Potential Allelopathic Candidates for Land Use and Possible Sustainable Weed Management in South Asian Ecosystem

 , ,
, ,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
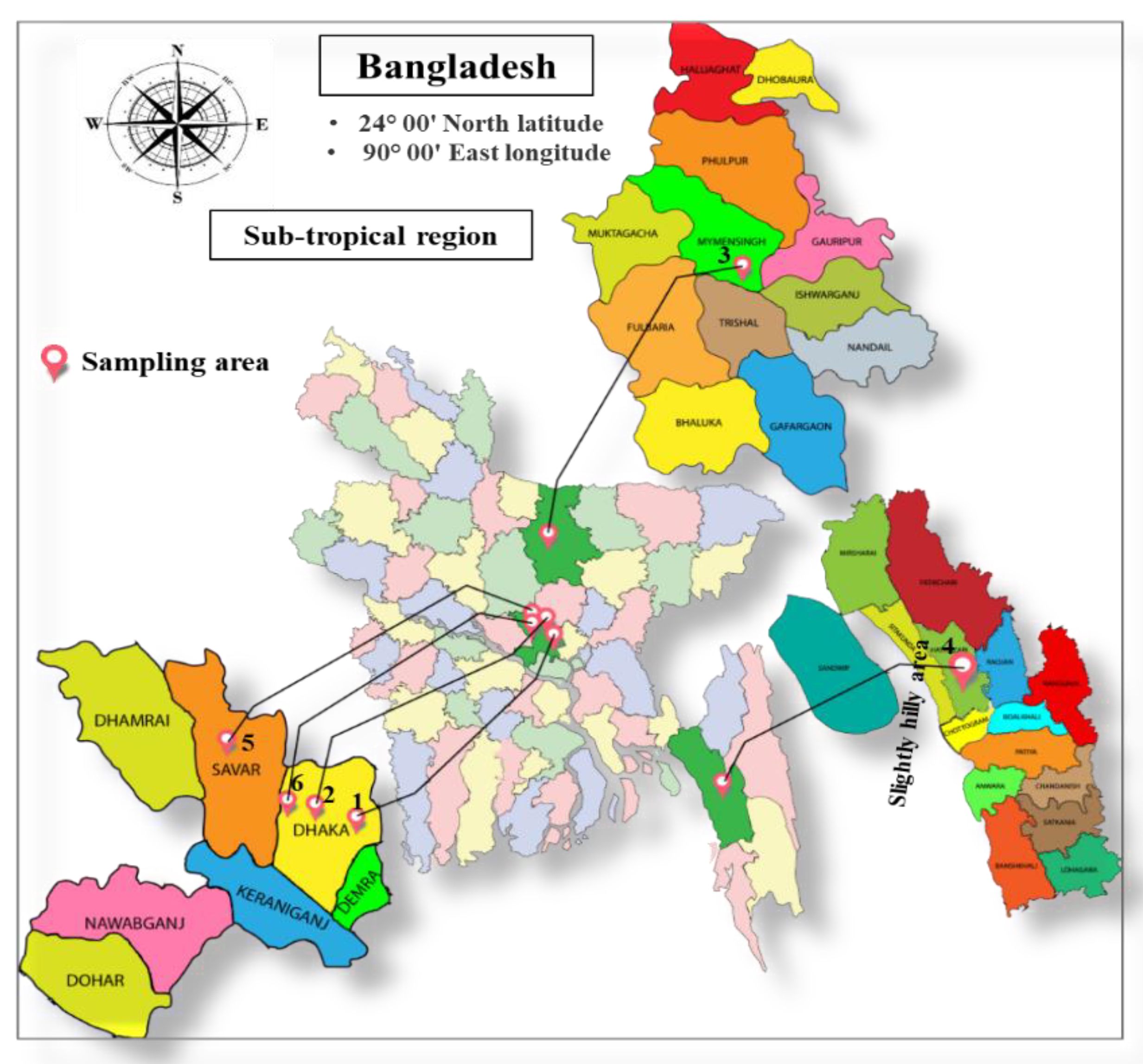
2.1. Study Area
2.2. Plant Samples and Preparation
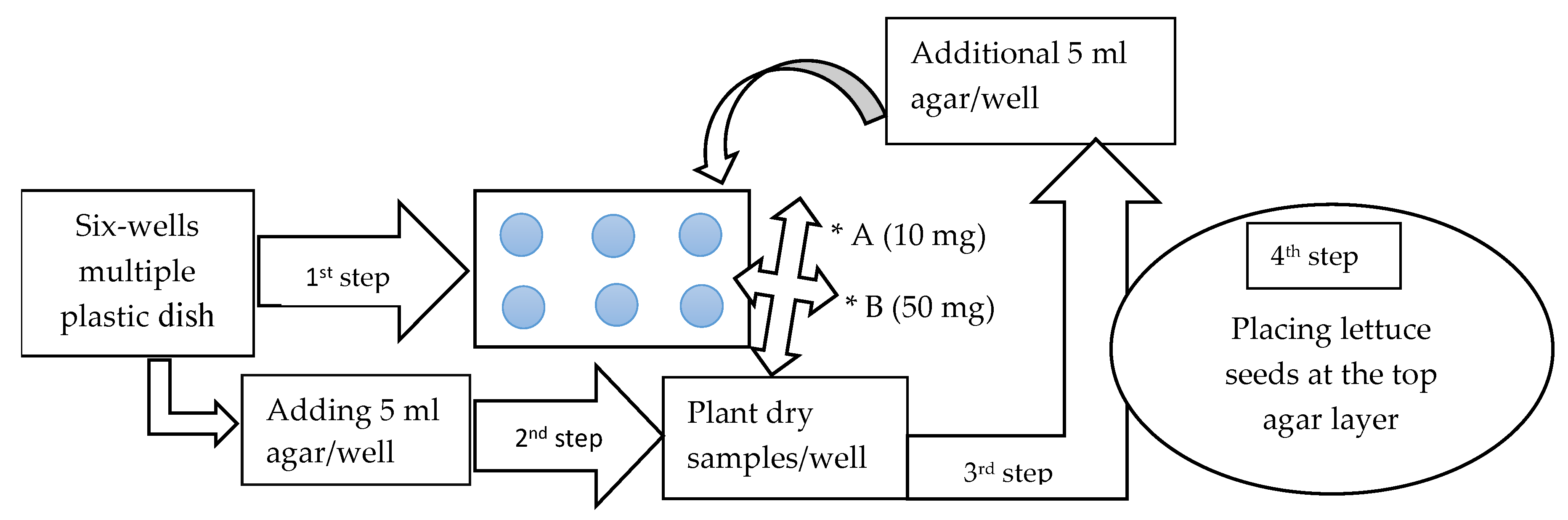
2.3. Sandwich Method
2.4. Analytical Study
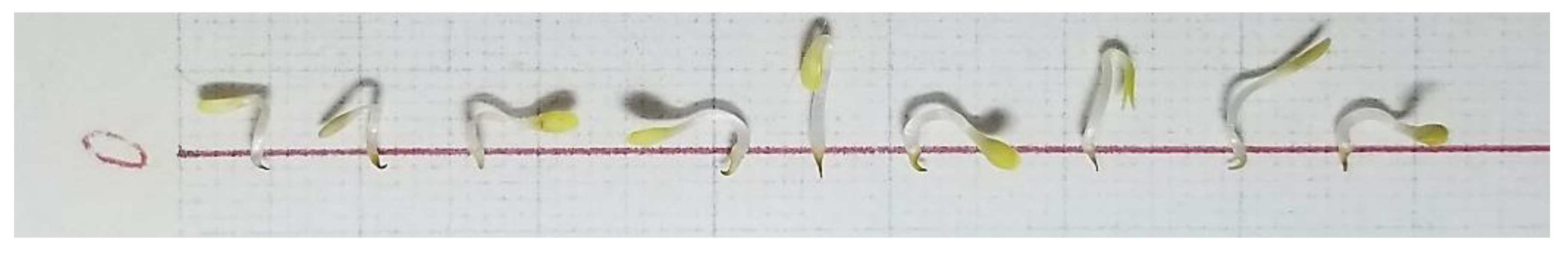
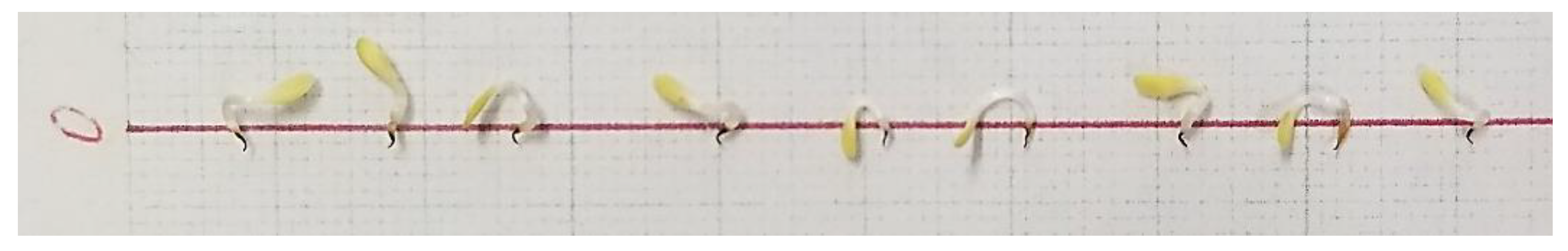
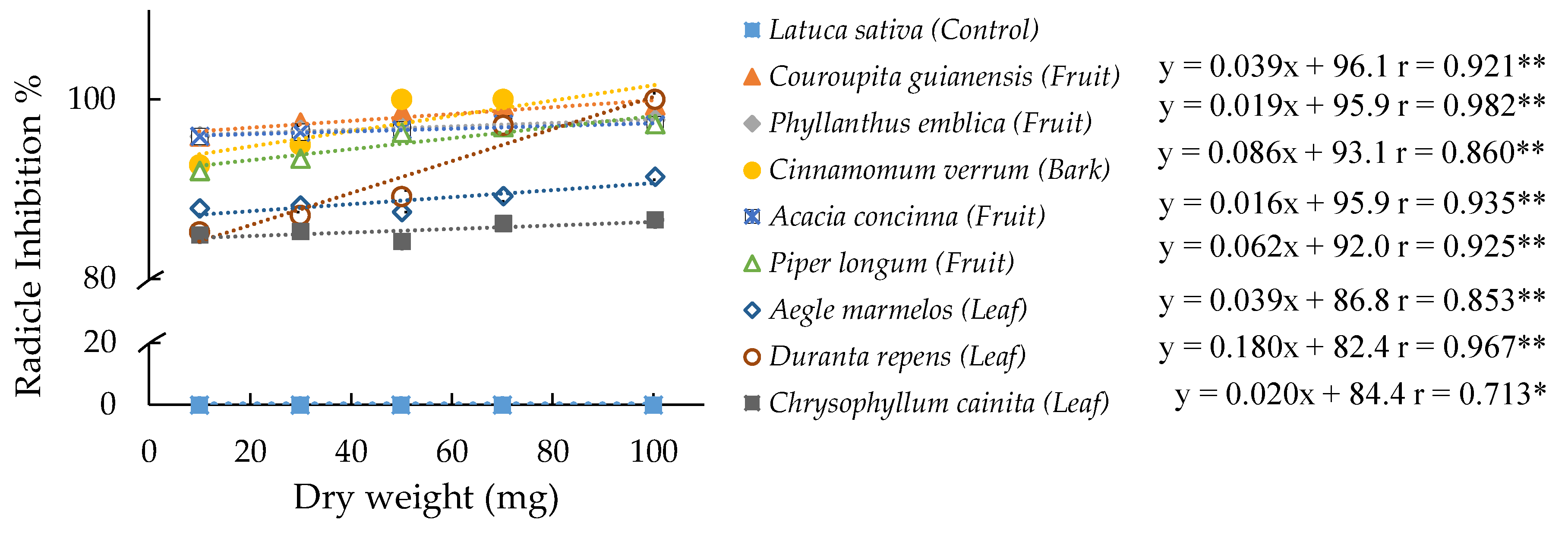
3. Results
3.1. Allelopathic Effect: Inhibition Diversity among the Plant Species
3.2. Optimal Inhibition Effect by Allelopathic Plants
3.3. Selective Species: Maximal Status of Inhibition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Minh, N.H.; Minh, T.B.; Kajiwara, N.; Kunisue, T.; Subramanian, A.; Iwata, H.; Tana, T.S.; Baburajendran, R.; Karuppiah, S.; Viet, P.H.; et al. Contamination by Persistent Organic Pollutants in Dumping Sites of Asian Developing Countries: Implication of Emerging Pollution Sources. Arch. Environ. Contam. Toxicol. 2006, 50, 474–481. [Google Scholar] [CrossRef]
- Shammi, M.; Sultana, A.; Hasan, N.; Mostafizur Rahman, M.; Saiful Islam, M.; Bodrud-Doza, M.; Khabir Uddin, M. Pesticide Exposures towards Health and Environmental Hazard in Bangladesh: A Case Study on Farmers’ Perception. J. Saudi Soc. Agric. Sci. 2018. [Google Scholar] [CrossRef]
- Zhang, M.; Zeiss, M.R.; Geng, S. Agricultural Pesticide Use and Food Safety: California’s Model. J. Integr. Agric. 2015, 14, 2340–2357. [Google Scholar] [CrossRef]
- Shammi, M.; Hasan, N.; Rahman, M.M.; Begum, K.; Sikder, M.T.; Bhuiyan, M.H.; Uddin, M.K. Sustainable Pesticide Governance in Bangladesh: Socio-Economic and Legal Status Interlinking Environment, Occupational Health and Food Safety. Environ. Syst. Decis. 2017, 37, 243–260. [Google Scholar] [CrossRef]
- Bangladesh Bureau of Statistics. Available online: http://203.112.218.65:8008/PageWebMenuContent.aspx?MenuKey=243 (accessed on 23 January 2019).
- World Bank. Population Density (People per sq. km of Land Area). Available online: https://data.worldbank.org/indicator/EN.POP.DNST (accessed on 22 January 2019).
- Bangladesh Bureau of Statistics, Statistics and Informatics Division, Ministry of Planning, Government of the People’s Republic of Bangladesh. Population Projection of Bangladesh: Dynamics and Trends 2011-2061; Dhaka, Bangladesh, 2015. Available online: http://203.112.218.65:8008/WebTestApplication/userfiles/Image/PopMonographs/PopulationProjection.pdf (accessed on 21 January 2019).
- Oerke, E.-C. Crop Losses to Pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Weed Science Society of North America (WSSA). WSSA Calculates Billions in Potential Economic Losses from Uncontrolled Weeds. Available online: http://wssa.net/2016/05/wssa-calculates-billions-in-potential-economic-losses-from-uncontrolled-weeds/ (accessed on 6 January 2019).
- Aziz, M.A. Proceedings of the Asia Regional Workshop on the Implementation, Monitoring and Observance of the International Code of Conduct on the Distribution and Use of Pesticides; Country Report; FAO-Regional Office for the Asia and the Pacific: Bangkok, Thailand, 2005. [Google Scholar]
- Matin, M.A. Pesticides in Bangladesh. In Pesticide Residues in Coastal Tropical Ecosystems: Distribution, Fate and Effects; Taylor and Francis Group: London, UK, 2002; pp. 137–158. [Google Scholar]
- Rahman, S. Pesticide Consumption and Productivity and the Potential of IPM in Bangladesh. Sci. Total Environ. 2013, 445–446, 48–56. [Google Scholar] [CrossRef]
- Rahman, M.S.; Malek, M.A.; Matin, M.A. Trend of Pesticide Usage in Bangladesh. Sci. Total Environ. 1995, 159, 33–39. [Google Scholar] [CrossRef]
- Vencill, W.K.; Nichols, R.L.; Webster, T.M.; Soteres, J.K.; Mallory-Smith, C.; Burgos, N.R.; Johnson, W.G.; McClelland, M.R. Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef] [Green Version]
- Heap, I. Herbicide Resistant Weeds. In Integrated Pest Management: Pesticide Problems; Pimentel, D., Peshin, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 3, pp. 281–301. [Google Scholar]
- Weston, L.A.; Duke, S.O. Weed and Crop Allelopathy. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Fujii, Y. Allelopathy in the Natural and Agricultural Ecosystems and Isolation of Potent Allelochemicals from Velvet Bean (Mucuna pruriens) and Hairy Vetch (Vicia villosa). Biol. Sci. Space 2003, 17, 6–13. [Google Scholar] [CrossRef]
- Fujii, Y.; Hiradate, S. Allelopathy: New Concepts & Methodology; Science Publishers, Inc.: Enfield, NH, USA, 2007. [Google Scholar]
- Leather, G.R. Weed Control Using Allelopathic Crop Plants. J. Chem. Ecol. 1983, 9, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Rawat, L.S.; Maikhuri, R.K.; Bahuguna, Y.M.; Jha, N.K.; Phondani, P.C. Sunflower Allelopathy for Weed Control in Agriculture Systems. J. Crop Sci. Biotechnol. 2017, 20, 45–60. [Google Scholar] [CrossRef]
- Sujeeun, L.; Thomas, S.C. Potential of Biochar to Mitigate Allelopathic Effects in Tropical Island Invasive Plants: Evidence from Seed Germination Trials. Trop. Conserv. Sci. 2017, 10. [Google Scholar] [CrossRef]
- Carmello, C.R.; Cardoso, J.C. Effects of Plant Extracts and Sodium Hypochlorite on Lettuce Germination and Inhibition of Cercospora longissima in Vitro. Sci. Hortic. 2018, 234, 245–249. [Google Scholar] [CrossRef]
- Cortés-Rojas, D.F.; de Souza, C.R.F.; Oliveira, W.P. Clove (Syzygium aromaticum): A Precious Spice. Asian Pac. J. Trop. Biomed. 2014, 4, 90–96. [Google Scholar] [CrossRef]
- Mazzafera, P. Efeito Alelopático Do Extrato Alcoólico Do Cravo-Da-Índia e Eugenol. Braz. J. Bot. 2003, 26, 231–238. [Google Scholar] [CrossRef]
- Shafique, S.; Bajwa, R.; Javaid, A.; Shafique, S. Biological Control of Parthenium IV: Suppressive Ability of Aqueous Leaf Extracts of Some Allelopathic Trees against Germination and Early Seedling Growth of Parthenium hysterophorus L. Pak. J. Weed Sci. Res. 2005, 11, 75–79. [Google Scholar]
- Abugre, S.; Apetorgbor, A.K.; Antwiwaa, A.; Apetorgbor, M.M. Allelopathic Effects of Ten Tree Species on Germination and Growth of Four Traditional Food Crops in Ghana. J. Agric. Technol. 2011, 7, 825–834. [Google Scholar]
- Chowdhury, H.; Singh, R.D.; Kumar, P. Phytotoxic Effects of Aqueous Extracts of Neem (Azadirachta indica A. Juss.) and Chinaberry (Melia azedarach L.) on Some Crop and Weed Species. Available online: https://eurekamag.com/research/003/525/003525586.php (accessed on 15 April 2019).
- Hazra, C.R.; Tripathi, S.B. Performance of Oat Genotypes under Different Tree Covers and Open. Agric. Sci. Dig. 1989, 9, 146–148. [Google Scholar]
- Jalal, S.; Roy, B.; Sarker, B.C.; Uddin, M.N.; Roy, S.R. Growth Inhibitory Effect of Ten Fruit Leaves on Selected Weeds. J. Agrofor. Environ. 2010, 4, 101–104. [Google Scholar]
- Rai, J.P.N.; Tripathi, R.S. Allelopathic Effects of Eupatorium riparium on Population Regulation of Two Species of Galinsoga and Soil Microbes. Plant Soil 1984, 80, 105–117. [Google Scholar] [CrossRef]
- Sahid, I.B.; Sugau, J.B. Allelopathic Effect of Lantana (Lantana camara) and Siam Weed (Chromolaena odorata) on Selected Crops. Weed Sci. 1993, 41, 303–308. [Google Scholar] [CrossRef]
- Zeng, R.S.; Mallik, A.U.; Luo, S. Allelopathy in Sustainable Agriculture and Forestry; Springer: New York, NY, USA, 2008. [Google Scholar]
- EI-Khatib, A.A.; Hegazy, A.K.; Galal, H.K. Does Allelopathy Have a Role in the Ecology of Chenopodium murale. Ann. Bot. Fenn. 2004, 41, 37–45. [Google Scholar]
- Mansour, M.M.F. Plasma Membrane Permeability as an Indicator of Salt Tolerance in Plants. Biol. Plant. 2013, 57, 1–10. [Google Scholar] [CrossRef]
- Fujii, Y.; Shibuya, T.; Nakatani, K.; Itani, T.; Hiradate, S.; Parvez, M.M. Assessment Method for Allelopathic Effect from Leaf Litter Leachates. Weed Biol. Manag. 2004, 4, 19–23. [Google Scholar] [CrossRef]
- de Albuquerque, U.P.; de Medeiros, P.M.; de Almeida, A.L.S.; Monteiro, J.M.; de Freitas Lins Neto, E.M.; de Melo, J.G.; dos Santos, J.P. Medicinal Plants of the Caatinga (Semi-Arid) Vegetation of NE Brazil: A Quantitative Approach. J. Ethnopharmacol. 2007, 114, 325–354. [Google Scholar] [CrossRef]
- Bhadoria, P. Allelopathy: A Natural Way towards Weed Management. Am. J. Exp. Agric. 2011, 1, 7–20. [Google Scholar] [CrossRef]
- Boonmee, S.; Kato-Noguchi, H. Allelopathic Activity of Acacia concinna Pod Extracts. Emir. J. Food Agric. 2017, 29, 250–255. [Google Scholar] [CrossRef]
- Rietveld, W.J. Allelopathic Effects of Juglone on Germination and Growth of Several Herbaceous and Woody Species. J. Chem. Ecol. 1983, 9, 295–308. [Google Scholar] [CrossRef]
- Hong, N.H.; Xuan, T.D.; Tsuzuki, E.; Terao, H.; Matsuo, M.; Khanh, T.D. Weed Control of Four Higher Plant Species in Paddy Rice Fields in Southeast Asia. J. Agron. Crop Sci. 2004, 190, 59–64. [Google Scholar] [CrossRef]
- Heywood, V.H.; Chant, S.R. Popular Encyclopedia of Plants; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Geetha, M.; Saluja, A.K.; Shankar, M.B.; Mehta, R.S. Analgesic and Anti-Inflammatory Activity of Couroupita guianensis Aubl. J. Nat. Remedies 2004, 4, 4. [Google Scholar]
- Khan, M.S.I.; Kato-Noguchi, H. Assessment of Allelopathic Potential of Couroupita guianensis Aubl. Plant Omics 2016, 9, 115–120. [Google Scholar] [CrossRef]
- Umachigi, S.P.; Jayaveera, K.; Kumar, A.; Kumar, G. Antimicrobial, Wound Healing and Antioxidant Potential of Couroupita guianensis in Rats. Pharmacol. Online 2007, 3, 269–281. [Google Scholar]
- Patel, S.H.; Suthar, J.V.; Patel, R.K.; Zankharia, U.S.; Jani, V.R.; Gajjar, K.N. Antimicrobial Activity Investigation of Aegle marmelos, Couroupita guianesis, Manilkara hexandra, Cow Urine and Dung. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1014–1022. [Google Scholar]
- Shivashankar, M.; Rajeshwari, S.; Nagananda, G.; Rajath, S.; Chandan, N. Comparative Antioxidant and Antimicrobial Studies of Cold and Hot Bark Hydromethanolic Extract of Couroupita guianensis Aubl. Res. Pharm. 2013, 3, 6–13. [Google Scholar]
- Manimegalai, S.; Sridharan, T.; Rameshpathy, M.; Devi Rajeswari, V. Antioxidant, Phytochemical Screening and Antimicrobial Activity of Couroupita guianensis Flower Extract. Der Pharm. Lett. 2014, 6, 251–256. [Google Scholar]
- Reinhardt, C.F.; Bezuidenhout, S.R. Growth Stage of Cyperus Esculentus Influences Its Allelopathic Effect on Ectomycorrhizal and Higher Plant Species. J. Crop Prod. 2001, 4, 323–333. [Google Scholar] [CrossRef]
- Begum, R.; Rahman, M.S.; Chowdhury, A.M.S.; Hasan, C.M.; Rashid, M.A. Secondary Metabolites (Triterpenes) from Couroupita guianensis. Orient. Pharm. Exp. Med. 2009, 9, 200–205. [Google Scholar] [CrossRef]
- Gousia, S.K.; Kumar, K.A.; Kumar, T.V.; Latha, J.N.L. Biological Activities and Medicinal Properties of Couroupita guianensis. Int. J. Pharm. Pharm. Sci. Res. 2013, 3, 140–143. [Google Scholar]
- Burden, R.S.; Cooke, D.T.; Carter, G.A. Inhibitors of Sterol Biosynthesis and Growth in Plants and Fungi. Phytochemistry 1989, 28, 1791–1804. [Google Scholar] [CrossRef]
- Sodaeizadeh, H.; Rafieiolhossaini, M.; Havlík, J.; Van Damme, P. Allelopathic Activity of Different Plant Parts of Peganum harmala L. and Identification of Their Growth Inhibitors Substances. Plant Growth Regul. 2009, 59, 227–236. [Google Scholar] [CrossRef]
- Kaur, R.; Callaway, R.M.; Inderjit. Soils and the Conditional Allelopathic Effects of a Tropical Invader. Soil Biol. Biochem. 2014, 78, 316–325. [Google Scholar] [CrossRef]
- Verma, R.C.; Gupta, A. Effect of Pre-Treatments on Quality of Solar-Dried Amla. J. Food Eng. 2004, 65, 397–402. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Tanaka, T.; Iwamoto, Y.; Yang, C.-R.; Kouno, I. Phyllaemblic Acid, a Novel Highly Oxygenated Norbisabolane from the Roots of Phyllanthus emblica. Tetrahedron Lett. 2000, 41, 1781–1784. [Google Scholar] [CrossRef]
- Chaudhuri, K.; Ratan, K. Standardised Extract of Phyllanthus emblica: A Skin Lightener with Anti-Aging Benefits. In Proceedings of the PCIA Conference, Guangzhou, China, 9–11 March 2004. [Google Scholar]
- Dutta, B.K.; Rahman, I.; Das, T.K. Antifungal Activity of Indian Plant Extracts. Mycoses 1998, 41, 535–536. [Google Scholar] [CrossRef]
- Rani, P.; Khullar, N. Antimicrobial Evaluation of Some Medicinal Plants for Their Anti-Enteric Potential against Multi-Drug Resistant Salmonella Typhi. Phytother. Res. 2004, 18, 670–673. [Google Scholar] [CrossRef]
- Asmawi, M.Z.; Kankaanranta, H.; Moilanen, E.; Vapaatalo, H. Anti-Inflammatory Activities of Emblica officinalis Gaertn Leaf Extracts. J. Pharm. Pharmacol. 1993, 45, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Perianayagam, J.B.; Sharma, S.K.; Joseph, A.; Christina, A.J.M. Evaluation of Anti-Pyretic and Analgesic Activity of Emblica officinalis Gaertn. J. Ethnopharmacol. 2004, 95, 83–85. [Google Scholar] [CrossRef]
- Jose, J.K.; Kuttan, G.; Kuttan, R. Antitumour Activity of Emblica officinalis. J. Ethnopharmacol. 2001, 75, 65–69. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Nagao, T.; Tanaka, T.; Yang, C.-R.; Okabe, H.; Kouno, I. Antiproliferative Activity of the Main Constituents from Phyllanthus emblica. Biol. Pharm. Bull. 2004, 27, 251–255. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant Activity of Plant Extracts Containing Phenolic Compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Mijangos, F.; Varona, F.; Villota, N. Changes in Solution Color During Phenol Oxidation by Fenton Reagent. Environ. Sci. Technol. 2006, 40, 5538–5543. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.D.; Shinkichi, T.; Khanh, T.D.; Chung, I.M. Biological Control of Weeds and Plant Pathogens in Paddy Rice by Exploiting Plant Allelopathy: An Overview. Crop Prot. 2005, 24, 197–206. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, USA, 1984; p. 422. [Google Scholar]
- Cheng, F.; Cheng, Z. Research Progress on the Use of Plant Allelopathy in Agriculture and the Physiological and Ecological Mechanisms of Allelopathy. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Yadav, R.; Yadav, K.; Wani, F.A.; Meghvansi, M.K.; Sharma, S.; Jabeen, F. Allelopathic Potentialities of Different Concentration of Aqueous Leaf Extracts of Some Arable Trees on Germination and Radicle Growth of Cicer arietinum Var. – C-235. Glob. J. Mol. Sci. 2009, 4, 91–95. [Google Scholar]
- Fujii, Y.; Parvez, S.S.; Parvez, M.M.; Ohmae, Y.; Iida, O. Screening of 239 Medicinal Plant Species for Allelopathic Activity Using the Sandwich Method. Weed Biol. Manag. 2003, 3, 233–241. [Google Scholar] [CrossRef]
- Balan, R. Investment Reduction and Increase in Service Life of Kattumaram Logs. Available online: http://www.fao.org/3/ad922e/ad922e00.pdf (accessed on 7 May 2019).
- Pandya, D.J.; Desai, T.R.; Nadpara, N.P.; Mehta, H.A.; Modi, A.M. Pharmacognostic Study and Establishment of Quality Parameters of Leaves of Bombax insigne Linn. Int. J. Pharmacogn. Phytochem. Res. 2010, 2, 1–5. [Google Scholar]
- Li, B.; Yang, C.; Lin, P. Ecology; High. Educ. Press: Beijing, China, 1999; pp. 97–100. [Google Scholar]
- Bai, Y.; Thompson, D.; Broersma, K. Early Establishment of Douglas-Fir and Ponderosa Pine in Grassland Seedbeds. J. Range Manag. 2000, 53, 511–517. [Google Scholar] [CrossRef]
- Parvez, S.S.; Parvez, M.M.; Fujii, Y.; Gemma, H. Differential Allelopathic Expression of Bark and Seed of Tamarindus indica L. Plant Growth Regul. 2004, 42, 245–252. [Google Scholar] [CrossRef]
- Mousavi, S.H.; Alami-Saeid, K.; Moshatati, A. Effect of Leaf, Stem and Root Extract of Alfalfa (Melilotus indicus) on Seed Germination and Seedling Growth of Wheat (Triticum aestivum). Int. J. Agric. Crop Sci. 2013, 5, 44–49. [Google Scholar]
- Aslani, F.; Juraimi, A.S.; Ahmad-Hamdani, M.S.; Omar, D.; Alam, M.A.; Hashemi, F.S.G.; Hakim, M.A.; Uddin, M.K. Allelopathic Effect of Methanol Extracts from Tinospora tuberculata on Selected Crops and Rice Weeds. Acta Agric. Scand. Sect. B Soil Plant Sci. 2014, 64, 165–177. [Google Scholar] [CrossRef]
- Abu-Romman, S.; Ammari, T. Allelopathic Effect of ‘Arundo donax’, a Mediterranean Invasive Grass. Plant Omics 2015, 8, 287. [Google Scholar]
- Silva, R.M.G.; Brigatti, J.G.F.; Santos, V.H.M.; Mecina, G.F.; Silva, L.P. Allelopathic Effect of the Peel of Coffee Fruit. Sci. Hortic. 2013, 158, 39–44. [Google Scholar] [CrossRef]
- Barbosa, L.C.A.; Demuner, A.J.; Clemente, A.D.; de Paula, V.F.; Ismail, F.M.D. Seasonal Variation in the Composition of Volatile Oils from Schinus terebinthifolius Raddi. Química Nova 2007, 30, 1959–1965. [Google Scholar] [CrossRef]
- Fujii, Y.; Matsuyama, M.; Hiradate, S.; Shimozawa, H. Dish Pack Method: A New Bioassay for Volatile Allelopathy. In Proceedings of the 4th World Congress on Allelopathy, Establishing the Scientific Base, Wagga Wagga, NSW, Australia, 21–26 August 2005; pp. 493–497. [Google Scholar]
- Liu, X.; Chen, Q.; Wang, Z.; Xie, L.; Xu, Z. Allelopathic Effects of Essential Oil from Eucalyptus grandis × E. urophylla on Pathogenic Fungi and Pest Insects. Front. For. China 2008, 3, 232–236. [Google Scholar] [CrossRef]
- Ahmed, R.; Uddin, M.B.; Khan, M.A.S.A.; Mukul, S.A.; Hossain, M.K. Allelopathic Effects of Lantana camara on Germination and Growth Behavior of Some Agricultural Crops in Bangladesh. J. For. Res. 2007, 18, 301–304. [Google Scholar] [CrossRef]
- Mushtaq, W.; Siddiqui, M.B. Allelopathy in Solanaceae Plants. J. Plant Prot. Res. 2018, 58, 1–7. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Site Code | Botanical Name | Plant Part | Dry Samples Content (10 mL Agar−1) | Score | |||
|---|---|---|---|---|---|---|---|---|
| 10 mg | 50 mg | |||||||
| R% | H% | R% | H% | |||||
| Acanthaceae | 1 | Rungia pectinate | Leaf | 78.3 | −27.2 | 77.1 | −4.67 | ** * |
| Acanthaceae | 4 | Justicia adhatoda | Leaf | 76.4 | −11.4 | 82.8 | 44.5 | ** |
| Acanthaceae | 4 | Asystasia gangetica | Leaf | 72.7 | −19.0 | 17.0 | −66.4 | ** |
| Acanthaceae | 3 | Phaulopsis imbricate | Leaf | 67.7 | −12.6 | 75.7 | 17.7 | ** |
| Acanthaceae | 5 | Adhatoda vasica | Leaf | 56.7 | 10.4 | 71.5 | 44.9 | * |
| Acanthaceae | 3 | Nelsonia canescens | Leaf | 50.0 | −78.8 | 63.7 | 16.0 | - |
| Acanthaceae | 1 | Justicia gendarussa | Leaf | 45.5 | 8.69 | 62.3 | 14.4 | - |
| Acanthaceae | 1 | Justicia peruviana | Leaf | 34.5 | 13.0 | 68.7 | 46.2 | - |
| Acanthaceae | 3 | Justicia peruviana | Stem | 30.9 | −48.5 | 80.7 | 17.3 | - |
| Acanthaceae | 4 | Acanthus ilicifolius | Leaf | 27.5 | −84.3 | 62.0 | −45.9 | - |
| Acanthaceae | 3 | Hygrophila schulli | Leaf | 19.0 | −54.7 | 61.0 | −16.8 | - |
| Acanthaceae | 1 | Andrographis paniculata | Stem | 17.0 | −60.8 | 49.6 | −6.71 | - |
| Acanthaceae | 1 | Andrographis paniculata | Leaf | 4.79 | −72.0 | 36.8 | −43.2 | - |
| Achariaceae | 2 | Hydnocarpus kurzii | Leaf | 64.0 | −0.18 | 78.6 | 29.2 | * |
| Amaranthaceae | 5 | Alternanthera philoxeroides | Leaf | 68.9 | 29.2 | 82.7 | 41.0 | ** |
| Amaranthaceae | 2 | Cyathula prostrata | Leaf | 64.1 | 2.11 | 73.1 | 24.0 | * |
| Anacardiaceae | 1 | Mangifera indica | seed | 37.3 | 9.11 | 86.0 | 52.0 | - |
| Anacardiaceae | 2 | Mangifera indica | Leaf | 19.5 | −8.72 | 54.7 | 30.4 | - |
| Anacardiaceae | 1 | Mangifera indica | Peel | 18.4 | −19.7 | 69.6 | 42.4 | - |
| Anacardiaceae | 1 | Buchanania lanzan | Stem | −6.8 | −29.9 | 2.56 | −53.9 | - |
| Annonaceae | 4 | Artabotrys hexapetalus | Leaf | 29.5 | −4.53 | 55.1 | 14.7 | - |
| Apiaceae | 2 | Centella asiatica | Leaf | 75.5 | −37.3 | 85.1 | 10.3 | ** |
| Apiaceae | 2 | Cuminum cyminum | seed | 8.37 | −37.3 | 44.9 | −13.9 | - |
| Aplaceae | 3 | Foeniculum vulgare | seed | 28.7 | 0.53 | 62.9 | 31.0 | - |
| Apocynaceae | 1 | Holarrhena pubescens | Leaf | 74.7 | 44.8 | 92.3 | 79.0 | ** |
| Apocynaceae | 1 | Rauvolfia serpentine | Leaf | 72.7 | 16.2 | 81.0 | 28.0 | ** |
| Apocynaceae | 1 | Calotropis gigantean | Leaf | 65.9 | −1.49 | 82.3 | 16.6 | ** |
| Apocynaceae | 1 | Tabernaemontana divaricate | Leaf | 44.3 | −18.8 | 75.3 | 18.6 | - |
| Apocynaceae | 1 | Tabernaemontana dichotoma | Flower | 39.0 | −38.8 | 60.0 | −15.4 | - |
| Apocynaceae | 1 | Calotropis gigantean | Stem | 37.8 | −11.1 | 75.5 | 50.5 | - |
| Apocynaceae | 2 | Nerium oleander | Leaf | 36.8 | −30.4 | 53.1 | −17.0 | - |
| Apocynaceae | 1 | Allamanda cathartica | Leaf | 33.0 | −38.9 | 62.8 | −37.8 | - |
| Apocynaceae | 3 | Holarrhena pubescens | Leaf | 22.4 | −53.7 | 23.3 | −53.4 | - |
| Apocynaceae | 1 | Hemidesmus indicus | Leaf | 13.2 | −67.5 | 48.6 | −17.3 | - |
| Apocynaceae | 1 | Nerium album | Leaf | 10.0 | −44.2 | 41.7 | 16.8 | - |
| Apocynaceae | 5 | Alstonia macrophylla | Leaf | −6.05 | −89.7 | 32.4 | −33.1 | - |
| Araceae | 2 | Dieffenbachia seguine | Leaf | 61.9 | −6.76 | 70.4 | −1.83 | * |
| Asparagaceae | 1 | Asparagus racemosus | Root | 13.2 | −46.2 | 70.6 | 18.5 | - |
| Asteraceae | 5 | Eupatorium triplinerve | Leaf | 80.2 | 33.6 | 90.3 | 56.9 | ** * |
| Asteraceae | 4 | Ageratum conyzoides | Leaf | 77.2 | 17.4 | 88.5 | 67.7 | ** * |
| Asteraceae | 4 | Vernonia cinerea | Leaf | 71.0 | 11.6 | 76.3 | 6.26 | ** |
| Asteraceae | 2 | Brickellia baccharidea | Leaf | 62.3 | −10.3 | 76.3 | 7.77 | * |
| Asteraceae | 5 | Spilanthes cliata | Leaf | 58.7 | −7.72 | 77.6 | 35.4 | * |
| Asteraceae | 2 | Dittrichia viscosa | Leaf | 58.6 | −5.94 | 85.0 | 43.0 | * |
| Asteraceae | 2 | Tagetes erecta | Leaf | 56.9 | −34.2 | 82.4 | 2.84 | * |
| Asteraceae | 2 | Helianthus annuus | Leaf | 56.3 | −33.9 | 68.4 | −2.67 | * |
| Asteraceae | 1 | Tagetes erecta | Flower | 51.4 | −25.6 | 73.9 | 5.47 | - |
| Asteraceae | 4 | Artemisia nilagirica | Leaf | 43.3 | −9.74 | 100 | 100 | - |
| Asteraceae | 1 | Vernonia patula | Leaf | 43.0 | −4.58 | 69.2 | −17.4 | - |
| Asteraceae | 4 | Gigantochloa apus | Leaf | 37.5 | 13.6 | 66.4 | −11.3 | - |
| Asteraceae | 4 | Wedelia chinensis | Leaf | 36.5 | −65.8 | 55.2 | −25.2 | - |
| Asteraceae | 4 | Mikania cordata | Leaf | 34.3 | −13.2 | 84.4 | 40.1 | - |
| Asteraceae | 4 | Mikania micrantha | Leaf | 20.7 | −46.7 | 28.7 | −17.3 | - |
| Asteraceae | 4 | Blumea lacera | Leaf | 11.5 | −80.6 | 35.3 | −73.0 | - |
| Asteraceae | 4 | Bidens pilosa | Leaf | −4.74 | −82.6 | 29.9 | −84.0 | - |
| Betulaceae | 1 | Betula alnoides | Leaf | 14.5 | −36.0 | 19.4 | −41.7 | - |
| Bignoniaceae | 1 | Parmentiera cereifera | Leaf | 58.6 | −12.8 | 85.7 | 42.9 | * |
| Bignoniaceae | 1 | Stereospermum angustifolium | Leaf | 44.8 | −18.1 | 68.3 | 12.6 | - |
| Bignoniaceae | 1 | Jacaranda mimosifolia | Leaf | 6.07 | −48.9 | 46.3 | −24.9 | - |
| Bombacaceae | 1 | Bombax ceiba | Root | 22.4 | −58.8 | 47.0 | −28.8 | - |
| Bombacaceae | 3 | Bombax ceiba | Leaf | 21.7 | −41.5 | 29.8 | −27.6 | - |
| Boraginaceae | 5 | Cordia dichotoma | Leaf | 65.1 | −25.6 | 87.4 | 26.9 | ** |
| Boraginaceae | 2 | Coldenia procumbens | Leaf | 21.3 | −45.1 | 51.7 | −43.0 | - |
| Boraginaceae | 4 | Heliotropium indicum | Leaf | 19.0 | −66.0 | 42.7 | −12.5 | - |
| Bromeliaceae | 4 | Ananas comosus | Leaf | 13.2 | −7.36 | 38.9 | 1.40 | - |
| Burseraceae | 4 | Canarium resiniferum | Leaf | 51.5 | −32.9 | 67.2 | −0.07 | - |
| Burseraceae | 1 | Protium serratum | Leaf | 24.6 | −51.5 | 58.9 | −15.3 | - |
| Caesalpiniaceae | 1 | Cassia renigera | Leaf | 52.5 | −3.40 | 84.9 | 25.4 | - |
| Caesalpiniaceae | 1 | Pongamia pinnata | Leaf | 33.4 | −16.7 | 68.6 | 36.3 | - |
| Caesalpiniaceae | 1 | Senna tora | Leaf | 30.5 | −57.0 | 61.2 | −1.59 | - |
| Caesalpiniaceae | 1 | Cassia angustifolia | Leaf | 20.1 | −16.7 | 64.0 | 13.4 | - |
| Calophyllaceae | 1 | Calophyllum inophyllum | Leaf | 26.0 | −22.7 | 40.1 | −0.42 | - |
| Calophyllaceae | 1 | Mesua ferrea | Leaf | −1.01 | −55.5 | 26.2 | −27.2 | - |
| Calophyllaceae | 1 | Mesua ferrea | Flower | 29.0 | −23.9 | 62.8 | 6.68 | - |
| Caricaceae | 2 | Carica papaya | Leaf | 60.0 | −26.0 | 83.1 | 13.2 | * |
| Combertaceae | 1 | Terminalia catappa | Leaf | 63.2 | 20.5 | 81.1 | 24.0 | * |
| Combretaceae | 1 | Terminalia chebula | Leaf | 64.7 | −6.28 | 89.7 | 63.1 | * |
| Combretaceae | 1 | Terminalia belerica | Leaf | 16.9 | −36.1 | 64.1 | −4.34 | - |
| Combretaceae | 1 | Terminalia arjuna | Leaf | 16.4 | −24.8 | 83.1 | 12.8 | - |
| Combretaceae | 3 | Terminalia chebula | Fruit | 0.82 | −62.3 | 49.4 | −3.26 | - |
| Commelinaceae | 2 | Commelina benghalensis | Leaf | 44.0 | −60.2 | 69.6 | −17.2 | - |
| Commelinaceae | 1 | Commelina diffusa | Leaf | 37.0 | −62.5 | 66.3 | 2.32 | - |
| Crassulaceae | 4 | Bryophyllum pinnatum | Leaf | 65.8 | 30.2 | 79.3 | 28.1 | ** |
| Cucurbitaceae | 2 | Cucumis sativa | Peel | 77.8 | −100 | 83.1 | 27.8 | ** * |
| Cucurbitaceae | 2 | Cucurbita moschata | Peel | 61.6 | 21.3 | 74.3 | 43.1 | * |
| Cucurbitaceae | 1 | Gynostemma pentaphyllum | Leaf | 43.4 | −29.3 | 49.4 | 11.1 | - |
| Cucurbitaceae | 4 | Coccinia grandis | Leaf | 40.2 | −7.44 | 89.1 | 37.9 | - |
| Cucurbitaceae | 2 | Benincasa hispida | Peel | 10.1 | −42.2 | 57.4 | 2.71 | - |
| Dilleniaceae | 1 | Dillenia indica | Leaf | 14.2 | −77.1 | 29.7 | −47.8 | - |
| Dipterocarpaceae | 1 | Dipterocarpus turbinatus | Leaf | 62.0 | −27.8 | 74.7 | 20.6 | * |
| Dipterocarpaceae | 1 | Hopea odorata | Leaf | 40.1 | −10.3 | 68.6 | −5.85 | - |
| Dipterocarpaceae | 1 | Anisoptera scaphula | Leaf | 36.5 | −66.3 | 56.4 | −5.16 | - |
| Ebenaceae | 1 | Diospyros montana | Leaf | 68.5 | 15.4 | 76.2 | 56.5 | ** |
| Euphorbiaceae | 4 | Manihot esculenta | Leaf | 71.0 | −13.6 | 84.0 | −5.29 | ** |
| Euphorbiaceae | 3 | Euphorbia neriifolia | Leaf | 61.2 | −3.95 | 89.3 | 46.2 | * |
| Euphorbiaceae | 2 | Euphorbia tithymaloides | Seed | 60.4 | −29.3 | 64.7 | 22.1 | * |
| Euphorbiaceae | 4 | Euphorbia tirucalli | Seed | 53.6 | 17.5 | 76.4 | 41.7 | * |
| Euphorbiaceae | 4 | Cnesmone javanica | Leaf | 46.1 | −31.4 | 81.2 | 55.3 | - |
| Euphorbiaceae | 4 | Macaranga tanarius | Leaf | 43.1 | −42.6 | 69.8 | −30.6 | - |
| Euphorbiaceae | 2 | Ricinus communis | Seed | 40.1 | −47.9 | 54.8 | 28.5 | - |
| Euphorbiaceae | 2 | Ricinus communis | Root | 28.3 | −38.4 | 84.9 | −15.8 | - |
| Euphorbiaceae | 3 | Pedilanthus tithymaloides | Leaf | 25.6 | −58.5 | 67.2 | −25.2 | - |
| Euphorbiaceae | 5 | Croton roxburghii | Leaf | 29.3 | −13.1 | 42.1 | −13.9 | - |
| Fabaceae | 2 | Acacia concinna | Fruit | 95.4 | 64.2 | 96.3 | 72.8 | ** ** |
| Fabaceae | 2 | Saraca asoca | Bark | 81.5 | 18.4 | 87.4 | 40.7 | ** * |
| Fabaceae | 4 | Mucuna pruriens | Leaf | 76.8 | 33.0 | 88.9 | 52.5 | ** * |
| Fabaceae | 4 | Tephrosia candida | Leaf | 75.5 | −42.4 | 86.4 | 23.3 | ** |
| Fabaceae | 4 | Senna alexandrina | Leaf | 74.8 | 17.3 | 83.9 | 60.9 | ** |
| Fabaceae | 1 | Senna alata | Leaf | 70.5 | −7.62 | 73.5 | 0.99 | ** |
| Fabaceae | 1 | Senna siamea | Leaf | 58.7 | −19.1 | 72.6 | −10.5 | * |
| Fabaceae | 5 | Pisum sativum | Peel | 56.6 | −9.73 | 77.4 | 49.6 | * |
| Fabaceae | 2 | Trigonella foenum-graecum | Fruit | 53.7 | 15.1 | 75.9 | 46.7 | * |
| Fabaceae | 1 | Indigo fera | Leaf | 50.5 | −41.1 | 51.0 | 5.71 | - |
| Fabaceae | 1 | Cicer arietinum | Leaf | 48.5 | −94.5 | 71.5 | −13.3 | - |
| Fabaceae | 1 | Dalbergia Sissoo | Leaf | 46.3 | −20.0 | 75.3 | 36.1 | - |
| Fabaceae | 2 | Millettia peguensis | Leaf | 40.2 | −30.3 | 68.4 | 24.5 | - |
| Fabaceae | 1 | Cassia nodosa | Leaf | 38.7 | −24.5 | 78.4 | 21.9 | - |
| Fabaceae | 5 | Glycyrrhiza glabra | Stem | 35.2 | 4.74 | 48.2 | 25.6 | - |
| Fabaceae | 1 | Vachellia nilotica | Bark | 12.4 | −56.3 | 10.2 | −67.9 | - |
| Fabaceae | 2 | Saraca asoca | Leaf | 11.6 | −73.0 | 12.5 | 0.47 | - |
| Fabaceae | 4 | Cajanus cajan | Leaf | 3.75 | −84.8 | 7.94 | −113 | - |
| Flacourtiaceae | 1 | Flacourtia jangomas | Leaf | 28.0 | −3.40 | 62.1 | −35.8 | - |
| Fumariaceae | 4 | Fumaria indica | Leaf | 20.3 | −82.4 | 70.2 | −50.4 | - |
| Gentianaceae | 6 | Swertia chirayita | Flower | 24.8 | −2.35 | 53.1 | 16.2 | - |
| Gentianaceae | 6 | Swertia chirayita | Leaf | 9.47 | −32.1 | 32.0 | −18.1 | - |
| Gentianaceae | 6 | Swertia chirayita | Stem | 4.00 | −25.1 | 33.7 | 5.47 | - |
| Lamiaceae | 2 | Mentha spicata | Leaf | 47.3 | −0.25 | 69.8 | 36.0 | - |
| Lamiaceae | 3 | Ocimum Sanctum | Leaf | 44.9 | −62.5 | 49.1 | −30.2 | - |
| Lamiaceae | 1 | Premna latifolia | Leaf | 42.1 | −13.5 | 67.1 | −8.64 | - |
| Lamiaceae | 1 | Tectona grandis | Leaf | 42.0 | −49.8 | 52.0 | −26.4 | - |
| Lamiaceae | 4 | Clerodendrum infortunatum | Leaf | 38.5 | −20.0 | 69.1 | 32.2 | - |
| Lamiaceae | 6 | Hyptis suaveolens | Fruit | 32.7 | −21.6 | 68.4 | 10.6 | - |
| Lamiaceae | 4 | Ocimum gratissimum | Leaf | 15.3 | −92.1 | 46.5 | −66.4 | - |
| Lamiaceae | 4 | Ocimum basilicum | Leaf | 12.6 | −72.0 | 49.0 | −55.5 | - |
| Lamiaceae | 4 | Vitex trifolia | Leaf | −2.60 | −57.1 | 47.5 | −22.2 | - |
| Lamiaceae | 6 | Gmelina arborea | Bark | 73.9 | 1.47 | 89.0 | 26.7 | ** |
| Lauraceae | 1 | Cinnamomum verrum | Bark | 91.8 | 80.0 | 100 | 100 | ** ** |
| Lauraceae | 3 | Litsea glutinosa | Leaf | 41.4 | −11.8 | 61.7 | 39.3 | - |
| Lauraceae | 4 | Cinnamomum camphora | Leaf | 36.6 | −24.5 | 84.9 | 85.9 | - |
| Lauraceae | 1 | Cinnamomum verum | Leaf | 25.1 | −21.9 | 62.6 | 21.3 | - |
| Lauraceae | 2 | Cinnamomum tamala | Leaf | 7.72 | −28.4 | 35.1 | −3.46 | - |
| Lecythidaceae | 2 | Couroupita guianensis | Fruit | 95.4 | 65.1 | 98.5 | 69.5 | ** ** |
| Lecythidaceae | 2 | Couroupita guianensis | Leaf | 71.9 | 11.8 | 80.3 | 46.0 | ** |
| Lecythidaceae | 2 | Couroupita guianensis | Flower | 44.4 | −26.5 | 63.7 | 13.2 | - |
| Lecythidaceae | 4 | Careya arborea | Leaf | 67.8 | 5.42 | 66.6 | −24.0 | ** |
| Lecythidaceae | 4 | Barringtonia acutangula | Leaf | 46.2 | −67.2 | 76.1 | −21.1 | - |
| Lecythidaceae | 5 | Gustavia superba | Leaf | 20.7 | −29.2 | 40.6 | −38.5 | - |
| Liliaceae | 1 | Asparagus racemosus | Root | 37.9 | −29.9 | 54.3 | −4.53 | - |
| Linderniaceae | 1 | Lindernia procumbens | Leaf | 2.50 | −56.3 | 41.8 | −17.7 | - |
| Lythraceae | 1 | Lawsonia inermis | Leaf | 37.3 | −6.13 | 90.5 | 53.4 | - |
| Lythraceae | 2 | Punica granatum | Leaf | 35.7 | −65.9 | 72.1 | −16.2 | - |
| Lythraceae | 4 | Lagerstroemia speciosa | Leaf | 10.6 | −72.6 | 65.9 | −0.59 | - |
| Malvaceae | 4 | Urena lobate | Leaf | 53.7 | 9.15 | 73.4 | 18.6 | * |
| Malvaceae | 1 | Sida acuta | Leaf | 51.2 | −30.9 | 75.1 | 9.84 | - |
| Malvaceae | 4 | Urena lobate | Stem | 45.0 | −37.2 | 73.5 | −17.7 | - |
| Malvaceae | 1 | Sida cordifolia | Leaf | 44.9 | −16.5 | 72.1 | −20.0 | - |
| Malvaceae | 2 | Pterospermum semisagittatum | Leaf | 43.9 | −45.3 | 69.1 | −29.0 | - |
| Malvaceae | 1 | Heritiera fomes | Leaf | 15.1 | −28.6 | 60.5 | −5.27 | - |
| Malvaceae | 1 | Hibiscus cannabinus | Leaf | 8.25 | −150 | 44.6 | −121 | - |
| Malvaceae | 1 | Bombax insigne | Leaf | −23.1 | −80.0 | 6.99 | −71.9 | - |
| Meliaceae | 1 | Azadirachta indica | Leaf | 83.3 | 77.3 | 89.0 | 81.3 | ** * |
| Meliaceae | 6 | Chukrasia tabularis | Leaf | 65.5 | −17.7 | 78.2 | −3.26 | ** |
| Meliaceae | 2 | Aphanamixis polystachya | Leaf | 37.6 | −58.4 | 100 | 100 | - |
| Meliaceae | 1 | Swietenia macrophylla | Seed | 4.82 | −31.1 | 0.92 | −50.3 | - |
| Mimosaceae | 1 | Entada rheedei | Fruit | 58.4 | 17.6 | 78.5 | 46.3 | * |
| Mimosaceae | 1 | Mimosa pudica | Leaf | 45.8 | −1.12 | 63.2 | 20.6 | - |
| Mimosaceae | 2 | Adenanthera pavonina | Leaf | 41.0 | −74.8 | 69.3 | 1.62 | - |
| Mimosaceae | 1 | Xylia xylocarpa | Leaf | 35.4 | −52.4 | 60.5 | −20.5 | - |
| Mimosaceae | 1 | Calliandra ruba | Leaf | 17.3 | −22.8 | 52.8 | −0.88 | - |
| Moraceae | 6 | Artocarpus lacucha | Leaf | 47.9 | −27.9 | 66.9 | −5.39 | - |
| Moraceae | 1 | Artocarpus altilis | Leaf | 45.0 | −18.2 | 57.9 | 28.8 | - |
| Moraceae | 6 | Ficus benghalensis | Leaf | 30.9 | −33.8 | 47.1 | −18.7 | - |
| Musaceae | 4 | Musa spp. | Peel | 6.02 | −90.8 | 41.0 | −33.3 | - |
| Myristicaceae | 1 | Myristica fragrance | Leaf | 28.0 | 7.85 | 58.9 | 44.4 | - |
| Myristicaceae | 1 | Myristica fragrance | Fruit | 11.5 | 18.9 | 51.7 | 57.0 | - |
| Myrtaceae | 1 | Syzygium aromaticum | Flower bud | 76.2 | 60.0 | 94.4 | 100 | ** |
| Myrtaceae | 1 | Syzygium firmum | Leaf | 55.9 | 12.1 | 79.3 | 33.6 | * |
| Myrtaceae | 1 | Psidium guajava | Leaf | 54.4 | −20.1 | 76.9 | 13.6 | * |
| Myrtaceae | 1 | Psidium guajava | Bark | 11.6 | −22.1 | 51.0 | −9.76 | - |
| Myrtaceae | 4 | Melaleuca citrina | Leaf | 9.03 | −26.6 | 48.4 | 2.39 | - |
| Myrtaceae | 4 | Syzygium cumini | Seed | 2.32 | −31.3 | 40.9 | 2.07 | - |
| Myrtaceae | 2 | Syzygium fruticosum | Leaf | −0.62 | −37.9 | 50.2 | −13.3 | - |
| Oleaceae | 2 | Jasminum scandes | Leaf | 37.3 | −7.50 | 69.7 | 26.2 | - |
| Onagraceae | 1 | Ludwigia octovalvis | Leaf | 49.6 | 27.3 | 84.7 | 42.6 | - |
| Orchidaceae | 6 | Geodorum densiflorum | Leaf | −16.9 | −62.9 | 22.6 | −39.4 | - |
| Oxalidaceae | 2 | Averrhoa bilimbi | Leaf | 28.1 | −15.8 | 57.6 | 5.49 | - |
| Pandanaceae | 2 | Pandanus amaryllifolius | Leaf | 32.3 | −40.9 | 62.3 | 16.2 | - |
| Pandanaceae | 6 | Pandanus tectorius | Leaf | 18.3 | −46.6 | 32.7 | −31.4 | - |
| Phyllanthaceae | 6 | Phyllanthus emblica | Fruit | 95.4 | 76.1 | 96.1 | 78.0 | ** ** |
| Phyllanthaceae | 3 | Phyllanthus urinaria | Leaf | 23.6 | −93.4 | 30.6 | −76.7 | - |
| Phyllanthaceae | 6 | Phyllanthus niruri | Leaf | 4.93 | −59.6 | 31.9 | −21.9 | - |
| Piperaceae | 1 | Piper longum | Fruit | 91.3 | 62.9 | 96.0 | 77.8 | ** ** |
| Piperaceae | 1 | Piper nigrum | Fruit | 77.5 | 59.2 | 87.0 | 74.2 | ** * |
| Piperaceae | 1 | Piper chaba | Leaf | 71.6 | 22.1 | 87.1 | 56.9 | ** |
| Plantagenaceae | 1 | Plantago scabra | Seed | 14.9 | −40.4 | 24.8 | −60.4 | - |
| Plumbaginaceae | 2 | Aegialitis rotundifolia | Leaf | 43.2 | −22.6 | 76.7 | 31.2 | - |
| Poaceae | 1 | Axonopus compressus | Leaf | 75.6 | 20.3 | 83.9 | 43.6 | ** |
| Poaceae | 1 | Dendrocalamus longispathus | Leaf | 50.6 | −5.73 | 55.2 | −5.50 | - |
| Poaceae | 1 | Dendrocalamus giganteus | Leaf | 41.6 | −38.2 | 91.0 | 66.2 | - |
| Poaceae | 2 | Cymbopogon citratus | Leaf | 34.8 | −27.6 | 100 | 100 | - |
| Poaceae | 1 | Cynodon dactylon | Leaf | 31.0 | −54.8 | 38.0 | −37.3 | - |
| Poaceae | 2 | Dactyloctenium aegyptium | Leaf | 18.0 | −45.4 | 57.9 | −17.5 | - |
| Podocarpacea | 1 | Podocarpus neriifolius | Leaf | 74.7 | 53.6 | 99.2 | 100 | ** |
| Ranunculaceae | 6 | Nigella sativa | Seed | 68.2 | 39.2 | 70.7 | 41.4 | ** |
| Rhamnaceae | 1 | Ziziphus mauritiana | Leaf | 48.5 | −43.0 | 64.2 | −19.1 | - |
| Rhizophoraceae | 1 | Carallia brachiate | Leaf | 34.3 | −23.2 | 62.4 | −19.2 | - |
| Rubiaceae | 5 | Spermacoce mauritiana | Leaf | 54.7 | 8.17 | 66.6 | −5.86 | * |
| Rubiaceae | 4 | Morinda citrifolia | Leaf | 52.4 | −7.77 | 82.5 | 52.8 | - |
| Rubiaceae | 3 | Paederia foetida | Leaf | 51.7 | −13.2 | 74.4 | −7.43 | - |
| Rubiaceae | 1 | Mitragyna parvifolia | Leaf | 47.4 | −5.50 | 73.0 | 18.4 | - |
| Rubiaceae | 1 | Exeocaicaria bi-color | Leaf | 34.2 | −29.0 | 80.7 | 18.3 | - |
| Rubiaceae | 1 | Haldina cordifolia | Leaf | 34.0 | −20.3 | 56.0 | 11.0 | - |
| Rubiaceae | 1 | Gardenia coronaria | Leaf | 17.1 | −66.2 | 51.4 | −37.2 | - |
| Rutaceae | 1 | Aegle marmelos | Leaf | 86.1 | 14.3 | 74.8 | 0.79 | ** * |
| Rutaceae | 6 | Aegle marmelos | Fruit | 75.5 | 7.29 | 84.8 | 38.2 | ** |
| Rutaceae | 4 | Citrus medica | Leaf | 64.6 | −1.33 | 74.9 | 34.8 | * |
| Rutaceae | 2 | Melicope triphylla | Leaf | 56.1 | −3.30 | 87.2 | 44.2 | * |
| Rutaceae | 1 | Glycosmis pentaphylla | Leaf | 52.8 | 17.7 | 82.5 | 70.2 | - |
| Rutaceae | 1 | Murraya paniculata | Leaf | 49.0 | −2.09 | 77.9 | 37.8 | - |
| Rutaceae | 1 | Limonia acidissima | Leaf | 47.4 | −15.1 | 65.1 | 11.2 | - |
| Rutaceae | 4 | Clausena heptaphylla | Leaf | 39.3 | −10.3 | 75.1 | 35.2 | - |
| Rutaceae | 4 | Citrus medica | Bark | 31.2 | −29.2 | 43.8 | −13.8 | - |
| Rutaceae | 1 | Zanthoxylum rhetsa | Leaf | 18.7 | −31.7 | 49.2 | 10.3 | - |
| Sapindaceae | 6 | Sapindus mukorossi | Fruit | 82.8 | 39.0 | 91.1 | 52.5 | ** * |
| Sapindaceae | 5 | Lepisanthes rubiginosa | Leaf | 64.7 | −11.2 | 71.1 | −13.6 | * |
| Sapindaceae | 1 | Dimocarpus longan | Leaf | 63.3 | −10.2 | 76.4 | 6.74 | * |
| Sapindaceae | 1 | Nephelium longana | Flower | 57.2 | −11.8 | 71.9 | 26.2 | * |
| Sapindaceae | 1 | Litchi chinensis | Leaf | 55.2 | 16.1 | 75.2 | 48.5 | * |
| Sapindaceae | 1 | Lepisanthes alata | Leaf | 37.0 | −33.3 | 62.4 | −7.30 | - |
| Sapindaceae | 1 | schleichera oleosa | Leaf | 30.6 | −16.2 | 76.8 | 53.4 | - |
| Sapindaceae | 1 | Dodonaea viscosa | Leaf | 3.49 | −65.1 | 79.5 | 27.6 | - |
| Sapotaceae | 2 | Chrysophyllum cainita | Leaf | 83.5 | 5.51 | 76.8 | −0.46 | ** * |
| Sapotaceae | 1 | Mahua longifolia | Flower | 60.3 | −37.4 | 65.5 | −30.3 | * |
| Sapotaceae | 1 | Mimusops elegi varigata | Leaf | 34.3 | −24.7 | 78.2 | 32.8 | - |
| Sapotaceae | 1 | Mimusops elengi | Leaf | 29.6 | −30.2 | 44.6 | −23.5 | - |
| Scrophulariaceae | 2 | Limnophila repens | Leaf | 25.6 | −60.6 | 34.0 | −27.6 | - |
| Solanaceae | 6 | Datura metel | Seed | 33.7 | −18.3 | 49.0 | 4.63 | - |
| Solanaceae | 2 | Solanum tuberosum | Peel | 23.9 | −14.3 | 56.2 | 23.7 | - |
| Sterculiaceae | 2 | Abroma augustum | Leaf | 62.5 | −68.3 | 66.4 | −33.8 | * |
| Sterculiaceae | 2 | Sterculia villosa | Leaf | 37.2 | 15.4 | 55.2 | 32.1 | - |
| Theaceae | 5 | Camellia sinensis | Leaf | 39.3 | −41.2 | 63.1 | 40.1 | - |
| Thymelaeaceae | 1 | Aquilaria khasiana | Leaf | 28.2 | −44.7 | 78.3 | −10.0 | - |
| Urticaceae | 4 | Boehmeria macrophylla | Leaf | 59.7 | −3.22 | 74.6 | 11.3 | * |
| Verbenaceae | 1 | Duranta repens | Leaf | 84.2 | 39.3 | 80.8 | 27.1 | ** * |
| Verbenaceae | 4 | Lantana camara | Leaf | 57.4 | 19.7 | 85.2 | 61.8 | * |
| Verbenaceae | 3 | Clerodendrum indicum | Leaf | 42.9 | −51.6 | 77.8 | −23.0 | - |
| Verbenaceae | 2 | Lippia gerinate | Leaf | 30.8 | −40.3 | 62.0 | 34.5 | - |
| Verbenaceae | 1 | Nyctanthes arbortristis | Leaf | 28.4 | −67.2 | 56.1 | −19.8 | - |
| Verbenaceae | 2 | Vitex negundo | Leaf | 24.1 | −44.1 | 50.4 | −7.20 | - |
| Zingiberaceae | 6 | Kaempferia galanga | Root | 71.3 | 29.4 | 82.1 | 68.9 | ** |
| Zingiberaceae | 4 | Curcuma roxburghii | Leaf | 38.5 | −16.1 | 72.6 | 40.7 | - |
| Zingiberaceae | 3 | Curcuma aromatica | Leaf | 19.6 | −8.74 | 55.8 | 37.3 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Begum, K.; Shammi, M.; Hasan, N.; Asaduzzaman, M.; Appiah, K.S.; FUJII, Y. Potential Allelopathic Candidates for Land Use and Possible Sustainable Weed Management in South Asian Ecosystem. Sustainability 2019, 11, 2649. https://doi.org/10.3390/su11092649
Begum K, Shammi M, Hasan N, Asaduzzaman M, Appiah KS, FUJII Y. Potential Allelopathic Candidates for Land Use and Possible Sustainable Weed Management in South Asian Ecosystem. Sustainability. 2019; 11(9):2649. https://doi.org/10.3390/su11092649
Chicago/Turabian StyleBegum, Kohinoor, Mashura Shammi, Nazmul Hasan, Md. Asaduzzaman, Kwame Sarpong Appiah, and Yoshiharu FUJII. 2019. "Potential Allelopathic Candidates for Land Use and Possible Sustainable Weed Management in South Asian Ecosystem" Sustainability 11, no. 9: 2649. https://doi.org/10.3390/su11092649