

New Inonotus Polysaccharides: Characterization and Anticomplementary Activity of Inonotus rheades Mycelium Polymers


1
Laboratory of Biomedical Research, Institute of General and Experimental Biology, Siberian Division, Russian Academy of Science, Sakh’yanovoy Street 6, 670047 Ulan-Ude, Russia
2
Laboratory of Plant Physiological Genetics, Siberian Institute of Plant Physiology and Biochemistry, Siberian Division, Russian Academy of Science, Lermontova Street 132, 664033 Irkutsk, Russia
*
Author to whom correspondence should be addressed.
Polymers 2023, 15(5), 1257; https://doi.org/10.3390/polym15051257
Submission received: 28 January 2023
/
Revised: 27 February 2023
/
Accepted: 28 February 2023
/
Published: 1 March 2023
(This article belongs to the Special Issue Applications of Polysaccharide-Based Materials)
Abstract
:Inonotus is a small genus of xylotrophic basidiomycetes and a source of bioactive fungochemicals among which a special place is occupied by polymeric compounds. In this study, polysaccharides that are widespread in Europe, Asia, and North America and a poorly understood fungal species, I. rheades (Pers.) Karst. (fox polypore), were investigated. Water-soluble polysaccharides of I. rheades mycelium were extracted, purified, and studied using chemical reactions, elemental and monosaccharide analysis, UV–Vis and FTIR spectroscopy, gel permeation chromatography, and linkage analysis. Five homogenic polymers (IRP-1–IRP-5) with molecular weights of 110–1520 kDa were heteropolysaccharides that consist mainly of galactose, glucose, and mannose. The dominant component, IRP-4, was preliminary concluded to be a branched (1→3,6)-linked galactan. Polysaccharides of I. rheades inhibited the hemolysis of sensitized sheep erythrocytes by complement from human serum, signifying anticomplementary activity with the greatest effects for the IRP-4 polymer. These findings suggest that I. rheades mycelium is a new source of fungal polysaccharides with potential immunomodulatory and anti-inflammatory properties.
1. Introduction
Xylotrophic basidiomycetes are a large ecological group of saprotrophic fungi that are often found on wood [1]. The ability to accumulate lignocelluloses is a special feature of xylotrophic basidiomycetes, which include fungi of the genus Inonotus consisting of approximately 20 species included in the Basidiomycota division of the Aphyllophorales order and the Hymenochaetaceae family [2]. Currently, basidiomycetes are being investigated as a source of various biologically active compounds. The most promising and widely significant in fungotherapy is the natural sclerotia of Inonotus obliquus (Ach. ex Pers.) Pilat or Chaga, which is formed mainly on living birch trunks [3]. Various biological activities of the extracts and compounds from I. obliquus have been revealed [4]. Based on Chaga, a number of mushroom-derived medicines have been developed [5]. However, the natural resource of I. obliquus is limited, and a wide geographical distribution and various natural conditions have resulted in difficulties in standardizing the raw materials. Many species of the genus Inonotus have still been poorly investigated.
Among the poorly studied species is I. rheades (Pers.) Karst. (fox polypore), a circumboreal species distributed in Europe from north to south, as well as in Asia and North America, and growing more often on species of Populus tremula (in Europe) and rarely on Quercus, Salix, and Sorbus (Figure 1) [6]. This annual species has a pileus, is solitary or grows in groups of several pilei, is medium in size, is flat or triangular, and has a thick base, with a uniformly colored upper surface. The old base is rough brown, and the young outer zones are a hairy yellow-brown. The upper surface is dark, and the actively growing edge is pale, soft, and matte. The pores are 2–3 × 1 mm in size and angular or tortuous in shape, with a brown surface. The hyphal system monomitic hyphae are without clamps and are brown without setae. Basidiospores are egg-shaped, in the form of an ellipsoid of 5–6 × 3.5–4 µm [6]. As a rule, this fungal species is considered inedible, although it is not poisonous.
The diverse range of chemical components found in I. rheades includes styrylpyrones and bis(styrylpyrones), in particular 1,1-distyrylpyrylethane, trans- and cis-hispidin, trans- and cis-bisnoryangonin, phellinins A1 and A2, hypholomine A, 3-bisnoryangonyl-14′-hispidin, hypholomine B, 3,14′-bishispidinyl, rheadinin [7,8], triterpenes as inotodiol, lanosterol, ergosterol peroxide, lupeol, betulin, betulinic acid, betulone, betulonic aldehyde, betulonic acid, and betulinic aldehyde [9], and fatty acids [10] detected in the mycelium. Regarding bioactivity, the extracts of I. rheades and pure compounds possess antiglucosidase [9], antioxidant [11], antibacterial, and fungistatic activity [12].
Among the variety of Inonotus constituents, polysaccharides are perhaps the most relevant as bioactive compounds of this fungal genus [13]. Structurally, Inonotus polysaccharides are homo- and heteropolymers with core chains of α-glucans [14], (1→ 3)- and (1→6)-linked β-glucans [15,16], (1→6)-linked α-galactans, (1 →4)-linked β-xylans [17], and mannans [18]. Various biological studies of Inonotus polysaccharides have demonstrated their effectiveness as antitumor [19,20,21], lipid-lowering [22], hypoglycemic [23,24,25], immunomodulatory [26], antioxidant [27], anti-fatigue [28], antiviral [29], gut microbiota-regulated [30], and anti-Toxoplasma agents [31]. The polysaccharides of I. rheades mycelium have not been previously investigated.
Both the fruiting bodies and mycelium are of practical importance in fungi. The use of pure mycelial mass is relevant for several reasons. First, there is no seasonal dependence on obtaining raw materials compared to fruiting bodies that are collected in nature. It is known that the fruiting period of mushrooms is very limited (1–3 weeks), and some species cannot bear fruit every year [5]. Second, there are no quantitative restrictions for obtaining mycelial mass, which may be associated with the individual characteristics of the fruiting of mushrooms, as well as with changes in climatic and weather conditions. Given the physiological characteristics of fungi and their distribution area, it should be remembered that some fungi do not form a sufficient number of fruiting bodies and may be rare [32]. In nature, the development of a fungal organism also directly depends on climatic factors; for example, during periods of drought, the development of the mycelium slows down, and the formation of primordia and fruiting bodies does not occur [33]. Third, the resulting mycelial mass is microbiologically pure and standardized in quality. Under natural conditions, any organism is subject to contamination, and fungi are no exception. With the multiple actions of various environmental factors, raw materials collected in natural conditions have variations in chemical composition [34]. Fourth, it is more convenient to regulate properties of a mycelium. Using ecological parameters for growing a mycelium, it is possible to regulate its chemical composition; for example, lighting parameters have a direct impact on the morphology of development, physiology, and metabolism of fungi.
As part of the ongoing study of bioactive polysaccharides from Asian fungi [16,35,36,37], this paper presents the results of the isolation and anticomplementary potential of soluble polysaccharides from I. rheades mycelium (IRP). The fraction IRP was studied using chemical analysis, elemental analysis, HPLC-UV, ultraviolet–visible and Fourier transform infrared spectroscopy, gel permeation chromatography, and linkage analysis, followed by the bioactivity in vitro assay to determine the ability of biopolymers to interact with the complement cascade reaction.
2. Materials and Methods
2.1. Fungal Material
The mycelium of the basidiomycete I. rheades (Pers.) Bondartsev & Singer, strain 0186, was purchased from the higher basidiomycete culture collection of the Komarov’s Botanical Institute (Russian Academy of Sciences, St. Petersburg, Russia). The culture was stored in the Bioresource Center (Siberian Institute of Plant Physiology and Biochemistry, Irkutsk, Russia). The pure culture was stored on a lean yeast medium at a temperature of 4 °C [38]. The mycelium was grown on wood discs of Betula pendula Roth. (Betulaceae) in a sterile plastic 5-L container for 30 days, in KBW 720 stationary chambers (Binder, Tuttlingen, Germany) at 25 ± 1 °C. Light-emitting diodes SMD-5050 (Rubicon, Moscow, Russia) were used as a source of radiation (465–470 nm, luminous flux 12.88 W/m2, flux intensity 48.3 µmol/m2·s). The resulting pure mycelium mass was dried to an air-dry state at a temperature no higher than 45 °C.
2.2. Isolation of Total Polysaccharide Fraction of I. rheades Mycelium (IRP)
The previous method of isolation natural polysaccharides [35,39] was used with modifications. Air-dried mycelium (480 g) was milled, then was extracted with hexane (70 °C; mycelium:hexane ratio 1:10; 2 h; two times), 70% ethanol (80 °C; mycelium:ethanol ratio 1:20; 2 h; five times), and hot water (90 °C; mycelium:water ratio 1:15; 2 h; three times). The water extract was centrifuged (6000 rpm, 30 min), concentrated in vacuo (30 times), and concentrated extract was precipitated using acetone (1:5). After 24 h, the precipitate was separated by centrifugation (9000 rpm, 15 min), suspended in warm distilled water (50 °C, 2 L), and the solution was passed through a polyamide column (0.5 kg; Sigma-Aldrich, cat. No. 02395, St. Louis, MI, USA) followed by cation-exchanging column filtration (KU-2-8, H+-form; Eco-Vita, St. Petersburg, Russia; 2 kg). Both columns were eluted with distilled water (3 L). The final water eluate was concentrated to 200 mL (30 °C; in vacuo), and the residue was deproteinated using the Sevag method [35] and protease-assisted deproteination with Streptomyces griseus pronase (type XIV, ≥3.5 units/mg; 1 unit per 1 mL of polysaccharide solution; Sigma-Aldrich, cat. No. P5147) [36]. After dialysis in benzoylated dialysis tubes (cut-off of 2 kDa; Sigma-Aldrich, cat. No. D2272) against distilled water (48 h), the non-dialyzed residue was freeze-dried to give a total polysaccharide fraction of I. rheades mycelium (IRP) as an off-white powder with a yield of 6.14 g.
2.3. Chemical Analysis and Elemental Composition of IRP
Commercial kits were used for spectrophotometric analysis of total carbohydrate content (High Sensitivity Carbohydrate Assay Kit, BioVision, Inc., Milpitas, CA, USA; cat. No. K2049-100), uronic acids (D-Glucuronic/D-Galacturonic Acid Assay Kit, Megazyme, Bray, Ireland; cat. No. K-URONIC), protein (Pierce™ BCA Protein Assay Kit, Thermo Fisher Scientific, Waltham, MA, USA), and phenolics (Phenolic Compounds Assay Kit, Sigma-Aldrich; cat. MAK365). Ash content was determined by the AOAC Official MethodSM 942.05 using muffle furnace ignition at 600 °C [40]. Reaction with Yariv reagent was performed by the Lamport et al. assay [41]. All chemical analyses were performed five times, and the data were expressed as the mean value ± standard deviation (S.D.). For the analysis of carbon, hydrogen, oxygen, and nitrogen contents, a 2400 Series II elemental analyzer (Perkin Elmer, Waltham, MA, USA) was used.
2.4. Monosaccharide Composition
The monosaccharide composition was determined after hydrolysis with 2 M trifluoroacetic acid [42], 1-phenyl-3-methyl-5-pyrazolone (PMP) labeling, and HPLC-UV separation of PMP-labeled monosaccharides as described previously [39]. Water solutions of reference monosaccharides (mannose, ribose, rhamnose, glucose, galactose, xylose, arabinose, fucose, galacturonic acid, glucuronic acid; all 1 mg/mL) after PMP-labeling were used to build calibration curves (peak area vs. the concentration levels). All analyses were performed in triplicate.
2.5. Ultraviolet (UV) and Fourier Transform Infrared (FTIR) Spectroscopy
A Spectrophotometer SF-2000 UV–Vis (OKB Spectr, St. Petersburg, Russia) was used to study the UV spectra of water polysaccharide solutions (5 mg/mL) in the spectral range of 190–1000 nm using a quartz cell (10 mm). To study FTIR spectra, an FT-801 Fourier transform infrared spectrometer (Simex, Novosibirsk, Russia) was used (frequency 4000–600 cm−1, 200 scans, 2-cm−1 resolution) and samples were tableted with potassium bromide (sample:KBr 1:100).
2.6. Molecular Weight Determination and Linkage Analysis
To determine molecular weights, a gel permeation–high performance liquid chromatography procedure was used, with an LCMS 8050 liquid chromatograph coupled with a Shim-pack Diol-150 column (250 × 7.9 mm, ∅ 5 μm; Shimadzu) as described previously [39]. A calibration curve was produced via analysis of dextran series (10–410 kDa; Sigma-Aldrich). All analyses were performed in duplicate. The procedure of linkage analysis was described in a previous study [39], and used methyl iodide methylation, 90% formic acid and 2 M TFA hydrolysis, NaBH4 reduction, and acetic anhydride acetylation [43], followed by gas chromatography–mass spectrometry analysis with a 5973N gas chromatograph mass spectrometer (Agilent Technologies, Santa-Clara, CA, USA) equipped with a 6890N mass selective detector, a diffusion pump, and an HP-Innowax capillary column (Agilent Technologies; 30 m × 250 μm × 0.50 μm) [37].
2.7. Sephacryl 400HR Gel Fractionation of IRP
The water solution of IRP (5 g in 200 mL) was passed through a Sephacryl 400HR gel column (GE Healthcare, Chicago, IL, USA) and eluted with water, detecting the elution progress spectrophotometrically (λ 190, 260 nm). The separate eluates were collected and acetone-precipitated to give fractions of IRP-1 (290 mg), IRP-2 (555 mg), IRP-3 (565 mg), IRP-4 (3.585 g), and IRP-5 (5 mg).
2.8. Partial Hydrolysis of IRP-4 by 0.5% TFA
A sample of IRP-7 (1 g) was incubated at 80 °C with 0.5% TFA (50 mL, 2 h), followed by the TFA elimination in vacuo, dialysis of the residue in dialysis tubes with cut-off 2 kDa (48 h), and freeze-drying of the non-dialyzed sample. The yield of partially hydrolyzed polymer IRP-4d was 320 mg.
2.9. Anticomplementary Activity
The anticomplementary activity of polysaccharides was studied using the method of Samuelson et al. [44] with sheep erythrocytes sensitized with rabbit anti-sheep erythrocyte antibodies (BioTrend, Köln, Deutschland, cat. No 113-4139) and human serum with intact complement proteins from healthy adults. Briefly, sheep erythrocytes pre-washed with 0.9% NaCl and veronal buffer (pH 7.2) were sensitized using rabbit anti-sheep erythrocyte antibodies, incubated (37 °C, 30 min), washed, and used to prepare 1% suspension in veronal buffer. Healthy adult human serum with removed antibodies against sheep erythrocytes after absorption on human red blood cells (5 mM EDTA, 0 °C) was diluted with veronal buffer to give serum showing 50% hemolysis. Polysaccharide solutions in in veronal buffer were mixed with serum (1:1), incubated at 37 °C (30 min), mixed with sensitized sheep erythrocytes (2:1), incubated again (30 min), and centrifuged (2000 rpm, 10 min). Absorbance of the sample supernatant was measured at 405 nm. Distilled water was used as negative control, giving 100% lysis, and MPP-2 polysaccharide from Mentha × piperita was a positive control. Anticomplementary activity was calculated as in the original research [44].
2.10. Statistical and Multivariate Analysis
Statistical analyses were performed using one-way analysis of variance, and the significance of the mean difference was determined using Duncan’s multiple range test. Differences at p < 0.05 were considered statistically significant. The results are presented as the mean ± S.D.
3. Results and Discussion
Water-soluble polysaccharides of I. obliquus have been extensively studied and concluded to be bioactive polymers [I7]; therefore, this was the main reason why water was chosen as the extraction medium for isolation of I. rheades mycelium polysaccharides. The total polysaccharide fraction (IRP) was isolated from I. rheades mycelium with a 1.28% yield, and showed a high carbohydrate content (82.67%) with low levels of protein, uronic acids, phenolics, and ash (Table 1). Negative reactions with iodine, resorcinol, Fehling’s reagent, and Yariv’s reagent indicated a lack of starch [45], fructans [46], galacto/gluco-mannans [47], and arabinogalactan–protein complexes [40], respectively. The elemental composition of IRT showed that the levels for carbon (40.20%), hydrogen (6.68%), and oxygen (53.01%) are typical for neutral polysaccharides [39].
The dominant monosaccharides of IRP were galactose, glucose, and mannose in a ratio of 4.4:1.9:1, and minor components were arabinose, fucose, rhamnose, xylose, and uronic acids with content variations of 0.6–6.5 mol%. The known data on Inonotus polysaccharides demonstrated the domination of glucose (68–97 mol%), mannose (2–19 mol%), and galactose (3–6 mol%) in exopolysaccharides of a submerged culture of the I. obliquus BELYU1102 strain [48]. Cultivated mycelium of the same strain of I. obliquus gave endopolysaccharides containing mannose (23–74 mol%) and glucose (1–15 mol%) [49]. Commercially available wild I. obliquus sclerotium was the source of Un-IOPS polysaccharide with levels of glucose, mannose, and galactose at 48, 21, and 17 mol%, respectively [50]. A series of heteropolysaccharide isolated from Chinese I. obliquus sclerotium contained glucose (31–40 mol%) and galactose (8–14 mol%) [51]. A submerged culture of I. levis gave a polysaccharide fraction that mainly included mannose, galactose, and glucose in a ratio of 3.3:2.3:1 [52]. Thus, given the abovementioned information, galactose, glucose, and mannose are the usual constituents of water-soluble polysaccharides found in the Inonotus genus.
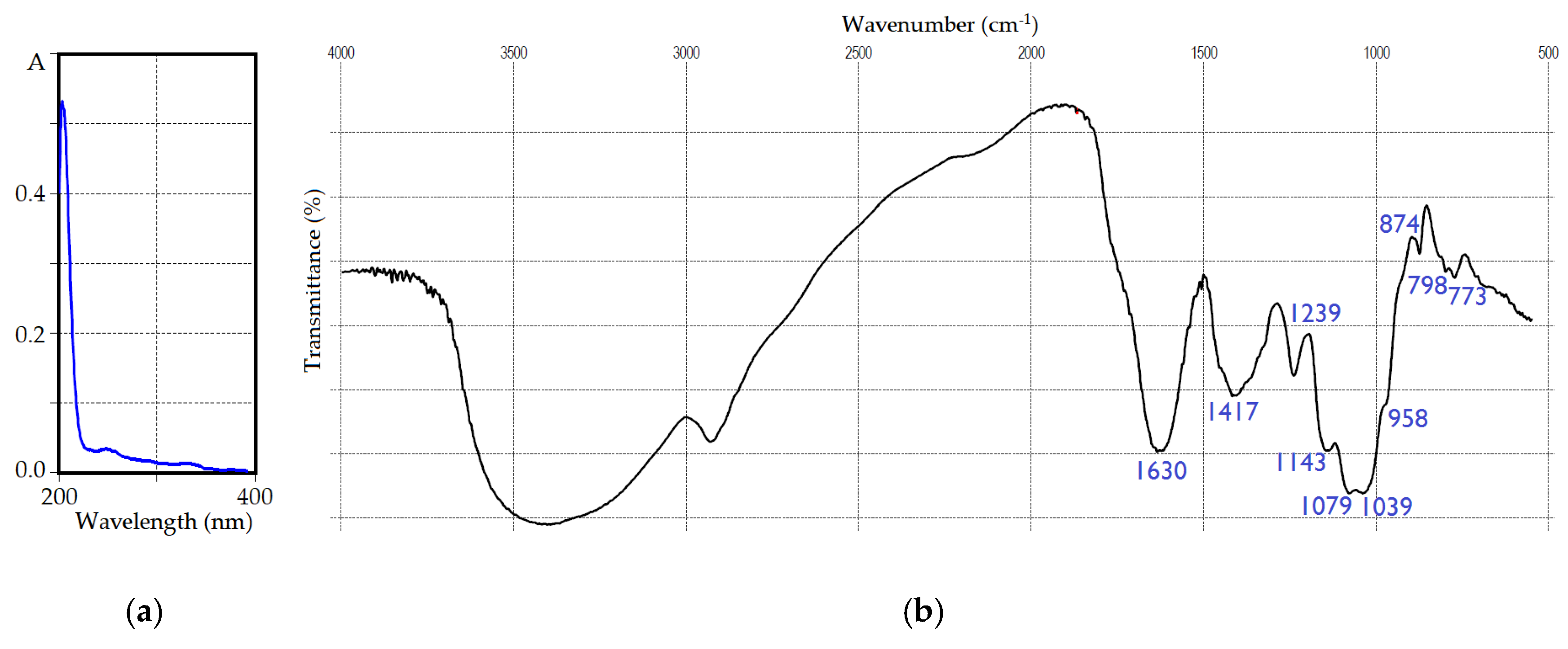
The spectral pattern of IRP in the ultra-violet region (Figure 2a) showed no distinct peaks, owing to its low phenolic and protein contents [53,54,55]. FTIR spectral data (Figure 2b) were typical for natural polysaccharides [56] with bands in a skeletal region (<800 cm−1) attributed to the vibrations of “breathing” rings at 773 cm−1 and sugar cycles at 798 cm−1 [57]. The fingerprint region (1200–800 cm−1) demonstrated vibrations of β-anomeric galactose/mannose aldopyranose rings at 874 cm−1, in-plane bending vibrations of C-OH and C-H linkages of C-1 and symmetric stretching vibrations of glycosidic linkages C-O-C at 958 cm−1, stretching vibrations of glycosidic C-O and C-C at 1039 cm−1, and in-plane bending vibrations of C-OH fragments at 1079 cm−1 [58]. The local symmetry region (1500–1200 cm−1) included asymmetric vibrations of C-O-C linkages at 1143 cm−1, and scissoring and in-plane bending vibrations of CH and CH2 at 1239 and 1417 cm−1, respectively [59]. The intense band at 1630 cm−1 of scissoring vibrations of hydrated water molecules was found in the double bond stretching region [60]. A broad and strong band of stretching vibrations of hydroxyl groups was observed at 3425 cm−1, accompanied by a band of symmetric and asymmetric stretching vibrations of skeletal C-H and CH2 fragments [61].
Gel permeation chromatography (Sephacryl 400HR) indicated the heterogeneity of IRP (Figure 3a), and then five homogenic fraction were isolated. Three polymers with low retention were IRP-1, IRP-2, and IRP-3 with molecular weights of 1520, 1150, and 820 kDa, respectively (Table 2). The IRP-1 polymer was characterized by the predominance of glucose, galactose, and mannose in the ratio of 6.1:1.2:1, while IRP-2 also contained glucose, galactose, and mannose as the main monosaccharides but in a different ratio of 2.9:2.2:1. For IRP-3, an increased content of xylose (15.9 mol%) was noted; the ratio of the main monosaccharides galactose, mannose, and xylose was 3.6:1.2:1.
The dominant polymer, IRP-4, with a 71.7% yield of IRP weight, had a molecular weight of 148 kDa and was a heteropolysaccharide containing galactose, mannose, and glucose as the main monosaccharides in a ratio of 4.2:1.4:1. The level of minor monosaccharides accounted for no more than 4.5 mol%. The polymer IRP-5 with the lowest molecular weight (110 kDa) was a glucan with a glucose content of 81.7 mol%. Thus, the complex of water-soluble polysaccharides of I. rheades mycelium is a mixture of predominantly glucans (IRP-1, IRP-2, and IRP-5) and galactans (IRP-3 and IRP-4) with varied molecular weights.
Considering the high content of IRP-4 in the IRP total fraction, we studied its structure after methylation and analysis of O-methylated alditol acetates using GC–MS. According to methylation data, IRP-4 was a highly branched polysaccharide containing residues of galactose (2,3,4,6-Me4-Gal), mannose (2,3,4,6-Me4-Man), glucose (2,3,4,6-Me4-Glc), xylose (2,3,4-Me3-Xyl), arabinose (2,3,4-Me3-Ara), and fucose (2,3,4-Me3-Fuc) at the non-reducing ends of the chains (Table 3). A high content of 6-O-(2,3,4-Me3-Gal) and 3,6-di-O-substituted galactose residues (2,4-Me2-Gal) was revealed, resulting in 56.3% residues in the polymer molecule. The presence of 3-O-substituted galactose (2,4,6-Me3-Gal) and mannose (3,4,6-Me3-Man), as well as 6-O-substituted glucose (2,3,4-Me3-Glc), was also established. These last residues likely form the side chains of the polysaccharide.
To determine the type of monosaccharides that form the main chain of the polymer molecule, partial hydrolysis was performed, which led to the formation of the IRP-4d fragment with a molecular weight of 25 kDa, the only component of which was galactose.
Analysis of alditol acetates after methylation showed that the main fragment of IRP-4d was 6-O-substituted galactose (2,3,4-Me3-Gal; 92.6%), which constructs the core of the polymer molecule in the form of a chain [→6)-Gal-(1→]. Thus, the performed studies allowed the preliminary structure of the dominant polymer IRP-4 as (1→6)-linked galactan to be established for the first time, in which more than 60% of galactose residues are substituted at the C-3 position by various carbohydrate fragments.
To study the biological effects of individual polysaccharides of I. rheades mycelium, their anticomplementary activities were determined on the model of inhibition of hemolysis of sensitized sheep erythrocytes with complement from human serum [44]. This method allows the study of the ability of polysaccharides to participate in the cascade of reactions of the complement system.
The IRP-1, IRP-2, and IRP-5 polysaccharides had a weak complement-fixing activity, demonstrating binding efficiencies of 12, 18, and 12%, respectively (Figure 4), at concentrations of 500 μg/mL. The binding index for the reference polysaccharide used as a positive control (MPP′-2 polysaccharide from Mentha × piperita) was 75% [62].
Polysaccharides IRP-4 and IRP-5 showed the highest anti-complementary activities, amounting to 38% and 49% at concentrations of 500 μg/mL, respectively. Thus, considering the chemical composition data of the studied polysaccharides, it can be argued that the biological activities of the components that are glucans (IRP-1, IRP-2, and IRP-5) were significantly lower than those of polymers enriched in galactose, the galactans (IRP-3 and IRP-4). A comparative analysis of the anti-complementary activity of the highly branched galactan IRP-4 and the unbranched fragment of its polysaccharide core IRP-4d indicated the low efficiency of the latter (21% at 500 μg/mL), which showed the important role of the degree of branching for polysaccharide biological activity.
The early knowledge about the structures of Inonotus polysaccharides revealed that homogenous polymer IOI-WN of I. obliquus sclerotia is (1→3)-linked β-glucan branched at every fifth C-6 position and that the IOE-WN polymer is 3-O-methylated (1→6)-linked α-galactan branched at every third galactose fragment [17]. Cultivated mycelium and wild sterile conk of I. obliquus gave the mixture of (1→3,6)-linked β-glucans and methylated galactans as minor components [14]. Mixed (1→3,6)-linked galactan consisting of galactose and 3-O-methylated galactose in molecular weight was isolated from a submerged culture of I. levis together with the minor (1→2,6)-linked mannan [52]. Thus, the isolation of glucans and galactans from I. rheades mycelium in our study was not uncommon. Despite the lack of knowledge, the most likely explanation is that glucans and galactans are the principal polysaccharides of the Inonotus genus. This makes even more sense if we remember that other fungi of the Hymenochaetaceae family were determined to be sources of polysaccharides; a good example is the Phellinus genus enriched with (1→3,6)-linked glucans and (1→3,6)-linked galactans with various fine structures [63].
The complement system is an important part of the body′s immune system, which includes various serum protein components C1–C9 that are activated according to the cascade principle by classical and alternative pathways [64]. The activation of the complement system plays an important role in initiating inflammatory and immune responses including leukocyte activation and degranulation of basophils and mast cells. The anticomplementary potential of natural polysaccharides is known for the polymers isolated from Echinacea purpurea (L.) Moench (Asteraceae) [65], Panax ginseng C.A. Meyer (Araliaceae) [66], Abelmoschus esculentus L. (Malvaceae) [67], and many others [64]. The complement-fixing activity of Inonotus polysaccharides has not been previously studied, but it is known that triterpenoids 3β-hydroxy-8,24-dien-21-al and inotodiol and the total melanin fraction of I. obliquus sclerotia are potent anticomplementary agents [68].
Anticomplementary potential was reported for (1→3,6)-linked galactans from Angelica acutiloba Kitagawa (Apiaceae) [69], Terminalia macroptera Guill. & Perr. (Combretaceae) [70], and Panax L. plants (Araliaceae) [71]. Arabino-3,6-galactans from Angelica acutiloba roots showed potent anticomplementary activity through the classical pathway expressed by the neutral galactan chains to a greater extent than arabinan chains [72]. The elimination of arabinose from arabino-3,6-galactans increased the activity of polysaccharides, demonstrating the essential role of the galactan core for the expression of anticomplementary potential. A similar mode of activity was demonstrated by neutral 3,6-galactans from Malva verticillata L. (Malvaceae) seeds [73] and Angelica gigas Nakai (Apiaceae) roots [74]. Anticomplementary 3,6-O-linked galactan TM4a from Teucrium viscidum Blume leaves used the classical and alternative pathways for the activation of the immune system and interacted with C3 and C5 components [75]. Two 4,6-galactans from the herb of Eclipta prostrata (L.) L. (Asteraceae) interacted with C1, C2, C4, C5, C7, and C9 components, resulting in the inhibition of the activation of the complement system [76].
However, despite the well-known fact that some natural galactans possessed anticomplementary activity, there are no any studies aimed at the identification of specific targets in the complement system interacting with fungal galactans. Regardless, it may be suggested that 3,6-linked galactan from I. rheades mycelium can provide anticomplementary activity through the classical pathway, as preliminary data demonstrated the similarity of its structure to the known bioactive plant polysaccharides. It is most likely that galactan IRP-4 can interact with serum protein components C1–C9, which will be studied later. However, nothing can be concluded regarding the structural similarity of polysaccharides of I. rheades mycelium and plant polymers until additional studies of I. rheades polysaccharides are completed. The indisputable fact remains that natural galactans are prospective bioactive agents due to considerable evidence. The obtained results showed that galactan IRP-4 is an anticomplementary polysaccharide, indicating its potential immunostimulatory and anti-inflammatory properties.
4. Conclusions
Fox polypore or Inonotus rheades (Pers.) Karst. is a known xylotrophic basidiomycete occurring in the deciduous forests of Europe and Asia. In this study, we demonstrated that the mycelium of I. rheades is able to accumulate a heterogenic complex of five polysaccharides with high amounts of galactose, glucose, and mannose and varied molecular weights for the first time. The dominant polymer of I. rheades mycelium IRP-4 with a molecular weight of 148 kDa was purified, and linkage analysis showed that it is a branched (1→3,6)-linked galactan. The results of the bioactivity study showed that I. rheades polymers are anticomplementary polysaccharides, and galactan IRP-4 was the most active. Our work demonstrated for the complement-fixing potential of polysaccharides from Inonotus fungi the first time. These findings suggest that polysaccharides may be active components of the I. rheades mycelium, which, in turn, can be a source of new anticomplementary agents.
Author Contributions
Conceptualization, methodology, software, validation, formal analysis, investigation, resources, data curation, writing—original draft preparation, writing—review and editing, visualization, supervision, project administration, funding acquisition, D.N.O. and T.G.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education and Science of Russia, grant number 121030100227-7.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are very thankful to Marina S. Polyakova (Siberian Institute of Plant Physiology and Biochemistry, Irkutsk, Russia) for kindly providing the photo of I. rheades in its natural habitat.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Mukhin, V.A.; Diyarova, D.K. Eco-physiological adaptations of the xylotrophic basidiomycetes fungi to CO2 and O2 mode in the woody habitat. J. Fungi 2022, 8, 1296. [Google Scholar] [CrossRef] [PubMed]
- Góes-Neto, A.; Loguercio-Leite, C.; Guerrero, R.T. Molecular phylogeny of tropical Hymenochaetales (Basidiomycota). Mycotaxon 2002, 84, 337–354. [Google Scholar]
- Balandaykin, M.E.; Zmitrovich, I.V. Review on chaga medicinal mushroom, Inonotus obliquus (higher basidiomycetes): Realm of medicinal applications and approaches on estimating its resource potential. Int. J. Med. Mushrooms 2015, 17, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.W.; Elkhateeb, W.A.; Daba, G.M. Chaga (Inonotus obliquus): A medical marvel becomes a conservation dilemma. Sydowia 2020, 72, 123–130. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Bartosz, S.; Tadeusz, P.; Jan, G. Inonotus obliquus–from folk medicine to clinical use. J. Tradit. Complement. Med. 2020, 11, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Ryvarden, L.; Gilbertson, R.L. European Polypores Part 1: Abortiporus-Lindtneria. In Synopsis Fungorum; Fungiflora A/S: Oslo, Norway, 1993; Volume 6, pp. 387–393. [Google Scholar]
- Olennikov, D.N.; Gornostai, T.G.; Penzina, T.A. Rheadinin, a new bis(styrylpyrone) from mycelium of Inonotus rheades. Chem. Nat. Compd. 2017, 53, 629–631. [Google Scholar] [CrossRef]
- Gornostai, T.G.; Borovskii, G.G.; Kashchenko, N.I.; Olennikov, D.N. Phenolic compounds of Inonotus rheades (Agaricomycetes) mycelium: RP-UPLC-DAD-ESI/MS profile and effect of light wavelength on styrylpyrone content. Int. J. Med. Mushrooms 2018, 20, 637–645. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Gornostai, T.G.; Penzina, T.A.; Borovskii, G.B. Lupane triterpenoids and sterols from Inonotus rheades mycelium and their anti-glucosidase activity. Chem. Nat. Compd. 2017, 53, 988–990. [Google Scholar] [CrossRef]
- Gornostay, T.G.; Polyakova, M.S.; Borovskii, G.B.; Olennikov, D.N. Lipids of Inonotus rheades (Hymenochaetaceae): Influence of substrate and light mode on fatty acid profile of mycelium. Chem. Plant Raw Mater. 2018, 22, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Gornostay, T.G.; Penzina, T.A.; Chkhenkeli, V.A.; Polyakova, M.S.; Osipenko, S.N.; Borovskii, G.B.; Olennikov, D.N. Research of antioxidant and antimicrobial activity of water-alcohol extractions of mycothallus and mycelium of Inonotus rheades. Basic Clin. Pharmacol. Toxicol. 2014, 115, 10. [Google Scholar] [CrossRef] [Green Version]
- Gornostai, T.; Stepanov, A.; Olennikov, D.; Borovskii, G. Antioxidant activity of styrylpyrones from mycelium of Inonotus rheades (Agaricomycetes) under oxidative stress of Thellungiella salsuginea cell suspension culture. Int. J. Med. Mushrooms 2021, 23, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Jia, Y.; Xue, Z.; Li, N.; Liu, J.; Chen, H. Recent developments in Inonotus obliquus (chaga mushroom) polysaccharides: Isolation, structural characteristics, biological activities and application. Polymers 2021, 13, 1441. [Google Scholar] [CrossRef] [PubMed]
- Beltrame, G.; Trygg, J.; Hemming, J.; Han, Z.; Yang, B. Comparison of polysaccharides extracted from cultivated mycelium of Inonotus obliquus with polysaccharide fractions obtained from sterile conk (chaga) and birch heart rot. J. Fungi 2021, 7, 189. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Zhuang, C.; Abe, K.; Okamoto, H.; Kiho, T.; Ukai, S.; Leclerc, S.; Meijer, L. Antitumor and hypoglycemic activities of polysaccharides from the sclerotia and mycelia of Inonotus obliquus (Pers.:Fr.) Pil. (Aphyllophoromycetideae). Int. J. Med. Mushrooms 1999, 1, 301–316. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Rokhin, A.V.; Agafonova, S.V. Base-soluble polysaccharides of Inonotus obliquus sclerotia. Chem. Nat. Compd. 2011, 47, 971–972. [Google Scholar] [CrossRef]
- Wold, C.W.; Kjeldsen, C.; Corthay, A.; Rise, F.; Christensen, B.E.; Duus, J.Ø.; Inngjerdingen, K.T. Structural characterization of bioactive heteropolysaccharides from the medicinal fungus Inonotus obliquus (chaga). Carbohydr. Polym. 2018, 185, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.O.; Park, H.W.; Kim, J.H.; Lee, J.Y.; Moon, S.H.; Shin, C.S. Anti-cancer effect and structural characterization of endo-polysaccharide from cultivated mycelia of Inonotus obliquus. Life Sci. 2006, 79, 72–80. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Huang, Y.R.; Cui, Z.M.; Liu, J.J. Purification, characterization and biological activity of a novel polysaccharide from Inonotus obliquus. Int. J. Biol. Macromol. 2015, 79, 587–594. [Google Scholar] [CrossRef]
- Lee, K.R.; Lee, J.S.; Lee, S.; Son, Y.K.; Kim, G.R.; Sim, Y.C.; Song, J.E.; Ha, S.J.; Hong, E.K. Polysaccharide isolated from the liquid culture broth of Inonotus obliquus suppresses invasion of B16-F10 melanoma cells via AKT/NF-kappa B signaling pathway. Mol. Med. Rep. 2016, 14, 4429–4435. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.R.; Lee, J.S.; Song, J.E.; Ha, S.J.; Hong, E.K. Inonotus obliquus-derived polysaccharide inhibits the migration and invasion of human non-small cell lung carcinoma cells via suppression of MMP-2 and MMP-9. Int. J. Oncol. 2014, 45, 2533–2540. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Hu, D.; Cui, Z.; Li, H.; Man, C.; Jiang, Y. Lipid-lowering effects of Inonotus obliquus polysaccharide in vivo and in vitro. Foods 2021, 10, 3085. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, Z.; Zhou, X.; Hu, J.; Xue, J.; Liu, X.; Zhang, J.; Liu, P.; Tong, S. Simultaneous use of stimulatory agents to enhance the production and hypoglycaemic activity of polysaccharides from Inonotus obliquus by submerged fermentation. Molecules 2019, 24, 4400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Tong, S.; Wang, Z.; Liu, P. Chemical characterization and hypoglycaemic activities in vitro of two polysaccharides from Inonotus obliquus by submerged culture. Molecules 2018, 23, 3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Xue, J.; Tong, S.; Dong, W.; Wu, P. Structure characterization and hypoglycaemic activities of two polysaccharides from Inonotus obliquus. Molecules 2018, 23, 1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Yu, D.; Li, L.; Liu, X.; Zhang, H.; Sun, W.; Lin, C.-C.; Chen, J.; Chen, Z.; Wang, W.; et al. Three-phase partitioning for the extraction and purification of polysaccharides from the immunomodulatory medicinal mushroom Inonotus obliquus. Molecules 2019, 24, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, H.; Zhang, A.; Zhang, W.; Cui, G.; Wang, S.; Duan, J. Antioxidative properties of crude polysaccharides from Inonotus obliquus. Int. J. Mol. Sci. 2012, 13, 9194–9206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.J.; Guo, J.Y.; Cheng, H.; Li, L.; Liu, Y.; Shi, Y.; Xu, J.; Yu, H.T. Spatial structure and anti-fatigue of polysaccharide from Inonotus obliquus. Int. J. Biol. Macromol. 2020, 151, 855–860. [Google Scholar] [CrossRef]
- Tian, J.; Hu, X.L.; Liu, D.F.; Wu, H.X.; Qu, L.D. Identification of Inonotus obliquus polysaccharide with broad-spectrum antiviral activity against multi-feline viruses. Int. J. Biol. Macromol. 2017, 95, 160–167. [Google Scholar] [CrossRef]
- Chen, Y.F.; Zheng, J.J.; Qu, C.; Xiao, Y.; Li, F.F.; Jin, Q.X.; Li, H.H.; Meng, F.P.; Jin, G.H.; Jin, D. Inonotus obliquus polysaccharide ameliorates dextran sulphate sodium induced colitis involving modulation of Th1/Th2 and Th17/Treg balance. Artif. Cell. Nanomed. Biotechnol. 2019, 47, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Sang, R.; Yu, Y.F.; Li, J.X.; Ge, B.J.; Zhang, X.M. The polysaccharide from Inonotus obliquus protects mice from Toxoplasma gondii-induced liver injury. Int. J. Biol. Macromol. 2019, 125, 1–8. [Google Scholar] [CrossRef]
- Jiao, S.; Lu, Y. Abundant fungi adapt to broader environmental gradients than rare fungi in agricultural fields. Glob. Chang. Biol. 2020, 26, 4506–4520. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.-M.; Zhang, L.-C.; Liang, H.-Q.; Lv, J.; Song, C.; Guo, S.-X. Sclerotial formation of Polyporus umbellatus by low temperature treatment under artificial conditions. PLoS ONE 2013, 8, e56190. [Google Scholar] [CrossRef]
- Zhou, S.; Raouche, S.; Grisel, S.; Navarro, D.; Sigoillot, J.-C.; Herpoël-Gimbert, I. Solid-state fermentation in multi-well plates to assess pretreatment efficiency of rot fungi on lignocellulose biomass. Microb. Biotechnol. 2015, 8, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Agafonova, S.V.; Borovskii, G.B.; Penzina, T.A.; Rokhin, A.V. Water-soluble polysaccharides of Laetiporus sulphureus (Bull.: Fr.) Murr. fruit bodies. Appl. Biochem. Microbiol. 2009, 45, 536–543. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Agafonova, S.V.; Borovskii, G.B.; Penzina, T.A.; Rokhin, A.V. Alkali-soluble polysaccharides of Laetiporus sulphureus (Bull.: Fr.) Murr. fruit bodies. Appl. Biochem. Microbiol. 2009, 45, 626–630. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Agafonova, S.V.; Rokhin, A.V.; Penzina, T.A.; Borovskii, G.B. Branched glucan from the fruiting bodies of Piptoporus betulinus (Bull.:Fr) Karst. Appl. Biochem. Microbiol. 2012, 48, 65–70. [Google Scholar] [CrossRef]
- Richardson, L.T. A simple culture tube closure method or prevention of contamination by airborne fungi and mites. Phytopathology 1975, 65, 833–834. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chemposov, V.V.; Chirikova, N.K. Polymeric compounds of lingonberry waste: Characterization of antioxidant and hypolipidemic polysaccharides and polyphenol-polysaccharide conjugates from Vaccinium vitis-idaea press cake. Foods 2022, 11, 2801. [Google Scholar] [CrossRef]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of ash in animal Feed: AOAC Official Method 942.05 revisited. J. AOAC Int. 2012, 95, 1392–1397. [Google Scholar] [CrossRef]
- Lamport, D.T.A.; Kieliszewski, M.J.; Showalter, A.M. Salt stress upregulates periplasmic arabinogalactan proteins: Using salt stress to analyse AGP function. New Phytol. 2006, 169, 479–492. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Gornostai, T.G.; Selyutina, I.Yu.; Zilfikarov, I.N. Effect of low temperature cultivation on the phytochemical profile and bioactivity of Arctic plants: A case of Dracocephalum palmatum. Int. J. Mol. Sci. 2017, 18, 2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Needs, P.W.; Selvendran, R.R. Avoiding oxidative degradation during sodium hydroxide/methyl iodide-mediated carbohydrate methylation in dimethyl sulfoxide. Carbohydr. Res. 1993, 245, 1–10. [Google Scholar] [CrossRef]
- Samuelsen, A.; Lund, I.; Djahromi, J.; Paulsen, B.; Wold, J.; Knutsen, S. Structural features and anti-complementary activity of some heteroxylan polysaccharide fractions from the seeds of Plantago major L. Carbohydr. Polym. 1999, 38, 133–143. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. A quantitative starch–iodine method for measuring alpha-amylase and glucoamylase activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Tankhaev, L.M. A quantitative assay for total fructans in burdock (Arctium spp.) roots. Russ. J. Bioorg. Chem. 2011, 37, 893–898. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Rokhin, A.V. Polysaccharides of Fabaceae. I. Galactomannan of Astragalus sericeocanus seeds. Chem. Nat. Compd. 2008, 44, 685–687. [Google Scholar] [CrossRef]
- Kim, Y.O.; Han, S.B.; Lee, H.W.; Ahn, H.J.; Yoon, Y.D.; Jung, J.K.; Kim, H.M.; Shin, C.S. Immuno-stimulating effect of the endo-polysaccharide produced by submerged culture of Inonotus obliquus. Life Sci. 2005, 77, 2438–2456. [Google Scholar] [CrossRef]
- Sun, Y.; He, H.; Wang, Q.; Yang, X.; Jiang, S.; Wang, D. A review of development and utilization for edible fungal polysaccharides: Extraction, chemical characteristics, and bioactivities. Polymers 2022, 14, 4454. [Google Scholar] [CrossRef]
- Ma, L.; Chen, H.; Zhang, Y.; Zhang, N.; Fu, L. Chemical modification and antioxidant activities of polysaccharide from mushroom Inonotus obliquus. Carbohydr. Polym. 2012, 89, 371–378. [Google Scholar] [CrossRef]
- Du, X.; Mu, H.; Zhou, S.; Zhang, Y.; Zhu, X. Chemical analysis and antioxidant activity of polysaccharides extracted from Inonotus obliquus sclerotia. Int. J. Biol. Macromol. 2013, 62, 691–696. [Google Scholar] [CrossRef]
- Vinogradov, E.; Wasser, S.P. The structure of a polysaccharide isolated from Inonotus levis P. Karst. mushroom (Heterobasidiomycetes). Carbohydr. Res. 2005, 340, 2821–2825. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk-Czepas, J.; Bijak, M.; Saluk, J.; Ponczek, M.B.; Zbikowska, H.M.; Nowak, P.; Tsirigotis-Maniecka, M.; Pawlaczyk, I. Radical scavenging and antioxidant effects of Matricaria chamomilla polyphenolic–polysaccharide conjugates. Int. J. Biol. Macromol. 2015, 72, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, W.; Zhao, J.; Zhang, Z.; Zhang, Y. Some physical properties of protein moiety of alkali-extracted tea polysaccharide conjugates were shielded by its polysaccharide. Molecules 2017, 22, 914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.-Q.; Zhang, Z.-F.; Gao, Z.-M.; Huang, Y.; Wu, Z.-Q. Physicochemical properties and cell-based bioactivity of Pu’erh tea polysaccharide conjugates. Int. J. Biol. Macromol. 2017, 104, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Gomba, G.K.; Synytsya, A.; Švecová, P.; Coimbra, M.A.; Čopíková, J. Distinction of fungal polysaccharides by N/C ratio and mid infrared spectroscopy. Int. J. Biol. Macromol. 2015, 80, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Robic, A.; Bertrand, D.; Sassi, J.-F.; Lerat, Y.; Lahaye, M. Determination of the chemical composition of ulvan, a cell wall polysaccharide from Ulva spp. (Ulvales, Chlorophyta) by FT-IR and chemometrics. J. Appl. Phycol. 2008, 21, 451–456. [Google Scholar] [CrossRef]
- Wiercigroch, E.; Szafraniec, E.; Czamara, K.; Pacia, M.Z.; Majzner, K.; Kochan, K.; Malek, K. Raman and infrared spectroscopy of carbohydrates: A review. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 185, 317–335. [Google Scholar] [CrossRef]
- Kacuráková, M. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym. 2000, 43, 195–203. [Google Scholar] [CrossRef]
- Borovkova, V.S.; Malyar, Y.N.; Sudakova, I.G.; Chudina, A.I.; Skripnikov, A.M.; Fetisova, O.Y.; Kazachenko, A.S.; Miroshnikova, A.V.; Zimonin, D.V.; Ionin, V.A.; et al. Molecular characteristics and antioxidant activity of spruce (Picea abies) hemicelluloses isolated by catalytic oxidative delignification. Molecules 2022, 27, 266. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Xin, Y.; Yin, J.-Y.; Huang, X.-J.; Wang, J.-Q.; Hu, J.-L.; Nie, S.-P. Revealing the architecture and solution properties of polysaccharide fractions from Macrolepiota albuminosa (Berk.) Pegler. Food Chem. 2022, 368, 130772. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Tankhaeva, L.M. Lamiaceae carbohydrates. I. Pectinic substances and hemicelluloses from Mentha × piperita. Chem. Nat. Compd. 2007, 43, 501–507. [Google Scholar] [CrossRef]
- Luan, F.; Peng, X.; Zhao, G.; Zeng, J.; Zou, J.; Rao, Z.; Liu, Y.; Zhang, X.; Ma, H.; Zeng, N. Structural diversity and bioactivity of polysaccharides from medicinal mushroom Phellinus spp.: A review. Food Chem. 2022, 397, 133731. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Kiyohara, H. Complement-activating polysaccharides from medicinal herbs. In Immunomodulatory Agents from Plants; Wagner, H., Ed.; Birkhäuser: Basel, Switzerland, 1999; pp. 161–202. [Google Scholar] [CrossRef]
- Stimple, M.; Proksch, A.; Wagner, H.; Lohmann-Matthes, M.L. Macrophage activation and induction of macrophage cytotoxicity by purified polysaccharide fractions from the plant Echinacea purpurea. Infect. Immun. 1984, 46, 845–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.P.; Kiyohara, H.; Cyong, J.C.; Yamada, H. Characterization of anti-complementary acidic heteroglycans from the leaves of Panax ginseng C. A. Meyer. Carbohydr. Res. 1988, 181, 175–187. [Google Scholar] [CrossRef]
- Tomoda, M.; Shimizu, N.; Gonda, R.; Kanari, M.; Yamada, H.; Hikino, H. Anti-complementary and hypoglycemic activities of okra and hibiscus mucilages. Carbohydr. Res. 1989, 190, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wold, C.W.; Gerwick, W.H.; Wangensteen, H.; Inngjerdingen, K.T. Bioactive triterpenoids and water-soluble melanin from Inonotus obliquus (chaga) with immunomodulatory activity. J. Funct. Foods 2020, 71, 104025. [Google Scholar] [CrossRef]
- Zhang, Y.; Kiyohara, H.; Sakurai, M.H.; Yamada, H. Complement activating galactan chains in a pectic arabinogalactan (AGIIb-1) from the roots of Angelica acutiloba Kitagawa. Carbohydr. Polym. 1996, 31, 149–156. [Google Scholar] [CrossRef]
- Zou, Y.-F.; Zhang, B.-Z.; Barsett, H.; Inngjerdingen, K.T.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. Complement fixing polysaccharides from Terminalia macroptera root bark, stem bark and leaves. Molecules 2014, 19, 7440–7458. [Google Scholar] [CrossRef]
- He, Y.; Niu, Z.; Shen, T.; Zhang, J.; Wang, X.; Hu, W.; Cho, J.Y. A review of the immunomodulatory activities of polysaccharides isolated from Panax species. J. Ginseng Res. 2022, 46, 23–32. [Google Scholar] [CrossRef]
- Kiyohara, H.; Cyong, J.-C.; Yamada, H. Relationship between structure and activity of an anticomplementary arabinogalactan from the roots of Angelica acutiloba Kitagawa. Carbohydr. Res. 1989, 193, 193–200. [Google Scholar] [CrossRef]
- Tomoda, M.; Shimizu, N.; Gonda, R.; Kanari, M.; Yamada, H.; Hikino, H. Anti-complementary and hypoglycemic activities of the glycans from the seeds of Malva verticillata. Planta Medica 1990, 56, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.S. Roles of sugar chains in immunostimulatory activity of the polysaccharide isolated from Angelica gigas. Korean J. Food Sci. Technol. 2019, 51, 336–342. [Google Scholar] [CrossRef]
- Min, J.G. Purification and characterization of anti-complementary polysaccharides from Teucrium viscidum. J. Food Nutr. Res. 2017, 5, 399–405. [Google Scholar] [CrossRef]
- Wang, H.; Li, N.; Zhu, C.; Shi, S.; Jin, H.; Wang, S. Anti-complementary activity of two homogeneous polysaccharides from Eclipta prostrata. Biochem. Biophys. Res. Commun. 2017, 493, 887–893. [Google Scholar] [CrossRef]
Figure 1.
Fruit bodies of Inonotus rheades (Pers.) Karst. (fox polypore) in its natural habitat (Kadinskii reserve, Kuitun District, Irkutsk Oblast, Russia) (a) and lab-grown I. rheades mycelium (b).
Figure 1.
Fruit bodies of Inonotus rheades (Pers.) Karst. (fox polypore) in its natural habitat (Kadinskii reserve, Kuitun District, Irkutsk Oblast, Russia) (a) and lab-grown I. rheades mycelium (b).

Figure 2.
UV (a) and FTIR spectra (b) of IRP polysaccharide.

Figure 3.
Elution curve of IRP fractions on the Sephacryl 400HR (a) and HPLC-UV chromatograms of PMP-labeled monosaccharides in IRP fraction hydrolysates (b). IRP fractions: 1—IRP-1, 2—IRP-2, 3—IRP-3, 4—IRP-4, 5—IRP-5.
Figure 3.
Elution curve of IRP fractions on the Sephacryl 400HR (a) and HPLC-UV chromatograms of PMP-labeled monosaccharides in IRP fraction hydrolysates (b). IRP fractions: 1—IRP-1, 2—IRP-2, 3—IRP-3, 4—IRP-4, 5—IRP-5.

Figure 4.
Anticomplementary activity of IRP-1–IRP-5, and IRP-4d polysaccharides. Positive control—MPP-2 polysaccharide from Mentha × piperita. Asterisks indicate significant difference (p < 0.05) vs. the MPP-2 group, 50 μg/mL (*); vs. the MPP-2 group, 200 μg/mL (**); vs. the MPP-2 group, 500 μg/mL (***).
Figure 4.
Anticomplementary activity of IRP-1–IRP-5, and IRP-4d polysaccharides. Positive control—MPP-2 polysaccharide from Mentha × piperita. Asterisks indicate significant difference (p < 0.05) vs. the MPP-2 group, 50 μg/mL (*); vs. the MPP-2 group, 200 μg/mL (**); vs. the MPP-2 group, 500 μg/mL (***).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Yield and chemical composition of total polysaccharide fraction of I. rheades mycelium (IRP).
Table 1.
Yield and chemical composition of total polysaccharide fraction of I. rheades mycelium (IRP).
| Parameter | Value |
|---|---|
| IRP yield, % of dry mycelium weight | 1.28 |
| Carbohydrates, % of IRP weight | 82.67 ± 2.14 |
| Protein, % of IRP weight | 0.54 ± 0.00 |
| Uronic acids, % of IRP weight | 2.40 ± 0.06 |
| Phenolics, % of IRP weight | 0.05 ± 0.00 |
| Ash, % of IRP weight | 1.24 ± 0.02 |
| Reaction with iodine | negative |
| Reaction with resorcinol | negative |
| Reaction with Fehling’s reagent | negative |
| Reaction with Yariv’s reagent | negative |
| Elemental composition, % | |
| Carbon | 40.20 |
| Hydrogen | 6.68 |
| Oxygen | 53.01 |
| Nitrogen | 0.11 |
| Monosaccharide composition, mol% | |
| Ara | 3.7 |
| Fuc | 6.5 |
| Gal | 49.1 |
| Glc | 21.5 |
| Man | 11.1 |
| Rha | 1.2 |
| Xyl | 4.7 |
| GalA | 1.5 |
| GlcA | 0.6 |
Table 2.
Yield, molecular weight, and monosaccharide composition of IRP fractions after Sephacryl 400HR separation.
Table 2.
Yield, molecular weight, and monosaccharide composition of IRP fractions after Sephacryl 400HR separation.
| Parameter | IRP-1 | IRP-2 | IRP-3 | IRP-4 | IRP-5 |
|---|---|---|---|---|---|
| Yield, % of IRP weight | 5.8 | 11.1 | 11.3 | 71.7 | 0.1 |
| Mw, kDa | 1520 (±1.2%) | 1150 (±1.7%) | 820 (±1.0%) | 148 (±2.9%) | 110 (±2.7%) |
| Mw/Mn | 1.26 (±1.5%) | 1.40 (±1.8%) | 1.1 (±2.1%) | 1.62 (±1.9%) | 1.57 (±1.6%) |
| Monosaccharide composition, mol% | |||||
| Ara | 2.8 | 2.2 | - | 1.7 | 2.4 |
| Fuc | - | 0.8 | 3.4 | 1.0 | - |
| Gal | 13.8 | 33.7 | 57.5 | 60.9 | 8.7 |
| Glc | 71.0 | 44.5 | 4.9 | 14.5 | 81.7 |
| Man | 11.6 | 15.3 | 18.3 | 20.3 | 7.1 |
| Rha | 0.7 | - | - | - | - |
| Xyl | - | 3.4 | 15.9 | 1.5 | - |
Table 3.
Content of O-methyl-alditol acetates in hydrolysates of IRP-4 and IRP-4d polymers.
| O-Methyl-Alditol Acetate | Content in Hydrolisates, % | Linkage Type | |
|---|---|---|---|
| IRP-4 | IRP-4d | ||
| 2,3,4,6-Me4-Gal | 0.9 | 3.6 | Gal-(1→ |
| 2,4,6-Me3-Gal | 7.2 | →3)-Gal-(1→ | |
| 2,3,4-Me3-Gal | 21.6 | 92.9 | →6)-Gal-(1→ |
| 2,4-Me2-Gal | 34.7 | 3.5 | →3,6)-Gal-(1→ |
| 2,3,4,6-Me4-Man | 17.8 | Man-(1→ | |
| 3,4,6-Me3-Man | 2.5 | →2)-Man-(1→ | |
| 2,3,4,6-Me4-Glc | 14.0 | Glc-(1→ | |
| 2,3,4-Me3-Glc | 0.5 | →6)-Glc-(1→ | |
| 2,3,4-Me3-Xyl | 0.5 | Xyl-(1→ | |
| 2,3,4-Me3-Ara | 0.2 | Ara-(1→ | |
| 2,3,4-Me3-Fuc | 0.1 | Fuc-(1→ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Olennikov, D.N.; Gornostai, T.G. New Inonotus Polysaccharides: Characterization and Anticomplementary Activity of Inonotus rheades Mycelium Polymers. Polymers 2023, 15, 1257. https://doi.org/10.3390/polym15051257
AMA Style
Olennikov DN, Gornostai TG. New Inonotus Polysaccharides: Characterization and Anticomplementary Activity of Inonotus rheades Mycelium Polymers. Polymers. 2023; 15(5):1257. https://doi.org/10.3390/polym15051257
Chicago/Turabian StyleOlennikov, Daniil N., and Tatyana G. Gornostai. 2023. "New Inonotus Polysaccharides: Characterization and Anticomplementary Activity of Inonotus rheades Mycelium Polymers" Polymers 15, no. 5: 1257. https://doi.org/10.3390/polym15051257
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.