Effect of Novel Penicillium verruculosum Enzyme Preparations on the Saccharification of Acid- and Alkali-Pretreated Agro-Industrial Residues

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Enzyme Preparations
2.3. Pretreatment Conditions
2.4. Saccharification Conditions
2.5. Analytical Procedures
2.5.1. Glucose Concentration
2.5.2. Reducing Sugars Concentration
2.5.3. High Performance Liquid Chromatography (HPLC) Analysis of Monosaccharides Composition
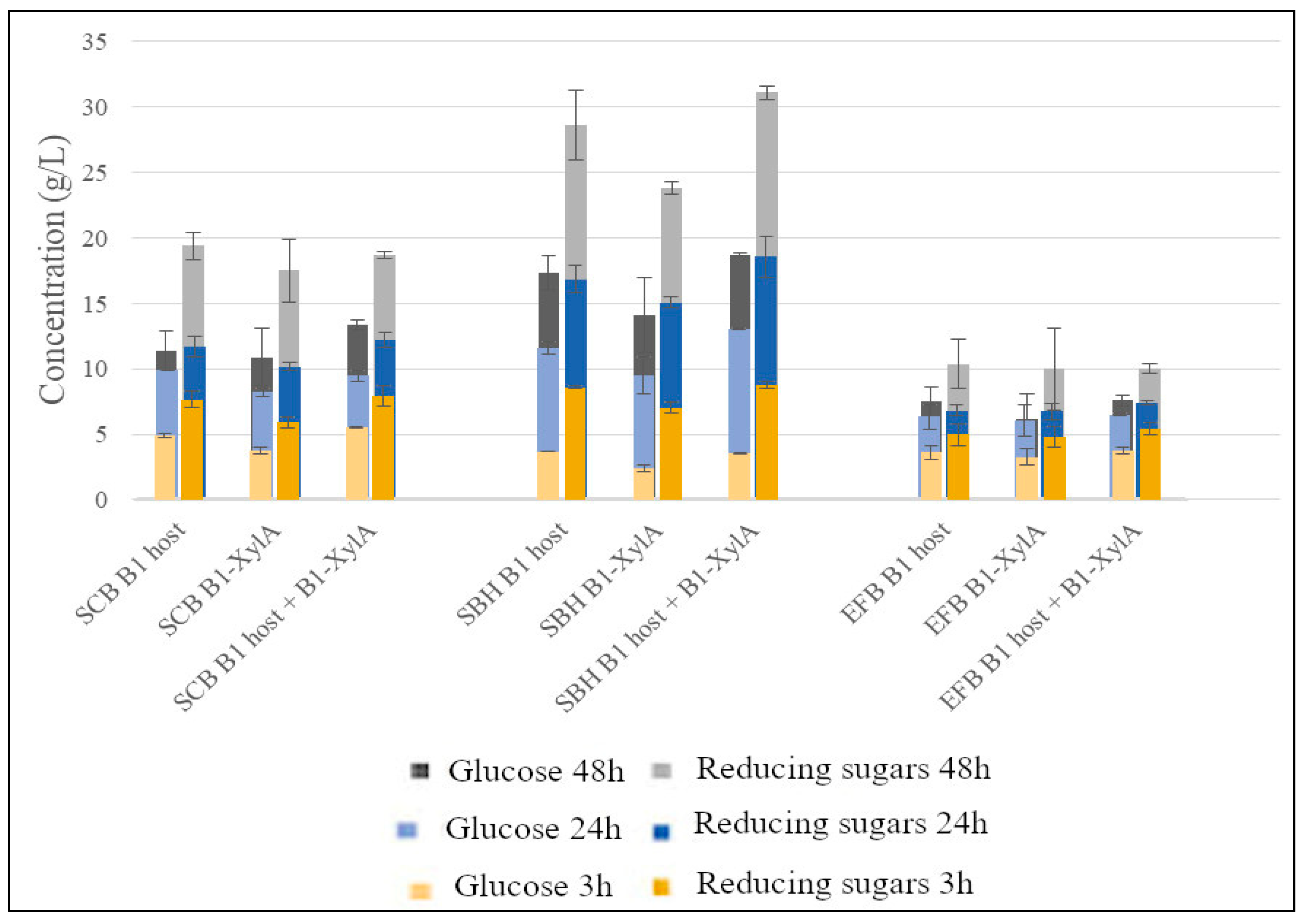
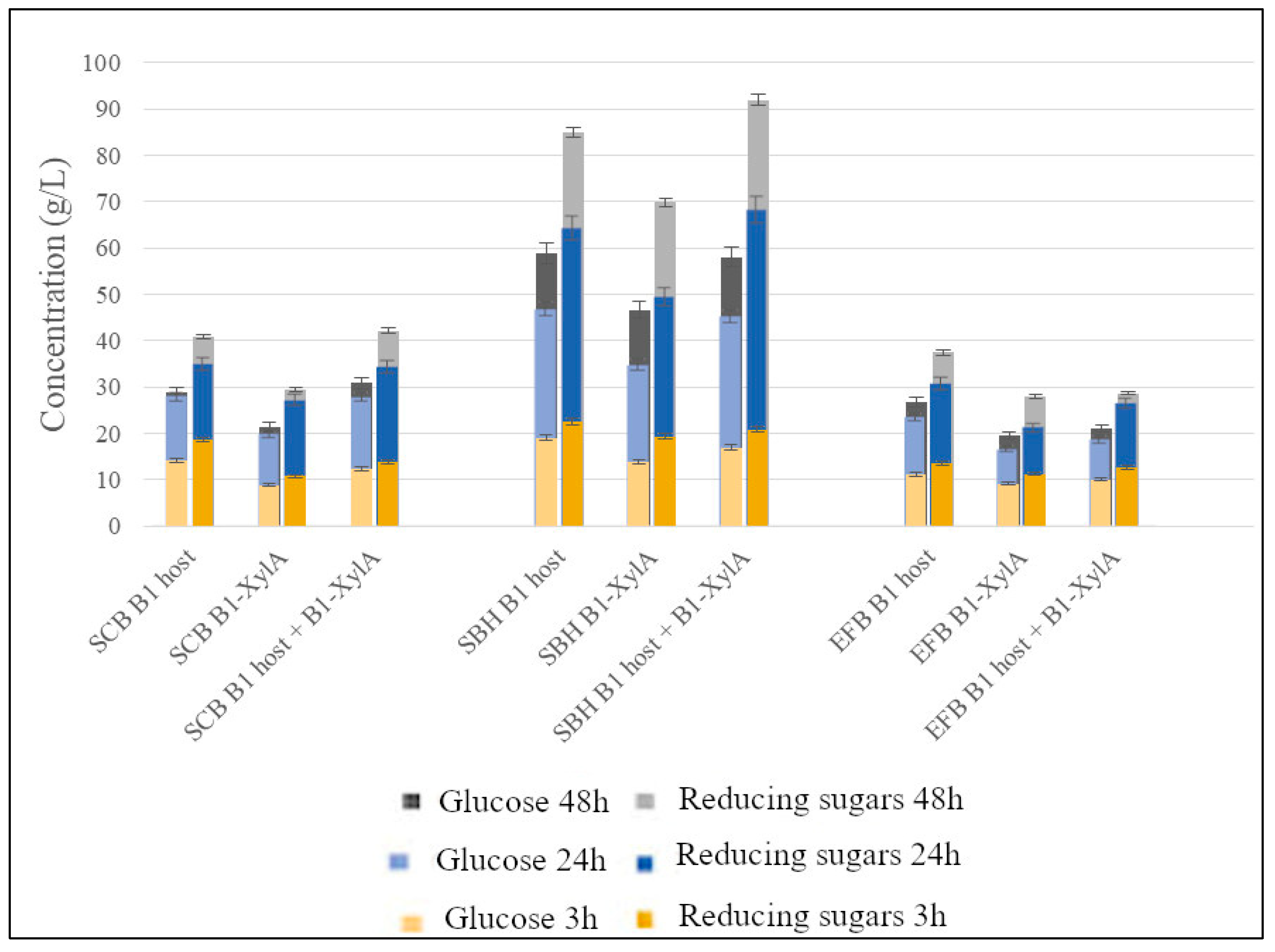
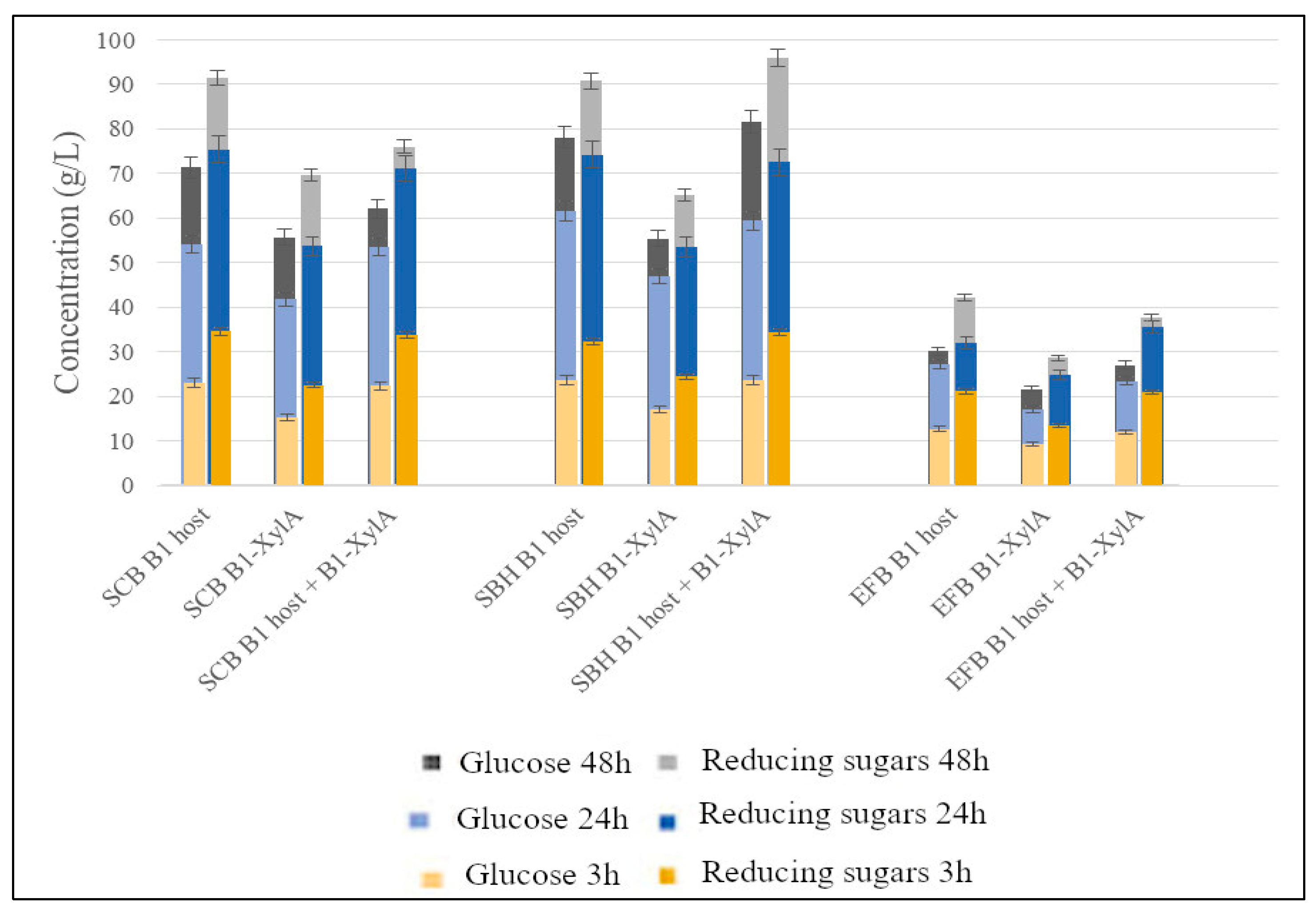
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Magalhães, A.I. Jr.; Carvalho, J.C.; Pereira, G.V.M.; Karp, S.G.; Câmara, M.C.; Medina, J.D.C.; Soccol, C.R. Lignocellulosic biomass from agro-industrial residues in South America: Current developments and perspectives. Biofuels Bioprod. Bioref. 2019, 13, 1505–1519. [Google Scholar] [CrossRef]
- Prajapati, B.P.; Jana, U.K.; Suryawanshi, R.K.; Kango, N. Sugarcane bagasse saccharification using Aspergillus tubingensis enzymatic cocktail for 2G bio-ethanol production. Renew. Energ. 2020, 152, 653–663. [Google Scholar] [CrossRef]
- Karp, S.G.; Woiciechowski, A.L.; Soccol, V.T.; Soccol, C.R. Pretreatment strategies for delignification of sugarcane bagasse: A review. Braz. Arch. Biol. Technol. 2013, 56, 679–689. [Google Scholar] [CrossRef]
- FAO–Food and Agriculture Organization of the United Nations. FAOSTAT Crops. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 May 2020).
- USDA–United States Department of Agriculture. Sugar: World Markets and Trade. Available online: https://apps.fas.usda.gov/psdonline/circulars/sugar.pdf (accessed on 10 May 2020).
- CONAB–Companhia Nacional de Abastecimento. Safra Brasileira de Cana de Açúcar. Available online: https://www.conab.gov.br/info-agro/safras/cana (accessed on 10 May 2020).
- Rodrigues, R.C.L.B.; Felipe, M.G.A.; Sil, J.B.A.; Vitolo, M. Response surface methodology for xylitol production from sugarcane bagasse hemicellulosic hydrolyzate using controlled vacuum evaporation process variables. Proc. Biochem. 2003, 38, 1231–1237. [Google Scholar] [CrossRef]
- Soccol, C.R.; Vandenberghe, L.P.S.; Medeiros, A.B.P.; Karp, S.G.; Buckeridge, M.; Ramos, L.P.; Pitarelo, A.P.; Ferreira-Leitão, V.; Gottschalk, L.M.F.; Ferrara, M.A.; et al. Bioethanol from lignocelluloses: Status and perspectives in Brazil. Bioresour. Technol. 2010, 101, 4820–4825. [Google Scholar] [CrossRef]
- USDA–United States Department of Agriculture. World Agricultural Production. Available online: https://apps.fas.usda.gov/psdonline/circulars/production.pdf (accessed on 10 May 2020).
- Karp, S.G.; Igashiyama, A.H.; Siqueira, P.F.; Carvalho, J.C.; Vandenberghe, L.P.S.; Thomaz-Soccol, V.; Coral, J.; Tholozan, J.L.; Pandey, A.; Soccol, C.R.; et al. Application of the biorefinery concept to produce L-lactic acid from the soybean vinasse at laboratory and pilot scale. Bioresour. Technol. 2011, 102, 1765–1772. [Google Scholar] [CrossRef]
- Liu, H.-M.; Li, H.-Y. Application and conversion of soybean hulls. In Soybean-The Basis of Yield, Biomass and Productivity, 1st ed.; Kasai, M., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Medina, J.D.C.; Woiciechowski, A.L.; Zandona Filho, A.; Brar, S.K.; Magalhães Júnior, A.I.; Soccol, C.R. Energetic and economic analysis of ethanol, xylitol and lignin production using oil palm empty fruit bunches from a Brazilian factory. J. Clean. Prod. 2018, 195, 44–55. [Google Scholar] [CrossRef]
- Morozova, V.V.; Gusakov, A.V.; Andrianov, R.M.; Pravilnikov, A.G.; Osipov, D.O.; Sinitsyn, A.P. Cellulase complex of the fungus Penicillium verruculosum: Properties of major endoglucanases and cellobiohydrolases. Biotechnol. J. 2010, 5, 871–880. [Google Scholar] [CrossRef]
- Gusakov, A.V.; Sinitsyn, A.P. Cellulases from Penicillium species for producing fuel from biomass. Biofuels 2012, 3, 463–477. [Google Scholar] [CrossRef]
- Osipov, D.O.; Rozhkova, A.M.; Matys, V.Y.; Koshelev, A.V.; Okunev, O.N.; Rubtsova, E.A.; Pravilnikov, A.G.; Zorov, I.N.; Oveshnikov, I.N.; Davidov, E.R.; et al. Production of biocatalysts on the basis of recombinant heterologous xylanase producer strains in the Penicillium verruculosum fungus: Their application in the hydrolysis of timber and wood processing industry wastes. Catal. Ind. 2011, 3, 34–40. [Google Scholar] [CrossRef]
- Dotsenko, G.S.; Gusakov, A.V.; Rozhkova, A.M.; Korotkova, O.G.; Sinitsyn, A.P. Heterologous beta-glucosidase in a fungal cellulase system: Comparison of different methods for development of multienzyme cocktails. Proc. Biochem. 2015, 50, 1258–1263. [Google Scholar] [CrossRef]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of sugars. J. Biol. Chem. 1944, 153, 375–379. [Google Scholar]
- Somogyi, M. A new reagent for the determination of sugars. J. Biol. Chem. 1945, 160, 61–68. [Google Scholar]
- Somogyi, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar]
- Sinitsyna, O.A.; Bukhtoyarov, E.F.; Gusakov, A.V.; Okunev, O.N.; Bekkarevitch, A.O.; Vinetsky, Y.P.; Sinitsyn, A.P. Isolation and properties of major components of Penicillium canescens extracellular enzyme complex. Biochemistry 2003, 68, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Bulakhov, A.G.; Volkov, P.V.; Rozhkova, A.M.; Gusakov, A.V.; Nemashkalov, V.A.; Sinitsyn, A.P. Using an inducible promoter of a gene encoding Penicillium verruculosum glucoamylase for production of enzyme preparations with enhanced cellulase performance. PLoS ONE 2017, 12, e0170404. [Google Scholar] [CrossRef]
- Kaputska, L.A.; Annala, A.E.; Swanson, W.C. The peroxidase-glucose oxidase system: A new method to determine glucose liberated by carbohydrate degrading soil enzymes. Plant Soil 1981, 63, 487–490. [Google Scholar] [CrossRef]
- Woiciechowski, A.L.; Dalmas Neto, C.J.; Vandenberghe, L.P.S.; Carvalho Neto, D.P.; Sydney, A.C.N.; Letti, L.A.J.; Karp, S.G.; Torres, L.A.Z.; Soccol, C.R. Lignocellulosic biomass: Acid and alkaline pretreatments and their effects on biomass recalcitrance–Conventional processing and recent advances. Bioresour. Technol. 2020, 304, 122848. [Google Scholar] [CrossRef]
- Rojas, M.J.; Siqueira, P.F.; Miranda, L.C.; Tardioli, P.W.; Giordano, R.L.C. Sequential proteolysis and cellulolytic hydrolysis of soybean hulls for oligopeptides and ethanol production. Ind. Crop. Prod. 2014, 61, 202–210. [Google Scholar] [CrossRef]
- Scarpa, J.C.P.; Marques, N.P.; Monteiro, D.A.; Martins, G.M.; de Paula, A.V.; Boscolo, M.; Silva, R.; Gomes, E.; Bocchini, D.A. Saccharification of pretreated sugarcane bagasse using enzymes solution from Pycnoporus sanguineus MCA 16 and cellulosic ethanol production. Ind. Crop. Prod. 2019, 111795. [Google Scholar] [CrossRef]
- Martin, C.; Volkov, P.V.; Rozhkova, A.M.; Puls, J.; Sinitsyn, A.P. Comparative study of the enzymatic convertibility of glycerol- and dilute acid-pretreated sugarcane bagasse using Penicillium-and Trichoderma-based cellulase preparations. Ind. Crop. Prod. 2015, 77, 382–390. [Google Scholar] [CrossRef]
- Hickert, L.R.; Cruz, M.M.; Dillon, A.J.P.; Fontana, R.C.; Rosa, C.A.; Ayub, M.A.Z. Fermentation kinetics of acid–enzymatic soybean hull hydrolysate in immobilized-cell bioreactors of Saccharomyces cerevisiae, Candida shehatae, Spathaspora arborariae, and their co-cultivations. Biochem. Eng. J. 2014, 88, 61–67. [Google Scholar] [CrossRef]
- Qing, Q.; Guo, Q.; Zhou, L.; Gao, X.; Lu, X.; Zhang, Y. Comparison of alkaline and acid pretreatments for enzymatic hydrolysis of soybean hull and soybean straw to produce fermentable sugars. Ind. Crop. Prod. 2017, 109, 391–397. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Substrate | SCB | SBH | EFB |
|---|---|---|---|
| Cellulose | 36 | 37 | 28 |
| Hemicellulose | 31 | 27 | 24 |
| Lignin | 23 | 8.7 | 20 |
| Lipids | 2.3 | 1.9 | 6.3 |
| Proteins | 2.2 | 14 | 3.4 |
| Ash | 2.7 | 4.2 | 3.2 |
| Extractives/others | 2.2 | 6.8 | 15 |
| Name | Protein, mg/g | CMC, U/g | β-Glucan, U/g | Xylan, U/g | pNPG, U/g |
|---|---|---|---|---|---|
| B1 host | 970 | 16,542 | 15,062 | 17,532 | 1074 |
| B1-XylA | 441 | 2240 | 2240 | 60,000 | 610 |
| F10 | 655 | 7007 | 6797 | 3800 | 39,852 |
| Condition | Enzyme Combinations |
|---|---|
| B1 host | B1 host 10 mg/g substrate + F10 40 U/g substrate |
| B1-XylA | B1-XylA 10 mg/g substrate + F10 40 U/g substrate |
| B1 host + B1-XylA | B1 host 8 mg/g substrate + B1-XylA 2 mg/g substrate + F10 40 U/g substrate |
| Substrate-Pretreatment | Solids Recovery in Dry Mass Basis (%) | Sugar Loss in Dry Mass Basis (%) |
|---|---|---|
| SCB—Acid | 63.6 | 24.1 |
| SCB—Alkaline | 65.2 | N. d. |
| SBH—Acid | 56.1 | 11.36 |
| SBH—Alkaline | 48.7 | N. d. |
| EFB—Acid | 63.4 | 18.14 |
| EFB—Alkaline | 67.4 | N. d. |
| Substrate-Pretreatment | Enzyme Combination | Glucose Concentration (g/L) | Reducing Sugars Concentration (g/L) | Glucose Yield (g/100 g Substrate) | Reducing Sugars Yield (g/100 g Substrate) |
|---|---|---|---|---|---|
| SCB—Untreated | B1 host, B1-XylA or B1 host + B1-XylA | 12 | 18 | 11 | 16 |
| SCB—Acid | B1 host or B1 host + B1-XylA | 30 | 40 | 17 (27) | 23 (36) |
| SCB—Alkaline | B1 host | 71 | 91 | 42 (65) | 54 (83) |
| SBH—Untreated | B1 host + B1-XylA | 19 | 31 | 17 | 28 |
| SBH—Acid | B1 host + B1-XylA | 58 | 92 | 30 (53) | 47 (84) |
| SBH—Alkaline | B1 host + B1-XylA | 81 | 96 | 36 (74) | 42 (87) |
| EFB—Untreated | B1 host or B1 host + B1-XylA | 7.5 | 10 | 6.8 | 9 |
| EFB—Acid | B1 host | 27 | 37 | 16 (25) | 21 (34) |
| EFB—Alkaline | B1 host | 30 | 42 | 18 (27) | 26 (38) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karp, S.G.; Osipov, D.O.; Semenova, M.V.; Rozhkova, A.M.; Zorov, I.N.; Sinitsyna, O.A.; Soccol, C.R.; Sinitsyn, A.P. Effect of Novel Penicillium verruculosum Enzyme Preparations on the Saccharification of Acid- and Alkali-Pretreated Agro-Industrial Residues. Agronomy 2020, 10, 1348. https://doi.org/10.3390/agronomy10091348
Karp SG, Osipov DO, Semenova MV, Rozhkova AM, Zorov IN, Sinitsyna OA, Soccol CR, Sinitsyn AP. Effect of Novel Penicillium verruculosum Enzyme Preparations on the Saccharification of Acid- and Alkali-Pretreated Agro-Industrial Residues. Agronomy. 2020; 10(9):1348. https://doi.org/10.3390/agronomy10091348
Chicago/Turabian StyleKarp, Susan G., Dmitrii O. Osipov, Margarita V. Semenova, Alexandra M. Rozhkova, Ivan N. Zorov, Olga A. Sinitsyna, Carlos R. Soccol, and Arkady P. Sinitsyn. 2020. "Effect of Novel Penicillium verruculosum Enzyme Preparations on the Saccharification of Acid- and Alkali-Pretreated Agro-Industrial Residues" Agronomy 10, no. 9: 1348. https://doi.org/10.3390/agronomy10091348