
Oak Decline Caused by Biotic and Abiotic Factors in Central Europe: A Case Study from the Czech Republic
by
 , , and
, , and
Markéta Macháčová
1,*, 
Oto Nakládal
1,
Michal Samek
2,
Daniel Baťa
1,
Václav Zumr
1 and
Vítězslava Pešková
1 1
Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Kamýcká 129, 165 00 Prague, Czech Republic
2
Department of Forest Protection Service, Forestry and Game Management Research Institute, Strnady 136, 252 02 Jíloviště, Czech Republic
*
Author to whom correspondence should be addressed.
Forests 2022, 13(8), 1223; https://doi.org/10.3390/f13081223
Submission received: 24 June 2022
/
Revised: 24 July 2022
/
Accepted: 27 July 2022
/
Published: 2 August 2022
(This article belongs to the Special Issue Ecology and Management of Forest Pests)
Abstract
:Oak decline presenting symptoms of tracheomycosis have been observed globally over long periods of time. Since the 1990s, oak decline has been considered to be a multifactorial process where abiotic predisposing factors play a significant role. Nevertheless, biotic factors, such as subcortical insects and fungal pathogens, may influence the decline process, as some insect species transmit spores of tracheomycotic pathogens. We investigated biotic agents (subcortical insects, Ophiostoma spp. and Phytophthora spp.) in six declining oak stands located within mesophytic and thermophytic zones in the Czech Republic, where five dry years occurred from 2015 to 2019. The spectrum of insect pests of oak stands was investigated using two methods: reared from logs from each stand and window traps. The presence of Ophiostoma fungi was assessed from cultivated woodblocks; Phytophthora pathogens were isolated from soil samples. In total, 2931 subcortical insect individuals were captured during the 2021 vegetation period. The most species-rich group of the subcortical insect complex involved in oak decline consisted of the families Cerambycidae: Xylotrechus antilope; Curculionidae: Scolytus intricatus, Xyleborus monographus; and Buprestidae: Agrilus biguttatus, A. sulcicollis. The presence of Ophiostoma was detected in all stands with different intensities between tree parts. Several important oak pests were positively correlated with the occurrence of Ophiostoma. On the contrary, from all soil samples, Phytophthora plurivora was found at only one studied oak stand. Based on the results, a new modified model of biotic and abiotic factors involved in the oak decline is proposed.
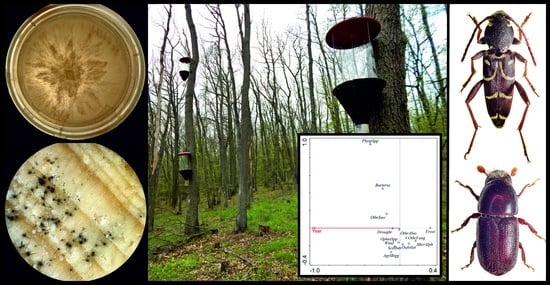
1. Introduction
Since the beginning of the 20th century, oak decline has occurred frequently across Europe in periodic waves preceded by periods of climatic extremes. Nevertheless, cases of oak decline have been reported regularly since the 18th century. Decline frequently affects Quercus robur L. and, to a lesser extent, Q. petraea (Matt.) Liebl., the two dominant species in Europe, but also other oak species. The phenomenon of oak decline has been studied in many oak species around the world (America, Asia) [1,2,3,4,5,6,7,8].
Episodes of decline have occurred sporadically, and since the 1980s (due to dry seasons), it has been recorded from many countries across Europe in a wide range of oak species (both evergreen and deciduous) (see Figure 1 and Figure S1). Decline has occurred in individual trees, groups, or whole forest stands, and various theories have gradually emerged to explain the nature of the problem. Research findings have shown, however, that neither symptoms nor causes may be the same in all cases [1,9,10,11,12,13]. A wide range of abiotic and biotic factors, including frost, drought, air pollutants, decreased groundwater levels, silvicultural mismanagement, insect defoliators, bark and woodboring beetles, fungal species such as Ophiostoma spp. and Ceratocystis spp., bacteria, mycoplasma-like organisms and viruses, have been discussed as predisposing and initiating factors of this phenomenon [1,3,9,10,13,14,15,16,17]. Last but not least, a factor that has also been studied in relation to oak decline is oak phenology and its changes resulting from climate change. The phenological variability in oak species has an important adaptive significance to abiotic and biotic factors, and climate change can cause higher susceptibility to oak decline [18,19,20,21,22].
The first scientific publication on the subject, in 1926, described “vascular mycosis of oak” in Yugoslavia [23,24]. Between the 1940s and 1960s, research in Romania and Russia led to the theory that fungi of the genera Ophiostoma and Ceratocystis were the cause of vascular mycoses, later known as tracheomycosis [25]. Frequent isolation of these pathogens from dying oak trees in other countries supported this theory for a long time. From the 1990s onwards, however, views emerged that did not accept this theory [10,11,26] causes were looked for elsewhere, often as a combination of multiple factors [12]. New theories argued that oak decline is not a single disease but rather a process driven by several diseases involving site factors, environmental factors, pests, and pathogens [12]. Fungi were most frequently seen as a possible cause, yet only a few of these fungi have actually been shown to cause tree mortality, and then only in trees already weakened by other factors [3].
Abiotic stress factors (seasonal dry periods and exposure to severe frost in winter) were considered critical predisposing factors for oak decline leading to tree mortality through the action of subcortical insects and root pathogens on weakened trees [12,25,57]. Other theories suggested that the cause of dieback in European temperate forests was repeated defoliation by insect larvae, the effect of which was compounded by powdery mildew [13].
During the 1990s in Czechoslovakia, tracheomycosis—a disease of vascular bundles caused by fungi—was often studied. This disease has long been considered the main cause of oak decline [74]. The causal agents are mainly fungi from Ophiostomatales, which cause damage by secreting toxic metabolites into living vascular bundle tissues [75]. These pathogens were found on both weakened and healthy trees [76]. Other studies, however, have shown the presence of these fungi only on some symptomatic trees [77]. Recent evidence suggests that these fungi are non-pathogenic or weakly pathogenic to healthy oaks but may be a significant pathogen when these trees are weakened by water deficit [9,78,79]. Many studies showed that ophiostomatoid fungi do not have the ability to invade healthy trees [56,67,80,81,82,83,84,85]. The occurrence of fungi of the genus Ophiostoma seemed to be conditional upon dead or dying tissues [86]. Ophiostomatoid fungi occur naturally in woody plants, where many of them live in the galleries of bark beetles and are quite often observed in necrotic bark parts and wood of various species of trees, where they act as secondary or, more rarely, primary parasites [87]. Thus, they are a rather endophytic species that can switch to parasitism and cause damage only under suitable conditions when the tree is weakened [83,86,88]. However, the interaction between fungi and insects, and especially the order Ophiostomatales and bark beetles, began to receive more attention in the 1930s in the context of Dutch Elm Disease and the causal agent Ophiostoma ulmi transmitted by insect vectors. Most Ophiostoma species produce spores in sticky droplets that can easily attach to the exoskeletons of their insect vectors [89]. The genus Ophiostoma includes plant-associated species with varying degrees of pathogenicity. Most species are regarded as non-pathogenic, especially in their endemic range, where they have co-evolved with their host tree species and are mainly responsible for causing blue-stain in freshly exposed sapwood [90]. In Czechoslovakia, studies were carried out on this topic in the 1950s [91]. The transmission of ophiostomatoid fungi was observed in several subcortical species [92,93]. This view is supported by the fact that members of the Ophiostomatales exist in symbiosis with bark beetles in Central Europe, as confirmed by many studies [94,95,96,97,98]. Similar transmission mechanisms could operate in other subcortical insect–fungal interactions. Some studies suggest that fungi of the genus Ophiostoma are not common endophytes of oaks and that none of them are closely related to Scolytus intricatus (Ratzeburg, 1837).
Other fungal pathogens were also considered to be involved in the oak decline phenomenon. Diplodia mutila (Fr.) Mont., Fusarium spp., Fusicoccum quercus Oudem., Phomopsis quercella (Sacc. and Roum.) Died. and Biscogniauxia mediterranea (De Not.) Kuntze [3,6], and especially Armillaria spp. pathogens were often studied [3,70,86,99,100,101]. Interestingly, infections with Armillaria mostly occur after the trees have been infested by Agrilus spp. [43]. Other wood-decay fungi, such as Stereum sp., Phellinus robustus (P. Karst.) Bourdot and Galzin, or Fistulina hepatica (Schaeff.) With., have also often been associated with oak decline as agents attacking frost-damaged trees in particular [102]. Collybia fusipes (Bull.) Quél. also has been shown to attack weakened trees [3].
In the 1990s, pathogens of the genus Phytophthora were identified as new biotic factors in oak decline [4,28,29,30,31,32]. Forest stands of Quercus robur (pedunculate oak) and Q. ilex (holm oak), widespread in temperate and Mediterranean regions of Europe, respectively, are highly sensitive to the impact of root rot caused by Phytophthora species. These pathogens cause many major plant disease epidemics and are among the most destructive plant pathogens in forests worldwide [103,104]. Since the 1990s, several important episodes of oak decline have occurred where Phytophthora were identified as causal agents. Numerous surveys in oak stands throughout Europe have uncovered a diverse assemblage of Phytophthora taxa. Several of these Phytophthora species, including P. cambivora (Petri) Buisman, P. cinnamomi Rands, P. cryptogea Pethybr. and Laff., P. drechsleri Tucker, P. multivora Scott and Jung, P. plurivora Jung et Burgess, and P. ramorum Werres, de Cock and Man in ’t Veld, are introduced invasive pathogens in Europe, whereas most other species are considered to be native or of cryptic origin [105]. Phytophthora quercina Jung is an example of an oak pathogen contributing significantly to oak decline in dry periods [30]. Phytophthora cinnamomi is an aggressive pathogen that can cause oak decline on its own, apparently therefore without the contribution of additional factors [4,106]. The results of current scientific studies suggest that root diseases caused by Phytophthora pathogens in combination with climatic extremes are causing severe forest dieback across Europe [107].
Oaks are hosts to a wide variety of insect species [108]. Among the bark insects, oak provides a haven for a wide range of representatives, some of which can cause severe damage leading to the mortality of entire trees [109]. An attack by a subcortical insect often is the main cause of tree mortality [110]. Various species of subcortical insects (e.g., Curculionidae: Scolytinae [111,112] or Xiphydria spp. [113]) act as vectors of ophiostomatoid and other fungal species. Nevertheless, these and other species of cambiophagous and subcortical insects also can cause significant damage in oak stands by their direct action. Soukup (1995) found in his study (1991–1995) that even repeated heavy insect defoliations did not cause significant deterioration in experimental plots, but monitoring showed locally high abundance of Scolytus intricatus, Agrilus sulcicollis (Boisduval and Lacordaire, 1835), Xiphydria species, longhorn beetles of the genera Clytus spp. and Xylotrechus spp. and some other species [114]. Another important subcortical pest, Agrilus biguttatus (Fabricius, 1776), occurs as the earliest one in the progression of decline and has a strong impact on tree vigor [49,115].
The aim of this study was to determine the causes of oak decline in six oak stands in the Czech Republic. The goal was to find out how the dry seasons during 2015 –2019 [116] and biotic factors such as insect pests and fungal pathogens contributed to the oak decline.
2. Materials and Methods
2.1. Review Methodology and Data Analyses
A literature review was conducted based primarily on a collection of publications on the Web of Science database concerning the topic of “oak decline” in Europe (keywords: “oak decline”, “oak dieback”, “oak wilt”, “acute oak decline”, “sudden oak death”, “tracheomycosis”, “Ophiostoma”). In addition, conference proceedings from the 1990s and early 2000s were used as sources, as well as references from publications and proceedings from other relevant sources. The review’s aim was to capture the history of decline episodes in Europe and their causes. We did not consider publications concerning damage caused by storms, browsing by herbivorous vertebrates, or management practices.
Based on the data obtained from the literature, a database was created to determine the most common and important causes of oak decline. The results of a multivariate statistical analysis then show which factors occur together over time. Using CANOCO 5 software, canonical correspondence analysis (CCA) was used to evaluate the chronology of oak decline factors.
Also processed were meteorological data available in European databases from 1901 to 2020. These data were examined in the context of oak decline episodes. Because drought is reported as the main driver of decline in most episodes, we compared oak decline incidence data with precipitation data. The aim was to reveal whether there is a relationship between the occurrence of drought and oak decline. The source of the precipitation data was the data set published by [117]. The dataset consists of gridded month-by-month variations in climate over the period from 1901 to 2020, with spatial resolution of 0.5° × 0.5°. Monthly precipitation data were summarized over the hydrological year (1 November to 31 October), average annual precipitation over the entire period of record was calculated, and the variations relative to the 1902–2020 mean also were calculated.
2.2. Study Plots
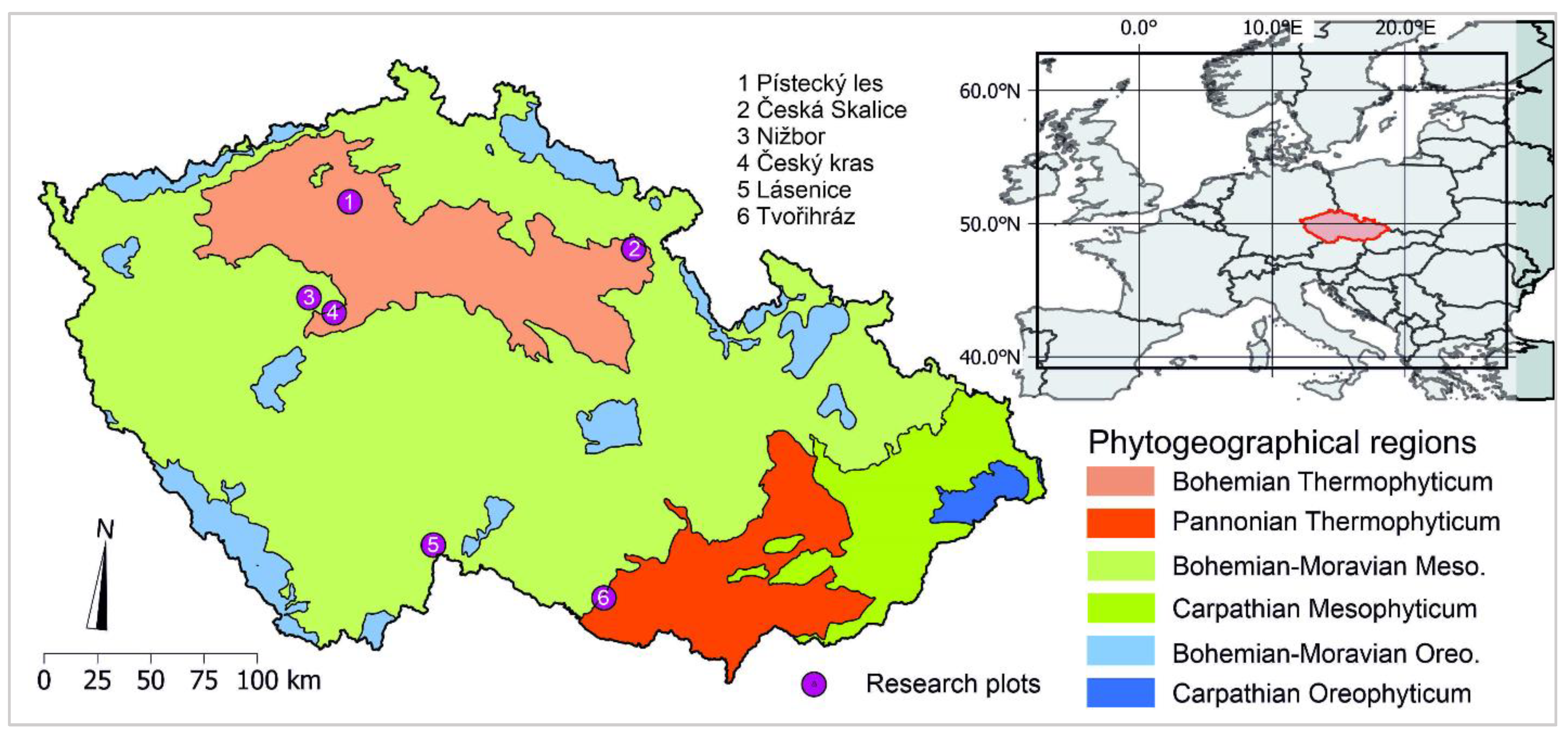
The research on the current oak decline situation in the Czech Republic was conducted during 2020/2021 and involved six oak stands (Quercus robur and Q. petraea) in various locations across the country. All the selected stands are managed forests originating from artificial regeneration. The plots were selected based on different levels of decline symptoms that included defoliation in the crown, formation of epicormic shoots, high crown transparency, and tarry exudates on the bark. Although the stands were characterized by weakened conditions and contained symptomatic trees, decline symptoms were not uniformly observed in all stands. Plot altitudes ranged from 198–530 m a.s.l. (Figure 2, Table 1). The studied plots are located in the mesophytic (Nižbor, Lásenice) and thermophytic (Pístecký les, Český kras, Tvořihráz, Česká Skalice) zones.
2.3. Insect Trapping
The spectrum of insect species in oak stands was investigated using two methods: (A) by rearing from trees (logs/branches) collected at each site, and (B) by capture with window traps placed on trees with visible symptoms of decline. Tree samples were collected from all sites during 5–12 March 2021. At each location, the tree showing the most significant evidence of wood-boring insects’ infestation was felled. From such a tree, 2 m were cut from the basal, middle, and crown portions of each tree. Heights for each section were relative to each tree’s absolute height (see Table 1: Average Height—the absolute height of the sampled tree corresponds to the average height of the studied oak plot). These samples were placed in rearing cages separately and all reared insects were collected regularly until September 2021. The reared insects were counted and are reported herein as their numbers per 1 m of tree stem within the three stem sections (basal, middle, crown) for each site. (Thus, results from rearing cages will be stated decimally, as opposed to in whole numbers.)
Two window traps were installed on three trees per site showing symptoms of decline (at heights 2 and 7 m). Five collections were made at regular monthly intervals, the last one during 13–15 September 2021. All Coleoptera (except Staphylinidae) and Hymenoptera: Xiphydriidae insects were determined to species level. Among the beetles, only primary wood-boring species directly associated with oak were selected and presented in the results section. All the insects were identified by the second author.
In this study, two methods for insect trapping were also compared. Correlation between numbers of reared specimens and captured specimens to the window traps was tested by correlation method from transformed data by function y = ln(x + 1) (software Statistica 14.0).
2.4. Sampling, Isolation, and Identification of Ophiostoma and Phytophthora
Tree samples were used to study the presence of Ophiostoma species. The modified isolation and identification methodology was performed according to [118]. Woodblocks 1–3 cm thick were cut from the tree’s basal, middle, and crown parts and incubated in moist chambers for 8 weeks at 23 °C. After fructification, the presence of perithecia growing from the wood was assessed according to [76]. Intensity of the perithecia presence was assessed by observing them under a binocular microscope (Olympus SZ61) on a scale of 0–3 (0 = no occurrence, 1 = weak occurrence, 2 = medium occurrence, 3 = strong occurrence).
A sampling of rhizosphere soil was used to determine presence of Phytophthora species in the oak stands. The methodology was performed according to [30]. Three to four soil monoliths (size about 20 · 30 · 30 cm) were taken from the rhizosphere of 10 trees per plot at distance 50–150 cm from the stem bases. Both symptomatic and asymptomatic trees were sampled. Each soil sample was thoroughly mixed and a subsample of ca 200 mL was taken for isolation testing that used young leaves of Quercus suber L. and Fagus sylvatica L. as baits. The leaflet baiting test was performed at 20 °C in natural light. After the appearance of the first necrotic spots, baiting leaves were examined for the presence of Phytophthora sporangia under a light microscope. Small pieces from the necroses were then plated onto selective PARPNH agar [119]. The first hyphae from plated leaves were subcultured onto V8-agar and carrot juice agar (800 mL distilled water, 200 mL carrot or vegetable juice), 18 g agar (Sigma-Aldrich, St. Louis, MO, USA), and 3 g CaCO3 [120], then stored at 20 °C for further examination. Three- to four-week-old colonies were observed at 400× magnification for the presence of typical Phytophthora sexual and asexual structures using a light microscope (Olympus BX41). The Phytophthora cultures were grouped according to morphological characters and compared with species descriptions in the literature [120,121]. Molecular identification of the isolates was performed according to [122] using specific primers ITS4 and ITS6. The nucleotide Basic Local Alignment Search Tool (blastn) was used to compare obtained sequences with those in the National Center for Biotechnology Information (NCBI) database (http://www.ncbi.nlm.nih.gov, accessed on 15 February 2022).
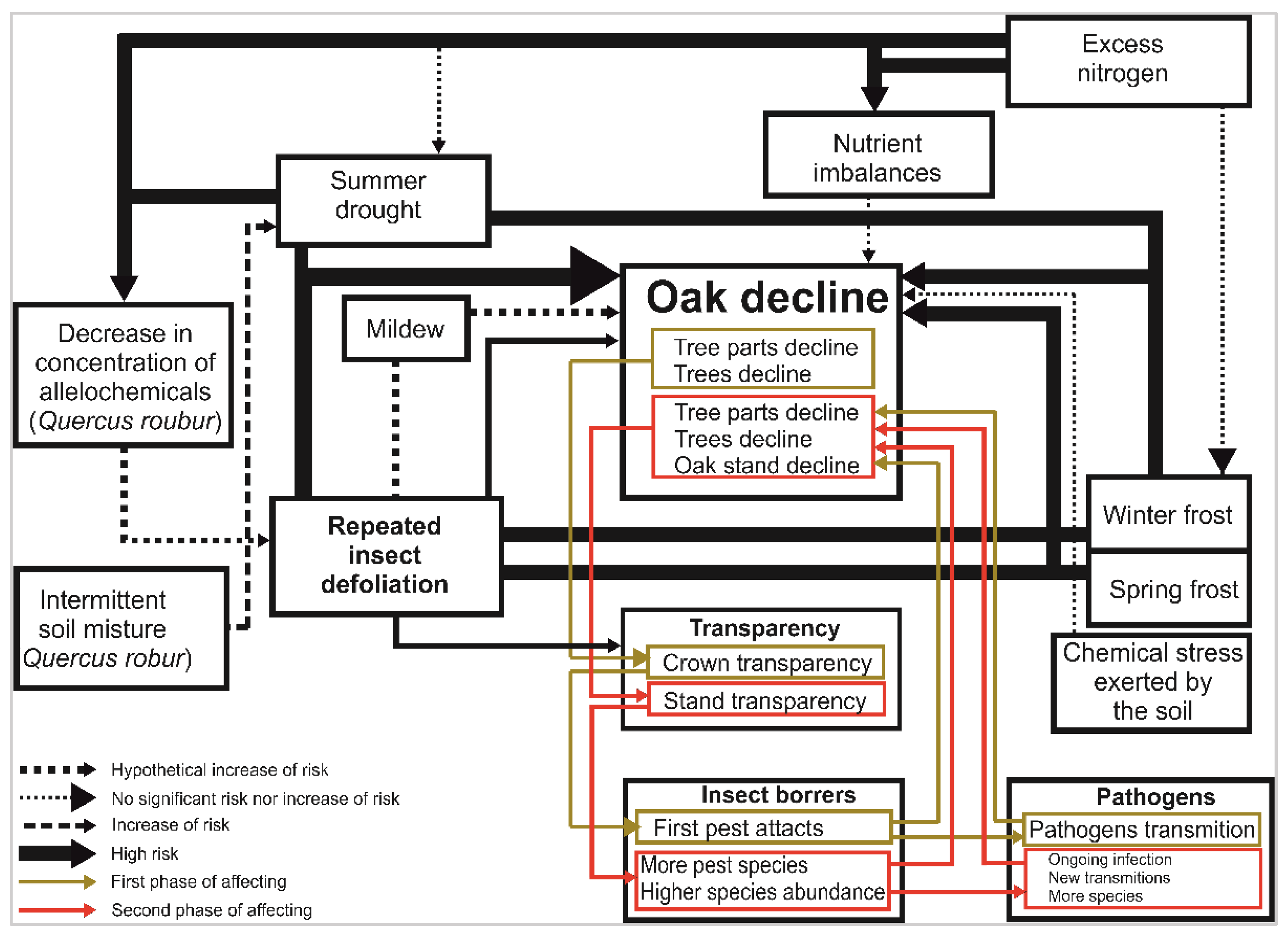
2.5. A New Modified Model of Factors Involved in Oak Decline
The new model was based on the publication by [13] and the “Conceptual model of the interaction of significant abiotic and biotic factors in the onset of the decline of oaks in Central Europe” model included there [13]. We elaborate more in the section regarding insect pests based on the results of this case study. Based on the literature search, we concluded that changes in crown transparency due to defoliation or inappropriate cultivation interventions can lead to different light and microclimatic conditions in the remaining forest vegetation and, therefore, that new insect pests, such as the wood-boring insect Agrilus biguttatus, can suddenly be attracted to that vegetation [123].
3. Results
3.1. Insect Pests
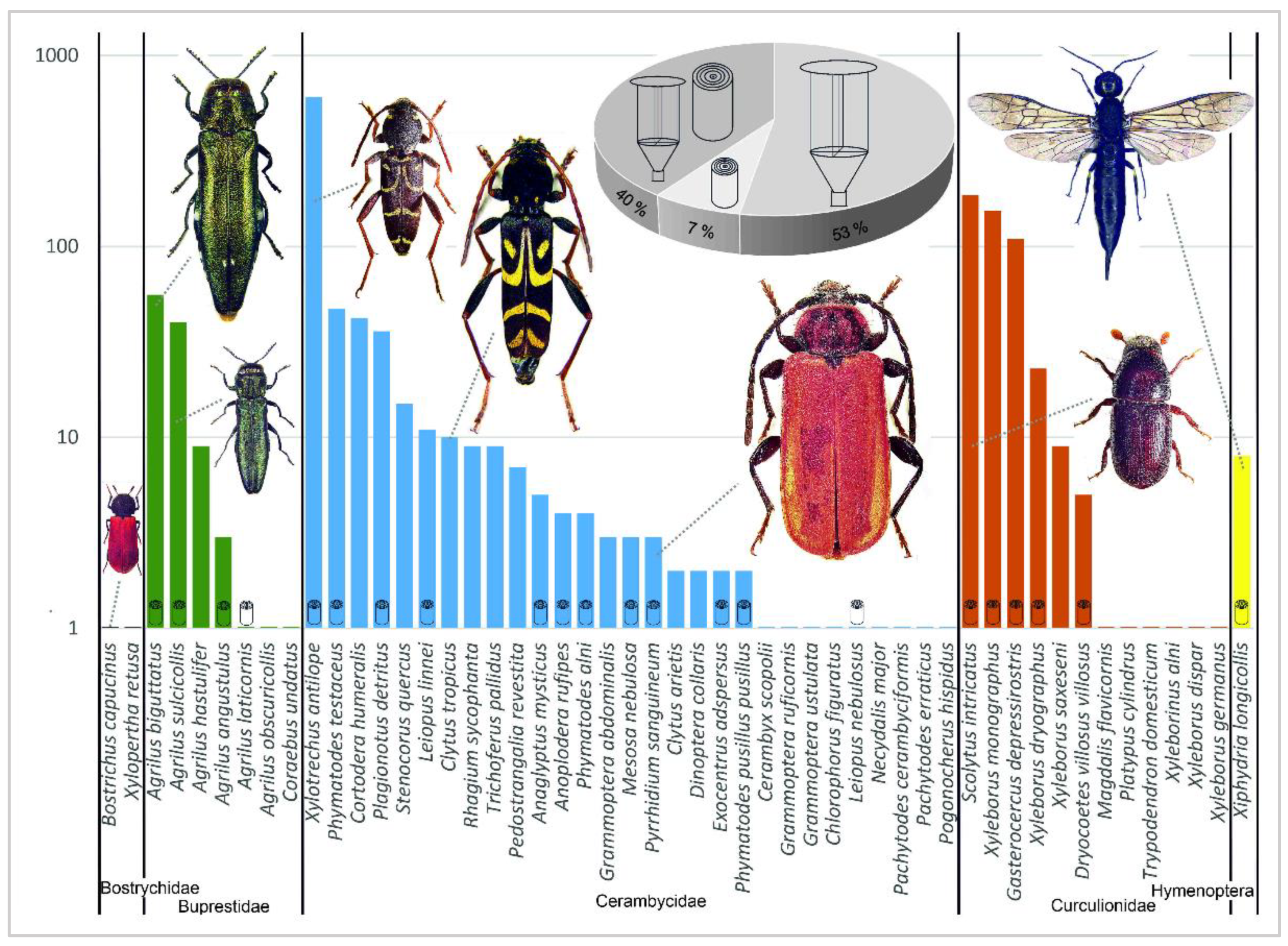
During the study, a total of 2931 subcortical insect individuals were captured. Of these, 1476 subcortical insects were reared from the infested logs and branches using the rearing cages (RC) and 1455 subcortical insects were captured in window traps (WT). A total of 55 subcortical insect species belonging to two orders were recorded: Coleoptera (54 species) and Hymenoptera (1 species). The specific species for each family are given in Appendix A. The most species-rich group of the subcortical insect complex involved in oak decline consisted of the family Cerambycidae: 30 species (29 from WT, 13 from RC), Curculionidae: 14 (13, 6), Buprestidae: 8 (7, 5), Bostrichidae: 2 (1, 1), and Xiphydriidae: 1 (1, 1). Species abundance showed the same order for both passive windows trapping and rearing from the logs + branches (p < 0.001, r = 0.694). Data regarding the method of capture (trapping vs. rearing) to specific species are presented in Figure 3 and Figure 4 (see Table A1 for the absolute number of records).
From these 2931 individuals, 93% of all insect species were captured in the window traps. The use of rearing cages to rear imagoes from the infested trees thus increased the total number of species captured by only 7%. In contrast, had rearing been the main collection method, the addition of window traps would have increased the species contribution by 53%. The use of traps appears to be a significantly more effective method of studying the spectrum of subcortical forest insect when six traps (hung on three trees at two height levels) are used compared to rearing from 6 m of logs (taken from a single tree at three height levels) (see pie chart in Figure 4).
Within the window traps, Xylotrechus antilope (Schönherr, 1817) (Cerambycidae) was the most abundant species (604 individuals captured in all plots) and could therefore be considered the most damaging longhorn beetle species. The common bark beetle species Scolytus intricatus was captured in the highest numbers both in WT (186) and RC (214.1 in three 1 m sections from basal + middle + crown parts (see Section 2.3 in Materials and Methods)). The second most abundant species of Curculionidae was Xyleborus monographus (Fabricius, 1792), which was found in all the plots (154 from WT, 48.4 from RC). Another important curculionid beetle, Gasterocercus depressirostris (Fabricius, 1792), was captured mainly at “Český kras” and “Tvořihráz” (83/25 in WT) and, interestingly, at “Lásenice” only in RC (46.8). Agrilus biguttatus (56) and A. sulcicollis (52) from the Buprestidae family were captured in the highest numbers in WT mostly in Český kras and Nižbor. These were very dry locations with the highest numbers of declining and dead trees (Figure 3 and Figure 4).
The other species were represented by fewer than 50 trapped specimens in WT per plot. The majority (73.4%) of all subcortical insect individuals (1068) were captured in WT at Český kras (Table A1).
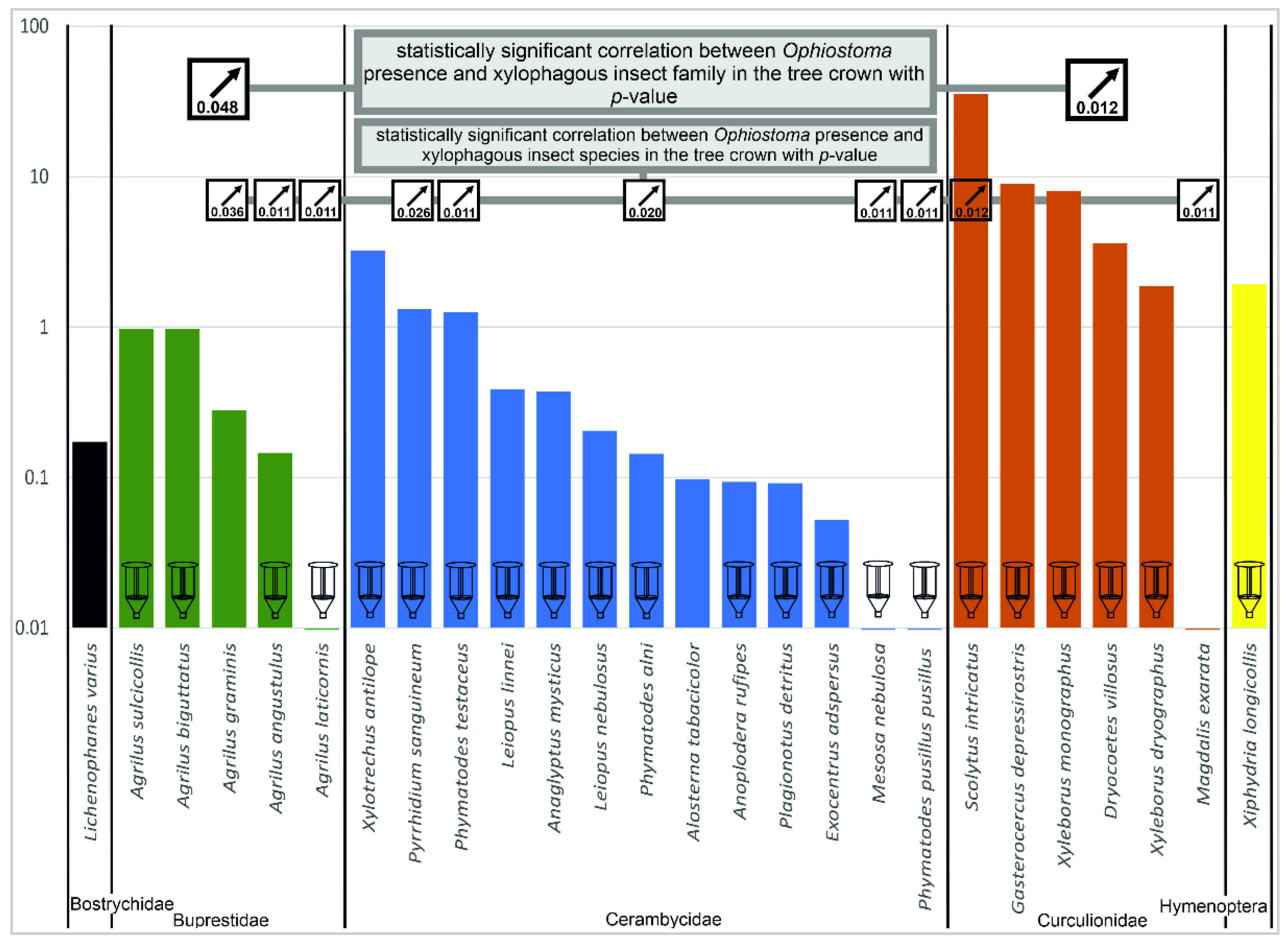
The correlation between Ophiostoma presence and subcortical insects is shown in Figure 3. Statistically significant correlation was found for Buprestidae (Agrilus graminis, A. angustulus, A. laticornis), Curculionidae (Scolytus intricatus), and for several Cerambycidae species. Notably, significant correlation between presence of Ophiostoma and the insects (both on species and family levels) was observed only in crown samples.
3.2. Presence of Ophiostoma and Phytophthora
The presence of Ophiostoma fungi in sexual stages was detected in samples from all oak stands. Differences were found among individual stands and among different parts of the trees (see Table 2).
Phytophthora species were isolated from a total of 24 soil samples collected at all six stands (four mixed samples per stand near two declining and two healthy trees). The isolation was performed in autumn 2020 using the leaf-baiting method. Positive isolation was made at only one stand out of six (Pístecký les), even though the oak stand there showed the least signs of decline. In total, 12 isolates were obtained and identified as Phytophthora plurivora based on morphological characteristics [120] and internal transcribed spacer (ITS) sequence. Phytophthora plurivora was isolated from rhizosphere soil samples at both declining (8 isolates) and healthy (4) trees.
3.3. A New Modified Model of Abiotic and Biotic Factors Involved in Oak Decline
Our results suggest a new model summarizing oak decline’s predisposing and triggering factors (Figure 5). This model considers the influence of abiotic factors that weaken oak trees and the impact of biotic agents that weaken oak stands secondarily. These include defoliators, which cause light penetration of stands and thus affect microclimatic conditions in the habitat. Moreover, weakening of the trees also occurs with inappropriate cultivation interventions. When many trees are removed, the stand may become significantly thinned, and crown transparency changes. The trunks of the remaining trees are suddenly more exposed to sunlight, which attracts subcortical insect such as A. biguttatus or S. intricatus. Especially bark beetles act as vectors of ophiostomatoid fungi, which further contribute to the oak decline.
The most common chronology and important causes of oak decline were studied based on the oak decline episodes database. Multivariate statistical analysis showed that factors involved in oak decline changed over time (Figure 6). In the oldest records, usually there was a single factor, or a couple of factors occurred together (e.g., frost, drought, Microsphaera alphitoides Griffon and Maubl. + defoliators). Gradually, more factors began to be cited as acting together, some of which (usually drought) were presented as the main factor and others as secondary factors. In the 1990s, also new biotic factors appeared, such as Phytophthora or bacteria. The results showed a significant relationship between factors and the timescale of oak decline records.
4. Discussion
Numerous studies showed that abiotic stresses are among the most important factors in oak decline and include summer droughts, changes in precipitation, excess precipitation and summer droughts, cold winters, as well as winter and/or spring frosts [13,42,57,124,125]. All these factors are becoming more important as the oak phenology is shifted due to global warming. Climate change causes the initiation of cambial activity and leaf development earlier than several decades ago. This results, for example, in higher risk exposure to spring defoliation caused by late spring frost in the case of early oaks [19,20]. In the conceptual model for the interaction of abiotic and biotic factors crucial in the emergence of oak decline by [13], the three factors considered to be decisive causal factors for oak decline are insect defoliation, summer drought, as well as winter and/or spring frost. Normally, at least two of these factors (always including defoliation) have to occur simultaneously to trigger decline. Our results show that abiotic factors (especially drought) cause weakening of individual trees or groups of trees, but mainly it is the reaction to the subsequently occurring biotic agents that can cause severe dieback and mortality of the tree. First, defoliators cause lightening of the stand and thus change microclimatic conditions, which together with inappropriate cultivation interventions [70,126], make the remaining weakened trees attractive to attack by bark beetles [127]. This process was particularly evident at dry sites (Český kras), where the largest numbers of important wood borers were captured (589 Xylotrechus antilope, 140 Scolytus intricatus). Especially in very dry years, this scenario has been prone to occur, for example in Slovakia or Hungary, since the late 1970s [72,128].
The oak decline phenomenon has always existed, just not exactly in the range where it began to appear in the 1980s and still appears to this day. Probably factors that formerly acted individually have accumulated and begun to act in synergy. Therefore, oak decline is considered a dynamic process with a complexity of interrelationships between the host and its environment [3,85] and a synergistic complex of abiotic, biotic, and anthropogenic factors acting together [12,25,56,57]. The oak decline episodes database showed that different factors occurred together over time (Figure 6). The analyses revealed that the first records usually showed single factors involved in oak decline. Then, gradually, more factors began to act together. Drought is nonetheless reported as one of the most critical factors, and more recent studies consider extreme climatic events [129], especially high temperature stress and summer droughts, to constitute the main factor triggering oak decline in many places within Europe [39,58]. Recognizing this, the relationship between the oak decline incidence and years with below-average total precipitation was evaluated for Central Europe (see Figures S2 and S3). The results show that the duration of below-average annual precipitation is what matters. When approximately five of these dry years occur in succession, oak decline follows quite regularly. In northern Germany, droughts between 1976 and 1983 predisposed sessile and pedunculate oaks to subsequent frost damage in the winters of 1984–1987 [43]. Likewise, in former Czechoslovakia, the first very dry year occurred in 1976 and since then the precipitation deficit has gradually increased in the following years. Consequently, severe oak decline appeared gradually across the whole country [73,130]. Later, in the late 1980s, this scenario repeated, as in other European countries [5].
Due to unfavorable climatic conditions (dry seasons) from 2015 to 2019 [116], it could be assumed that abiotic factors played a significant role in the weakening of the studied oak stands. The subsequent development of the stands could have been influenced by the intensive development of biotic agents, including insect pests. At two studied locations (Tvořihráz, Nižbor), weakening of oaks has been exacerbated by defoliators attack. The subcortical insect species seem to favor such damaged trees, as was observed in Tvořihráz (a thermophytic area in South Moravia), which had the richest complex of subcortical species (29 species). Nevertheless, drought is the initializing factor that induces changes in the ability of oak to resist wood-boring insects, such as by decreasing its production of callus tissue [131,132] and also by increasing the sensitivity to drought by changes in phenology [19,21]. Severe droughts that lead to weakening of oak trees benefit pathogens and pests, and especially buprestids of genus Agrilus. Old oak stands have traditionally been regarded as being susceptible to Agrilus infestation [49], but also much younger trees (about 30–40 years old) have suffered in Poland from severe attacks due to weakening by drought [133]. That finding is consistent with our results showing the greatest occurrence of Agrilus sp. at Český kras and Nižbor (oak stands 42 and 48 years old, respectively). However, as some studies suggest, episodes of drought can affect the health of oaks with delay of several years, making it very difficult to determine the causes of dieback [27,134].
Subcortical insects have been demonstrated to participate frequently in the final phase of the causative chain that leads to mortality of oaks [12,115]. The results of our study show an important significance of these pests in oak decline. The most abundant species included S. intricatus, A. biguttatus, X. antilope, and X. monographus. In all the study plots, we found the smaller black exudates that were associated with A. biguttatus. These symptoms on the bark, together with entry or exit holes, usually indicate the presence of various insect species of which the most common are subcortical beetles, such as buprestids and cerambycids [135,136]. Nevertheless, the dark-colored leaking exudates on the bark surface—often without under-bark insect galleries—were also present in all six study oak stands. However, the origin of the dark exudations remains unclear. They might originate from necrotizing tunneled tissues as a result of larval activity or from colonization by opportunist saprophytes or pathogenic bacteria [123,137].
Some oak pest species that have been considered insignificant or even endangered in the past may be gaining in importance in response to climate change and declining oak stands. Scolytus intricatus was once thought to be of a little significance in forestry [138], but its impact increased considerably in the early 1980s when this species was found to be the most abundant insect pest in a 3-year survey in Slovakia during the most extensive oak decline episode known there [139]. Scolytus intricatus causes damage not only directly by feeding but also as a vector of ophiostomatoid fungi [112,140]. According to our results, S. intricatus was the most abundant species at all the studied sites. Correlation between the occurrence of S. intricatus and Ophiostoma spp. was also confirmed. Xyleborus monographus, too, was recorded on tree samples where Ophiostoma spp. were also identified. This could suggest that both species can act as important vectors of Ophiostoma fungi when attacking the moist wood: S. intricatus during the maturation feeding and X. monographus during the boring of maternal galleries.
The weevil Gasterocercus depressirostris was found to be relatively abundant at 4 out of 6 studied plots. It was both captured in window traps and reared in rearing cages. Interestingly, in the Czech Republic this species was in 2005 included in the Red List of Invertebrates with the status of “Vulnerable species” [141], as well as in 2017 [142]. In Poland it is listed as an “Endangered species” [143] and in Germany as “Critically endangered” [144]. Similarly, the longhorn beetle Clytus tropicus is considered “Near Threatened” in the Czech Republic [142] and “Endangered” in Germany [144]. Other red-listed species, such as Trichoferus pallidus (Cerambycidae) or Agrilus hastulifer (Buprestidae), have been recorded to a lesser extent [142]. The common species A. biguttatus was likely overlooked in the past and was once considered a rare species deserving of protection [42,49], but today it is considered one of the most dangerous pests of oak stands, especially on trees weakened by drought stress [133]. It is the first secondary insect to arrive and one of the most important among the final contributors to oak mortality [129]. Moreover, A. biguttatus appears to be a major player in a new type of oak decline in Great Britain (termed acute oak decline) that appeared in 2008 [145]. Symptoms of the infested trees included bleeding lesions from longitudinal cracks on the trunks with necrotic underlying tissues and a presence of A. biguttatus larval galleries. Bacteria of the Enterobacteriaceae family have been isolated from the infected tissue [146,147]. Another pest, X. antilope (Cerambycidae), was the most abundant species in the present research (604 individuals captured in all window traps) and can be considered an important pest, as mentioned in other studies [148,149].
Sampling of oak rhizospheres for Phytophthora species showed positive results only at one locality (Pístecký les), where P. plurivora was identified. This oak stand is located in the thermophytic zone and is characterized by balanced soil moisture. The oaks there showed relatively less severe symptoms of decline. Moreover, the lowest number of subcortical insects were captured at this oak plot. In the other studied oak plots, the conditions are probably too dry to be suitable for Phytophthora to survive. Phytophthora plurivora was first identified in the Czech Republic in 2010, but it is very likely that this species was introduced much earlier, and it is now considered to be domesticated in many habitat types [150]. In neighboring countries, there are records of the association between oak decline and the presence of Phytophthora spp. pathogens from earlier times (e.g., in Germany [31] and Austria [28]). Phytophthora plurivora is the most widespread alien representative of the Phytophthora spp. in natural forest stands in the Czech Republic [151,152,153]. This species was isolated quite recently from oak stands in other central European countries, such as Poland [154] and Slovakia [155], where symptoms of sampled oaks included crown transparency, as in our case. Pathogenicity tests using artificial inoculations have demonstrated aggressiveness of P. plurivora in different hosts to occur at different levels [155,156], but the pathogen is generally considered to represent a serious threat to the health of oak forests [152,155].
The role of pathogens of the genus Ophistoma has been unclear for a long time, and in the 1990s this topic was often studied because of their possible transmission by subcortical insects [157,158]. Our results showed positive correlation between the presence of ophiostomatoid fungi and some insect species. Specifically, it was confirmed in three species of buprestid beetles, five of longhorn beetles, and two species of weevils. Nevertheless, the intensity of Ophiostoma spp. presence varied significantly among the localities and also among the trunk parts (basal, middle, crown). This can be explained by possible vectors that transmit the spores to different tree parts, such as S. intricatus in small branches [66]. However, some studies suggested that fungi of the genus Ophiostoma are not common endophytes of oaks and that none of them are closely related to the oak bark beetle (S. intricatus). This microscopic fungus acts as a non-pathogenic or weakly pathogenic species against healthy oaks, but, in the case of these trees being weakened by water deficit, it can manifest itself as a stronger pathogen [3,78,79,88,114]. On the other hand, some studies confirmed the transmission of ophiostomatoid fungal spores by insects, such as by Scolytinae [111,112] or Xiphydria species [73,113]. Physiologically weakened broadleaved trees are often the primary target of wood wasps (Hymenoptera: Xiphydriidae). In Ukraine, the wood wasp Xiphydria longicollis caused mass dieback of oak stands [159]. In the present study, this species was found at three localities but in very low numbers. Only at a very dry site—Nižbor—were a higher number of individuals (11) found, and these were reared from the logs.
5. Conclusions
Several synergistic factors are usually involved in oak decline, as indicated by Figure 5. These factors cyclically cause gradual dieback of additional tree parts and then the whole stand. Abiotic factors (mainly drought) and defoliators are considered to act as stress-inducing factors, whereas others, such as oak powdery mildew, Armillaria spp., bark beetles and borers, are mostly regarded as secondary agents killing already weakened trees. Based on our research data, the individual causes of dieback may not act with the same intensity throughout the decline period. In the case of subcortical insects, fewer species are active in the initial phase of decline than in the phase of severely damaged stands, where species normally classified as rare and even in the highest categories of red lists may be involved to a large extent in the final phase of oak stand decline (e.g., Gasterocercus depressirostris). In this study, the most abundant species of subcortical insect were Xylotrechus antilope, Scolytus intricatus, Xyleborus monographus, Agrilus biguttatus, and A. sulcicollis. Most individuals were captured in window traps at very dry sites.
The presence of Ophiostoma species was confirmed in all six oak stands with different levels of intensity both among stands and among tree parts (basal/middle/crown). Correlation between Ophiostoma presence and subcortical insects was found in three families: Buprestidae, Curculionidae, and Cerambycidae, from which S. intricatus and X. monographus seem to be the most important vectors of these pathogens. Noteworthy is that this correlation was observed only in the crown parts. Another pathogen, P. plurivora, was detected from rhizosphere samples in only one oak stand, indicating a humid environment suitable for this pathogen to survive. Other oak stands, by contrast, are probably too dry or else the pathogen has not yet been introduced there. In this single plot, and despite showing symptoms of decline (crown transparency), the lowest intensity of Ophiostoma presence was recorded as well as the lowest number of subcortical insects. Therefore, P. plurivora might be the cause of the oak decline there.
Climate extremes such as precipitation deficit and high temperatures seem to play a crucial role in the oak decline phenomenon. Together with direct damage effects there might be changes in phenology resulting from global warming. The results comparing the relationship between annual precipitation and oak decline episodes show that oak decline occurs regularly after droughts lasting ca. five years in a row, as recorded in the 1980s in Central Europe. Drought is usually presented as the main or primary cause of decline, but usually additional factors are involved, such as defoliators, bark beetles and wood-boring beetles (S. intricatus, A. biguttatus etc.), pathogens (Ophiostoma spp., Phytophthora spp., Microsphaera alphitoides etc.), or other abiotic factors (mainly frost). Every incident of oak decline should therefore be evaluated individually and precisely and studying all the possible aspects.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13081223/s1, Figure S1: The map of oak decline’s occurrence in Europe and its driving factors [160]; Figure S2: Timescale of oak decline episodes and causal factors in Central European countries (Austria, Czechia, Slovakia) in context of annual precipitation and difference from mean value (1902–2020).; Figure S3: Timescale of oak decline episodes and causal factors in Central European countries (Hungary, Poland, Germany) in context of annual precipitation and difference from mean value (1902–2020).
Author Contributions
Conceptualization, V.P., O.N., M.M. and M.S.; Methodology, V.P., O.N. and M.M.; Formal analysis, O.N., D.B. and V.Z.; Investigation, V.P., O.N., M.M. and M.S.; Resources, M.M.; Data curation, M.M. and O.N.; Writing—original draft preparation, M.M.; Writing—review and editing, M.M., V.P., O.N., M.S. and D.B.; Visualization, O.N. and D.B.; Supervision, V.P. and O.N.; Project administration, V.P. and O.N.; Funding acquisition, V.P. and O.N. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Internal Grant Agency of Faculty of Forestry and Wood Sciences Czech University of Life Sciences in Prague–project GA FLD CZU A20/07 and by the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0118. Infrastructural support and salary for M.M., V.P. and O.N. were also obtained from “EVA4.0”, No. CZ.02.1.01/0.0/0.0/16_019/0000803 financed by OP RDE.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the reported results can be found at the owner—Markéta Macháčová, email address: [email protected].
Acknowledgments
The authors thank to Johana Šindelářová for technical support and resources completion, Barbora Dvořáková and Adéla Juříková for technical support and Jiří Trombik for help with data processing. English Editorial Services is acknowledged for their valuable comments on the revised version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Number of individuals captured in window traps (WT) and rearing cages (RC) at or from the six studied plots.
Table A1.
Number of individuals captured in window traps (WT) and rearing cages (RC) at or from the six studied plots.
| Study Plot | Pístecký les | Česká Skalice | Nižbor | Český Kras | Lásenice | Tvořihráz | Total | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Family | Species | WT | RC | WT | RC | WT | RC | WT | RC | WT | RC | WT | RC | WT | RC |
| Bostrichidae | Bostrichus capucinus | 1 | 1 | 0.0 | |||||||||||
| Lichenophanes varius | 0.8 | 0.2 | 0 | 1.0 | |||||||||||
| Xylopertha retusa | 1 | 1 | 0.0 | ||||||||||||
| Buprestidae | Agrilus angustulus | 1 | 0.3 | 1 | 0.6 | 1 | 3 | 0.9 | |||||||
| Agrilus biguttatus | 3 | 7 | 5.3 | 45 | 1 | 0.6 | 56 | 5.8 | |||||||
| Agrilus graminis | 0.5 | 0.3 | 0.9 | 0 | 1.7 | ||||||||||
| Agrilus hastulifer | 9 | 9 | 0.0 | ||||||||||||
| Agrilus laticornis | 0.0 | 1 | 1 | 0.0 | |||||||||||
| Agrilus obscuricollis | 1 | 1 | 0.0 | ||||||||||||
| Agrilus sulcicollis | 2 | 0.4 | 7 | 5.1 | 42 | 0.4 | 1 | 52 | 5.9 | ||||||
| Coraebus undatus | 1 | 1 | 0.0 | ||||||||||||
| Cerambycidae | Alosterna tabacicolor | 0.6 | 0 | 0.6 | |||||||||||
| Anaglyptus mysticus | 4 | 1 | 2.3 | 5 | 2.3 | ||||||||||
| Anoplodera rufipes | 4 | 0.6 | 4 | 0.6 | |||||||||||
| Cerambyx scopolii | 1 | 1 | 0.0 | ||||||||||||
| Clytus arietis | 1 | 1 | 2 | 0.0 | |||||||||||
| Clytus tropicus | 9 | 1 | 10 | 0.0 | |||||||||||
| Cortodera humeralis | 1 | 10 | 13 | 18 | 42 | 0.0 | |||||||||
| Dinoptera collaris | 0.3 | 2 | 21.4 | 2 | 21.7 | ||||||||||
| Exocentrus adspersus | 0.1 | 0.2 | 2 | 2 | 0.3 | ||||||||||
| Grammoptera abdominalis | 1 | 1 | 1 | 3 | 0.0 | ||||||||||
| Grammoptera ruficornis | 1 | 1 | 0.0 | ||||||||||||
| Grammoptera ustulata | 1 | 1 | 0.0 | ||||||||||||
| Chlorophorus figuratus | 1 | 1 | 0.0 | ||||||||||||
| Leiopus linnei | 2 | 3 | 1.9 | 5 | 0.4 | 1 | 11 | 2.3 | |||||||
| Leiopus nebulosus | 0.4 | 1 | 0.2 | 0.7 | 1 | 1.2 | |||||||||
| Mesosa nebulosa | 1 | 1 | 0.0 | 1 | 3 | 0.0 | |||||||||
| Necydalis major | 1 | 1 | 0.0 | ||||||||||||
| Pachytodes cerambyciformis | 1 | 1 | 0.0 | ||||||||||||
| Pachytodes erraticus | 1 | 1 | 0.0 | ||||||||||||
| Pedostrangalia revestita | 1 | 6 | 7 | 0.0 | |||||||||||
| Phymatodes alni | 1 | 0.2 | 0.6 | 3 | 4 | 0.9 | |||||||||
| Phymatodes pusillus pusillus | 2 | 0.0 | 2 | 0.0 | |||||||||||
| Phymatodes testaceus | 11 | 0.6 | 4 | 17 | 6.9 | 2 | 13 | 47 | 7.5 | ||||||
| Plagionotus detritus | 29 | 0.6 | 7 | 36 | 0.6 | ||||||||||
| Pogonocherus hispidus | 1 | 1 | 0.0 | ||||||||||||
| Pyrrhidium sanguineum | 0.2 | 2 | 1 | 7.7 | 3 | 7.9 | |||||||||
| Rhagium sycophanta | 2 | 2 | 1 | 4 | 9 | 0.0 | |||||||||
| Stenocorus quercus | 7 | 8 | 15 | 0.0 | |||||||||||
| Trichoferus pallidus | 9 | 9 | 0.0 | ||||||||||||
| Xylotrechus antilope | 1.0 | 9 | 13.2 | 589 | 1.0 | 4.4 | 6 | 604 | 19.5 | ||||||
| Curculionidae | Dryocoetes villosus villosus | 2 | 3 | 5 | 0.0 | ||||||||||
| Gasterocercus depressirostris | 1 | 1 | 7.1 | 83 | 46.8 | 25 | 110 | 53.9 | |||||||
| Magdalis exarata | 0.0 | 0 | 0.0 | ||||||||||||
| Magdalis flavicornis | 1 | 1 | 0.0 | ||||||||||||
| Platypus cylindrus | 1 | 1 | 0.0 | ||||||||||||
| Scolytus intricatus | 4 | 2 | 3.8 | 22 | 94.8 | 140 | 114.0 | 1.4 | 18 | 186 | 214.1 | ||||
| Trypodendron domesticum | 1 | 1 | 0.0 | ||||||||||||
| Xyleborinus alni | 1 | 1 | 0.0 | ||||||||||||
| Xyleborus dispar | 1 | 1 | 0.0 | ||||||||||||
| Xyleborus dryographus | 22 | 11.3 | 1 | 23 | 11.3 | ||||||||||
| Xyleborus germanus | 1 | 1 | 0.0 | ||||||||||||
| Xyleborus monographus | 1 | 45 | 4.1 | 6 | 9.2 | 37 | 0.6 | 32.7 | 65 | 1.7 | 154 | 48.3 | |||
| Xyleborus saxeseni | 1 | 2 | 6 | 9 | 0.0 | ||||||||||
| Hymenoptera | Xiphydria longicollis | 1 | 11.0 | 7 | 0.6 | 8 | 11.6 | ||||||||
| Total WT | 18 | 72 | 89 | 1068 | 5 | 203 | 1455 | ||||||||
| Total RC | 0 | 12.7 | 148.9 | 134.0 | 119.7 | 4.5 | 419.9 | ||||||||
RC = sum of individuals numbers calculated from 1 m of crown parts + 1 m of central parts + 1 m of basal parts.
References
- Delatour, C. Les dépérissements de chênes en Europe. Biol. For. 1983, 35, 265–282. [Google Scholar] [CrossRef] [Green Version]
- Schütt, P. Oak decline in central and eastern Europe. A critical review of a little understood phenomenon. In Recent Advances in Studies on Oak Decline, Proceedings of the International Congress, Brindisi, Italy, 13–18 September, 1992; Luisi, N., Vannini, A., Eds.; Dipartimento di Pathologia Vegetale Università Degli Studi; International Union of Forestry Research Organizations (IUFRO): Bari, Italia, 1993; pp. 235–240. [Google Scholar]
- Ragazzi, A.; Vagniluca, S.; Moricca, S. European expansion of oak decline, involved microorganisms and methodological approaches. Phytopathol. Mediterr. 1995, 34, 207–226. [Google Scholar]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Oszako, T. Oak declines in Europe’s forests—History, causes and hypothesis. In Recent Advances on Oak Health in Europe, Proceedings of the International Scientific Conference, Warsaw, Poland, 22–24 November 1999; Oszako, T., Delatour, C., Eds.; Forest Research Institute: Warsaw, Poland, 2000; pp. 11–40. [Google Scholar]
- Thomas, F.M. Recent advances in cause-effect research on oak decline in Europe. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2008, 3, 1–12. [Google Scholar] [CrossRef]
- Millers, I.; Shriner, D.S.; Rizzo, D. History of Hardwood Decline in the Eastern United States; General Technical Report, NE-126; U.S. Department of Agriculture, Forest Service: Broomall, PA, USA, 1989; pp. 1–78. [CrossRef]
- Attarod, P.; Rostami, F.; Dolatshahi, A.; Sadeghi, S.M.M.; Zahedi Amiri, G.; Bayramzadeh, V. Do Changes in Meteorological Parameters and Evapotranspiration Affect Declining Oak Forests of Iran? J. For. Sci. 2016, 62, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Oleksyn, J.; Przybil, K. Oak decline in the Soviet Union—Scale and hypotheses. Eur. J. Plant Pathol. 1987, 17, 321–336. [Google Scholar] [CrossRef]
- Siwecki, R.; Liese, W. Oak Decline in Europe. In Proceedings of the International Symposium, Kórnik, Poland, 15–18 May 1990; Polish Academy of Sciences Institute of Dendrology: Kórnik, Poland, 1991; pp. 1–360. [Google Scholar]
- Luisi, N.; Lerario, P.; Vannini, A. Recent Advances in Studies on Oak Decline. In Proceedings of the International Congress, Brindisi, Italy, 13–18 September 1992; Dipartimento di Pathologia Vegetale Università Degli Studi; International Union of Forestry Research Organizations (IUFRO): Bari, Italia, 1993; pp. 1–541. [Google Scholar]
- Führer, E. Oak decline in Central Europe: A synopsis of hypotheses. In Proceedings of the Population Dynamics, Impacts, and Integrated Management of Forest Defoliating Insects, Banská Štiavnica, Slovakia, 18–23 August 1996; McManus, M.L., Liebhold, A.M., Eds.; USDA Forest Service General Technical Report NE-247. USDA: Washington, DC, USA, 1998; pp. 7–24. [Google Scholar]
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. For. Pathol. 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Manion, P.D. Tree Disease Concepts, 2nd ed.; Prentice Hall: Englewood Cliffs, NJ, USA, 1981. [Google Scholar]
- Čapek, M.; Findo, S.; Brutovský, D. Možnosti biologickej ochrany proti vektorom hromadného hynutia dubov. In Problematika Hynutia Dubov na Slovensku, Vedecké práce VÚLH vo Zvolene; Príroda v Bratislave: Bratislava, Slovakia, 1987; Volume 36, pp. 169–182. [Google Scholar]
- Nienhaus, F. Viren und primitive Prokaryonten in Eichen. Osterr. Forstztg. 1987, 3, 64–65. [Google Scholar]
- Ahrens, U.; Seemüller, E. Detection of mycoplasmalike organisms in declining oaks by polymerase chain reaction. Eur. J. Plant Pathol. 1994, 24, 55–63. [Google Scholar] [CrossRef]
- Šafar, J. Problem fizioloških, ekoloških i ekonomskih karakteristika kasnoga i ranog hrasta lužnjaka. Sumar. List 1966, 90, 503–515. [Google Scholar]
- Dantec, C.F.; Ducasse, H.; Capdevielle, X.; Fabreguettes, O.; Delzon, S.; Desprez-Loustau, M.-L. Escape of spring frost and disease through phenological variations in oak populations along elevation gradients. J. Ecol. 2015, 103, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Puchałka, R.; Koprowski, M.; Gričar, J.; Przybylak, R. Does tree-ring formation follow leaf phenology in Pedunculate oak (Quercus robur L.)? Eur. J. For. Res. 2017, 136, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Utkina, I.A.; Rubtsov, V.V. Studies of Phenological Forms of Pedunculate Oak. Contemp. Probl. Ecol. 2017, 10, 804–811. [Google Scholar] [CrossRef]
- Wesołowski, T.; Rowiński, P. Late leaf development in pedunculate oak (Quercus robur): An antiherbivore defence? Scand. J. For. Res. 2008, 23, 386–394. [Google Scholar] [CrossRef]
- Georgevitch, P. Armillaria mellea (Vahl) Quel. cause du dessechement forets de chene en Yougoslavie. Compte-Rendus Achademic Sci. 1926, 182, 289–491. [Google Scholar]
- Georgevitch, P. Ceratostomella quercus n. sp., Ein parasite der slawonischen Eichen. Biol. Gen. 1927, 3, 245–252. [Google Scholar]
- Marcu, G. Ursachen des Eichensterbens in Rumänien und Gegenmassnahmen. Österr. Forstz 1987, 98, 53–54. [Google Scholar]
- Brasier, C.M. Phytophthora cinnamomi as a contributory factor on European oak declines. In Recent Advances in Studies on Oak Decline, Proceedings of the International Congress, Brindisi, Italy, 13–18 September 1992; Luisi, N., Vannini, A., Eds.; Dipartimento di Patholog ia Vegetale Università Degli Studi; International Union of Forestry Research Organizations (IUFRO): Bari, Italia, 1993; pp. 49–58. [Google Scholar]
- Andersson, M.; Milberg, P.; Bergman, K.O. Low pre-death growth rates of oak (Quercus robur L.)—Is oak death a long-term process induced by dry years? Ann. For. Sci. 2011, 68, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Balci, Y.; Halmschlager, E. Incidence of Phytophthora species in oak forests in Austria and their possible involvement in oak decline. For. Pathol 2003, 33, 157–174. [Google Scholar] [CrossRef]
- Hansen, E.; Delatour, C. Phytophthora species in oak forests of north-east France. Ann. For. Sci. 1999, 56, 539–547. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. Plant Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Barzanti, G.P.; Bianco, M.C.; Ragazzi, A.; Capretti, P.; Paoletti, E.; Luisi, N.; Anselmi, N.; Vannini, A. Occurrence of Phytophthora species in oak stands in Italy and their association with declining oak trees. For. Pathol 2002, 32, 19–28. [Google Scholar] [CrossRef]
- Biosca, E.G.; Gonzalez, R.; Lopez-Lopez, M.J.; Soria, S.; Monton, C.; Perez-Laorga, E.; Lopez, M.M. Isolation and characterization of Brenneria quercina, causal agent for bark canker and drippy nut of Quercus spp. in Spain. Phytopathology 2003, 93, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.; Jeger, M.; Kirk, S.; Williams, D.; Xu, X.; Pautasso, M.; Denman, S. Acute oak decline and Agrilus biguttatus: The co-occurrence of stem bleeding and D-shaped emergence holes in Great Britain. Forests 2017, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gentilesca, T.; Oliva, J.; Redondo, M.A.; Ripullone, F. Drought and Phytophthora are associated with the decline of oak species in southern Italy. Front. Plant Sci. 2018, 9, 1595. [Google Scholar] [CrossRef]
- Conte, A.L.; Di Pietro, R.; Iamonico, D.; Di Marzio, P.; Cillis, G.; Lucia, D.; Fortini, P. Oak decline in the Mediterranean Basin: A Study Case from the Southern Apennines (Italy). Plant Sociol. 2019, 56, 69–80. [Google Scholar] [CrossRef]
- Correia, A.C.; Galla, A.; Nunes, A.; Pereira, J.S. Ecological interactions between cork oak (Quercus suber L.) and stone pine (Pinus pinea L.): Results from a pot experiment. Forests 2019, 9, 534. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.; Brown, N.; Kirk, S.; Jeger, M.; Webber, J. A description of the symptoms of acute oak decline in Britain and a comparative review on causes of similar disorders on oak in Europe. Forestry 2014, 87, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Doležal, J.; Mazůrek, P.; Klimešová, J. Oak decline in southern Moravia: The association between climate change and early and late wood formation in oaks. Preslia 2010, 82, 289–306. [Google Scholar]
- Donaubauer, E. Auftreten von Krankheiten und Schädlingen der Eiche und ihr Bezug zum Eichensterben. Osterreichische Forstztg. 1987, 3, 46. [Google Scholar]
- Frisullo, S.; Lima, G.; Magnano di San Lio, G.; Camele, I.; Melissano, L.; Puglisi, I.; Pane, A.; Agosteo, G.E.; Prudente, L.; Cacciola, S.O. Phytophthora cinnamomi involved in the decline of holm oak (Quercus ilex) stands in southern Italy. For. Sci. 2018, 64, 290–298. [Google Scholar] [CrossRef]
- Gibbs, J.N.; Greig, B.J.W. Biotic and abiotic factors affecting the dying back of pedunculate oak Quercus robur L. Forestry 1997, 70, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, G.; Blank, R. Winterfrost, Kahlfraß und Pracht Käferbefall als Faktoren im Ursachenkomplex des Eichensterbens in Norddeutschland. Forst Und Holz 1992, 47, 443–452. [Google Scholar]
- Jurc, D. Oak Decline and the Status of Ophiostoma spp. on Oak in Europe (Yugoslavia). Bull. OEPP/EPPO Bull. 1990, 20, 419–422. [Google Scholar]
- Jurc, M.; Bojović, S.; Komjanc, B.; Krč, J. Xylophagous entomofauna in branches of oak (Quercus spp.) and its significance for oak health in the Karst region of Slovenia. Biologia 2009, 64, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Klepáč, D. Les forêts de Chêne en Slavonie. Rev. For. Fr. 1981, 33, 86–106. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, T. Oak Decline and the Status of Ophiostoma spp. on Oak in Europe (Poland). Bull. OEPP/EPPO Bull. 1990, 20, 417–418. [Google Scholar]
- Losseau, J.; Jonard, M.; Vincke, C. Pedunculate oak decline in southern Belgium: A long-term process highlighting the complex interplay among drought, winter frost, biotic attacks, and masting. Can. J. For. Res. 2020, 50, 380–389. [Google Scholar] [CrossRef]
- Moraal, L.G.; Hilszczánski, J. The oak buprestid beetle, Agrilus biguttatus (F.) (Col., Buprestidae), a recent factor in oak decline in Europe. Anz. Für Schädlingskunde 2000, 73, 134–138. [Google Scholar]
- Mora-Sala, B.; Berbegal, M.; Abad-Campos, P. The use of qPCR reveals a high frequency of Phytophthora quercina in two Spanish holm oak areas. Forests 2018, 9, 697. [Google Scholar] [CrossRef] [Green Version]
- Oosterbaan, A. Oak Decline and the Status of Ophiostoma spp. on Oak in Europe (Netherlands). Bull. OEPP/EPPO Bull. 1990, 20, 414–417. [Google Scholar]
- Redondo, M.Á.; Thomsen, I.M.; Oliva, J. First report of Phytophthora uniformis and P. plurivora causing stem cankers on Alnus glutinosa in Denmark. Plant Dis. 2017, 101, 512. [Google Scholar] [CrossRef]
- Rossnev, B.; Petkov, P.; Georgiev, D. Importance and character of the tracheomycotic disease in the oak forests of Bulgaria. Nauka Za Gorata 1994, 31, 49–54. [Google Scholar]
- Ruffner, B.; Schneider, S.; Meyer, J.B.; Queloz, V.; Rigling, D. First report of acute oak decline disease of native and nonnative oaks in Switzerland. New Dis. Rep. 2020, 41, 18. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Gómez, F.J.; Pérez-de-Luque, A.; Navarro-Cerrillo, R.M. The involvement of Phytophthora root rot and drought stress in holm oak decline: From ecophysiology to microbiome influence. Curr. For. Rep. 2019, 5, 251–266. [Google Scholar] [CrossRef]
- Selochnik, N.N.; Pashenova, N.V.; Sidorov, E.; Wingfield, M.J.; Linnakoski, R. Ophiostomatoid fungi and their roles in Quercus robur die-back in Tellermann forest, Russia. Silva Fenn. 2015, 49, 16. [Google Scholar] [CrossRef] [Green Version]
- Siwecki, R.; Ufnalski, K. Review of oak stand decline with special reference to the role of drought in Poland. Eur. J. Plant Pathol. 1998, 28, 99–112. [Google Scholar] [CrossRef]
- Sohar, K.; Helama, S.; Läänelaid, A.; Raisio, J.; Tuomenvirta, H. Oak decline in a southern Finnish forest as affected by a drought sequence. Geochronometria 2014, 41, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Sonesson, K.; Drobyshev, I. Recent advances on oak decline in southern Sweden. Ecol. Bull. 2010, 53, 197–208. [Google Scholar]
- Spaič, I. O sušenju hrastika. Šumarski List. 1974, 78, 273–284. [Google Scholar]
- Stojanović, D.; Levanič, T.; Matović, B.; Bravo-Oviedo, A. Climate change impact on a mixed lowland oak stand in Serbia. Ann. Silvic. Res. 2015, 39, 94–99. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Celma, L.; Ruņģis, D.E.; Bokuma, G. First report on Brenneria goodwinii and Gibbsiella quercinecans bacteria, detected on weaken oak trees in Poland. Balt. For. 2021, 27, 1. [Google Scholar] [CrossRef]
- Tomiczek, C. Oak decline in Austria and Europe. Arboric. J. 1993, 19, 71–73. [Google Scholar] [CrossRef]
- Vannini, A.; Luisi, N. Oak Decline and the Status of Ophiostoma spp. on Oak in Europe (Italy). Bull. OEPP/EPPO Bull. 1990, 20, 413–414. [Google Scholar]
- Yde-Andersen, A. Oak Decline and the Status of Ophiostoma spp. on Oak in Europe. Bull. OEPP/EPPO Bull. 1990, 20, 405–422. [Google Scholar]
- Zumr, V. Hmyz jako potenciální vektor tracheomykózního odumírání dubů. In Aktuální Problémy Ochrany Dřevin: Sborník Referátů z Odb. Semináře; Čížková, D., Švecová, M., Eds.; Ministerstvo Životního Prostředí ČR: Prague, Czech Republic, 1995; Volume 1, pp. 89–92. [Google Scholar]
- Petrescu, M. Dieback of oak in Romania. Eur. J. For. Path. 1974, 4, 222–227. [Google Scholar] [CrossRef]
- Oszako, T. Protection of Forests Against Pest Insects and Diseases. European Oak Decline Study Case; Forests Research Institute (IBL): Warsaw, Poland, 2004; ISBN 83-87-647-37-3. [Google Scholar]
- Varga, F. Disease and dying of trees in Hungarian oak pedunculate stands. Osterr. Forstztg. 1987, 3, 57–58. [Google Scholar]
- Yakoylev, A.I.; Yakovlev, S.A. Oak decline in the Middle Povolzhje region of Russia. Recent Advances on Oak Health in Europe. In Proceedings of the International Scientific Conference, Warsaw, Poland, 22–24 November 1999; Oszako, T., Delatour, C., Eds.; Forest Research Institute: Warsaw, Poland, 2000; pp. 73–82. [Google Scholar]
- Uroševič, B. Tracheomycotic diseases in oak. Commun. Inst. For. Cechoslov. 1957, 13, 85–100. [Google Scholar]
- Igmandy, Z.; Pagony, H.; Szoutagh, P.; Varga, F. A report on the mortality in national sessile oak stands 1978–1983. Acta Fac. For. 1985; 5–16. [Google Scholar]
- Příhoda, A. Hynutí dubů ve středních Čechách. Bohemia Cent. 1990, 19, 81–90. [Google Scholar]
- Jančařík, V. Tracheomykózní onemocnění lesních dřevin—Realita a hrozba. In Proceedings of the Aktuální Problémy Ochrany Dřevin. Sborník Referátů z Odb. Semináře, Prachatice, Czech Republic, 3–4 October 1995; Čížková, D., Švecová, M., Eds.; Ministerstvo Životního Prostředí ČR: Prague, Czech Republic, 1995; Volume 2, pp. 5–17. [Google Scholar]
- Švecová, M.; Skalický, V. Problematika hub řádu Ophiostomatales a jejich výskyt v českých zemích. In Proceedings of the Ophiostomatales—Výsledky Současného Taxonomického a Fytopatologického Výzkumu. Sborník Referátů, Prague, Czech Republic, 29 May 1991; Holubová, V., Prášil, K., Eds.; Československá Vědecká Společnost pro Mykologii při ČSAV: Prague, Czech Republic, 1992; pp. 19–29. [Google Scholar]
- Kubátová, A.; Prášil, K. Ophiostomatální a další mikroskopické houby lesních dřevin s příznaky tracheomykózního onemocnění. In Proceedings of the Aktuální Problémy Ochrany Dřevin. Sborník Referátů, Prachatice, Czech Republic, 3–4 October 1995; Čížková, D., Švecová, M., Eds.; Ministerstvo Životního Prostředí ČR: Prague, Czech Republic, 1995; Volume 2, pp. 89–92. [Google Scholar]
- Kowalski, T.; Bartnik, C. Ceratocystis Species on Quercus robur with Oak Decline Symptoms in Southern Poland. Bull. OEPP/EPPO Bull. 1990, 20, 221–228. [Google Scholar] [CrossRef]
- Kowalski, T.; Butin, H. Taxonomie bekannter und neuer Ceratocystis-Arten an Eiche (Quercus robur L.). J. Phytopathol. 1989, 124, 236–248. [Google Scholar] [CrossRef]
- Kowalski, T.; Domanski, S. Preliminary results of artificial inoculations of Quercus robur L. with different species of Ceratocystis. Oak Decline in Europe. In Proceedings of the International Symposium, Kórnik, Poland, 15–18 May 1990; Siwecki, R., Liese, W., Eds.; Polish Academy of Sciences Institute of Dendrology: Kórnik, Poland, 1991; pp. 93–104. [Google Scholar]
- Rohde, T. Beitrag zur Kenntnis Einer Krebsartigen Eichenkrankheit und Ihrer Pilzflora. Ph.D. Thesis, M. &. H. Schaper, Hannover, Germany, 1936. [Google Scholar]
- Leontovyč, R.; Gontková, E. Napadnutie, náchylnosť a odolnosť jednotlivých druhov k hromadnému hynutiu dubov. In Problematika Hynutia Dubov na Slovensku, Vedecké Práce VÚLH vo Zvolene; Príroda v Bratislave: Bratislava, Slovakia, 1987; Volume 36, pp. 211–232. [Google Scholar]
- Beranová, J. Umělá infekce dubových sazenic houbovými původci tracheomykózního onemocnění. Zprávy Lesn. Výzkumu 1989, 3, 38–41. [Google Scholar]
- Oszako, T. Influence of water stress, defoliation and inoculation with Ophiostoma querci on pedunculate oak seedlings. Folia For. Pol. A For. 1997, 39, 5–15. [Google Scholar]
- Novotný, D. Ophiostomatales a lesní dřeviny (zvláště dub). In Proceedings of the Houby a les, Sborník z Konference s Mezinárodní Účastí, Brno, Czech Republic, 3–4 June 1999; MZLU: Brno, Czech Republic, 1999; pp. 89–94. [Google Scholar]
- Somogyi, Z. Oak decline in Hungary: Case study. Recent Advances on Oak Health in Europe. In Proceedings of the International Scientific Conference, Warsaw, Poland, 22–24 November 1999; Oszako, T., Delatour, C., Eds.; Forest Research Institute: Warsaw, Poland, 2000; pp. 73–82, 91–103. [Google Scholar]
- Novotný, D. Příspěvek k mykoflóře dubů s tracheomykózními příznaky. In Proceedings of the Aktuální problémy ochrany dřevin. Sborník referátů, Prachatice, Czech Republic, 3–4 October 1995; Čížková, D., Švecová, M., Eds.; Ministerstvo Životního Prostředí ČR: Prague, Czech Republic, 1995; Volume 2, pp. 52–64. [Google Scholar]
- Novotný, D. Endofyty a ophiostomatální houby ve vztahu k listnatým dřevinám. Zprávy Lesn. Výzkumu 2003, 48, 126–129. [Google Scholar]
- Przybyl, K. On the pathogenicity of Ophiostoma piceae. Oak decline in Europe. In Proceedings of the International Symposium, Kórnik, Poland, 15–18 May 1990; Siwecki, R., Liese, W., Eds.; Polish Academy of Sciences Institute of Dendrology: Kórnik, Poland, 1991; pp. 83–89. [Google Scholar]
- Malloch, D.; Blackwell, M. Dispersal biology of the ophiostomatoid fungi. In Ceratocystis and Ophiostoma: Taxonomy, Ecology, and Pathogenicity; Wingfield, M.J., Seifert, K.A., Webber, J.F., Eds.; American Phytopathological Society Press: St Paul, MN, USA, 1993; pp. 195–206. ISBN 978-0890541562. [Google Scholar]
- Wingfield, M.J.; Barnes, I.; De Beer, Z.W.; Roux, J.; Wingfield, B.D.; Taerum, S.J. Novel associations between ophiostomatoid fungi, insects and tree hosts: Current status—Future prospects. Biol. Invasions 2017, 19, 3215–3228. [Google Scholar] [CrossRef]
- Fassatiová, O. Houby v chodbách kůrovců. Česká Mykol. 1954, 8, 138–143. [Google Scholar]
- Heško, J. Diagnostické znaky tracheomykóznych dubov. Les 1985, 9, 391–394. [Google Scholar]
- Příhoda, A.; Jančařík, V. Tracheomykózy dubů. In TEI—Bulletin Technickoekonomických Informací 1; VÚLHM: Jíloviště-Strnady, Prague, Czech Republic, 1988; pp. 1–8. [Google Scholar]
- Kirisits, T. Studies on the Association of Ophiostomatoid Fungi with Bark Beetles in Austria with Special Emphasis on Ips typographus and Ips cembrae and Their Associated Fungi Ceratocystis polonica and Ceratocystis laricicola. Ph.D. Thesis, Universität für Bodenkultur (BOKU), Wien, Austria, 2001. [Google Scholar]
- Kirschner, R. Diversity of filamentous fungi in bark beetle galleries in central Europe. In Trichomycetes and Other Fungal Groups, 1st ed.; Lichtwardt Commemoration Volume; Robert, W., Misra, J.K., Horn, B.W., Eds.; Science Publishers, Inc.: Plymouth, UK, 2001; pp. 175–196. ISBN 9781578081325. [Google Scholar]
- Kotýnková-Sychrová, E. Mykoflóra chodeb kůrovců v Československu. Česká Mykol. 1966, 20, 45–53. [Google Scholar]
- Jankowiak, R. Fungi associated with Ips typographus on Picea abies in Southern Poland and their succession into the phloem and sapwood of beetle-infested trees and logs. For. Pathol. 2005, 35, 37–55. [Google Scholar] [CrossRef]
- Jankovský, L.; Mrkva, R. Přenos patogenů vaskulárních pletiv na smrku lýkožroutem smrkovým Ips typographus L. In Proceedings of the Les, Drevo, Životné Prostredie, Zbornik Referátov, Zvolen, Slovakia, 8–11 September 1997; Križová, E., Kodrík, M., Eds.; Technická Univerzita vo Zvolene: Zvolen, Slovakia, 1997; pp. 209–218. [Google Scholar]
- Żółciak, A. Occurrence of Armillaria species in oak stands in Poland. In Recent Advances on Oak Health in Europe, Proceedings of the International Scientific Conference, Warsaw, Poland, 22–24 November 1999; Oszako, T., Delatour, C., Eds.; Forest Research Institute: Warsaw, Poland, 2000; pp. 243–248. [Google Scholar]
- Kunca, A.; Leontovyč, R. Dieback of Broadleaved Trees in Slovakia. In Possible Limitation of Decline Phenomena in Broadleaved Stands; Oszako, T., Woodward, S.W., Eds.; Forest Research Institute: Warsaw, Poland, 2006; pp. 29–33. ISBN 83-87647-56-X. [Google Scholar]
- Szewczyk, W.; Kwaśna, H.; Behnke-Borowczyk, J. Armillaria Population in Flood-Plain Forest of Natural Pedunculate Oak Showing Oak Decline. Pol. J. Environ. Stud. 2016, 25, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Mrkva, R. Praskliny kůry suchem jako poškození a dosud neznámý symptom chřadnutí listnatých dřevin. Zprávy Lesn. Výzkumu 2003, 48, 136–142. [Google Scholar]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil—And airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, P.; Bader, M.K.-F.; Burgess, T.; Hardy, G.; Williams, N. Global biogeography and invasion risk of the plant pathogen genus Phytophthora. Environ. Sci. Policy 2019, 101, 175–182. [Google Scholar] [CrossRef]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Anguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Corcobado, T.; Solla, A.; Madeira, A.M.; Moreno, G. Combined effects of soil properties and Phytophthora cinnamomi infections on Quercus ilex decline. Plant Soil 2013, 373, 403–413. [Google Scholar] [CrossRef]
- Szabó, I.; Lakatos, F.; Sipos, G. Occurrence of soilborne Phytophthora species in declining broadleaf forest in Hungary. Eur. J. Plant Pathol. 2013, 137, 159–168. [Google Scholar] [CrossRef]
- Southwood, T.R.E. The number of species of insect associated with various trees. J. Anim. Ecol. 1961, 30, 1–8. [Google Scholar] [CrossRef]
- Knížek, M. Polník dvojtečný Agrilus biguttatus (Fabricius, 1777) (a ostatní krasci rodu Agrilus na dubech). Lesn. Práce 2011, 90, 1–4. [Google Scholar]
- Liška, J. Role Listožravého Hmyzu v Procesu Odumírání; Sub-final report to the project no. 329-91-9106; VÚLHM: Jíloviště-Strnady, Prague, Czech Republic, 1995. [Google Scholar]
- Jankovský, L.; Novotný, D.; Mrkva, R. Doprovodná mykoflóra. Ips typographus a ranové reakce smrku na umělou inokulaci imágy lýkožrouta smrkového. In Ochrana lesa a lesnická fytopatológia, Proceedings of the Conference, Zvolen, Sielnica, Slovakia, 4–6 September 2000; Hlaváč, P., Reinprecht, L., Gáper, J., Eds.; Technická Universita vo Zvolene: Zvolen, Slovakia, 2001. [Google Scholar]
- Kubátová, A.; Novotný, D.; Prášil, K. Microscopic fungi associated with oak bark beetle (Scolytus intricatus) in the Czech Rebublic. In IMC 7 Book of Abstracts, Proceedings of the 7th International Mycological Congress, Oslo, Norway, 11–17 August 2002; Universitat für Bodenkultur: Vienna, Austria; pp. 1–300.
- Šrůtka, P.; Pažoutová, S.; Kolařík, M. Daldinia decipiens and Entonaema cinnabarina as fungal symbionts of Xiphydria wood wasps. Mycol. Res. 2007, 111, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Soukup, F. Odumírání Dubů a Dalších Dřevin v Lesních Porostech s Příznaky Tracheomykózního Onemocnění; Final report to the project no. 329-91-9106; VÚLHM: Jíloviště-Strnady, Prague, Czech Republic, 1995. [Google Scholar]
- Sallé, A.; Nageleisen, L.M.; Lieutier, F. Bark and wood boring insects involved in oak declines in Europe: Current knowledge and future prospects in a context of climate change. For. Ecol. Manag. 2014, 328, 79–93. [Google Scholar] [CrossRef]
- Czech Hydrometeorological Institute (CHMI). Historical Data—Meteorology and Climatology. Daily Data. Available online: https://www.chmi.cz/historicka-data/pocasi/denni-data/Denni-data-dle-z.-123-1998-Sb (accessed on 1 June 2022).
- Harris, I.C.; Jones, P.D.; Osborn, T. CRU TS4.05: Climatic Research Unit (CRU) Time-Series (TS) Version 4.05 of High-Resolution Gridded Data of Month-by-Month Variation in Climate (January 1901–December 2020); University of East Anglia Climatic Research Unit; NERC EDS Centre for Environmental Data Analysis: Norwich, UK, 2021. [Google Scholar]
- Gregus, L.; Heško, J.; Hruška, J.; Jančařík, V.; Mráček, Z.; Pokorný, J.; Potoček, J.; Příhoda, A.; Zumr, V. Záchrana Jilmů—Metodická Příručka č. 9; ÚV Českého Svazu Ochránců Přírody: Prague, Czech Republic, 1986; pp. 1–46. [Google Scholar]
- Jung, T.; Blaschke, M. Phytophthora root and collar rot of alders in Bavaria: Distribution, modes of spread and possible management strategies. Plant Pathol. 2004, 53, 197–208. [Google Scholar] [CrossRef]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; The American Phytopathological Society Press: St Paul, MN, USA, 1996; ISBN 0-89054-212-0. [Google Scholar]
- Grünwald, N.J.; Martin, F.N.; Larsen, M.M.S.; Press, C.M.; Coffey, M.D.; Hansen, E.M.; Parke, J.L. Phytophthora-ID.org: A sequence-based Phytophthora identification tool. Plant Dis. 2011, 95, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vansteenkiste, D.; Tirry, L.; Van Acker, J.; Stevens, M. Predispositions and symptoms of Agrilus borer attack in declining oak trees. Ann. For. Sci. 2004, 61, 815–823. [Google Scholar] [CrossRef] [Green Version]
- Pilcher, J.R.; Gray, B. The relationships between oak tree growth and climate in Britain. J. Ecol. 1982, 70, 297–304. [Google Scholar] [CrossRef]
- Thomas, F.M.; Hartmann, G. Tree rooting patterns and soil water relations of healthy 4 and damaged stands of mature oak (Quercus robur L. and Quercus petraea [Matt.] Liebl.). Plant Soil 1998, 203, 145–158. [Google Scholar] [CrossRef]
- Bréda, N. Water shortage as a key factor in the case of the oak dieback in the Harth Forest (Alsatian plain, France) as demonstrated by dendroecological and ecophysiological study. In Recent Advances on Oak Health in Europe, Proceedings of the International Scientific Conference, Warsaw, Poland, 22–24 November 1999; Oszako, T., Delatour, C., Eds.; Forest Research Institute: Warsaw, Poland, 2000; pp. 157–159. [Google Scholar]
- Wermelinger, B.; Rigling, A.; Schneider, M.D.; Dobbertin, M. Assessing the role of bark-and wood-boring insects in the decline of Scots pine (Pinus sylvestris) in the Swiss Rhone valley. Ecol. Entomol. 2008, 33, 239–249. [Google Scholar] [CrossRef]
- Leontovyč, R.; Kunca, A. Health condition of oak stands in Slovakia. In Recent Advances on Oak Health in Europe, Proceedings of the International Scientific Conference, Warsaw, Poland, 22–24 November 1999; Oszako, T., Delatour, C., Eds.; Forest Research Institute: Warsaw, Poland, 2000; pp. 105–106. [Google Scholar]
- Haavik, L.J.; Billings, S.A.; Guldin, J.M.; Stephen, F.M. Emergent insects, pathogens and drought shape changing patterns in oak decline in North America and Europe. For. Ecol. Manag. 2015, 354, 190–205. [Google Scholar] [CrossRef]
- Leontovyč, R. Výskyt rodu Ophiostoma na listnatých dřevinách v súčasných zmenených ekologických podmienkach Slovenska. In Proceedings of the Ophiostomatales—Výsledky současného taxonomického a fytopatologického výzkumu. Sborník referátů, Praha, Czech Republic, 29 May 1991; Holubová, V., Prášil, K., Eds.; Československá Vědecká Společnost pro Mykologii při ČSAV: Prague, Czech Republic, 1992; pp. 30–35. [Google Scholar]
- McIntyre, G.A.; Jacobi, W.R.; Ramaley, A.W. Factors affecting cytospora canker occurrence on aspen. J. Arboric. 1996, 22, 229–233. [Google Scholar] [CrossRef]
- Haavik, L.J.; Stephen, F.M. Factors that affect compartmentalization and wound closure of Quercus rubra infested with Enaphalodes rufulus. Agric. For. Entomol 2011, 13, 291–300. [Google Scholar] [CrossRef]
- Sierpinski, A.; Hilszczański, J. Agrilus spp. The Main Factor of Oak Decline In Poland. In Possible Limitation of Decline Phenomena in Broadleaved Stands; Oszako, T., Woodward, S.W., Eds.; Forest Research Institute: Warsaw, Poland, 2006; pp. 159–161. ISBN 83-87647-56-X. [Google Scholar]
- Drobyshev, I.; Linderson, H.; Sonesson, K. Temporal mortality pattern of pedunculate oaks in southern Sweden. Dendrochronologia 2007, 24, 97–108. [Google Scholar] [CrossRef]
- Evans, H.F.; Moraal, L.; Pajares, J.A. Biology, ecology and economic importance of Buprestidae and Cerambycidae. In European Bark and Wood Boring Insects in Living Trees: A Synthesis; Lieutier, F., Day, K., Battisti, A., Gregoire, J.C., Evans, H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 447–474. ISBN 978-1-4020-2240-1. [Google Scholar]
- Brown, N.; Inward, D.J.G.; Jeger, M.; Denman, S. A Review of Agrilus Biguttatus in UK Forests and Its Relationship with Acute Oak Decline. Forestry 2015, 88, 53–63. [Google Scholar] [CrossRef]
- Denman, S.; Kirk, S.A.; Webber, J.F. Managing Acute Oak Decline; Forestry Commission: Farnham, UK, 2010; 6p, ISBN 978-0-85538-802-7. [Google Scholar]
- Knížek, M. Bělokaz dubový. Scolytus intricatus (Ratzeburg). Lesn. Práce 2002, 81, 12, Annex I–Annex IV. [Google Scholar]
- Patočka, J.; Novotný, J. Účasť hmyzu na hromadnom hynutí dubov na Slovensku. In Problematika Hynutia Dubov na Slovensku, Vedecké Práce VÚLH vo Zvolene; Príroda v Bratislave: Bratislava, Slovakia, 1987; Volume 36, pp. 59–90. [Google Scholar]
- Gogola, E.; Chovanec, D. Podkornik Dubový a Tracheomykóza Dubov; Videopress MON: Bratislava, Slovakia, 1987. [Google Scholar]
- Farkač, J.; Král, D.; Škorpík, M. Červený Seznam Ohrožených Druhů České Republiky—Bezobratlí; Agentura ochrany přírody a krajiny ČR: Prague, Czech Republic, 2005; ISBN 80-86064-96-4. [Google Scholar]
- Hejda, R.; Farkač, J.; Chobot, K. Červený Seznam Ohrožených Druhů České republiky—Bezobratlí; Agentura ochrany přírody a krajiny ČR: Prague, Czech Republic, 2017; ISBN 978-80-88076-53-7. [Google Scholar]
- Głowaciński, Z.; Nowacki, J. Polska Czerwona Księga Zwierząt—Bezkręgowce; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2004; ISBN 83-88934-60-0. [Google Scholar]
- Binot, M.; Bless, R.; Boye, P.; Gruttke, H.; Pretscher, P. Rote Liste Gefährdeter Tiere Deutschlands; Bundesamt für Naturschutz: Leipzig, Germany, 1998; ISBN 3-89624-110-9. [Google Scholar]
- Denman, S.; Webber, J. Oak declines: New definitions and new episodes in Britain. Q. J. For. 2009, 103, 285–290. [Google Scholar]
- Brady, C.; Denman, S.; Kirk, S.; Venter, S.; Rodriguez-Palenzuela, P.; Coutinho, T. Description of Gibbsiella quercinecans gen. nov., sp. nov., associated with acute oak decline. Syst. Appl. Microbiol. 2010, 33, 444–450. [Google Scholar] [CrossRef]
- Denman, S.; Brady, C.; Kirk, S.; Cleenwerck, I.; Venter, S.; Countinho, T.; De Vos, P. Brenneria goodwinii sp. Nov., Associated with Acute Oak Decline in the UK. Int. J. Syst. Evol. Microbiol. 2012, 62, 2451–2456. [Google Scholar] [CrossRef]
- Szontagh, P. Die Rolle der Insektengradation im Verlauf der Krankheiten von Trauben-eichenbeständen. Osterreichische Forstztg. 1987, 98, 65–66. [Google Scholar]
- Schopf, A. Rinden-und holzbriitende Schädlinge an erkrankten Eichen in Österreich. Osterreichische Forstztg. 1992, 103, 33–35. [Google Scholar]
- Mrázková, M.; Černý, K.; Tomšovský, M.; Holub, V.; Strnadová, V.; Zlatohlávek, A.; Gabrielová, S. First Report of Root Rot of Pedunculate Oak and Other Forest Tree Species Caused by Phytophthora plurivora in the Czech Republic. Plant Dis. 2010, 94, 2. [Google Scholar] [CrossRef] [PubMed]
- Černý, K.; Tomšovský, M.; Mrázková, M.; Strnadová, V. The present state of knowledge of Phytophthora spp. Diversity in forest and ornamental woody plants in the Czech Republic. N. Z. J. For. Sci. 2011, 41, S75–S82. [Google Scholar]
- Mrázková, M.; Černý, K.; Strnadová, V.; Filipová, N. Identifikace Symptomů Napadení Dřevin a Okrasných Rostlin Patogeny z rodu Phytophthora de Bary. Certifikovaná metodika; VÚKOZ: Průhonice, Czech Republic, 2012. [Google Scholar]
- Mrázková, M.; Černý, K.; Tomšovský, M.; Strnadová, V.; Gregorová, B.; Holub, V.; Pánek, M.; Havrdová, L.; Hejná, M. Occurrence of Phytophthora multivora and Phytophthora plurivora in the Czech Republic. Plant Protect. Sci. 2013, 49, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Jankowiak, R.; Stępniewska, H.; Bilański, P.; Kolařík, M. Occurrence of Phytophthora Plurivora and Other Phytophthora Species in Oak Forests of Southern Poland and Their Association with Site Conditions and the Health Status of Trees. Folia Microbiol. 2014, 59, 531–542. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Sikora, K.; Galko, J.; Kunca, A.; Milenković, I. Isolation and Pathogenicity of Phytophthora Species from Sessile Oak (Quercus Petraea (Matt.) Liebl.) Stands in Slovakia. For. Pathol. 2020, 50, 330. [Google Scholar] [CrossRef]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Nowakovska, J.A.; Oszako, T.; Sikora, K.; Corcobado, T.; Jung, T. Isolation and Pathogenicity of Phytophthora Species from Poplar Plantations in Serbia. Forests 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Jančařík, V. Fytopatologické problémy působené houbami rodu Ophiostoma a možnosti ochrany. In Proceedings of the Ophiostomatales—Výsledky Současného Taxonomického a Fytopatologického Výzkumu. Sborník Referátů, Prague, Czech Republic, 29 May 1991; Holubová, V., Prášil, K., Eds.; Československá Vědecká Společnost pro Mykologii při ČSAV: Prague, Czech Republic, 1992; pp. 95–110. [Google Scholar]
- Čížková, D.; Švecová, M. Gradual oaks decline with tracheomycotic symptoms in green areas outside forests in Southern Bohemia. In Aktuální problémy ochrany dřevin. Sborník referátů, Prachatice, Czech Republic, 3–4 October 1995; Čížková, D., Švecová, M., Eds.; Ministerstvo Životního Prostředí ČR: Prague, Czech Republic, 1995; Volume 1, pp. 18–23. [Google Scholar]
- Dominik, J.; Starczyk, J.R. Owady Niszczące Drewno; Panstwowe Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 1988. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef]
Figure 1.
Timescale of oak decline episodes in European countries (according to [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73]).

Figure 2.
Studied oak plots with declining trees: 1 = Pístecký les, 2 = Česká Skalice, 3 = Nižbor, 4 = Český kras, 5 = Lásenice, 6 = Tvořihráz.
Figure 2.
Studied oak plots with declining trees: 1 = Pístecký les, 2 = Česká Skalice, 3 = Nižbor, 4 = Český kras, 5 = Lásenice, 6 = Tvořihráz.

Figure 3.
Subcortical insects reared from rearing cages. The individual species are grouped into families. The y-axis represents the logarithmic number of records of each species. Boxes with arrows represent the positive correlation between Ophiostoma presence and subcortical insects (with p-values).
Figure 3.
Subcortical insects reared from rearing cages. The individual species are grouped into families. The y-axis represents the logarithmic number of records of each species. Boxes with arrows represent the positive correlation between Ophiostoma presence and subcortical insects (with p-values).

Figure 4.
Subcortical insects captured in window traps. Individual species are grouped by family. The y-axis represents the logarithmic number of records for each species. Pie chart represents percentages for the number of species captured in each trapping method and by both methods together.
Figure 4.
Subcortical insects captured in window traps. Individual species are grouped by family. The y-axis represents the logarithmic number of records for each species. Pie chart represents percentages for the number of species captured in each trapping method and by both methods together.

Figure 5.
A conceptual model of abiotic and biotic factors involved in oak decline (modified from Thomas et al., 2002 [13]).
Figure 5.
A conceptual model of abiotic and biotic factors involved in oak decline (modified from Thomas et al., 2002 [13]).

Figure 6.
Chronology of cited factors in oak decline episodes over time (1739–2020), (pF = 35.4 p < 0.002). CCA = canonical correspondence analysis (CANOCO 5).
Figure 6.
Chronology of cited factors in oak decline episodes over time (1739–2020), (pF = 35.4 p < 0.002). CCA = canonical correspondence analysis (CANOCO 5).

Table 1.
Characteristics of the studied oak plots.
| Study Plot | Latitude; Longitude | Altitude (m a.s.l.) | Age | Average DBH (cm) | Average Height (m) |
|---|---|---|---|---|---|
| Pístecký les | 50°25′25.3″ N 14°08′2.7″ E | 198 | 60 | 31.4 | 26.8 |
| Tvořihráz | 48°54′27.5″ N 16°06′31.6″ E | 310 | 38 | 31 | 19.6 |
| Česká Skalice | 50°22′55.2″ N 16°02′27.2″ E | 318 | 40 | 19 | 19.5 |
| Nižbor | 50°00′09.9″ N 13°57′50.4″ E | 351 | 48 | 14.4 | 12.3 |
| Český kras | 49°56′50.3″ N 14°08′41.0″ E | 363 | 42 | 18.9 | 18.5 |
| Lásenice | 49°02′14.2″ N 14°58′54.3″ E | 530 | 110 | 36 | 29.2 |
a.s.l. = above sea level. DBH = diameter at breast height.
Table 2.
Intensity of Ophiostoma spp. presence identified at individual forest stands (Locality) and in different tree parts.
Table 2.
Intensity of Ophiostoma spp. presence identified at individual forest stands (Locality) and in different tree parts.
| Locality | Tree Part | Intensity (0–3) |
|---|---|---|
| Pístecký les | basal | 0 |
| middle | 1 | |
| crown | 1 | |
| Tvořihráz | basal | 1 |
| middle | 3 | |
| crown | 1 | |
| Česká Skalice | basal | 2 |
| middle | 1 | |
| crown | 1 | |
| Nižbor | basal | 3 |
| middle | 1 | |
| crown | 0 | |
| Český kras | basal | 0 |
| middle | 0 | |
| crown | 3 | |
| Lásenice | basal | 3 |
| middle | 1 | |
| crown | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Macháčová, M.; Nakládal, O.; Samek, M.; Baťa, D.; Zumr, V.; Pešková, V. Oak Decline Caused by Biotic and Abiotic Factors in Central Europe: A Case Study from the Czech Republic. Forests 2022, 13, 1223. https://doi.org/10.3390/f13081223
AMA Style
Macháčová M, Nakládal O, Samek M, Baťa D, Zumr V, Pešková V. Oak Decline Caused by Biotic and Abiotic Factors in Central Europe: A Case Study from the Czech Republic. Forests. 2022; 13(8):1223. https://doi.org/10.3390/f13081223
Chicago/Turabian StyleMacháčová, Markéta, Oto Nakládal, Michal Samek, Daniel Baťa, Václav Zumr, and Vítězslava Pešková. 2022. "Oak Decline Caused by Biotic and Abiotic Factors in Central Europe: A Case Study from the Czech Republic" Forests 13, no. 8: 1223. https://doi.org/10.3390/f13081223
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.