Identification of Black Rot Resistance in a Wild Brassica Species and Its Potential Transferability to Cauliflower

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Inoculation and Disease Resistance Assay
2.3. Cross-Compatibility Survey and F1 Production
2.4. F1 Hybrid Identification
2.5. Inoculation of Parents and F1, and F2 Generations
2.6. Data Analysis
3. Results
3.1. Investigation of Resistance to Xcc race 4 in Cauliflower and Related Wild Accessions
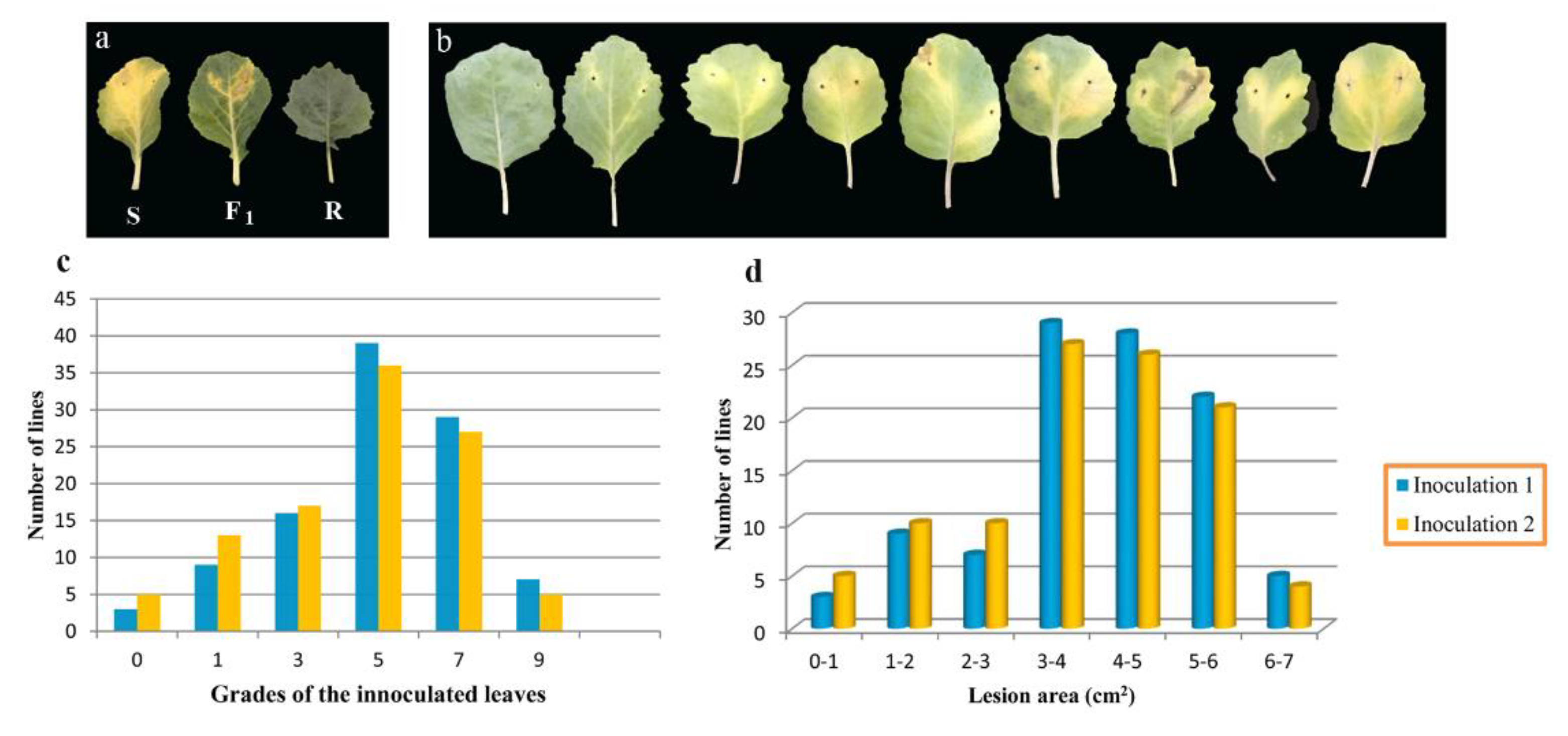
3.1.1. Investigation Based on the Traditional Grading Method
3.1.2. Investigation Based on the PPMS
3.2. Cross-Compatibility Survey and F1 production
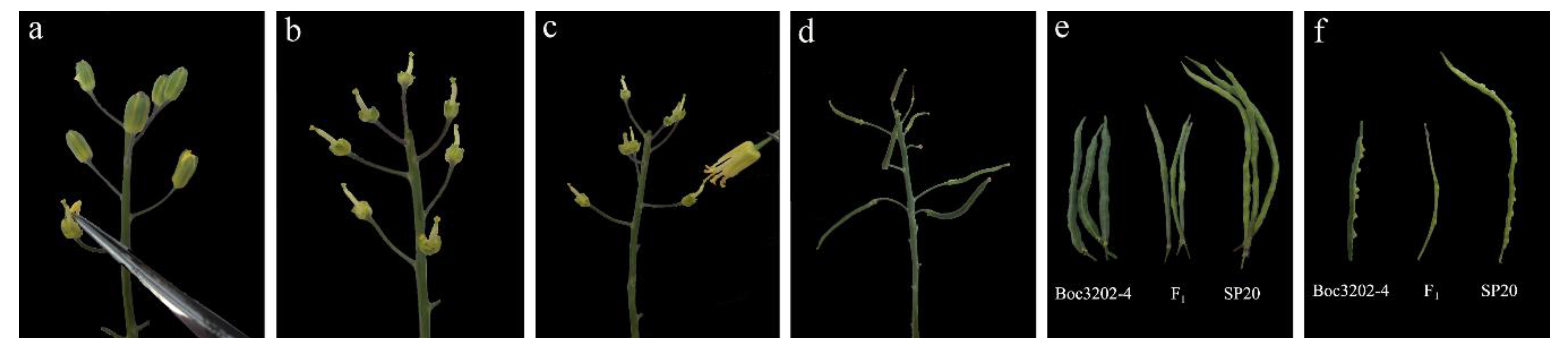
3.3. F1 Hybrid Identification
3.4. Inoculation of Parents, F1 and F2 Generations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keck, A.; Finley, J. Cruciferous vegetables: Cancer protective mechanisms of glucosinalate hydrolysis products and selenium. Integr. Cancer Ther. 2004, 3, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalik, A.; Pascual, B.; Nájera, I.; Baixauli, C.; Pascual-Seva, N. Deficit irrigation as a sustainable practice in improving irrigation water use efficiency in cauliflower under mediterranean conditions. Agronomy 2019, 9, 732. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Thiele, B.; Santhiraraja-Abresch, S.; Watt, M.; Kraska, T.; Ulbrich, A.; Kuhn, A. Effects of Root Temperature on the Plant Growth and Food Quality of Chinese Broccoli (Brassica oleracea var. alboglabra Bailey). Agronomy 2020, 10, 702. [Google Scholar] [CrossRef]
- Williams, P.H. Black rot: A continuing threat to world crucifers. Plant Dis. 1980, 64, 736–742. [Google Scholar] [CrossRef]
- Vicente, J.G.; Holub, E.B. Xanthomonas campestris pv. campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to Brassica crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef]
- Bella, P.; Moretti, C.; Strano, C.P.; Zaccardelli, M.; Branca, F.; Buonaurio, R.; Vincente, J.C.; Catara, V. Race typing and molecular characterization of Xanthomonas campestris pv campestris strains occuring in Italy. J. Plant Pathol. 2015, 97, S17. [Google Scholar]
- Bella, P.; Moretti, C.; Licciardello, G.; Branca, F.; Vincente, J.G.; Catara, V. Multilocus sequence typing analysis of Italian Xanthomonas campestris pv. Campestris strains suggested the evolution of local endemic populations of the pathogen and does not correlate with rce distribution. Plant Pathol. 2019, 68, 278–287. [Google Scholar] [CrossRef]
- Ragasová, L.; Penázová, E.; Gazdík, F.; Pecenka, J.; Cechová, J.; Pokluda, R.; Eichmeier, A. The Change of Bacterial Spectrum after Storage of X. campestris pv. campestris Inoculated Cabbage Heads (Brassica oleracea var. capitata L.). Agronomy 2020, 10, 443. [Google Scholar]
- Tonguc, M.; Griffiths, P.D. Evaluation of Brassica carinata accessions for resistance to black rot (Xanthomonas campestris pv. campestris). HortScience 2004, 39, 952–954. [Google Scholar] [CrossRef]
- Vicente, J.G.; Conway, J.; Roberts, S.J. Identification and origin of Xanthomonas campestris pv. campestris races and related pathovars. Phytopathology 2001, 91, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Fargier, E.; Manceau, C. Pathogenicity assays restrict the species Xanthomonas campestris into three pathovars and reveal nine races within X. campestris pv. campestris. Plant Pathol. 2007, 56, 805–818. [Google Scholar] [CrossRef]
- Vicente, J.G.; Tayor, J.D.; Sharpe, A.G. Inheritance of race-specific resistance to Xanthomonas campestris pv. campestris in Brassica genomes. Phytopathology 2002, 92, 1134–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westman, A.L.; Kresovich, S.; Dickson, M.H. Regional variation in Brassica nigra and other weedy crucifers for disease reaction to Alternaria brassicicola and Xanthomonas campestris pv. campestris. Euphytica 1999, 106, 253–259. [Google Scholar] [CrossRef]
- Taylor, J.D.; Conway, J.; Roberts, S.J.; Astley, D.; Vicente, J.G. Sources and origin of resistance to Xanthomonas campestris pv. campestris in Brassica genomes. Phytopathology 2002, 92, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, B.B.; Kalia, P.; Yadava, D.K.; Singh, D.; Sharma, T.R. Genetics and molecular mapping of black rot resistance locus Xca1bc on chromosome B-7 in Ethiopian Mustard (Brassica carinata A. Braun). PLoS ONE 2016, 11, e0152290. [Google Scholar] [CrossRef]
- Sharma, S.R.; Kapoor, K.S.; Gill, H.S. Screening against sclerotinia rot (Sclerotinia sclerotiorum), downy mildew (Peronospora parasitica) and black rot (Xanthomonas campestris) in cauliflower (Brassica oleracea var. botrytis subvar cauliflora. Indian J. Agri. Sci. 1995, 65, 916–918. [Google Scholar]
- Griesbach, E.; Loptien, H.; Miersch, U. Resistance to Xanthomonas campestris pv. campestris (Pammel) Dowson in cabbage Brassica oleracea L. J. Plant Dis. Prot. 2003, 110, 461–475. [Google Scholar] [CrossRef]
- Jensen, B.D.; Massomo, S.M.; Ignas, S.S.; Hockenhull, J.; Andersen, S.B. Field evaluation for resistance to the black rot pathogen Xanthomonas campestris pv. campestris in cabbage (Brassica oleracea). Eur. J. Plant Pathol. 2005, 113, 297–308. [Google Scholar] [CrossRef]
- Saha, P.; Kalia, P.; Sharma, M.; Singh, D. New source of black rot disease resistance in Brassica oleracea and genetic analysis of resistance. Euphytica 2016, 207, 35–48. [Google Scholar] [CrossRef]
- Mei, J.Q.; Liu, Y.; Wei, D.Y.; Wittkop, B.; Ding, Y.J.; Li, Q.F.; Li, J.N.; Wan, H.F.; Li, Z.Y.; Ge, X.H. Transfer of sclerotinia resistance from wild relative of Brassica oleracea into Brassica napus using a hexaploidy step. Theor. Appl. Genet. 2015, 128, 639–644. [Google Scholar] [CrossRef]
- Wang, G.X.; Tang, Y.; Yan, H.; Sheng, X.G.; Hao, W.W.; Zhang, L.; Lu, K.; Liu, F. Production and characterization of interspecific somatic hybrids between Brassica oleracea var. botrytis and B. nigra and their progenies for the selection of advanced pre-breeding materials. Plant Cell Rep. 2011, 30, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Rvon, B.; Gustafsson, M.; Snogerup, S. Brassica sect. Brassica (Brassicaceae). II. Inter- and intraspecific crosses with cultivars of B. oleracea. Genet. Resour. Crop Evol. 1995, 42, 165–178. [Google Scholar] [CrossRef]
- Singh, R.; Ellis, P.R.; Pink, A.C.; Phelps, K. An investigation of the resistance to cabbage aphid in Brassica species. Ann. Appl. Biol. 2010, 125, 457–465. [Google Scholar] [CrossRef]
- Happstadius, I.; Ljungberg, A.; Kristiansson, B.; Dixelius, C. Identification of Brassica oleracea germplasm with improved resistance to Verticillium wilt. Plant Breed. 2010, 122, 30–34. [Google Scholar] [CrossRef]
- Ding, Y.; Mei, J.; Liu, Y.; Wang, L.; Li, Y.; Wan, H.; Li, J.; Qian, W. Transfer of sclerotinia stem rot resistance from wild Brassica oleracea into B. rapa. Mol. Breed. 2015, 35, 225–229. [Google Scholar] [CrossRef]
- Mithen, R.F.; Lewis, B.G.; Heaney, R.K.; Fenwick, G.R. Resistance of leaves of Brassica species to Leptosphaeria maculans. Trans. Br. Mycol. Soc. 1987, 88, 525–531. [Google Scholar] [CrossRef]
- Mithen, R.F.; Lewis, B.G.; Heaney, R.K.; Fenwick, G.R. Glucosinolates of wild and cultivated Brassica species. Phytochemistry 1987, 26, 1969–1973. [Google Scholar]
- Bradburne, R.; Majer, D.; Magrath, R.; Werner, C.P.; Lewis, B.; Mithen, R. Winter oilseed rape with high levels of resistance to Pyrenopeziza brassicae derived from wild Brassica species. Plant Pathol. 1999, 48, 550–558. [Google Scholar] [CrossRef]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgenson, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley. Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.Q.; Gu, H.H.; Sheng, X.G.; Yu, H.F.; Wang, J.S.; Cao, J.S. Genetic diversity and relationships among loose-curd cauliflower and related varieties as revealed by microsatellite markers. Sci. Hortic. 2014, 166, 105–110. [Google Scholar] [CrossRef]
- Panaud, O.; Chen, X.; McCouch, S.R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol. Gen. Genet. 1996, 252, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Soengas, P.; Hand, P.; Vicente, J.G.; Pole, J.M.; Pink, A.C. Identification of quantitative trait loci for resistance to Xanthomonas campestris pv. campestris in Brassica rapa. Theor. Appl. Genet. 2007, 114, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Dickson, M.H.; Hunter, J.E. Brassica napus sources of resistance to black rot of crucifers and inheritance of resistance. HortScience 1991, 26, 1545–1547. [Google Scholar] [CrossRef]
- Hunter, J.E.; Dickson, M.H.; Ludwig, J.W. Source of resistance to black rot of cabbage expressed in seedlings and adult plants. Plant Dis. 1987, 71, 263–266. [Google Scholar] [CrossRef]
- Bain, D. Reaction of Brassica seedlings to black rot. Phytopathology 1952, 42, 316–319. [Google Scholar]
- Jamwal, R.S.; Sharma, P.P. Inheritance of resistance to black rot (Xanthomonas campestris pv. campestris) in cauliflower (Brassica oleracea var. botrytis). Euphytica 1986, 35, 941–943. [Google Scholar] [CrossRef]
- Saha, P.; Kalia, P.; Sonah, H.; Sharma, T. Molecular mapping of black rot resistance locus Xca1bo on chromosome 3 in Indian cauliflower (Brassica oleracea var. botrytis L.). Plant Breed. 2014, 133, 268–274. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name | Species | Type | DI Score | Resistance | AIP (cm2) | PTL (%) |
|---|---|---|---|---|---|---|---|
| 1 | SH-80 | Cauliflower | F1 | 33.8 ± 2.2 p | IR | 1.4 ± 0.2 op | 14.4 ± 2.5 |
| 2 | SH-88 | Cauliflower | F1 | 45.7 ± 2.8 klm | IR | 1.8 ± 0.3 n | 17.9 ± 2.2 |
| 3 | QN-65 | Cauliflower | F1 | 50.6 ± 2.5 hij | S | 4.3 ± 0.3 def | 40.2 ± 2.8 |
| 4 | QN-80 | Cauliflower | F1 | 43.8 ± 2.4 lm | IR | 2.9 ± 0.2 k | 28.0 ± 2.0 |
| 5 | QN-90 | Cauliflower | F1 | 37.8 ± 2.3 o | IR | 1.6 ± 0.2 no | 15.9 ± 2.1 |
| 6 | ZS-50 | Cauliflower | F1 | 56.1 ± 1.2 fg | S | 4.7 ± 0.2 c | 46.3 ± 2.1 |
| 7 | ZS-60 | Cauliflower | F1 | 45.7 ± 1.5 klm | IR | 3.1 ± 0.1 j | 32.7 ± 1.1 |
| 8 | BY-80 | Cauliflower | F1 | 33.9 ± 2.9 p | IR | 1.4 ± 0.1 op | 14.3 ± 1.4 |
| 9 | Boc3202-4 | Cauliflower | DH | 83.2 ± 2.5 a | HS | 5.8 ± 0.3 a | 53.1 ± 2.8 |
| 10 | Boc3226-4A | Cauliflower | DH | 56.9 ± 2.3 fg | S | 4.5 ± 0.3 cd | 46.1 ± 3.0 |
| 11 | Boc3206-1 | Cauliflower | DH | 41.7 ± 2.2 mn | IR | 2.2 ± 0.3 m | 21.2 ± 2.9 |
| 12 | Boc3206-4 | Cauliflower | DH | 58.5 ± 3.1 ef | S | 4.2 ± 0.4 efg | 45.3 ± 4.4 |
| 13 | Boc3225-21 | Cauliflower | DH | 39.3 ± 1.8 no | IR | 1.7 ± 0.1 n | 17.2 ± 1.1 |
| 14 | Boc3201-1 | Cauliflower | DH | 47.2 ± 3.2 jkl | IR | 3.4 ± 0.3 ij | 24.9 ± 2.3 |
| 15 | Boc3005-1 | Cauliflower | DH | 43.7 ± 2.4 lm | IR | 2.6 ± 0.2 l | 28.4 ± 2.2 |
| 16 | Boc3228-4 | Cauliflower | DH | 76.4 ± 3.1 b | HS | 5.3 ± 0.4 b | 52.8 ± 4.1 |
| 17 | Boc3203-4 | Cauliflower | DH | 54.3 ± 4.1 gh | S | 3.9 ± 0.4 h | 42.1 ± 4.4 |
| 18 | Boc4259 | Cauliflower | SI | 65.5 ± 3.1 c | S | 4.6 ± 0.3 cd | 44.5 ± 2.9 |
| 19 | Boc4258 | Cauliflower | SI | 64.3±2.6 cd | S | 4.5 ± 0.3 cde | 42.2 ± 2.8 |
| 20 | Boc4222-3 | Cauliflower | SI | 61.4 ± 3.3 de | S | 5.0 ± 0.3 b | 51.4 ± 2.9 |
| 21 | Boc4229 | Cauliflower | SI | 56.8 ± 1.9 fg | S | 4.4 ± 0.2 cdef | 45.2±2.1 |
| 22 | Boc4710-1 | Cauliflower | SI | 53.8 ± 2.3 gh | S | 4.3 ± 0.2 def | 43.3 ± 1.9 |
| 23 | Boc4601 | Cauliflower | SI | 22.9 ± 1.3 q | R | 1.0 ± 0.1 q | 12.2 ± 1.2 |
| 24 | Boc4604 | Cauliflower | SI | 65.3 ± 1.3 cd | S | 5.2 ± 0.2 b | 51.1 ± 2.2 |
| 25 | Boc4605 | Cauliflower | SI | 51.6 ± 3.0 hi | S | 3.9 ± 0.4 gh | 39.9 ± 3.0 |
| 26 | Boc4251 | Cauliflower | SI | 48.8 ± 2.5 ijk | IR | 3.5 ± 0.3 i | 35.8 ± 2.7 |
| 27 | PI435896 | Brassica balearica | WD | 11.2 ± 0.6 r | R | 0.5 ± 0.08 r | 4.6 ± 0.9 |
| 28 | PI662587 | Brassica insularis | WD | 38.8 ± 2.6 no | IR | 1.7 ± 0.2 n | 18.6 ± 2.3 |
| 29 | UNICT4785 | Brassica macrocarp | WD | 42.4 ± 1.8 mn | IR | 4.2 ± 0.2 fg | 42.9 ± 2.1 |
| 30 | UNICT5168 | Brassica villosa | WD | 26.0 ± 1.7 q | R | 1.2 ± 0.1 pq | 11.3 ± 1.1 |
| 31 | UNICT3512 | Brassica incana | WD | 63.7 ± 2.4 cd | S | 5.2 ± 0.3 b | 44.5 ± 2.6 |
| 32 | UNICT5169 | Brassica montana | WD | 7.8 ± 0.5 r | HR | 0.3 ± 0.06 r | 2.9 ± 1.2 |
| DI | AIP | PTL | I2-DI | I2-AIP | |
|---|---|---|---|---|---|
| DI | / | 0.93 ** | 0.91 ** | / | / |
| AIP | 0.93 ** | / | 0.98 ** | / | / |
| PTL | 0.91 ** | 0.98 ** | / | / | / |
| I1-DI | / | / | / | 0.97 ** | / |
| I1-AIP | / | / | / | / | 0.98 ** |
| Accessions | Type | DI | AIP |
|---|---|---|---|
| Boc3202-4 | female parent | 80.8 a | 5.6 a |
| UNICT5169 | male parent | 8.1 d | 0.3 d |
| Mid-parent value | / | 44.4 b | 2.9 b |
| F1 | F1 hybrids | 39.4 c | 1.9 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, X.-G.; Branca, F.; Zhao, Z.-Q.; Wang, J.-S.; Yu, H.-F.; Shen, Y.-S.; Gu, H.-H. Identification of Black Rot Resistance in a Wild Brassica Species and Its Potential Transferability to Cauliflower. Agronomy 2020, 10, 1400. https://doi.org/10.3390/agronomy10091400
Sheng X-G, Branca F, Zhao Z-Q, Wang J-S, Yu H-F, Shen Y-S, Gu H-H. Identification of Black Rot Resistance in a Wild Brassica Species and Its Potential Transferability to Cauliflower. Agronomy. 2020; 10(9):1400. https://doi.org/10.3390/agronomy10091400
Chicago/Turabian StyleSheng, Xiao-Guang, Ferdinando Branca, Zhen-Qing Zhao, Jian-Sheng Wang, Hui-Fang Yu, Yu-Sen Shen, and Hong-Hui Gu. 2020. "Identification of Black Rot Resistance in a Wild Brassica Species and Its Potential Transferability to Cauliflower" Agronomy 10, no. 9: 1400. https://doi.org/10.3390/agronomy10091400