
Poriella subacida Gen. & Comb Nov. for Perenniporia subacida (Peck) Donk



1
Key Laboratory for Forest Resources Conservation and Utilization in the Southwest Mountains of China, Ministry of Education, Southwest Forestry University, Kunming 650224, China
2
College of Biodiversity Conservation, Southwest Forestry University, Kunming 650224, China
3
CAS Key Laboratory for Plant Biodiversity and Biogeography of East Asia (KLPB), Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(7), 1308; https://doi.org/10.3390/agronomy11071308
Submission received: 11 May 2021
/
Revised: 20 June 2021
/
Accepted: 22 June 2021
/
Published: 27 June 2021
(This article belongs to the Special Issue Research Progress on Added-Value Fungi)
Abstract
:Poriella subacida gen. & comb. nov., previously known as Perenniporia subacida, which causes white rot, has been documented in temperate and tropical forests. Specimens from Asia, North America, and Europe were examined, including the type specimen of Polylorus subacidus. Sequences of the ITS1-5.8S-ITS2 region, the 28S rDNA, the mitochondrial rDNA small subunit (mtSSU), and the gene encoding the translation elongation factor 1-α (EF1) were generated. In multigene phylogenies (maximum parsimony, maximum likelihood, Bayesian inferences), “Perenniporia subacida” formed a well-supported lineage, distinct from the core “Perenniporia” clade (type species: “P. medulla-panis”), and sister to the “Yuchengia narymica” lineage. We therefore conclude that “P. subacida” should be placed in the new genus “Poriella”gen. nov. Morphologically, “Poriella” is characterized by a di- to trimitic hyphal system, non-truncate basidiospores, and strongly dextrinoid, cyanophilic skeletal hyphae.
1. Introduction
Polyporales is one of the most intensively studied clades of fungi, being of interest to both fungal ecologists and applied scientists [1,2]. The family Polyporaceae accommodates 92 genera [3], and recently many new genera of polypores have been described, such as Amylosporia (B.K. Cui, C.L. Zhao & Y.C. Dai), Crystallicutis (El-Gharabawy and Griffith), Hirticrusta Matozaki (T. Hatt. and Sotome), and Murinicarpus (B.K. Cui and Y.C. Dai) [4,5,6,7]. The family is typified by Polyporus P. Micheli ex Adans. (1763) and many taxa have been separated from Polyporus as new genera over time [1,4,6].
Perenniporia subacida (Peck) Donk (basionym Polyporus subacidus Peck, 1885) is a widespread species of wood-decaying polypores with perennial, resupinate to effused-reflex basidiomes and thick-walled, non-truncate basidiospores. The species has been reported to occur in many forest ecosystems, in boreal, temperate, subtropical, and tropical regions [8,9,10,11,12,13,14,15]. This species was described by Peck (1885) as Polyporus subacidus, but its taxonomic position has long been debated. It has been treated in several genera, but none of these placements have been fully satisfactory [16,17,18,19,20]. Currently, the species is widely accepted in Perenniporia Murrill, which is typed by P. medulla-panis (Jacq.) Donk [8,9,10,11,12,13]. However, Decock and Stalpers argued for a different placement [13].
Molecular studies involving Polyporaceae, mainly based on ITS and/or nLSU sequences, have been carried out [21,22,23,24,25,26,27]. Further studies employing a six-gene dataset have helped to clarify the generic relationships of polyporoid fungi for 373 taxa. The latter study showed that Perenniporia subacida clustered in the core polyporoid clade in which it was related to P. medulla-panis (Jacq.) Donk [23]. Further studies of Perenniporia inferred from nuclear ribosomal 28S and ITS sequence data revealed that P. subacida formed a monophyletic lineage distant from the Perenniporia s.s. [15,16,28].
To resolve the placement of this species, phylogenetic research was carried out employing the ITS, 28S, TEF1 and mtSSU and a comparative morphological study of the type specimen was conducted. We conclude that Perenniporia subacida should be treated as a distinct, new genus as described below.
2. Materials and Methods
2.1. Morphological Studies
Specimens studied are deposited at the Farlow Herbarium of Harvard University (FH), MA, USA, Beijing Forestry University (BJFC), Beijing, China and United States National Fungus Collections (BPI), New York, NY, USA. Macromorphological descriptions were on the basis of field notes and study of specimens. Color terms followed previous studies [29]. Microscopic measurements were made from slide preparations of dried specimens stained with Cotton Blue and Melzer’s reagent by light microscopy [30,31]. Sections were studied using an Olympus BX40 compound microscope (Tokyo, Japan). In presenting spore size variation, 5% of measurements were excluded from each end of the range. The following abbreviations are used: KOH = 5% potassium hydroxide, CB = Cotton Blue, CB+ = cyanophilous; IKI = Melzer’s reagent, IKI– = both non-amyloid and non-dextrinoid, L = mean spore length (arithmetic average of all spores), W = mean spore width (arithmetic average of all spores), Q = mean each spore length/width ratio, n (a/b) = number of spores (a) measured from given number of specimens (b).
2.2. Molecular Techniques and Phylogenetic Analyses
CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to extract DNA from dried specimens from China and other regions, and 2xTaq PCR Mix (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to perform PCR products [30]. The primer pair ITS5 and ITS4 were amplified for the ITS region [32]. The primer pair LR0R and LR7 were for the nuclear r28S region [33]. The primer pair MS1 and MS2 were for the mitochondrial SSU region [32]. The primer pair EF1-983F and EF1-2218R were for Tef1 [34]. The PCR cycling program for ITS, mtSSU, 28S and TEF1 followed previous studies [27]. The Beijing Genomics Institute (Beijing, China) was in charge of purifying and direct sequencing of the PCR products. All sequences used in the phylogeny were downloaded from GenBank (Table 1) with references. The sequence alignment was deposited in TreeBase (submission ID 23826).
A previous study [30] was followed for maximum parsimony analysis. The combined multiple genes dataset was analyzed under heuristic search and 1000 homogeneity replicates, giving a P value of 1.000, which is much greater than the 0.01 used in PAUP* version 4.0b10, which means there is no discrepancy among the four loci in reconstructing phylogenetic trees. Trees were constructed in PAUP* version 4.0b10 [44]. All characters were equally weighted and gaps were treated as missing data. Trees were inferred using the heuristic search option with TBR branch swapping and 1000 bootstraps. Max-trees were set to 5000, branches of zero length were collapsed, and all parsimonious trees were saved. Clade robustness was assessed using a bootstrap (BT) analysis with 1000 replicates [45]. DNA sequence data was also analyzed using Maximum Likelihood (ML) with RAxML-HPC2 on Abe through the Cipres Science Gateway [46], with default settings except that branch support was obtained with 1000 rapid bootstrap replicates.
Mr Modeltest 2.3 was used to estimate the best-fit evolution model for each data set for Bayesian inference (BI). The best fit models were general time reversible (GTR)+G for ITS, and general time reversible (GTR)+I+G for nr28S, mtSSU, the exons of Tef1, and the combined dataset. The partitioned mixed model, which allows for model parameters to be estimated separately for each genetic marker, was used in the Bayesian analysis. BI was performed using MrBayes 3.1.2 [47,48,49]. Two runs of four Markov chains were run from random starting trees for two datasets: (ITS+28S) dataset for 5 million generations and (ITS+28S+mtSSU+Tef1) dataset for 10 million generations. Trees were sampled every 100 generations. The first quarter of the generations was by default discarded as burn-in. A majority rule consensus tree of all remaining trees was calculated. Branches that received bootstrap support for maximum likelihood (ML-BS), maximum parsimony (MP-BT), and Bayesian posterior probabilities (BPP) greater than or equal to 75% (ML-BS and MP-BS) and 0.95 (BPP) were considered significantly supported.
3. Results
3.1. Molecular Phylogeny
The ITS+28S dataset included sequences from 63 fungal specimens representing 56 taxa. The dataset had an aligned length of 2112 characters, of which 1216 characters are constant, 256 are variable and parsimony-uninformative, and 640 are parsimony-informative. Maximum parsimony analysis yielded 12 equally parsimonious trees (TL = 4377, CI = 0.337, HI = 0.664, RI = 0.525, RC = 0.177). The best model for the ITS+28S dataset estimated and applied in the Bayesian analysis is general time reversible (GTR)+I+G, lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1). Bayesian analysis and ML analysis resulted in a similar topology to the MP analysis, with an average standard deviation of split frequencies = 0.005569 (BI).
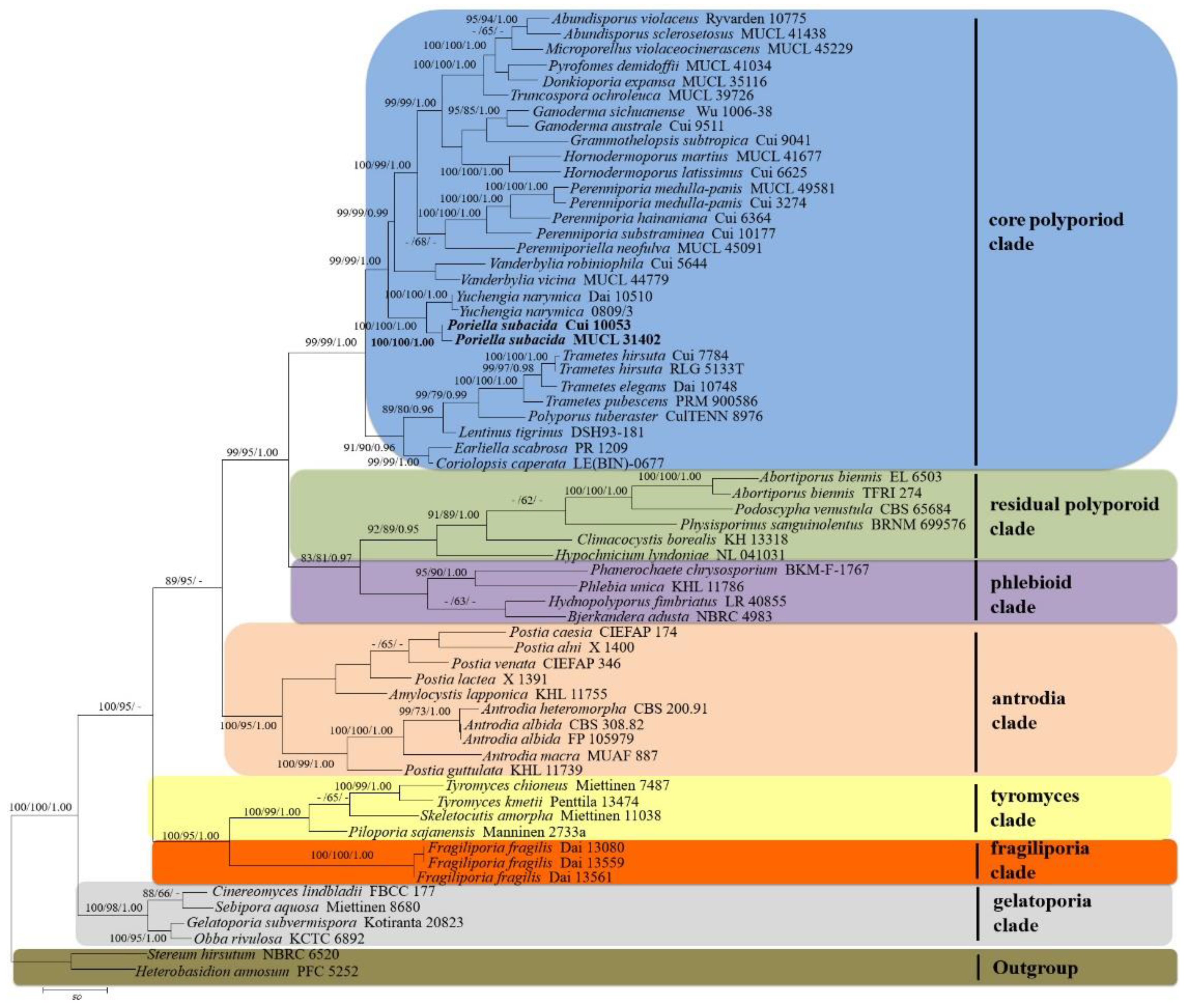
The phylogeny (Figure 1) inferred from ITS+28S sequences resolves seven major clades for 56 species of the Polyporales. Collections of Perenniporia subacida formed a lineage distinct from the Perenniporia s.s. lineage, within the core polyporoid clade. The P. subacida lineage is closely related to the Yuchengia lineage based on Y. narymica (Pilát) (B.K. Cui, C.L. Zhao, and Steffen) with good support (100% BS, 100% BP, 1.00 BPP).
The four gene (ITS+28S+mtSSU+Tef1) sequence dataset did not show any conflicts in tree topology for the reciprocal bootstrap trees, which allowed us to combine them. The combined dataset included sequences from 61 specimens representing 29 species. The dataset had an aligned length of 3515 characters, of which 2489 characters are constant, 219 are variable and parsimony-uninformative, and 807 are parsimony-informative. Maximum parsimony analysis yielded 10 equally parsimonious trees (TL = 3075, CI = 0.498, HI = 0.502, RI = 0.747, RC = 0.372). Best model for the combined ITS+28S+mtSSU+Tef1 estimated and applied in the Bayesian analysis: general time reversible (GTR)+I+G, lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1). Bayesian analysis and ML analyses resulted in a topology similar to the MP analysis, with an average standard deviation of split frequencies = 0.003420.
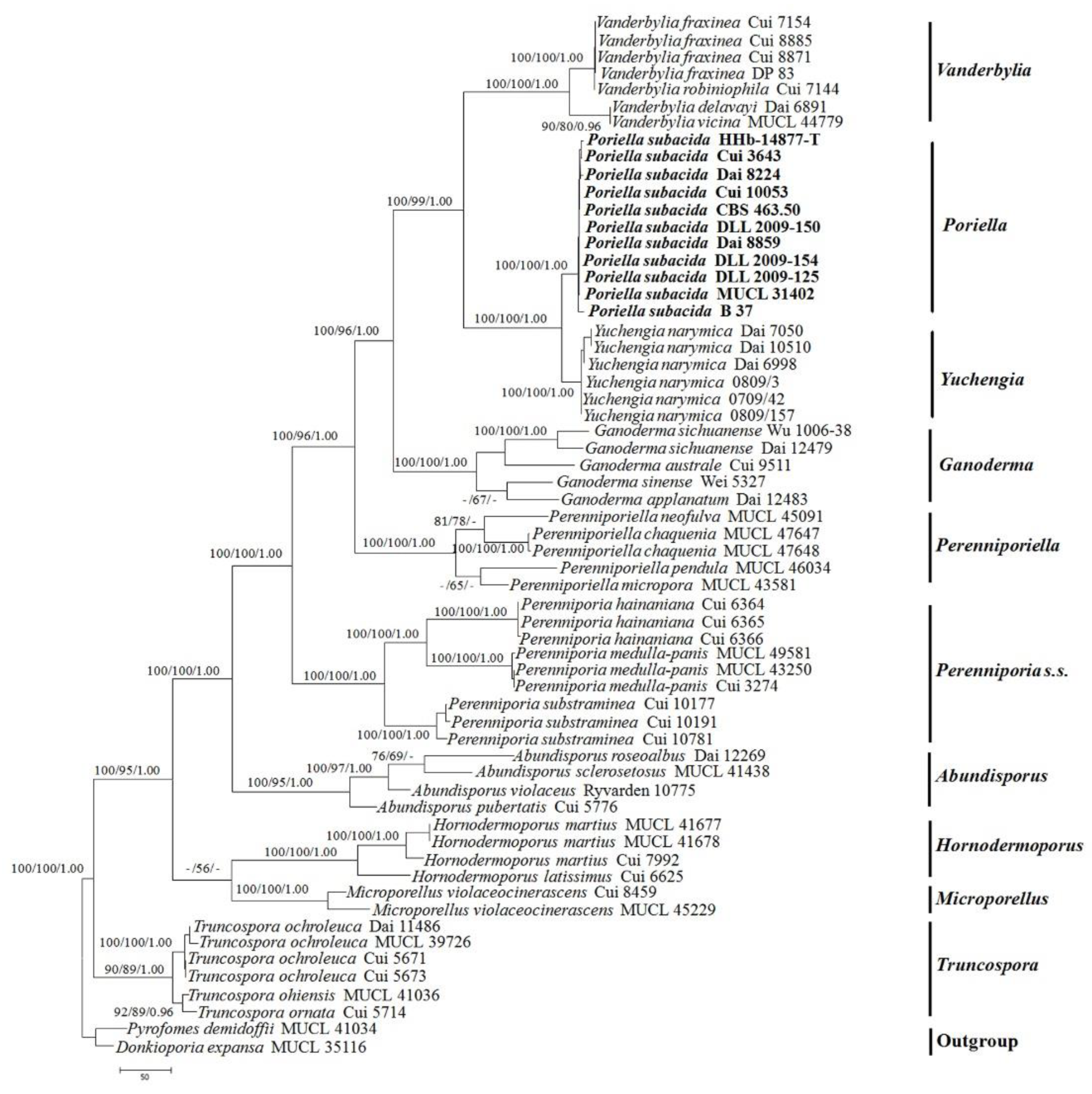
A further phylogeny (Figure 2) inferred from the combined ITS+28S+mtSSU+Tef1 sequences was obtained for ten genera in Perenniporia s.l. and demonstrated that this taxon formed a clade together with Y. narymica, with strong support (100% BS, 100% BP, 1.00 BPP). The clade is distinct from P. medulla-panis (Jacq.) We, along with Donk, conclude that it belongs to a distinct, new genus, hereafter called Poriella.
3.2. Taxonomy
Poriella C.L. Zhao, gen. nov.
MycoBank: 840061.
It is characterized by producing resupinate to effused-reflex basidiomata with dingy-yellowish to pale tan to ochraceous surface and a di-trimitic hyphal system with unbranched and strongly dextrinoid skeletal hyphae, and thick-walled, non-dextrinoid, cyanophilous basidiospores.
Type species: Poriella subacida (Peck) C.L. Zhao.
Etymology: Poriella (Lat.): referring to its similar appearance to the genus Perenniporia.
Basidiomata perennial, resupinate to effused-reflex. Pore surface is dingy-yellowish to pale tan to ochraceous. Pores round to angular. Subiculum cream to buff, thin. Tubes concolorous with pore surface, corky. Hyphal system di-trimitic, generative hyphae hyaline, thin-walled, with clamp connections; skeletal hyphae predominant, unbranched, strongly dextrinoid, cyanophilous, rarely dissolving in KOH. Basidiospores ellipsoid, non-truncate, hyaline, thick-walled, smooth, non-dextrinoid, CB+.
- MycoBank: 840062.
- Basionym: Polyporus subacidus Peck, Ann. Rep. N.Y. St. Mus. nat. Hist. 38: 92, 1885.
- =Poria subacida (Peck) Sacc., Syll. fung. (Abellini) 6: 325 (1888).
- =Chaetoporus subacidus (Peck) Bondartsev & Singer, Annls mycol. 39(1): 51 (1941).
- =Oxyporus subacidus (Peck) Komarova, Mycoth. Eston. 3: 13 (1961).
- =Perenniporia subacida (Peck) Donk, Persoonia 5(1): 76 (1967).
- =Poria colorea Overh. & Englerth, Bull. Yale Univ. School For. 50: 21 (1942).
- =Poria fuscomarginata Berk. ex Cooke, Grevillea 15(no. 73): 24 (1886).
- =Poria subaurantia Berk. ex Cooke, Grevillea 15(no. 73): 27 (1886).
Fruiting body: Perennial, resupinate to effused-reflex, becomes corky when dried, about 22 cm or more at the longest dimension, 15 cm or more at the widest dimension, and up to 17 mm thick at the center. Pore surface pale yellowish to dingy-yellowish when fresh, dingy-yellowish to pale tan to ochraceous when dry; pores round to angular, 4–6 per mm; dissepiments thin, entire. Subiculum thin, cream to buff, up to 1 mm thick. Tubes concolorous with pore surface, up to 16 mm thick.
Hyphal structure: Hyphal system di-trimitic; generative hyphae with clamp connections; skeletal hyphae strongly dextrinoid, CB+; dissolving in KOH.
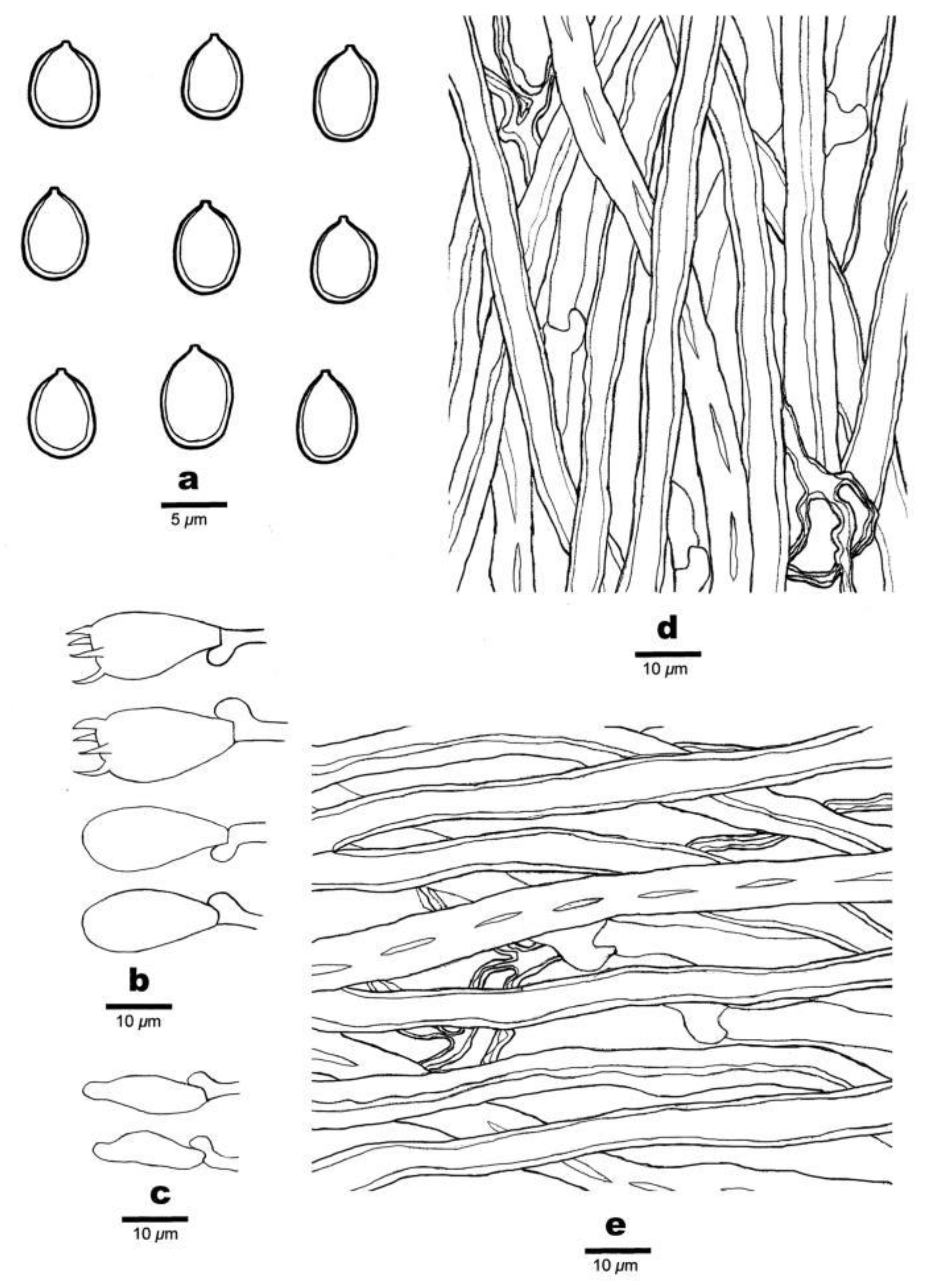
Context: Generative hyphae infrequent, hyaline, thin-walled, 2.5–4 µm in diameter; skeletal hyphae dominant, thick-walled with a wide lumen, unbranched, subparallel, 3–5.5 µm in diameter; skeletal-binding hyphae hyaline, thick-walled, frequently branched, flexuous, interwoven, 1–2 µm in diameter.
Tubes: Generative hyphae infrequent, hyaline, thin-walled, 2.5–3.5 µm in diameter; skeletal hyphae dominant, thick-walled with a wide lumen, unbranched, subparallel, 3–4.5 µm in diameter; skeletal-binding hyphae hyaline, thick-walled, frequently branched, flexuous, interwoven, 0.7–1.7 µm in diameter. Cystidia absent, but fusoid cystidioles present, hyaline, thin-walled, 13–16 × 3–4.5 µm. Basidia barrel-shaped, with four sterigmata and a basal clamp connection, 20–22.5 × 7–8 µm; basidioles similar in shape to basidia, but slightly smaller.
Basidiospores: Ellipsoid, not truncate, hyaline, thick-walled, smooth, non-dextrinoid, CB+, (4.2–)4.5–6.2(–6.4) × (3.2–)3.6–4.6(–4.9) µm, L = 5.25 µm, W = 3.92 µm, Q = 1.13–1.42 (n = 450/15).
Associated wood-rot: White.
Substrates and distribution: Mainly on conifers, but also on hardwood, causes white rot in Abies Mill and Tsuga Carr. and common on dead fallen trees in many areas. A boreal eastern species in Europe, widely distributed in forest regions of Asia and North America [8,10,11].
Additional specimen examined: CANADA, Ontario, Gull lake, on Thuja occidentalis L., July 25 1919, J.H. Faull (5088) (FH 00605379); Lake Timagami, Timagami Is., on fallen trunk of Abies balsamea (L.) Mill., September 10 1918, J.H. Faull (3365) (FH 00605372); Lake Rosseau, on Tsuga Carr., September 1902, Harper (587) (FH 00605380); Lake Rosseau, on Pinus Linn log, October 1903, S.A. Haper (832) (FH); Lake Rosseau, on fallen log of Abies balsamea (L.) Mill., 18 August 1921, J.H. Faull (6013) (FH 00605381); Humber Valley, on roots of Thuja occidentalis L., September 1914, J.H. Faull (169) (FH 00605373); Humber Valley, on dead hardwood, September 1914, J.H. Faull (156) (FH 00605375); Sudbury District, Cleland Tp., on dead coniferous wood, 15 September 1918, J.H. Faull (3502) (FH 00605377). CHINA, Fujian Province, Wuyishan Nature Reserve, on fallen angiosperm trunk, 21 October 2005, Dai 7316 (IFP); Guizhou Province, Jiangkou County, Fanjingshan Nature Reserve, on fallen angiosperm trunk, 21 August 2010, Yuan 5511 (IFP); Heilongjiang Province, Yichun, Fenglin Nature Reserve, on fallen trunk of Picea Dietr., 2 August 2011, Cui 9849, 9853 (BJFC); Yunnan Province, Baoshan, Gaoligong Mountain, on angiosperm trunk, 23 September 2007, Yuan 3850, 3854 (IFP); Zhejiang Province, Linan, Tianmushan Nature Reserve, on fallen trunk of Picea Dietr., 11 October 2005, Cui 2705 2712 (BJFC). FINLAND, Pera-Pohjanma, Pisavaara National Park, on fallen trunk of Picea Dietr., 14 September 1997, Dai 2648 (BJFC); Pisavaara National Park, on Picea Dietr., 4 November 2011, Dai 12619 (BJFC). USA, Arizona, Coronado National Forest, on conifer, 28 August 1958, Lowe (9407) (FH 00605393); Connecticut, New Haven, Sleeping Giant State Park, on fallen trunk of Betula L., 22 July 2012, Dai 12773 (BJFC); as above, on fallen trunk of Tsuga Carr., 22 July 2012, Dai 12785 (BJFC); as above, 24 July 2012, Dai 10287 (BJFC); as above, on fallen trunk of Pinus Linn, 24 July 2012, Dai 10285 (BJFC); Idaho, Bovill County, on dead fallen trunk of Pinus Linn, 2 October 1920, A.S. Rhoads (15891)(FH 00605391); Priest River, on Picea engelmanni, September 1915, J.R. Weir (9551) (FH 00605392); Priest River, on Abies grandis (Dougl ex D Don) Lindl, September 1915, J.R. Weir (8152) (FH 00605394); Kansas, Rooks County, Rockport, on underside of old log, 9 December 1893, (1315) (FH 00605389); Maine, Norcross, on fallen Populus L. log, 20 August 1940, D.H. Linder s.n. (FH 00605344), Kittery Point, on log of Quercus L., 28 July 1922, J.R. Weir (897)(FH 00605347); Linekin, on Picea Dietr., September 1899, Burt (2125B) (FH); as above, 16 August 1899, Burt 2127 (FH); Massachusetts, Norfolk County, Sharon, 28 June 1946, A.P.D.Piguet (54) (FH 00605353); Michigan, Neebish, September 1911, E.T.Harper & S.A.Harper s.n. (FH 00605383); Isle Royal, Rock Harbor, August 1904, E.T.Harper & S.A.Harper s.n. (FH 00605384); Pleasant Ridge, on fallen Tsuga Carr., 25 September 1915, P.Spaulding & J.F.Collins s.n. (FH 00605345); New York, Osceola, on Tsuga Carr., August 1885, C.H. Peck, BPI 844697 (Holotype, BPI); as above, on Picea Dietr. log, FH 0053717 (FH); Dryden, Ringwood Preserve, on old wood, 9 June 1952, W.B.Cooke & V.G.Cooke (29105) (FH 00605356); Ithaca, on Betula lutea F.Michx., nom. illeg. 21 March 1935, W.L. White (1568) (FH 00605360); Floodwood, on conifer, August 1900, Burt 2122B (FH); Floodwood, on Picea Dietr. log, 20 August 1900, 2123B (FH); Pennsylvania: Bedford County, New Paris, on fallen Quercus L., 1 September 1916, J.H. Faull (1490) (FH 00605366); Lackawanna County, Carbondale, on Tsuga Carr. log, 13 December 1898, Burt 2124C (FH); Tennessee, Knox County, Ball Camp Pike, on Pinus Linn log, 18 December 1938, L.R. Hesler (11907) (FH 00605387); Vermont: Addison County, Ripton, on hardwood log, 31 October 1896, (FH 00605346); Addison County, Ripton, on Picea Dietr., 4 November 1896, Burt 2126A (FH); Addison County, Ripton, on Picea Dietr., 31 October 1896, Burt 2126B (FH); Abby, on Tsuga Carr. log, 31 October 1896, Burt 2123A (FH); Underhill, on hardwood, 18 May 1880, Pringle (1025) (FH 00605350); as above, on dead conifer, August 1897, Burt 2121A, B, C (FH); as above, on fallen trunk of Tsuga Carr., September 1902, Harper 625 (FH); Dunamase, on Tsuga Carr. log, 14 September 1896, Burt 2121E (FH); Dunamase, on Tsuga Carr., October 1896, Burt 2122A (FH); Snake Mountain, January 1896, 2124A (FH); as above, September 1902, Harper 746 (FH); Washington: Marymere Falls, Olympic National Park, on Pseudotsuga taxifolia (Lamb.) Britton, WB (27590) (FH 00605395); Clallam County, Sol Duc Hot Springs, on fallen Tsuga heterophylla (Raf.) Sarg., 4 July 1920, J.R. Weir (650) (FH 00605396); as above, on fallen trunk of Picea sitchensis (Bong.) Carr., 14 July 1920, J.R. Weir (609) (FH 00605397); Wisconsin, June 21 1946, Neuman (162) (FH 00605386).
4. Discussion
The taxonomic position of this taxon has been long debated [16,17,18,19,20]. It was transferred to Perenniporia by Donk [20], and this placement has been generally accepted [8,9,10,11,12]. Due to a combination of characteristics (completely unbranched skeletal hyphae and ellipsoid and non-truncate basidiospores), Decock and Staplers [13] mentioned that the species did not belong to Perenniporia. Based on phylogenetic and morphological grounds, Robledo et al. confirmed that this taxon should be separated from Perenniporia, and could be recognized as a distinct genus morphologically as well [28]. Previous studies also confirmed that this species formed a clade distinct from the P. medulla-panis clade [5,6,28]. The present study confirms these results from previous studies and formally describes the new genus, Poriella with P. subacida as type species.
Other species of Perenniporia and Poriella subacida share the morphological features of unbranched skeletal hyphae and non-truncate basidiospores, such as Perenniporia africana (Ipulet and Ryvarden) in Uganda [50,51], P. contraria (Berk. and M.A. Curtis) Ryvarden in Cuba, and P. ellipsospora (Ryvarden and Gilb.) in North America [10]. A comparison of Poriella and related genera is presented in Table 2.
Three species have been mentioned as possible synonyms of this highly variable species. Poria colorea Overh. & Englerth, described from Western Tsuga Carr. and has generally been considered to be conspecific with Polyporus subacidus. Poria fuscomarginata Berk. ex Cooke was commented on by Murrill [52] who found the type material to be badly preserved and scanty. He concluded that “it was suggested little.” Poria subaurantia Berk. ex Cooke was considered a synonym of Polyporus subacidus by Murrill [53]. Judging by their descriptions, these fungi show a range of variation that is acceptable for P. subacidus.
Our molecular phylogenetic analyses revealed that the genus Poriella is closely related to Yuchengia narymica and then grouped with Vanderbylia based on the ITS+28S gene regions and the combined ITS+28S+mtSSU+Tef1 sequences (Figure 1 and Figure 2). Morphologically, two species share the same hyphal system and basidiospores. However, Y. narymica differs markedly from the acyanophilous, amyloid skeletal hyphae [38,53,54,55]. Vanderbylia mainly differs in its pileate basidiomata, a dimitic hyphal system with distinctly arboriform vegetative hyphae in the hymenophoral trama and obovoid basidiospores [10,56].
Poriella subacida is primarily a boreal taxon and is widely distributed in forest regions of northern Asia, North America and Europe [8,10,11]. It mainly grows on conifers, especially Picea Dietr., but also on Larix and Pinus Linn. In Europe, it has also been found occasionally on hardwoods like Populus L. and Prunus L. [8], and other hardwoods in North America and Asia [10,11]. The species is also present in tropical areas (e.g., in Africa) [57], but these records should be treated with caution. Ipulet and Ryvarden [50] recently described Perenniporia africana Ipulet and Ryvarden, from Uganda, with seemingly the same combination of characteristicss, i.e., unbranched skeletal hyphae and non-truncate basidiospores [57,58]. A list of characteristics of Poriella subacida comb. nov. from different regions is presented in Table 3.
Poriella subacida causes white rot of conifers and hardwoods and also butt and root rots of living conifers. Due to the cream to golden yellow mycelia felts that develop in the decayed wood, this rot is commonly called “feather rot” [10].
Author Contributions
Conceptualization, C.-L.Z. and R.C.; methodology, C.-L.Z. and R.C.; software, R.C.; validation, R.C., C.-L.Z. and S.C.K.; formal analysis, R.C.; resources, C.-L.Z.; data curation, R.C.; writing—original draft preparation, R.C.; writing—review and editing, C.-L.Z. and S.C.K.; visualization, R.C.; supervision, C.-L.Z. and S.C.K.; project administration, C.-L.Z.; funding acquisition, C.-L.Z. and S.C.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Yunnan Fundamental Research Project (Grant No. 202001AS070043) and the High-level Talents Program of Yunnan Province (YNQR-QNRC-2018-111).
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ncbi.nlm.nih.gov/; https://www.mycobank.org/page/Simple%20names%20search; http://purl.org/phylo/treebase, submission ID 23826; accessed on 25 June 2021.
Acknowledgments
Special thanks are conveyed to Yu-Cheng Dai (BJFU, China), Bao-Kai Cui (BJFU, China), and Donald H. Pfister (Harvard University) for allowing me to study their specimens.
Conflicts of Interest
The authors declare no conflict of interest. All the experiments undertaken in this study comply with the current laws of the People’s Republic of China.
References
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a fully resolved fungal tree of life. Annu. Rev. Microbiol. 2020, 15, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.H.; Ryvarden, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Minter, D.W.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CAB International Press: Wallingford, UK, 2008. [Google Scholar] [CrossRef]
- Cui, B.K.; Li, H.J.; Ji, X.; Zhou, J.L.; Song, J.; Si, J.; Yang, Z.L.; Dai, Y.C. Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Divers. 2019, 97, 137–392. [Google Scholar] [CrossRef]
- El-Gharabawy, H.M.; Leal-Dutra, C.A.; Griffith, G.W. Crystallicutis gen. nov. (Irpicaceae, Basidiomycota), including C. damiettensis sp. nov., found on Phoenix dactylifera (date palm) trunks in the Nile Delta of Egypt. Fungal Biol. 2021, 125, 447–458. [Google Scholar] [CrossRef]
- Matozaki, T.; Hattori, T.; Maekawa, N.; Nakagiri, A.; Ishikawa, N.K.; Sotome, K. Hirticrusta gen. nov. segregated from Neofomitella in Polyporaceae (Polyporales). Mycoscience 2020, 5, 240–248. [Google Scholar] [CrossRef]
- Ma, X.; Zhao, C.L. Crepatura ellipsospora gen. et sp. nov. in Phanerochaetaceae (Polyporales, Basidiomycota) bearing a tuberculate hymenial surface. Mycol. Prog. 2019, 18, 785–793. [Google Scholar] [CrossRef]
- Ryvarden, L.; Melo, I. Poroid fungi of Europe. Syn. Fung. 2014, 31, 1–455. [Google Scholar]
- Peck, C.H. Report of the botanist. Ann. Rep. N. Y. State Mus. Nat. Hist. 1885, 38, 77–138. [Google Scholar]
- Gramss, G.R.; Gilbertson, L.; Ryvarden, L. North American Polypores. Volume 1: Abortiporus—Lindtneria. 433 S., 209 Abb. Oslo 1986. Fungiflora A/S. J. Basic Microbiol. 1987. [Google Scholar] [CrossRef]
- Núñez, M.; Ryvarden, L. East Asian polypores 2. Syn. Fung. 2001, 14, 165–522. [Google Scholar]
- Dai, Y.C.; Niemelä, T.; Kinnunen, J. The polypore genera Abundisporus and Perenniporia (Basidiomycota) in China, with notes on Haploporus. Ann. Bot. Fenn. 2002, 39, 169–182. [Google Scholar]
- Decock, C.; Stalpers, J. Studies in Perenniporia: Polyporus unitus, Boletus medulla-panis, the nomenclature of Perenniporia, Poria and Physisporus, and a note on European Perenniporia with a resupinate basidiome. Taxon 2006, 53, 759–778. [Google Scholar] [CrossRef]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Zhao, C.L.; Cui, B.K.; Dai, Y.C. New species and phylogeny of Perenniporia based on morphological and molecular characters. Fungal Divers. 2013, 58, 47–60. [Google Scholar] [CrossRef]
- Shen, S.; Xu, T.M.; Jason, K.; Zhao, C.L. Morphological and molecular identification of a new species of Perenniporia (Polyporales, Basidiomycota) in North America. Phytotaxa 2018, 351, 63–71. [Google Scholar] [CrossRef]
- Saccardo, P.A. Sylloge hymenomycetum, Vol. II. Polyporeae, Hydneae, Thelephoreae, Clavarieae, Tremellineae. Sylloge Fungorum 1888, 6, 1–928. [Google Scholar]
- Bondartsev, A.; Singer, R. Zur systematik der Polyporaceae. Ann. Mycol. 1941, 39, 43–65. [Google Scholar]
- Parmasto, E. Eesti seente eksikaat. Mycotheca Est. 1961, 3, 51–75. [Google Scholar]
- Donk, M.A. Notes on European polypores II. Persoonia 1967, 5, 47–130. [Google Scholar]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef]
- Miettinen, O.; Larsson, K.H.; Sjökvist, E.; Larsson, K.L. Comprehensive taxon sampling reveals unaccounted diversity and morphological plasticity in a group of dimitic polypores (Polyporales, Basidiomycota). Cladistics 2012, 28, 251–270. [Google Scholar] [CrossRef] [Green Version]
- Binder, M.; Justo, A.; Riley, R.; Salamov, A.; López-Giráldez, F.; Sjökvist, E.; Copeland, A.; Foster, B.; Sun, H.; Larsson, E.; et al. Phylogenetic and phylogenomic overview of the Polyporales. Mycologia 2013, 105, 1350–1373. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C.; Cui, B.K.; Si, J.; He, S.H.; Hyde, K.D.; Yuan, H.S.; Lui, X.Y.; Zhou, L.W. Dynamics of the worldwide number of fungi with emphasis on fungal diversity in China. Mycol. Prog. 2015, 14, 62. [Google Scholar] [CrossRef]
- Miettinen, O.; Spirin, V.; Vlasák, J.; Rivoire, B.; Stenroos, S.; Hibbett, D. Polypores and genus concepts in Phanerochaetaceae (Polyporales, Basidiomycota). MycoKeys 2016, 17, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Dai, Y.C.; Vlasák, J.; Yuan, Y. Molecular phylogeny and global diversity of the genus Haploporus (Polyporales, Basidiomycota). J. Fungi 2021, 7, 96. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.B.; Zhou, M.; Yuan, Y.; Dai, Y.C. Global diversity and taxonomy of Sidera (Hymenochaetales, Basidiomycota): Four new species and keys to species of the genus. J. Fungi. 2021, 7, 251. [Google Scholar] [CrossRef]
- Robledo, G.L.; Amalfi, M.; Castillo, G.; Rajchenberg, M.; Decock, C. Perenniporiella chaquenia sp. nov. and further notes on Perenniporiella and its relationships with Perenniporia (Poriales, Basidiomycota). Mycologia 2009, 101, 657–673. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort: The Danish Mycological Society’s Colour Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996. [Google Scholar]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- Huang, R.X.; Zhao, C.L. Three new species of Phlebia (Polyporales, Basidiomycota) based on the evidence from morphology and DNA sequence data. Mycol. Prog. 2020, 19, 753–767. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Lutzoni Lab. Available online: http://lutzonilab.org (accessed on 4 June 2021).
- Rehner, S.A.; Buckley, E.A. Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [PubMed]
- Kim, K.M.; Lee, J.S.; Jung, H.S. Fomitopsis incarnatus sp. nov. based on generic evaluation of Fomitopsis and Rhodofomes. Mycologia 2007, 99, 833–841. [Google Scholar] [CrossRef]
- Vlasak, J.; Vlasa, K.J., Jr.; Cui, B.K. Antrodia kmetii, a new European polypore similar to Antrodia variiformis. Cryptogam. Mycol. 2013, 34, 203–209. [Google Scholar] [CrossRef]
- Tomšovský, M.; Menkis, A.; Vasaitis, R. Phylogenetic relationships in European Ceriporiopsis species inferred from nuclear and mitochondrial ribosomal DNA sequences. Fungal Biol. 2010, 114, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Justo, A.; Hibbett, D.S. Phylogenetic classification of Trametes (Basidiomycota, Polyporales) based on a five-marker dataset. Taxon 2011, 60, 1567–1583. [Google Scholar] [CrossRef]
- Zhao, C.L.; Cui, B.K.; Steffen, K.T. Yuchengia, a new polypore genus segregated from Perenniporia (Polyporales, Basidiomycota) based on morphological and molecular characters. Nord. J. Bot. 2013, 31, 331–338. [Google Scholar] [CrossRef]
- Sotome, K.; Hattori, T.; Ota, Y.; Toanun, C.; Salleh, B.; Kakishima, M. Phylogenetic relationships of Polyporus and morphologically allied genera. Mycologia 2008, 100, 603–615. [Google Scholar] [CrossRef]
- Brazee, N.J.; Lindner, D.L.; Fraver, S.; D’Amato, A.W.; Milo, A.M. Wood-inhabiting, polyporoid fungi in aspen-dominated forests managed for biomass in the U.S. Lake States. Fungal Ecol. 2012, 5, 600–609. [Google Scholar] [CrossRef]
- Tabata, M.; Harrington, T.C.; Chen, W.; Abe, Y. Molecular phylogeny of species in the genera Amylostereum and Echinodontium. Mycoscience 2000, 41, 585–593. [Google Scholar] [CrossRef]
- Pildain, M.B.; Rajchenberg, M. The phylogenetic position of Postia s.l. (Polyporales, Basidiomycota) from Patagonia, Argentina. Mycologia 2013, 105, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. (Archived by WebCite(r)). 4 August 2009. Available online: http://www.phylo.org/sub_sections/portal; http://www.webcitation.org/5imQlJeQa (accessed on 4 June 2021).
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author. Evolutionary Biology Centre; Uppsala University: Uppsala, Sweeden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipulet, P.; Ryvarden, L. New and interesting polypores from Uganda. Syn. Fung. 2005, 20, 87–99. [Google Scholar]
- Decock, C.; Ryvarden, L. Perenniporiella gen. nov. segregated from Perenniporia, including key to neotropical Perenniporia species with pileate basidiomes. Mycol. Res. 2003, 107, 93–103. [Google Scholar] [CrossRef]
- Murrill, W.A. Some described species of Poria. Mycologia 1919, 11, 231–244. [Google Scholar] [CrossRef]
- Murrill, W.A. Light-colored resupinate polypores II. Mycologia 1920, 12, 299–308. [Google Scholar]
- Pilát, A. Additamenta ad floram Sibiriae Asiaeque orientalis mycologicam. Pars Tertia. Bull. Soc. Mycolog. 1936, 51, 351–426. [Google Scholar]
- Pouzar, Z. Notes on four European polypores. Česká Mykol. 1984, 38, 203–204. [Google Scholar]
- Decock, C.; Ryvarden, L. Studies in neotropical polypores. Some coloured resupinate Perenniporia species. Mycol. Res. 1999, 103, 1138–1144. [Google Scholar] [CrossRef]
- Ryvarden, L.; Johansen, I. A Preliminary Polypore Flora of East Africa; Fungiflora: Oslo, Norway, 1980. [Google Scholar]
- Ryvarden, L. Genera of polypores: Nomenclature and taxonomy. Syn. Fung. 1991, 5, 1–363. [Google Scholar]
Figure 1.
Maximum parsimony strict consensus tree illustrating the phylogeny of Poriella subacida and related species in Polyporales based on ITS+28S sequences. Branches are labeled with maximum likelihood bootstrap higher than 70%, parsimony bootstrap proportions higher than 50% and Bayesian posterior probabilities more than 0.95, respectively. Clade names [23].
Figure 1.
Maximum parsimony strict consensus tree illustrating the phylogeny of Poriella subacida and related species in Polyporales based on ITS+28S sequences. Branches are labeled with maximum likelihood bootstrap higher than 70%, parsimony bootstrap proportions higher than 50% and Bayesian posterior probabilities more than 0.95, respectively. Clade names [23].

Figure 2.
Maximum Parsimony strict consensus tree illustrating the phylogeny of Poriella subacida, and related species in Perenniporia s.l. based on the combined ITS+28S+mtSSU+TEF1 sequence datasets. Branches are labeled with maximum likelihood bootstrap higher than 70%, parsimony bootstrap proportions higher than 50% and Bayesian posterior probabilities more than 0.95, respectively. Clade names follow [15].
Figure 2.
Maximum Parsimony strict consensus tree illustrating the phylogeny of Poriella subacida, and related species in Perenniporia s.l. based on the combined ITS+28S+mtSSU+TEF1 sequence datasets. Branches are labeled with maximum likelihood bootstrap higher than 70%, parsimony bootstrap proportions higher than 50% and Bayesian posterior probabilities more than 0.95, respectively. Clade names follow [15].

Figure 3.
Basidiomata of Poriella subacida (Holotype, BPI 844697). Scale bar = 1 cm.

Figure 4.
Microscopic structures of Poriella subacida (Holotype). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidioles. (d) Hyphae from trama. (e) Hyphae from context. Bars: A = 5 µm; B–E = 10 µm.
Figure 4.
Microscopic structures of Poriella subacida (Holotype). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidioles. (d) Hyphae from trama. (e) Hyphae from context. Bars: A = 5 µm; B–E = 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Information of the sequences used in this study.
| Species Name | Sample No. | GenBank Accessions | References | |||
|---|---|---|---|---|---|---|
| ITS | LSU | mtSSU | TEF1 | |||
| Abortiporus biennis | TFRI 274 | EU232187 | EU232235 | [21] | ||
| A. biennis | EL65-03 | JN649325 | JN649325 | [21] | ||
| Abundisporus roseoalbus | Dai 12269 | KC415908 | KC415910 | KF051037 | KF181131 | [5] |
| A. pubertatis | Cui 5776 | KC787565 | KC787572 | KF051029 | KF181129 | [5] |
| A. sclerosetosus | MUCL 41438 | FJ411101 | FJ393868 | [28] | ||
| A. violaceus | Ryvarden 10775 | KF018126 | KF018134 | KF051058 | KF181152 | [5] |
| Amylocystis lapponica | KHL 11755 | EU118603 | EU118603 | [21] | ||
| Antrodia albida | CBS 308.82 | DQ491414 | [35] | |||
| A. albida | FP 105979 | EU232272 | EU232272 | [36] | ||
| A. heteromorpha | CBS 200.91 | DQ491415 | AY515350 | [35] | ||
| A. macra | MUAF 887 | EU340898 | [23] | |||
| Bjerkandera adusta | NBRC 4983 | AB733156 | AB733333 | [23] | ||
| Cinereomyces lindbladii | FBCC 177 | HQ659223 | HQ659223 | [23] | ||
| Climacocystis borealis | KH 13318 | JQ031126 | JQ031126 | [23] | ||
| Coriolopsis caperata | LE(BIN)-0677 | AB158316 | AB158316 | [37] | ||
| Donkioporia expansa | MUCL 35116 | FJ411104 | FJ393872 | [28] | ||
| Earliella scabrosa | PR1209 | JN165009 | JN164793 | [38] | ||
| Fragiliporia fragilis | Dai 13080 | KJ734260 | KJ734264 | KJ734268 | KJ790245 | [5] |
| F. fragilis | Dai 13559 | KJ734261 | KJ734265 | KJ734269 | KJ790246 | [5] |
| F. fragilis | Dai 13561 | KJ734262 | KJ734266 | KJ734270 | KJ790247 | [5] |
| Ganoderma sichuanense | Wu 1006-38 | JQ781858 | JX029989 | JX029976 | [5] | |
| G. sichuanense | Dai 12479 | JQ781864 | JX029988 | JX029975 | [5] | |
| G. australe | Cui 9511 | JN048773 | JN048792 | [39] | ||
| G. sinense | Wei 5327 | KF494998 | KF495008 | KF494976 | [5] | |
| G. applanatum | Dai 12483 | KF494999 | KF495009 | KF494977 | [5] | |
| Gelatoporia subvermispora | BRNU 592909 | FJ496694 | FJ496706 | [37] | ||
| Grammetheliopsis subtropica | Cui 9041 | JQ845096 | JQ845099 | KF051039 | KF181133 | [39] |
| Heterobasidion annosum | PFC 5252 | KC492906 | KC492906 | [23] | ||
| Hornodermoporus latissima | Cui 6625 | HQ876604 | JF706340 | KF051040 | KF181134 | [5] |
| H. martius | Cui 7992 | HQ876603 | HQ654114 | KF051041 | KF181135 | [5] |
| H. martius | MUCL 41677 | FJ411092 | FJ393859 | [28] | ||
| H. martius | MUCL 41678 | FJ411093 | FJ393860 | [28] | ||
| Hydnopolyporus fimbriatus | LR 40855 | JN649347 | JN649347 | [23] | ||
| Hypochnicium lyndoniae | NL 041031 | JX124704 | JX124704 | [23] | ||
| Lentinus tigrinus | DSH93-181 | AY218419 | AF518627 | U27050 | [38] | |
| Microporellus violaceocinerascens | MUCL 45229 | FJ411106 | FJ393874 | [28] | ||
| M. violaceocinerascens | Cui 8459 | HQ876606 | HQ654113 | KF051042 | KF181136 | [5] |
| Obba rivulosa | KCTC 6892 | FJ496693 | FJ496710 | [4] | ||
| Perenniporia hainaniana | Cui 6364 | JQ861743 | JQ861759 | KF051044 | KF181138 | [5] |
| P. hainaniana | Cui 6365 | JQ861744 | JQ861760 | KF051045 | KF181139 | [5] |
| P. hainaniana | Cui 6366 | JQ861745 | JQ861761 | KF494996 | KF494981 | [5] |
| P. medulla-panis | MUCL 49581 | FJ411088 | FJ393876 | [28] | ||
| P. medulla-panis | MUCL 43250 | FJ411087 | FJ393875 | [28] | ||
| P. medulla-panis | Cui 3274 | JN112792 | JN112793 | KF051043 | KF181137 | [5] |
| P. substraminea | Cui 10177 | JQ001852 | JQ001844 | KF051046 | KF181140 | [5] |
| P. substraminea | Cui 10191 | JQ001853 | JQ001845 | KF051047 | KF181141 | [5] |
| P. substraminea | Dai 10781 | KF495007 | KF495018 | KF494995 | KF494983 | [5] |
| Perenniporiella chaquenia | MUCL 47647 | FJ411083 | FJ393855 | HM467609 | [28] | |
| P. chaquenia | MUCL 47648 | FJ411084 | FJ393856 | HM467610 | [28] | |
| P. micropora | MUCL43581 | FJ411086 | FJ393858 | HM467608 | [28] | |
| P. neofulva | MUCL 45091 | FJ411080 | FJ393852 | HM467599 | [28] | |
| P. pendula | MUCL 46034 | FJ411082 | FJ393853 | HM467601 | [28] | |
| Phanerochaete chrysosporium | BKM-F-1767 | HQ188436 | GQ470643 | [37] | ||
| Phlebia unica | KHL 11786 | EU118657 | EU118657 | [37] | ||
| Physisporinus sanguinolentus | BRNM 699576 | FJ496671 | FJ496725 | [37] | ||
| Piloporia sajanensis | Mannine 2733a | HQ659239 | HQ659239 | [37] | ||
| Podoscypha venustula | CBS 65684 | JN649367 | JN649367 | [23] | ||
| Polyporus tuberaster | CulTENN 8976 | AF516598 | AJ488116 | [40] | ||
| Poriella subacida | Dai 8224 | HQ876605 | JF713024 | KF218322 | KF286328 | [15] |
| P. subacida | Cui 3643 | FJ613655 | AY336753 | KF218320 | KF286326 | [15] |
| P. subacida | Cui 10053 | KF495006 | KF495017 | KF218321 | KF286327 | [15] |
| P. subacida | MUCL 31402 | FJ411103 | FJ393880 | [28] | ||
| P. subacida | CBS 463.50 | FJ805245 | Direct submission | |||
| P. subacida | DLL 2009-125 | JQ673136 | [41] | |||
| P. subacida | DLL 2009-150 | JQ673014 | [41] | |||
| P. subacida | DLL 2009-154 | JQ673015 | [41] | |||
| P. subacida | Dai 8859 | FJ613656 | [15] | |||
| P. subacida | HHb-14877-T | AY089739 | AY089739 | Direct submission | ||
| P. subacida | B 37 | AF218403 | [42] | |||
| Postia alni | X 1400 | KC595932 | KC595932 | [23] | ||
| P. caesia | CIEFAP 174 | JX090109 | JX090129 | [23] | ||
| P. guttulata | KHL 11739 | EU11865 | EU11865 | [43] | ||
| P. venata | CIEFAP 346 | JX090113 | JX090133 | [43] | ||
| P. lactea | X 1391 | KC595939 | KC595939 | [23] | ||
| Pyrofomes demidoffii | MUCL 41034 | FJ411105 | FJ393873 | [28] | ||
| Sebipora aquosa | Miettinen 8680 | HQ659240 | HQ659240 | [22] | ||
| Skeletocutis amorpha | Miettinen 11038 | FN907913 | FN907913 | [37] | ||
| Stereum hirsutum | NBRC 6520 | AB733150 | AB733325 | [23] | ||
| Trametes elegans | FP105679 | JN048766 | JN048785 | [15] | ||
| T. hirsuta | Cui 7784 | JN048768 | JN048787 | [15] | ||
| T. hirsuta | RLG5133T | JN164854 | JN164801 | AF042154 | JN164891 | [38] |
| T. pubescens | PRM 900586 | AY684173 | AY855906 | [37] | ||
| Truncospora ochroleuca | Dai 11486 | HQ654105 | JF706349 | KF051048 | KF181142 | [5] |
| T. ochroleuca | MUCL 39726 | FJ411098 | FJ393865 | [28] | ||
| T. ochroleuca | Cui 5671 | JX941584 | JX941602 | KF218309 | KF286315 | [5] |
| T. ochroleuca | Cui 5673 | JX941585 | JX941603 | KF218308 | KF286314 | [5] |
| T. ornata | Cui 5714 | HQ654103 | HQ654116 | KF051056 | KF181150 | [5] |
| T. ohiensis | MUCL 41036 | FJ411096 | FJ393863 | [28] | ||
| Tyromyces chioneus | Cui 10225 | KF698745 | KF698756 | [5] | ||
| T. kmetii | Penttila 13474 | KF705040 | KF705041 | [5] | ||
| Vanderbylia delavayi | Dai 6891 | JQ861738 | KF495019 | KF218287 | KF286293 | [5] |
| V. fraxinea | DP 83 | AM269789 | AM269853 | [28] | ||
| V. fraxinea | Cui 7154 | HQ654095 | HQ654110 | KF218288 | KF286294 | [5] |
| V. fraxinea | Cui 8885 | HQ876611 | JF706344 | KF218289 | KF286295 | [5] |
| V. fraxinea | Cui 8871 | JF706329 | JF706345 | KF051050 | KF181144 | [5] |
| V. robiniophila | Cui 5644 | HQ876609 | JF706342 | KF051051 | KF181145 | [5] |
| V. vicina | MUCL 44779 | FJ411095 | AF518666 | [28] | ||
| Yuchengia narymica | Dai 7050 | JN048776 | JN048795 | KF051053 | KF181147 | [39] |
| Y. narymica | Dai 10510 | HQ654101 | JF706346 | KF051054 | KF181148 | [39] |
| Y. narymica | Dai 6998 | JN048775 | JN048794 | KF051055 | KF181149 | [5] |
| Y. narymica | 0709/42 | JN641258 | JN641265 | [39] | ||
| Y. narymica | 0709/157 | JN641259 | JN641266 | [39] | ||
| Y. narymica | 0809/3 | JN641261 | JN641268 | KF051049 | KF181143 | [39] |
Table 2.
A comparison of Poriella subacida and related genera and species.
| Genera | Hyphal System | Basidiospore Morphology | Chemical Reactions | Reference | |
|---|---|---|---|---|---|
| Skeletal Hyphae | Basidiospores | ||||
| Perenniporia s.s. | dimitic | ellipsoid, truncate or not | dextrinoid or not, CB+ | variable dextrinoid, CB+ | [13] |
| Perenniporiopsis | trimitic | oblong-ellipsoid, truncate | dextrinoid, CB– in the context, CB+ in the trama | dextrinoid, CB+ | [6] |
| Perenniporiella | dimitic | globose to subglobose, non-truncate | non-dextrinoid to strongly dextrinoid, slightly to distinctly cyanophilous | slightly dextrinoid, CB+ | [51] |
| Poriella subacida | di-trimitic | ellipsoid, non-truncate | strongly dextrinoid, CB+ | non-dextrinoid, CB+ | This study |
| Yuchengia narymica | dimitic | ellipsoid, non-truncate | amyloid, CB– | IKI–, CB+ | [38] |
CB+ = cyanophilous, CB– = acyanophilous.
Table 3.
A list of characteristics of Poriella subacida comb. nov. from different regions.
| Specimens | Locality | Basidiospores (µm) | Average | Q | Pores/mm | Substrate |
|---|---|---|---|---|---|---|
| BPI 844697 (Type) | USA, NY | (4.2–)4.4–5.7(–6.2) × (3.1–)3.5–4.3(–4.5) | 5.2 × 3.9 | 1.35 | 4–5 | on Tsuga Carr. |
| BPI 844698 | USA, NY | (4.7–)5–6(–6.4) × (3.6–)3.9–4.6(–4.8) | 5.4 × 4.1 | 1.36 | 4–5 | on Tsuga Carr. |
| BPI 844699 | USA, NY | (4.3–)4.5–5.7(–6.2) × (3.2–)3.6–4.4(–4.6) | 5.2 × 3.9 | 1.35 | 4–5 | on Tsuga Carr. |
| BPI 885858 | USA, NY | (4.9–)5.1–6.2(–6.5) × (3.4–)3.7–4.6(–4.8) | 5.6 × 4.2 | 1.3 | 4–5 | on Tsuga Carr. |
| FH0053717 | USA, NY | (4.8–)5–5.6(–5.8) × (3.7–)3.9–4.5(–4.7) | 5.3 × 4.2 | 1.33 | 4–5 | on the log of Picea asperata Mast. |
| Hesler 11907 | USA, TN | (4.4–)4.6–5.3(–5.5) × (3.5–)3.7–4.3(–4.6) | 4.9 × 4 | 1.2 | 4–5 | on the log of Pinus Linn |
| 00605389 | USA, KS | (4.1–)4.5–5.1(–5.3) × (3–)3.4–4.5(–4.8) | 4.8 × 3.8 | 1.3 | 4–6 | on underside of old log |
| Lowe 9407 | USA, AZ | (4.6–)4.9–5.8(–6) × (3.9–)4.3–4.9(–5.1) | 5.4 × 4.5 | 1.2 | 4–6 | on the log of Picea asperata Mast. |
| 00605344 | USA, ME | (4.3–)4.6–5.2(–5.4) × (3.4–)3.6–4.2(–4.6) | 4.9 × 3.9 | 1.2 | 4–5 | on fallen Populus L. log |
| 00605372 | Canada, Ontario | (4.4–)4.6–5.8(–6) × (3–)3.3–4.2(–4.5) | 5.3 × 3.8 | 1.2 | 4–6 | on fallen trunk of Abies Mill |
| Cui 9849 | China, Heilongjiang | (4.9–)5.2–5.7(–5.9) × (3.9–)4.1–4.4(–4.6) | 5.4 × 4.2 | 1.3 | 4–5 | on fallen trunk of Picea Dietr. |
| Yuan 3854 | China, Yunnan | (5–)5.2–5.8(–6) × (3.9–)4.1–4.4(–4.5) | 5.6 × 4.2 | 1.3 | 4–6 | on angiosperm trunk |
| Cui 2712 | China, Zhejiang | (4.9–)5.1–5.7(–6) × (3.9–)4–4.4(–4.6) | 5.4 × 4.2 | 1.3 | 4–5 | on fallen trunk of Picea Dietr. |
| Dai 2648 | Finland, Pohjanmaa | (4.3–)4.7–5.7(–6.1) × (3–)3.3–3.9(–4.1) | 5.3 × 3.6 | 1.3 | 4–6 | on fallen trunk of Picea Dietr. |
| Dai 12619 | Finland, Pohjanmaa | (4.5–)4.6–5.8(–6.4) × (3.1–)3.5–4.1(–4.6) | 5.2 × 3.8 | 1.3 | 4–5 | on Picea Dietr. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, R.; Karunarathna, S.C.; Zhao, C.-L. Poriella subacida Gen. & Comb Nov. for Perenniporia subacida (Peck) Donk. Agronomy 2021, 11, 1308. https://doi.org/10.3390/agronomy11071308
AMA Style
Chen R, Karunarathna SC, Zhao C-L. Poriella subacida Gen. & Comb Nov. for Perenniporia subacida (Peck) Donk. Agronomy. 2021; 11(7):1308. https://doi.org/10.3390/agronomy11071308
Chicago/Turabian StyleChen, Rui, Samantha C. Karunarathna, and Chang-Lin Zhao. 2021. "Poriella subacida Gen. & Comb Nov. for Perenniporia subacida (Peck) Donk" Agronomy 11, no. 7: 1308. https://doi.org/10.3390/agronomy11071308
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.