
Roles of Medicinal Mushrooms as Natural Food Dyes and Dye-Sensitised Solar Cells (DSSC): Synergy of Zero Hunger and Affordable Energy for Sustainable Development
 , ,
, ,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Medicinal Mushroom Dyes
- Polyketide synthetic pathway; Antrodia cinnamomea [34]—allowing the formation of melanin, flavins, ankaflavin, quinones, and azaphilones. Aromatic ketides or fatty acids are formed via this pathway. Cyclisation reactions and partial reductions lead to stabilisation of the growing chain in the synthesis of aromatic ketides. In the case of fatty acid synthesis, the carbonyl groups of the chain are reduced prior to binding to the next C2 group. This process produces tetra-, hepta-, octa-, and higher number of aromatic ketides as well as fatty acid-derived compounds;
- Shikimate pathway; Agaricus bisporus [35]—the key intermediates of shikimic and chorismic acids are used to manufacture the important amino acids tyrosine, tryptophan, and phenylalanine. Various important building components of many colours found in fungi, such as benzoic, arylpyruvic, and cinnamic acids, are produced from tyrosine and phenylalanine precursors. Tyrosine is crucial in the phylum Basidiomycota because it is a precursor of betalaine, a pigment found solely in the genera Hygrocybe and Amanita;
- Terpenoid synthetic pathway; Hypsizygus marmoreus [36]—significant for the formation of carotenoids belonging to terpenoids. The condensation routes of C5 isoprene units and terpenoid synthesis are triggered during carotenoids production. A common precursor to terpenoid formation is isopentenyl pyrophosphate, which is formed by mevalonate pathway;
- Nitrogen-containing metabolite pathway; Agaricus bisporus [37]—ensure the formation of various fungal pigments.
3. Medicinal Mushroom Pigments in Food Production
4. Isolation of Mushroom Dyes
5. Natural Colourants in the Dye-Sensitized Solar Cell (DSSC)
6. Medicinal Mushroom Dye as an Active Photo-Energy Conversion Molecules in DSSCs
7. Safety, Production, Limitations, and Future Strategy of Medicinal Mushroom Dyes in DSSC
- Dye-sensitised solar cells (DSSC);
- Solar cells with bulk heterojunction (BHJ);
- Solar cells with quantum dots (QDSSC);
- Solar cells made of perovskite.
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Summary for Policymakers; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- Eisenmenger, N.; Pichler, M.; Krenmayr, N.; Noll, D.; Plank, B.; Schalmann, E.; Wandl, M.-T.; Gingrich, S. The Sustainable Development Goals prioritize economic growth over sustainable resource use: A critical reflection on the SDGs from a socio-ecological perspective. Sustain. Sci. 2020, 15, 1101–1110. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, K. Sustainable development: Dimensions, intersections and knowledge platform. Sustain. Fundam. Appl. 2020, 43–68. [Google Scholar] [CrossRef]
- Sachs, J.D. From Millennium Development Goals to Sustainable Development Goals. Lancet 2012, 379, 2206–2211. [Google Scholar] [CrossRef]
- Le Blanc, D. Towards Integration at Last? The Sustainable Development Goals as a Network of Targets. Sustain. Dev. 2015, 23, 176–187. [Google Scholar] [CrossRef]
- Gratzer, G.; Winiwarter, V. Chancen und Herausforderungen bei der Umsetzung der UN-Nachhaltigkeitsziele aus österreichischer Sicht. KIOES Opin 2018, 8, 13–26. [Google Scholar]
- Kalra, R.; Conlan, X.A.; Goel, M. Fungi as a Potential Source of Pigments: Harnessing Filamentous Fungi. Front. Chem. 2020, 8, 369. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Garcia, J.A.A.; Correa, V.G.; Vieira, T.F.; Bracht, A.; Peralta, R.M. Pigments and vitamins from plants as functional ingredients: Current trends and perspectives. Adv. Food Nutr. Res. 2019, 90, 259–303. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J. Fungal Pigments and Their Roles Associated with Human Health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef]
- Galaffu, N.; Bortlik, K.; Michel, M. 5—An industry perspective on natural food colour stability. In Colour Additives for Foods and Beverages; Woodhead Publishing Series in Food Science, Technology and Nutrition; Elsevier: Amsterdam, The Netherlands, 2015; pp. 91–130. [Google Scholar]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous ascomycetes fungi as a source of natural pigments. Fungal Biol. Biotechnol. 2017, 4, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Méndez, A.; Pérez, C.; Montañez, J.; Martínez, G.; Aguilar, C.N. Red pigment production by Penicillium purpurogenum GH2 is influenced by pH and temperature. J. Zhejiang Univ. Sci. B 2011, 12, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Lagashetti, A.C.; Räisänen, R.; Robinson, S.C.; Singh, S.K.; Dufossé, L. Colorful Macrofungi and their Pigment Structures. In Advances in Macrofungi; CRC Press: Boca Raton, FL, USA, 2021; pp. 252–322. [Google Scholar]
- Suthar, M.; Lagashetti, A.C.; Räisänen, R.; Singh, P.N.; Dufossé, L.; Robinson, S.C.; Singh, S.K. Industrial Applications of Pigments from Macrofungi. In Advances in Macrofungi; CRC Press: Boca Raton, FL, USA, 2021; pp. 223–251. [Google Scholar]
- Akilandeswari, P.; Pradeep, B.V. Exploration of industrially important pigments from soil fungi. Appl. Microbiol. Biotechnol. 2015, 100, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Albertsen, J.; Albertsen, J.; Sørensen, P.G. Chemistry of Dyeing. In Proceedings of the 17th International Fungi & Fibre Symposium, Madeira Park, BC, Canada, 17–22 October 2016. [Google Scholar]
- Dong, J.Z.; Wang, S.H.; Ai, X.R.; Yao, L.; Sun, Z.W.; Lei, C.; Wang, Y.; Wang, Q. Composition and characterization of cordyxanthins from Cordyceps militaris fruit bodies. J. Funct. Foods 2013, 5, 1450–1455. [Google Scholar] [CrossRef]
- Spiteller, P.; Arnold, N.; Spiteller, M.; Steglich, W. Lilacinone, a Red Aminobenzoquinone Pigment from Lactarius lilacinus. J. Nat. Prod. 2003, 66, 1402–1403. [Google Scholar] [CrossRef] [PubMed]
- Prados-Rosales, R.; Toriola, S.; Nakouzi, A.; Chatterjee, S.; Stark, R.; Gerfen, G.; Tumpowsky, P.; Dadachova, E.; Casadevall, A. Structural Characterization of Melanin Pigments from Commercial Preparations of the Edible Mushroom Auricularia auricula. J. Agric. Food Chem. 2015, 63, 7326–7332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvakumar, P.; Rajasekar, S.; Periasamy, K.; Raaman, N. Isolation and characterization of melanin pigment from Pleurotus cystidiosus (telomorph of Antromycopsis macrocarpa). World J. Microbiol. Biotechnol. 2008, 24, 2125–2131. [Google Scholar] [CrossRef]
- Ribera, J.; Panzarasa, G.; Stobbe, A.; Osypova, A.; Rupper, P.; Klose, D.; Schwarze, F.W.M.R. Scalable Biosynthesis of Melanin by the Basidiomycete Armillaria cepistipes. J. Agric. Food Chem. 2018, 67, 132–139. [Google Scholar] [CrossRef]
- Weijn, A.; Bastiaan-Net, S.; Wichers, H.; Mes, J. Melanin biosynthesis pathway in Agaricus bisporus mushrooms. Fungal Genet. Biol. 2013, 55, 42–53. [Google Scholar] [CrossRef]
- Weijn, A.; Berg-Somhorst, D.B.P.M.V.D.; Slootweg, J.C.; Vincken, J.-P.; Gruppen, H.; Wichers, H.; Mes, J.J. Main Phenolic Compounds of the Melanin Biosynthesis Pathway in Bruising-Tolerant and Bruising-Sensitive Button Mushroom (Agaricus bisporus) Strains. J. Agric. Food Chem. 2013, 61, 8224–8231. [Google Scholar] [CrossRef]
- Yan, D.; Liu, Y.; Rong, C.; Song, S.; Zhao, S.; Qin, L.; Wang, S.; Gao, Q. Characterization of brown film formed by Lentinula edodes. Fungal Biol. 2020, 124, 135–143. [Google Scholar] [CrossRef]
- Orona-Navar, A.; Aguilar-Hernández, I.; Nigam, K.; Cerdán-Pasarán, A.; Ornelas-Soto, N. Alternative sources of natural pigments for dye-sensitized solar cells: Algae, cyanobacteria, bacteria, archaea and fungi. J. Biotechnol. 2021, 332, 29–53. [Google Scholar] [CrossRef]
- Kumara, N.; Lim, A.; Lim, C.M.; Petra, M.I.; Ekanayake, P. Recent progress and utilization of natural pigments in dye sensitized solar cells: A review. Renew. Sustain. Energy Rev. 2017, 78, 301–317. [Google Scholar] [CrossRef]
- O’Regan, B.; Grätzel, M. A low-cost, high-efficiency solar cell based on dye-sensitized colloidal TiO2 films. Nature 1991, 353, 737–740. [Google Scholar] [CrossRef]
- Rao, M.P.N.; Xiao, M.; Li, W.-J. Fungal and Bacterial Pigments: Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.; Shabbir, M.; Mohammad, F. Natural colorants: Historical, processing and sustainable prospects. Nat. Prod. Prod. Prod. Bioprospecting 2017, 7, 123–145. [Google Scholar] [CrossRef] [Green Version]
- Ntrallou, K.; Gika, H.; Tsochatzis, E. Analytical and Sample Preparation Techniques for the Determination of Food Colorants in Food Matrices. Foods 2020, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Velíšek, J.; Cejpek, K. Pigments of higher fungi—A review. Czech J. Food Sci. 2011, 29, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Dufossé, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef]
- Yu, P.-W.; Chang, Y.-C.; Liou, R.-F.; Lee, T.-H.; Tzean, S.-S. pks63787, a Polyketide Synthase Gene Responsible for the Biosynthesis of Benzenoids in the Medicinal Mushroom Antrodia cinnamomea. J. Nat. Prod. 2016, 79, 1485–1491. [Google Scholar] [CrossRef]
- Yang, X.; Yang, K.; Wang, X.; Wang, Y.; Zhao, Z.; Meng, D. Transcriptomic analysis reveals the mechanism of bacterial disease resistance of postharvest button mushroom (Agaricus bisporus). Physiol. Mol. Plant Pathol. 2022, 122, 101903. [Google Scholar] [CrossRef]
- Min, B.; Kim, S.; Oh, Y.-L.; Kong, W.-S.; Park, H.; Cho, H.; Jang, K.-Y.; Kim, J.-G.; Choi, I.-G. Genomic discovery of the hypsin gene and biosynthetic pathways for terpenoids in Hypsizygus marmoreus. BMC Genom. 2018, 19, 789. [Google Scholar] [CrossRef]
- Baars, J.J.P.; Camp, H.J.M.O.D.; Hermans, J.M.H.; Mikeš, V.; Van Der Drift, C.; Van Griensven, L.J.L.D.; Vogels, G.D. Nitrogen assimilating enzymes in the white button mushroom Agaricus bisporus. Microbiology 1994, 140, 1161–1168. [Google Scholar] [CrossRef]
- Klaus, A.; Wan-Mohtar, W.A.A.Q.I.; Nikolić, B.; Cvetković, S.; Vunduk, J. Pink oyster mushroom Pleurotus flabellatus mycelium produced by an airlift bioreactor—The evidence of potent in vitro biological activities. World J. Microbiol. Biotechnol. 2021, 37, 1–14. [Google Scholar] [CrossRef]
- Ma, G.; Yang, W.; Zhao, L.; Pei, F.; Fang, D.; Hu, Q. A critical review on the health promoting effects of mushrooms nutraceuticals. Food Sci. Hum. Wellness 2018, 7, 125–133. [Google Scholar] [CrossRef]
- Yang, X.-L.; Qin, C.; Wang, F.; Dong, Z.-J.; Liu, J.-K. A New Meroterpenoid Pigment from the Basidiomycete Albatrellus confluens. Chem. Biodivers. 2008, 5, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, A.; Li, W.M.; Liu, V.; Wang, C.; Mackedenski, S.; Tackaberry, L.E.; Massicotte, H.B.; Egger, K.N.; Reimer, K.; Lee, C.H. Grifolin, neogrifolin and confluentin from the terricolous polypore Albatrellus flettii suppress KRAS expression in human colon cancer cells. PLoS ONE 2020, 15, e0231948. [Google Scholar] [CrossRef] [PubMed]
- Bin, L.; Wei, L.; Xiaohong, C.; Mei, J.; Mingsheng, D. In vitro antibiofilm activity of the melanin from Auricularia auricula, an edible jelly mushroom. Ann. Microbiol. 2012, 62, 1523–1530. [Google Scholar] [CrossRef]
- Zou, Y.; Hu, W.; Ma, K.; Tian, M. Physicochemical properties and antioxidant activities of melanin and fractions from Auricularia auricula fruiting bodies. Food Sci. Biotechnol. 2015, 24, 15–21. [Google Scholar] [CrossRef]
- Song, J.; Manir, M.; Moon, S.-S. Cytotoxic Grifolin Derivatives Isolated from the Wild Mushroom Boletus pseudocalopus (Basidiomycetes). Chem. Biodivers. 2009, 6, 1435–1442. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Vunduk, J.; Zizak, Z.; Niksic, M.; Jakovljevic, D.; Vrvic, M.M.; Van Griensven, L.J.L.D. Nutraceutical properties of the methanolic extract of edible mushroom Cantharellus cibarius (Fries): Primary mechanisms. Food Funct. 2015, 6, 1875–1886. [Google Scholar] [CrossRef] [Green Version]
- Tudor, D.; Robinson, S.C.; Cooper, P.A. The influence of pH on pigment formation by lignicolous fungi. Int. Biodeterior. Biodegradation 2013, 80, 22–28. [Google Scholar] [CrossRef]
- Velmurugan, P.; Lee, Y.H.; Nanthakumar, K.; Kamala-Kannan, S.; Dufossé, L.; Mapari, S.A.; Oh, B.T. Water-soluble red pigments from Isaria farinosa and structural characterization of the main colored component. J. Basic Microbiol. 2010, 50, 581–590. [Google Scholar] [CrossRef]
- Lee, J.-H.; Hyun, C.-K. Insulin-Sensitizing and Beneficial Lipid-Metabolic Effects of the Water-Soluble Melanin Complex Extracted from Inonotus obliquus. Phytother. Res. 2014, 28, 1320–1328. [Google Scholar] [CrossRef]
- Caro, Y.; Venkatachalam, M.; Lebeau, J.; Fouillaud, M.; Dufossé, L. Pigments and Colorants from Filamentous Fungi. Fungal Metab. 2017, 25, 499–568. [Google Scholar] [CrossRef]
- Kim, K.H.; Noh, H.J.; Choi, S.U.; Park, K.M.; Seok, S.-J.; Lee, K.R. Lactarane sesquiterpenoids from Lactarius subvellereus and their cytotoxicity. Bioorganic Med. Chem. Lett. 2010, 20, 5385–5388. [Google Scholar] [CrossRef] [PubMed]
- Davoli, P.; Mucci, A.; Schenetti, L.; Weber, R.W. Laetiporic acids, a family of non-carotenoid polyene pigments from fruit-bodies and liquid cultures of Laetiporus sulphureus (Polyporales, Fungi). Phytochemistry 2005, 66, 817–823. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, X.; Huang, C.; Zhang, Z.; Gao, W. Isolation and identification of pigments from oyster mushrooms with black, yellow and pink caps. Food Chem. 2021, 372, 131171. [Google Scholar] [CrossRef]
- De Souza, R.A.; Kamat, N.M.; Nadkarni, V.S. Purification and characterisation of a sulphur rich melanin from edible mushroom Termitomyces albuminosus Heim. Mycology 2018, 9, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Harki, E.; Talou, T.; Dargent, R. Purification, characterisation and analysis of melanin extracted from Tuber melanosporum Vitt. Food Chem. 1997, 58, 69–73. [Google Scholar] [CrossRef]
- Dufossé, L.; Galaup, P.; Yaron, A.; Arad, S.M.; Blanc, P.; Murthy, K.N.C.; Ravishankar, G.A. Microorganisms and microalgae as sources of pigments for food use: A scientific oddity or an industrial reality? Trends Food Sci. Technol. 2005, 16, 389–406. [Google Scholar] [CrossRef]
- Barredo, J.L.; García-Estrada, C.; Kosalkova, K.; Barreiro, C. Biosynthesis of Astaxanthin as a Main Carotenoid in the Heterobasidiomycetous Yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-Y.; Liu, J.-K. Pigments of fungi (macromycetes). Nat. Prod. Rep. 2010, 27, 1531–1570. [Google Scholar] [CrossRef]
- Pacelli, C.; Cassaro, A.; Maturilli, A.; Timperio, A.M.; Gevi, F.; Cavalazzi, B.; Stefan, M.; Ghica, D.; Onofri, S. Multidisciplinary characterization of melanin pigments from the black fungus Cryomyces antarcticus. Appl. Microbiol. Biotechnol. 2020, 104, 6385–6395. [Google Scholar] [CrossRef]
- Wold, C.W.; Gerwick, W.H.; Wangensteen, H.; Inngjerdingen, K.T. Bioactive triterpenoids and water-soluble melanin from Inonotus obliquus (Chaga) with immunomodulatory activity. J. Funct. Foods 2020, 71, 104025. [Google Scholar] [CrossRef]
- Räisänen, R. Fungal colorants in applications–focus on Cortinarius species. Color. Technol. 2019, 135, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Mattoon, E.; Cordero, R.; Casadevall, A. Fungal Melanins and Applications in Healthcare, Bioremediation and Industry. J. Fungi 2021, 7, 488. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Martins, A.; Vasconcelos, M.H.; Morales, P.; Ferreira, I.C.F.R. Functional foods based on extracts or compounds derived from mushrooms. Trends Food Sci. Technol. 2017, 66, 48–62. [Google Scholar] [CrossRef]
- Wan-Mohtar, W.A.A.Q.I.; Taufek, N.M.; Yerima, G.; Rahman, J.; Thiran, J.P.; Subramaniam, K.; Sabaratnam, V. Effect of bioreactor-grown biomass from Ganoderma lucidum mycelium on growth performance and physiological response of red hybrid tilapia (Oreochromis sp.) for sustainable aquaculture. Org. Agric. 2020, 11, 327–335. [Google Scholar] [CrossRef]
- Neves, M.I.L.; Silva, E.K.; Meireles, M.A.A. Natural blue food colorants: Consumer acceptance, current alternatives, trends, challenges, and future strategies. Trends Food Sci. Technol. 2021, 112, 163–173. [Google Scholar] [CrossRef]
- Sulieman, A.A.; Zhu, K.-X.; Peng, W.; Shoaib, M.; Obadi, M.; Hassanin, H.A.M.; Alahmad, K.; Zhou, H.-M. Assessment of rheological, physicochemical, and staling characteristics of gluten-free dough and bread containing Agaricus bisporus polysaccharide flour and inulin. J. Food Meas. Charact. 2018, 12, 2032–2044. [Google Scholar] [CrossRef]
- Ulziijargal, E.; Yang, J.-H.; Lin, L.-Y.; Chen, C.-P.; Mau, J.-L. Quality of bread supplemented with mushroom mycelia. Food Chem. 2013, 138, 70–76. [Google Scholar] [CrossRef]
- Szydłowska-Tutaj, M.; Złotek, U.; Combrzyński, M. Influence of addition of mushroom powder to semolina on proximate composition, physicochemical properties and some safety parameters of material for pasta production. LWT 2021, 151, 112235. [Google Scholar] [CrossRef]
- Parvin, R.; Farzana, T.; Mohajan, S.; Rahman, H.; Rahman, S.S. Quality improvement of noodles with mushroom fortified and its comparison with local branded noodles. NFS J. 2020, 20, 37–42. [Google Scholar] [CrossRef]
- Olawuyi, I.F.; Lee, W.Y. Quality and antioxidant properties of functional rice muffins enriched with shiitake mushroom and carrot pomace. Int. J. Food Sci. Technol. 2019, 54, 2321–2328. [Google Scholar] [CrossRef]
- Salehi, F.; Kashaninejad, M.; Asadi, F.; Najafi, A. Improvement of quality attributes of sponge cake using infrared dried button mushroom. J. Food Sci. Technol. 2016, 53, 1418–1423. [Google Scholar] [CrossRef] [Green Version]
- Wan-Mohtar, W.A.A.Q.I.; Mahmud, N.; Supramani, S.; Ahmad, R.; Zain, N.A.M.; Hassan, N.A.M.; Peryasamy, J.; Halim-Lim, S.A. Fruiting-body-base flour from an oyster mushroom—A waste source of antioxidative flour for developing potential functional cookies and steamed-bun. AIMS Agric. Food 2018, 3, 481–492. [Google Scholar] [CrossRef]
- Kurt, A.; Gençcelep, H. Enrichment of meat emulsion with mushroom (Agaricus bisporus) powder: Impact on rheological and structural characteristics. J. Food Eng. 2018, 237, 128–136. [Google Scholar] [CrossRef]
- Pérez-Montes, A.; Rangel-Vargas, E.; Lorenzo, J.M.; Romero, L.; Santos, E.M. Edible mushrooms as a novel trend in the development of healthier meat products. Curr. Opin. Food Sci. 2020, 37, 118–124. [Google Scholar] [CrossRef]
- Wan-Mohtar, W.A.A.Q.I.; Halim-Lim, S.A.; Kamarudin, N.Z.; Rukayadi, Y.; Rahim, M.H.A.; Jamaludin, A.A.; Ilham, Z. Fruiting-body-base flour from an Oyster mushroom waste in the development of antioxidative chicken patty. J. Food Sci. 2020, 85, 3124–3133. [Google Scholar] [CrossRef]
- Cerón-Guevara, M.I.; Santos, E.M.; Lorenzo, J.M.; Pateiro, M.; Bermúdez-Piedra, R.; Rodríguez, J.A.; Castro-Rosas, J.; Rangel-Vargas, E. Partial replacement of fat and salt in liver pâté by addition of Agaricus bisporus and Pleurotus ostreatus flour. Int. J. Food Sci. Technol. 2021, 56, 6171–6181. [Google Scholar] [CrossRef]
- Stephan, A.; Ahlborn, J.; Zajul, M.; Zorn, H. Edible mushroom mycelia of Pleurotus sapidus as novel protein sources in a vegan boiled sausage analog system: Functionality and sensory tests in comparison to commercial proteins and meat sausages. Eur. Food Res. Technol. 2017, 244, 913–924. [Google Scholar] [CrossRef]
- Keerthana, K.; Anukiruthika, T.; Moses, J.; Anandharamakrishnan, C. Development of fiber-enriched 3D printed snacks from alternative foods: A study on button mushroom. J. Food Eng. 2020, 287, 110116. [Google Scholar] [CrossRef]
- Pecić, S.; Nikićević, N.; Veljović, M.; Jardanin, M.; Tešević, V.; Belović, M.; Nikšić, M. The influence of extraction parameters on physicochemical properties of special grain brandies with Ganoderma lucidum. License 2015, 22, 181–189. [Google Scholar]
- Nguyen, A.N.; Capone, D.L.; Johnson, T.E.; Jeffery, D.W.; Danner, L.; Bastian, S.E. Volatile Composition and Sensory Profiles of a Shiraz Wine Product Made with Pre- and Post-Fermentation Additions of Ganoderma lucidum Extract. Foods 2019, 8, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vunduk, J.; Veljović, S. Macrofungi in the Production of Alcoholic Beverages Beer, Wine, and Spirits. In Advances in Macrofungi; CRC Press: Boca Raton, FL, USA, 2021; pp. 108–141. [Google Scholar]
- Belscak-Cvitanovic, A.; Nedovic, V.; Salevic, A.; Despotovic, S.; Komes, D.; Niksic, M.; Bugarski, B.; Leskosek-Cukalovic, I. Modification of functional quality of beer by using microencapsulated green tea (Camellia sinensis L.) and Ganoderma mushroom (Ganoderma lucidum L.) bioactive compounds. Chem. Ind. Chem. Eng. Q. 2017, 23, 457–471. [Google Scholar] [CrossRef] [Green Version]
- Martins, N.; Roriz, C.L.; Morales, P.; Barros, L.; Ferreira, I.C. Food colorants: Challenges, opportunities and current desires of agro-industries to ensure consumer expectations and regulatory practices. Trends Food Sci. Technol. 2016, 52, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhang, M.; Yang, H.; Zhou, H.; Yang, H. Production, physico-chemical characterization and antioxidant activity of natural melanin from submerged cultures of the mushroom Auricularia auricula. Food Biosci. 2018, 26, 49–56. [Google Scholar] [CrossRef]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal Pigments and Their Prospects in Different Industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, H.; Lou, Q.; Liu, H.; Han, H.; Wang, Q.; Tang, Z.; Ma, Y.; Wang, H. Differential Regulation of Anthocyanins in Green and Purple Turnips Revealed by Combined De Novo Transcriptome and Metabolome Analysis. Int. J. Mol. Sci. 2019, 20, 4387. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, S.; Xu, X.; Lowry, D.; Jackson, J.C.; Roberson, R.W.; Lin, X. Subcellular Compartmentalization and Trafficking of the Biosynthetic Machinery for Fungal Melanin. Cell Rep. 2016, 14, 2511–2518. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, J.R.; Junior, R.N.D.C. Occurrence and possible roles of polysaccharides in fungi and their influence on the development of new technologies. Carbohydr. Polym. 2020, 246, 116613. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kempf, V.R.; Nofsinger, J.B.; Weinert, E.E.; Rudnicki, M.; Wakamatsu, K.; Ito, S.; Simon, J.D. Comparison of the Structural and Physical Properties of Human Hair Eumelanin Following Enzymatic or Acid/Base Extraction. Pigment Cell Res. 2003, 16, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.-G.; Sun, Y.-Z.; Tian, Y.-G.; Xie, M.-Y.; Chen, J. Physicochemical characterisation and antioxidant activity of melanin from the muscles of Taihe Black-bone silky fowl (Gallus gallus domesticus Brisson). Food Chem. 2009, 114, 1345–1350. [Google Scholar] [CrossRef]
- Grätzel, M. Recent Advances in Sensitized Mesoscopic Solar Cells. Accounts Chem. Res. 2009, 42, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Yahya, M.; Bouziani, A.; Ocak, C.; Seferoğlu, Z.; Sillanpää, M. Organic/metal-organic photosensitizers for dye-sensitized solar cells (DSSC): Recent developments, new trends, and future perceptions. Dye. Pigment. 2021, 192, 109227. [Google Scholar] [CrossRef]
- Al-Alwani, M.A.; Mohamad, A.B.; Ludin, N.A.; Kadhum, A.A.H.; Sopian, K. Dye-sensitised solar cells: Development, structure, operation principles, electron kinetics, characterisation, synthesis materials and natural photosensitisers. Renew. Sustain. Energy Rev. 2016, 65, 183–213. [Google Scholar] [CrossRef]
- Jamalullail, N.; Smohamad, I.; Nnorizan, M.; Mahmed, N. Enhancement of Energy Conversion Efficiency for Dye Sensitized Solar Cell Using Zinc Oxide Photoanode. IOP Conf. Ser. Mater. Sci. Eng. 2018, 374, 012048. [Google Scholar] [CrossRef]
- Solaiyammal, T.; Murugakoothan, P. Green synthesis of Au and the impact of Au on the efficiency of TiO2 based dye sensitized solar cell. Mater. Sci. Energy Technol. 2019, 2, 171–180. [Google Scholar] [CrossRef]
- Cherepy, N.J.; Smestad, G.P.; Grätzel, A.M.; Zhang, J.Z. Ultrafast Electron Injection: Implications for a Photoelectrochemical Cell Utilizing an Anthocyanin Dye-Sensitized TiO2 Nanocrystalline Electrode. J. Phys. Chem. B 1997, 101, 9342–9351. [Google Scholar] [CrossRef]
- Gong, J.; Sumathy, K.; Qiao, Q.; Zhou, Z. Review on dye-sensitized solar cells (DSSCs): Advanced techniques and research trends. Renew. Sustain. Energy Rev. 2016, 68, 234–246. [Google Scholar] [CrossRef]
- Ahmed, U.; Anwar, A. Chapter 3-Application of natural dyes in dye-sensitized solar cells. In Dye-Sensitized Solar Cells; Pandey, A.K., Shahabuddin, S., Ahmad, M.S., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 45–73. [Google Scholar]
- Sharma, K.; Sharma, V.; Sharma, S.S. Dye-Sensitized Solar Cells: Fundamentals and Current Status. Nanoscale Res. Lett. 2018, 13, 1–46. [Google Scholar] [CrossRef]
- Akhtaruzzaman, M.; Shahiduzzaman, M.; Selvanathan, V.; Sopian, K.; Hossain, M.I.; Amin, N.; Hasan, A.M. Enhancing spectral response towards high-performance dye-sensitised solar cells by multiple dye approach: A comprehensive review. Appl. Mater. Today 2021, 25, 101204. [Google Scholar] [CrossRef]
- Calogero, G.; Citro, I.; Crupi, C.; Carini Jr, G.; Arigò, D.; Spinella, G.; Bartolotta, A.; Di Marco, G. Absorption spectra, thermal analysis, photoelectrochemical characterization and stability test of vegetable-based dye-sensitized solar cells. Opt. Mater. 2019, 88, 24–29. [Google Scholar] [CrossRef]
- Ayalew, W.A.; Ayele, D.W. Dye-sensitized solar cells using natural dye as light-harvesting materials extracted from Acanthus sennii chiovenda flower and Euphorbia cotinifolia leaf. J. Sci. Adv. Mater. Devices 2016, 1, 488–494. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Bin, Y.; Karthick, S.; Hemalatha, K.; Raj, C.J.; Venkatesan, S.; Park, S.; Vijayakumar, G. Natural dye extracted from Rhododendron species flowers as a photosensitizer in dye sensitized solar cell. Int. J. Electrochem. Sci. 2013, 8, 6734–6743. [Google Scholar]
- Iqbal, M.Z.; Ali, S.R.; Khan, S. Progress in dye sensitized solar cell by incorporating natural photosensitizers. Sol. Energy 2019, 181, 490–509. [Google Scholar] [CrossRef]
- Chang, H.; Lo, Y.-J. Pomegranate leaves and mulberry fruit as natural sensitizers for dye-sensitized solar cells. Sol. Energy 2010, 84, 1833–1837. [Google Scholar] [CrossRef]
- Ninis, O.; Kacimi, R.; Bouaamlat, H.; Abarkan, M.; Bouachrine, M. Theoretical studies of photovoltaic properties for design of new Azo-Pyrrole photo-sensitizer materials as dyes in solar cells. J. Mater. Environ. Sci. 2017, 8, 2572–2578. [Google Scholar]
- Xu, D.; Li, Z.; Peng, Y.-X.; Geng, J.; Qian, H.-F.; Huang, W. Post-modification of 2-formylthiophene based heterocyclic azo dyes. Dye. Pigment. 2016, 133, 143–152. [Google Scholar] [CrossRef]
- Toor, R.A.; Sayyad, M.H.; Shah, S.A.A.; Nasr, N.; Ijaz, F.; Munawar, M.A. Synthesis, computational study and characterization of a 3-{[2,3-diphenylquinoxalin-6-yl] diazenyl}-4-hydroxy-2H-chromen-2-one azo dye for dye-sensitized solar cell applications. J. Comput. Electron. 2018, 17, 821–829. [Google Scholar] [CrossRef]
- Golshan, M.; Osfouri, S.; Azin, R.; Jalali, T. Fabrication of optimized eco-friendly dye-sensitized solar cells by extracting pigments from low-cost native wild plants. J. Photochem. Photobiol. A Chem. 2019, 388, 112191. [Google Scholar] [CrossRef]
- Ekanayake, P.; Kooh, M.R.R.; Kumara, N.; Lim, A.; Petra, M.I.; Voo, N.Y.; Lim, C.M. Combined experimental and DFT–TDDFT study of photo-active constituents of Canarium odontophyllum for DSSC application. Chem. Phys. Lett. 2013, 585, 121–127. [Google Scholar] [CrossRef]
- Le Quéré, C.; Andres, R.J.; Boden, T.A.; Conway, T.; Houghton, R.A.; House, J.I.; Marland, G.; Peters, G.P.; van der Werf, G.R.; Ahlström, A. The global carbon budget 1959–2011. Earth Syst. Sci. 2013, 5, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Hsu, S.-C.; Zheng, S.; Chen, J.-H.; Li, X.I. Renewable energy microgrids: Economic evaluation and decision making for government policies to contribute to affordable and clean energy. Appl. Energy 2020, 274, 115287. [Google Scholar] [CrossRef]
- Hao, S.; Wu, J.; Huang, Y.; Lin, J. Natural dyes as photosensitizers for dye-sensitized solar cell. Sol. Energy 2006, 80, 209–214. [Google Scholar] [CrossRef]
- Wan-Mohtar, W.; Ilham, Z.; Jamaludin, A.; Rowan, N. Use of Zebrafish Embryo Assay to Evaluate Toxicity and Safety of Bioreactor-Grown Exopolysaccharides and Endopolysaccharides from European Ganoderma applanatum Mycelium for Future Aquaculture Applications. Int. J. Mol. Sci. 2021, 22, 1675. [Google Scholar] [CrossRef] [PubMed]
- Usuldin, S.R.A.; Mahmud, N.; Ilham, Z.; Ikram, N.K.K.; Ahmad, R.; Wan, W.A.A.Q.I. In-depth spectral characterization of antioxidative (1,3)-β-D-glucan from the mycelium of an identified tiger milk mushroom Lignosus rhinocerus strain ABI in a stirred-tank bioreactor. Biocatal. Agric. Biotechnol. 2020, 23, 101455. [Google Scholar] [CrossRef]
- Supramani, S.; Jailani, N.; Ramarao, K.; Zain, N.A.M.; Klaus, A.; Ahmad, R.; Wan-Mohtar, W.A.A.Q.I. Pellet diameter and morphology of European Ganoderma pfeifferi in a repeated-batch fermentation for exopolysaccharide production. Biocatal. Agric. Biotechnol. 2019, 19, 101118. [Google Scholar] [CrossRef]
- Arulraj, A.; Senguttuvan, G.; Veeramani, S.; Sivakumar, V.; Subramanian, B. Photovoltaic performance of natural metal free photo-sensitizer for TiO2 based dye-sensitized solar cells. Optik 2018, 181, 619–626. [Google Scholar] [CrossRef]
- Zalas, M.; Gierczyk, B.; Bogacki, H.; Schroeder, G. The Cortinarius Fungi Dyes as Sensitizers in Dye-Sensitized Solar Cells. Int. J. Photoenergy 2015, 2015, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Júnior, A.M.; Ribani, R.H. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef]
- Knop, D.; Yarden, O.; Hadar, Y. The ligninolytic peroxidases in the genus Pleurotus: Divergence in activities, expression, and potential applications. Appl. Microbiol. Biotechnol. 2014, 99, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Huang, C.; Zhang, J. Genome-wide functional analysis of SSR for an edible mushroom Pleurotus ostreatus. Gene 2016, 575, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, J.R.; Junior, R.N.D.C. Polysaccharides obtained from natural edible sources and their role in modulating the immune system: Biologically active potential that can be exploited against COVID-19. Trends Food Sci. Technol. 2021, 108, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Papaspyridi, L.-M.; Aligiannis, N.; Christakopoulos, P.; Skaltsounis, A.-L.; Fokialakis, N. Production of bioactive metabolites with pharmaceutical and nutraceutical interest by submerged fermentation of Pleurotus ostreatus in a batch stirred tank bioreactor. Procedia Food Sci. 2011, 1, 1746–1752. [Google Scholar] [CrossRef]
- Barbosa, J.R.; Freitas, M.M.D.S.; Martins, L.; de Carvalho, R.N. Polysaccharides of mushroom Pleurotus spp.: New extraction techniques, biological activities and development of new technologies. Carbohydr. Polym. 2019, 229, 115550. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Pan, Y.; Tang, X.; Huang, Z. Isolation and characterization of melanin from Osmanthus fragrans’ seeds. LWT 2006, 39, 496–502. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, X.; Sun, S.; Zhang, L.; Shan, S.; Zhu, H. Production of natural melanin by Auricularia auricula and study on its molecular structure. Food Chem. 2016, 190, 801–807. [Google Scholar] [CrossRef]
- Ko, K.H.; Lee, Y.C.; Jung, Y.J. Enhanced efficiency of dye-sensitized TiO2 solar cells (DSSC) by doping of metal ions. J. Colloid Interface Sci. 2005, 283, 482–487. [Google Scholar] [CrossRef]
- Ayare, N.N.; Sharma, S.; Sonigara, K.K.; Prasad, J.; Soni, S.S.; Sekar, N. Synthesis and computational study of coumarin thiophene-based D-π-A azo bridge colorants for DSSC and NLOphoric application. J. Photochem. Photobiol. A Chem. 2020, 394, 112466. [Google Scholar] [CrossRef]
- Madurai, V.; Natarajan, M.; Santhanam, A.; Asokan, V.; Velauthapillai, D. Size controlled synthesis of TiO2 nanoparticles by modified solvothermal method towards effective photo catalytic and photovoltaic applications. Mater. Res. Bull 2018, 97, 351–360. [Google Scholar]
- Balis, N.; Zaky, A.A.; Perganti, D.; Kaltzoglou, A.; Sygellou, L.; Katsaros, F.; Stergiopoulos, T.; Kontos, A.G.; Falaras, P. Dye Sensitization of Titania Compact Layer for Efficient and Stable Perovskite Solar Cells. ACS Appl. Energy Mater. 2018, 1, 6161–6171. [Google Scholar] [CrossRef]
- Amchova, P.; Kotolova, H.; Ruda-Kucerova, J. Health safety issues of synthetic food colorants. Regul. Toxicol. Pharmacol. 2015, 73, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Faustino, M.; Veiga, M.; Sousa, P.; Costa, E.M.; Silva, S.; Pintado, M. Agro-Food Byproducts as a New Source of Natural Food Additives. Molecules 2019, 24, 1056. [Google Scholar] [CrossRef] [Green Version]
- Neves, M.I.L.; Desobry-Banon, S.; Perrone, I.T.; Desobry, S.; Petit, J. Encapsulation of curcumin in milk powders by spray-drying: Physicochemistry, rehydration properties, and stability during storage. Powder Technol. 2019, 345, 601–607. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Natural food pigments and colorants. Curr. Opin. Food Sci. 2016, 7, 20–26. [Google Scholar] [CrossRef]
- Rehman, A.Q.T.; Jafari, S.M.; Assadpour, E.Q.S.; Aadil, R.M.; Iqbal, M.W.; Rashed, M.M.A.; Sajid, B.; Mushtaq, W.A. Carotenoid-loaded nanocarriers: A comprehensive review. Adv. Colloid Interface Sci. 2020, 275, 102048. [Google Scholar] [CrossRef]
- Comunian, T.A.; Silva, M.; Moraes, I.C.F.; Favaro-Trindade, C.S. Reducing carotenoid loss during storage by co-encapsulation of pequi and buriti oils in oil-in-water emulsions followed by freeze-drying: Use of heated and unheated whey protein isolates as emulsifiers. Food Res. Int. 2019, 130, 108901. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Rahman, S.U.; Bilal, M.; Iqbal, H.; Huang, D. Biosynthesis and biomedical perspectives of carotenoids with special reference to human health-related applications. Biocatal. Agric. Biotechnol. 2018, 17, 399–407. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mushroom Species | Family | Key Uses | Bioactive Pigment | Ref. |
|---|---|---|---|---|
| Agaricus bisporus | Agaricaceae | Bruising-tolerant | γ-glutaminyl-4-hydroxybenzene | [24] |
| Agrocybe cylindracea | Strophariaceae | Immunity Immunity | 6-hydroxy-1H-indole-3-carboxaldehyde, 6-hydroxy-1H-indole-3-acetamide | [10] |
| Albatrellus confluens | Albatrellaceae | Anti-cancer | albatrellin | [40] |
| Albatrellus flettii | Albatrellaceae | Anti-cancer | grifolin, neogrifolin, confluentin | [41] |
| Auricularia auricula | Auriculariaceae | Antibiofilm Antioxidant | melanin pheomelanin | [42] [43] |
| Boletus pseudocalopus | Boletaceae | Anti-cancer | grifolin derivatives 1–3 | [44] |
| Cantharelluus cibarius | Cantharellaceae | Antihypertensive | lycopene and β-carotene | [45] |
| Cerioporus squamosus | Polyporaceae | Blood regulation | melanin | [46] |
| Chalciporus piperatus | Boletaceae | Bio deterrent | chalciporone | [10] |
| Cordyceps farinosa | Cordycipitaceae | Protecion | anthraquinone derivative | [47] |
| Fomes fomentarius | Polyporaceae | Anti-tumour | melanin | [46] |
| Inonotus hispidus | Hymenochaetaceae | Antiviral | melanin | [46] |
| Inonotus obliquus | Hymenochaetaceae | Antioxidant | melanin, inotodiol | [48] |
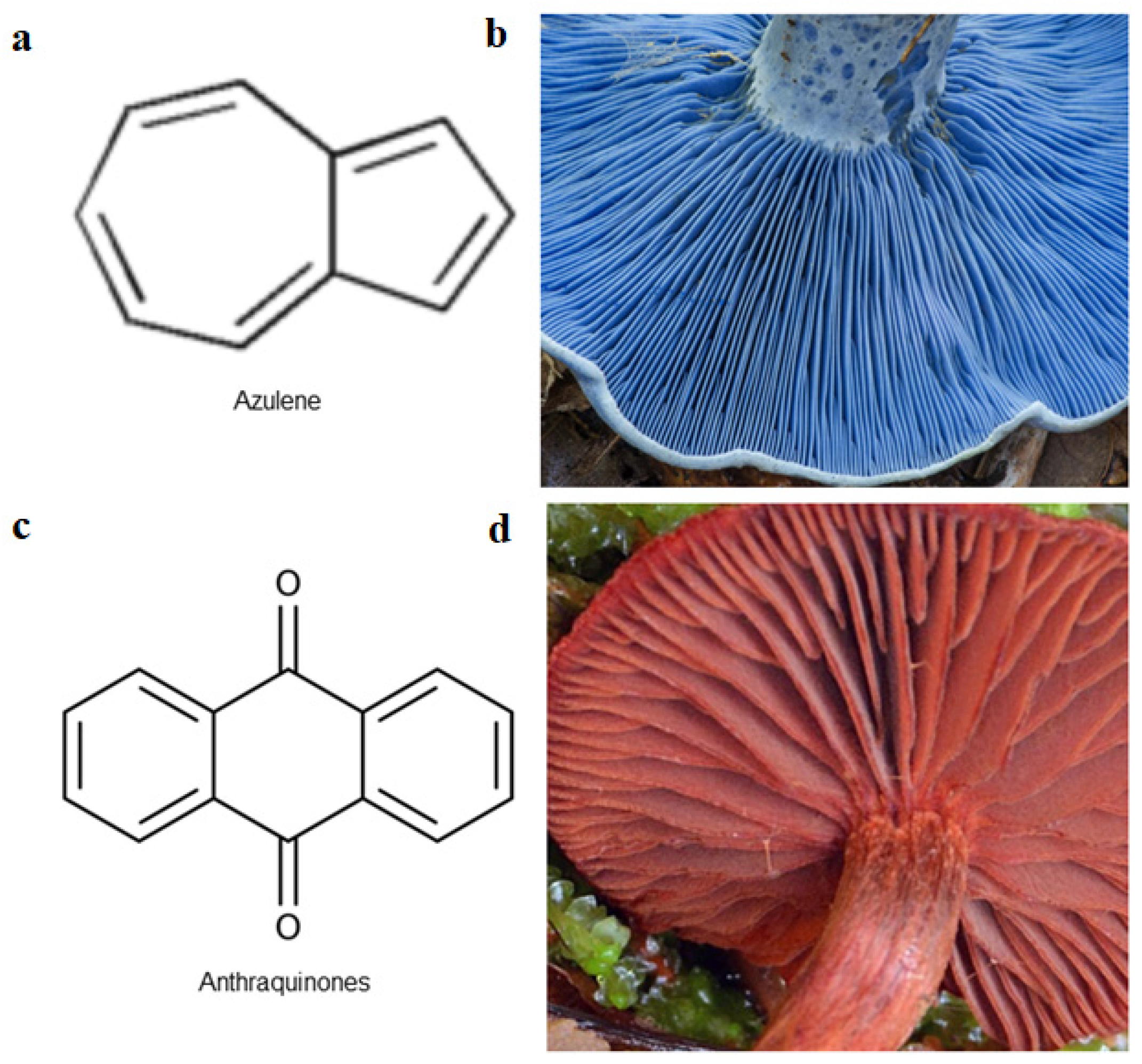
| Lactarius spp. | Russulaceae | Antimicrobial | azulenes | [49] |
| Lactarius subvellereus | Russulaceae | Anti-cancer | subvellerolactone B, subvellerolactone D, subvellerolactone E, | [50] |
| Laetiporus sulphureus | Fomitopsidaceae | Antioxidant | laetiporic acid A, 2-dehydro-3-deoxylaetiporic acid A, laetiporic acids B, laetiporic acids C | [51] |
| Lentinus brumalis | Polyporaceae | Not available | melanin | [46] |
| Ophiocordyceps unilateralis | Ophiocordycipitaceae | Antimalarial | Erythrostominone, 3,5,8–TMON, deoxyerythrostominone, deoxyerythrostominol, 4–O–methyl, erythrostominone, epierythrostominol, naphthoquinones | [49] |
| Pleurotus citrinopileatus | Pleurotaceae | Antioxidant | eumelanin, phaeomelanin | [52] |
| Pleurotus cornucopiae | Pleurotaceae | Antioxidant | eumelanin, phaeomelanin | [52] |
| Pleurotuis djamor | Pleurotaceae | Antioxidant | eumelanin, phaeomelanin | [52] |
| Pycnoporus cinnabarinus | Polyporaceae | Antibacterial | cinnabarinic acid | [10] |
| Termitomyces albuminosus | Agaricaceae | Antioxidant | melanin | [53] |
| Trametes versicolor | Polyporaceae | Breast cancer | melanin | [46] |
| Tuber melanosporum | Tuberaceae | Anti-fatigue | melanin | [54] |
| Compounds Name | Molecular Weight [g/mol] and Formula | Chemical Structure | Species |
|---|---|---|---|
| Emodin | 270.0528 C15O5H10 |  | Cortinarius sanguineus, C. semisanguineus |
| Dermocybin-1-β-D-glucopyranoside | 478.1111 C22O12H22 |  | C. vitiosus, C. sanguineus, C. semisanguineus |
| Icterinoidin B1 | 528.1056 C29H20O10 |  | C. icterinoides |
| Austrocolorin A1 | 602.2152 C34H34O10 |  | Dermocybe sp. WAT 26641 |
| Hypericin | 504.0845 C30H16O8 |  | C. austrovenetus |
| Species | Biomass Source | Technique Details | Dye Colour | Reference |
|---|---|---|---|---|
| Cordyceps militaris | Fruiting body | 0.50 g of sample mixed with 10 mL 80% ethanol | Brown | [18] |
| Lactarius lilacinus | Fruiting body | 500 g of frozen mushroom mash with MeOH (2 × 500 mL) for 30 min | Red | [19] |
| Lentinula edodes | Fruiting body | In 15 mL of 1 M NaOH, 1 g of sample is dispersed. The mixture is cooked for 3 h in water and then strained. The filtrate is acidified with 1 M HCl to a pH of 1.7 and incubated at 25 °C overnight | Brown | [25] |
| Pleurotus cystidiosus | Mycelium | Autoclaved at 120 °C for 20 min after a 1 M NaOH wash. With concentrated HCl, the alkaline pigment centrifuge-supernatant extract is acidified to pH 2 | Black | [21] |
| Armillaria cepistipes | Mycelium | A 0.45 m nitrocellulose membrane is used to filter the pH 6 sample. The filtrate is sterilised by autoclaving at 121 °C and 1 bar for 20 min. Lyophilisation is used to obtain “raw melanin” | Black | [22] |
| Pleurotus cornucopiae | Fruiting body | 1:30 of (mushroom mash/1.5 mol/L NaOH solution) | Black | [52] |
| Pleurotus citrinopileatus | Fruiting body | 1:30 of (mushroom mash/1.5 mol/L NaOH solution) | Yellow | [52] |
| Pleurotus djamor | Fruiting body | 1:30 of (mushroom mash/1.5 mol/L NaOH solution) | Pink | [52] |
| Species | Melanin Extract | DSSC Status | References |
|---|---|---|---|
| Agaricus bisporus | (White-brown) Riboflavin | TiO2 sintered at 500 °C for 30 min | [116] |
| Cortinarius sp. | Brown | TiO2 sintered at 500 °C for 30 min | [117] |
| Pleurotus cornucopiae CCMSSC 00406 | Black | Untested | [52] |
| Pleurotus citrinopileatus CCMSSC 04208 | Yellow | Untested | [52] |
| Pleurotus djamor CCMSSC 00450 | Pink | Untested | [52] |
| Lentinula edodes | Brown | Untested | [25] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, N.; Vunduk, J.; Klaus, A.; Dahlan, N.Y.; Ghosh, S.; Muhammad-Sukki, F.; Dufossé, L.; Bani, N.A.; Wan-Mohtar, W.A.A.Q.I. Roles of Medicinal Mushrooms as Natural Food Dyes and Dye-Sensitised Solar Cells (DSSC): Synergy of Zero Hunger and Affordable Energy for Sustainable Development. Sustainability 2022, 14, 13894. https://doi.org/10.3390/su142113894
Ahmad N, Vunduk J, Klaus A, Dahlan NY, Ghosh S, Muhammad-Sukki F, Dufossé L, Bani NA, Wan-Mohtar WAAQI. Roles of Medicinal Mushrooms as Natural Food Dyes and Dye-Sensitised Solar Cells (DSSC): Synergy of Zero Hunger and Affordable Energy for Sustainable Development. Sustainability. 2022; 14(21):13894. https://doi.org/10.3390/su142113894
Chicago/Turabian StyleAhmad, Nurfadzilah, Jovana Vunduk, Anita Klaus, Nofri Yenita Dahlan, Soumya Ghosh, Firdaus Muhammad-Sukki, Laurent Dufossé, Nurul Aini Bani, and Wan Abd Al Qadr Imad Wan-Mohtar. 2022. "Roles of Medicinal Mushrooms as Natural Food Dyes and Dye-Sensitised Solar Cells (DSSC): Synergy of Zero Hunger and Affordable Energy for Sustainable Development" Sustainability 14, no. 21: 13894. https://doi.org/10.3390/su142113894