Comparative Analysis of Secondary Metabolites Produced by Ascochyta fabae under In Vitro Conditions and Their Phytotoxicity on the Primary Host, Vicia faba, and Related Legume Crops

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
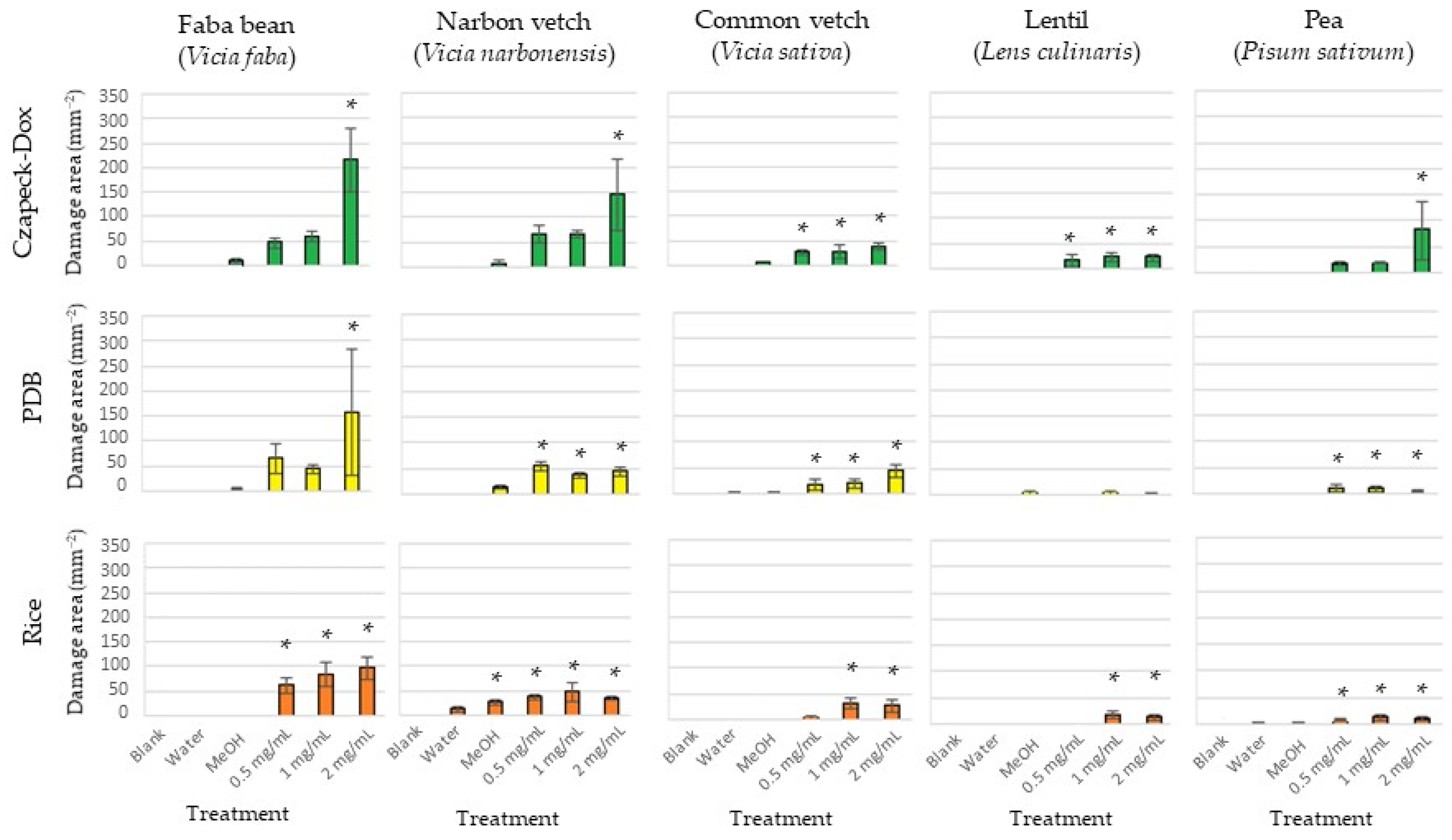
2.1. Bioassays of Fungal Organic Extract
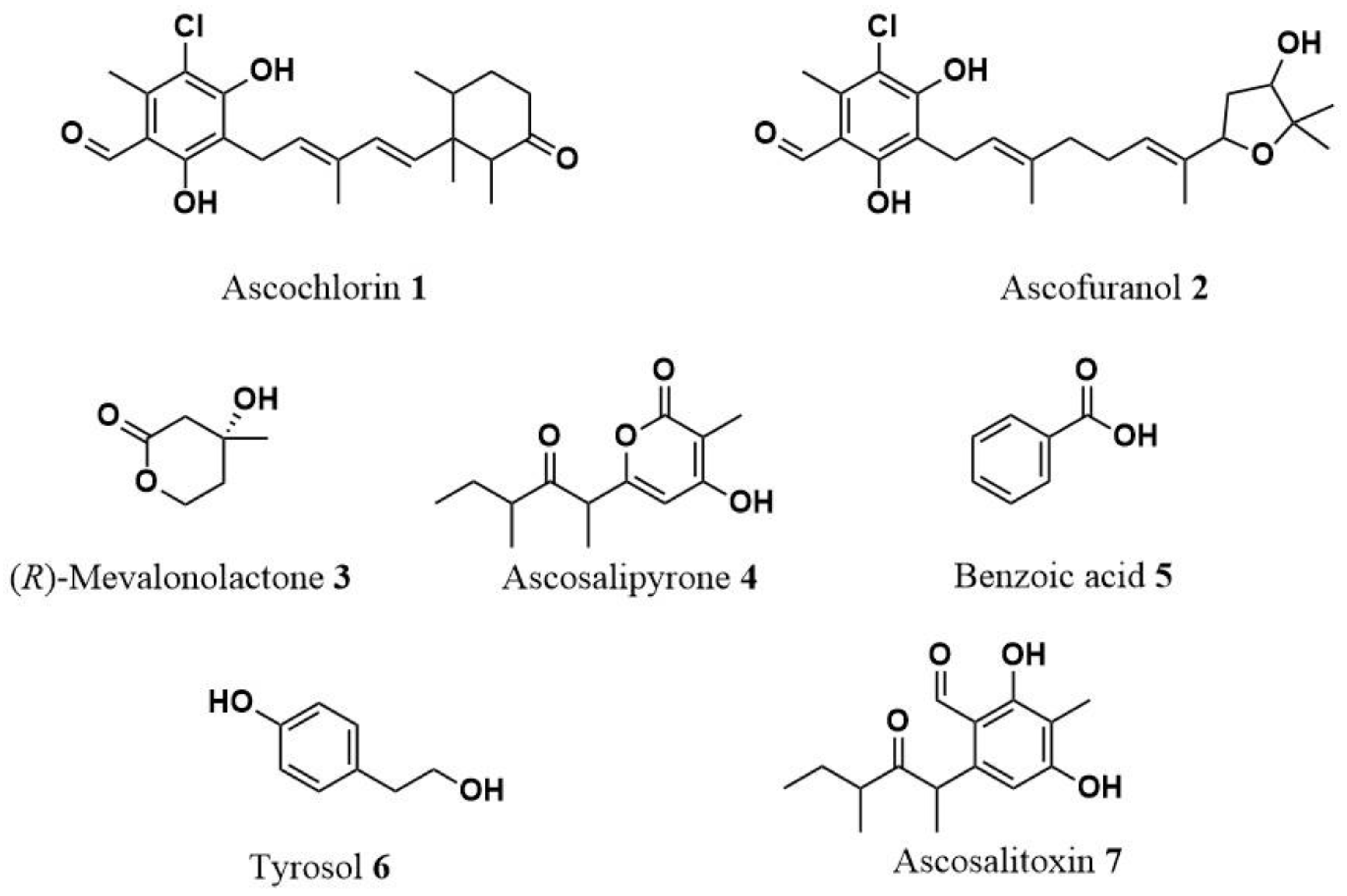
2.2. Identification of Secondary Metabolites from Culture Filtrates of A. fabae Cultures
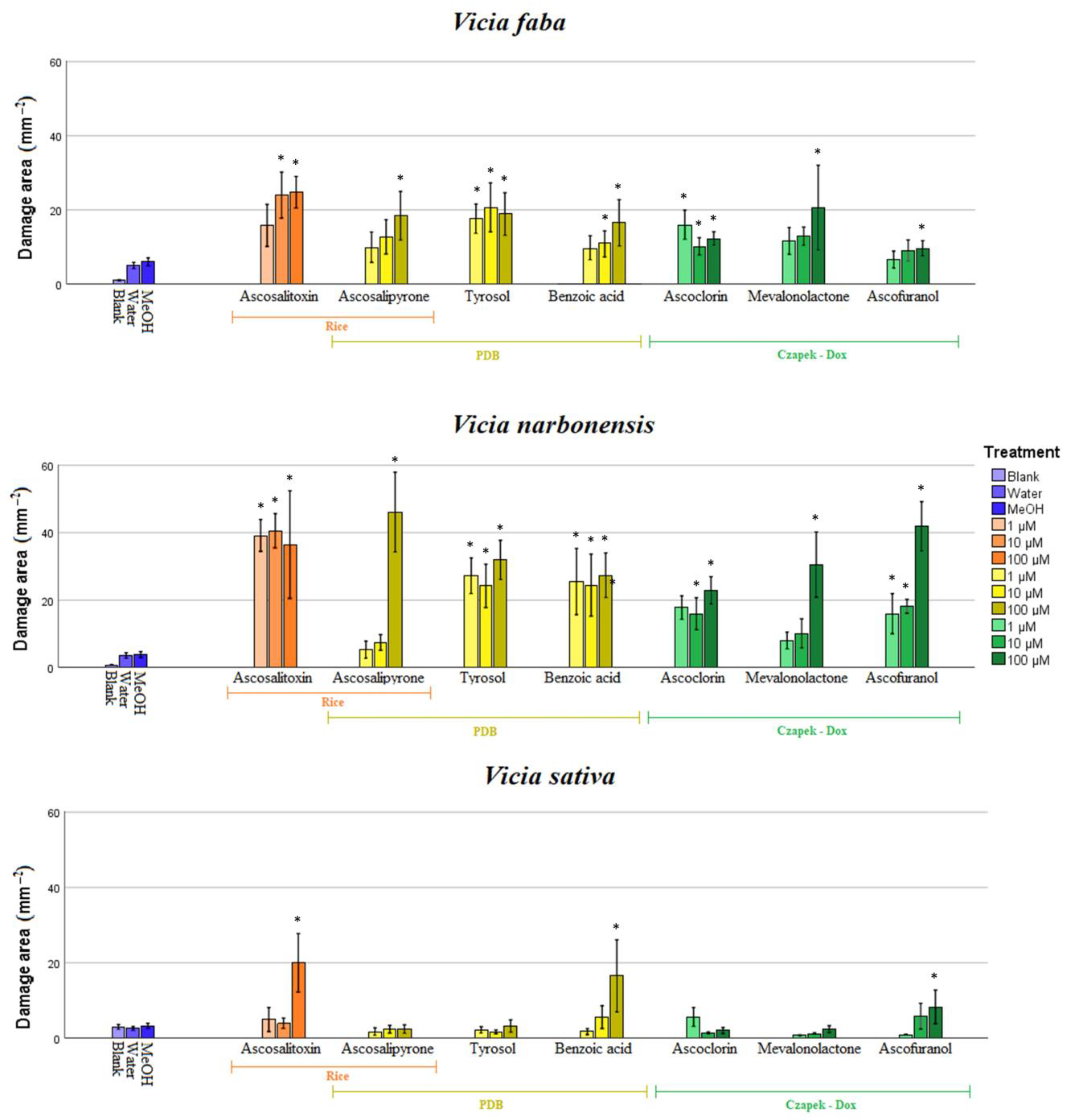
2.3. Bioassay of Pure Compounds
3. Discussion
4. Materials and Methods
4.1. Fungal Strain, Culture Medium and Growth Conditions
4.2. General Experimental Procedure for Chemical Analysis
4.3. Extraction and Purification of Secondary Metabolites Produced in Czapek-Dox Culture
4.4. Extraction and Purification of Secondary Metabolites Produced in PDB Culture
4.5. Extraction and Purification of Secondary Metabolites Produced in Rice Culture
4.6. Bioassays
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rubiales, D.; Annicchiarico, P.; Vaz Patto, M.C.; Julier, B. Legume breeding for the agroecological transition of global agri-food systems: A European perspective. Front. Plant Sci. 2021, 12, 782574. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, H.; Pinto, E.; Vasconcelos, M.W. Legumes as a cornerstone of the transition toward more sustainable agri-food systems and diets in Europe. Front. Sustain. Food Syst. 2021, 5, 694121. [Google Scholar] [CrossRef]
- Rubiales, D.; Duc, G.; Stoddard, F. Faba beans in sustainable agriculture. Field Crops Res. 2010, 115, 201–233. [Google Scholar] [CrossRef]
- Gharzeddin, K.; Maalouf, F.; Khoury, B.; Abou Khater, L.; Christmann, S.; Jamal El Dine, N.A. Efficiency of different breeding strategies in improving the faba bean productivity for sustainable agriculture. Euphytica 2019, 215, 203. [Google Scholar] [CrossRef]
- Yitayih, G.; Fininsa, C.; Terefe, H.; Shibabaw, A. Integrated management approaches reduced yield loss, and increased productivity in faba bean, due to gall disease in northwestern Ethiopia. Arch. Phytopathol. Plant Prot. 2022, 55, 1592–1610. [Google Scholar] [CrossRef]
- Karkanis, A.; Ntatsi, G.; Lepse, L.; Fernández, J.A.; Vågen, I.M.; Rewald, B.; Alsina, I.; Kronberga, A.; Balliu, A.; Olle, M.; et al. Faba bean cultivation-revealing novel managing practices for more sustainable and competitive European cropping systems. Front. Plant Sci. 2018, 9, 1115. [Google Scholar] [CrossRef]
- Tivoli, B.; Banniza, S. Comparison of the Epidemiology of Ascochyta Blights on Grain Legumes; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Hernandez-Bello, M.; Chilvers, M.; Akamatsu, H.; Peever, T. Host specificity of Ascochyta spp. infecting legumes of the Viciae and Cicerae tribes and pathogenicity of an interspecific hybrid. Phytopathology 2006, 96, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, W.; Sankaran, S. High-throughput field phenotyping of Ascochyta blight disease severity in chickpea. Crop Prot. 2019, 125, 104885. [Google Scholar] [CrossRef]
- Bretag, T.W.; Keane, P.J.; Price, T.V. The epidemiology and control of ascochyta blight in field peas: A review. Aust. J. Agric. Res. 2006, 57, 883–902. [Google Scholar] [CrossRef]
- Barilli, E.; Cobos, M.J.; Rubiales, D. Clarification on host range of Didymella pinodes the causal agent of pea Ascochyta blight. Front. Plant Sci. 2016, 7, 592. [Google Scholar] [CrossRef]
- Ahmed, S.; Abang, M.; Maalouf, F. Integrated management of Ascochyta blight (Didymella fabae) on faba bean under Mediterranean conditions. Crop Prot. 2016, 81, 65–69. [Google Scholar] [CrossRef]
- Koder, S.B.; Nawale, R.; Katyayani, K.K.S.; Rana, M.; Srivastava, S. Symptoms, biology and management of ascochyta blight (Phoma exigua) of French beans: A review. Agric. Sci. Dig. Res. J. 2022, 42, 657–664. [Google Scholar] [CrossRef]
- Trapero-Casas, A.; Kaiser, W.J. Influence of temperature, wetness period, plant age, and inoculum concentration on infection and development of Ascochyta blight of chickpea. Phytopathology 1992, 82, 589–596. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef]
- Pratap, A.; Douglas, C.; Prajapati, U.; Kumari, G.; War, A.R.; Tomar, R.; Pandey, A.K.; Dubey, S. Breeding progress and future challenges: Biotic stresses. In The Mungbean Genome. Compendium of Plant Genomes; Springer: Cham, Switzerland, 2020; pp. 55–80. [Google Scholar]
- Rubiales, D.; Ávila, C.M.; Sillero, J.C.; Hybl, M.; Narits, L.; Sass, O.; Flores, F. Identification and multi-environment validation of resistance to Ascochyta fabae in faba bean (Vicia faba). Field Crops Res. 2012, 126, 165–170. [Google Scholar] [CrossRef]
- Mengiste, T. Plant immunity to necrotrophs. Annu. Rev. Phytopathol. 2012, 50, 267–294. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Chen, W. Phytotoxic metabolites produced by legume-associated Ascochyta and its related genera in the Dothideomycetes. Toxins 2019, 11, 627. [Google Scholar] [CrossRef] [PubMed]
- Oku, H.; Nakanishi, T. A toxic metabolite from Ascochyta fabae having antibiotic activity. Phytopathology 1963, 53, 1321–1325. [Google Scholar]
- Kim, W.; Lichtenzveig, J.; Syme, R.A.; Williams, A.H.; Peever, T.L.; Chen, W. Identification of a polyketide synthase gene responsible for Ascochitine biosynthesis in Ascochyta fabae and its abrogation in sister taxa. Msphere 2019, 4, e00622-19. [Google Scholar] [CrossRef]
- Agudo-Jurado, F.J.; Reveglia, P.; Rubiales, D.; Evidente, A.; Barilli, E. Status of Phytotoxins Isolated from Necrotrophic Fungi Causing Diseases on Grain Legumes. Int. J. Mol. Sci. 2023, 24, 5116. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Vurro, M.; Bottalico, A. Ascosalitoxin, a phytotoxic trisubstituted salicylic aldehyde from Ascochyta pisi. Phytochemistry 1993, 34, 995–998. [Google Scholar] [CrossRef]
- Beed, F.; Strange, R.; Onfroy, C.; Tivoli, B. Virulence for faba bean and production of ascochitine by Ascochyta fabae. Plant Pathol. 1994, 43, 987–997. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Farinella, V.F.; Kawafune, E.S.; Tangerina, M.M.; Domingos, H.V.; Costa-Lotufo, L.V.; Ferreira, M.J. OSMAC strategy integrated with molecular networking for accessing griseofulvin derivatives from endophytic fungi of Moquiniastrum polymorphum (Asteraceae). Molecules 2021, 26, 7316. [Google Scholar] [CrossRef] [PubMed]
- Tamura, G.; Suzuki, S.; Takatsuki, A.; Ando, K.; Arima, K. Ascochlorin, a new antibiotic, found by paper-disc agar-diffusion method. I isolation, biological and chemical properties of ascochlorin (studies on antiviral and antitumor antibiotics. I). J. Antibiot. 1968, 21, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Seephonkai, P.; Isaka, M.; Kittakoop, P.; Boonudomlap, U.; Thebtaranonth, Y. A novel ascochlorin glycoside from the insect pathogenic fungus Verticillium hemipterigenum BCC 2370. J. Antibiot. 2004, 57, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Mogi, T.; Miyoshi, H. Properties of cytochrome bd plastoquinol oxidase from the cyanobacterium Synechocystis sp. PCC 6803. J. Biochem. 2009, 145, 395–401. [Google Scholar] [CrossRef]
- Berry, E.A.; Huang, L.S.; Lee, D.W.; Daldal, F.; Nagai, K.; Minagawa, N. Ascochlorin is a novel, specific inhibitor of the mitochondrial cytochrome bc1 complex. Biochim. Biophys. Acta BBA Bioenerg. 2010, 1797, 360–370. [Google Scholar] [CrossRef]
- Jeong, J.H.; Jeong, Y.J.; Cho, H.J.; Shin, J.M.; Kang, J.H.; Park, K.K.; Park, Y.Y.; Chung, I.K.; Chang, H.W.; Magae, J. Ascochlorin inhibits growth factor-induced HIF-1α activation and tumor-angiogenesis through the suppression of EGFR/ERK/p70S6K signaling pathway in human cervical carcinoma cells. J. Cell. Biochem. 2012, 113, 1302–1313. [Google Scholar] [CrossRef]
- Sasaki, H.; Hosokawa, T.; Sawada, M.; Ando, K. Isolation and structure of ascofuranone and ascofranol, antibiotics with hypolipidemic activity. J. Antibiot. 1973, 26, 676–680. [Google Scholar] [CrossRef]
- Hamid, K. Separation and Phytotoxicity of Solanapyrone Compounds Produced by Ascochyta rabiei (Pass.) Labr. and Their Metabolism by Chickpea (Cicer arietinum L.); University of London, University College London (United Kingdom): London, UK, 1999. [Google Scholar]
- Leyte-Lugo, M.; González-Andrade, M.; González, M.d.C.; Glenn, A.E.; Cerda-García-Rojas, C.M.; Mata, R. (+)-Ascosalitoxin and vermelhotin, a calmodulin inhibitor, from an endophytic fungus isolated from Hintonia latiflora. J. Nat. Prod. 2012, 75, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Nian, Y.; Chen, H.; Liang, S.; Sun, M.; Pei, Y.; Wang, H. Pyranone Derivatives with antitumor activities, from the endophytic fungus Phoma sp. YN02-P-3. Front. Chem. 2022, 10, 950726. [Google Scholar] [CrossRef] [PubMed]
- Osterhage, C.; Kaminsky, R.; König, G.M.; Wright, A.D. Ascosalipyrrolidinone a, an antimicrobial alkaloid, from the obligate marine fungus Ascochyta salicorniae. J. Org. Chem. 2000, 65, 6412–6417. [Google Scholar] [CrossRef] [PubMed]
- Seibert, S.F.; Eguereva, E.; Krick, A.; Kehraus, S.; Voloshina, E.; Raabe, G.; Fleischhauer, J.; Leistner, E.; Wiese, M.; Prinz, H. Polyketides from the marine-derived fungus Ascochyta salicorniae and their potential to inhibit protein phosphatases. Org. Biomol. Chem. 2006, 4, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, M.M.; Alves, A.; Andolfi, A. Secondary metabolites of Lasiodiplodia theobromae: Distribution, chemical diversity, bioactivity, and implications of their occurrence. Toxins 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Fujita, M.; Feng, Q.; Clardy, J.; Fink, G.R. Tyrosol is a quorum-sensing molecule in Candida albicans. Proc. Natl. Acad. Sci. USA 2004, 101, 5048–5052. [Google Scholar] [CrossRef] [PubMed]
- Muriana, F.J.; Montserrat-de la Paz, S.; Lucas, R.; Bermudez, B.; Jaramillo, S.; Morales, J.C.; Abia, R.; Lopez, S. Tyrosol and its metabolites as antioxidative and anti-inflammatory molecules in human endothelial cells. Food Funct. 2017, 8, 2905–2914. [Google Scholar] [CrossRef]
- Loru, D.; Incani, A.; Deiana, M.; Corona, G.; Atzeri, A.; Melis, M.; Rosa, A.; Dessì, M. Protective effect of hydroxytyrosol and tyrosol against oxidative stress in kidney cells. Toxicol. Ind. Health 2009, 25, 301–310. [Google Scholar] [CrossRef]
- Karković Marković, A.; Torić, J.; Barbarić, M.; Jakobušić Brala, C. Hydroxytyrosol, tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef]
- Bu, Y.; Rho, S.; Kim, J.; Kim, M.Y.; Lee, D.H.; Kim, S.Y.; Choi, H.; Kim, H. Neuroprotective effect of tyrosol on transient focal cerebral ischemia in rats. Neurosci. Lett. 2007, 414, 218–221. [Google Scholar] [CrossRef]
- Ohtsuki, T.; Sato, K.; Sugimoto, N.; Akiyama, H.; Kawamura, Y. Absolute quantification for benzoic acid in processed foods using quantitative proton nuclear magnetic resonance spectroscopy. Talanta 2012, 99, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Uyeda, M.; Kido, Y.; Toya, N.; Nakashima, R.; Terazumi, R. A new antibiotic K-82 A and minor components, produced by Streptomyces lavendulae, strain no. K-82. J. Antibiot. 1980, 33, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Guo, Y.; Li, Y.; Lv, J.; Dong, K.; Dong, Y. Benzoic Acid Phytotoxicity the Structural Resistance and Photosynthetic Physiology of Faba Bean and Promotes Fusarium Wilt Incidence. SSRN Electron. J. 2022. [Google Scholar] [CrossRef]
- Zhang, K.; Sun, P.; Khan, A.; Zhang, Y. Photochemistry of biochar during ageing process: Reactive oxygen species generation and benzoic acid degradation. Sci. Total Environ. 2021, 765, 144630. [Google Scholar] [CrossRef] [PubMed]
- Buhaescu, I.; Izzedine, H. Mevalonate pathway: A review of clinical and therapeutical implications. Clin. Biochem. 2007, 40, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Lasunción, M.A.; Martinez-Botas, J.; Martin-Sanchez, C.; Busto, R.; Gómez-Coronado, D. Cell cycle dependence on the mevalonate pathway: Role of cholesterol and non-sterol isoprenoids. Biochem. Pharm. 2022, 196, 114623. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Nocera, P.; Boari, A.; Zonno, M.C.; Pescitelli, G.; Sarrocco, S.; Baroncelli, R.; Vannacci, G.; Vurro, M.; Evidente, A. Secondary metabolites produced by Colletotrichum lupini, the causal agent of anthachnose of lupin (Lupinus spp.). Mycologia 2020, 112, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Khamthong, N.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. An antibacterial cytochalasin derivative from the marine-derived fungus Diaporthaceae sp. PSU-SP2/4. Phytochem. Lett. 2014, 10, 5–9. [Google Scholar] [CrossRef]
- Hemtasin, C.; Kanokmedhakul, S.; Kanokmedhakul, K.; Hahnvajanawong, C.; Soytong, K.; Prabpai, S.; Kongsaeree, P. Cytotoxic pentacyclic and tetracyclic aromatic sesquiterpenes from Phomopsis archeri. J. Nat. Prod. 2011, 74, 609–613. [Google Scholar] [CrossRef]
- Varejão, E.V.V.; Demuner, A.J.; Barbosa, L.C.d.A.; Barreto, R.W. Phytotoxic effects of metabolites from Alternaria euphorbiicola against its host plant Euphorbia heterophylla. Quím. Nova 2013, 36, 1004–1007. [Google Scholar] [CrossRef]
- Junio, H.A.; Sy-Cordero, A.A.; Ettefagh, K.A.; Burns, J.T.; Micko, K.T.; Graf, T.N.; Richter, S.J.; Cannon, R.E.; Oberlies, N.H.; Cech, N.B. Synergy-directed fractionation of botanical medicines: A case study with goldenseal (Hydrastis canadensis). J. Nat. Prod. 2011, 74, 1621–1629. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed]
- Gauglitz, J.M.; West, K.A.; Bittremieux, W.; Williams, C.L.; Weldon, K.C.; Panitchpakdi, M.; Di Ottavio, F.; Aceves, C.M.; Brown, E.; Sikora, N.C. Enhancing untargeted metabolomics using metadata-based source annotation. Nat. Biotechnol. 2022, 40, 1774–1779. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fiehn, O. Flash entropy search to query all mass spectral libraries in real time. Nat. Methods 2023, 20, 1475–1478. [Google Scholar] [CrossRef]
- Burns, D.C.; Mazzola, E.P.; Reynolds, W.F. The role of computer-assisted structure elucidation (CASE) programs in the structure elucidation of complex natural products. Nat. Prod. Rep. 2019, 36, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Marcarino, M.O.; Zanardi, M.M.; Cicetti, S.; Sarotti, A.M. NMR calculations with quantum methods: Development of new tools for structural elucidation and beyond. Acc. Chem. Res. 2020, 53, 1922–1932. [Google Scholar] [CrossRef]
- Beniddir, M.A.; Kang, K.B.; Genta-Jouve, G.; Huber, F.; Rogers, S.; Van Der Hooft, J.J. Advances in decomposing complex metabolite mixtures using substructure-and network-based computational metabolomics approaches. Nat. Prod. Rep. 2021, 38, 1967–1993. [Google Scholar] [CrossRef]
- Mori, K.; Takechi, S. Synthesis of the natural enantiomers of ascochlorin, ascofuranone and ascofuranol. Tetrahedron 1985, 41, 3049–3062. [Google Scholar] [CrossRef]
- Menna, M.; Imperatore, C.; Mangoni, A.; Della Sala, G.; Taglialatela-Scafati, O. Challenges in the configuration assignment of natural products. A case-selective perspective. Nat. Prod. Rep. 2019, 36, 476–489. [Google Scholar] [CrossRef]
- Bitchagno, G.T.M.; Nchiozem-Ngnitedem, V.-A.; Melchert, D.; Fobofou, S.A. Demystifying racemic natural products in the homochiral world. Nat. Rev. Chem. 2022, 6, 806–822. [Google Scholar] [CrossRef]
- Pereda-Miranda, R.; Bautista, E.; Martínez-Fructuoso, L.; Fragoso-Serrano, M. From Relative to Absolute Stereochemistry of Secondary Metabolites: Applications in Plant Chemistry. Rev. Bras. Farmacogn. 2023, 33, 1–48. [Google Scholar] [CrossRef]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Singh, H.; Bhardwaj, N.; Bhardwaj, S.K.; Khatri, M.; Kim, K.H.; Peng, W. An exploration on the toxicity mechanisms of phytotoxins and their potential utilities. Crit. Rev. Environ. Sci. Technol. 2022, 52, 395–435. [Google Scholar] [CrossRef]
- Guo, J.; Cheng, Y. Advances in fungal elicitor-triggered plant immunity. Int. J. Mol. Sci. 2022, 23, 12003. [Google Scholar] [CrossRef] [PubMed]
- Sillero, J.C.; Rojas-Molina, M.M.; Ávila, C.M.; Rubiales, D. Induction of systemic acquired resistance against rust, ascochyta blight and broomrape in faba bean by exogenous application of salicylic acid and benzothiadiazole. Crop Prot. 2012, 34, 65–69. [Google Scholar] [CrossRef]
- Gulder, T.A.; Hong, H.; Correa, J.; Egereva, E.; Wiese, J.; Imhoff, J.F.; Gross, H. Isolation, structure elucidation and total synthesis of lajollamide A from the marine fungus Asteromyces cruciatus. Mar. Drugs. 2012, 10, 2912–2935. [Google Scholar] [CrossRef] [PubMed]
- Reveglia, P.; Agudo-Jurado, F.J.; Barilli, E.; Masi, M.; Evidente, A.; Rubiales, D. Uncovering phytotoxic compounds produced by Colletotrichum spp. involved in legume diseases using an OSMAC–metabolomics approach. J. Fungi 2023, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hosokawa, T.; Nawata, Y.; Ando, K. Isolation and structure of ascochlorin and its analogs. Agric. Biol. Chem. 1974, 38, 1463–1466. [Google Scholar] [CrossRef]
- Kimura, Y.; Tamura, S. Isolation of L-β-phenyllactic acid and tyrosol as plant growth regulators from Gloeosporium laeticolor. Agric. Biol. Chem. 1973, 37, 2925. [Google Scholar] [CrossRef]
- Sillero, J.; Fondevilla, S.; Davidson, J.; Patto, M.V.; Warkentin, T.; Thomas, J.; Rubiales, D. Screening techniques and sources of resistance to rusts and mildews in grain legumes. Euphytica 2006, 147, 255–272. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plant | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | Faba Bean | Narbon Vetch | Common Vetch | Lentil | Pea | |||||
| Blank | 0 ± 0 | d | 0 ± 0 | c | 0 ± 0 | c | 0 ± 0 | c | 0 ± 0 | c |
| Water | 0 ± 0 | d | 1.8 ± 1.6 | c | 0.2 ± 0.1 | c | 0 ± 0 | c | 0.3 ± 0.2 | c |
| MeOH | 3.4 ± 1.5 | c | 7.3 ± 3.3 | bc | 0.2 ± 0.05 | c | 0.5 ± 0.3 | c | 0.4 ± 0.2 | c |
| Czapek-Dox | 110.6 ± 3.9 | a | 96.4 ± 8.8 | a | 27.9 ± 5.5 | ab | 19.9 ± 5.2 | a | 29.2 ± 3.2 | a |
| PDB | 95.6 ± 5.4 | b | 45.2 ± 4.7 | b | 30.2 ± 7.3 | a | 1.0 ± 0.6 | c | 7.5 ± 2.6 | b |
| Rice | 81.6 ± 2.6 | b | 38.9 ± 5.1 | b | 17.1 ± 5.1 | b | 9.1 ± 3.4 | b | 9.9 ± 2.1 | b |
| Legume | Plant Specie | Genotype |
|---|---|---|
| Faba bean | Vicia faba | Baraca |
| Narbon vetch | V. narbonensis | VN01 |
| Common vetch | V. sativa | Buzza |
| Lentil | Lens culinaris | Pardina |
| Pea | Pisum sativum | Messire |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barilli, E.; Reveglia, P.; Agudo-Jurado, F.J.; Cañete García, V.; Cimmino, A.; Evidente, A.; Rubiales, D. Comparative Analysis of Secondary Metabolites Produced by Ascochyta fabae under In Vitro Conditions and Their Phytotoxicity on the Primary Host, Vicia faba, and Related Legume Crops. Toxins 2023, 15, 693. https://doi.org/10.3390/toxins15120693
Barilli E, Reveglia P, Agudo-Jurado FJ, Cañete García V, Cimmino A, Evidente A, Rubiales D. Comparative Analysis of Secondary Metabolites Produced by Ascochyta fabae under In Vitro Conditions and Their Phytotoxicity on the Primary Host, Vicia faba, and Related Legume Crops. Toxins. 2023; 15(12):693. https://doi.org/10.3390/toxins15120693
Chicago/Turabian StyleBarilli, Eleonora, Pierluigi Reveglia, Francisco J. Agudo-Jurado, Vanessa Cañete García, Alessio Cimmino, Antonio Evidente, and Diego Rubiales. 2023. "Comparative Analysis of Secondary Metabolites Produced by Ascochyta fabae under In Vitro Conditions and Their Phytotoxicity on the Primary Host, Vicia faba, and Related Legume Crops" Toxins 15, no. 12: 693. https://doi.org/10.3390/toxins15120693