
Involvement of Lophodermella sulcigena in Endemic Disease of Pinus mugo Needles in the Polish Tatra Mountains


Department of Forest Ecosystems Protection, University of Agriculture in Krakow, Al. 29 Listopada 46, 31-425 Krakow, Poland
*
Author to whom correspondence should be addressed.
Forests 2024, 15(3), 422; https://doi.org/10.3390/f15030422
Submission received: 9 January 2024
/
Revised: 13 February 2024
/
Accepted: 20 February 2024
/
Published: 22 February 2024
(This article belongs to the Section Forest Health)
Abstract
:Pinus mugo plays a significant ecological role in the natural environment at high altitudes in the mountains including the Alps, Pyrenees, Carpathians, and Balkans. In such severe conditions, it is subjected to the harmful effects of various abiotic and biotic factors. In one of the areas of its natural occurrence in Tatra Mts. (southern Poland), for the last few years, a significant intensification of needle disease has been observed. Symptoms similar to those recorded in Tatra Mts. also occur on other Pinus species in Europe and North America, where they are caused by fungi belonging to the genus Elytroderma, Lophodermella, Lophophacidium or Ploioderma (Rhytismataceae). The current paper presents the results of research which was mainly aimed at characterization of disease symptoms observed for the first time in Poland on P. mugo needles, and identification of the main causal agent with use of the morphological and molecular technique. Based on the analyses performed at different times of the year (2015–2020), it was found that dieback symptoms initially appeared only on first-year needles, a few weeks after their development. Symptoms occur on one or both needles in the bundle. The distal parts of the needles died, while the basal parts remained green. In the following year, mainly in June and July, on the previous year’s needles attached to the shoots, mature ascomata can be seen. The fungus Lophodermella sulcigena has been identified as the cause of these symptoms. So far, the related species L. conjuncta has not been found. The morphological features of the pathogen microstructure produced on P. mugo needles are presented. Attention was drawn to certain features that may make its identification difficult, especially in terms of shapes and sizes of ascospores. The phylogenetic position of the identified causal agent in relation to closely related other species was determined. The current results confirmed that L. sulcigena shows great phylogenetic similarity to L. montivaga, which is found in North America. Nine rDNA barcode sequences of L. sulcigena obtained in this work will enrich the NCBI GenBank database. The obtained results, indicating the presence of other fungi in L. sulcigena ascomata, which may limit the spread of its ascospores, were also discussed.
1. Introduction
The dwarf mountain pine (Pinus mugo Turra) mainly grows as a shrub adapted to various rocky habitats in the high-altitude mountains, including the Alps, Pyrenees, Carpathians, and Balkans [1]. It shows a high capacity for vegetative reproduction and is characterized by a large morphological diversity [2,3]. A related species found in the Pyrenees and Western Alps is distinguished as a separate taxon, P. uncinata, which develops a 12–20 m high tree form [2,4]. In the Tatra Mountains (southern Poland), P. mugo forms compact groups of shrubs mainly above the montane forest zone, ca. 1.550–1.850 m a.s.l. [3,5].
P. mugo plays a significant ecological role in establishing a natural mountain environment. It protects soil from erosion, decreases the avalanche hazard and promotes formation of soil layer. The strong root system of P. mugo increases soil pores, thereby improving the hydrological condition of soils [1,2,6,7].
P. mugo in severe mountain conditions may be subjected to various types of damage caused by abiotic factors. Stem damage can be caused by rockfall, snow pressure, and wind abrasion [8,9]. Winter drying in dwarf pine clusters can be caused by unfavorable weather conditions. Reduced snow cover exposes plants to increased temperature stress and hinders recovery processes [10]. Reductions in snow cover heights and durations may lead to embolism in P. mugo, similar to the seasonal patterns observed in other conifers [11]. At high elevation, drought and frost can co-occur in circumstances where towards the end of winter air temperature increases while soil is still frozen, inducing the so-called winter drought [12]. These factors may lead to the death of needles and shoots and increase of the disease predisposition of plants [11,12].
P. mugo also suffers from biotic factors that cause various types of diseases. One of the most important diseases is brown felt blight caused by Herpotrichia juniperi, which can lead to destruction of natural regenerations in large areas, especially when thick snow cover persists for a long time [13,14]. In places, similar symptoms are caused by Allantophomopsis cytisporea [15]. In some mountain sites the dieback of P. mugo is largely due to the fungi of the genus Armillaria and Heterobasidion, which infect plant roots and cause their rot [16,17]. In turn, the most known diseases of P. mugo needles are red band disease caused by Dothistroma septosporum and brown spot needle blight caused by Lecanosticta acicola [14,18,19,20,21,22,23,24,25,26]. Among the insects that damage the needles of P. mugo, needle shortening gall midge (Thecodiplosis brachyntera) can cause significant losses [27].
For several years in Polish Tatra, there has been a high intensity of P. mugo needle diseases in some regions [28]. During the preliminary analysis, several different types of disease symptoms on the needles were distinguished, and an attempt was made to determine which fungi can be associated with these symptoms. In total, more than fifty fungal taxa were found in symptomatic and symptomless needles [28]. Three species, Lophodermium corconticum, Lo. pini-mugonis and Leptomelanconium allescheri, have been detected for P. mugo in this region for the first time [28,29]. The only fungal species that was found in each symptomatic needle group and also in living symptomless needles was Lo. conigenum. Of the biotrophic fungi, only one species has been found: Coleosporium tussilaginis. During the in situ analysis, it was found that the most common symptom was the death of the distal parts of this year’s needles just a few weeks after their development, while the green basal parts were unscathed. This type of symptom on the needles of various pine species can be caused by fungi of the genus Elytroderma, Lophodermella, Lophophacidium and Ploioderma [14,30,31,32,33]. In Poland, their occurrence on the needles of P. mugo has so far not been recorded [34,35,36]. Our preliminary study of P. mugo needles with this type of symptom showed significant differences between the result of morphological analyses and the result obtained with the use of molecular technique. Due to the great importance of this disease for the reduction of the health status of P. mugo in the Tatra Mountains, it was considered justified to conduct more detailed studies. The current paper presents the results of research that had two primary objectives: (i) documentation of disease symptoms observed for the first time in Poland on P. mugo needles, and (ii) identification and characterization of the main causal agent on the morphological and molecular basis. An additional goal was to determine the phylogenetic position of identified causal agent in relation to closely related other species. Many rDNA barcode sequences of identified causal agents sent to the NCBI GenBank database can be used for comparison purposes for other researchers. The obtained results should provide a basis for further considerations on the possibilities of protecting P. mugo in the high-altitude mountains.
2. Materials and Methods
2.1. Field Observations
The object of the research was P. mugo growing in the Polish Tatra in Chochołowska Valley, where in 2015 a high intensity of the needle disease was noticed, which lasted for the following years. Field observations were carried out on seven plots (1.5–5.5 a) located in dense clumps of P. mugo at an altitude of 1556 to 1746 m a.s.l., where an average of 44%–65% needles with discoloration and necrosis were present. The location of these plots and the severity of the needle disease were presented by Bartnik et al. [28]. The samples were collected in autumn 2015 and at least four times in the period from May to November in 2016, 2017, and 2020, and were also used for conducting research on other diseases of P. mugo needles [28]. For the purposes of the current study, a detailed description and documentation of selected types of symptoms on this year’s needles, and their changes on two-year and three-year-old needles, was performed.
2.2. Symptom Analysis
In 2016, a detailed quantitative analysis of the previous year’s needles and current year’s needles was carried out. For this purpose, 14 twigs in June and 7 twigs in September 2016 were collected, respectively two and one twig, on each of the 7 study plots. The twigs representing the most common disease severity came from the upper part of the crown. They were selected as follows: 10 trees were randomly selected on each plot and the discoloration of needles on the selected twigs was analyzed by assessing its intensity according to the adopted five-point scale. Then, on the basis of the weighted average, the average degree of needle discoloration was determined and the twig that best represented the calculated degree of needle damage was taken for analysis. The twigs were placed separately in plastic bags, transported to the Department’s laboratory and placed in a cold store at −25 °C. Then, the selected parameters of the needles were measured: the number of needles was calculated depending on the type of shoot (main, lateral), the length of the needles was measured, the degree of discoloration of each needle, the color of the necrotic part and the presence of hysterothecia characteristic of Lophodermella species were assessed. Statistica 13.1 [37] software was used for statistical analyses. The Student’s T-test was used to compare two independent means.
In addition, an analysis of the width of annual rings was carried out in order to determine whether needle disease is accompanied by a decrease in thickness. For this purpose, in September 2020, 35 apical shoots of P. mugo, each 50 cm long (7 plots, 5 shoots each), were randomly sampled. In the laboratory, 3 cm thick fragments were cut from the base of these shoots, the cross-sectional surface was sanded and scanned with a resolution of 2400 dpi. Measurements were made with the accuracy of one hundredth of a mm using CooRecoder 7.6 and CDendro 7.6 [38,39]. Thickness increments in a given year were compared in relation to such increments in the previous year.
2.3. Fungal Isolation and Microscopic Analysis
To assess the occurrence of hysterothecia of Lophodermella type, observations were made using the Zeiss Discovery V12 stereomicroscope (Zeiss, Göttingen, Germany) on needles attached to shoots collected at different times of the year, as well as on needles already fallen from shoots. For microscopic analysis, 1–3 slides from the given fruit body, immersed in water, were prepared. Occasionally, hand-cut sections were made. Great attention was paid to the fact that hysterothecia of various lengths and at various stages of maturity should be included in the microscopic analyses. Measurements of morphological structures were made using the Zeiss Axiophot light microscope with DIC (differential interference contrast) illumination under 1000× enlargement. Numerous cross-sections and analyses of dark spots on needles adjacent to hysterothecia and before their development were also performed. Fungi co-occurring on needles infected by Lophodermella and inside hysterothecia were also identified. Among the fungi associated with Lophodermella, only Lophodermium spp. produced sexual and asexual morphs, and the others only produced asexual stages. For the microscopical analyses, appropriate mycological keys and monographs were used [14,28,31,35,40,41,42,43,44,45].
Needles with a dead distal part and a living basal part were used for isolation. Surface sterilization was obtained by rinsing the needles in 96% ethanol (1 min), then in a solution of NaOCl (ca. 4% available chlorine, 30 s), 96% ethanol (30 s), sterile water (3 min) and drying in sterilized filter paper. From 50 needles treated in this way, 180 fragments were excised and placed in the Petri dishes on malt extract agar supplemented with tetracycline to inhibit bacterial growth (MEA: 20 g/L malt extract, 15 g/L agar, Difco, Sparks, MD, USA; 200 mg/L tetracycline, TZF Polfa, Tarchomin, Poland). Petrie dishes were incubated in the dark for at least 10 weeks at 20 °C. Growing colonies were transferred to MEA for new dishes. For the initial identification of L. sulcigena cultures, experience obtained during previous research has proven to be useful [46,47].
2.4. DNA Extraction, PCR, Sequencing and Phylogenetic Analyses
To verify the morphology-based identification, the nucleotide sequences for four gene fragments of representative cultures were determined: 18S; the internal transcribed spacer regions ITS1 and ITS2, including the 5.8S gene (ITS); 28S region of the ribosomal RNA (rRNA) and translation elongation factor 1-α (TEF1). Genomic DNA extraction, polymerase chain reaction (PCR) amplification and sequencing reactions of the isolates were carried out according to the procedure described in detail by Kowalski et al. [48]. Primers used include NS1 and NS4 [49], ITS1-F [50] and ITS4 [49], LR0R [51] and LR6 [52], and EF1-983F and EF-gr [53]. The resulting ITS and 28S sequences were concatenated because they contained overlapping fragments. All obtained gene fragment sequences in this study were deposited in GenBank with the accession numbers shown in Table 1. The sequences obtained from representative cultures were used as queries in searches using the BLASTn algorithm [54] to retrieve sequences of taxa closely related to them from GenBank (http://www.ncbi.nlm.nih.gov, accessed on 29 December 2022) for phylogenetic analysis. Phylogenetic analyses were performed for concatenated ITS-28S-TEF1 sequences, using methods following Kowalski et al. [48]. The dataset contained all species of the family Rhytismataceae for which three gene sequences were available in the GenBank with Bisporella discedens MFLU 18-0691 included as an outgroup following Ekanayaka et al. [55]. The best evolutionary substitution model for the combined datasets was GTR + I + G.
3. Results
3.1. Symptoms
P. mugo needle disease in Chochołowska Valey in Tatra Mountains persisted throughout the study period, from 2015 to 2020. The disease affected the needles on the main and lateral shoots, mainly in the upper part of the crowns (Figure 1). Based on the analyses performed at different times of the year, it can be determined that the disease had the following course. Dieback symptoms initially appeared only on first-year needles, a few weeks after their development. From July to late summer, dying needles were generally reddish brown in color (Figure 2a). Symptoms occur on one or both needles in the bundle (Figure 2a–c). The distal parts of the needles died, while the basal parts, 0.3–0.8 (1.5) cm long, remained green. Since autumn, numerous needles have become white-gray (Figure 2b). In the following year, in June and July, ascomata continued to ripen on such previous year needles attached to the shoots (Figure 2c).
Other symptoms on some of the needles have also been detected. Transverse red-brown bands were often visible between the green living part and the dead part of the needle (Figure 2d). In turn, dark spots appeared on the dead parts of the needles (Figure 2e-g). They were particularly numerous on the needles of some shoots in the summer of 2016 (Figure 2e). In addition, transverse black stripes were formed on some needles at the periphery of zones with developed ascomata (Figure 2f). Basal parts of previous year’s needles began to die off in late summer and autumn which was followed by falling off of these needles.
The characteristics of the needles on the shoots and the severity of some symptoms in 2016 are given in Supplementary Materials Table S1. The density of the needles was higher on the main than on the lateral shoots. In total, needle discoloration was found in 65.8% of the previous year’s needles and in 77.8% of current year’s needles. In general, symptoms of fungal infection were slightly more common on only one needle in the bundle (Supplementary Materials Table S1). Great variation in the severity of the disease process between individual shoots was found. On the main shoots, the average length of discolored needles was significantly shorter than that of green needles. The type of discoloration covering more than ¾ of the length of the needle occurred in 60.7% of the previous year’s needles and in 59.7% of current year’s needles. These symptoms were slightly more common on the needles on the lateral shoots than on the main shoots. The vast majority of these needles showed a white-gray discoloration during the analyzed period. Hysterothecia of L. sulcigena were present on 41.6% of such needles from last year. In the autumn, the initial stage of such fruitbody could be seen on 3.4% of current year’s needles (Supplementary Materials Table S1).
The analyses of the width of annual rings on 35 shoots showed that 42.9% of them had a reduction in growth in 2016 compared to 2015 (Table 2). In the space of the following four years, the share of shoots with decrease in growth compared to the previous year varied from 56.7 to 65.7% (Table 2). The decrease in increment of individual shoots ranged from 2.3% to 72.6%, while the average value of decrease in increment for all shoots between individual years ranged from 20% to 25.8% (Table 2).
3.2. Morphology of Lophodermella sulcigena
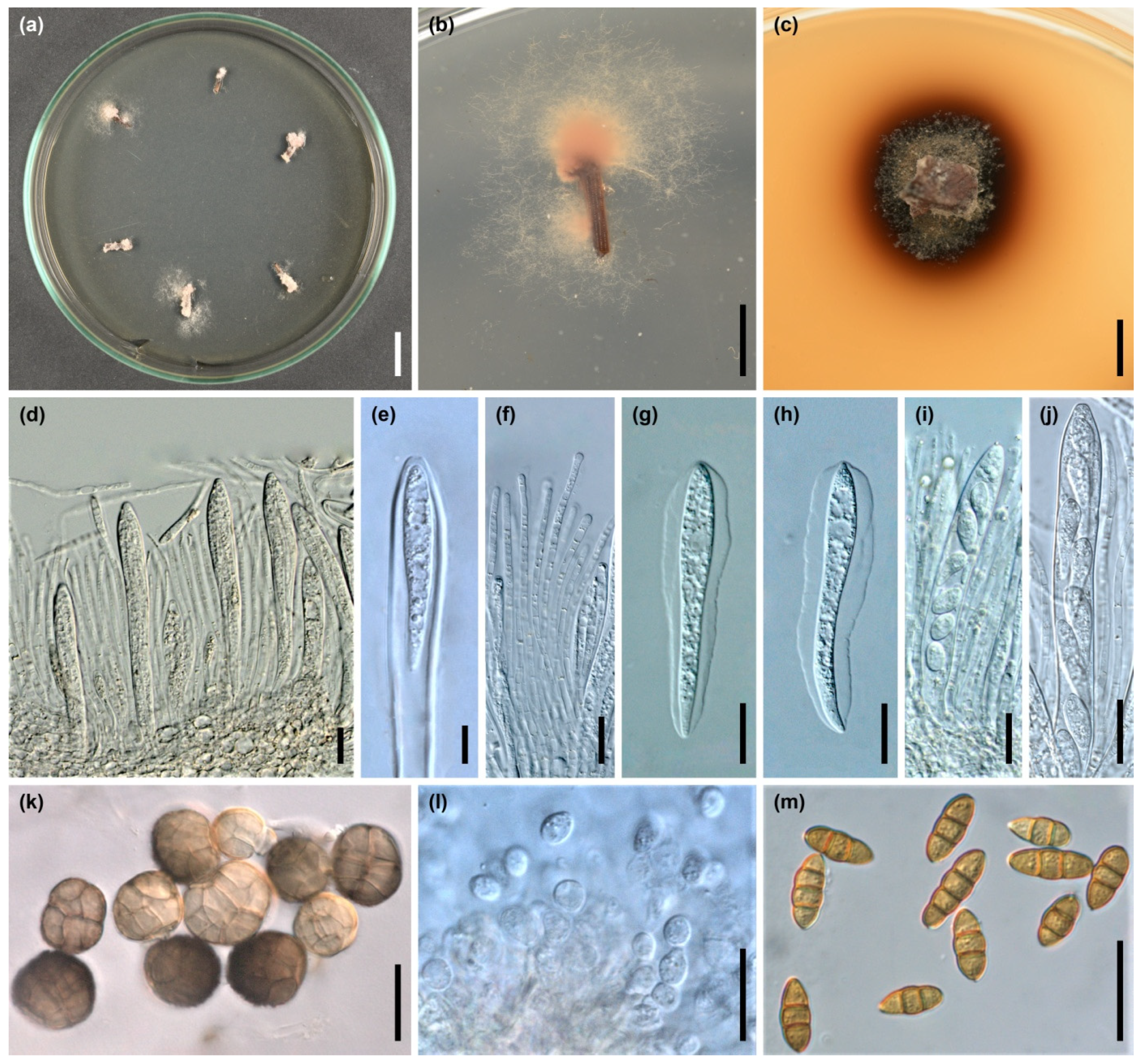
The basis for the identification of fungal species associated with disease symptoms on a morphological basis were hysterothecia (Figure 2h–j) and, to some extent, cultures isolated on agar medium (Figure 3a–c). Hysterothecia matured mainly in June–July on needles that had been infected the year before. They developed on both sides of the needles, on uniformly white-gray needles (Figure 2c) and those that showed numerous dark spots (Figure 2e–g). Hysterothecia were produced below the host hypodermis. They were elliptical or elongated in shape, 1.5–12 mm long, or longer if connected to each other, 0.3–0.5 mm wide and 0.16–0.3 mm deep. In mature ascocarps, a single longitudinal split appeared (Figure 2i,j), which is particularly well visible in the state of high humidity. Asci clavate, 110–140 × 12–15 µm (average 126.3 × 13.2 µm), 8 spored, rarely 4 spored (Figure 3d,e). Paraphyses filiform, unbranched, with numerous septa, 100–120 µm long, 2.0–3.0 µm wide, sometimes swollen at the tip, surrounded by a thin layer of mucus (Figure 3f). Ascospores clavate, tapering abruptly towards the base, straight or slightly curved, hyaline, often filled with granular content, 1-celled, 35–64 × 4.5–6.0 µm (average 45.5 × 4.7 µm), surrounded by a 2–4 µm thick gelatinous sheath, especially wider from both sides, much thinner on both ends of the spore (Figure 3g,h). Occasionally, single asci containing eight or six ascospores of different shapes and sizes were found within the hymenium. These were 1-celled, hyaline, subglobose, ellipsoidal, pyriform or clavate spores with dimensions of 16–33 × 8–10 µm (Figure 3i,j). Based on the morphological characteristics of the ascomata and dimensions of ascospores compared with literature (Table 3), the fungus causing the type of symptoms analyzed on P. mugo needles was identified as Lophodermella sulcigena. No other Lophodermella species were found during the research.
Out of 180 needle fragments obtained from needles with typical symptoms of the disease laid out on MEA in Petrie dishes, L. sulcigena colonies were obtained from only 20 (11.1%) fragments, mainly from current year’s needles without ascomata from the border of the living and dead parts. The first colonies were obtained in November 2015 (Figure 3a–c). The colonies were mainly concentrated on the pine needle fragment, while they showed very limited growth on the MEA (Figure 3a,b). After a fragment of the medium with mycelium was transferred to the new Petrie dishes, the colony either did not grow at all or again the growth was very limited, and the medium became dark brown after about 10 weeks (Figure 3c). Colonies were gray-brown with a flocculent structure, adjacent to the medium towards the periphery, and the reverse was gray-pink (Figure 3b). Aerial hyphae were hyaline to olive-brown, 2.0 to 7.5 µm thick with circular or oval, intercalary or apical, thin-walled swollen cells 6–17.5 µm in diameter. In the substrate, numerous olive-brown swollen cells, irregular or circular, 8–20.0 µm in diameter or elongated, 15–25 × 10–12.5 µm were observed. Sexual or asexual morph in vitro were not seen.
PCR resulted in fragments of 1132 base pairs (bp) for the 18S, 1655 bp for the appended ITS-28S, and 827 bp for the TEF1 gene regions of the subset of isolates sequenced. The sequences of the TEF1 fragment included only an exon without introns. All sequences within the analyzed gene fragments showed 100% similarity.
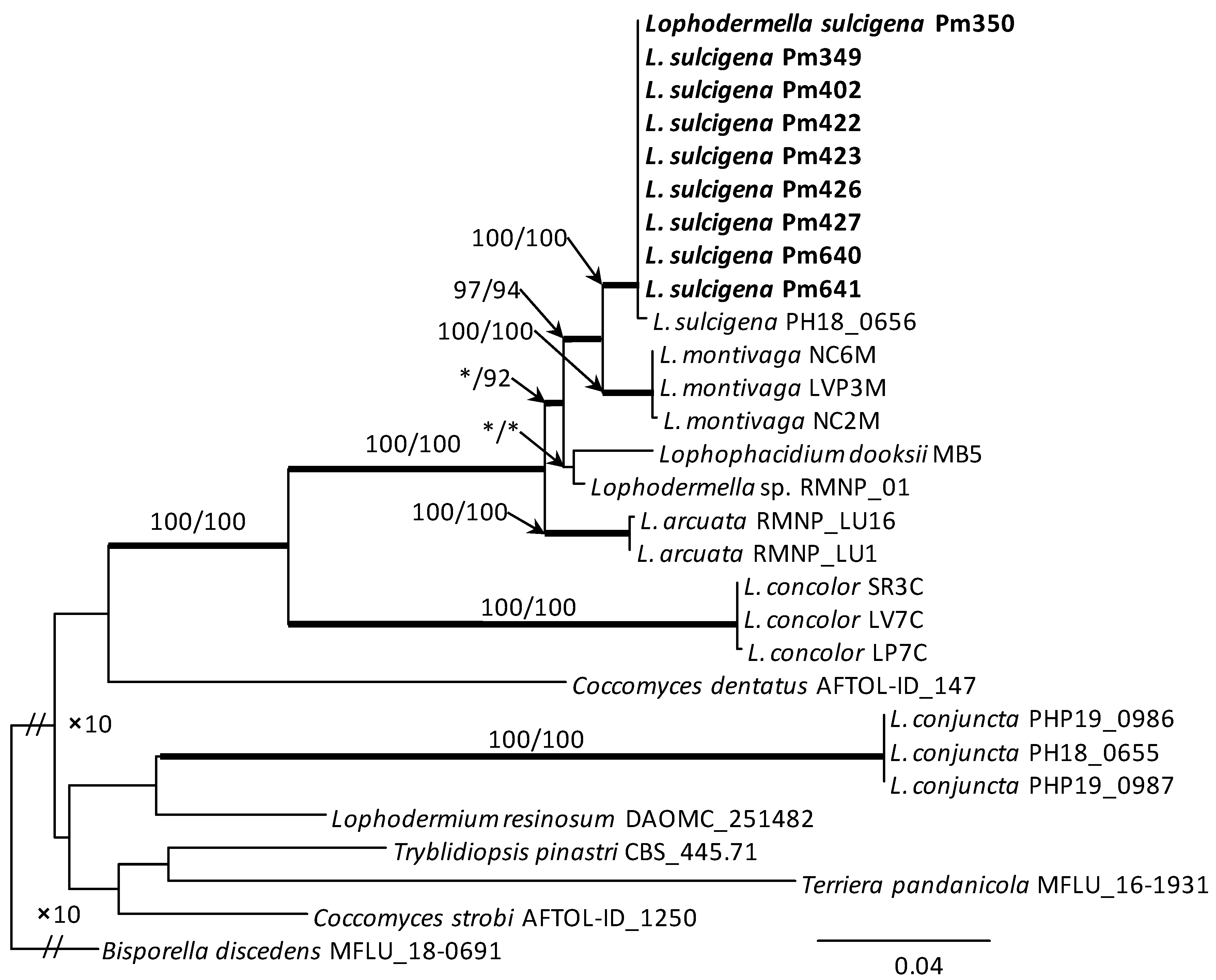
The aligned data set for the concatenated ITS-28S-TEF1 gene region used in phylogenetic analysis included 29 taxa and 2190 characters (with gaps). The conducted phylogenetic analysis, based on the data available in GenBank for representatives of the Rhytismataceae family, showed the high usefulness of this method for identifying closely related species forming it. Members of the genus Lophodermella together with Lophophacidium dooksii form a clade with strong statistical support. The only exception is Lophodermella conjuncta forming a separate clade. The sequences of L. sulcigena isolates form a distinct group on the phylogenetic tree. L. sulcigena shows the greatest phylogenetic similarity to Lophodermella montivaga, with which it forms a sister clade. The comparison of the DNA sequences of concatenated ITS-28S-TEF1 gene of selected isolates from Poland confirmed their high affinity to L. sulcigena PH18 0656 (Figure 4).
3.3. Secondary Fungi
Fruit bodies of other fungi were also found on the needles of P. mugo primary killed by L. sulcigena; the more common of them are listed in Table 4. Hendersonia sp. were found at various locations on the needle, while Leptomelanconium allescheri and Lophodermium species were more often present in the apical part of the needle, and this zone was often separated by a brown fuzzy stripe. The second needle in the bundle was not infected by L. sulcigena, which underwent secondary death; no Hendersonia sp. was found, and apart from Lophodermium species, asexual morph of Sydowia polyspora appeared (Table 4). Various fungi were also found inside hysterothecia developed on killed needles (Table 4). Some of them were found already in the early stage of hysterothecium development, when numerous ascospores were still present in the asci (Table 4, Figure 3k–m). In the upper part of the hymenium, sporodochia with conidia (Epicoccum nigrum) (Figure 2l and Figure 3k), stromatic colonies with torulose cells in a gelatinous matrix (asexual morph of Seuratia millardetii) (Figure 3l), or pycnidia with conidia (Hendersonia sp.) (Figure 3m) were identified. Some fungi were encountered in a later stage when ascospores from asci were mostly or all ejaculated. A cluster of their conidiophores and conidia could be observed outside the hysterothecia (Figure 2k).
4. Discussion
The results of this study have shown that a serious needle blight outbreak that occurred from 2015–2020 among P. mugo in high elevation forests of southern Poland was caused by L. sulcigena. Current analyses have not shown the occurrence of L. conjuncta in this area, despite the fact it was recorded both in the past [57] and recently on the needles of P. mugo in the Swiss Alps [58,59]. This species, unlike L. sulcigena, produces shorter hysterothecia and significantly longer ascospores [44,58]. Morphologically, L. sulcigena is very similar to L. montivaga found on pines in the USA. It was even speculated whether they were the same species [30,40,57]. However, recent molecular studies have shown that they are separate species [60]. Also, the current results indicate that L. sulcigena shows the greatest phylogenetic similarity to L. montivaga.
It was noticed relatively long ago that the most destructive of the Lophodermella species are those capable of completing their life cycles in one year, and L. sulcigena was included in this group [40]. L. sulcigena has so far been observed in Europe on two-needle pines: P. mugo, P. nigra, and Pinus sylvestris [41,44,57,58,59,61,62,63,64,65]. In Poland, this is the first observation of this pathogen on P. mugo. So far, L. sulcigena has been found in Poland only on P. sylvestris [46,47,66].
The currently observed symptoms on P. mugo were generally very similar to those of L. sulcigena on other pine species [41,44,62,64]. However, it has now been observed that the infestation of needles by L. sulcigena was occasionally accompanied by an unusually high density of dark spots, which was not observed to such an extent in P. sylvestris needles [46,62]. According to Campbell [67], the small dark areas on the needle are the result of the accumulation of dark hyphae of L. sulcigena which develops later into the hysterothecium. Such dark spots were also found on needles affected by L. conjuncta [57].
Other authors [32] observed the transverse black bands at the edge of the fruiting areas on P. contorta infected by L. montivaga. Such symptoms have also been reported now, but sporadically. Needles infected by L. sulcigena take on a white-gray color. However, this is not unique to this pathogen. Such discoloration accompanies the infection of needles by Leptomelanconium allescheri as well [29]. Also secondary fungi, e.g., Hendersonia acicola, contribute to such a change in color of the needles [44,68]. Different reactions between various tree species are also observed in case of other plant pathogens. For example, needles infected by Dothistroma septosporum usually develop red bands, but depending on the host species and on the environment, in which the tree grows, may have a different color [69,70,71]. This is also reflected in different in vitro cultivation conditions [72].
It is generally known that Lophodermella does not grow well in agar culture, even with the addition of pine needle extract, and attempts to obtain pure cultures from single-ascospore isolations have failed [44,60]. Due to this fact, some authors classify them as fungi with potentially obligate lifestyle [60]. However, it was quite often possible to isolate L. sulcigena on agar medium from symptomatic needles of P. sylvestris [47,62]. The present study confirmed that it was possible to obtain cultures of L. sulcigena from P. mugo needles, although the growth of L. sulcigena on MEA was very limited. However, it should be concluded that the isolation of L. sulcigena on agar media cannot be the basis for the assessment of the frequency of this pathogen in needles.
Most of the morphological features of L. sulcigena on P. mugo in Polish Tatra were consistent with previous observations by other authors [41,43,44,58]. However, there are some differences in the length of ascospores. According to some authors, ascospores reach up to 35–40 µm in length [14,42,43], while according to other authors, ascospores reach up to 50–65 µm in length [41,46,56,58]. These differences may indicate that there are two subpopulations of this pathogen in Europe. However, it cannot be excluded that the different data are the result of analyses of the pathogen at different stages of development. In addition, the measurement results can be affected by the use of different mounting media and the measurement of dried but not living samples [73,74]. The present study also indicated that L. sulcigena hysterothecia may contain asci with eight or six ascospores of different shapes and sizes. This suggests that there may be some disturbance in the formation of spores. Terrier [57] stated that in some L. sulcigena asci instead of eight ascospores there were four because the other four degenerated. Instability of sporogenesis was also sometimes observed in other fungal species [75]. For example, in some Pezicula species, most asci contain eight spores, and some contain six or four spores. There are known cases that the four spores were not degenerated but showed spores smaller than usual. Verkley [75] considers that these changes may be influenced by adverse environmental conditions or some kind of genetic degeneration.
Possibilities of comparing L. sulcigena strains on a genetic basis are limited so far [60,76]. For the first time, one L. sulcigena sequence from P. sylvestris was deposited in 2021 [60]. These data have been enriched as a result of the present study with nine sequences from P. mugo growing in the Polish Tatra, obtained both on the basis of cultures and direct DNA isolation from symptomatic needles. Enlarging the collections of this type of data from different regions of Europe and different plant hosts should enable the analysis of the genetic diversity of this pathogen. Genetic analysis of another dangerous pathogen of Pinus species, Gremmeniella abietina, in Poland has shown that mountain populations differ from lowland populations [77].
Based on the results of the phylogenetic analysis, it can be concluded that the genus Lophodermella is not monophyletic. Lophodermella conjuncta individuals form a well-defined and externally located clade in relation to other species included in the genus Lophodermella. A similar conclusion can be drawn from the work of Ata et al. [60]. This may indicate the need for changes in the taxonomy of species currently belonging to the genus Lophodermella.
Numerous fungal species associated with needles infected primary by L. sulcigena are known. On P. sylvestris, it is mainly Hendersonia acicola [44,47,68,78]. On P. nigra, heavy secondary colonization was shown by Lophodermium seditiosum and Lo. conigenum [44]. The currently observed colonization of Lo. conigenum and Lo. corconticum on P. mugo needles already infected by L. sulcigena may have been facilitated by the fact that both species occur in symptomless needles as endophytes [28]. In turn, Leptomelanconium allescheri is rather not dependent on the attack of other primary pathogens, as it can independently cause local necrotic lesion [29]. Hendersonia acicola and H. pinicola, through the secondary invasion of needles, may prevent the fructification of some Lophodermella species, which may contribute to the biological elimination of the disease [44,61,68,79]. On needles of P. sylvestris infected by L. sulcigena, the fungus Epicoccum nigrum was six times more common than in symptomless needles [47]. This fungal species, now found for the first time in L. sulcigena hysterothecia, is known for antifungal activities. It produces, i.al., flavipin, epicorazine A, epicorazine B, epipyrone and fungal cell wall degrading enzymes which take part in lysis of pathogen hyphae [80,81,82]. Also known among the Cladosporium genus are mycoparasites, as well as species producing various antibiotics with antifungal properties [83,84]. Another fungus, Seuratia millardetii, found presently in asexual morph, is a representative of the sooty mold that mostly grows superficially on plant tissue [45]. It can be assumed that the numerous gelatinous structures of this species found inside L. sulcigena hysterothecium may limit the spread of the pathogen’s ascospores. Reducing the development of L. sulcigena by other fungi would be of great ecological importance, as L. sulcigena contributes to significant losses in attacked pine cultures [68]. Mitchell et al. [78] showed that the volume of diseased Corsican pine trees was reduced by 59% following a six-year epidemic. The reduction of the radial increment of P. mugo shoots was also confirmed in the present study. This was probably also influenced by other less frequent fungi on the needles of P. mugo [28,29].
5. Conclusions
For several years, a high intensity of P. mugo needle disease has been observed in one of the areas of its natural occurrence in the Tatra Mts. Previous preliminary studies showed that the mycobiota on diseased and dead needles of P. mugo is quite diverse. The current work was devoted to explaining the causal agent of the most common disease symptom on P. mugo needles. It was characterized by the fact that current year’s needles died a few weeks after development, with their basal parts remaining green. The following year, numerous mature fungal fruit bodies (hysterothecia) appeared on such needles in early summer. Later, after the death of the basal parts, such needles gradually fell. Morphological analyses of microstructures of fungal fruit bodies produced on needles together with the use of molecular techniques showed that Lophodermella sulcigena is the cause of these symptoms. No other species of this genus were found on the examined needles. This is the first record of L. sulcigena on P. mugo in Poland. So far, it has only been found locally on P. sylvestris. Careful analysis of diseased needles was necessary for two main reasons: (a) symptoms on P. mugo needles were slightly different compared to P. sylvestris, especially regarding the intensity of dark spots, and (b) data on the dimensions of L. sulcigena ascospores are contradictory in the literature. The present study has provided further evidence of variation within L. sulcigena in terms of the presence of different numbers of spores in one ascus and the occurrence of ascospores of substantially different shapes and sizes. This is important data, as identification of this species can sometimes be problematic. In the course of the current research, cases of the presence of other, hitherto unknown fungi in the fruit body of L. sulcigena have been found. This creates an interesting direction of research on the possibilities of biological protection of P. mugo against L. sulcigena.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f15030422/s1, Table S1: Characteristics of Pinus mugo needles with disease symptoms from previous year and current year, studied in 2016 in Chochołowska Valley (Tatra Mountains).
Author Contributions
Conceptualization, T.K., P.B. and C.B.; methodology, C.B. (field studies and sampling), T.K. (mycological aspects), and P.B. (molecular aspects); investigation, T.K., P.B. and C.B.; data curation, T.K. and P.B.; writing—original draft preparation, review and editing T.K. and P.B.; software, P.B.; visualization T.K. and P.B. All authors have read and agreed to the published version of the manuscript.
Funding
The research was partly financed by a subsidy from the forest fund transferred by the State Forests to the Tatra National Park in 2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the NCBI GenBank database.
Acknowledgments
The authors thank T. Zwijacz-Kozica for enabling field research in the National Park.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ballian, D.; Ravazzi, C.; de Rigo, D.; Caudullo, G. Pinus mugo in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; European Commission: Luxemburg, 2016; pp. 124–125. [Google Scholar]
- Bączkiewicz, A.; Prus-Głowacki, W. Variability of Pinus mugo Turra clones from Ostry Wierch peat bog. Acta Soc. Bot. Pol. 1997, 66, 79–82. [Google Scholar] [CrossRef]
- Hamerník, J.; Musil, I. The Pinus mugo complex—Its structuring and general overview of the used nomenclature. J. For. Sci. 2007, 53, 253–266. [Google Scholar] [CrossRef]
- Sokołowska, J.; Fuchs, H.; Celiński, K. New insight into taxonomy of European mountain pines, Pinus mugo complex, based on complete chloroplast genomes sequencing. Plants 2021, 10, 1331. [Google Scholar] [CrossRef] [PubMed]
- Boratyńska, K. Needle variability of Pinus mugo Turra in the West Tatra Mts. Dendrobiology 2002, 48, 3–8. [Google Scholar]
- Dirnböck, T.; Grabherr, G. GIS assessment of vegetation and hydrological change in a high mountain catchment of the northern limestone alps. Mt. Res. Dev. 2000, 20, 172–179. [Google Scholar] [CrossRef]
- Švajda, J.; Solár, J.; Janiga, M.; Buliak, M. Dwarf pine (Pinus mugo) and selected abiotic habitat conditions in the Western Tatra Mountains. Mt. Res. Dev. 2011, 31, 220–228. [Google Scholar] [CrossRef]
- Bitterli, P. Jahrringe in Legföhren (Pinus mugo Turra ssp. mugo ) als Ausdruck lokaler mechanischer Faktoren. Dendrochronologia 1987, 5, 79–95. [Google Scholar]
- Innes, J.L.; Schwyzer, A. Stem damage in Swiss forests: Incidence, causes and relations to crown transparency. For. Pathol. 1994, 24, 20–31. [Google Scholar] [CrossRef]
- Bencheva, S.; Doychev, D. Winter desiccation of dwarf pine (Pinus mugo Turra) needles in the area of Belmeken Dam. Silva Balc. 2022, 23, 79–88. [Google Scholar] [CrossRef]
- Mayr, S.; Schmid, P.; Rosner, S. Winter embolism and recovery in the conifer shrub Pinus mugo L. Forests 2019, 10, 941. [Google Scholar] [CrossRef]
- Charrier, G.; Martin-StPaul, N.; Damesin, C.; Delpierre, N.; Hänninen, H.; Torres-Ruiz, J.M.; Davi, H. Interaction of drought and frost in tree ecophysiology: Rethinking the timing of risks. Ann. For. Sci. 2021, 78, 40. [Google Scholar] [CrossRef]
- Schneider, M.; Grünig, C.R.; Holdenrieder, O.; Sieber, T.N. Cryptic speciation and community structure of Herpotrichia juniperi, the causal agent of brown felt blight of conifers. Mycol. Res. 2009, 113, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Butin, H. Krankheiten der Wald- und Parkbäume; Ulmer Verlag: Stuttgart, Germany, 2011; ISBN 978-3-8001-7636-6. [Google Scholar]
- Sieber, T.N. The endophyte Allantophomopsis cytisporea is associated with snow blight on Calluna vulgaris in the Alps—An effect of climate change? Arctic, Antarct. Alp. Res. 2019, 51, 460–470. [Google Scholar] [CrossRef]
- Dobbertin, M.; Baltensweiler, A.; Rigling, D. Tree mortality in an unmanaged mountain pine (Pinus mugo var. uncinata) stand in the Swiss National Park impacted by root rot fungi. For. Ecol. Manag. 2001, 145, 79–89. [Google Scholar] [CrossRef]
- Cherubini, P.; Fontana, G.; Rigling, D.; Dobbertin, M.; Brang, P.; Innes, J.L. Tree-life history prior to death: Two fungal root pathogens affect tree-ring growth differently. J. Ecol. 2002, 90, 839–850. [Google Scholar] [CrossRef]
- Hintsteiner, M.; Cech, T.L.; Halmschlager, E.; Stauffer, C.; Kirisits, T. First report of Mycosphaerella dearnessii on Pinus nigra var. nigra in Austria. For. Pathol. 2012, 42, 437–440. [Google Scholar] [CrossRef]
- Holdenrieder, O.; Sieber, T.N. First report of Mycosphaerella dearnessii in Switzerland. For. Pathol. 1995, 25, 293–295. [Google Scholar] [CrossRef]
- La Porta, N.; Capretti, P. Mycosphaerella dearnessii, a needle-cast pathogen on mountain pine (Pinus mugo) in Italy. Plant Dis. 2000, 84, 922. [Google Scholar] [CrossRef]
- Jankovský, L.; Palovčíková, D.; Dvořák, M.; Tomšovský, M. Records of Brown spot needle blight related to Lecanosticta acicola in the Czech Republic. Plant Prot. Sci. 2009, 45, 16–18. [Google Scholar] [CrossRef]
- Boroń, P.; Lenart-Boroń, A.; Mullett, M. The distribution of Dothistroma septosporum and its mating types in Poland. For. Pathol. 2016, 46, 489–496. [Google Scholar] [CrossRef]
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.E.; Vahalík, P.; Mullett, M.S.; Martín-García, J.; Bulman, L.S.; Wingfield, M.J.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: A comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- Ondrušková, E.; Hečková, Z.; Horáková, M.K.; Koltay, A.; Ostrovský, R.; Pažitný, J.; Adamčíková, K. Distribution and characterization of Dothistroma needle blight pathogens on Pinus mugo in Slovakia. Eur. J. Plant Pathol. 2017, 148, 283–294. [Google Scholar] [CrossRef]
- Cleary, M.; Laas, M.; Oskay, F.; Drenkhan, R. First report of Lecanosticta acicola on non-native Pinus mugo in southern Sweden. For. Pathol. 2019, 49, e12507. [Google Scholar] [CrossRef]
- Sadiković, D.; Piškur, B.; Barnes, I.; Hauptman, T.; Diminić, D.; Wingfield, M.J.; Jurc, D. Genetic diversity of the pine pathogen Lecanosticta acicola in Slovenia and Croatia. Plant Pathol. 2019, 68, 1120–1131. [Google Scholar] [CrossRef]
- Skuhravý, V.; Skuhravá, M. Zur Verbreitung und Schädlichkeit der Gallmücken (Cecidomyiidae, Diptera) an Waldbäumen in Mitteleuropa. Anz. Schädlingskd. Pflanzenschutz Umweltschutz 1993, 66, 134–140. [Google Scholar] [CrossRef]
- Bartnik, C.; Kowalski, T.; Bilański, P.; Zwijacz-Kozica, T. Fungi associated with disease symptoms on Pinus mugo needles in the Polish Tatra Mountains. Plant Fungal Syst. 2021, 66, 53–65. [Google Scholar] [CrossRef]
- Kowalski, T.; Boroń, P.; Bartnik, C.; Rossa, R. Morphological and molecular characterization of Leptomelanconium allescheri associated with necrotic lesions on Pinus mugo needles in the Polish Tatra Mountains. For. Pathol. 2018, 48, e12420. [Google Scholar] [CrossRef]
- Funk, A. Foliar Fungi of Western Trees. Information Report BC-X-265; Agriculture Canada, Minister of Supply & Services Canada, Pacific Forest Research Centre: Victoria, BC, Canada, 1985. [Google Scholar]
- Sinclair, W.A.; Lyon, H.H. Diseases of Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 2005; ISBN 978-0-8014-4371-8. [Google Scholar]
- Worrall, J.; Marchetti, S.; Mask, R. An Epidemic of Needle Cast on Lodgepole Pine in Colorado. Biological Evaluation R2-12-01; USDA Forest Service, Rocky Mountain Region, Forest Health Protection: Gunnison, CO, USA, 2012; pp. 1–16. [Google Scholar]
- Broders, K.; Munck, I.; Wyka, S.; Iriarte, G.; Beaudoin, E. Characterization of fungal pathogens associated with White Pine Needle Damage (WPND) in Northeastern North America. Forests 2015, 6, 4088–4104. [Google Scholar] [CrossRef]
- Lutyk, P. Diseases of the mountain pine (Pinus mughus Scop.) on the area of the Tatra National Park. Stan zdrowotny sosny kosówki (Pinus mughus Scop.) na terenie Tatrzańskiego Parku Narodowego. Sylwan 1978, 122, 51–57. [Google Scholar]
- Koukol, O.; Pusz, W.; Minter, D. A new species of Lophodermium on needles of mountain pine (Pinus mugo) from the Giant Mountains in Poland. Mycol. Prog. 2015, 14, 23. [Google Scholar] [CrossRef]
- Pusz, W.; Zwijacz-Kozica, T.; Kita, W. Assessment of the mountain pine (Pinus mugo) needles health at selected locations in the Tatra National Park. Sylwan 2015, 159, 411–418. [Google Scholar]
- TIBCO Software Inc. Data Science Workbench. Statistica Version 13.1. 2020. Available online: http://tibco.com (accessed on 15 September 2020).
- Cybis Electronik & Data AB. Cybis CooRecoder—Image Coordinate Recording Program Version:7.6. 2012. Available online: http://www.cybis.se (accessed on 15 September 2020).
- Cybis Electronik & Data AB. CDendro 7.6 TEST—Cybis Dendro Dating Program Version:7.6. 2012. Available online: http://www.cybis.se (accessed on 15 September 2020).
- Darker, G.D. The Hypodermataceae of Conifers; Jamaica Plain, Mass; The Arnold Arboretum of Harvard University: Boston, MA, USA, 1932; Volume 1. [Google Scholar]
- Moriondo, F. La diffusione della ruggine curvatrice nelle pinete italiane. Ann. Accad. Ital. Sci. 1963, 11, 247–263. [Google Scholar]
- Dennis, R.W.G. British Ascomycetes; Verlag von J. Cramer: Lehre, Germany, 1968. [Google Scholar]
- Millar, C.; Minter, D. Lophodermella sulcigena. In CMI Descriptions of Pathogenic Fungi and Bacteria; Commonwealth Mycological Institute: Kew, UK, 1978; pp. 45–55. [Google Scholar]
- Millar, C.S. Lophodermella species on pines. In Proceedings of the Recent Research on Conifer Needle Diseases; Peterson, G.W., Ed.; General Technical Report GTR-WO 50; USDA Forest Service: Gulfport, MS, USA, 1984; pp. 45–55. [Google Scholar]
- Gillis, I.; Glawe, D.A. Characterization of Seuratia millardetii on Camellia species and in artificial culture. N. Am. Fungi 2008, 3, 215–229. [Google Scholar] [CrossRef]
- Kowalski, T. Erstnachweis von Lophodermella sulcigena in Polen. For. Pathol. 1988, 18, 445–447. [Google Scholar] [CrossRef]
- Kowalski, T.; Krygier, J. Mycological study on symptomless and diseased needles in pine stand attacked by Lophodermella sulcigena. Phytopathol. Pol. 1996, 11, 159–168. [Google Scholar]
- Kowalski, T.; Bilański, P.; Grad, B. The occurrence of Apiognomonia hystrix and its pathogenicity towards Acer pseudoplatanus and Fraxinus excelsior under field conditions. Forests 2022, 13, 35. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R. Conserved Primer Sequences for PCR Amplification and Sequencing from Nuclear Ribosomal RNA. Available online: https://sites.duke.edu/vilgalyslab/files/2017/08/rDNA-primers-for-fungi.pdf (accessed on 20 January 2023).
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ekanayaka, A.; Hyde, K.; Gentekaki, E.; McKenzie, E.; Zhao, Q.; Bulgakov, T.; Camporesi, E. Preliminary classification of Leotiomycetes. Mycosphere 2019, 10, 310–489. [Google Scholar] [CrossRef]
- Lagerberg, T. Om grabarrsjukan host allen, dess orsak och verkningar. Medd. Från Statens Skogsförsöksanstalt 1910, 7, 127–174. [Google Scholar]
- Terrier, C. Über zwei in der Schweiz bisher wenig bekannte Schiittepilze der Kiefern: Hypodermella sulcigena (Rostr.) v.Tub. und Hypodermella conjuncta Darker. Phytopathol. Z. 1944, 14, 442–449. [Google Scholar]
- Beenken, L. Lophodermella-Nadelschütte. In Waldschutzüberblick 2018; Queloz, V., Forster, B., Beenken, L., Stroheker, S., Odermatt, O., Hölling, D., Meyer, J., Dubach, V., Eds.; Forschungsanstalt WSL: Birmensdorf, Switzerland, 2019; pp. 18–19. [Google Scholar]
- Dubach, V.; Queloz, V.; Stroheker, S. Needle and shoot diseases of pine. WSL Fact Sheet 2022, 70, 1–12. [Google Scholar]
- Ata, J.P.; Burns, K.S.; Marchetti, S.; Munck, I.A.; Beenken, L.; Worrall, J.J.; Stewart, J.E. Molecular characterization and phylogenetic analyses of Lophodermella needle pathogens (Rhytismataceae) on Pinus species in the USA and Europe. PeerJ 2021, 9, e11435. [Google Scholar] [CrossRef] [PubMed]
- Darker, G.D. A revision of the genera of the Hypodermataceae. Can. J. Bot. 1967, 45, 1399–1444. [Google Scholar] [CrossRef]
- Jalkanen, R. The occurrence and importance of Lophodermella sulcigena and Hendersonia acicola on Scots pine in Finland. Karstenia 1985, 25, 53–61. [Google Scholar] [CrossRef]
- Ranta, H.; Saloniemi, I. Distribution of fungal foliage and shoot pathogens in a natural Scots pine population in relation to environmental variables. Can. J. For. Res. 2005, 35, 503–510. [Google Scholar] [CrossRef]
- Karadžić, D.; Milijašević, T. The most important parasitic and saprophytic fungi in Austrian pine and Scots pine plantations in Serbia. Bull. Fac. For. 2008, 97, 147–170. [Google Scholar]
- Cech, T.L. Österreich: Phytopathologische Notizen 2009. Forstsch. Aktuell 2009, 47, 6–8. [Google Scholar]
- Sadłowski, W.; Kowalski, T. Variability of the Lophodermella sulcigena infection severity in a young pine stand. In Shoot and foliage diseases in forest trees, Proceedings of the Joint Meeting of the IUFRO Working Parties S2.06.02 and S2.06.04, Florence, Italy, 6–11 June 2014; Capretti, P., Heinige, U., Stephan, R., Eds.; Istituto di Patologia e Zoologia Forestale e Agraria: Florence, Italy, 1994; pp. 51–56. [Google Scholar]
- Campbell, R. Ultrastructure of asci, ascospores, and spore release in Lophodermella sulcigena (Rostr.) v. Höhn. Protoplasma 1973, 78, 69–80. [Google Scholar] [CrossRef]
- Jalkanen, R.; Laakso, R. Hendersonia acicola in an epidemic caused by Lophodermella sulcigena with special reference to biological control. Karstenia 1986, 26, 49–56. [Google Scholar] [CrossRef]
- Pehl, L.; Butin, H. Dothistroma septospora, ein neuer Schadpilz an der Bergkiefer. Allg. Forst Z. 1992, 14, 758–760. [Google Scholar]
- Kirisits, T.; Cech, T. Auffälliges Vorkommen der Dothistroma-Nadelbräune an Zirben im oberen Murtal. Forstsch. Aktuell 2007, 41, 13–15. [Google Scholar]
- Drenkhan, R.; Hanso, M. Recent invasion of foliage fungi of pines (Pinus spp.) to the Northern Baltics. For. Stud. 2009, 51, 49–64. [Google Scholar] [CrossRef]
- Kowalski, T.; Nawrot-Chorabik, K.; Grad, B.; Leszczyński, K. Influence of culture conditions on medium discoloration and mycelial growth of Dothistroma septosporum. For. Pathol. 2016, 46, 507–514. [Google Scholar] [CrossRef]
- Baral, H.O. Vital versus herbarium taxonomy: Morphological differences between living and dead cells of ascomycetes, and their taxonomic implications. Mycotaxon 1992, 45, 333–390. [Google Scholar]
- Brown, P.A. A review of techniques used in the preparation, curation and conservation of microscope slides at the Natural History Museum, London. Biol. Curator 1997, 10, 1–34. [Google Scholar]
- Verkley, G.J.M. A Monograph of the Genus Pezicula and Its Anamorphs; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 1999; Volume 44, pp. 1–180. [Google Scholar]
- Robert, V.; Vu, D.; Amor, A.B.H.; van de Wiele, N.; Brouwer, C.; Jabas, B.; Szoke, S.; Dridi, A.; Triki, M.; ben Daoud, S.; et al. MycoBank gearing up for new horizons. IMA Fungus 2013, 4, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Kraj, W.; Kowalski, T. Genetic variation in Polish strains of Gremmeniella abietina. For. Pathol. 2008, 38, 203–217. [Google Scholar] [CrossRef]
- Mitchell, C.P.; Millar, C.S.; Haworth, M.N. Effect of the needle-cast fungus Lophodermella sulcigena on growth of Corsican pine. Forestry 1976, 49, 153–158. [Google Scholar] [CrossRef]
- Mitchell, C.P.; Williamson, B.; Millar, C.S. Hendersonia acicola on pine needles infected by Lophodermella sulcigena. For. Pathol. 1976, 6, 92–102. [Google Scholar] [CrossRef]
- Brown, A.E.; Finlay, R.; Ward, J.S. Antifungal compounds produced by Epicoccum purpurascens against soil-borne plant pathogenic fungi. Soil Biol. Biochem. 1987, 19, 657–664. [Google Scholar] [CrossRef]
- Madrigal, C.; Tadeo, J.L.; Melgarejo, P. Relationship between flavipin production by Epicoccum nigrum and antagonism against Monilinia laxa. Mycol. Res. 1991, 95, 1375–1381. [Google Scholar] [CrossRef]
- Lee, A.J.; Cadelis, M.M.; Kim, S.H.; Swift, S.; Copp, B.R.; Villas-Boas, S.G. Epipyrone A, a broad-spectrum antifungal compound produced by Epicoccum nigrum ICMP 19927. Molecules 2020, 25, 5997. [Google Scholar] [CrossRef] [PubMed]
- Assante, G.; Maffi, D.; Saracchi, M.; Farina, G.; Moricca, S.; Ragazzi, A. Histological studies on the mycoparasitism of Cladosporium tenuissimum on urediniospores of Uromyces appendiculatus. Mycol. Res. 2004, 108, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wedge, D.E.; Cutler, S.J. Chemical and biological study of cladosporin, an antimicrobial inhibitor: A review. Nat. Prod. Commun. 2016, 11, 1595–1600. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Pinus mugo in Chochołowska Valley in Tatra Mountains, general view of infested thickets (July 2016).
Figure 1.
Pinus mugo in Chochołowska Valley in Tatra Mountains, general view of infested thickets (July 2016).

Figure 2.
Symptoms on needles of Pinus mugo and etiological signs of Lophodermella sulcigena (a) reddish-brown current year needles (August 2017), (b) partly white-gray discolored previous year needles (May 2016), (c) hysterothecia on white-gray needles (July 2016), (d) red-brown band between living and dead parts of the needle (arrow), (e) numerous dark spots on dead distal parts of the needles, (f) black transverse stripe on dead part (arrow), (g) hysterothecium developed within dark spot (arrow), (h–j) hysterothecia: in dry condition (arrow) (h), in humid conditions (arrows) (i), after ejecting most of the ascospores (arrows) (j), (k,l) secondary fungi in/on hysterothecia: Cladosporium herbarum conidiophores and conidia (arrows) (k), Epicoccum nigrum sporodochia (arrow) (l). Scale bars: (a–c) = 1 cm, (d,e) = 0.2 cm, (f,g) = 0.1 cm, (h) = 0.2 cm, (i,j) = 0.1 cm, (k) = 0.2 cm, (l) = 0.1 cm.
Figure 2.
Symptoms on needles of Pinus mugo and etiological signs of Lophodermella sulcigena (a) reddish-brown current year needles (August 2017), (b) partly white-gray discolored previous year needles (May 2016), (c) hysterothecia on white-gray needles (July 2016), (d) red-brown band between living and dead parts of the needle (arrow), (e) numerous dark spots on dead distal parts of the needles, (f) black transverse stripe on dead part (arrow), (g) hysterothecium developed within dark spot (arrow), (h–j) hysterothecia: in dry condition (arrow) (h), in humid conditions (arrows) (i), after ejecting most of the ascospores (arrows) (j), (k,l) secondary fungi in/on hysterothecia: Cladosporium herbarum conidiophores and conidia (arrows) (k), Epicoccum nigrum sporodochia (arrow) (l). Scale bars: (a–c) = 1 cm, (d,e) = 0.2 cm, (f,g) = 0.1 cm, (h) = 0.2 cm, (i,j) = 0.1 cm, (k) = 0.2 cm, (l) = 0.1 cm.

Figure 3.
Colonies and microstructures of Lophodermella sulcigena and secondary fungi: (a) colonies growing from needle fragments (8 weeks, MEA, 20 °C), (b) reverse of two colonies, (c) 10-week old subculture on MEA, (d–j)—microstructures of L. sulcigena: fragment of hymenium with asci and paraphyses (d), ascus with one unejected ascospore (e), paraphyses (f), ascospores with gelatinous sheath (g,h), ascus with six unusually shaped ascospores (i), ascus with eight unusually shaped ascospores (j), (k,l)—secondary fungi in hysterothecia of L. sulcigena: Epicoccum nigrum conidia (k), Seuratia millardetii asexual stage, conidia and torulose cells in gelatinous matrix (l), Hendersonia sp. conidia (m). Scale bars: (a–c) = 1 cm, (d) = 20 µm, (e) = 10 µm, (f) = 20 µm, (g,h) = 10 µm, (i–m) = 20 µm.
Figure 3.
Colonies and microstructures of Lophodermella sulcigena and secondary fungi: (a) colonies growing from needle fragments (8 weeks, MEA, 20 °C), (b) reverse of two colonies, (c) 10-week old subculture on MEA, (d–j)—microstructures of L. sulcigena: fragment of hymenium with asci and paraphyses (d), ascus with one unejected ascospore (e), paraphyses (f), ascospores with gelatinous sheath (g,h), ascus with six unusually shaped ascospores (i), ascus with eight unusually shaped ascospores (j), (k,l)—secondary fungi in hysterothecia of L. sulcigena: Epicoccum nigrum conidia (k), Seuratia millardetii asexual stage, conidia and torulose cells in gelatinous matrix (l), Hendersonia sp. conidia (m). Scale bars: (a–c) = 1 cm, (d) = 20 µm, (e) = 10 µm, (f) = 20 µm, (g,h) = 10 µm, (i–m) = 20 µm.

Figure 4.
Phylogram from Maximum Likelihood (ML) analyses of the combined datasets of ITS + 28S + TEF1 for selected Rhytismataceae. Sequences obtained in this study are indicated with bold text. Bootstrap values ≥ 75% for ML and Maximum Parsimony (MP) analyses are presented at nodes as follows: ML/MP. Bold branches indicate posterior probability values ≥ 0.95 obtained from Bayesian Inference (BI) analyses. * Bootstrap values < 75%. The tree is drawn to scale (see bar) with branch lengths measured in the number of substitutions per site. Bisporella discedens was used as outgroup taxon.
Figure 4.
Phylogram from Maximum Likelihood (ML) analyses of the combined datasets of ITS + 28S + TEF1 for selected Rhytismataceae. Sequences obtained in this study are indicated with bold text. Bootstrap values ≥ 75% for ML and Maximum Parsimony (MP) analyses are presented at nodes as follows: ML/MP. Bold branches indicate posterior probability values ≥ 0.95 obtained from Bayesian Inference (BI) analyses. * Bootstrap values < 75%. The tree is drawn to scale (see bar) with branch lengths measured in the number of substitutions per site. Bisporella discedens was used as outgroup taxon.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fungal isolates obtained in the present study from Pinus mugo in Chochołowska Valley (Tatra Mountains).
Table 1.
Fungal isolates obtained in the present study from Pinus mugo in Chochołowska Valley (Tatra Mountains).
| Species | Isolate Number 1 | Collection Date | GenBank Accession Number | ||
|---|---|---|---|---|---|
| 18S | ITS-28S | TEF1 | |||
| Lophodermella sulcigena | Pm349 | 17 May 2016 | OQ288928 | OQ288965 | OQ303894 |
| Pm350 | 17 May 2016 | OQ288929 | OQ288966 | OQ303895 | |
| Pm402 1 | 10 November 2015 | OQ288930 | OQ288967 | OQ303896 | |
| Pm422 | 7 July 2016 | OQ288931 | OQ288968 | OQ303897 | |
| Pm423 | 7 July 2016 | OQ288932 | OQ288969 | OQ303898 | |
| Pm426 | 12 July 2016 | OQ288933 | OQ288970 | OQ303899 | |
| Pm427 | 13 September 2016 | OQ288934 | OQ288971 | OQ303900 | |
| Pm640 | 23 September 2017 | OQ288935 | OQ288972 | OQ303901 | |
| Pm641 1 | 24 September 2017 | OQ288936 | OQ288973 | OQ303902 | |
1 For molecular studies fungal cultures were used, in other cases DNA was extracted directly from the needles.
Table 2.
Decrease in radial growth of Pinus mugo shoots with diseased needles in 2016–2020.
| Years | 2016 | 2017 | 2018 | 2019 | 2020 |
|---|---|---|---|---|---|
| % of shoots showing a decrease in radial growth compared to the previous year | 42.9 | 57.1 | 65.7 | 62.9 | 56.7 |
| Average decrease in radial growth of shoots (%) | 22.1 | 20.0 | 22.2 | 24.7 | 25.8 |
| Minimal decrease in radial growth in a single shoot (%) | 4.1 | 2.3 | 3.8 | 2.7 | 4.6 |
| Maximal decrease in radial growth in a single shoot (%) | 68.1 | 57.1 | 65.7 | 72.6 | 57.9 |
Table 3.
Dimensions of ascospores given in the literature for Lophodermella sulcigena on Pinus species in Europe.
Table 3.
Dimensions of ascospores given in the literature for Lophodermella sulcigena on Pinus species in Europe.
| Reference | Tree Species | Ascospores Length × Width (µm) |
|---|---|---|
| Lagerberg (1910) [56] | P. sylvestris | 44–58 × 6 |
| Darker (1932) [40] | P. mugo, P. sylvestris | 27–35 × 4–5 |
| Terrier (1944) [57] | P. mugo, P. sylvestris | 35-(44.5–54)-65 × 4–6 |
| Moriondo (1963) [41] | P. sylvestris | 35–55 × 4–6 |
| Dennis (1968) [42] | Pinus spp. | 27–35 × 4–5 |
| Millar & Minter (1978) [43] | P. mugo, P. nigra, P. sylvestris | 27–40 × 4–5 |
| Kowalski (1988) [46] | P. sylvestris | 35–57 × 3.7–5 |
| Butin (2011) [14] | P. mugo, P. nigra, P. sylvestris | 27–35 × 4–5 |
| Beenken (2019) [58] | P. mugo | 35–65 × 4–6 |
| Kowalski et al. (2024) (Present study) | P. mugo | 35–64 × 4.5–6.0 |
Table 4.
Fungi associated Lophodermella sulcigena on Pinus mugo needles.
| Substrate Type | Fungi 1 |
|---|---|
| Needle killed by L. sulcigena | Hendersonia sp. Leptomelanconium allescheri Lophodermium conigenum Lophodermium corconticum |
| The second dead needle in the bundle not infected by L. sulcigena | Lophodermium conigenum Lophodermium corconticum Sydowia polyspora |
| Hysterothecium of L. sulcigena | Alternaria sp. (B) Cladosporium herbarum (B) Cladosporium sp. (B) Epicoccum nigrum (A) Hendersonia sp. (A) Seuratia millardetii (A) |
1 (A) early stage, second year of needles, summer; (B) late stage, second year of needles, autumn, and third year of needles.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kowalski, T.; Bartnik, C.; Bilański, P. Involvement of Lophodermella sulcigena in Endemic Disease of Pinus mugo Needles in the Polish Tatra Mountains. Forests 2024, 15, 422. https://doi.org/10.3390/f15030422
AMA Style
Kowalski T, Bartnik C, Bilański P. Involvement of Lophodermella sulcigena in Endemic Disease of Pinus mugo Needles in the Polish Tatra Mountains. Forests. 2024; 15(3):422. https://doi.org/10.3390/f15030422
Chicago/Turabian StyleKowalski, Tadeusz, Czesław Bartnik, and Piotr Bilański. 2024. "Involvement of Lophodermella sulcigena in Endemic Disease of Pinus mugo Needles in the Polish Tatra Mountains" Forests 15, no. 3: 422. https://doi.org/10.3390/f15030422
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.