
Reaction of Oat Genotypes to Fusarium equiseti (Corda) Sacc. Infection and Mycotoxin Concentrations in Grain

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experiment
2.1.1. Infectious Material
2.1.2. Panicle Inoculation
2.1.3. Fungus Re-Isolation
2.2. Chemical Analyses
2.3. Statistical Analysis
2.4. Weather Conditions
3. Results
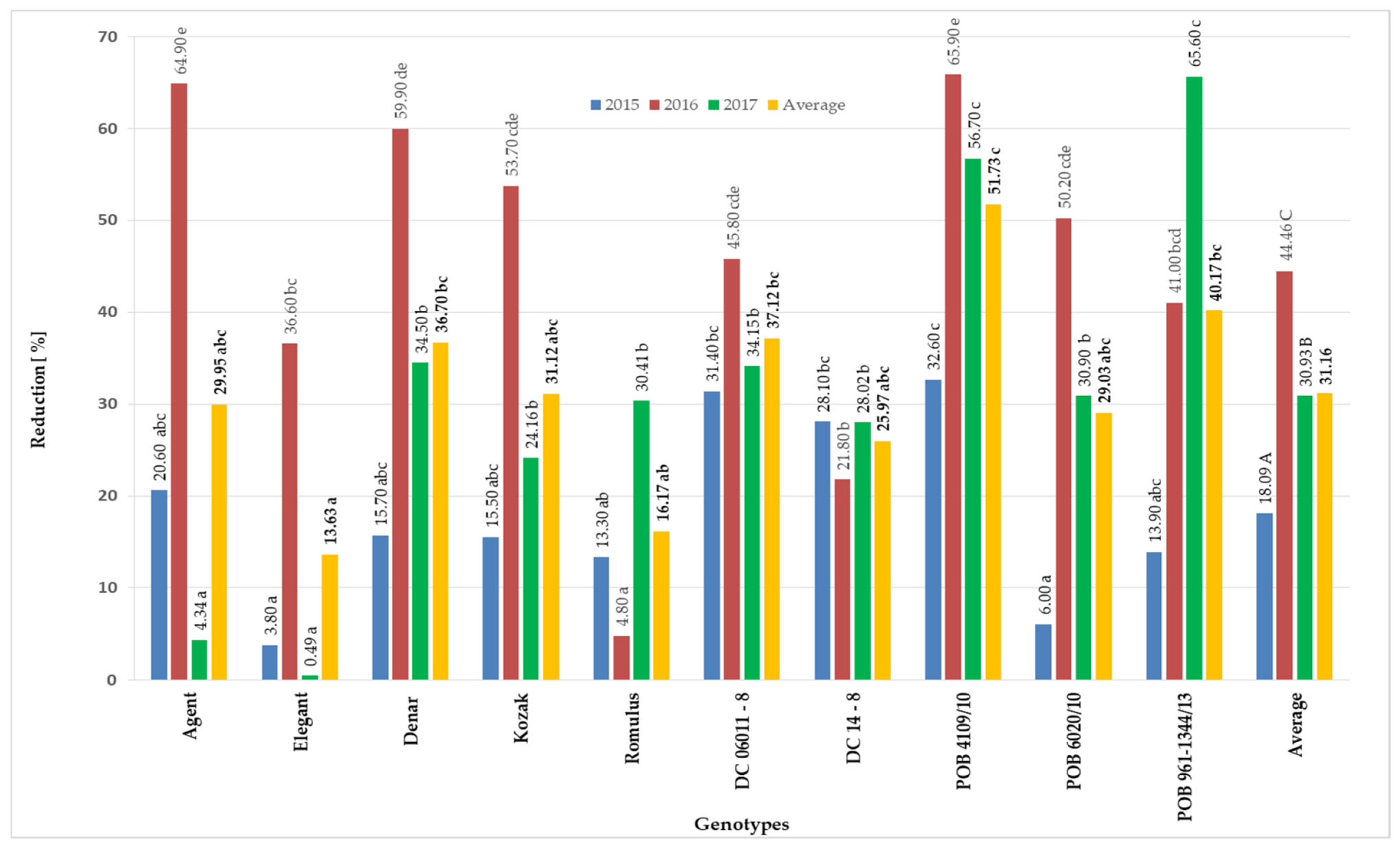
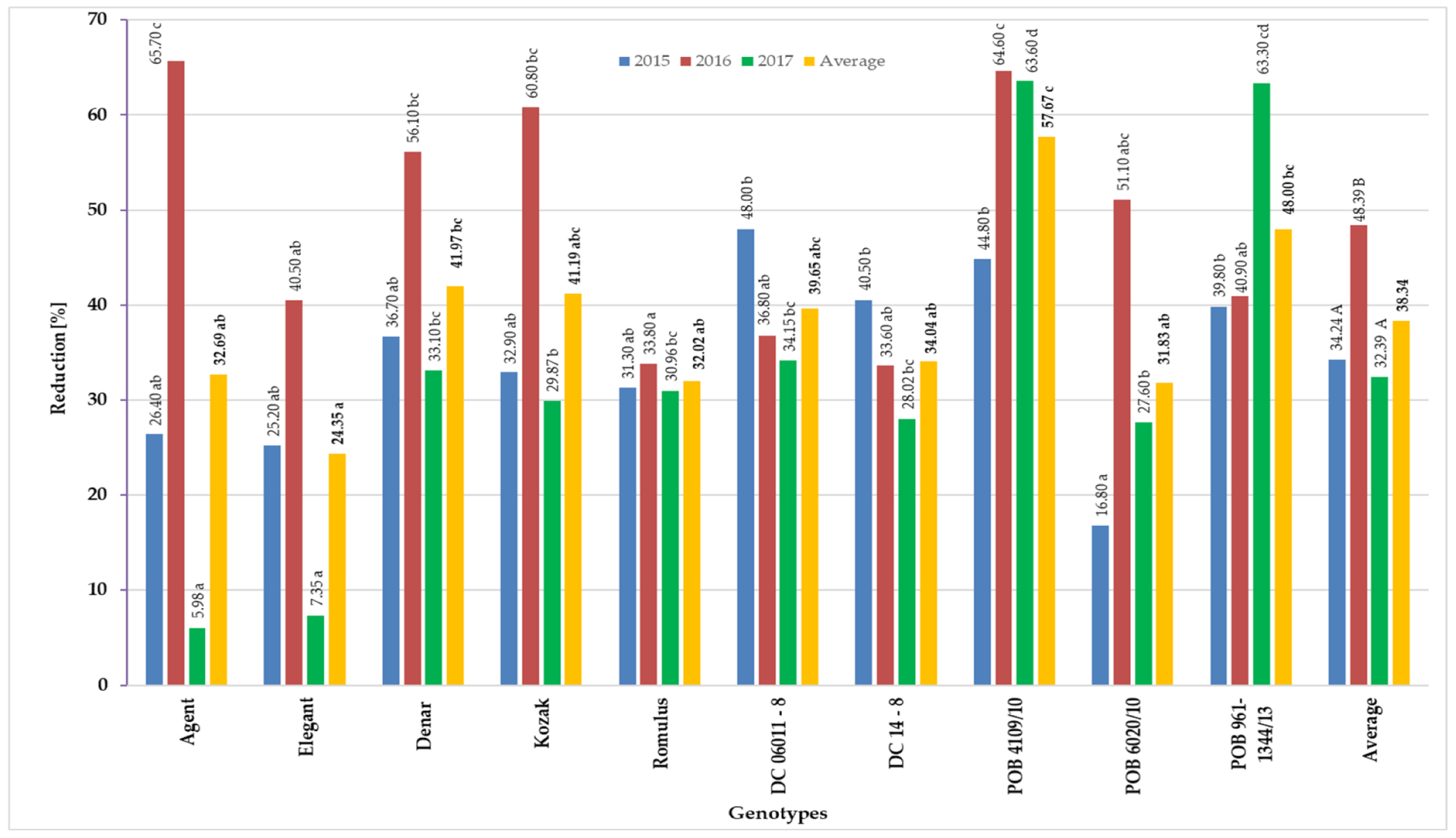
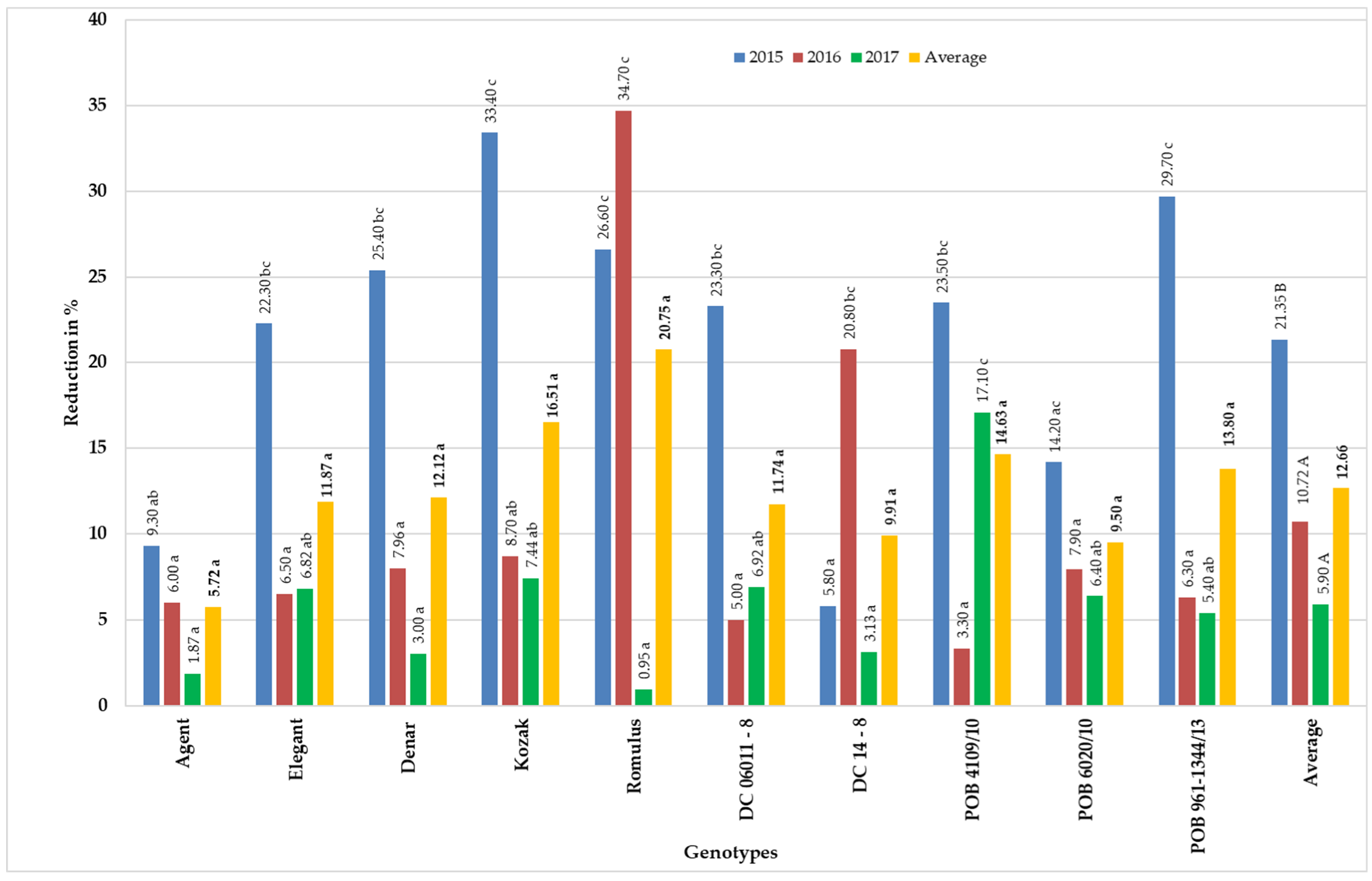
3.1. Mycotoxin Contents in Grain Obtained from Panicles Inoculated with F. equiseti at the Flowering Stage
3.2. Weather Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Parry, D.W.; Jenkinson, P.; McLeoad, L. Fusarium ear blight (scab) in small grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Logrieco, A.; Bottalico, A.; Mule, G.; Morietti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some miediterranean crops. Eur. J. Plant Pathol. 2003, 109, 557–645. Available online: https://www.researchgate.net/publication/225231648 (accessed on 6 February 2020). [CrossRef]
- McMullen, M.; Bergstrom, G.C.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium head blight, mycotoxins and strategies for their reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef] [Green Version]
- Bottallico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. J. Plant. Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Cooke, B.M.; Doohan, F.M.; Edwards, S.G. Relationship between the fungal complex causing Fusarium head blight of wheat and environmental conditions. Phytopathology 2008, 98, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium species, chemotype characterisation and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef]
- Backhouse, D.; Burgess, L.W.; Summerell, B.A. Biogeography of Fusarium. In Fusarium Nelson Memorial Symposium; Summerell, B.A., Leslie, J.F., Backhouse, D., Bryden, W.L., Burgess, L.W., Eds.; APS Press: St Paul, MI, USA, 2012; pp. 122–137. [Google Scholar]
- Miedaner, T.; Reinbrecht, C.; Lauber, U.; Schollenberger, M.; Geiger, H.H. Effects of genotype and genotype-environment interaction on deoxynivalenol accumulation and resistance to Fusarium head blight in rye, triticale, and wheat. Plant Breed. 2001, 120, 97–105. [Google Scholar] [CrossRef]
- Kosiak, B.; Trop, M.; Skjerve, E.; Andersen, B. Alternaria and Fusarium in Norwegian grains of reduced quality-a matched pair sample study. Int. J. Food Microbiol. 2004, 93, 51–62. [Google Scholar] [CrossRef]
- How to cite this resource - Schoch CL, et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database (Oxford). Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?mode=Tree&id=450425&lvl=3&lin=f&unlock (accessed on 8 December 2021).
- Fernandez, M.R.; Chen, Y. Pathogenicity of Fusarium species on different plan parts of spring wheat under controlled conditions. Plant Dis. 2005, 89, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamburic-Ilincic, L. Fusarium species and mycotoxins associated with oat in southwestern Ontario, Canada. Can. J. Plant Sci. 2010, 90, 211–216. [Google Scholar] [CrossRef]
- Kosiak, B.; Trop, M.; Skjerwe, E.; Thrane, U. The prevalence and distribution of Fusarium species in Norwegian cereals: A survey. Acta Agric. Scand. Sect. B Soil Plant Sci. 2003, 53, 168–176. [Google Scholar]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gräfenhan, T.L.; Patrick, S.K.; Roscoe, M.; Trelka, R.; Gaba, D.; Chan, J.M.; McKendry, T.; Clear, R.M.; Tittlemier, S.A. Fusarium damage in cereal grains from Western Canada. 1. Phylogenetic analysis of moniliformin-producing Fusarium species and their natural occurrence in mycotoxin-contaminated wheat, oats, and rye. J. Agric. Food Chem 2013, 61, 5425–5437. [Google Scholar] [CrossRef] [PubMed]
- Buerstmayr, H.; Lemmens, M. Breeding healthy cereals: Genetic improvement of Fusarium resistance and consequences for mycotoxins. World Mycotox. J. 2015, 8, 591–602. [Google Scholar] [CrossRef]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.-L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European Database Fusarium graminearum and F. culmorum Trichothecene Genotypes. Front. Microbiol. 2016, 7, 406. [Google Scholar] [CrossRef] [Green Version]
- Yli-Mattila, T. Ecology and evolution of toxigenic Fusarium species in cereals in Northern Europe and Asia. J. Plant Pathol. 2010, 92, 7–18. Available online: https://www.jstor.org/stable/41998764 (accessed on 8 December 2021).
- Janssen, E.M.; Mourits, M.C.M.; Fels-Klerx, H.J.; Oude Lansink, A.G.J.M. Pre-harvest measures against Fusarium spp. Infection and related mycotoxins implemented by Dutch wheat farmers. Crop Prot. 2019, 122, 9–18. [Google Scholar] [CrossRef]
- Weber, R.; Kita, W. Effects of Tillage system and forecrop type on frequency of Fusarium culmorum and F. avenaceum occurrence on culm base of some winter wheat (Triticum aestivum L.) cultivars. Acta Agrobot. 2010, 63, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Langseth, W.; Bernhoft, A.; Rundberget, T.; Kosiak, B.; Gareis, M. Mycotoxin production and cytotoxity of Fusarium strains isolated from Norwegian cereals. Mycopathologia 1999, 144, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Hestbjerg, H.; Nielsen, K.F.; Thrane, U.; Elmoholt, S. Production of trichothecenes and other secondary metabolites by Fusarium culmorum and Fusarium equiseti on common laboratory media and a soil organic matter agar an ecological interpretation. J. Agric Food Chem. 2002, 50, 7593–7599. [Google Scholar] [CrossRef]
- Morrison, E.; Rundberget, T.; Kosiak, B.; Aastveit, A.H.; Bernhoft, A. Cytotoxicity of trichothecenes and fusarochromanone produced by Fusarium equiseti strains isolated from Norwegian cereals. Mycopathologia 2002, 153, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Moss, M.; Thrane, H. Fusarium taxonomy with relations to trichothecene formation. Toxicol. Lett. 2004, 153, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kosiak, E.B.; Holst-Jensen, A.; Runtberget, T.; Jaen, M.T.G.; Torp, M. Morphological, chemical and molecular differentiation of Fusarium equiseti isolated from Norwegian cereals. Int. J. Food Microbiol. 2005, 99, 195–206. [Google Scholar] [CrossRef]
- Mielniczuk, E. The occurrence of Fusarium spp. on panicles of oat (Avena sativa L.). J. Plant Prot. Res. 2001, 41, 173–180. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium species. In An Illustrated Manual of Identification; The Pensylvania State University: State College, PA, USA; Press University Park and London: London, UK, 1983. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- Mishra, C.B.P.; Behr, L. Der Einfluss von Kulturfiltraten von Fusarium culmorum (W.G.Sm.) Sacc., Fusarium avenaceum (Fr.) Sacc. und Fusarium nivale (Fr.) Ces., Griphosphaeria nivalis Müller et v. Arx auf die Keimung des Weizen. Arch. Phytopathol. Plant Prot. 1976, 12, 373–377. [Google Scholar] [CrossRef]
- Kiecana, I.; Mielniczuk, E.; Kaczmarek, Z.; Kostecki, M.; Goliński, P. Scab response and moniliformin accumulation in kernels of oat genotypes inoculated with Fusarium avenaceum in Poland. Eur. J. Plant Pathol. 2002, 108, 245–251. [Google Scholar] [CrossRef]
- Mielniczuk, E.; Cegiełko, M. Fungi occurred on marigold (Tagetes L.) and harmfulness of Fusarium species to selected cultivars. Acta Sci. Pol. Hortorum Cultus 2019, 18, 193–203. [Google Scholar] [CrossRef]
- Perkowski, J.; Kiecana, I.; Kaczmarek, Z. Natural occurrence and distribution of Fusarium toxins in contaminated barley cultivars. Eur. J. Plant Pathol. 2003, 109, 331–339. [Google Scholar] [CrossRef]
- Lemmens, M.; Krska, R.; Buerstmayr, H.; Josephs, R.; Schumacher, R.; Grausgruber, H.; Ruckenbauer, P. Fusarium head blight reactions and accumulation of deoxynivalenol, moniliformin and zearalenone in wheat grains. Cer. Res. Comm. 2003, 31, 407–414. [Google Scholar] [CrossRef]
- Goliński, P.; Waśkiewicz, A.; Wiśniewska, H.; Kiecana, I.; Mielniczuk, E.; Gromadzka, M.; Kostecki, M.; Bocianowski, J.; Rymaniak, E. Reaction of winter wheat (Triticum aestivum L.) cultivars to infection with Fusarium spp. mycotoxins contamination in grain and chaff. Food Add. Contam. A 2010, 27, 1015–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christ, D.S.; Gödecke, R.; von Tiedemann, A.; Varrelmann, M. Pathogenicity, symptom development and mycotoxin formation in wheat by Fusarium species frequently isolated from sugar beet. Phytopathology 2011, 101, 1338–1345. [Google Scholar] [CrossRef] [Green Version]
- Gagkaeva, T.; Gavrilova, O.; Yli-Mattila, T.; Loskutov, I. Evaluation of oat germplasm for resistance to Fusarium head blight. Plant Breed. Seed Sci. 2011, 64, 15–22. Available online: http://ojs.ihar.edu.pl/index.php/pbss/article/view/338/290 (accessed on 8 December 2021). [CrossRef]
- Linkmeyer, A.; Götz, M.; Hu, L.; Asam, S.; Rychlik, M.; Hausladen, H.; Hess, M.; Hückelhoven, R. Assessment and introduction of quantitative resistance to Fusarium head blight in elite spring barley. Phytopathology 2013, 103, 1252–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Hwang, S.F.; Eudes, F.; Chang, K.F.; Howard, R.J.; Turnbul, G.D. Trichothecenes and aggressiveness of Fusarium graminearum causing seedling blight and root rot in cereals. Plant Pathol. 2006, 55, 224–230. [Google Scholar] [CrossRef]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Sullyok, M.; Hecker, A.; Jenny, E.; Krska, R.; Schuhmacher, R.; Forrer, H.-R. Toxigenicity and pathogenicity of Fusarium poae and Fusarium avenaceum on wheat. Eur. J. Plant Pathol. 2008, 122, 265–276. [Google Scholar] [CrossRef]
- Beccari, G.; Covarelli, L.; Nicholson, P. Infection process and soft wheat response to root rot and crown rot caused by Fusarium culmorum. Plant Pathol. 2011, 60, 671–684. [Google Scholar] [CrossRef]
- McCallum, B.D.; Tekauz, A. Influence of inoculation method and growth stage on fusarium head blight in barley. Can. J. Plant Pathol. 2002, 24, 77–80. [Google Scholar] [CrossRef]
- Vančo, B.; Šliková, S.; Šudyova, V.; Šrobárová, A. Response to Fusarium culmorum inoculation in barley. Biologia 2007, 62, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Kiecana, I.; Cegiełko, M.; Mielniczuk, E.; Perkowski, J. The occurrence of Fusarium poae (Peck) Wollenw. on oat (Avena sativa L.) panicles and its harmfulness. Acta Agrobot. 2012, 65, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Mielniczuk, E.; Kiecana, I.; Perkowski, J. Susceptybility of oat genotypes to Fusarium crookwellense Burgess, Nelson and Toussoun infection and mycotoxin accumulation in kernels. Biologia 2004, 59, 809–816. [Google Scholar]
- Kiecana, I.; Mielniczuk, E.; Cegiełko, M.; Perkowski, J. Panicles blight and biosynthesis of Fusarium toxins in oats kernels field inoculated with Fusarium culmorum (W.G.Sm.) Sacc. In Proceedings of the 11th European Fusarium Seminar “Fusarium—Mycotoxins, Taxonomy, Pathogenicity and Host Resistance”, Book of Abstracts, Radzików, Poland, 20–23 September 2010; Plant Breeding and Acclimatization Institute—National Research Institute: Radzików, Poland, 2010; p. 281. [Google Scholar]
- Van Coller, G.J.; Sedeman, Z.A.R.; Boutigny, A.L.; Rose, L.; Lamprecht, S.C.; Viljoen, A. Fusarium species associated with head blight of wheat in South Africa. In Proceedings of the 11th European Fusarium Seminar “Fusarium—Mycotoxins, Taxonomy, Pathogenicity and Host Resistance”, Book of Abstracts, Radzików, Poland, 20–23 September 2010; Plant Breeding and Acclimatization Institute—National Research Institute: Radzików, Poland, 2010; pp. 217–218. [Google Scholar]
- Kiecana, I.; Mielniczuk, E.; Cegiełko, M.; Pastucha, A. The occurrence of Fusarium spp. on oat (Avena sativa L.) and susceptibility of seedlings of selected genotypes to infection with Fusarium graminearum Schwabe. Acta Agrobot. 2014, 67, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Majchrzak, B.; Kurowski, T.P.; Okorski, A. Fungi isolated from the roots and stem bases of spring wheat grown after different cruciferous plants as forecrops. Pol. J. Natur. Sci. 2008, 23, 299–309. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium–Mycotoxins Chemistry Genetics and Biology; APS Press: St. Paul, MI, USA, 2006. [Google Scholar]
- Chełkowski, J.; Kaptur, P.; Tomkowiak, M.; Kostecki, M.; Goliński, P.; Ponitka, A.; Ślusarkiewicz-Jarzina, A.; Bocianowski, J. Moniliformin accumulation in kernels of triticale accessions inoculated with Fusarium avenaceum, in Poland. J. Phytopathol. 2000, 148, 433–439. [Google Scholar] [CrossRef]
- Perkowski, J.; Miedaner, T.; Geiger, H.H.; Müller, H.N.; Chełkowski, J. Occurrence of deoxynivalenol (DON), 3-acetyl-DON, zearalenone and ergosterol in winter rye inoculated with Fusarium culmorum. Cereal Chem. 1995, 72, 205–209. [Google Scholar]
- Wiśniewska, H.; Perkowski, J.; Kaczmarek, Z. Scab Response and deoxyniva-lenol accumulation in spring wheat kernels of different geographical origins following inoculation with Fusarium culmorum. J. Phytopathol. 2004, 152, 613–621. [Google Scholar] [CrossRef]
- Snijders, C.H.A.; Perkowski, J. Effect of head blight caused by Fusarium culmorum on toxin content and weight of wheat kernels. Phytopathology 1990, 80, 556–570. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Elements of Yield Structure | |||||
|---|---|---|---|---|---|---|
| Number of Kernels | Kernels Yield (g) | 1000 Kernels Weight (g) | ||||
| F. eq. Inoculated | Control | F. eq. Inoculated | Control | F. eq. Inoculated | Control | |
| Agent | 48.70 * | 71.57 | 18.31 * | 27.89 | 37.57 * | 39.86 |
| Elegant | 61.48 | 69.12 | 19.42 * | 25.14 | 30.66 * | 36.26 |
| Denar | 45.38 * | 72.65 | 14.30 * | 24.37 | 30.84 * | 35.60 |
| Kozak | 59.24 * | 79.58 | 18.21 * | 29.38 | 33.27 * | 38.17 |
| Romulus | 61.95 * | 74.75 | 19.35 * | 28.37 | 30.32 * | 38.24 |
| DC06011-8 | 58.62 * | 91.30 | 18.53 * | 32.11 | 32.73 * | 37.02 |
| DC 14-8 | 54.21 * | 73.50 | 15.93 * | 24.58 | 30.63 * | 33.99 |
| POB 4109/10 | 35.07 * | 69.47 | 10.87 * | 24.77 | 24.60 * | 33.07 |
| POB 6020/10 | 60.17 * | 83.63 | 18.71 * | 27.09 | 25.71 * | 30.08 |
| POB 961-1344/13 | 48.87 * | 83.84 | 14.45 * | 27.94 | 23.99 * | 34.62 |
| Average | 53.37 * | 76.94 | 16.81 * | 27.16 | 30.03 * | 35.69 |
| Variabiles Analyzed Parametrs | RKY | RNKP | RTKW |
|---|---|---|---|
| RKY | 1 | 0.821 * | 0.091 |
| RNKP | 0.821 * | 1 | −0.338 * |
| RTKW | 0.091 | −0.338 * | 1 |
| Fungus Species | Number of Isolates in Each Study Year | Total Number of Isolates | ||
|---|---|---|---|---|
| 2015 | 2016 | 2017 | ||
| Alternaria alternata (Fr.) Keissler | 5 | 8 | 10 | 23 |
| Aspergillus niger Link | 0 | 2 | 0 | 2 |
| Cladosporium cladosporioides (Fresen.) G.A. de Vries | 1 | 6 | 4 | 11 |
| Drechslera avenae (Edima) Scharif | 2 | 0 | 0 | 2 |
| Epicoccum nigrum Link ex Link | 34 | 29 | 12 | 75 |
| Fusarium equiseti (Fr.) Sacc. | 433 | 396 | 401 | 1230 |
| Fusarium poae (Peck.) Wr. | 1 | 2 | 1 | 4 |
| Mucor hiemalis Wehmer | 2 | 0 | 0 | 2 |
| Penicillium spp. | 9 | 2 | 5 | 16 |
| Stemphylium botryosum Wallr. | 0 | 2 | 1 | 3 |
| Total | 487 | 447 | 434 | 1368 |
| Genotypes | Average Mycotoxin Content over 3 Years of Research [mg·kg−1] | Sum of Group A Trichothecenes | |||||
|---|---|---|---|---|---|---|---|
| T-2 | HT-2 | DAS | STO | T-2 Triol | T-2 Tetraol | ||
| Agent | 0.0000 | 0.0043 | 0.0023 | 0.0334 | 0.0023 | 0.0020 | 0.0442 a |
| Elegant | 0.0009 | 0.0128 | 0.0056 | 0.0299 | 0.0017 | 0.0029 | 0.0538 b |
| Denar | 0.0050 | 0.0083 | 0.0014 | 0.0308 | 0.0010 | 0.0082 | 0.0547 b |
| Kozak | 0.0000 | 0.0055 | 0.0247 | 0.0290 | 0.0003 | 0.0014 | 0.0610 b |
| Romulus | 0.0007 | 0.0282 | 0.0162 | 0.0325 | 0.0005 | 0.0132 | 0.0913 c |
| DC06011-8 | 0.0000 | 0.0067 | 0.0029 | 0.0298 | 0.0003 | 0.0050 | 0.0448 a |
| DC 14-8 | 0.0003 | 0.0100 | 0.0009 | 0.0289 | 0.0002 | 0.0067 | 0.0470 a |
| POB 4109/10 | 0.0147 | 0.0113 | 0.0033 | 0.0391 | 0.0010 | 0.0179 | 0.0872 c |
| POB 6020/10 | 0.0020 | 0.0075 | 0.0010 | 0.0305 | 0.0007 | 0.0091 | 0.0507 b |
| POB 961-1344/13 | 0.0003 | 0.0156 | 0.0015 | 0.0429 | 0.0010 | 0.0213 | 0.0827 c |
| Average mycotoxin content | 0.0024 | 0.0110 | 0.0059 | 0.0327 | 0.0009 | 0.0088 | 0.0616 |
| Years | Mycotoxin Contents [mg·kg−1], Average for All Genotypes | ||||||
|---|---|---|---|---|---|---|---|
| T-2 toxin | HT-2 toxin | DAS | STO | T-2 Triol | T-2 Tetraol | Sum of Group A Trichothecenes | |
| 2015 | 0.0001 | 0.0138 | 0.0003 | 0.0798 | 0.0001 | 0.0056 | 0.0997 c |
| 2016 | 0.0003 | 0.0075 | 0.0022 | 0.005 | 0.0002 | 0.0058 | 0.0210 a |
| 2017 | 0.0068 | 0.0118 | 0.0151 | 0.0133 | 0.0024 | 0.0149 | 0.0643 b |
| Average mycotoxin content | 0.0024 | 0.0110 | 0.0059 | 0.0327 | 0.0009 | 0.0088 | 0.0616 |
| Genotypes | Average Mycotoxin Contents over 3 Years of Research [mg·kg−1] | |||||
|---|---|---|---|---|---|---|
| DON | 3-ACDON | 15-AcDON | NIV | FUS X | Sum of Group B Trichothecenes | |
| Agent | 0.0227 | 0.0073 | 0.0067 | 0.1860 | 0.0094 | 0.2322 b |
| Elegant | 0.0160 | 0.0077 | 0.0130 | 0.1793 | 0.0105 | 0.2264 b |
| Denar | 0.0177 | 0.0157 | 0.0055 | 0.0634 | 0.0078 | 0.1101 a |
| Kozak | 0.0495 | 0.0113 | 0.0053 | 0.2827 | 0.0085 | 0.3574 c |
| Romulus | 0.0365 | 0.0173 | 0.0198 | 0.2564 | 0.0084 | 0.3384 c |
| DC06011-8 | 0.0277 | 0.0087 | 0.0105 | 0.1456 | 0.0301 | 0.2226 b |
| DC 14-8 | 0.0209 | 0.0770 | 0.0070 | 0.0859 | 0.0106 | 0.2014 b |
| POB 4109/10 | 0.0116 | 0.0153 | 0.0058 | 0.0537 | 0.0082 | 0.0945 a |
| POB 6020/10 | 0.0517 | 0.0150 | 0.0055 | 0.0343 | 0.0081 | 0.1146 a |
| POB 961-1344/13 | 0.0343 | 0.0153 | 0.0053 | 0.0745 | 0.0081 | 0.1376 a |
| Average mycotoxin content | 0.0287 | 0.0191 | 0.0084 | 0.1362 | 0.0110 | 0.2034 |
| Years | Mycotoxin Contents [mg·kg−1] | |||||
|---|---|---|---|---|---|---|
| DON | 3-Ac DON | 15-Ac DON | NIV | FUS X | Sum of Group B Trichothecenes | |
| 2015 | 0.0194 | 0.0421 | 0.0159 | 0.0692 | 0.0242 | 0.1708 a |
| 2016 | 0.0190 | 0.0000 | 0.0032 | 0.0938 | 0.0040 | 0.1201 a |
| 2017 | 0.0477 | 0.0151 | 0.0062 | 0.2455 | 0.0047 | 0.3192 b |
| Average | 0.0287 | 0.0191 | 0.0084 | 0.1362 | 0.0110 | 0.2034 |
| Month | Air Temperature [°C] | |||
|---|---|---|---|---|
| Average for the Years 1971–2005 | 2015 | 2016 | 2017 | |
| April | 7.9 | 7.5 | 9.1 | 7.5 |
| May | 14.1 | 13.0 | 14.7 | 14.31 |
| June | 16.8 | 17.0 | 18.5 | 17.5 |
| July | 18.4 | 20.0 | 19.8 | 18.7 |
| August | 17.8 | 21.5 | 18.3 | 19.5 |
| Rainfall [mm] | ||||
| Average for the Years 1971–2005 | 2015 | 2016 | 2017 | |
| April | 44.1 | 40.0 | 90.0 | 35.0 |
| May | 65.5 | 114.8 | 85.0 | 54.0 |
| June | 78.9 | 55.0 | 48.0 | 45.0 |
| July | 98.4 | 64.8 | 99.0 | 83.0 |
| August | 54.3 | 4.8 | 50.0 | 36.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mielniczuk, E.; Wit, M.; Patkowska, E.; Cegiełko, M.; Wakuliński, W. Reaction of Oat Genotypes to Fusarium equiseti (Corda) Sacc. Infection and Mycotoxin Concentrations in Grain. Agronomy 2022, 12, 295. https://doi.org/10.3390/agronomy12020295
Mielniczuk E, Wit M, Patkowska E, Cegiełko M, Wakuliński W. Reaction of Oat Genotypes to Fusarium equiseti (Corda) Sacc. Infection and Mycotoxin Concentrations in Grain. Agronomy. 2022; 12(2):295. https://doi.org/10.3390/agronomy12020295
Chicago/Turabian StyleMielniczuk, Elżbieta, Marcin Wit, Elżbieta Patkowska, Małgorzata Cegiełko, and Wojciech Wakuliński. 2022. "Reaction of Oat Genotypes to Fusarium equiseti (Corda) Sacc. Infection and Mycotoxin Concentrations in Grain" Agronomy 12, no. 2: 295. https://doi.org/10.3390/agronomy12020295