

Fungal Presence and Changes of Wood Structure in Bark Stripping Wounds Made by Red Deer (Cervus elaphus L.) on Stems of Fraxinus angustifolia (Vahl)
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Measurement and Sampling
- tree species;
- diameter at breast height (DBH, cm), measured at the 1.30 m mark of stem height;
- presence of red deer damage on stem (visible bark stripping, yes/no) (Figure 2);
- bark stripping intensity (1—less than 1/3 of circumference damaged, 2—1/3 to 2/3 of circumference damaged, 3—more than 2/3 of circumference damaged);
- height at which bark stripping occurred (the lowest and the highest point on each stem, m);
- total length of visible bark stripping on a stem (m).
2.2. Isolation and Identification of Fungi
2.3. Sample Preparation and Wood Structure Analysis
2.4. Statistical Analysis
3. Results
3.1. Inventory of Bark Stripping in the Analyzed Forest Stands
3.2. Fungi Present in Bark Stripping Wounds
3.3. Differences in Wood Structure in Relation to Zones, Trees, and Stem Height
4. Discussion
4.1. Bark Stripping Inventory
4.2. Fungal Presence in Bark Stripping Wounds
4.3. Wood Structure Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Temunović, M.; Franjić, J.; Satovic, Z.; Grgurev, M.; Frascaria-Lacoste, N.; Fernández-Manjarrés, J.F. Environmental Heterogeneity Explains the Genetic Structure of Continental and Mediterranean Populations of Fraxinus angustifolia Vahl. PLoS ONE 2012, 7, e42764. [Google Scholar] [CrossRef]
- Gašparović, M.; Pilaš, I.; Klobučar, D.; Gašparović, I. Monitoring Ash Dieback in Europe—An Unrevealed Perspective for Remote Sensing? Remote Sens. 2023, 15, 1178. [Google Scholar] [CrossRef]
- Kowalski, T. Chalara fraxinea sp. Nov. Associated with Dieback of Ash (Fraxinus excelsior) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the Causal Agent of E Uropean Ash Dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bottecchia, F.; Bregant, C.; Maddau, L.; Montecchio, L. Diplodia fraxini and Diplodia subglobosa: The Main Species Associated with Cankers and Dieback of Fraxinus excelsior in North-Eastern Italy. Forests 2020, 11, 883. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Brglez, A.; Piškur, B.; Ogris, N. First Report of Diplodia fraxini and Diplodia subglobosa Causing Canker and Dieback of Fraxinus excelsior in Slovenia. Plant Dis. 2022, 106, 26–29. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on Stems and Twigs in Initial and Advanced Stages of Dieback of European Ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef]
- Kowalski, T.; Bilański, P.; Kraj, W. Pathogenicity of Fungi Associated with Ash Dieback towards Fraxinus excelsior. Plant Pathol. 2017, 66, 1228–1238. [Google Scholar] [CrossRef]
- Kranjec Orlović, J.; Moro, M.; Diminić, D. Role of Root and Stem Base Fungi in Fraxinus angustifolia (Vahl) Dieback in Croatian Floodplain Forests. Forests 2020, 11, 607. [Google Scholar] [CrossRef]
- Meyn, R.; Langer, G.J.; Gross, A.; Langer, E.J. Fungal Colonization Patterns in Necrotic Rootstocks and Stem Bases of Dieback-Affected Fraxinus excelsior L. For. Pathol. 2019, 49, e12520. [Google Scholar] [CrossRef]
- Vemić, A. The Ecological and Histological Investigation of Species Complex Diaporthe Eres on Fraxinus excelsior Seed from Montenegro. Reforesta 2021, 11, 19–26. [Google Scholar] [CrossRef]
- Lygis, V.; Vasiliauskas, R.; Larsson, K.-H.; Stenlid, J. Wood-Inhabiting Fungi in Stems of Fraxinus excelsior in Declining Ash Stands of Northern Lithuania, with Particular Reference to Armillaria cepistipes. Scand. J. For. Res. 2005, 20, 337–346. [Google Scholar] [CrossRef]
- Bakys, R.; Vasiliauskas, A.; Ihrmark, K.; Stenlid, J.; Menkis, A.; Vasaitis, R. Root Rot, Associated Fungi and Their Impact on Health Condition of Declining Fraxinus excelsior Stands in Lithuania. Scand. J. For. Res. 2011, 26, 128–135. [Google Scholar] [CrossRef]
- Lenz, H.D.; Bartha, B.; Straßer, L.; Lemme, H. Development of Ash Dieback in South-Eastern Germany and the Increasing Occurrence of Secondary Pathogens. Forests 2016, 7, 41. [Google Scholar] [CrossRef]
- Bence, M.; Katona, K. Vadrágás Szerepe a Magas Kőris Hajtás- És Vesszőpusztulásában (Hymenoscyphus fraxineus). Vadbiológia 2020, 20, 31. [Google Scholar]
- Verheyden, H.; Ballon, P.; Bernard, V.; SAINT-ANDRIEUX, C. Variations in Bark-stripping by Red Deer Cervus elaphus across Europe. Mammal Rev. 2006, 36, 217–234. [Google Scholar] [CrossRef]
- Gill, R.; Beardall, V. The Impact of Deer on Woodlands: The Effects of Browsing and Seed Dispersal on Vegetation Structure and Composition. Forestry 2001, 74, 209–218. [Google Scholar] [CrossRef]
- Vasiliauskas, R.; Stenlid, J. Discoloration Following Bark Stripping Wounds on Fraxinus excelsior. Eur. J. For. Pathol. 1998, 28, 383–390. [Google Scholar] [CrossRef]
- Vasiliauskas, R. Patterns of Wounding and Decay in Stems of Quercus Robur Due to Bark Peeling. Scand. J. For. Res. 1998, 13, 437–441. [Google Scholar] [CrossRef]
- Putman, R.; Staines, B.W. Supplementary Winter Feeding of Wild Red Deer Cervus elaphus in Europe and North America: Justifications, Feeding Practice and Effectiveness. Mammal Rev. 2004, 34, 285–306. [Google Scholar] [CrossRef]
- Candaele, R.; Lejeune, P.; Licoppe, A.; Malengreaux, C.; Brostaux, Y.; Morelle, K.; Latte, N. Mitigation of Bark Stripping on Spruce: The Need for Red Deer Population Control. Eur. J. For. Res. 2021, 140, 227–240. [Google Scholar] [CrossRef]
- Vospernik, S. Probability of Bark Stripping Damage by Red Deer (Cervus elaphus) in Austria. Silva Fenn. 2006, 40, 589. [Google Scholar] [CrossRef]
- Månsson, J.; Jarnemo, A. Bark-Stripping on Norway Spruce by Red Deer in Sweden: Level of Damage and Relation to Tree Characteristics. Scand. J. For. Res. 2013, 28, 117–125. [Google Scholar] [CrossRef]
- Renaud, P.-C.; Verheyden-Tixier, H.; Dumont, B. Damage to Saplings by Red Deer (Cervus elaphus): Effect of Foliage Height and Structure. For. Ecol. Manag. 2003, 181, 31–37. [Google Scholar] [CrossRef]
- Kiffner, C.; Rossiger, E.; Trisl, O.; Schulz, R.; Rühe, F. Probability of Recent Bark Stripping Damage by Red Deer (Cervus elaphus) on Norway Spruce (Picea abies) in a Low Mountain Range in Germany—A Preliminary Analysis. Silva Fenn. 2008, 42, 125. [Google Scholar] [CrossRef]
- Hahn, C.; Vospernik, S. Position, Size, and Spatial Patterns of Bark Stripping Wounds Inflicted by Red Deer (Cervus elavus L.) on Norway Spruce Using Generalized Additive Models in Austria. Ann. For. Sci. 2022, 79, 13. [Google Scholar] [CrossRef]
- White, T. The Cause of Bark Stripping of Young Plantation Trees. Ann. For. Sci. 2019, 76, 105. [Google Scholar] [CrossRef]
- Maria Arnold, J.; Gerhardt, P.; Steyaert, S.M.; Hochbichler, E. Diversionary Feeding Can Reduce Red Deer Habitat Selection Pressure on Vulnerable Forest Stands, but Is Not a Panacea for Red Deer Damage. For. Ecol. Manag. 2017, 407, 166–173. [Google Scholar] [CrossRef]
- Jerina, K.; Dajčman, M.; Adamič, M. Red Deer (Cervus elaphus) Bark Stripping on Spruce with Regard to Spatial Distribution of Supplemental Feeding Places. Zb. Gozdarstva Lesar. 2008, 86, 33–43. [Google Scholar]
- Boddy, L. Fungal Community Ecology and Wood Decomposition Processes in Angiosperms: From Standing Tree to Complete Decay of Coarse Woody Debris. Ecol. Bull. 2001, 49, 43–56. [Google Scholar]
- Bässler, C.; Müller, J.; Dziock, F.; Brandl, R. Effects of Resource Availability and Climate on the Diversity of Wood-decaying Fungi. J. Ecol. 2010, 98, 822–832. [Google Scholar] [CrossRef]
- Marčiulynas, A.; Sirgedaitė-Šėžienė, V.; Menkis, A. Fungi Inhabiting Stem Wounds of Quercus Robur Following Bark Stripping by Deer Animals. Forests 2023, 14, 2077. [Google Scholar] [CrossRef]
- Menkis, A.; Redr, D.; Bengtsson, V.; Hedin, J.; Niklasson, M.; Nordén, B.; Dahlberg, A. Endophytes Dominate Fungal Communities in Six-Year-Old Veteranisation Wounds in Living Oak Trunks. Fungal Ecol. 2022, 59, 101020. [Google Scholar] [CrossRef]
- Parfitt, D.; Hunt, J.; Dockrell, D.; Rogers, H.J.; Boddy, L. Do All Trees Carry the Seeds of Their Own Destruction? PCR Reveals Numerous Wood Decay Fungi Latently Present in Sapwood of a Wide Range of Angiosperm Trees. Fungal Ecol. 2010, 3, 338–346. [Google Scholar] [CrossRef]
- Cline, L.C.; Schilling, J.S.; Menke, J.; Groenhof, E.; Kennedy, P.G. Ecological and Functional Effects of Fungal Endophytes on Wood Decomposition. Funct. Ecol. 2018, 32, 181–191. [Google Scholar] [CrossRef]
- Arhipova, N.; Jansons, A.; Zaluma, A.; Gaitnieks, T.; Vasaitis, R. Bark Stripping of Pinus Contorta Caused by Moose and Deer: Wounding Patterns, Discoloration of Wood, and Associated Fungi. Can. J. For. Res. 2015, 45, 1434–1438. [Google Scholar] [CrossRef]
- Hecht, U.; Kohnle, U.; Nill, M.; Grüner, J.; Metzler, B. Bark Wounds Caused by Felling Are More Susceptible to Discoloration and Decay than Wounds Caused by Extraction in European Beech. Ann. For. Sci. 2015, 72, 731–740. [Google Scholar] [CrossRef]
- Moore, G. Ring-Barking and Girdling: How Much Vascular Connection Do You Need between Roots and Crown; University of Melbourne Adelaide: Docklands, Australia, 2013; pp. 87–96. [Google Scholar]
- Rößner, M.; Vor, T.; Seidel, D. Quantifying Bark-Stripping Damages to Address the Relationship between External and Internal Wood Defects. Trees For. People 2023, 13, 100415. [Google Scholar] [CrossRef]
- Čermák, P.; Beranová, P.; Orálková, J.; Horsák, P.; Plšek, J. Relationships between Browsing Damage and the Species Dominance by the Highly Food-Attractive and Less Food-Attractive Trees. Acta Univ. Agric. Silvic. Mendel. Brun. 2014, 59, 4. [Google Scholar] [CrossRef]
- Schwartz, M.K.; Luikart, G.; Waples, R.S. Genetic Monitoring as a Promising Tool for Conservation and Management. Trends Ecol. Evol. 2007, 22, 25–33. [Google Scholar] [CrossRef]
- Welch, D.; Scott, D. Observations on Bark-Stripping by Red Deer in a Picea Sitchensis Forest in Western Scotland over a 35-Year Period. Scand. J. For. Res. 2017, 32, 473–480. [Google Scholar] [CrossRef]
- Zobel, B.J.; Van Buijtenen, J.P. Wood Variation: Its Causes and Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 3-642-74069-3. [Google Scholar]
- Panshin, A.J.; de Zeeuw, C. Textbook of Wood Technology. Volume I. Structure, Identification, Uses, and Properties of the Commercial Woods of the United States and Canada; CABI: Wallingford, UK, 1970. [Google Scholar]
- Wagenführ, R.; Scheiber, C. Holzatlas; Fachbuchverlag Leipzig: Munchen, Germany, 1974. [Google Scholar]
- Tsoumis, G. Science and Technology of Wood: Structure, Properties, Utilization; Van Nostrand Reinhold: New York, NY, USA, 1991; Volume 115. [Google Scholar]
- Anić, I. Uspijevanje i Pomlađivanje Sastojina Poljskog Jasena (Fraxinus angustifolia Vahl) u Posavini. Ph.D. Thesis, University of Zagreb Faculty of Forestry, Zagreb, Croatia, 2001. [Google Scholar]
- Cenis, J.L. Rapid Extraction of Fungal DNA for PCR Amplification. Nucleic Acids Res. 1992, 20, 2380. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes-application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Elsevier BV: Amsterdam, The Netherlands, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucl Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tomljanović, K.; Grubešić, M.; Diminić, D.; Poljak, M.; Kranjec Orlović, J. Red Deer (Cervus elaphus L.) Damage on Stands of Narrow-Leaved Ash (Fraxinus angustifolia Vahl) of Middle Posavina. Šumar. List 2022, 146, 117–124. [Google Scholar] [CrossRef]
- Čermák, P.; Horsák, P.; Špiřík, M.; Mrkva, R. Relationships between Browsing Damage and Woody Species Dominance. J. For. Sci. 2009, 55, 23–31. [Google Scholar] [CrossRef]
- Volk, F. Bark Peeling Frequency in the Alpine Provinces of Austria: The Importance of Forest Structure and Red Deer Management. Z. Fûr Jagdwiss. 1999, 45, 1–16. [Google Scholar]
- Gill, R. A Review of Damage by Mammals in North Temperate Forests: 1. Deer. For. Int. J. For. Res. 1992, 65, 145–169. [Google Scholar] [CrossRef]
- Jelonek, T.; Tomczak, K.; Naskrent, B.; Klimek, K.; Tomczak, A.; Lewandowski, K. The Effect of Beech (Fagus sylvatica L.) Bark Stripping by Deer on Depreciation of Wood. Forests 2022, 13, 1531. [Google Scholar] [CrossRef]
- Krisans, O.; Saleniece, R.; Rust, S.; Elferts, D.; Kapostins, R.; Jansons, A.; Matisons, R. Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests 2020, 11, 357. [Google Scholar] [CrossRef]
- Hart, J.H.; Hart, D. Interaction among Cervids, Fungi, and Aspen in Northwest Wyoming; Shepperd, W.D., Binkley, D., Bartos, D.L., Stohlgren, T.J., Eskew, L.G., Eds.; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2001; pp. 197–205. [Google Scholar]
- Burneviča, N.; Jansons, Ā.; Zaļuma, A.; Kļaviņa, D.; Jansons, J.; Gaitnieks, T. Fungi Inhabiting Bark Stripping Wounds Made by Large Game on Stems of Picea abies (L.) Karst. in Latvia. Balt. For. 2016, 22, 2–7. [Google Scholar]
- Peters, S.; Fuchs, S.; Bien, S.; Bußkamp, J.; Langer, G.J.; Langer, E.J. Fungi Associated with Stem Collar Necroses of Fraxinus excelsior Affected by Ash Dieback. Mycol. Prog. 2023, 22, 52. [Google Scholar] [CrossRef]
- Ghaedi, M.; Bolboli, Z.; Mostowfizadeh-Ghalamfarsa, R. First Report of Peroneutypa Scoparia Associated with Canker Disease on Ficus Carica in Northern Iran. New Dis. Rep. 2023, 48, e12201. [Google Scholar] [CrossRef]
- Travadon, R.; Baumgartner, K. Molecular Polymorphism and Phenotypic Diversity in the Eutypa Dieback Pathogen Eutypa Lata. Phytopathology 2015, 105, 255–264. [Google Scholar] [CrossRef]
- Pusz, W.; Baturo-Cieśniewska, A.; Kaczmarek-Pieńczewska, A.; Patejuk, K.; Czarnota, P. Is the Survivability of Silver Fir under Condition of Strong Ungulate Pressure Related to Mycobiota of Bark-Stripping Wounds? Forests 2021, 12, 976. [Google Scholar] [CrossRef]
- Nemesio-Gorriz, M.; McGuinness, B.; Grant, J.; Dowd, L.; Douglas, G.C. Lenticel Infection in Fraxinus excelsior Shoots in the Context of Ash Dieback. IForest-Biogeosciences For. 2019, 12, 160. [Google Scholar] [CrossRef]
- Langer, G.J.; Peters, S.; Bußkamp, J.; Bien, S. Cryptostroma Corticale and Fungal Endophytes Associated with Fraxinus excelsior Affected by Ash Dieback. J. Plant Dis. Prot. 2023, 1–11. [Google Scholar] [CrossRef]
- Kowalski, T.; Bilański, P. Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus. Forests 2021, 12, 1412. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Proctor, R.H.; Moretti, A. Mycotoxin Production in Fusarium According to Contemporary Species Concepts. Annu. Rev. Phytopathol. 2021, 59, 373–402. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma Species—Opportunistic, Avirulent Plant Symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Elad, Y.; Chet, I.; Henis, Y. Degradation of Plant Pathogenic Fungi by Trichoderma Harzianum. Can. J. Microbiol. 1982, 28, 719–725. [Google Scholar] [CrossRef]
- Bauch, J.; Shigo, A.; Starck, M. Wound Effects in the Xylem of Acer and Betula Species. Holzforschung 1980, 34, 153–160. [Google Scholar] [CrossRef]
- Rademacher, P.; Bauch, J.; Shigo, A. Characteristics of Xylem Formed after Wounding in Acerbetula, Andfagus. LAWA Bull. 1984, 5, 141–150. [Google Scholar]
- Lowerts, G.; Wheeler, E.; Kellison, R.C. Characteristics of Wound-Associated Wood of Yellow-Poplar (Liriodendron tulipifera L.). Wood Fiber Sci. 1986, 18, 537–552. [Google Scholar]
- Morris, H.; Brodersen, C.; Schwarze, F.W.; Jansen, S. The Parenchyma of Secondary Xylem and Its Critical Role in Tree Defense against Fungal Decay in Relation to the CODIT Model. Front. Plant Sci. 2016, 7, 1665. [Google Scholar] [CrossRef]
- Shigo, A.L.; Marx, H.G. Compartmentalization of Decay in Trees; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 1977. [Google Scholar]
- Welch, D.; Scott, D. An Estimate of Timber Degrade in Sitka Spruce Due to Bark Stripping by Deer in a Scottish Plantation. Forestry 2008, 81, 489–497. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Link, R.; Li, R.; Deng, L.; Schuldt, B.; Jiang, X.; Zhao, R.; Zheng, J.; Li, S. Influence of Cambial Age and Axial Height on the Spatial Patterns of Xylem Traits in Catalpa Bungei, a Ring-Porous Tree Species Native to China. Forests 2019, 10, 662. [Google Scholar] [CrossRef]
- Anfodillo, T.; Petit, G.; Crivellaro, A. Axial Conduit Widening in Woody Species: A Still Neglected Anatomical Pattern. In Wood Structure in Plant Biology and Ecology; Brill: Leiden, The Netherlands, 2013; pp. 24–36. [Google Scholar]
- Anfodillo, T.; Carraro, V.; Carrer, M.; Fior, C.; Rossi, S. Convergent Tapering of Xylem Conduits in Different Woody Species. New Phytol. 2006, 169, 279–290. [Google Scholar] [CrossRef]
- Tippett, J.T.; Shigo, A.L. Barrier Zone Formation: A Mechanism of Tree Defense against Vascular Pathogens. IAWA J. 1981, 2, 163–168. [Google Scholar] [CrossRef]
- Luley, C.J. Biology and Assessment of Callus and Woundwood. Arborist News 2015, 24, 12–21. [Google Scholar]
- Fonti, P.; Macchioni, N. Ring Shake in Chestnut: Anatomical Description, Extent and Frequency of Failures. Ann. For. Sci. 2003, 60, 403–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
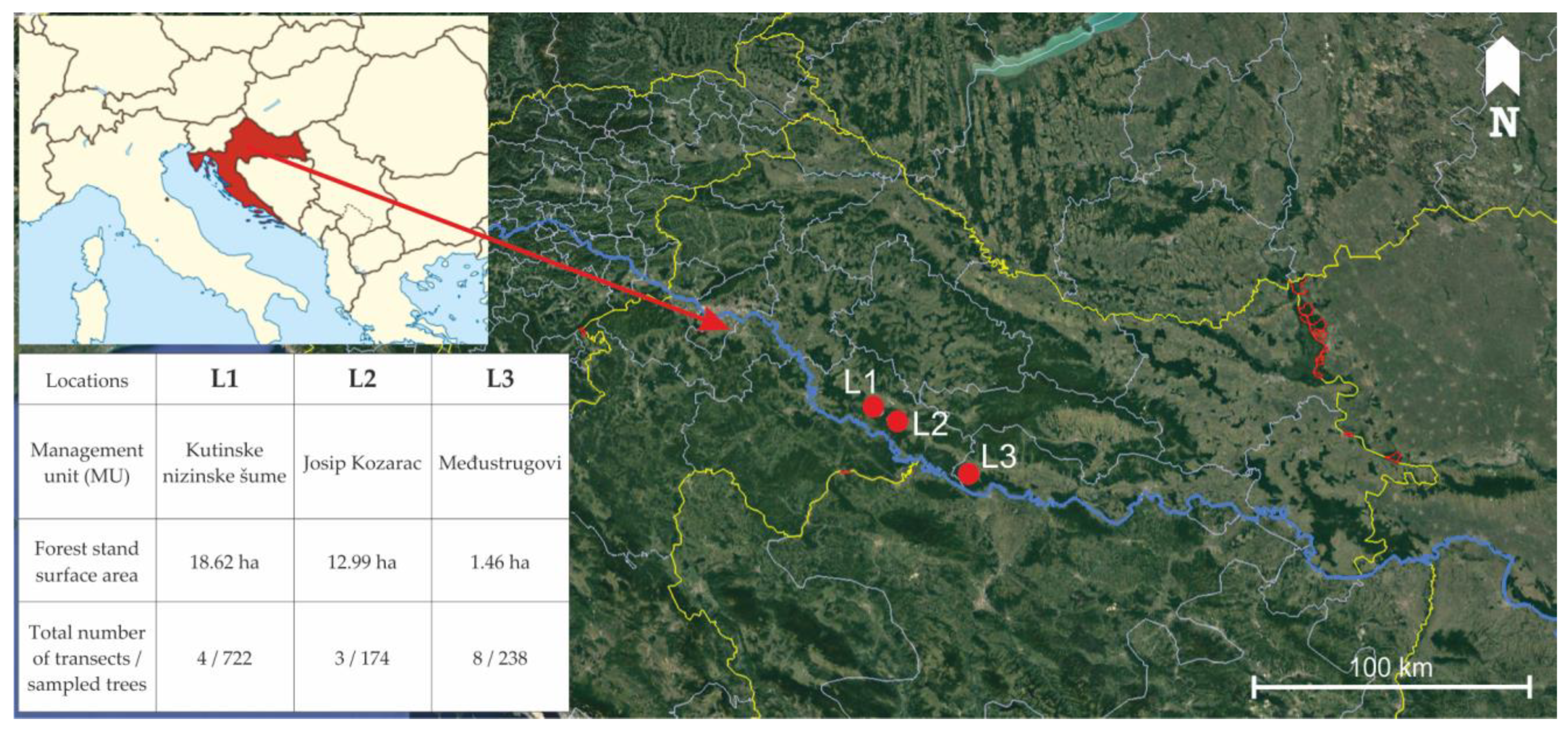
| L1 | L2 | L3 | |
|---|---|---|---|
| Coordinates | 45.431646 N, 16.735151 E | 45.403425 N, 16.787225 E | 45.203113 N, 17.152394 E |
| Elevation (above sea level, m) | 94–95 | 94–95 | 91–92 |
| Slope (°) | 0 | 0 | 0 |
| Phytocenosis | forest of narrow-leaved ash with autumn snowflake (As. Leucojo-Fraxinetum angustifoliae Glavač 1959) | forest of pedunculate oak with Genista elata (As. Genisto elatae-Quercetum roboris Ht. 1938) | forest of narrow-leaved ash with autumn snowflake (As. Leucojo-Fraxinetum angustifoliae Glavač 1959) |
| Forest type | regular, even aged forest of narrow-leaved ash | regular, even aged forest of pedunculate oak with narrow-leaved ash | regular, even aged forest of narrow-leaved ash |
| Average tree age | <15 years | <15 years | <15 years |
| Density (No. of trees/ha) | 388 | 134 | 1698 |
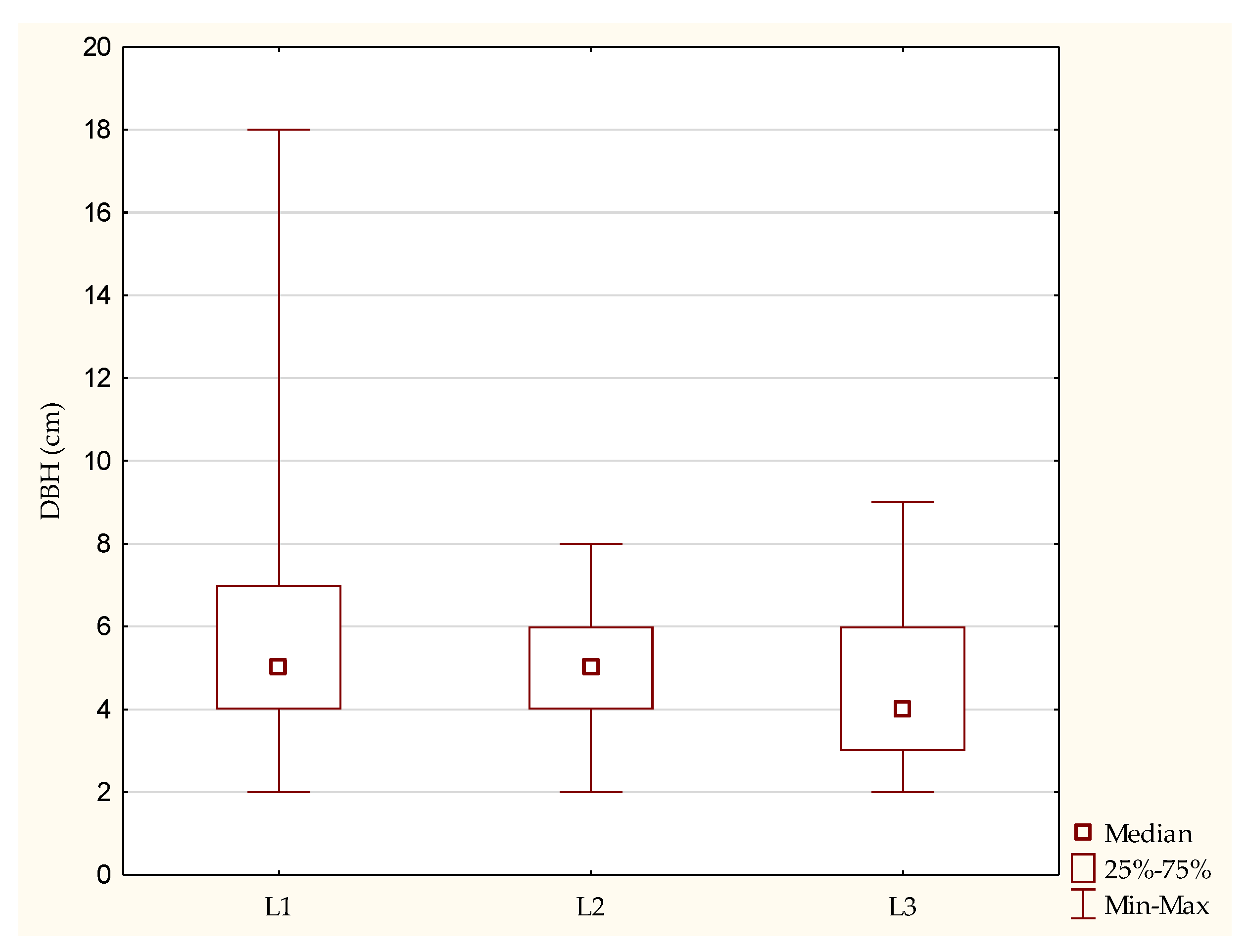
| The average DBH of narrow-leaved ash trees (cm) | 10 | 7 | 5 |
| Soil type | amphigley | amphigley | amphigley |
| Mean annual temperature (°C) | 11.01 | 11.01 | 11.1 |
| Mean temperature in the vegetation period (°C) | 17.6 | 17.6 | 17.7 |
| Mean annual precipitation (mm) | 861.3 | 861.3 | 770.3 |
| Mean precipitation in the vegetation period (mm) | 506.7 | 506.7 | 441.0 |
| Understory vegetation | shrubs rarely present, ground vegetation mostly presented by Leucojum aestivum | rich shrub layer mostly presented by Genista elata | shrubs rarely present, ground vegetation mostly presented by Leucojum aestivum |
| L1 | L2 | L3 | |
|---|---|---|---|
| The average lowest point of bark stripping (±SD) (m) | 0.4 (±0.2) | 0.5 (±0.3) | 0.3 (±0.2) |
| The average highest point of bark stripping (±SD) (m) | 1.0 (±0.3) | 1.0 (±0.3) | 0.9 (±0.2) |
| The average length of visible bark stripping (±SD) (m) | 0.6 (±0.3) | 0.5 (±0.2) | 0.6 (±0.2) |
| Variable | DBH | Lowest Point of Bark Stripping on a Stem | Highest Point of Bark Stripping on a Stem | Length of Bark Stripping on a Stem |
|---|---|---|---|---|
| DBH | 1.000 | 0.181 | 0.153 | 0.004 |
| Lowest point of bark stripping on a stem | 0.181 | 1.000 | 0.441 | −0.394 |
| Highest point of bark stripping on a stem | 0.153 | 0.441 | 1.000 | 0.651 |
| Length of bark stripping on a stem | 0.004 | −0.394 | 0.651 | 1.000 |
| Phylum | Fungal Taxon | GenBank Accession No. | No. of Obtained Isolates | No. of Colonized Stems |
|---|---|---|---|---|
| A | Aspergillus sp. | OR808055 | 3 | 2 |
| A | Boeremia exigua | OR808056 | 1 | 1 |
| A | Boeremia sp. | OR808057 | 5 | 2 |
| A | Cadophora sp. | OR808058 | 1 | 1 |
| A | Cladosporium sp. | OR808059 | 1 | 1 |
| A | Clonostachys rosea | OR808060 | 17 | 3 |
| A | Dothiorella viticola | OR808061 | 1 | 1 |
| A | Eutypa lata | OR808062 | 29 | 1 |
| A | Fusarium acuminatum | OR808063 | 13 | 4 |
| A | Fusarium sp. 31c_36 | OR808064 | 3 | 2 |
| A | Fusarium sp. 31c_42 | OR808065 | 2 | 2 |
| A | Fusarium sp. 31c_4 | OR808066 | 12 | 2 |
| A | Fusarium sp. 31c_41 | OR808067 | 2 | 2 |
| A | Fusarium sp. 31c_40 | OR808068 | 1 | 1 |
| A | Fusarium sp. 31c_39 | OR808069 | 12 | 3 |
| A | Neofabraea vagabunda | OR808070 | 2 | 1 |
| A | Paracucurbitaria corni | OR808071 | 2 | 2 |
| A | Penicillium brevicompactum | OR808072 | 2 | 1 |
| A | Penicillium sizovae | OR808073 | 2 | 1 |
| A | Peroneutypa scoparia | OR808074 | 6 | 1 |
| A | Phaeoacremonium sp. | OR808075 | 3 | 1 |
| A | Phialemonium sp. | OR808076 | 12 | 1 |
| A | Pleosporales sp. | OR808077 | 2 | 1 |
| A | Trichoderma spp. | – | 32 | 5 |
| B | Bjerkandera adusta | OR808078 | 1 | 1 |
| B | Filobasidium stepposum | OR808079 | 2 | 1 |
| B | Hyphodermella rosae | OR808080 | 4 | 1 |
| B | Papiliotrema laurentii | OR808081 | 1 | 1 |
| Tree | Property | Data | Type | Stem Height (m) | ||||
|---|---|---|---|---|---|---|---|---|
| 0.25 | 0.75 | 1.25 | 1.75 | Total (All Heights) | ||||
| 1 | EVLD (µm) | MEAN ± SD | C | 151.24 ± 36.53 | 183.09 ± 33.02 | 171.20 ± 35.50 | 238.93 ± 41.98 | 186.12 ± 36.76 |
| D | 148.38 ± 27.92 | 80.63 ± 25.92 | 97.25 ± 23.88 | 119.71 ± 64.41 | 111.49 ± 35.53 | |||
| D (cm) | MEAN | S | 35.00 | 31.00 | 29.00 | 22.00 | 29.25 | |
| 2 | EVLD (µm) | MEAN ± SD | C | 168.43 ± 43.83 | 118.14 ± 34.67 | 174.50 ± 35.21 | 141.31 ± 29.15 | 150.60 ± 35.72 |
| D | 91.90 ± 20.86 | 72.54 ± 24.75 | 88.83 ± 30.54 | 120.49 ± 32.41 | 93.44 ± 27.14 | |||
| D (cm) | MEAN | S | 24.00 | 21.00 | 19.50 | 16.50 | 20.25 | |
| 3 | EVLD (µm) | MEAN ± SD | C | 177.60 ± 28.67 | 140.91 ± 33.95 | 159.54 ± 29.50 | 176.62 ± 24.96 | 163.67 ± 29.27 |
| D | 162.91 ± 35.36 | 101.29 ± 32.18 | 126.86 ± 28.60 | 88.94 ± 23.12 | 120.00 ± 29.82 | |||
| D (cm) | MEAN | S | 27.00 | 22.00 | 20.50 | 17.50 | 21.75 | |
| 4 | EVLD (µm) | MEAN ± SD | C | 151.96 ± 35.98 | 178.13 ± 27.38 | 168.26 ± 36.34 | 235.87 ± 39.29 | 183.56 ± 34.75 |
| D | 147.81 ± 29.65 | 81.46 ± 23.04 | 99.41 ± 19.22 | 118.39 ± 54.14 | 111.77 ± 31.51 | |||
| D (cm) | MEAN | S | 33.50 | 30.50 | 29.00 | 24.00 | 29.25 | |
| 5 | EVLD (µm) | MEAN ± SD | C | 167.34 ± 37.15 | 120.81 ± 29.97 | 172.54 ± 33.83 | 141.81 ± 28.15 | 150.63 ± 32.28 |
| D | 87.78 ± 18.91 | 71.42 ± 20.15 | 84.04 ± 29.21 | 119.25 ± 29.92 | 90.62 ± 24.55 | |||
| D (cm) | MEAN | S | 25.50 | 22.50 | 19.00 | 16.50 | 20.88 | |
| Property | Source | SS | d.f. | MS | F | p |
|---|---|---|---|---|---|---|
| EVLD (µm) | Tree × Zone | 4281 | 4 | 1070 | 2.94 | 0.0365 |
| Stem height × Tree | 21514 | 12 | 1793 | 9.1 | 0.0000 | |
| Stem height × Zone | 10429 | 3 | 3476 | 17.6 | 0.0000 | |
| Stem height × Tree × Zone | 35529 | 12 | 2961 | 15.0 | 0.0000 |
| Property | Stem Height (m) | Source | SS | d.f. | MS | F | p |
|---|---|---|---|---|---|---|---|
| EVLD (µm) | 0.25 | Zone | 12,883.8 | 1 | 12,883.8 | 4.8 | 0.0594 |
| 0.75 | Zone | 42,432.8 | 1 | 42,432.8 | 21.2 | 0.0017 | |
| 1.25 | Zone | 49,862.0 | 1 | 49,862.0 | 71.8 | 0.0001 | |
| 1.75 | Zone | 61,930.6 | 1 | 61,930.0 | 12.0 | 0.0085 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kranjec Orlović, J.; Diminić, D.; Ištok, I.; Volenec, I.; Hodak, L.; Grubešić, M.; Tomljanović, K. Fungal Presence and Changes of Wood Structure in Bark Stripping Wounds Made by Red Deer (Cervus elaphus L.) on Stems of Fraxinus angustifolia (Vahl). Forests 2024, 15, 314. https://doi.org/10.3390/f15020314
Kranjec Orlović J, Diminić D, Ištok I, Volenec I, Hodak L, Grubešić M, Tomljanović K. Fungal Presence and Changes of Wood Structure in Bark Stripping Wounds Made by Red Deer (Cervus elaphus L.) on Stems of Fraxinus angustifolia (Vahl). Forests. 2024; 15(2):314. https://doi.org/10.3390/f15020314
Chicago/Turabian StyleKranjec Orlović, Jelena, Danko Diminić, Iva Ištok, Ida Volenec, Luka Hodak, Marijan Grubešić, and Kristijan Tomljanović. 2024. "Fungal Presence and Changes of Wood Structure in Bark Stripping Wounds Made by Red Deer (Cervus elaphus L.) on Stems of Fraxinus angustifolia (Vahl)" Forests 15, no. 2: 314. https://doi.org/10.3390/f15020314