
Prospects for Increasing the Efficacy of Plant Resistance Inducers Stimulating Salicylic Acid
Unité Propre de Recherche Innovante, ERIT Plant Science, Interactions and Innovation, Avignon Université, 301 rue Baruch de Spinoza, 84140 Avignon, France
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(12), 3151; https://doi.org/10.3390/agronomy12123151
Submission received: 25 November 2022
/
Revised: 6 December 2022
/
Accepted: 7 December 2022
/
Published: 12 December 2022
(This article belongs to the Special Issue Phytoalexins, Resistance Inducers, and Sustainable Control Measures in Crop Protection Strategies)
Abstract
:Systemic acquired resistance is a powerful mechanism, based on the salicylic acid (SA) signaling pathway, which allows plants to resist to a wide range of pathogens. High SA, moreover, plays a key role in plant tolerance to abiotic stress. It seems, therefore, desirable to supply analogs of SA or stimulate the production of endogenous SA. Unfortunately, the chemical substances or physical means used for this effect often display a variable efficacy. After providing a review of them, we defend three major ideas: (i) plant resistance inducers (PRIs) must be combined for higher efficacy, notably for exploiting synergic effects between the SA and other signaling pathways, (ii) disease pressure can be reduced by exploiting the fungicidal properties displayed by some PRIs, (iii) biostimulants and crop management techniques should be used to ensure that plants have the resources they need to synthesize the compounds and structures required for efficient and lasting resistance. Some PRIs could also be used for their biostimulant effects in stress conditions. It could be concluded that holistic approaches which jointly address the issues of defense and tolerance stimulation, disease pressure and resource availability in plants are the ones that will allow for substantial reduction in fungicide use without sacrificing crop performance.
1. Introduction
In addition to resistance issues that may be exacerbated by climate change [1], pesticides have well-documented negative effects on the environment and human health [2]. However, pesticides cannot be dismissed as quickly as one would wish. They are generally not easily replaced by alternative techniques capable of controlling pests and diseases as efficiently and therefore securing the high yields needed by farmers for mere economic survival and human consumers for their daily supply in nutrients and other commodities. In addition to resistant or tolerant cultivars, biological control of pests and the development of complex cropping systems that are intrinsically less fragile and more resilient when confronted with aggressors and stressing conditions, plant resistance inducers (PRIs), alias elicitors, are on the rise [3]. Whereas techniques of biological control are more specifically designed for replacing insecticides and acaricides, PRIs represent the alternative to fungicides, including copper and sulfur. Several PRIs have, moreover, been found to be effective not only against fungi but also bacteria and viruses [4].
The PRIs that seem to be the most effective are the ones that stimulate Systemic Acquired Resistance (SAR), which is based on salicylic acid (SA) [5]. SAR is generally believed to be the most important mechanism of induced resistance (IR) in plants. Besides being systemic and more efficient than basal resistance (BR), SAR has lasting effects and presents a very broad spectrum of efficacy against numerous biotrophic and hemi-biotrophic pathogens in potentially all vascular plants. Induced Systemic Resistance (ISR), the other major form of IR, depends on the JA and ET pathways [6] and is basically efficient against necrotrophic pathogens and pests of the arthropod class [7]. Compared to ISR, SAR appears more universal and may therefore be considered as the “Holy Grail” in plant immunity. Not surprisingly, a vast majority of chemical PRIs target SAR [8].
Unfortunately, for all their promises, SA-based PRIs have so far proven less efficient than fungicides, or, more to the point, less reliable. Efficiency seems, indeed, highly dependent on disease pressure, which varies a lot from one cropping season to another [9], the climatic conditions prevailing at the time of treatments [10], and the physiological status of plants [11], to name only the three most obvious limiting factors. The challenge of fungicide reduction or replacement is clearly a steep one, rendered even steeper by climate change, which is characterized by an increase in frequency, intensity and duration of extreme climatic events such as drought, frost and heat waves, depending on years and locations [12]. One of the negative effects of stress is that it decreases carbon acquisition and accumulation, and therefore the level of resources available for the synthesis of the structures and molecules needed for defense. Resource availability has been stressed for being one of the major limiting factors of the efficacy of PRIs in the field [13,14].
After a brief reminder of the role played by SA in plant immunity and a more detailed reminder of its role in photosynthesis and crop performance in standard and stress conditions, we review the chemical and physical means that exist for increasing SA levels in plants. We then develop the point of view that the full potential of SA-based PRIs will be revealed by addressing the three following ideas: (i) chemical, biological and physical means of stimulation of plant defense must be combined for higher efficacy, possibly with the aim of exploiting synergic effects between the SA and other signaling pathways, (ii) disease pressure must and can be reduced, notably by exploiting the fungicide or the disinfecting properties displayed by some of the PRIs, (iii) crop management techniques and biostimulants, including some PRIs, can and must be used to ensure that plants have the resources required for adequate and lasting resistance.
The objective of this review article is to propose an organized vision for the way PRIs could be used, along with other levers, with the exclusion of genetic levers and canopy management techniques [15], to reap all the benefits associated with increased concentrations of SA in plants and SAR.
2. The Role of the SA Signaling Pathway in Plant Immunity
Plants have an innate immunity system capable of producing defense compounds and structures in response to aggression. Even though lines are blurry, a distinction is generally made between effector-triggered immunity (ETI) and pattern-triggered immunity (PTI). ETI represents a form of resistance founded on the existence of unique resistance genes (R) and their corresponding resistance proteins (R proteins). The latter are involved in the direct or indirect recognition of effectors and the rapid activation of defense responses, typically associated with the so-called hypersensitive response (HR) and cell death. ETI is only efficient in the short term, since the pathogen is bound to evolve and produce effectors that are no more recognized by R proteins. Whereas ETI can be considered as a qualitative form of resistance, PTI corresponds more to a quantitative form of resistance. PTI is triggered by multiple molecular patterns that are conserved in case of infection or infestation, and capable of activating a whole range of defense pathways and mechanisms. The latter can be modulated, moreover, as a function of the stage of interaction between the host plant and its aggressor. PTI is under the control of a complex signaling network that can be activated after recognition of molecular patterns associated with pathogenic microorganisms or the damage they cause. The corresponding concepts are called pathogen-associated molecular patterns (PAMPs), alias microbe-associated molecular patterns (MAMPs), and damage-associated molecular patterns (DAMPs). PTI broadens the response spectrum of plants undergoing an aggression, but it is not efficient enough in the presence of virulent pathogens producing PTI-suppressing effectors and even effectors capable of suppressing ETI. In the much-cited zig-zag model [16], the level of resistance left when ETI and PTI have been repressed is called basal resistance (BR).
Even though there are differences among ETI, PTI and BR, these three forms of resistance share the same signaling pathways and defense mechanisms, including the production of toxic reactive oxygen species (ROS), wall reinforcement and the production of hormones, salicylic acid (SA), jasmonic acid (JA), ethylene (ET), and abscisic acid (ABA), which interact among themselves to modulate the defense response.
To reduce the damage made by the most virulent pathogens, plants have developed the capacity to increase their immunity level by “memorizing” former aggressions. Priming helps them to increase the speed and intensity of defense responses when dealing with the pathogens again. Induced resistance (IR) is not the result of an evolutionary process but part of phenotypic plasticity and involves epigenetic processes. IR is more efficient than BR. It often expresses itself systemically, has effects lasting over days and sometimes weeks, and is characterized by a broad spectrum of efficiency against a very large range of biotrophic and hemi-biotrophic pathogens.
There are two major forms of IR: induced systemic resistance (ISR) and systemic acquired resistance (SAR). ISR is triggered by beneficial microorganisms associated with the roots, such as plant growth promoting rhizobacteria (PGPR), plant growth promoting fungi (PGPF), and arbuscular mycorrhizal fungi (AMF). ISR depends on the JA and ET pathways [6] and it is basically efficient against necrotrophic pathogens and pests of the arthropod class [7].
SAR depends on SA and an SA receptor specific protein, NPR1 (for Nonexpresser of pathogenesis-related genes1). The activation of SAR results in priming of plants and the direct activation of numbers of defense mechanisms, including the production of pathogenesis-related (PR) proteins. SAR provides long-lasting protection against a large range of biotrophic and hemi-biotrophic pathogens [7]. Plant resistance inducers used for crop protection generally supply SA or analogs of SA, or target the activation of the SA pathway.
3. High Concentrations in SA Are also Associated with Increased Tolerance of Plants towards Abiotic Stresses
Tolerance against abiotic stress is understood here not as the capacity to survive in extreme adverse conditions, but the aptitude to maintain crop performance to an acceptable level when conditions deteriorate, or in the presence of pollutants such as so-called heavy metals. Depending on the crops, there is a more or less strong correlation between yield and growth (defined as accumulation of biomass). Considering that growth depends on the carbon budget and the latter on photosynthesis, it is safe to consider that photosynthesis generally determines yield. Photosynthesis also determines all quality criteria associated with carbon accumulation, such as fruit and grain size, dry matter and sugar content of harvested organs. Now, does increased SA, be it applied exogenously or triggered in plants, increase photosynthesis, growth, yield and quality of production? If so, under which conditions, and through which physiological mechanisms?
3.1. High SA Exerts Positive Effects on Photosynthesis of Stressed Plants
SA treatments seem to be systematically credited with positive effects in conditions of stress. There are many observations showing, for instance, positive effects of SA treatments on Rubisco and phosphoenolpyruvate carboxylase (PEPC) activity in maize plants submitted to cadmium stress [17], or on the net photosynthetic assimilation rate of wheat [18] and cucumber [19] in conditions of high salinity. Rubisco and PEPC are the two major enzymes of the dark reactions of photosynthesis. It was observed that SA applications do not improve grapevine photosynthesis in standard conditions, but that they do so in conditions of heat stress [20].
The effects of endogenous levels of SA on photosynthesis were reviewed in the literature [21], notably exploiting observations made on Arabidopsis or Populus mutants in stress conditions conducive to the synthesis of SA. In Arabidopsis, mutants with high levels of SA can display decreased net photosynthesis in addition to other parameters linked to photosynthesis, which is consistent with the SA-associated dwarfing symptoms observed by [22]. The level of light influences responses, arguably by determining the risk of photooxidative stress, i.e., the risk of production of toxic ROS. The inhibiting effect of high levels of SA on photosynthesis is less marked in conditions of low than of high levels of light. It was found that the induction threshold of Apx2 is lower in high SA mutants of Arabidopsis, which is consistent with better protection against photooxidative stress [22]. Apx2 encodes for a cytosolic ascorbate peroxidase, which plays a key role in the scavenging of ROS, which are typically produced in conditions of high light, when the imbalance increases between the amount of energy absorbed under the form of photons and the amount of energy used by photosynthesis under the form of electrons. One major hypothesis currently gaining momentum is that SA exert a protective effect against stress by stimulating antioxidant responses. In this respect, it is revealing that SA treatments were found to reduce toxicity of certain pesticides as it was shown in cucumber [23]. The toxicity of pesticides in plants is often attributed to oxidative effects. There is a wealth of scientific evidence that SA stimulates the activity of antioxidant enzymes (superoxide dismutase, peroxidases, catalase) or enzymes of antioxidant systems, such as glutathion reductase. See, for instance, Dong et al. on this topic [24].
3.2. High SA Exerts Positive Effects on Growth and Yield of Stressed Plants
The literature is nearly unanimous: exogenous applications of SA have beneficial effects on growth and yield in conditions of stress (Table 1).
3.3. High SA May Exert Positive Effects on Quality of Production
There are not many observations on the effect of SA on the quality of production. It was observed that low doses of SA stimulate translocations of sugars from leaves to fruits in pepper grown in conditions of moderate salinity [34]. Harvested fruits were also found to be richer in ascorbate and carotenoids. It must be noted that a stimulating effect of SA on translocations could explain its positive effect on photosynthesis, besides other mechanisms, since translocations, by emptying leaves of the sugars synthetized, prevent their accumulation to downregulate photosynthesis [35]. This, again, contributes to lowering the risk of photooxidative stress associated with reduced photosynthesis.
3.4. High SA Increases Plant Tolerance to Exceedingly High Temperatures
Depending on intensity, duration of exposition to unfavorable temperatures, as well as the rate of temperature increase or decrease, heat and cold stress, can more or less severely impact growth, development and crop performance [36,37].
High temperatures can result in denaturation and disassembling of proteins, or in an increase in membrane fluidity [36]. Some plants, when submitted to moderate heat stress, can acclimate progressively at the metabolic and cellular levels [37]. Thermotolerance will allow them to tolerate extreme temperatures that otherwise would be damaging. Adaptive responses include changes in phospholipid composition of membranes, activation of antioxidant systems under hormonal control, notably SA, and the production of heat shock proteins [36,37,38]. It was more specifically observed that exogenous applications of SA increase activity of antioxidant enzymes and reduce the deleterious effects of high temperatures [38].
3.5. High SA increases Plant Tolerance to Exceedingly Low Temperatures
See Saleem et al. for a recent review [39]. Low temperatures provoke an excessive excitation of electron transport systems, therefore increasing the risk of production of ROS. If controlling mechanisms do not eliminate ROS efficiently, membrane lipids, proteins and nucleic acids can be damaged and cell death may occur [40]. Temperatures close to zero and temperatures well below zero do not affect temperate species similarly. Near to zero temperatures, generally, are at the origin of reversible physiological disorders, whereas severe frost can be at the origin of the formation of ice crystals in cells. Notably, frost during springtime can be extremely harmful to fruit tree production and viticulture because it is concomitant with bud burst and the appearance of flowers. Frost can damage ovaries and entail the destruction of flowers and small fruits; even young developing leaves and vegetative buds can be damaged to the point of full destruction. Ice crystals are mainly damaging when they form inside cells, less so when they appear in the apoplast. When the rate of temperature decrease is not too fast and is limited to the near zero zone, ice forms only in the apoplast and plants can often deal with cold conditions. SA can play a positive role by stimulating the activity of antioxidant enzymes and the accumulation of protective proteins such as ice-binding proteins [41] in the apoplast [42,43,44,45]. Observations made on numerous species have shown that moderate cold results in an increase in SA endogenous concentrations in plants, and that exogenous SA treatments can supply protection against moderate forms of cold stress [46,47,48]. By using stress to stimulate SA production and paclobutrazol, an inhibitor of SA synthesis, it was shown that high levels of endogenous SA are required for cold resistance in cucumber [21].
3.6. High SA Can Help Crops to Overcome the Consequences of Frost
In conditions of severe frost, when ice crystals form inside cells, cell death may occur and primary buds can be completely destroyed. In certain species, such as grapevine, secondary buds can take over, securing minimal yield. Flashes of UV-C light, which are believed to stimulate the production and accumulation of SA in plants, might be able to stimulate secondary bud burst in grapevine after a severe frost episode [49]. This hypothesis is consistent with the findings of Orrantia-Araujo et al., who demonstrated that SA was involved in dormancy breaking in grapevine [50].
3.7. SA Acts as a Critical Hormone in Improving Uptake and Status of Mineral Nutrients under Stress Conditions
Mineral nutrition is essential for the survival of plants and crop performance in both unstressed and stressed conditions. It can have a mitigating effect on abiotic stress [39]. Sheteiwy et al. found that SA seed priming improved K and Na uptake in rice submitted to high salinity conditions [51]. SA application inhibited the uptake of Pb and partially reversed Pb-induced alterations in Mn, Ca and Fe content in leaves of Vallisneria natans (Lour.) Hara exposed to 50 μM Pb [52].
3.8. High SA Stimulates Flowering
The role of SA in the stimulation and regulation of flowering has been long known. It was observed that SA fosters formation of floral buds in tobacco calluses [53]. Since their pioneering work there have numerous studies on wild plants and SA mutants, which have confirmed the role played by SA in flower induction [54]. This role of SA in flower induction can be observed in stressed as well as in unstressed plants [55]. Plants submitted to UV-C light produce more SA and activate their flowering program earlier than control plants [56,57].
4. The Dark Side of High SA
Considering all the benefits associated with high SA levels in plants and the stimulation of the SA signaling pathway, it does not come as a surprise that SA itself, SA analogs such as acibenzolar-S-methyl (ASM, see below), and inducers of the SA signaling pathway have attracted a lot of attention from the industry as well as from researchers [4]. Unfortunately, SA, SA analogs and most chemical PRIs can exert negative effects on growth and yield.
In the absence of stress, exogenous supplies of SA can exert negative effects such as decreasing stomatal conductance [58,59,60]. Since the process allowing evacuation of the energy in excess under the form of heat, known as non-photochemical quenching (NPQ), may also be reduced [61], there can be an increase in the risk of photodamage [62]. In the long term, a decrease in Rubisco activity can be observed [58], or even down-regulation of Rubisco synthesis or a modification of the isoforms [63]. High levels of SA may also exert inhibiting effects on the electron flux at the photosystem I level and have been observed to be associated with a decline in cytochrome f554 at the thylakoids level. Consistent with the inhibiting effects of high SA treatments on photosynthesis, and their damaging effects to the photosynthetic machinery, their effects on growth and yield are not necessarily positive ones in the absence of stress [25]. They could even be negative, as observed in maize [17]. Phytotoxicity and growth reduction have been observed with ASM treatments on tobacco, tomato, pepper, onion and Amaranthus [64,65,66,67,68]. Such effects may be attributed to the allocation of resources to defense at the expense of growth [4,69]. However, these negative effects are not systematically observed [70]. Moreover, observations made on Populus mutants do not fully support the findings on annuals; they suggest that high SA levels do not necessarily result in lower photosynthesis and growth of ligneous plants, whereas the mitigating effects against stress consequences seem to be conserved [71]. This difference may be attributed to the existence of reserves in woody plants that can be mobilized in case of higher demand for defenses.
5. A Review of the Existing Means of Increasing SA in Plants and Crops
Since high SA seems generally desirable in plants, numerous solutions have been tested in research and by the industry. They fall into three major categories: (1) SA and its analogs and other molecules stimulating the endogenous synthesis of SA, (2) biological PRIs, and (3) physical PRIs. Interestingly, it was suggested that exogenous applications of SA on pea seeds can stimulate endogenous synthesis in plants [72].
5.1. Analogs of SA and Chemical Inducers/Stimulators of the SA Signaling Pathway
White observed that acetyl-salicylic acid induces resistance to tobacco mosaic virus in tobacco plants [5]. Since his pioneering work, numerous SA-analogs have been designed and tested, showing a high interest in them from both the researchers and the industry. For a recent review, synthetic functional SA analogs activate plant defense via SA-mediated signaling [4]. It has been established that SA-analogs such as INA (2,6-dichloroisonicotinic acid) and ASM (acibenzolar-S-methyl, a benzothiadiazole) are SAR inducers [73] and therefore potent stimulators of plant defenses against a large range of pathogenic microorganisms. ASM has notably been reported to be effective against plant diseases caused by fungal, bacterial and viral agents; to name only a few among many references, see, for instance, [3,64,66,74,75,76].
Besides SA and analogs of SA, the major documented chemical PRIs against fungal diseases are nearly all based on, or at least have links with, the SA signaling pathway (Table 2). These chemicals encompass chitin and chitosan, the phosphite ion (Phi), laminarin and sulfated laminarin, the chitooligosaccharides (COS) and oligogalacturonides (OGA), plant and yeast extracts, and some essential oils (Eos); see below.
5.2. Chitin and Chitosan
Chitin is a key component of the cuticle of insects, the exoskeleton of crustaceans and the cell wall of fungi and of some algae. Formulation problems are currently addressed by the design of complexes associating chitin with iron, for instance, under the form of nanoparticles. Chitin oligomers form MAMPs that induce PTI responses when recognized by plant cells. Chitin can be supplied to leaves or roots, as well as to harvested organs and seeds. For recent reviews, see [77,78].
Chitosan is a polymer resulting from the partial deacetylation of chitin. It generates a lot of interest in the scientific community for being safe, declining rapidly and being easily combined with other compounds or elements, such as SA, harpin or copper, notably in the form of nanoparticles. Besides exerting direct inhibiting effects on the germination and growth of some fungi, chitosan can also stimulate plant defenses and increase the expression of genes associated with resistance against fungal diseases. Chitosan induces callose deposition and the expression of genes coding for PR proteins and NPR1, which is pivotal in SAR. In addition to interacting with SAR, chitosan was found to be involved in the JA/ET signaling pathway in gray mold resistance of grapevine [79]. Observations of clear-cut disease controlling effects of chitosan are, however, lacking for field conditions, suggesting that there is still some way to go before chitosan can be considered as a major tool in the arsenal of substances and techniques that can be used to reduce or replace fungicides.
On the other hand, and by contrast with ASM, chitosan has a clear positive effect on the growth and development of plants, and consequently on related crop performance parameters such as seed germination rate, yield and quality of production, in standard as well as in stress conditions [80].
5.3. Phosphites
The H2PO3− ion (Phi) represents the reduced form of inorganic phosphate that must not be confused with the phosphate ions HPO42− and H2PO4−. The most common forms are potassium phosphonate and sodium diphosphonate. Fosetyl-Al is an ethyl phosphonate marketed under numerous brand names. Despite having a very limited impact on the environment, Phi is not on the list of approved substances by the International Federation of Organic Agriculture Movement. Besides being a fungistatic and fungicide, Phi is an inducer of plant defense, with demonstrated stimulating effects on the production of phytoalexins and chitinases, as well as wall strengthening [81,82]. Studying the Phytophthora cinnamomi/Arabidopsis thaliana pathosystem, Eshraghi et al. found that Phi induced genes tightly associated with the JA and the ET signaling pathways, such as PDF1.2 and THI2.1 [83]. Phi was found to be at the origin of an increase in the expression of defense genes against leaf rust in Arabidopsis, such as PR2 (β-1,3-glucanase), PR3, PR4 and PR8 [chitinases], and PR5 [thaumatin], as well as of genes associated with the JA signaling pathway [84]. Phi was observed to downregulate MAP kinase MPK4 in the Hyaloperonospora arabidopsidis/A. thaliana pathosystem [85]. MPK4 is a negative regulator of the defenses induced in the SA signaling pathway, suggesting that Phi could positively influence the latter.
The effect of Phi on plant growth and crop performance has been studied, first of all with the objective of assessing its role as a source of phosphorus in conditions of phosphorus limitation. It is now well assessed that Phi cannot be used as a source of phosphorus. Moreover, Phi does not have any beneficial effect on the growth of healthy plants; it may even exert negative effects depending on the phosphorus status of plants [86].
5.4. Laminarin and Sulfated Laminarin
Laminarin is a polymer of β-1,3-glucanes extracted from Laminaria digitata, a brown algae. It was found to be effective against grey mold and downy mildew in grapevine [87]. Laminarin has been approved against powdery mildew in grapevine and strawberry, scab and fire blight in apple, powdery mildew, foot rot, and leaf rust in field crops, among others. Laminarin mechanisms of action have been investigated for wheat [88] and grapevine [87]. It was found that hydrogen peroxide accumulates in the sites of potential penetration by the pathogen, and that lipoxygenase (LOX) activity increases. LOX is involved in the JA signaling pathway. Sulfated laminarin is considered as more effective than laminarin [89]. The former seems incapable of inducing the production of ROS, but is at the origin of lasting depolarization of plasmic membranes in grapevine cells while initiating SA-dependent defense mechanisms, but after inoculation by Plasmopara viticola.
Laminarin seems all the more interesting for crop performance in stress conditions; it can promote plant growth and enhance tolerance to heat and salt stress [90]. Laminarin-improved tolerance against stress is, among other things, associated with enhanced antioxidant activity in the chloroplasts. The way plants treated with laminarin arbitrate between the needs for defense against pathogens and those for growth under conditions of abiotic stress would need to be investigated in detail.
5.5. COS-OGA
The association of oligomers of chitooligosaccharides (COS) and oligogalacturonides (OGA) was approved for treatments of cucumber, strawberry, pepper, tomato and grapevine against powdery mildew [91]. It is also approved against downy mildew in grapevine. Whereas the COS mimic the chitooligosaccharides produced by the lytic activity of plants reacting to the aggression by the pathogen, the OGA mimic the oligogalacturonides produced by the lytic activity exerted by the pathogen on plant cell walls [92]. It has been observed in tomato that repeated applications of COS–OGA resulted in an increase in leaf concentration in SA. COS–OGA also results in an overexpression of genes and an accumulation of proteins associated with the SA signaling pathway, whereas genes associated with the JA/ET pathway are not affected. Interestingly, COS–OGA treatments seem to be at the origin of the production of ROS [93], which could well reinforce the stimulating effect on resistance mechanisms according to the crosstalk theory [94]. Being of natural origin, COS–OGA can be used in organic farming.
5.6. Plant and Yeast Extracts
There have been only a few attempts to use plant extracts with stimulating properties of plant defenses. However, there are many examples where added plant biomass of allelopathatic plants such as Coronopus didymus, Anagallis arvensis, Whithania somnifera and Datura metel released allelochemicals during leaching and decomposition, resulting in the activation of defense-related enzymes and successfully controlled diseases caused by Ascochyta rabiei [95], Macrophomina phaseolina [96] and Sclerotium rolfsii [97,98].
So far, not many plant and yeast extracts have made it to the market. The extract of Reynoutria sachalinensis (a Polygonaceae) was observed to be efficient against such diseases as powdery mildew. It was found that the effect of this extract was linked with the SA signaling pathway [99]. Crushed fenugreek seeds (Trigonella foenum-graecum) were found to be effective against powdery mildew in grapevine and melon, but it was demonstrated that they induce the release of p20 caspase-1 fragment, suggesting the involvement of the NLRP3 inflammasome [100]. It was even observed that crushed fenugreek seeds can induce cell death and, moreover, exert toxic effects on Zebrafish larvae [100].
Yeast wall extracts (more specifically Saccharomyces cerevisiae LAS117) were found to stimulate both the SA and the JA/ET signaling pathways, which explains why they have an efficacy against powdery mildew, downy mildew and gray mold [101]. This specific yeast extract was, moreover, found to act as a biostimulant of photosynthesis.
5.7. Essential Oils
Essential oils (EOs) constitute secondary metabolites, terpenoids and phenolic compounds, some of which are known to play direct and indirect (signaling) roles in defenses against pathogens and aggressors. EOs do not act exclusively as elicitors. It may even be said that their eliciting properties probably represent a minor aspect of their mode of action. Some EOs exert direct toxicity against insects by inhibiting P450 cytochromes, associating with GABA receptors, inhibiting acetyl-choline esterase or activating receptors of octopamine, a neuromodulator [102]. There are numerous references, essentially descriptive in nature, about the antifungal effects of different EOs, notably in post-harvest conditions; see, for instance, [103,104]. The purely eliciting properties of EOs are not much documented. Melaleuca [tea tree] EO exerts a stimulating effect on the synthesis of phenolic compounds in lettuce [105]. The eliciting properties of EOs are arguably attributable to the methyl-salicylate many of them contain. Gaultheria EO is made of 99.96% methyl-salicylate [106]. Gaultheria EO induced resistance of Arabidopsis thaliana against Colletotrichum higginsianum through the SA signaling pathway [107].
EOs can be used by organic farmers, but their efficacy seems variable and not very lasting. There are not many commercial products, probably because agreement procedures are lengthy [102]. Recently, sweet orange EO (several brands) emerged as especially promising, notably against fungal diseases. Sweet orange EO contains limonene, for which references of efficacy in the scientific literature are lacking, in contrast to naringenin [108].
So far, the interest of EOS does not seem to lie so much in their defense stimulating effects than in their potential for reducing disease pressure by exerting direct fungicidal effects. Moreover, their phytotoxic effects can make them difficult to use on crops.
5.8. Biological PRIs
Non-pathogenic microorganisms, saprophytes or non-virulent strains of pathogenic agents, are used as biocontrols; they are either sprayed on leaves or used in a drench to reach the roots. Bacillus subtilis and Trichoderma harzianum are arguably the most common microorganisms used to protect crops. They act by mycoparasitism, antibiosis and competition for nutritive resources, therefore exerting a direct antifungal effect. They display a large range of action that includes, in addition to fungi, pathogenic oomycetes, bacteria and viruses. Moreover, these microorganisms are elicitors of local and systemic induced resistance [109]. Interactions between these microorganisms and their host plants is at the origin of the synthesis and accumulation of peroxidases, chitinases, glucanases, phenylalanine and hydroperoxide lyases, and phytoalexines [110]. Studies have confirmed that the SA and JA signaling pathways are involved and regulate the interactions between pathogens and plants [111,112].
Several biological PRIs have been shown to be promoters of plant growth and development. This is notably the case for Trichoderma [113]. Enhancement of growth by Trichoderma in conditions of stress is linked to a stimulating effect on the antioxidant and ROS scavenging mechanisms, as shown in tomato plants submitted to water stress [114]. Epigenetic processes are probably involved in the heritable plant growth promoting and plant resistance stimulation effects of Trichoderma [115]. It can be beneficial to use microorganisms in association, as in the case of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 in the wheat/Fusarium graminearum pathosystem [116]. Interestingly, better disease control was found to be compatible with enhanced growth [116].
5.9. Physical PRIs
Physical PRIs (light and mechanical stress, mainly) have unique features: they can be applied in field conditions even in the presence of rain and wind, they do not need to be formulated, and they do not leave any residues on plants or in the soil. Lengthy procedures of homologation do not apply to physical PRIs, basically because they do not exert any toxic effects on humans and the environment. All the same, some of them, UV radiation, notably, have to be applied by following safety rules. However, these rules are easy to define and follow.
Mechanical stress, involving wounding or not, can stimulate plant defenses against a large range of necrotrophic pathogens and herbivores [117]. There are few observations of the differences existing between the different forms of mechanical stress, and the role of treatments intensity and frequency on efficacy and persistence of effects. The systemic effect of treatments has not been described. Coutand made observations about the effect of imposed curvatures and concluded that treatments repeated every 7 to 10 days are sufficient to induce resistance [117]. Since all studies have been conducted by comparing treated plants with unruffled controls, it remains to be assessed whether or not mechanical stress is still effective in field conditions where plants are nearly continuously agitated under the effect of wind spurs irregular in frequency, intensity and direction. Oxidative stress [118,119] and the JA [120,121] and ethylene [122] signaling pathways have been found to be involved, but otherwise not much is known about the underlying physiological mechanisms. The implication of the SA signaling pathway has not been investigated to our knowledge, possibly because studies about mechanical stress have been conducted on necrotrophic fungi [117]. It has been recognized for many years that mechanical stress can result in growth reduction [122,123]. In the case of some horticultural crops, mechanical stimulation has, indeed, a true potential for replacing plant growth regulators for control of plant size and quality [124].
The potential of some wavelengths to act as physical PRIs is now acknowledged [125], which includes blue and red light as well as UV-B and UV-C radiations [126,127]. Pulsed light was found to dramatically increase fruit concentrations in secondary metabolites in mango [128] and to stimulate plant defense against Fusarium pallidoroseum in melon [129]. Pulsed light consists of flashes of light of 300 to 500 μs, encompassing wavelengths from 200 to 1100 nm, but it is generally believed to act thanks to its high proportion of UV-C light. There is some evidence that blue photoreceptors are involved in the regulation of defense responses. Cryptochrome 1 (CRY1) was found to positively regulate SA-induced PR genes involved in the resistance of Arabidopsis thaliana L. against Pseudomonas syringae [130]. Cryptochromes are photoreceptors of UV-A and blue radiations with maximal action wavelengths at 370 and 450 nm. The blue-light photoreceptors, cryptochrome and phototropin 2, are required for the stability of protein HRT and thereby resistance to Turnip Crinkle Virus [131]. Protein HRT is involved in an SA signaling pathway independent of NPR1. UV-B radiation is proven to be a positive regulator of plant defenses through both JA-dependent [132,133] and JA-independent pathways [134]. On the dark side of UV-B light there is a well-documented negative impact on photosynthesis and growth [135]. UV-C light has even a better record of defense stimulating effects than UV-B light, well-documented in some species such as tomato against Alternaria alternata [136] and Botrytis cinerea [137,138], carrot and pepper against B. cinerea [139,140], and lettuce against B. cinerea and Sclerotinia minor [141]. UV-C light was also observed to stimulate plant defenses of tobacco against tobacco mosaic virus [56]. In their study they clearly attributed this effect with an increase in SA. This is nearly all that is known from the mechanisms of action of UV-C light. The effect of UV-C light on plant immunity and mechanisms of resistance to abiotic stresses are well described but not well understood [127,142]. See Figure 1.
5.10. Flashes of UV-C Light, a Newcomer among PRIs with Unique Features
The pioneering observations of Aarrouf and Urban have established that flashes of UV-C light stimulate defenses of lettuce and tomato against B. cinerea, pepper against Phytophthora capsici and grapevine against Plasmopara viticola better than conventional exposures for the same amount of energy delivered and the same wavelength, paving the way for the use of flashes of UV-C as a physical elicitor in cropping conditions [143]. Since then, confirming evidence has been accumulated that flashes of UV-C light strongly stimulate crop defenses in conditions of commercial production. Strong reductions in disease symptoms were observed on strawberry inoculated by powdery mildew in greenhouses similar to commercial greenhouses [144] and on grapevine inoculated by either powdery or downy mildew in regular vineyards [145].
The mechanisms of UV-C perception and of the signaling, regulatory and metabolic pathways involved downstream are not fully elucidated, even though there is some evidence that flashes of UV-C light increase SA content in plants [127,142].
Interestingly, all the plant resistance inducing effects of flashes of UV-C light were obtained at “hormetic doses”, i.e., at doses previously assessed as being harmless for plant integrity and growth. In other words, effective doses of UV-C light can be defined that do not exert any negative effects on plant photosynthesis and growth and crop performance, even in unstressed plants [144,145]. Flashes of UV-C light therefore appear superior to all the PRIs that negatively impact the photosynthesis and growth of unstressed plants, such as ASM, mechanical stress, UV-B light or phosphites.
6. Combining PRIs: Playing Complementarity or Reinforcement Effects?
It is generally accepted that the crosstalk between the SA and the JA/ET signaling pathways represents the backbone of the signaling network leading to plant immunity [146]. It has been found, for instance, that both the SA and the JA signaling pathways are involved in grapevine response to Plasmopara viticola and Botrytis cinerea, whereas the ET signaling pathway is, moreover, involved against Erysiphe necator (Table 3).
It may therefore seem tempting to simultaneously stimulate the SA, the JA/ET and possibly also the β-amino butyric acid (BABA) signaling pathways. It was found that the association of ASM and Bacillus subtilis strain B4 was efficient for controlling anthracnose in cucumber [152]. Faba bean chocolate spot disease was successfully controlled using ASM and Trichoderma harzianum in association [149]. Bacterial canker in kiwifruit was also controlled using ASM and Aureobasidium pullulans strain CG163 [153] jointly. ASM and BABA, each at half the recommended rate, provided an additive effect against Plasmopara viticola, and performed as well as the full rate of each compound alone [154]. Flashes of UV-C light (a stimulator of the SA pathway) controlled powdery mildew in strawberry better than sulfated laminarin (basically a stimulator of the JA pathway, even though stimulation of the SA pathway exists after inoculation), and their combination was even more effective than flashes of UV-C light alone [153].
Walters et al. tested a combination of ASM, BABA and cis-jasmone to control powdery mildew and leaf scald in two spring barley varieties under field conditions [155]. They observed an increase in levels of Ramularia leaf spot as a negative side-effect of their treatments. Moreover, they found in a greenhouse trial that treatments resulted in an up-regulation of the SAR marker gene PR1-b and to a concomitant and substantial down-regulation of LOX2, a gene involved in biosynthesis of JA. They hypothesized that the elicitor combination activates SAR in barley, but represses the JA signaling pathway. So far, scientific evidence for additive or synergic effects of combinations of elicitors remains scarce. The observations even suggest that counterproductive effects may be observed depending on pathogens [156].
Surprisingly, there are some references about positive effects of combinations of elicitors that one would not expect to be effective, considering that the elicitors duplicate themselves: MeJA + T. harzianum against Bipolaris sorokiniana in wheat [157], and MeJA + T. harzianum against Fusarium oxysporum in tomato [158]. However, again, such references are scarce. Moreover, they are all only about JA, not SA.
7. Reducing Disease Pressure
Reducing disease pressure represents an essential step to take when disease pressure is too high relative to the capacity of plants to produce the compounds and structures required for efficient control of disease development. Besides conventional fungicides, copper or sulfur, there are numerous substances that have fungicidal effects such as phosphites (Phi), which are also plant nutrients, essential oils and the so-called basic substances that comply with European Regulation (EC) No 1107/2009. Basic substances include lecithin, fructose, dairy milk, whey, sodium chloride, sodium bicarbonate, chitosan hydrochloride, onion (Allium cepa L.) bulb extract and macerates of plants such as willow (Salix spp.), horsetail (Equisetum arvense L.) and nettle (Urtica spp.), among others [159]. Physical methods of surface disinfection may be used in addition or as alternatives to chemical substances.
7.1. Chemical Substances with Antifungal Properties
Scientific references about efficacy or modes of action exist only for Phi, some essential oils and macerates, lecithin, chitosan and UV light. Phi has a recognized efficacy for control of the diseases caused by some bacteria and oomycetes of the Peronospora, Plasmopara, Phytophtora and Pythium genera [160], and fungi such as Phoma costarricensis [161], Colletotrichum gloeosporioides [162], Hemilia vastatrix [163], Fusarium solani [164] and Alternaria alternata [155]; see also the review which describes Phi as excellent fungicides [165]. It was observed that essential oils of Citrus sinensis, Cupressus sempervirens, Eucalyptus globulus, Lavendula angustifolia, Pelargonium graveolens, Pogostemon patchouli, Mentha piperita, Rosmarinus officinalis, Santolina chamaecyparissus, Syzygium aromaticum, Thymus mastichina and T. vulgaris inhibited the mycelial growth of Fusarium oxysporum, Phytophthora parasitica and Sclerotinia sclerotiorum [166]. The regular foliar sprays of horsetail macerate reduce late blight (Phythophtora infestans) and brown rust (Puccinia triticina) symptoms in tomato and durum wheat, respectively [167]. The mode of action of lecithin against several causal agents of plant diseases has been investigated to some extent. Studying powdery mildew in cucumber, it was observed that soy lecithin disturbs hyphal elongation and penetration in host plants of Sphaerotheca fuliginea [168]. This was confirmed by observations of the effect of lecithin on the morphology of Pyricularia oryzae [169]. Homma et al. found that lecithin is at the origin of hyphal anomalies, which consequently is an inhibitor of appressorium formation and penetration in the epiderm. Antiviral effects of lecithin were recently observed [170]. Lecithin being an emulsifier, it is used as an additive in many treatment products. Lecithin-based encapsulation was found to increase the efficacy of resveratrol against B. cinerea [171]. The antimicrobial properties of chitosan have attracted a lot of interest for many years because they seem to be associated with a complete lack of toxicity [172]. Although chitosan and chitosan-based polymers are involved in the stimulation of plant defenses [78], other reviews clearly deal with the direct antimicrobial properties of chitosan and its derivatives. See, for instance, the recent review [77].
7.2. Physical Methods of Surface Disinfection
The inhibitory and damaging effects of UV light on bacteria have been described and exploited for many years. This includes pulsed light, whose surface disinfecting properties are attributed to the high amount of UV, notably UV-C radiations, it supplies [173]. Absorption of UV radiations by conjugated carbon–carbon double bonds in proteins and nucleic acids may result in protein polymerization, enzyme inactivation and DNA structural changes [174,175]. Wuytack et al., moreover, stated that pulsed light induces abnormal ion flow, increased cell membrane permeability and depolarization of the cell membrane in Salmonella enterica [176]. Whereas UV-A and UV-B radiations are believed to exert lethal effects, mainly as a result of membrane damage and the formation of peroxides, UV-C radiation in the 250–260 nm range is often credited as carrying most of the bactericidal effects of UV light by damaging cellular DNA via pyrimidine dimer formation [174]. UV-C radiation is generally more effective at destroying bacteria and at lower doses than UV-B and UV-A [177]. It must, however, be stressed that nearly all articles found in the scientific literature about UV-B light are based on observations made using mercury vapor lamps with an emission peak around 310 nm, while it is established that the latter is much less absorbed by DNA than 280 nm [178].
Recent observations suggest that UV-C light can be used not only for food disinfection purposes against bacteria but also against pathogenic fungi affecting crops such as powdery mildew in cucumber [179] and strawberry [180], at doses that are compatible with normal plant functioning and crop performance. The highest efficacy was found for nighttime treatments of diseased strawberry plants, arguably because the light-dependent mechanisms of DNA repair were prevented from countering the damaging effects of UV-C light. Recent observations suggest that UV-C light can be used not only for food disinfection purposes against bacteria, but also against pathogenic fungi affecting crops such as powdery mildew in cucumber [179] and strawberry [180], at doses and frequencies that are compatible with normal plant functioning and crop performance. In fact, these doses are about ten times lower than the doses recommended for stimulating plant defenses. The highest efficacy was found for nighttime treatments of diseased strawberry plants, arguably because the light-dependent mechanisms of DNA repair were prevented from countering the damaging effects of UV-C light.
Based on these considerations, it is quite obvious that the lamps that are used for delivering high doses of UV-C light can also be used at lower doses for crop surface disinfection purposes; what can do more can do less. In greenhouses it would probably be smart to combine disinfecting treatments at low dose/high frequency with stimulating treatments at high dose/low frequency, using the same high-energy lamps.
8. Ensuring That Plants Have the Resources Required for Synthesis of Defense Compounds and Structures
Decreasing disease pressure by exploiting the antifungal and disinfecting properties of some PRIs is certainly recommendable. Even so, and even when stimulation of plant resistance by PRIs is maximized, the latter may appear not efficient enough in conditions of very high disease pressure or for a period of time of sufficient duration for successful completion of the cropping cycle.
It has been hypothesized that efficient and lasting resistance of plants is conditioned by the level of carbohydrate resources that can be mobilized for synthetizing the compounds and structures required by defense. Resources and, notably, carbohydrates play a pivotal role in plant–pathogen interactions [181]. Both plant and pathogen compete for utilizing [host] resources. Calonnec et al. have supplied arguments in favor of the idea that high carbohydrate availability decreases the susceptibility of grapevine leaves towards Erisyphe necator, rather than being favorable to the development of the fungus [182]. In the case of other fungal pathogens, such as B. cinerea, the role of sugars may be less straightforward than for E. necator [183]. Ecologists know that plants must find trade-offs for resource utilization between growth and defense functions [184]. In natural conditions, plants are competing among themselves for access to light and must therefore maximize photosynthesis and growth. At the same time, they must be capable of defending themselves when threatened by pests or diseases. Surprisingly, researchers studying mechanisms of plant defenses have paid little attention to the issue of competition between growth and defense functions, even though it is probable that they determine the level of resources and, therefore, the duration of the resistance induced by PRIs [185,186]. PRIs that are at the origin of a priming effect have been given preference recently, since they do not tap resources at the expense of growth and yield as much as PRIs directly stimulating the metabolic pathways of plant defense [13,187]. Then, in the presence of a pathogen or an herbivore, plants must eventually synthetize defense compounds, and, therefore, consume resources, anyway. It was, therefore, hypothesized that resource availability represents the most important factor limiting the development of PRIs as an alternative to pesticide use [14,185,186].
It would seem tempting to develop strategies for crop management that leave as much carbohydrate in plant organs, notably leaves, as possible for plant defense. Unfortunately, such strategies may come at a price since leaf carbohydrate accumulation can inhibit photosynthesis and increase the risk of photooxidative stress in leaves [35]. A decrease in leaf net photosynthesis may be at the origin of a decrease in growth and yield, and also may have a negative impact on the quality of production (fruit size, carbohydrate content, etc.), whereas photooxidative stress in leaves has been shown to impact the metabolism of nearby fruits [187].
Theoretically, it is possible to increase the carbohydrate level of leaves either by decreasing sink demand or increasing photosynthesis. Sink demand can be reduced by sink removal or the inhibition of sink activity. Removal of competing sinks is a common practice in viticulture and horticulture. Trimming of the vine, for instance, helps to control canopy development in vineyards. It is, however, necessary to assess whether this technique has a sufficient impact on the carbohydrate content of the remaining leaves as long as clusters are not trimmed themselves. Fruits are, indeed, very strong sinks [188,189] that accumulate carbohydrates nearly until the end of the cropping season. In the case of tomato crops, the removal of fruits eventually stimulates the development of the remaining fruits and may also result in an increase in vegetative growth [190], all of which tap carbohydrate resources in leaves. Sink activity per se can be controlled only in greenhouses, by reducing temperature setpoints for heating. This will, however, come at the price of a reduction in crop performance and an increase in the risk of gray mold development. Clearly, there are ways to influence the source–sinks relationships with the objective of leaving more carbohydrate resources in leaves, but strategies must be carefully designed to avoid the development of alternative sinks or a reduction in crop performance. Models of carbon gains and allocation could be used as part of advanced strategies of manipulation of source–sink relationships [191].
Since it is probable that the control of carbohydrates in leaves will require action on both ends, not only sink activity and volume, but also source activity, observations about the effects of foliar applications of nitrogen on photosynthetic capacity and activity would be required in the future. So far, the possibility of increasing photosynthesis by applying nitrogen directly to leaves has not been paid much attention. While there are several observations about the effects of foliar applications of nitrogen, for instance under the form of urea, on berry and juice composition (e.g., [192,193]), observations about their effects on photosynthetic capacity has not been made. Besides mineral sources of nitrogen, it would also be useful to consider biostimulants known for being sources of amino acids, peptides or proteins. The effects of some biostimulants on grape and wine quality were reviewed, but they did not provide any information about photosynthesis [194].
Eventually, it must be emphasized that abiotic stress deteriorates carbon assimilation by plants, therefore exerting a negative effect on resource availability, growth and the capacity of plants to produce defense compounds and structures. Since some PRIs, such as laminarin, chitosan, Trichoderma spp. and flashes of UV-C light (Table 2), can act as biostimulants (Figure 2), it would be interesting to assess their efficacy for stimulating defenses in conditions of stress and to test whether their efficacy is attributable to both a stimulating effect on plant defenses and on resource availability for synthesis of defense compounds and structures.
9. Conclusions
The idea is gaining credibility that it will be possible to control fungal diseases by using exclusively environmentally friendly techniques. This requires, however, the satisfactory addressing of the issue of variable efficacy of PRIs. We propose in this review to combine natural or ecofriendly substances with fungicide properties, physical means of plant surface disinfection, chemical, biological and physical PRIs alone or associated for potential additive or synergic effects on the SA and JA signaling pathways, biostimulants and crop management techniques. Clearly, the issue of plant defense and plant growth, especially in conditions of stress, needs to be addressed jointly. On the bright site of the upcoming challenges, many of the listed PRIs can be associated in cropping conditions; some of them even seem to do almost everything, from inducing plant defense to reducing disease pressure and promoting resource acquisition, notably in conditions of abiotic stress, which seems pivotal for strong and durable defenses. More field trials are nevertheless needed to obtain more references about the efficacy of some associations of means. There is also a more fundamental requirement for a better understanding of the interactions between growth and defense, and the arbiter role played by SA.
Author Contributions
Conceptualization, L.U., J.A. and F.L.; writing—original draft preparation, L.U.; writing—review and editing, L.U., J.A., F.L. and D.B.H.; supervision, L.U. and J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ma, C.-S.; Zhang, W.; Peng, Y.; Zhao, F.; Chang, X.-Q.; Xing, K.; Zhu, L.; Ma, G.; Yang, H.-P.; Rudolf, V.H.W. Climate warming promotes pesticide resistance through expanding overwintering range of a global pest. Nat. Commun. 2021, 12, 5351. [Google Scholar] [CrossRef] [PubMed]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2020, 283, 124657. [Google Scholar] [CrossRef]
- Marolleau, B.; Gaucher, M.; Heintz, C.; Degrave, A.; Warneys, R.; Orain, G.; Lemarquand, A.; Brisset, M.-N. When a Plant Resistance Inducer Leaves the Lab for the Field: Integrating ASM into Routine Apple Protection Practices. Front. Plant Sci. 2017, 8, 1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance—Salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- White, R. Acetylsalicylic acid (aspirin) induces resistance to tobacco mosaic virus in tobacco. Virology 1979, 99, 410–412. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Yassin, M.; Ton, J.; Rolfe, S.A.; Valentine, T.A.; Cromey, M.; Holden, N.; Newton, A.C. The rise, fall and resurrection of chemical-induced resistance agents. Pest Manag. Sci. 2021, 77, 3900–3909. [Google Scholar] [CrossRef]
- Launay, M.; Caubel, J.; Bourgeois, G.; Huard, F.; de Cortazar-Atauri, I.G.; Bancal, M.-O.; Brisson, N. Climatic indicators for crop infection risk: Application to climate change impacts on five major foliar fungal diseases in Northern France. Agric. Ecosyst. Environ. 2014, 197, 147–158. [Google Scholar] [CrossRef]
- ACTA. Stimulateurs de Défense des Plantes (SDP), Technique à l’étude. Guide Pour la Conception de Systèmes de Production Fruitière Economes en Produits Phytosanitaires; GIS et Ministère de l’Agriculture: Paris, France, 2014; pp. 104–106. [Google Scholar]
- Sandroni, M.; Liljeroth, E.; Mulugeta, T.; Alexandersson, E. Plant resistance inducers (PRIs): Perspectives for future disease management in the field. CAB Rev. Perspect. Agric. Vet.-Sci. Nutr. Nat. Resour. 2020, 15, 1–10. [Google Scholar] [CrossRef]
- Ripoll, J.; Urban, L.; Staudt, M.; Lopez-Lauri, F.; Bidel, L.P.; Bertin, N. Water shortage and quality of fleshy fruits—Making the most of the unavoidable. J. Exp. Bot. 2014, 65, 4097–4117. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.; Heil, M. Costs and trade-offs associated with induced resistance. Physiol. Mol. Plant Pathol. 2007, 71, 3–17. [Google Scholar] [CrossRef]
- Walters, D.R.; Fountaine, J.M. Practical application of induced resistance to plant diseases: An appraisal of effectiveness under field conditions. J. Agric. Sci. 2009, 147, 523–535. [Google Scholar] [CrossRef]
- Austin, C. Sunlight’s Influence on Grapevine Powdery Mildew: Direct Effects on Pathogen Development and Attendant Consequences of Canopy Management and Vineyard Variability. Ph.D. Dissertation, Cornell University, Ithaca, NY, USA, 2010. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Krantev, A.; Yordanova, R.; Janda, T.; Szalai, G.; Popova, L. Treatment with salicylic acid decreases the effect of cadmium on photosynthesis in maize plants. J. Plant Physiol. 2008, 165, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Arfan, M.; Athar, H.R.; Ashraf, M. Does exogenous application of salicylic acid through the rooting medium modulate growth and photosynthetic capacity in two differently adapted spring wheat cultivars under salt stress? J. Plant Physiol. 2007, 164, 685–694. [Google Scholar] [CrossRef]
- Miao, Y.; Luo, X.; Gao, X.; Wang, W.; Li, B.; Hou, L. Exogenous salicylic acid alleviates salt stress by improving leaf photosynthesis and root system architecture in cucumber seedlings. Sci. Hortic. 2020, 272, 109577. [Google Scholar] [CrossRef]
- Wang, L.-J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.-J.; Cheng, J.-S.; Luo, H.-B.; Li, S.-H. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Janda, T.; Gondor, O.K.; Yordanova, R.; Szalai, G.; Pál, M. Salicylic acid and photosynthesis: Signalling and effects. Acta Physiol. Plant. 2014, 36, 2537–2546. [Google Scholar] [CrossRef] [Green Version]
- Mateo, A.; Funck, D.; Mühlenbock, P.; Kular, B.; Mullineaux, P.M.; Karpinski, S. Controlled levels of salicylic acid are required for optimal photosynthesis and redox homeostasis. J. Exp. Bot. 2006, 57, 1795–1807. [Google Scholar] [CrossRef]
- Liu, T.; Li, T.; Zhang, L.; Li, H.; Liu, S.; Yang, S.; An, Q.; Pan, C.; Zou, N. Exogenous salicylic acid alleviates the accumulation of pesticides and mitigates pesticide-induced oxidative stress in cucumber plants (Cucumis sativus L.). Ecotoxicol. Environ. Saf. 2020, 208, 111654. [Google Scholar] [CrossRef]
- Dong, C.-J.; Li, L.; Shang, Q.-M.; Liu, X.-Y.; Zhang, Z.-G. Endogenous salicylic acid accumulation is required for chilling tolerance in cucumber (Cucumis sativus L.) seedlings. Planta 2014, 240, 687–700. [Google Scholar] [CrossRef]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.-J. Salicylic Acid Alleviates the Cadmium Toxicity in Barley Seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Wael, M.S.; Mostafa, M.R.; Taia, A.A.E.-M.; Saad, M.H.; Magdi, T.A. Alleviation of cadmium toxicity in common bean (Phaseolus vulgaris L.) plants by the exogenous application of salicylic acid. J. Hortic. Sci. Biotechnol. 2015, 90, 83–91. [Google Scholar] [CrossRef]
- Roustakhiz, J.; Saboki, E. Effect of salicylic acid on yield and yield component of grapevine (Vitis vinifera) under salinity stress condition. Int. J. Farming Allied Sci. 2017, 6, 39–42. [Google Scholar]
- Zamaninejad, M.; Khorasani, S.K.; Moeini, M.J.; Heidarian, A.R. Effect of salicylic acid on morphological characteristics, yield and yield components of corn (Zea mays L.) under drought condition. Eur. J. Exp. Biol. 2013, 3, 153–161. [Google Scholar]
- Tahjib-Ul-Arif, M.; Siddiqui, M.N.; Sohag, A.A.M.; Sakil, M.A.; Rahman, M.M.; Polash, M.A.S.; Mostofa, M.G.; Tran, L.-S.P. Salicylic Acid-Mediated Enhancement of Photosynthesis Attributes and Antioxidant Capacity Contributes to Yield Improvement of Maize Plants Under Salt Stress. J. Plant Growth Regul. 2018, 37, 1318–1330. [Google Scholar] [CrossRef]
- Qados, M.M. Effects of salicylic acid on growth, yield and chemical contents of pepper (Capsicum annuum L.) plants grown under salt stress conditions. Int. J. Agric. Crop. Sci. 2015, 8, 107–113. [Google Scholar]
- Karlidag, H.; Yildirim, E.; Turan, M. Exogenous applications of salicylic acid affect quality and yield of strawberry grown under antifrost heated greenhouse conditions. J. Plant Nutr. Soil Sci. 2009, 172, 270–276. [Google Scholar] [CrossRef]
- Azimi, M.S.; Daneshian, J.; Sayfzadeh, S.; Zare, S. Evaluation of amino acid and salicylic acid application on yield and growth of wheat under water deficit. Int. J. Agric. Crop. Sci. 2013, 5, 816. [Google Scholar]
- Munir, M.; Shabbir, G. Salicylic acid mediated heat stress tolerance in selected bread wheat genotypes of Pakistan. Pak. J. Bot. 2018, 50, 2141–2146. [Google Scholar]
- Elwan, M.; El-Hamahmy, M. Improved productivity and quality associated with salicylic acid application in greenhouse pepper. Sci. Hortic. 2009, 122, 521–526. [Google Scholar] [CrossRef]
- Urban, L.; Léchaudel, M.; Lu, P. Effect of fruit load and girdling on leaf photosynthesis in Mangifera indica L. J. Exp. Bot. 2004, 55, 2075–2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkindale, J.; Hall, J.D.; Knight, M.; Vierling, E. Heat Stress Phenotypes of Arabidopsis Mutants Implicate Multiple Signaling Pathways in the Acquisition of Thermotolerance. Plant Physiol. 2005, 138, 882–897. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.M.; Mur, L.A.J.; Wood, J.E.; Scott, I.M. Salicylic acid dependent signaling promotes basal thermotolerance but is not essential for acquired thermotolerance in Arabidopsis thaliana. Plant J. 2004, 38, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-J.; Li, S.-H. Salicylic acid-induced heat or cold tolerance in relation to Ca2+ homeostasis and antioxidant systems in young grape plants. Plant Sci. 2006, 170, 685–694. [Google Scholar] [CrossRef]
- Saleem, M.; Fariduddin, Q.; Janda, T. Multifaceted Role of Salicylic Acid in Combating Cold Stress in Plants: A Review. J. Plant Growth Regul. 2020, 40, 464–485. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2005, 126, 45–51. [Google Scholar] [CrossRef]
- Bredow, M.; Walker, V.K. Ice-Binding Proteins in Plants. Front. Plant Sci. 2017, 8, 2153. [Google Scholar] [CrossRef] [Green Version]
- Janda, T.; Szalai, G.; Antunocis, Z.; Horvath, E.; Paldi, E. Effect of benzoic acid and aspirin on chilling tolerance and photosynthesis in young maize plants. Maydica 2000, 45, 29–33. [Google Scholar]
- Griffith, M.; Yaish, M.W. Antifreeze proteins in overwintering plants: A tale of two activities. Trends Plant Sci. 2004, 9, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Tasgin, E.; Atici, O.; Nalbantoglu, B.; Popova, L. Effects of salicylic acid and cold treatments on protein levels and on the activities of antioxidant enzymes in the apoplast of winter wheat leaves. Phytochemistry 2006, 67, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Zhang, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Salicylic acid and cold priming induce late-spring freezing tolerance by maintaining cellular redox homeostasis and protecting photosynthetic apparatus in wheat. Plant Growth Regul. 2019, 90, 109–121. [Google Scholar] [CrossRef]
- Clarke, S.M.; Cristescu, S.M.; Miersch, O.; Harren, F.J.M.; Wasternack, C.; Mur, L.A.J. Jasmonates act with salicylic acid to confer basal thermotolerance in Arabidopsis thaliana. New Phytol. 2009, 182, 175–187. [Google Scholar] [CrossRef]
- López-Delgado, H.A.; Martínez-Gutiérrez, R.; Mora-Herrera, M.E.; Torres-Valdés, Y. Induction of Freezing Tolerance by the Application of Hydrogen Peroxide and Salicylic Acid as Tuber-Dip or Canopy Spraying in Solanum tuberosum L. Plants. Potato Res. 2018, 61, 195–206. [Google Scholar] [CrossRef]
- Zhang, B.B.; Guo, L.; Song, Z.Z.; Yu, M.L.; Ma, R.J. Effect of salicylic acid on freezing injury in peach floral organs and the expressions of CBF genes. Biol. Plant. 2017, 61, 622–630. [Google Scholar] [CrossRef]
- Matton, Y.; Sement, F.; Rouesné, B.; Ledermann, L.; Pascouau, N.; Charel, J.; Goncalves, H.; Urban, L.; Aaarrouf, J. Utilisation D’un Dispositif Mobile Délivrant Des Impulsions Lumineuses Sur Une Matière Végétale Pour La Prévention Contre Les Impacts Liés à Des Stress Abiotiques. European Patent PCT/EP2022/066721, 2021. [Google Scholar]
- Orrantia-Araujo, M.A.; Martínez-Téllez, M.; Rivera-Domínguez, M.; Hernández-Oñate, M.; Vargas-Arispuro, I. Changes in the Endogenous Content and Gene Expression of Salicylic Acid Correlate with Grapevine Bud Dormancy Release. J. Plant Growth Regul. 2020, 40, 254–262. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; An, J.; Yin, M.; Jia, X.; Guan, Y.; He, F.; Hu, J. Cold plasma treatment and exogenous salicylic acid priming enhances salinity tolerance of Oryza sativa seedlings. Protoplasma 2018, 256, 79–99. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Wang, P.; Hou, J.; Qian, J.; Ao, Y.; Lu, J.; Li, L. Salicylic acid involved in the regulation of nutrient elements uptake and oxidative stress in Vallisneria natans (Lour.) Hara under Pb stress. Chemosphere 2011, 84, 136–142. [Google Scholar] [CrossRef]
- Lee, T.T.; Skoog, F. Effect of substituted phenols on bud formation and growth of tobacco tissue and vineyard conditions. Physiol. Plant. 1965, 18, 386–402. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X. Salicylic acid: Biosynthesis, perception, and contributions to plant immunity. Curr. Opin. Plant Biol. 2019, 50, 29–36. [Google Scholar] [CrossRef]
- Yalpani, N.; Enyedi, A.J.; Raskin, I. Ultraviolet light and ozone stimulate accumulation of salicylic acid, pathogenesis-related proteins and virus resistance in tobacco. Planta 1994, 193, 372–376. [Google Scholar] [CrossRef]
- Martínez, C.; Pons, E.; Prats, G.; León, J. Salicylic acid regulates flowering time and links defence responses and reproductive development. Plant J. 2003, 37, 209–217. [Google Scholar] [CrossRef]
- Pancheva, T.; Popova, L.; Uzunova, A. Effects of salicylic acid on growth and photosynthesis in barley plants. J. Plant Physiol. 1996, 149, 57–63. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Tari, I.; Páldi, E. Hydroponic treatment with salicylic acid decreases the effects of chilling injury in maize (Zea mays L.) plants. Planta 1999, 208, 175–180. [Google Scholar] [CrossRef]
- Mateo, A.; Mühlenbock, P.; Rustérucci, C.; Chang, C.C.-C.; Miszalski, Z.; Karpinska, B.; Parker, J.E.; Mullineaux, P.M.; Karpinski, S. LESION SIMULATING DISEASE 1 Is Required for Acclimation to Conditions That Promote Excess Excitation Energy. Plant Physiol. 2004, 136, 2818–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, K.; Hideg, É.; Szalai, G.; Kovács, L.; Janda, T. Salicylic acid may indirectly influence the photosynthetic electron transport. J. Plant Physiol. 2012, 169, 971–978. [Google Scholar] [CrossRef]
- Poór, P.; Tari, I. Regulation of stomatal movement and photosynthetic activity in guard cells of tomato abaxial epidermal peels by salicylic acid. Funct. Plant Biol. 2012, 39, 1028–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zu, X.; Wang, X.; Sun, A.; Zhang, J.; Wang, S.; Chen, Y. Comparative Proteomic Analysis of the Effects of Salicylic Acid and Abscisic Acid on Maize (Zea mays L.) Leaves. Plant Mol. Biol. Rep. 2012, 31, 507–516. [Google Scholar] [CrossRef]
- Csinos, A.S.; Pappu, H.R.; McPherson, R.M.; Stephenson, M.G. Management of Tomato spotted wilt virus in Flue-Cured Tobacco with Acibenzolar-S-Methyl and Imidacloprid. Plant Dis. 2001, 85, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Louws, F.J.; Wilson, M.; Campbell, H.L.; Cuppels, D.A.; Jones, J.B.; Shoemaker, P.B.; Sahin, F.; Miller, S.A. Field Control of Bacterial Spot and Bacterial Speck of Tomato Using a Plant Activator. Plant Dis. 2001, 85, 481–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, A.M.; Kousik, C.S.; Ritchie, D.F. Resistance to Bacterial Spot in Bell Pepper Induced by Acibenzolar-S-Methyl. Plant Dis. 2001, 85, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Gent, D.H.; Schwartz, H.F. Management of Xanthomonas Leaf Blight of Onion with a Plant Activator, Biological Control Agents, and Copper Bactericides. Plant Dis. 2005, 89, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Nair, C.B.; Anith, K.N.; Sreekumar, J. Mitigation of growth retardation effect of plant defense activator, acibenzolar-S-methyl, in amaranthus plants by plant growth-promoting rhizobacteria. World J. Microbiol. Biotechnol. 2006, 23, 1183–1187. [Google Scholar] [CrossRef]
- Heil, M.; Hilpert, A.; Kaiser, W.; Linsenmair, K.E. Reduced growth and seed set following chemical induction of pathogen defence: Does systemic acquired resistance (SAR) incur allocation costs? J. Ecol. 2000, 88, 645–654. [Google Scholar] [CrossRef]
- Babu, B.; Paret, M.L.; Martini, X.; Knox, G.W.; Riddle, B.; Ritchie, L.; Aldrich, J.; Kalischuk, M.; Da Silva, S. Impact of Foliar Application of Acibenzolar-S-Methyl on Rose Rosette Disease and Rose Plant Quality. Plant Dis. 2022, 106, 818–827. [Google Scholar] [CrossRef]
- Xue, L.J.; Guo, W.; Yuan, Y.; Anino, E.O.; Nyamdari, B.; Wilson, M.C.; Frost, C.J.; Chen, H.Y.; Babst, B.A.; Harding, S.A.; et al. Constitutively Elevated Salicylic Acid Levels Alter Photosynthesis and Oxidative State but Not Growth in Transgenic Populus. Plant Cell 2013, 25, 2714–2730. [Google Scholar] [CrossRef] [Green Version]
- Szalai, G.; Horgosi, S.; Soós, V.; Majláth, I.; Balázs, E.; Janda, T. Salicylic acid treatment of pea seeds induces its de novo synthesis. J. Plant Physiol. 2011, 168, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, F.; Faoro, F. Systemic Acquired Resistance (50 Years after Discovery): Moving from the Lab to the Field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Cole, D.L. The efficacy of acibenzolar-S-methyl, an inducer of systemic acquired resistance, against bacterial and fungal diseases of tobacco. Crop Prot. 1999, 18, 267–273. [Google Scholar] [CrossRef]
- Babu, R.M.; Sajeena, A.; Samundeeswari, A.V.; Sreedhar, A.; Vidhyasekeran, P.; Reddy, M.S. Induction of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice by treatment with acibenzolar-S-methyl. Ann. Appl. Biol. 2003, 143, 333–340. [Google Scholar] [CrossRef]
- Ishiga, T.; Iida, Y.; Sakata, N.; Ugajin, T.; Hirata, T.; Taniguchi, S.; Hayashi, K.; Ishiga, Y. Acibenzolar-S-methyl activates stomatal-based defense against Pseudomonas cannabina pv. alisalensis in cabbage. J. Gen. Plant Pathol. 2019, 86, 48–54. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef] [Green Version]
- Malerba, M.; Cerana, R. Recent Applications of Chitin- and Chitosan-Based Polymers in Plants. Polymers 2019, 11, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bona, G.S.; Vincenzi, S.; De Marchi, F.; Angelini, E.; Bertazzon, N. Chitosan induces delayed grapevine defense mechanisms and protects grapevine against Botrytis cinerea. J. Plant Dis. Prot. 2021, 128, 715–724. [Google Scholar] [CrossRef]
- Chakraborty, M.; Hasanuzzaman, M.; Rahman, M.; Khan, A.R.; Bhowmik, P.; Mahmud, N.U.; Tanveer, M.; Islam, T. Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer. Agriculture 2020, 10, 624. [Google Scholar] [CrossRef]
- Lobato, M.C.; Machinandiarena, M.F.; Tambascio, C.; Dosio, G.A.A.; Caldiz, D.O.; Daleo, G.R.; Andreu, A.B.; Olivieri, F.P. Effect of foliar applications of phosphite on post-harvest potato tubers. Eur. J. Plant Pathol. 2011, 130, 155–163. [Google Scholar] [CrossRef]
- Olivieri, F.P.; Feldman, M.L.; Machinandiarena, M.F.; Lobato, M.C.; Caldiz, D.O.; Daleo, G.R.; Andreu, A.B. Phosphite applications induce molecular modifications in potato tuber periderm and cortex that enhance resistance to pathogens. Crop Prot. 2012, 32, 1–6. [Google Scholar] [CrossRef]
- Eshraghi, L.; Anderson, J.; Aryamanesh, N.; McComb, J.A.; Shearer, B.; Hardy, G. Defence Signalling Pathways Involved in Plant Resistance and Phosphite-Mediated Control of Phytophthora cinnamomi. Plant Mol. Biol. Rep. 2013, 32, 342–356. [Google Scholar] [CrossRef]
- Gill, U.S.; Sun, L.; Rustgi, S.; Tang, Y.; Wettstein, D.; Mysore, K.S. Transcriptome-based analyses of phosphite-mediated suppression of rust pathogens Puccinia emaculata and Phakopsora pachyrhizi and functional characterization of selected fungal target genes. Plant J. 2018, 93, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Massoud, K.; Barchietto, T.; Le Rudulier, T.; Pallandre, L.; Didierlaurent, L.; Garmier, M.; Ambard-Bretteville, F.; Seng, J.-M.; Saindrenan, P. Dissecting Phosphite-Induced Priming in Arabidopsis Infected with Hyaloperonospora arabidopsidis. Plant Physiol. 2012, 159, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Thao, H.T.B.; Yamakawa, T. Phosphite (phosphorous acid): Fungicide, fertilizer or bio-stimulator? Soil Sci. Plant Nutr. 2009, 55, 228–234. [Google Scholar] [CrossRef]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.-M.; Pugin, A. Laminarin Elicits Defense Responses in Grapevine and Induces Protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant-Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- Renard-Merlier, D.; Randoux, B.; Nowak, E.; Farcy, F.; Durand, R.; Reignault, P. Iodus 40, salicylic acid, heptanoyl salicylic acid and trehalose exhibit different efficacies and defence targets during a wheat/powdery mildew interaction. Phytochemistry 2007, 68, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, A.; Trouvelot, S.; Kelloniemi, J.; Frettinger, P.; Wendehenne, D.; Daire, X.; Joubert, J.-M.; Ferrarini, A.; Delledonne, M.; Flors, V.; et al. The Sulfated Laminarin Triggers a Stress Transcriptome before Priming the SA- and ROS-Dependent Defenses during Grapevine’s Induced Resistance against Plasmopara viticola. PLoS ONE 2014, 9, e88145. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-R.; Lin, Y.-C.; Chuang, H.-W. Laminarin modulates the chloroplast antioxidant system to enhance abiotic stress tolerance partially through the regulation of the defensin-like gene expression. Plant Sci. 2016, 247, 83–92. [Google Scholar] [CrossRef]
- Cabrera, J.-C.; Boland, A.; Cambier, P.; Frettinger, P.; Van Cutsem, P. Chitosan oligosaccharides modulate the supramolecular conformation and the biological activity of oligogalacturonides in Arabidopsis. Glycobiology 2010, 20, 775–786. [Google Scholar] [CrossRef] [Green Version]
- Van Cutsem, P.; Messiaen, J. Biological effects of pectin fragments in plant cells. Acta Bot. Neerl. 1994, 43, 231–245. [Google Scholar] [CrossRef]
- Moreau, S.; Van Aubel, G.; Janky, R.; Van Cutsem, P. Chloroplast Electron Chain, ROS Production, and Redox Homeostasis Are Modulated by COS-OGA Elicitation in Tomato (Solanum lycopersicum) Leaves. Front. Plant Sci. 2020, 11, 597589. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Javaid, A.; Munir, R.; Khan, I.H.; Shoaib, A. Control of the chickpea blight, Ascochyta rabiei, with the weed plant, Withania somnifera. Egypt. J. Biol. Pest Control. 2020, 30, 114. [Google Scholar] [CrossRef]
- Javaid, A.; Khan, I.; Shoaib, A. Management of Charcoal Rot of Mungbean by Two Trichoderma Species and Dry Biomass of Coronopus didymus. Planta Daninha 2018, 36, e018182795. [Google Scholar] [CrossRef]
- Jabeen, N.; Javaid, A.; Shoaib, A.; Khan, I.H. Management of southern blight of bell pepper by soil amendment with dry biomass of Datura metel. J. Plant Pathol. 2021, 103, 901–913. [Google Scholar] [CrossRef]
- Sharf, W.; Javaid, A.; Shoaib, A.; Khan, I.H. Induction of resistance in chili against Sclerotium rolfsii by plant growth promoting rhizobacteria and Anagallis arvensis. Egypt. J. Biol. Pest Control. 2021, 31, 16. [Google Scholar] [CrossRef]
- Margaritopoulou, T.; Toufexi, E.; Kizis, D.; Balayiannis, G.; Anagnostopoulos, C.; Theocharis, A.; Rempelos, L.; Troyanos, Y.; Leifert, C.; Markellou, E. Reynoutria sachalinensis extract elicits SA-dependent defense responses in courgette genotypes against powdery mildew caused by Podosphaera xanthii. Sci. Rep. 2020, 10, 3354. [Google Scholar] [CrossRef]
- Teysssier, L.; Colussi, J.; Delemasure, S.; Chluba, J.; Wendehenne, D.; LaMotte, O.; Connat, J.-L. Inflammatory Effects of the Plant Protection Product Stifenia (FEN560) on Vertebrates. Front. Public Health 2017, 5, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, R.M.D.M.; Rotolo, C.; Gerin, D.; Abate, D.; Pollastro, S.; Faretra, F. Global transcriptome analysis and differentially expressed genes in grapevine after application of the yeast-derived defense inducer cerevisane. Pest Manag. Sci. 2019, 75, 2020–2033. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Dubey, N.; Shukla, A.K. Use of some essential oils as post-harvest botanical fungicides in the management of grey mould of grapes caused by Botrytis cinerea. World J. Microbiol. Biotechnol. 2007, 24, 39–46. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential Oils: Sources of Antimicrobials and Food Preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viacava, G.E.; Goyeneche, R.; Goñi, M.G.; Roura, S.I.; Agüero, M.V. Natural elicitors as preharvest treatments to improve postharvest quality of Butterhead lettuce. Sci. Hortic. 2018, 228, 145–152. [Google Scholar] [CrossRef]
- Verdi, C.M.; Machado, V.S.; Machado, A.K.; Klein, B.; Bonez, P.C.; de Andrade, E.N.C.; Rossi, G.; Campos, M.M.; Wagner, R.; Sagrillo, M.R.; et al. Phytochemical characterization, genotoxicity, cytotoxicity, and antimicrobial activity of Gautheria procumbens essential oil. Nat. Prod. Res. 2020, 36, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Vergnes, S.; Ladouce, N.; Fournier, S.; Ferhout, H.; Attia, F.; Dumas, B. Foliar treatments with Gaultheria procumbens essential oil induce defense responses and resistance against a fungal pathogen in Arabidopsis. Front. Plant Sci. 2014, 5, 477. [Google Scholar] [CrossRef]
- Jamiołkowska, A. Natural Compounds as Elicitors of Plant Resistance against Diseases and New Biocontrol Strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced Systemic Resistance and Promotion of Plant Growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, P. Biological Management of Diseases of Crops; Springer: Dordrecht, The Netherlands, 2013; 673p. [Google Scholar]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. Effects of Bacillus subtilis on some physiological and biochemical parameters of Triticum aestivum L. (wheat) under salinity. Plant Physiol. Biochem. 2017, 121, 80–88. [Google Scholar] [CrossRef]
- Giraud, R. Bacillus: Une Bactérie Auxilliaire Pour la Santé du Gazon; Clinique du Gazon: Saint-Verand, France, 2019. [Google Scholar]
- Stewart, A.; Hill, R. Applications of Trichoderma in Plant Growth Promotion. In Biotechnology and Biology of Trichoderma; Elsevier: Amsterdam, The Netherlands, 2014; pp. 415–428. [Google Scholar] [CrossRef]
- Mastouri, F.; Björkman, T.; Harman, G.E. Trichoderma harzianum Enhances Antioxidant Defense of Tomato Seedlings and Resistance to Water Deficit. Mol. Plant-Microbe Interact. 2012, 25, 1264–1271. [Google Scholar] [CrossRef] [Green Version]
- Morán-Diez, M.E.; Martínez de Alba, Á.E.; Rubio, M.B.; Hermosa, R.; Monte, E. Trichoderma and the Plant Heritable Priming Responses. J. Fungi 2021, 7, 318. [Google Scholar] [CrossRef]
- Karuppiah, V.; Sun, J.; Li, T.; Vallikkannu, M.; Chen, J. Co-cultivation of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 Causes Differential Gene Expression and Improvement in the Wheat Growth and Biocontrol Activity. Front Microbiol. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Coutand, C. The Effect of Mechanical Stress on Plant Susceptibility to Pests: A Mini Opinion Review. Plants 2020, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- L’Haridon, F.; Besson-Bard, A.; Binda, M.; Serrano, M.; Mansour, E.A.; Balet, F.; Schoonbeek, H.-J.; Hess, S.; Mir, R.; León, J.; et al. A Permeable Cuticle Is Associated with the Release of Reactive Oxygen Species and Induction of Innate Immunity. PLoS Pathog. 2011, 7, e1002148. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Cardenas, M.; Ryan, C.A. Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6553–6557. [Google Scholar] [CrossRef] [Green Version]
- Mauch, F.; Kmecl, A.; Schaffrath, U.; Volrath, S.; Gorlach, J.; Ward, E.; Ryals, J.; Dudler, R. Mechanosensitive Expression of a Lipoxygenase Gene in Wheat. Plant Physiol. 1997, 114, 1561–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tretner, C.; Huth, U.; Hause, B. Mechanostimulation of Medicago truncatula leads to enhanced levels of jasmonic acid. J. Exp. Bot. 2008, 59, 2847–2856. [Google Scholar] [CrossRef]
- Mitchell, C.A.; Myers, P.N. Mechanical Stress Regulation of Plant Growth and Development. In Horticultural Reviews; Wiley: Hoboken, NJ, USA, 2010; Volume 17, pp. 1–42. [Google Scholar] [CrossRef]
- Turgeon, R.; Webb, J.A. Growth Inhibition by Mechanical Stress. Science 1971, 174, 961–962. [Google Scholar] [CrossRef] [Green Version]
- Börnke, F.; Rocksch, T. Thigmomorphogenesis—Control of plant growth by mechanical stimulation. Sci. Hortic. 2018, 234, 344–353. [Google Scholar] [CrossRef]
- Huché-Thélier, L.; Crespel, L.; Le Gourrierec, J.; Morel, P.; Sakr, S.; Leduc, N. Light signaling and plant responses to blue and UV radiations—Perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Ballaré, C.L. Light Regulation of Plant Defense. Annu. Rev. Plant Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef]
- Urban, L.; Sari, D.C.; Orsal, B.; Lopes, M.; Miranda, R.; Aarrouf, J. UV-C light and pulsed light as alternatives to chemical and biological elicitors for stimulating plant natural defenses against fungal diseases. Sci. Hortic. 2018, 235, 452–459. [Google Scholar] [CrossRef]
- Lopes, M.M.; Silva, E.O.; Canuto, K.M.; Silva, L.M.; Gallão, M.I.; Urban, L.; Ayala-Zavala, J.F.; Miranda, M.R.A. Low fluence pulsed light enhanced phytochemical content and antioxidant potential of ‘Tommy Atkins’ mango peel and pulp. Innov. Food Sci. Emerg. Technol. 2016, 33, 216–224. [Google Scholar] [CrossRef]
- Filho, F.O.; Silva, E.D.O.; Lopes, M.M.D.A.; Ribeiro, P.R.V.; Oster, A.H.; Guedes, J.A.C.; Zampieri, D.D.S.; Bordallo, P.D.N.; Zocolo, G.J. Effect of pulsed light on postharvest disease control-related metabolomic variation in melon (Cucumis melo) artificially inoculated with Fusarium pallidoroseum. PLoS ONE 2020, 15, e0220097. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Yang, H.-Q. CRYPTOCHROME 1 Is Implicated in Promoting R Protein-Mediated Plant Resistance to Pseudomonas syringae in Arabidopsis. Mol. Plant 2010, 3, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Jeong, R.-D.; Chandra-Shekara, A.C.; Barman, S.R.; Navarre, D.; Klessig, D.F.; Kachroo, A.; Kachroo, P. Cryptochrome 2 and phototropin 2 regulate resistance protein-mediated viral defense by negatively regulating an E3 ubiquitin ligase. Proc. Natl. Acad. Sci. USA 2010, 107, 13538–13543. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Zhang, M.; Lu, C.; Hettenhausen, C.; Tan, Q.; Cao, G.; Zhu, X.; Wu, G.; Wu, J. Ultraviolet-B enhances the resistance of multiple plant species to lepidopteran insect herbivory through the jasmonic acid pathway. Sci. Rep. 2018, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Bravo, R.; Cheng, G.; Kim, H.K.; Grosser, K.; Van Dam, N.M.; Leiss, K.A.; Klinkhamer, P.G.L. Ultraviolet radiation exposure time and intensity modulate tomato resistance to herbivory through activation of jasmonic acid signaling. J. Exp. Bot. 2018, 70, 315–327. [Google Scholar] [CrossRef]
- Demkura, P.V.; Ballaré, C.L. UVR8 Mediates UV-B-Induced Arabidopsis Defense Responses against Botrytis cinerea by Controlling Sinapate Accumulation. Mol. Plant 2012, 5, 642–652. [Google Scholar] [CrossRef]
- Teramura, A.H.; Sullivan, J. Effects of UV-B radiation on photosynthesis and growth of terrestrial plants. Photosynth. Res. 1994, 39, 463–473. [Google Scholar] [CrossRef]
- Liu, J.; Stevens, C.; Khan, V.A.; Lu, J.Y.; Wilson, C.L.; Adeyeye, O.; Kabwe, M.K.; Pusey, P.L.; Chalutz, E.; Sultana, T.; et al. Application of Ultraviolet-C Light on Storage Rots and Ripening of Tomatoes. J. Food Prot. 1993, 56, 868–873. [Google Scholar] [CrossRef]
- Charles, M.T.; Goulet, A.; Arul, J. Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit. Postharvest Biol. Technol. 2008, 47, 41–53. [Google Scholar] [CrossRef]
- Charles, M.T.; Tano, K.; Asselin, A.; Arul, J. Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit. V. Constitutive defence enzymes and inducible pathogenesis-related proteins. Postharvest Biol. Technol. 2009, 51, 414–424. [Google Scholar] [CrossRef]
- Mercier, J.; Arul, J.; Julien, C. Effect of UV-C on Phytoalexin Accumulation and Resistance to Botrytis cinerea in Stored Carrots. J. Phytopathol. 1993, 139, 17–25. [Google Scholar] [CrossRef]
- Mercier, J.; Baka, M.; Reddy, B.; Corcuff, R.; Arul, J. Shortwave Ultraviolet Irradiation for Control of Decay Caused by Botrytis cinerea in Bell Pepper: Induced Resistance and Germicidal Effects. J. Am. Soc. Hortic. Sci. 2001, 126, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Urban, L.; Charles, F.; de Miranda, M.R.A.; Aarrouf, J. Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol. Biochem. 2016, 105, 1–11. [Google Scholar] [CrossRef]
- Aarrouf, J.; Urban, L. Flashes of UV-C light: An innovative method for stimulating plant defences. PLoS ONE 2020, 15, e0235918. [Google Scholar] [CrossRef]
- Aarrouf, J.; Goncalves, H.; Ledermann, L.; Matton, Y.; Lizzi, Y.; Junglee, S.; Suchail, S.; Urban, L. Simulation des défenses du fraisier par les flashs d’UV-C. Phytoma 2020, 732, 48–51. [Google Scholar]
- Ledermann, L.; Daouda, S.; Gouttesoulard, C.; Aarrouf, J.; Urban, L. Flashes of UV-C Light Stimulate Defenses of Vitis vinifera L. ‘Chardonnay’ Against Erysiphe necator in Greenhouse and Vineyard Conditions. Plant Dis. 2021, 105, 2106–2113. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Dufour, M.C.; Lambert, C.; Bouscaut, J.; Mérillon, J.M.; Corio-Costet, M.F. Benzothiadiazole-primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2012, 62, 370–382. [Google Scholar] [CrossRef]
- Banani, H.; Roatti, B.; Ezzahi, B.; Giovannini, O.; Gessler, G.; Pertot, I.; Perazzolli, M. Characterization of resistance mechanisms activated by Trichoderma harzianum T39 and benzothiadiazole to downy mildew in different grapevine cultivars. Plant Pathol. 2013, 63, 334–343. [Google Scholar] [CrossRef]
- Bellée, A.; Cluzet, S.; Dufour, M.-C.; Mérillon, J.-M.; Corio-Costet, M.-F. Comparison of the Impact of Two Molecules on Plant Defense and on Efficacy against Botrytis cinerea in the Vineyard: A Plant Defense Inducer (Benzothiadiazole) and a Fungicide (Pyrimethanil). J. Agric. Food Chem. 2018, 66, 3338–3350. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, A.; Figueiredo, J.; Silva, M.S.; Figueiredo, A. Linking Jasmonic Acid to Grapevine Resistance against the Biotrophic Oomycete Plasmopara viticola. Front. Plant Sci. 2016, 7, 565. [Google Scholar] [CrossRef] [Green Version]
- Belhadj, A.; Telef, N.; Cluzet, S.; Bouscaut, J.; Corio-Costet, M.-F.; Mérillon, J.-M. Ethephon Elicits Protection against Erysiphe necator in Grapevine. J. Agric. Food Chem. 2008, 56, 5781–5787. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Park, J.-W.; Lee, S.-W.; Balaraju, K. Disease suppression and growth promotion in cucumbers induced by integrating PGPR agent Bacillus subtilis strain B4 and chemical elicitor ASM. Crop. Prot. 2013, 54, 199–205. [Google Scholar] [CrossRef]
- El-Rahman, S.S.A.; Mohamed, H.I. Application of benzothiadiazole and Trichoderma harzianum to control faba bean chocolate spot disease and their effect on some physiological and biochemical traits. Acta Physiol. Plant. 2013, 36, 343–354. [Google Scholar] [CrossRef]
- De Jong, H.; Reglinski, T.; Elmer, P.A.G.; Wurms, K.; Vanneste, J.L.; Guo, L.F.; Alavi, M. Integrated Use of Aureobasidium pullulans Strain CG163 and Acibenzolar-S-Methyl for Management of Bacterial Canker in Kiwifruit. Plants 2019, 8, 287. [Google Scholar] [CrossRef] [Green Version]
- Reuveni, M.; Sheglov, D.; Cohen, Y. Control of Moldy-Core Decay in Apple Fruits by β-Aminobutyric Acids and Potassium Phosphites. Plant Dis. 2003, 87, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R.; Havis, N.D.; Sablou, C.; Walsh, D.J. Possible trade-off associated with the use of a combination of resistance elicitors. Physiol. Mol. Plant Pathol. 2011, 75, 188–192. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Kumar, M.; Sahu, P.K.; Singh, H.V.; Kumar, S.; Roy, M.; Imran, M.; Rai, J.P.; et al. Trichoderma harzianum- and Methyl Jasmonate-Induced Resistance to Bipolaris sorokiniana through Enhanced Phenylpropanoid Activities in Bread Wheat (Triticum aestivum L.). Front. Microbiol. 2019, 10, 1697. [Google Scholar] [CrossRef] [Green Version]
- Zehra, A.; Meena, M.; Dubey, M.K.; Aamir, M.; Upadhyay, R.S. Synergistic effects of plant defense elicitors and Trichoderma harzianum on enhanced induction of antioxidant defense system in tomato against Fusarium wilt disease. Bot. Stud. 2017, 58, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langa-Lomba, N.; Buzón-Durán, L.; Martín-Ramos, P.; Casanova-Gascón, J.; Martín-Gil, J.; Sánchez-Hernández, E.; González-García, V. Assessment of Conjugate Complexes of Chitosan and Urtica dioica or Equisetum arvense Extracts for the Control of Grapevine Trunk Pathogens. Agronomy 2021, 11, 976. [Google Scholar] [CrossRef]
- Gómez-Merino, F.C.; Trejo-Téllez, L.I. Biostimulant activity of phosphite in horticulture. Sci. Hortic. 2015, 196, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Nojosa, G.B.A.; Resende, M.L.V.; Barguil, B.M.; Moraes, S.A.G.; Vilas Boas, C.H. Effect of resistance inducers on coffee against Phoma leaf spot. Summa Phytopathol. 2009, 35, 60–62. [Google Scholar] [CrossRef] [Green Version]
- Ogoshi, C.; de Abreu, M.S.; da Silva, B.M.; Neto, H.S.; Junior, P.M.R.; de Resende, M.L.V. Potassium phosphite: A promising product in the management of diseases caused by Colletotrichum gloeosporioides in coffee plants. Biosci. J. 2013, 29, 1558–1565. [Google Scholar]
- Monteiro, A.C.A.; de Resende, M.L.V.; Valente, T.C.T.; Junior, P.M.R.; Pereira, V.F.; da Costa, J.R.; da Silva, J.A.G. Manganese Phosphite in Coffee Defence against Hemileia vastatrix, the Coffee Rust Fungus: Biochemical and Molecular Analyses. J. Phytopathol. 2016, 164, 1043–1053. [Google Scholar] [CrossRef]
- Lobato, M.C.; Olivieri, F.P.; Altamiranda, E.A.G.; Wolski, E.A.; Daleo, G.R.; Caldiz, D.O.; Andreu, A.B. Phosphite compounds reduce disease severity in potato seed tubers and foliage. Eur. J. Plant Pathol. 2008, 122, 349–358. [Google Scholar] [CrossRef]
- Achary, V.M.M.; Ram, B.; Manna, M.; Datta, D.; Bhatt, A.; Reddy, M.K.; Agrawal, P.K. Phosphite: A novel P fertilizer for weed management and pathogen control. Plant Biotechnol. J. 2017, 15, 1493–1508. [Google Scholar] [CrossRef] [Green Version]
- Diánez, F.; Santos, M.; Parra, C.; Navarro, M.; Blanco, R.; Gea, F. Screening of antifungal activity of 12 essential oils against eight pathogenic fungi of vegetables and mushroom. Lett. Appl. Microbiol. 2018, 67, 400–410. [Google Scholar] [CrossRef]
- Trebbi, G.; Negri, L.; Bosi, S.; Dinelli, G.; Cozzo, R.; Marotti, I. Evaluation of Equisetum arvense (Horsetail Macerate) as a Copper Substitute for Pathogen Management in Field-Grown Organic Tomato and Durum Wheat Cultivations. Agriculture 2020, 11, 5. [Google Scholar] [CrossRef]
- Misato, T.; Homma, Y.; Kō, K. The development of a natural fungicide, soybean lecithin. Eur. J. Plant Pathol. 1977, 83, 395–402. [Google Scholar] [CrossRef]
- Homma, Y.; Takahashi, H.; Arimoto, Y. Studies on the Mode of Action of Soybean Lecithin. Part 3. Effects on the Infection Process of Rice Blast Fungus, Pyricularia oryzae. Jpn. J. Phytopathol. 1992, 58, 514–521. [Google Scholar] [CrossRef]
- Kang, B.R.; Park, J.S.; Jung, W.-J. Antiviral activity by lecithin-induced fengycin lipopeptides as a potent key substrate against Cucumber mosaic virus. Microb. Pathog. 2021, 155, 104910. [Google Scholar] [CrossRef] [PubMed]
- Salgado, M.; Rodríguez-Rojo, S.; Alves-Santos, F.M.; Cocero, M.J. Encapsulation of resveratrol on lecithin and β-glucans to enhance its action against Botrytis cinerea. J. Food Eng. 2015, 165, 13–21. [Google Scholar] [CrossRef]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro-and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Mahendran, R.; Ramanan, K.R.; Barba, F.J.; Lorenzo, J.M.; López-Fernández, O.; Munekata, P.E.S.; Roohinejad, S.; Sant’Ana, A.S.; Tiwari, B.K. Recent advances in the application of pulsed light processing for improving food safety and increasing shelf life. Trends Food Sci. Technol. 2019, 88, 67–79. [Google Scholar] [CrossRef]
- Bintsis, T.; Litopoulou-Tzanetaki, E.; Robinson, R.K. Existing and potential applications of ultraviolet light in the food industry—A critical review. J. Sci. Food Agric. 2000, 80, 637–645. [Google Scholar] [CrossRef]
- Jay, J.M.; Loessner, M.A.J.; Golden, D.A. Modern Food Microbiology; Springer Science and Business Media: New York, NY, USA, 2008. [Google Scholar]
- Wuytack, E.Y.; Phuong, L.D.T.; Aertsen, A.; Reyns, K.M.F.; Marquenie, D.; De Ketelaere, B.; Masschalck, B.; Van Opstal, I.; Diels, A.M.J.; Michiels, C.W. Comparison of Sublethal Injury Induced in Salmonella enterica Serovar Typhimurium by Heat and by Different Nonthermal Treatments. J. Food Prot. 2003, 66, 31–37. [Google Scholar] [CrossRef]
- Santos, A.L.; Oliveira, V.; Baptista, I.; Henriques, I.; Gomes, N.C.M.; Almeida, A.; Correia, A.; Cunha, Â. Wavelength dependence of biological damage induced by UV radiation on bacteria. Arch. Microbiol. 2012, 195, 63–74. [Google Scholar] [CrossRef]
- Vanhaelewyn, L.; Van Der Straeten, D.; De Coninck, B.; Vandenbussche, F. Ultraviolet Radiation From a Plant Perspective: The Plant-Microorganism Context. Front. Plant Sci. 2020, 11, 597642. [Google Scholar] [CrossRef]
- Patel, J.S.; Radetsky, L.C.; Nagare, R.; Rea, M.S. Nighttime Application of UV-C to Control Cucumber Powdery Mildew. Plant Health Prog. 2020, 21, 40–46. [Google Scholar] [CrossRef]
- Onofre, R.B.; Gadoury, D.M.; Stensvand, A.; Bierman, A.; Rea, M.; Peres, N.A. Use of Ultraviolet Light to Suppress Powdery Mildew in Strawberry Fruit Production Fields. Plant Dis. 2021, 105, 2402–2409. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, P.; Jha, G. Alterations in plant sugar metabolism: Signatory of pathogen attack. Planta 2018, 249, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Calonnec, A.; Jolivet, J.; Vivin, P.; Schnee, S. Pathogenicity Traits Correlate with the Susceptible Vitis vinifera Leaf Physiology Transition in the Biotroph Fungus Erysiphe necator: An Adaptation to Plant Ontogenic Resistance. Front. Plant Sci. 2018, 9, 1808. [Google Scholar] [CrossRef]
- Lecompte, F.; Nicot, P.C.; Ripoll, J.; Abro, M.A.; Raimbault, A.K.; Lopez-Lauri, F.; Bertin, N. Reduced susceptibility of tomato stem to the necrotrophic fungus Botrytis cinerea is associated with a specific adjustment of fructose content in the host sugar pool. Ann. Bot. 2017, 119, 931–943. [Google Scholar] [CrossRef] [Green Version]
- Herms, D.A.; Mattson, W.J. The Dilemma of Plants: To Grow or Defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced Resistance for Plant Disease Control: Maximizing the Efficacy of Resistance Elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R.; Ratsep, J.; Havis, N. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef]
- Van Hulten, M.; Pelser, M.; van Loon, L.C.; Pieterse, C.M.J.; Ton, J. Costs and benefits of priming for defense in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 5602–5607. [Google Scholar] [CrossRef] [Green Version]
- Poiroux-Gonord, F.; Santini, J.; Fanciullino, A.-L.; Lopez-Lauri, F.; Giannettini, J.; Sallanon, H.; Berti, L.; Urban, L. Metabolism in orange fruits is driven by photooxidative stress in the leaves. Physiol. Plant. 2013, 149, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Sanz, A.; Brenner, M.L.; Smith, A. Sucrose Synthase, Starch Accumulation, and Tomato Fruit Sink Strength. Plant Physiol. 1993, 101, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Aslani, L.; Gholami, M.; Mobli, M.; Sabzalian, M.R. The influence of altered sink-source balance on the plant growth and yield of greenhouse tomato. Physiol. Mol. Biol. Plants 2020, 26, 2109–2123. [Google Scholar] [CrossRef] [PubMed]
- Pawar, R.; Rana, V.S. Manipulation of Source-Sink Relationship in Pertinence to Better Fruit Quality and Yield in Fruit Crops: A Review. Agric. Rev. 2019, 40, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Lasa, B.; Menendez, S.; Sagastizabal, K.; Cervantes, M.E.C.; Irigoyen, I.; Muro, J.; Aparicio-Tejo, P.M.; Ariz, I. Foliar application of urea to “Sauvignon Blanc” and “Merlot” vines: Doses and time of application. Plant Growth Regul. 2012, 67, 73–81. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Gonzalo-Diago, A.; Moreno-Simunovic, Y.; Martínez-Gil, A.M. Effect of different foliar nitrogen applications on the must amino acids and glutathione composition in Cabernet Sauvignon vineyard. LWT Food Sci. Technol. 2017, 75, 147–154. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Romanazzi, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. A review of the use of biostimulants in the vineyard for improved grape and wine quality: Effects on prevention of grapevine diseases. J. Sci. Food Agric. 2018, 99, 1001–1009. [Google Scholar] [CrossRef]
- Massi, F.; Torriani, S.F.F.; Borghi, L.; Toffolatti, S.L. Fungicide Resistance Evolution and Detection in Plant Pathogens: Plasmopara viticola as a Case Study. Microorganisms 2021, 9, 119. [Google Scholar] [CrossRef]
Figure 1.
Biological and agronomic effects of UV-C light.

Figure 2.
Combining fungicides, elicitors/PRIs and biostimulants to increase disease control. An acceptable level of disease development is achieved when plants synthetize enough molecules and structures of defense relative to pathogen/disease pressure. The production of molecules and structures depends on both the level of defense stimulation and the level of resources, whereas the level of disease pressure can be controlled by crop management, specific fungicides and elicitors endowed with fungicide effects. A feedback effect of the level of disease pressure on fungicide efficacy has been added to this figure [195].
Figure 2.
Combining fungicides, elicitors/PRIs and biostimulants to increase disease control. An acceptable level of disease development is achieved when plants synthetize enough molecules and structures of defense relative to pathogen/disease pressure. The production of molecules and structures depends on both the level of defense stimulation and the level of resources, whereas the level of disease pressure can be controlled by crop management, specific fungicides and elicitors endowed with fungicide effects. A feedback effect of the level of disease pressure on fungicide efficacy has been added to this figure [195].


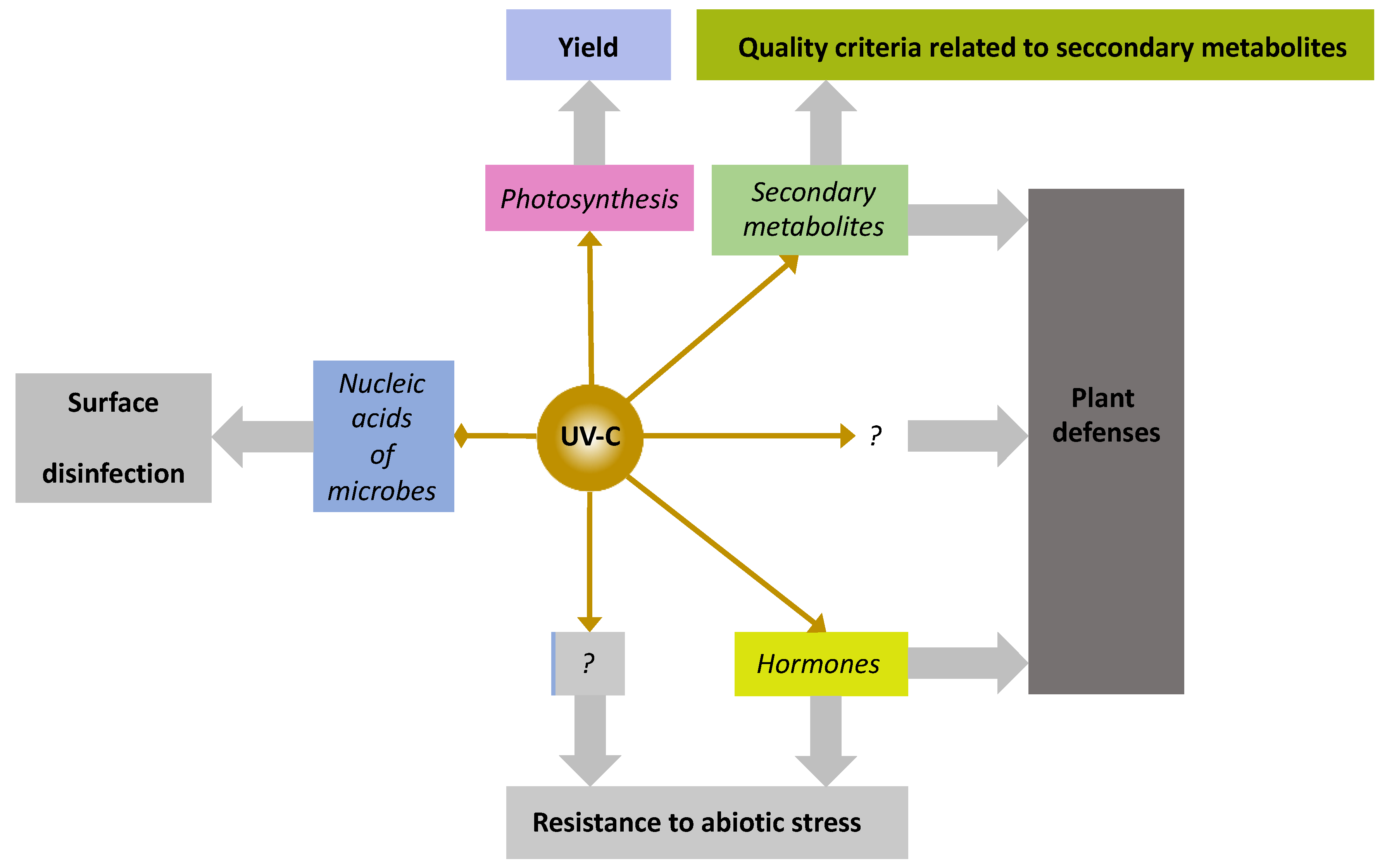{kind=link}
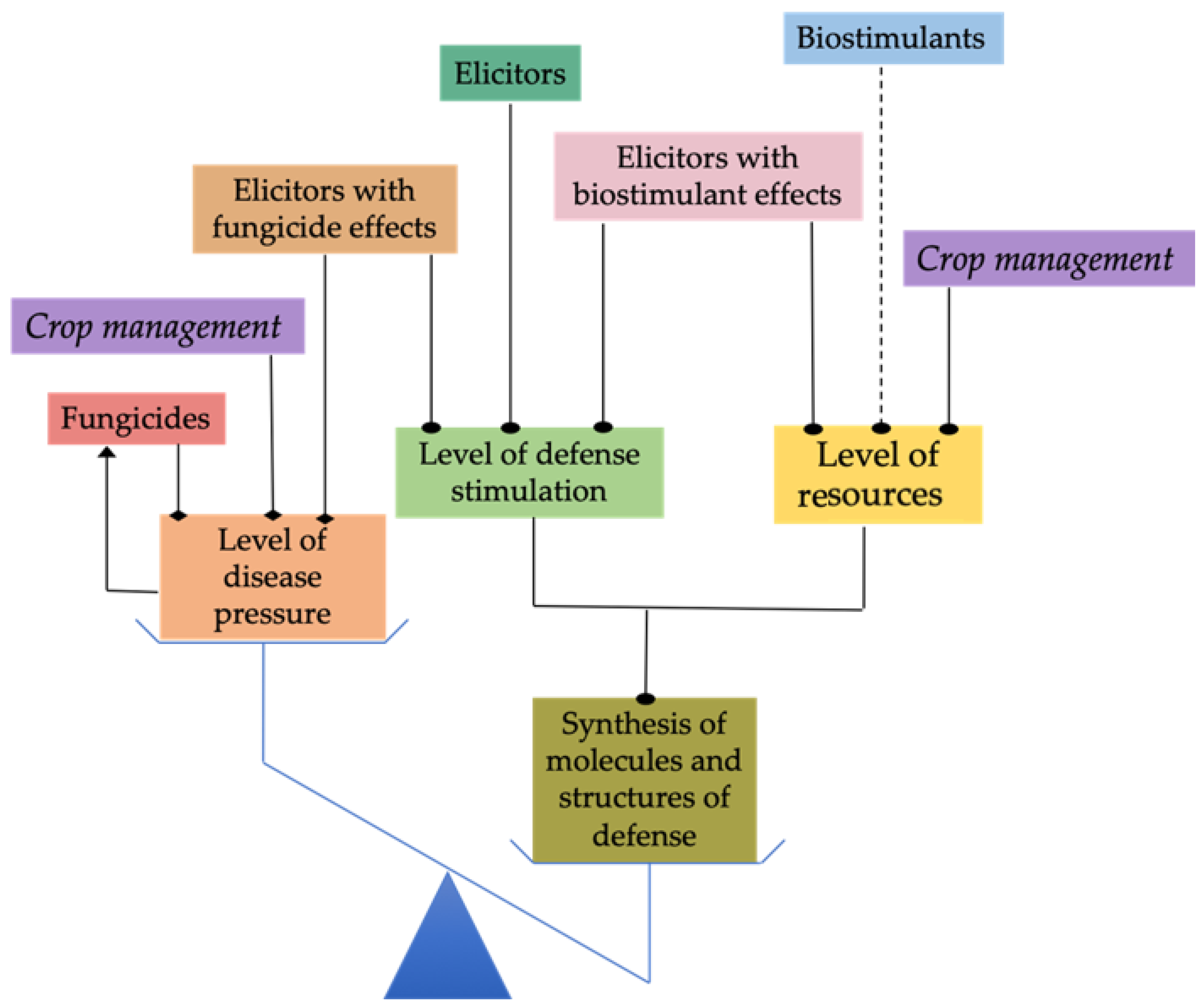{kind=link}
Table 1.
Some crops for which SA treatments have proved beneficial for stress tolerance. In the trial of [17], SA was supplied to seeds (seed priming).
Table 1.
Some crops for which SA treatments have proved beneficial for stress tolerance. In the trial of [17], SA was supplied to seeds (seed priming).
| Crop | Stress | Reference |
|---|---|---|
| Barley | High cadmium | [25] |
| Common bean | High cadmium | [26] |
| Cucumber | Cold | [24] |
| Grapevine | High salinity | [27] |
| Maize | Drought | [28] |
| Maize | High cadmium | [17] |
| Maize | High salinity | [29] |
| Pepper | High salinity | [30] |
| Strawberry | Cold | [31] |
| Wheat | Drought | [32] |
| Wheat | High temperature | [33] |
Table 2.
Some PRIs/elicitors used in cropping conditions. See text for details and references. The effect on growth was assessed considering the doses that are effective for stimulating plant resistance.
Table 2.
Some PRIs/elicitors used in cropping conditions. See text for details and references. The effect on growth was assessed considering the doses that are effective for stimulating plant resistance.
| Name | Category | SA Pathway | JA/ET Pathway | Fungicide Effect | Effect on Growth |
|---|---|---|---|---|---|
| ASM | Chemical elicitor | yes | yes | not documented | generally negative |
| Cerevisane | Chemical elicitor of natural origin | yes | yes | not documented | not documented |
| Chitin and chitosan | Chemical elicitor of natural origin | yes | yes | yes | positive |
| COS-OGA | Chemical elicitor of natural origin | yes | no | not documented | no negative effects reported |
| Essential oils | Chemical elicitor of natural origin | probable | not documented | some EOs are effective | not documented |
| Flashes of UV-C light | Physical elicitor | yes | (yes) | yes | no negative effect in the absence of stress |
| Laminarin/ PS3 | Chemical elicitor of natural origin | after inoculation | yes | not documented | positive in conditions of stress |
| Mechanical stress | Physical elicitor | unknown | yes | no | potentially negative |
| Phosphites | Chemical elicitor | indirectly | yes | yes | potentially negative |
| Trichoderma spp. | Biological elicitor | yes | yes | yes | positive, plays a role in the defense/growth balance |
| UV-B light | Physical elicitor | not documented | yes | yes | negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Urban, L.; Lauri, F.; Ben Hdech, D.; Aarrouf, J. Prospects for Increasing the Efficacy of Plant Resistance Inducers Stimulating Salicylic Acid. Agronomy 2022, 12, 3151. https://doi.org/10.3390/agronomy12123151
AMA Style
Urban L, Lauri F, Ben Hdech D, Aarrouf J. Prospects for Increasing the Efficacy of Plant Resistance Inducers Stimulating Salicylic Acid. Agronomy. 2022; 12(12):3151. https://doi.org/10.3390/agronomy12123151
Chicago/Turabian StyleUrban, Laurent, Félicie Lauri, Douae Ben Hdech, and Jawad Aarrouf. 2022. "Prospects for Increasing the Efficacy of Plant Resistance Inducers Stimulating Salicylic Acid" Agronomy 12, no. 12: 3151. https://doi.org/10.3390/agronomy12123151
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.