, M. Better Crops with Plant Food; IPNI: Norcross, GA, USA, 1998; Volume LXXXII. [Google Scholar]
- Houlden, A.; Timms-Wilson, T.M.; Day, M.J.; Bailey, M.J. Influence of plant developmental stage on microbial community structure and activity in the rhizosphere of three field crops. FEMS Microbiol. Ecol. 2008, 65, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mougel, C.; Offre, P.; Ranjard, L.; Corberand, T.; Gamalero, E.; Robin, C.; Lemanceau, P. Dynamic of the genetic structure of bacterial and fungal communities at different developmental stages of Medicago truncatula Gaertn. cv. Jemalong line J5. New Phyt. 2006, 170, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Marschner, P.; Crowley, D.; Rengel, Z. Rhizosphere interactions between microorganisms and plants govern iron and phosphorus acquisition along the root axis—Model and research methods. Soil Biol. Bioch. 2011, 43, 883–894. [Google Scholar] [CrossRef]
- Castano, C.; Hernandez-Rodriguez, M.; Geml, J.; Eberhart, J.; Olaizola, J.; Oria-de-Rueda, J.A.; Martin-Pinto, P. Resistance of the soil fungal communities to medium-intensity fire prevention treatments in a Mediterranean scrubland. For. Ecol. Manag. 2020, 472, 118–217. [Google Scholar] [CrossRef]
- Brabcová, V.; Nováková, M.; Davidová, A.; Baldrian, P. Dead fungal mycelium in forest soil represents a decomposition hotspot and a habitat for a specific microbial community. New Phytol. 2016, 210, 1369–1381. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L.O.; Wallander, H. Production of external mycelium by ectomycorrhizal fungi in a Norway spruce forest was reduced in response to nitrogen fertilization. New Phytol. 2003, 158, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Cigan, P.W.; Karst, J.; Cahill, J.F.; Sywenky, A.N.; Pec, G.J.; Erbilgin, N. Influence of bark beetle outbreaks on nutrient cycling in native pine stands in western Canada. Plant Soil. 2015, 390, 29–47. [Google Scholar] [CrossRef]
- Przybył, K.; Karolewski, P.; Oleksyn, J.; Łabędzki, A.; Reich, P.B. Fungal diversity of Norway spruce litter: Effects of site conditions and premature leaf fall caused by bark beetle outbreak. Microb Ecol. 2008, 56, 332–340. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Ahlholm, J.; Helander, M.; Elamo, P.; Saloniemi, I.; Neuvonen, S.; Hanhimäki, S.; Saikkonen, K. Micro-fungi and invertebrate herbivores on birch trees: Fungal mediated plant–herbivore interactions or responses to host quality? Ecology 2002, 5, 648–655. [Google Scholar] [CrossRef]
- Chareprasert, S.; Piapukiew, J.; Thienhirun, S.; Whalley, A.J.; Sihanonth, P. Endophytic fungi of teak leaves Tectona grandis L. and rain tree leaves Samanea saman Merr. World J. Microbiol. Biotechnol. 2006, 22, 481–486. [Google Scholar] [CrossRef]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Schjønning, P.; Thomsen, I.K.; Petersen, S.O.; Kristensen, K.; Christensen, B.T. Relating soil microbial activity to water content and tillage-induced differences in soil structure. Geoderma 2011, 163, 256–264. [Google Scholar] [CrossRef]
- Singh, B.K.; Nunan, N.; Ridgway, K.P.; McNicol, J.; Young, J.P.W.; Daniell, T.J.; Prosser, J.I.; Millard, P. Relationship between assemblages of mycorrhizal fungi and bacteria on grass roots. Envir. Microb. 2008, 10, 534–541. [Google Scholar] [CrossRef]
- Guo, X.; Gong, J. Differential effects of abiotic factors and host plant traits on diversity and community composition of root-colonizing arbuscular mycorrhizal fungi in a salt-stressed ecosystem. Mycorrhiza 2014, 24, 79–94. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Saikkonen, K. Forest structure and fungal endophytes. Fungal Biol. Rev. 2007, 21, 67–74. [Google Scholar] [CrossRef]
- James, T.Y.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Phillips, R.; & Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phyt. 2003, 157, 475–492. [Google Scholar] [CrossRef] [PubMed]
- Netherway, T.; Bengtsson, J.; Krab, E.J.; Bahram, M. Biotic interactions with mycorrhizal systems as extended nutrient acquisition strategies shaping forest soil communities and functions. Basic Appl. Ecol. 2021, 50, 25–42. [Google Scholar] [CrossRef]
- Arias, M.M.D.; Batzer, J.C.; Harrington, T.C.; Wong, A.W.; Bost, S.C.; Cooley, D.R.; Ellis, M.A.; Hartman, J.R.; Rosenberger, D.A.; Sundin, G.W.; et al. Diversity and biogeography of sooty blotch and flyspeck fungi on apple in the eastern and midwestern United States. Phytopathology 2010, 100, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Zhu, M.; Zhang, R.; Yin, H.; Lei, Y.; Sun, G.; Gleason, M.L. Phialophora sessilis, a species causing flyspeck signs on bamboo in China. Mycotaxon 2010, 113, 405–413. [Google Scholar] [CrossRef]
- Nguyen, D.; Boberg, J.; Ihrmark, K.; Stenström, E.; Stenlid, J. Do foliar fungal communities of Norway spruce shift along a tree species diversity gradient in mature European forests? Fungal Ecol. 2016, 23, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Livsey, S.; Barklund, P. Lophodermium piceae and Rhizosphaera kalkhoffii in fallen needles of Norway spruce (Picea abies). Eur. J. For. Pathol. 1992, 22, 204–216. [Google Scholar] [CrossRef]
- Scattolin, L.; Montecchio, L. Lophodermium piceae and Rhizosphaera kalkhoffii in Norway spruce: Correlations with host age and climatic features. Phytopathol. Mediterr. 2009, 48, 226–239. [Google Scholar]
- Hormazabal, E.; Astudillo, L.; Schmeda-Hirschmann, G.; Rodríguez, J.; Theoduloz, C. Metabolites from Microsphaeropsis olivacea, an endophytic fungus of Pilgerodendron uviferum. Z. Naturforsch. C 2005, 60, 11–21. [Google Scholar] [CrossRef]
- Lazarević, J.; Menkis, A. Fungal diversity in the phyllosphere of Pinus heldreichii H. Christ-an endemic and high-altitude pine of the Mediterranean Region. Diversity 2020, 12, 172. [Google Scholar] [CrossRef]
- Razaghi, P.; Zafari, D. First report of Microsphaeropsis olivacea causing brown spine rot on Alhagi maurorum in Iran. Plant Pathol. 2016, 98, 685. [Google Scholar] [CrossRef]
- Crous, P.W.; Quaedvlieg, W.; Hansen, K.; Hawksworth, D.L.; Groenewald, J.Z. Phacidium and Ceuthospora (Phacidiaceae) are congeneric: Taxonomic and nomenclatural implications. Ima Fungus 2014, 5, 173–193. [Google Scholar] [CrossRef] [PubMed]
- Di Cosmo, F.; Nag Raj, T.R.; Kendrick, W.B. A Revision of the Phacidiaceae and Related Anamorphs [New Taxa]; Mycotaxon: New York, NY, USA, 1984; p. 234. [Google Scholar]
- Ribeiro, T.H.C.; Fernandes-Brum, C.N.; de Souza, C.R.; Dias, F.A.N.; Almeida-Junior, O.D.; Regina, M.D.A.; Chalfun-Junior, A. Transcriptome analyses suggest that changes in fungal endophyte lifestyle could be involved in grapevine bud necrosis. Sci. Rep. 2020, 10, 9514. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Cacciola, S.O.; Sanz-Ros, A.V.; Garbelotto, M.; Aguayo, J.; Solla, A.; Díez, J.J. Potential interactions between invasive Fusarium circinatum and other pine pathogens in Europe. Forests 2020, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Orr, R.; Nelson, P.N. Impacts of soil abiotic attributes on Fusarium wilt, focusing on bananas. Appl. Soil Ecol. 2018, 132, 20–33. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenström, E.; Stenlid, J.; Finlay, R. Fungi in decayed roots of conifer seedlings from forest nurseries, afforested clearcuts and abandoned farmland. Plant Pathol. 2006, 55, 117–129. [Google Scholar] [CrossRef]
- Striganavičiūtė, G.; Žiauka, J.; Sirgedaitė-Šėžienė, V.; Vaitiekūnaitė, D. Priming of Resistance-Related Phenolics: A Study of Plant-Associated Bacteria and Hymenoscyphus fraxineus. Microorganisms 2021, 9, 2504. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Mauch-Mani, B. Priming: Getting ready for battle. Mol. Plant-Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site No. | Geographical Position | Stand Characteristics | Soil Chemical Parameters | Meteorological Data | Stand Sanitary Condition | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | E | Forest a/Vegetation Type b | Tree Species Composition, % c | Age (y) | pH (KCl), mol/L | P2O5, mg/kg | K2O, mg/kg | Ca, mg/kg | Mg, mg/kg | Cl, mg/kg | Salts, ms/cm | Average Temp., °C | Precipitation, mm/year | Defoliation, % | Dechromation, % | Dry Branches, % | |
| 1 | 55°37′ | 25°11′ | Ncl/ox | 100S | 50 | 3.9 | 306 | 43 | 267 | 72 | 3.6 | 3.35 | 8.0 | 508.8 | 11.2 | 5.8 | 14.5 |
| 2 | 55°01′ | 24°12′ | Lcl/mox | 100S | 52 | 3.3 | 56 | 39 | 242 | 56 | 5.3 | 4.33 | 8.3 | 614.9 | 29.0 | 13.5 | 26.0 |
| 3 | 54°52′ | 23°26′ | Pcn/fils | 100S | 61 | 2.4 | 65 | 296 | 774 | 279 | 8.9 | 11.7 | 8.6 | 519.8 | 13.2 | 2.3 | 12.8 |
| 4 | 56°12′ | 21°27′ | Lbp/m | 90S, 10P | 59 | 3.0 | 32 | 170 | 583 | 136 | 5.3 | 7.64 | 8.9 | 491.7 | 15.8 | 4.1 | 13.5 |
| 5 | 56°00′ | 21°07′ | Lbl/m | 100S | 59 | 2.8 | 12 | 69 | 264 | 73 | 5.3 | 8.83 | 8.9 | 491.7 | 16.3 | 3.7 | 13.5 |
| 6 | 55°54′ | 25°13′ | Lcs/mox | 90S, 10A | 67 | 5.8 | 15 | 47 | 2256 | 490 | 3.6 | 6.49 | 7.7 | 514.6 | 18.0 | 4.8 | 15.8 |
| 7 | 55°44′ | 24°43′ | Lcl/mox | 90S, 10P | 57 | 3.1 | 8 | 21 | 154 | 28 | 1.8 | 2.36 | 8.1 | 479.8 | 24.8 | 7.0 | 20.7 |
| 8 | 56°03′ | 22°56′ | Lds/oxn | 100S | 48 | 5.1 | 12 | 38 | 1472 | 329 | 5.3 | 13.6 | 8.1 | 505.5 | 16.5 | 6.8 | 14.7 |
| 9 | 56°11′ | 22°23′ | Lcp/mox | 90S, 10P | 58 | 5.2 | 16 | 144 | 1770 | 270 | 5.3 | 4.87 | 8.0 | 480.4 | 18.5 | 4.0 | 17.5 |
| 10 | 54°52′ | 25°42′ | Ncl/ox | 100S | 35 | 3.8 | 169 | 40 | 176 | 48 | 3.6 | 2.01 | 8.0 | 755.4 | 18.3 | 6.2 | 14.3 |
| 11 | 55°53′ | 24°12′ | Ldf/oxn | 80S, 20F | 62 | 5.7 | 17 | 75 | 2968 | 630 | 5.3 | 12.4 | 8.1 | 505.5 | 16.7 | 8.0 | 17.0 |
| 12 | 56°03′ | 23°40′ | Lds/oxn | 90S, 10A | 42 | 4.4 | 11 | 131 | 2952 | 542 | 7.1 | 26.0 | 8.1 | 505.5 | 9.0 | 1.5 | 11.2 |
| 13 | 55°29′ | 24°12′ | Lds/oxn | 100S | 53 | 4.8 | 25 | 76 | 1294 | 197 | 5.3 | 4.7 | 8.1 | 479.8 | 12.7 | 3.7 | 9.3 |
| 14 | 54°35′ | 23°57′ | Ncl/ox | 100S | 47 | 3.9 | 27 | 21 | 101 | 32 | 7.1 | 2.55 | 8.6 | 595.2 | 8.8 | 3.3 | 13.8 |
| 15 | 55°27′ | 23°27′ | Nds/hox | 100S | 50 | 4.0 | 19 | 77 | 942 | 147 | 5.3 | 3.42 | 7.8 | 458 | 18.5 | 5.3 | 14.8 |
| 16 | 56°03′ | 25°42′ | Lcp/mox | 60S, 30PT, 10B | 60 | 3.7 | 36 | 63 | 286 | 73 | 3.6 | 3.14 | 7.7 | 514.6 | 18.3 | 7.2 | 15.3 |
| 17 | 55°28′ | 21°57′ | Lcp/mox | 60S, 20B, 10Q, 10A | 59 | 3.8 | 11 | 107 | 791 | 122 | 3.6 | 4.95 | 8.4 | 617.0 | 14.3 | 3.7 | 12.2 |
| 18 | 55°02′ | 22°41′ | Lcl/mox | 60S, 40P | 23 | 2.8 | 15 | 86 | 277 | 75 | 3.6 | 3.64 | 8.6 | 519.8 | 19.3 | 7.7 | 16.8 |
| 19 | 54°18′ | 25°40′ | Ncl/ox | 80S, 10Q, 10PT | 39 | 3.8 | 71 | 43 | 157 | 42 | 5.3 | 3.06 | 8.0 | 530.6 | 15.8 | 2.0 | 12.7 |
| 20 | 54°18′ | 25°40′ | Nds/hox | 100S | 53 | 5.0 | 27 | 48 | 2584 | 374 | 7.1 | 11.1 | 8.1 | 479.8 | 25.3 | 16.5 | 25.7 |
| 21 | 55°10′ | 25°42′ | Nbl/v | 90S, 10P | 40 | 4.1 | 78 | 36 | 238 | 52 | 5.3 | 3.96 | 7.4 | 669.2 | 18.7 | 8.0 | 13.8 |
| 22 | 55°19′ | 22°27′ | Lcp/mox | 90S, 10B | 55 | 3.3 | 118 | 300 | 2092 | 566 | 7.1 | 18.5 | 8.6 | 591.3 | 8.7 | 0.3 | 12.7 |
| 23 | 55°45′ | 21°41′ | Ncs/ox | 100S | 50 | 4.1 | 71 | 123 | 2043 | 161 | 3.6 | 5.0 | 8.0 | 598.7 | 14.7 | 3.5 | 15.0 |
| 24 | 55°44′ | 22°25′ | Ncl/ox | 100S | 40 | 3.5 | 37 | 25 | 212 | 48 | 3.6 | 2.07 | 8.0 | 598.7 | 16.7 | 6.0 | 16.7 |
| 25 | 54°44′ | 24°42′ | Ncl/ox | 100S | 58 | 3.8 | 12 | 70 | 416 | 136 | 3.6 | 4.65 | 7.3 | 614.9 | 22.0 | 7.8 | 18.3 |
| 26 | 54°56′ | 24°42′ | Lbl/m | 80S, 10B, 10P | 60 | 4.7 | 26 | 68 | 2362 | 308 | 5.3 | 14.0 | 8.0 | 535.8 | 13.7 | 2.5 | 10.7 |
| 27 | 55°10′ | 24°41′ | Ncl/ox | 60S, 20P, 20PT | 50 | 6.6 | 544 | 183 | 11,508 | 1184 | 3.6 | 8.43 | 8.0 | 506.9 | 21.3 | 5.7 | 17.2 |
| 28 | 55°18′ | 25°42′ | Pcn/fils | 90S, 10B | 52 | 3.0 | 132 | 237 | 2584 | 520 | 14.2 | 33 | 7.7 | 505.4 | 19.3 | 5.2 | 17.7 |
| 29 | 54°25′ | 24°57′ | Nbl/v | 100S | 68 | 3.4 | 23 | 31 | 161 | 42 | 3.6 | 2.73 | 8.0 | 593.2 | 16.2 | 2.7 | 12.8 |
| 30 | 54°25′ | 24°27′ | Ncl/ox | 70S, 20B, 10P | 48 | 3.7 | 169 | 53 | 512 | 117 | 3.6 | 5.0 | 8.0 | 540.9 | 14.2 | 3.3 | 11.2 |
| Site | No. of Sequences/Fungal OTUs | Shannon Diversity Index (H) | ||||||
|---|---|---|---|---|---|---|---|---|
| No. | Needles | Shoots | Roots | Soil | Needles | Shoots | Roots | Soil |
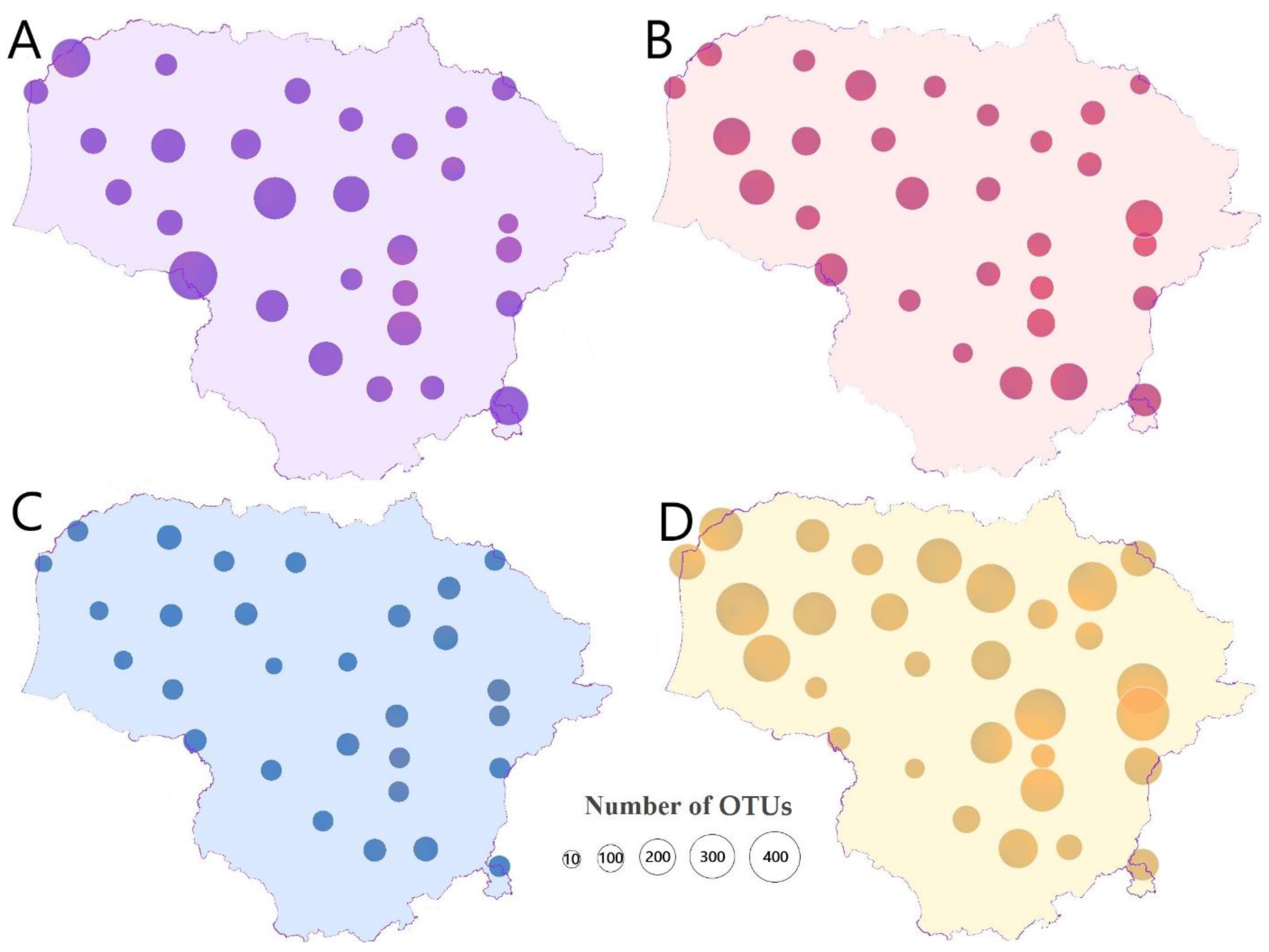
| 1 | 4222/65 | 636/132 | 3136/145 | 1792/160 | 0.34 | 3.95 | 2.77 | 3.52 |
| 2 | 2533/27 | 425/104 | 413/87 | 4586/295 | 0.14 | 4.01 | 3.27 | 4.38 |
| 3 | 1664/182 | 645/91 | 702/47 | 24/19 | 4.16 | 3.42 | 2.49 | 2.92 |
| 4 | 2605/241 | 562/136 | 139/31 | 1769/310 | 4.36 | 3.43 | 1.68 | 3.82 |
| 5 | 2215/94 | 406/72 | 21/7 | 2327/210 | 1.12 | 4.11 | 2.55 | 4.96 |
| 6 | 678/51 | 1088/120 | 706/79 | 2782/346 | 1.25 | 3.36 | 3.15 | 4.78 |
| 7 | 831/143 | 247/64 | 277/57 | 1079/171 | 4.28 | 3.66 | 3.03 | 3.85 |
| 8 | -/- | 1861/186 | 205/38 | 1104/185 | - | 3.81 | 2.92 | 4.13 |
| 9 | 38/22 | 594/73 | 1086/102 | 2212/192 | 2.89 | 2.87 | 3.22 | 3.00 |
| 10 | 722/132 | 645/117 | 128/39 | 1632/220 | 4.06 | 3.89 | 3.15 | 4.26 |
| 11 | 245/80 | 303/73 | -/- | 2796/338 | 3.67 | 3.44 | - | 4.89 |
| 12 | 1020/139 | 446/75 | 116/41 | 2275/313 | 3.52 | 3.57 | 3.16 | 5.01 |
| 13 | 2676/216 | 3314/145 | 1047/117 | 2011/292 | 3.78 | 3.18 | 3.07 | 4.67 |
| 14 | 2860/203 | 98/41 | 3136/149 | 1295/160 | 4.14 | 3.27 | 3.00 | 4.34 |
| 15 | 7090/300 | 1271/190 | 3/2 | 629/158 | 3.95 | 4.34 | 0.64 | 4.41 |
| 16 | 5675/84 | 109/36 | 60/22 | 2450/209 | 4.20 | 3.01 | 2.35 | 3.72 |
| 17 | 865/150 | 2173/216 | 53/18 | 2899/316 | 1.01 | 4.30 | 2.25 | 4.49 |
| 18 | 6501/316 | 11,512/201 | 273/59 | 119/61 | 4.18 | 3.25 | 3.12 | 3.62 |
| 19 | 2276/232 | 2269/189 | 46/26 | 850/184 | 4.29 | 4.09 | 3.08 | 4.56 |
| 20 | 1127/161 | 1312/100 | 1041/63 | 5301/269 | 4.06 | 2.53 | 2.41 | 3.82 |
| 21 | 1205/149 | 1171/146 | 178/24 | 8092/444 | 3.96 | 3.88 | 2.59 | 3.99 |
| 22 | 5757/102 | 1699/145 | 150/42 | 86/44 | 0.72 | 3.73 | 3.06 | 3.42 |
| 23 | 1447/155 | 3994/270 | 84/18 | 11,181/472 | 4.06 | 4.08 | 1.95 | 4.33 |
| 24 | 2898/202 | 2521/173 | 385/63 | 2354/309 | 4.09 | 3.21 | 3.03 | 4.87 |
| 25 | 2927/208 | 2070/162 | 135/25 | 3830/310 | 4.17 | 4.13 | 2.28 | 4.22 |
| 26 | 821/128 | 1642/103 | 417/51 | 368/80 | 3.89 | 2.99 | 2.68 | 3.73 |
| 27 | 1328/172 | 952/132 | 856/76 | 7218/410 | 4.05 | 3.91 | 3.06 | 4.12 |
| 28 | 58/19 | 6703/226 | 3314/94 | 8181/434 | 2.03 | 3.56 | 2.62 | 4.18 |
| 29 | 259/68 | 7618/242 | 1539/116 | 898/121 | 3.21 | 3.54 | 3.29 | 3.95 |
| 30 | 423/104 | 1802/192 | 807/62 | 6811/273 | 3.89 | 4.05 | 2.60 | 3.02 |
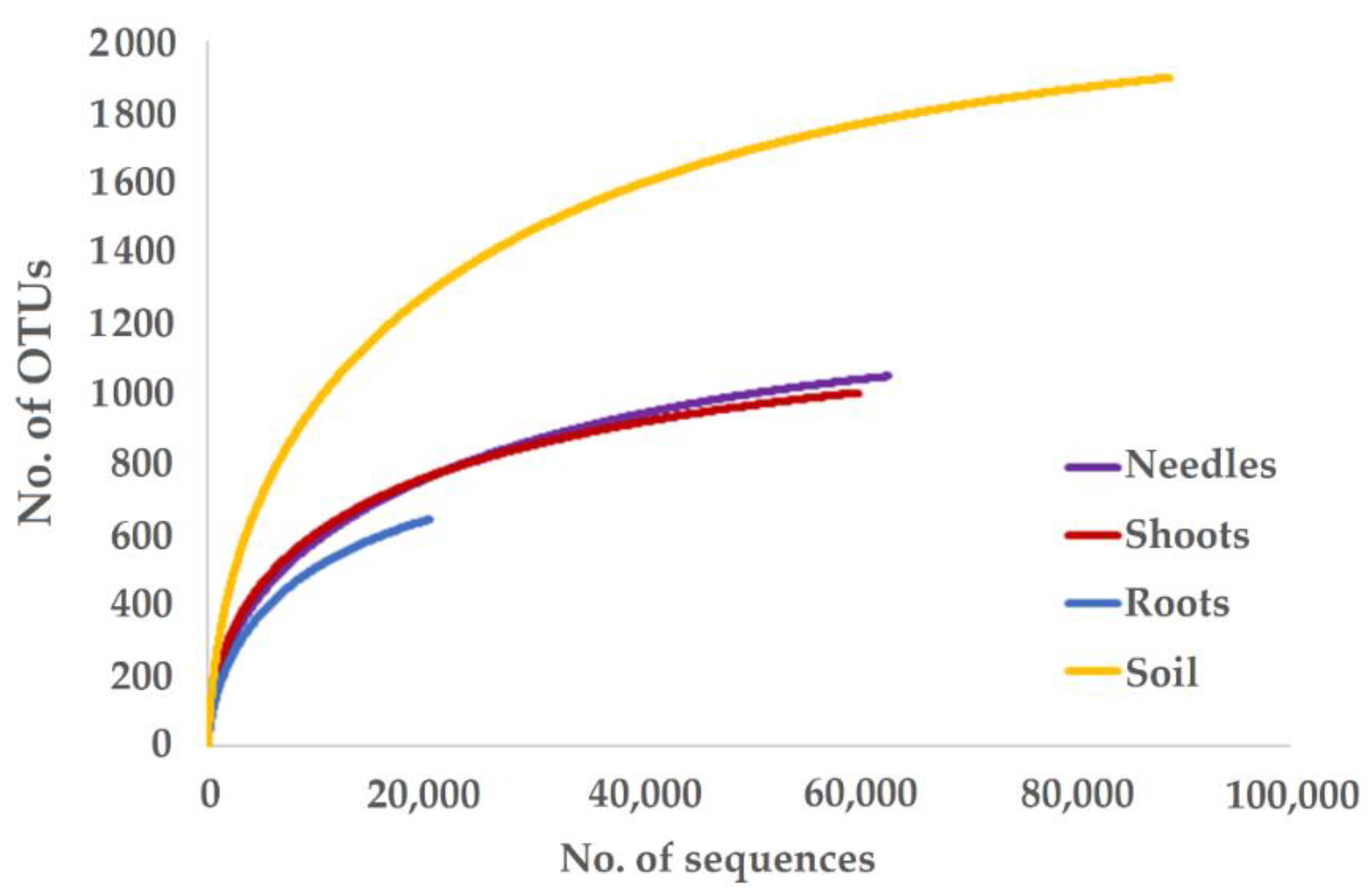
| Total | 62,966/1049 | 60,188/1002 | 20,443/641 | 88,951/1895 | ||||
| Fungal OTUs | Phylum * | Genbank Reference | Similarity, % | Needles, % | Shoots, % | Roots, % | Soil, % | All, % | Ecological Role |
|---|---|---|---|---|---|---|---|---|---|
| Aspergillus pseudoglaucus | A | MT582752 | 100 | 29.9 | 0.01 | 0.1 | 0.05 | 7.9 | Other ** |
| Archaeorhizomyces sp. 5208_0 | A | MH248043 | 100 | 2.4 | 0.3 | 18.4 | 3.3 | 3.6 | Unknown |
| Rhinocladiella sp. 5208_3 | A | KM056296 | 98 | 1.7 | 5.4 | 0.01 | 0.5 | 2.0 | Saprotroph |
| Arachnopeziza sp. 5208_27 | A | MH558278 | 97 | 0.7 | 6.0 | - | 0.5 | 1.9 | Saprotroph |
| Trichomerium sp. 5208_23 | A | NR137946 | 97 | 2.7 | 3.9 | - | 0.4 | 1.9 | Endophyte |
| Phacidium lacerum | A | MN588163 | 100 | 0.6 | 0.3 | 0.2 | 3.7 | 1.7 | Pathogen |
| Unidentified sp. 5208_1 | A | MT595563 | 92 | 1.2 | 3.7 | 0.03 | 1.0 | 1.6 | Unknown |
| Dothideomycetes sp. 5208_5 | A | KX908472 | 99 | 0.6 | 0.5 | 0.1 | 3.3 | 1,5 | Unknown |
| Cladosporium herbarum | A | MT635288 | 100 | 2.1 | 0.9 | 0.3 | 1.7 | 1.5 | Saprotroph |
| Cyphellophora sessilis | A | KP400571 | 100 | 3.8 | 1.4 | 0.01 | 0.1 | 1.4 | Pathogen |
| Rhizosphaera kalkhoffii | A | KY003236 | 100 | 3.5 | 1.1 | 0.07 | 0.5 | 1.4 | Pathogen |
| Lophium arboricola | A | MK159395 | 100 | 0.2 | 4.6 | 0.01 | 0.3 | 1.4 | Unknown |
| Chaetothyriales sp. 5208_15 | A | KP400572 | 100 | 1.6 | 3.0 | 0.01 | 0.4 | 1.3 | Unknown |
| Sporidesmium sp. 5208_41 | A | MT596057 | 100 | 2.1 | 2.8 | 0.01 | 0.1 | 1.3 | Saprotroph |
| Agaricomycetes sp. 5208_12 | A | FJ553582 | 99 | 0.1 | 0.01 | 0.01 | 3.4 | 1.3 | Unknown |
| Phialocephala fortinii | A | MN947395 | 100 | 0.4 | 0.04 | 9.8 | 0.5 | 1.2 | Endophyte |
| Microsphaeropsis olivacea | A | MT561396 | 100 | 0.6 | 1.5 | 0.1 | 1.5 | 1.1 | Other ** |
| Malassezia restricta | B | LT854697 | 100 | 1.0 | 0.6 | 0.4 | 1.7 | 1.1 | Other ** |
| Clavulina sp. 5208_24 | B | OU498806 | 99 | - | - | - | 2.9 | 1.1 | Mycorrhizal |
| Chaetothyriales sp. 5208_2 | A | JQ342183 | 99 | 0.2 | 0.4 | 0.04 | 2.4 | 1.1 | Unkonwn |
| Pezizomycotina sp. 5208_65 | A | KP843512 | 96 | 0.4 | 3.1 | - | 0.3 | 1.0 | Unkonwn |
| Mycena cinerella | B | KT900146 | 100 | 0.1 | 0.01 | 9.5 | 0.4 | 0.9 | Saprotroph |
| Blumeria graminis f. sp. tritici | A | MT162615 | 100 | 0.01 | 0.002 | - | 2.4 | 0.9 | Pathogen |
| Trechispora sp. 5208_19 | B | JX392812 | 99 | 0.002 | - | 9.4 | 0.1 | 0.8 | Saprotroph |
| Umbelopsis dimorpha | Z | MT138616 | 100 | 0.2 | 0.01 | 0.01 | 2.1 | 0.8 | Endophyte |
| Total of 25 OTUs, % | 54.9 | 39.4 | 48.4 | 33.3 | 42.1 |
| Fungi | Phylum * | Genbank Reference | Similarity, % | Needles, % | Shoots, % | Roots, % | Soil, % | All, % |
|---|---|---|---|---|---|---|---|---|
| Plant pathogens | ||||||||
| Phacidium lacerum | A | MN588163 | 100 | 0.6 | 0.3 | 0.2 | 3.7 | 1.7 |
| Cyphellophora sessilis | A | KP400571 | 100 | 3.8 | 1.4 | 0.01 | 0.1 | 1.4 |
| Rhizosphaera kalkhoffii | A | MN547387 | 100 | 3.5 | 1.1 | 0.07 | 0.5 | 1.4 |
| Microsphaeropsis olivacea | A | MT561396 | 100 | 0.6 | 1.5 | 0.1 | 1.5 | 1.2 |
| Exobasidium arescens | B | FJ896135 | 99 | 0.8 | 0.1 | - | 0.04 | 0.3 |
| Coniochaeta hoffmannii | A | MN341268 | 100 | 0.00 | 0.8 | - | 0.03 | 0.2 |
| Phaeomoniella pinifoliorum | A | MK762595 | 100 | - | - | - | 0.5 | 0.2 |
| Alternaria infectoria | A | MT635276 | 100 | 0.2 | 0.03 | - | 0.1 | 0.1 |
| Hendersonia pinicola | A | KT000192 | 100 | 0.05 | 0.03 | 0.01 | 0.2 | 0.1 |
| Typhula sp. 5208_156 | B | MN902561 | 95 | 0.07 | 0.2 | 0.1 | 0.03 | 0.07 |
| Neonectria sp. 5208_421 | A | LR603781 | 100 | - | - | 0.1 | 0.1 | 0.05 |
| Alternaria alternata | A | OL636518 | 100 | 0.1 | 0.03 | 0.1 | 0.04 | 0.05 |
| Ganoderma applanatum | B | MN906143 | 100 | 0.1 | 0.06 | 0.02 | 0.01 | 0.03 |
| Fusarium sp. 5208_517 | A | MT557415 | 99 | 0.01 | 0.01 | - | 0.06 | 0.03 |
| Hymenoscyphus fraxineus | A | MT155386 | 100 | 0.01 | 0.01 | 0.02 | 0.04 | 0.02 |
| Total of 15 plant pathogens, % | 9.8 | 5.6 | 0.7 | 6.9 | 6.9 | |||
| Mycorrhizal | ||||||||
| Clavulina sp. 5208_24 | B | OU498806 | 99 | - | - | - | 2.9 | 1.1 |
| Inocybe nitidiuscula | B | AM882913 | 99 | - | - | - | 1.9 | 0.7 |
| Cenococcum geophilum | A | HM189724 | 100 | 0.02 ** | 0.003 ** | 2.5 | 0.8 | 0.5 |
| Piloderma lanatum | B | KP783452 | 100 | - | 0.002 ** | 0.005 | 1.2 | 0.5 |
| Inocybe geophylla | B | MK961172 | 99 | - | - | - | 1.2 | 0.4 |
| Russula firmula | B | DQ422017 | 100 | - | - | 0.03 | 1.2 | 0.4 |
| Amphinema sp. 5208_105 | B | KP125811 | 100 | - | - | - | 0.9 | 0.3 |
| Lactarius rufus | B | MK838331 | 100 | 0.03 ** | 0.002 ** | 2.9 | 0.06 | 0.3 |
| Tricholoma sp. 5208_110 | B | MK607553 | 100 | - | - | - | 0.5 | 0.2 |
| Inocybe geophylla | B | MT594793 | 100 | - | - | 0.005 | 0.5 | 0.2 |
| Tylospora asterophora | B | MG597438 | 100 | - | - | 0.04 | 0.4 | 0.2 |
| Amphinema sp. 5208_188 | B | MF352678 | 99 | - | - | - | 0.4 | 0.1 |
| Cortinarius scotoides | B | MW555551 | 99 | - | - | - | 0.3 | 0.1 |
| Piloderma sphaerosporum | B | MK131527 | 100 | 0.05 ** | - | 0.2 | 0.2 | 0.1 |
| Inocybe pseudodestricta | B | JF908157 | 100 | - | - | - | 0.3 | 0.1 |
| Total of 15 mycorrhizal OTUs, % | 0.1 | 0.007 | 5.8 | 12.7 | 5.4 | |||
| Endophytes | ||||||||
| Trichomerium sp. 5208_23 | A | NR_137946 | 97 | 2.7 | 3.9 | - | 0.4 | 1.9 |
| Phialocephala fortinii | A | MN947395 | 100 | 0.4 | 0.0 | 9.8 | 0.5 | 1.2 |
| Umbelopsis dimorpha | Z | MT138616 | 100 | 0.2 | 0.005 | 0.01 | 2.1 | 0.8 |
| Mollisia scopiformis | A | OM337553 | 100 | 0.003 | 0.6 | - | 0.05 | 0.2 |
| Mollisia sp. 5208_387 | A | OK430930 | 97 | 0.003 | - | 0.7 | 0.007 | 0.06 |
| Mollisia sp. 5208_1830 | A | MG195564 | 96 | 0.003 | 0.002 | 0.03 | 0.002 | 0.005 |
| Mollisia novobrunsvicensis | A | MT026439 | 100 | 0.002 | 0.007 | - | - | 0.002 |
| Vestigium sp. 5208_802 | A | NR_121556 | 94 | - | 0.003 | - | 0.003 | 0.002 |
| Mollisia fusca | A | LC425049 | 98 | 0.005 | 0.002 | - | - | 0.002 |
| Phialocephala fusca | A | KU668953 | 99 | - | - | - | 0.004 | 0.002 |
| Phialocephala sp. 5208_2177 | A | AB671500 | 98 | - | - | - | 0.004 | 0.002 |
| Phialocephala bamuru | A | MN006138 | 97 | - | - | 0.02 | - | 0.002 |
| Mollisia sp. 5208_3430 | A | MG195527 | 98 | - | - | 0.02 | - | 0.002 |
| Mycroceros sp. 5208_4863 | B | KT186373 | 96 | - | 0.007 | - | - | 0.002 |
| Cadophora sp. 5208_2138 | A | KY987540 | 97 | 0.002 | - | - | 0.002 | 0.001 |
| Total of 15 endophyte OTUs, % | 3.4 | 4.5 | 10.5 | 3.0 | 4.2 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marčiulynas, A.; Marčiulynienė, D.; Mishcherikova, V.; Franić, I.; Lynikienė, J.; Gedminas, A.; Menkis, A. High Variability of Fungal Communities Associated with the Functional Tissues and Rhizosphere Soil of Picea abies in the Southern Baltics. Forests 2022, 13, 1103. https://doi.org/10.3390/f13071103
Marčiulynas A, Marčiulynienė D, Mishcherikova V, Franić I, Lynikienė J, Gedminas A, Menkis A. High Variability of Fungal Communities Associated with the Functional Tissues and Rhizosphere Soil of Picea abies in the Southern Baltics. Forests. 2022; 13(7):1103. https://doi.org/10.3390/f13071103
Chicago/Turabian StyleMarčiulynas, Adas, Diana Marčiulynienė, Valeriia Mishcherikova, Iva Franić, Jūratė Lynikienė, Artūras Gedminas, and Audrius Menkis. 2022. "High Variability of Fungal Communities Associated with the Functional Tissues and Rhizosphere Soil of Picea abies in the Southern Baltics" Forests 13, no. 7: 1103. https://doi.org/10.3390/f13071103