Cupuassu (Theobroma grandiflorum [Willd. ex Sprengel] Schumann) Fruit Development: Key Genes Involved in Primary Metabolism and Stress Response

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Cupuassu Pulp Quality Analyses
2.3. RNA Extraction and cDNA Synthesis
2.4. Sequence Selection, Functional Annotation and Interactomics Analysis
2.5. Primer Design and Quantitative PCR
3. Results
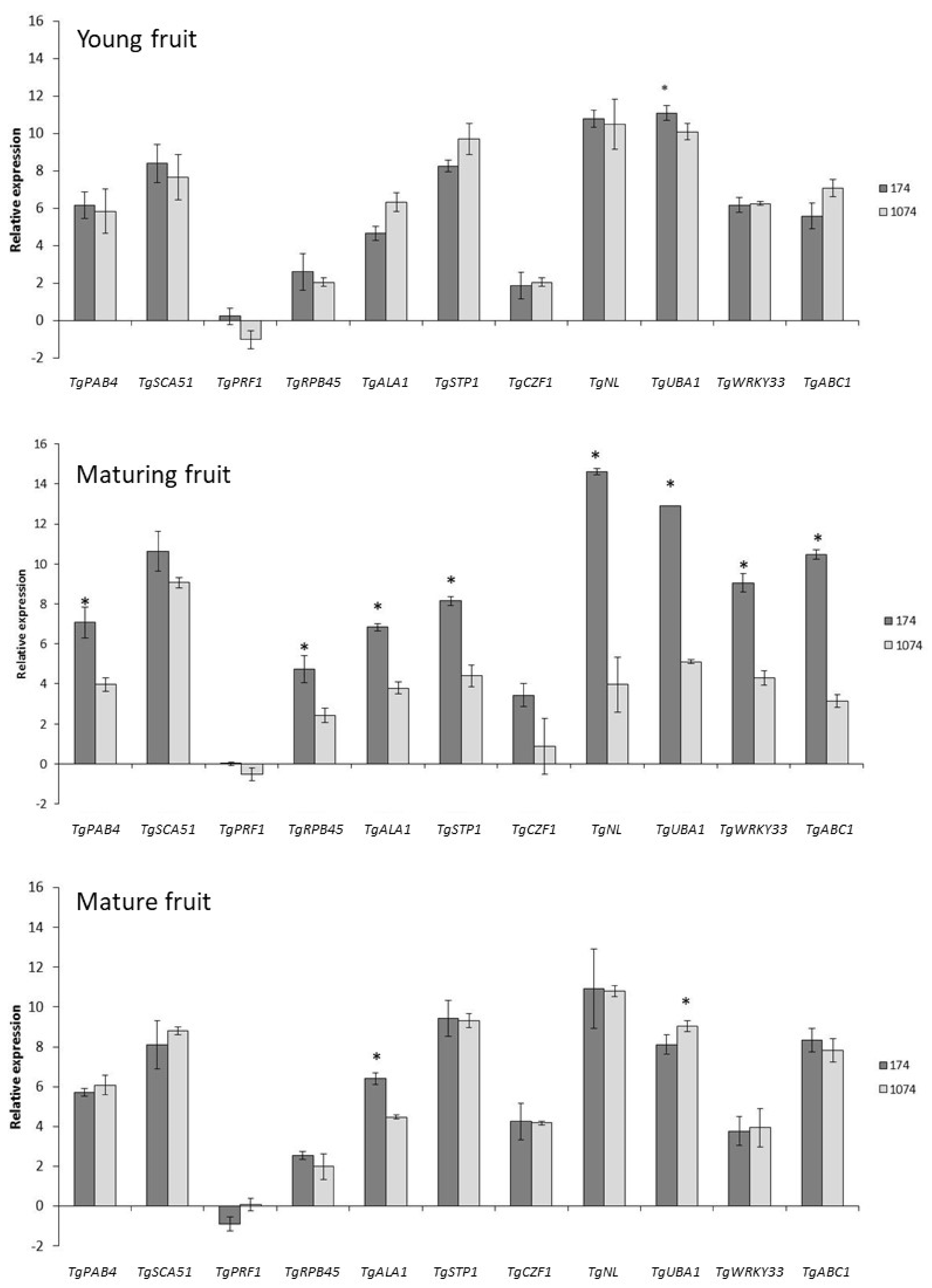
3.1. Gene Expression during Fruit Maturation
3.2. Interactomics and Ontology
4. Discussion
4.1. Cupuassu Genotypes Showed Different Pulp Characteristics and Gene Expression Patterns
4.2. Monosaccharide and Carbohydrate Transport Pathways Were Enhanced in Cupuassu Fruit during Maturation
4.3. In Maturing Cupuassu Fruits, Sugar Accumulation Participated in Signal Transduction Associated with Fruit Development and Stress Response
4.4. Maturing Cupuassu Pulp and Seeds Showed Increased Phospholipid Metabolism and Translocation, as Well as Immune System Activation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbalho, G.N.; Matos, B.N.; da Silva Brito, G.F.; da Cunha Miranda, T.; Alencar-Silva, T.; Sodré, F.F.; Gelfuso, G.M.; Cunha-Filho, M.; Carvalho, J.L.; da Silva, J.; et al. Skin Regenerative Potential of Cupuaçu Seed Extract (Theobroma grandiflorum), a Native Fruit from the Amazon: Development of a Topical Formulation Based on Chitosan-Coated Nanocapsules. Pharmaceutics 2022, 14, 207. [Google Scholar] [CrossRef]
- Gondim, T.M.S.; Thomazini, M.J.; Calvalcante, M.J.B.; Souza, J.M.L. Aspectos da Produção de Cupuaçu; Acre, E., Ed.; Embrapa: Rio Branco, Brazil, 2001; Volume 67. [Google Scholar]
- Nazaré, R.F.R.; Barbosa, W.C.; Viégas, R.M.F. Processamento das Sementes de Cupuaçu para Obtenção de Cupulate; Embrapa Amazônia Oriental: Belém, Brazil, 1990; p. 38. [Google Scholar]
- Barbosa, W.C.; Nazaré, R.F.R.; Nagata, I. Estudo Tecnolpogico de Frutas da Amazônia; Embrapa (Centro de Pesquisa Agropecuário do Trópico Úmido): Belém, Brazil, 1978; p. 19. [Google Scholar]
- Cohen, K.O.; Jackix, M.N.H. Estudo do liquor de cupuaçu. Food Sci. Technol. 2005, 25, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.M.; Resende, M.D.V.d.; Bandeira, B.d.S.; Pinheiro, T.M.; Farias, D.C.R. Avaliação e seleção de progênies de cupuaçuzeiro (Theobroma grandiflorum), em Belém, Pará. Rev. Bras. Frutic. 2010, 32, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.M.; Silva, C.R.S.; Silva, M.S.C.; Silva, D.C.S.; Sebbenn, A.M. Diversidade genética em coleções amazônicas de germoplasma de cupuaçuzeiro [Theobroma grandiflorum (Willd. ex Spreng.) Schum.]. Rev. Bras. Frutic. 2013, 35, 818–828. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.M.; Resende, M.D.V. Avaliação genética de individuos e progênies de cupuaçuzeiro no estado do Para e estimativas de parâmetros genéticos. Rev. Bras. Frutic. 2008, 30, 696–701. [Google Scholar] [CrossRef]
- Alves, R.M.; Resende, M.D.V.; Bandeira, B.S.; Pinheiro, T.M.; Farias, D.C.R. Evolução da vassoura-de-bruxa e avaliação da resistência em progênies de cupuaçuzeiro. Rev. Bras. Frutic. 2009, 31, 1022–1032. [Google Scholar] [CrossRef] [Green Version]
- Ferraz Dos Santos, L.; Moreira Fregapani, R.; Falcão, L.L.; Togawa, R.C.; Costa, M.M.d.C.; Lopes, U.V.; Peres Gramacho, K.; Alves, R.M.; Micheli, F.; Marcellino, L.H. First Microsatellite Markers Developed from Cupuassu ESTs: Application in Diversity Analysis and Cross-Species Transferability to Cacao. PLoS ONE 2016, 11, e0151074. [Google Scholar] [CrossRef] [Green Version]
- Falcao, L.L.; Werneck, J.O.S.; Togawa, R.C.; Costa, M.M.d.C.; Grynberg, P.; Silva Junior, O.B.d.; Alves, R.M.; Albuquerque, P.S.B.; Marcellino, L.H. Analyses of cupuassu (Theobroma grandiflorum) transcriptome during interaction with Moniliophthora perniciosa, the causal agent of Witches’ Broom disease. In Proceedings of the 7th Brazilian Biotechnology Congress and 2nd Biotechnology Ibero-American Congress, Brasília, Brazil, 18–21 November 2008. [Google Scholar]
- Santana Silva, R.J.; Alves, R.M.; Peres Gramacho, K.; Marcellino, L.H.; Micheli, F. Involvement of structurally distinct cupuassu chitinases and osmotin in plant resistance to the fungus Moniliophthora perniciosa. Plant Physiol. Biochem. 2020, 148, 142–151. [Google Scholar] [CrossRef]
- Chuang, H.-Y.; Hofree, M.; Ideker, T. A decade of systems biology. Annu. Rev. Cell Dev. Biol. 2010, 26, 721–744. [Google Scholar] [CrossRef]
- Dangi, A.K.; Sharma, B.; Khangwal, I.; Shukla, P. Combinatorial Interactions of Biotic and Abiotic Stresses in Plants and Their Molecular Mechanisms: Systems Biology Approach. Mol. Biotechnol. 2018, 60, 636–650. [Google Scholar] [CrossRef]
- Da Hora Junior, B.T.; Poloni, J.d.F.; Lopes, M.A.; Dias, C.V.; Gramacho, K.P.; Schuster, I.; Sabau, X.; Cascardo, J.C.D.M.; Mauro, S.M.Z.D.; Gesteira, A.d.S.; et al. Transcriptomics and systems biology analysis in identification of specific pathways involved in cacao resistance and susceptibility to witches’ broom disease. Mol. Biosyst. 2012, 8, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- De Andrade Silva, E.M.; Reis, S.P.M.; Argolo, C.S.; Gomes, D.S.; Barbosa, C.S.; Gramacho, K.P.; Ribeiro, L.F.; Silva, R.J.S.; Micheli, F. Moniliophthora perniciosa development: Key genes involved in stress-mediated cell wall organization and autophagy. Int. J. Biol. Macromol. 2020, 154, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.M.; Garcia, A.A.F.; Cruz, E.D.; Figueira, A. Seleção de descritores botânico-agronômicos para caracterização de germoplasma de cupuaçuzeiro. Pesqui. Agropecu. Bras. 2003, 38, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Report. Ispmb 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Logemann, J.; Schell, J.; Willmitzer, L. Improved method for the isolation of RNA from plant tissues. Anal. Biochem. 1987, 163, 16–20. [Google Scholar] [CrossRef]
- Santana, R.J.S.; Silva-Werneck, J.O.; Falcao, L.L.; Marcellino, L.H. Extração de RNA de cupuaçuzeiro (Theobroma grandiflorum) para estudos de expressão gênica. In Proceedings of the Congresso Nacional de Ciências Biológicas, Recife, Brazil, 7–11 November 2011; pp. 387–391. [Google Scholar]
- Argout, X.; Fouet, O.; Wincker, P.; Gramacho, K.; Legavre, T.; Sabau, X.; Risterucci, A.M.; Da Silva, C.; Cascardo, J.; Allegre, M.; et al. Towards the understanding of the cocoa transcriptome: Production and analysis of an exhaustive dataset of ESTs of Theobroma cacao L. generated from various tissues and under various conditions. BMC Genom. 2008, 9, 512. [Google Scholar] [CrossRef] [Green Version]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The Arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.J. BLAT—The BLAST-Like Alignment Tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology Consortium. Gene Ontology Consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Kucera, M.; Isserlin, R.; Arkhangorodsky, A.; Bader, G.D. AutoAnnotate: A Cytoscape app for summarizing networks with semantic annotations. F1000Research 2016, 5, 1717. [Google Scholar] [CrossRef]
- Morris, J.H.; Apeltsin, L.; Newman, A.M.; Baumbach, J.; Wittkop, T.; Su, G.; Bader, G.D.; Ferrin, T.E. clusterMaker: A multi-algorithm clustering plugin for Cytoscape. BMC Bioinform. 2011, 12, 436. [Google Scholar] [CrossRef] [Green Version]
- Cordoba, E.; Aceves-Zamudio, D.L.; Hernández-Bernal, A.F.; Ramos-Vega, M.; León, P. Sugar regulation of SUGAR TRANSPORTER PROTEIN 1 (STP1) expression in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Ferraz dos Santos, L.; Santana Silva, R.J.; Oliveira Jordão do Amaral, D.; Barbosa de Paula, M.F.; Ludke Falcão, L.; Legavre, T.; Alves, R.M.; Marcellino, L.H.; Micheli, F. Selection of Reference Genes for Expression Study in Pulp and Seeds of Theobroma grandiflorum (Willd. ex Spreng.) Schum. PLoS ONE 2016, 11, e0160646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Fernald, R.D. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Canteri, M.G.; Althaus, R.A.; das Virgens Filho, J.S.; Giglioti, E.A.; Godoy, C.V. SASM-AGRI—Sistema para analise e separação de médias em experimentos agricolas pelos métodos Scott-Knott, Tukey a Duncan. Rev. Bras. Agrocomputação 2001, 1, 18–24. [Google Scholar]
- Vriesmann, L.C.; de Oliveira Petkowicz, C.L. Polysaccharides from the pulp of cupuassu (Theobroma grandiflorum): Structural characterization of a pectic fraction. Carbohydr. Polym. 2009, 77, 72–79. [Google Scholar] [CrossRef]
- Büttner, M. The Arabidopsis sugar transporter (AtSTP) family: An update. Plant Biol. 2010, 12, 35–41. [Google Scholar] [CrossRef]
- Fujiki, Y.; Ito, M.; Nishida, I.; Watanabe, A. Multiple signaling pathways in gene expression during sugar starvation. Pharmacological analysis of din gene expression in suspension-cultured cells of Arabidopsis. Plant Physiol. 2000, 124, 1139–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, T.; Maekawa, S.; Yanagisawa, S. The Pre-rRNA Processing Complex in Arabidopsis Includes Two WD40-Domain-Containing Proteins Encoded by Glucose-Inducible Genes and Plant-Specific Proteins. Mol. Plant 2016, 9, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.C.; Hsu, Y.F.; Chen, Y.C.; Chang, Y.L.; Wang, C.S. A WD40 protein, AtGHS40, negatively modulates abscisic acid degrading and signaling genes during seedling growth under high glucose conditions. J. Plant Res. 2016, 129, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Yan, F.; Hu, Z.; Wei, S.; Lai, J.; Chen, G. Accumulation of Anthocyanin and Its Associated Gene Expression in Purple Tumorous Stem Mustard (Brassica juncea var. tumida Tsen et Lee) Sprouts When Exposed to Light, Dark, Sugar, and Methyl Jasmonate. J. Agric. Food Chem. 2019, 67, 856–866. [Google Scholar] [CrossRef]
- Sheen, J. Master Regulators in Plant Glucose Signaling Networks. J. Plant Biol. 2014, 57, 67–79. [Google Scholar] [CrossRef]
- Yu, J.Q.; Li, X.M.; Wang, W.Y.; Gu, K.D.; Sun, C.H.; You, C.X.; Hu, D.G. Glucose sensor MdHXK1 activates an immune response to the fungal pathogen Botryosphaeria dothidea in apple. Physiol. Plant 2021, 174, e13596. [Google Scholar] [CrossRef]
- Gai, X.T.; Jiang, N.; Ma, J.; Wang, A.; Lu, C.; Xuan, Y.H.; Xia, Z.Y. NtSWEET1 promotes tobacco resistance to Fusarium oxysporum-induced root rot disease. Plant Signal. Behav. 2021, 16, 1970940. [Google Scholar] [CrossRef]
- Yamada, K.; Saijo, Y.; Nakagami, H.; Takano, Y. Regulation of sugar transporter activity for antibacterial defense in Arabidopsis. Science 2016, 354, 1427–1430. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.-P.; Shen, J.-L.; Li, W.-J.; Wu, N.; Chen, C.; Hou, Y.-X. Evolutionary and Characteristic Analysis of RING-DUF1117 E3 Ubiquitin Ligase Genes in Gossypium Discerning the Role of GhRDUF4D in Verticillium dahliae Resistance. Biomolecules 2021, 11, 1145. [Google Scholar] [CrossRef]
- Sun, J.; Jiang, H.; Xu, Y.; Li, H.; Wu, X.; Xie, Q.; Li, C. The CCCH-type zinc finger proteins AtSZF1 and AtSZF2 regulate salt stress responses in Arabidopsis. Plant Cell Physiol. 2007, 48, 1148–1158. [Google Scholar] [CrossRef]
- Wibowo, A.; Becker, C.; Marconi, G.; Durr, J.; Price, J.; Hagmann, J.; Papareddy, R.; Putra, H.; Kageyama, J.; Becker, J.; et al. Hyperosmotic stress memory in Arabidopsis is mediated by distinct epigenetically labile sites in the genome and is restricted in the male germline by DNA glycosylase activity. eLife 2016, 5, e13546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roitsch, T. Source-sink regulation by sugar and stress. Curr. Opin. Plant Biol. 1999, 2, 198–206. [Google Scholar] [CrossRef]
- Sato, T.; Maekawa, S.; Yasuda, S.; Sonoda, Y.; Katoh, E.; Ichikawa, T.; Nakazawa, M.; Seki, M.; Shinozaki, K.; Matsui, M.; et al. CNI1/ATL31, a RING-type ubiquitin ligase that functions in the carbon/nitrogen response for growth phase transition in Arabidopsis seedlings. Plant J. 2009, 60, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Vargas, F.; Godoy, T.; Tejos, R.; O’Brien, J.A. Overexpression of the Auxin Receptor AFB3 in Arabidopsis Results in Salt Stress Resistance and the Modulation of NAC4 and SZF1. Int. J. Mol. Sci. 2020, 21, 9528. [Google Scholar] [CrossRef]
- Li, S.; Fu, Q.; Chen, L.; Huang, W.; Yu, D. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar] [CrossRef]
- Fan, S.; Wang, D.; Xie, H.; Wang, H.; Qin, Y.; Hu, G.; Zhao, J. Sugar Transport, Metabolism and Signaling in Fruit Development of Litchi chinensis Sonn: A Review. Int. J. Mol. Sci. 2021, 22, 11231. [Google Scholar] [CrossRef]
- Liu, N.J.; Bao, J.J.; Wang, L.J.; Chen, X.Y. Arabidopsis leaf extracellular vesicles in wound-induced jasmonate accumulation. Plant Signal. Behav. 2020, 15, 1833142. [Google Scholar] [CrossRef]
- Liu, N.J.; Wang, N.; Bao, J.J.; Zhu, H.X.; Wang, L.J.; Chen, X.Y. Lipidomic Analysis Reveals the Importance of GIPCs in Arabidopsis Leaf Extracellular Vesicles. Mol. Plant 2020, 13, 1523–1532. [Google Scholar] [CrossRef]
- Guo, Z.; Lu, J.; Wang, X.; Zhan, B.; Li, W.; Ding, S.W. Lipid flippases promote antiviral silencing and the biogenesis of viral and host siRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Gao, H.; Xu, G.; Wu, D.; Song, S.; Jiang, H.; Zhu, S.; Qi, T.; Xie, D. Arabidopsis ALA1 and ALA2 Mediate RNAi-Based Antiviral Immunity. Front. Plant Sci. 2017, 8, 422. [Google Scholar] [CrossRef] [Green Version]
- Goritschnig, S.; Zhang, Y.; Li, X. The ubiquitin pathway is required for innate immunity in Arabidopsis. Plant J. 2007, 49, 540–551. [Google Scholar] [CrossRef]
- Tiwari, S.; Schulz, R.; Ikeda, Y.; Dytham, L.; Bravo, J.; Mathers, L.; Spielman, M.; Guzmán, P.; Oakey, R.J.; Kinoshita, T.; et al. Maternally Expressed PAB C-Terminal, a novel imprinted gene in Arabidopsis, encodes the conserved C-terminal domain of polyadenylate binding proteins. Plant Cell 2008, 20, 2387–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peal, L.; Jambunathan, N.; Mahalingam, R. Phylogenetic and expression analysis of RNA-binding proteins with triple RNA recognition motifs in plants. Mol. Cells 2011, 31, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, Z.; Du, P.; Zhang, C.; Yan, H.; Wang, W.; Li, W. NtRBP45, a nuclear RNA-binding protein of Nicotiana tabacum, facilitates post-transcriptional gene silencing. Plant Direct 2020, 4, e00294. [Google Scholar] [CrossRef]
- Santana Silva, R.J.; Micheli, F. RRGPredictor, a set-theory-based tool for predicting pathogen-associated molecular pattern receptors (PRRs) and resistance (R) proteins from plants. Genomics 2020, 112, 2666–2676. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkhadir, Y.; Nimchuk, Z.; Hubert, D.A.; Mackey, D.; Dangl, J.L. Arabidopsis RIN4 Negatively Regulates Disease Resistance Mediated by RPS2 and RPM1 Downstream or Independent of the NDR1 Signal Modulator and Is Not Required for the Virulence Functions of Bacterial Type III Effectors AvrRpt2 or AvrRpm1. Plant Cell 2004, 16, 2822–2835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burla, B.; Pfrunder, S.; Nagy, R.; Francisco, R.M.; Lee, Y.; Martinoia, E. Vacuolar Transport of Abscisic Acid Glucosyl Ester Is Mediated by ATP-Binding Cassette and Proton-Antiport Mechanisms in Arabidopsis. Plant Physiol. 2013, 163, 1446–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Origin | Physiochemical Characteristics of the Pulp | Response to Witches’ Broom Disease | |||||
|---|---|---|---|---|---|---|---|---|
| pH | SST | ATT | Humidity | ST | SST/ATT | |||
| 174 | Coari-AM | 3.5 | 13.1 | 1.5 | 83.9 | 16.1 | 8.7 | R |
| 1074 | Parintins-AM | 3.5 | 10.7 | 1.8 | 86.0 | 14.0 | 5.9 | S |
| Locus Name 1 | T. grandiflorum Gene Name | Putative Function | T. cacao Orthologue 2 | A. thaliana Orthologue |
|---|---|---|---|---|
| C692 | TgPAB4 | Polyadenylate-binding protein 4 | Tc01v2_p005300.1 | PAB2 |
| C339 | TgSCA51 | Transcription factor SAC51-like | Tc08v2_p005820.1 | AT5G50010 |
| C70 | TgPRF1 | Profilin-1 | Tc09v2_p008530.1 | PRF1 |
| C203 | TgRBP45B | Polyadenylate-binding protein RBP45B | Tc06v2_p017980.1 | RBP45B |
| C366 | TgALA1 | Phospholipid-transporting ATPase 1 | Tc10v2_p010150.1 | ALA1 |
| C28 | TgSTP1 | Sugar carrier protein C | Tc06v2_p016570.1 | STP1 |
| C345 | TgCZF1 | Zinc finger CCCH domain-containing protein 29 | Tc10v2_p001340.1 | CZF1 |
| C5718 | TgNL | Disease resistance protein RPS2 | Tc07v2_p005320.2 | At4g26090 |
| C733 | TgUBA1 | Ubiquitin-activating enzyme E1 1 | Tc09v2_p029770.1 | UBA1 |
| C193 | TgWRKY33 | WRKY transcription factor | Tc09v2_p030160.1 | WRKY33 |
| C4546 | TgABC1 | ABC transporter C family | Tc02v2_p000950.1 | ABCC1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferraz dos Santos, L.; Santana Silva, R.J.; Falcão, L.L.; Alves, R.M.; Marcellino, L.H.; Micheli, F. Cupuassu (Theobroma grandiflorum [Willd. ex Sprengel] Schumann) Fruit Development: Key Genes Involved in Primary Metabolism and Stress Response. Agronomy 2022, 12, 763. https://doi.org/10.3390/agronomy12040763
Ferraz dos Santos L, Santana Silva RJ, Falcão LL, Alves RM, Marcellino LH, Micheli F. Cupuassu (Theobroma grandiflorum [Willd. ex Sprengel] Schumann) Fruit Development: Key Genes Involved in Primary Metabolism and Stress Response. Agronomy. 2022; 12(4):763. https://doi.org/10.3390/agronomy12040763
Chicago/Turabian StyleFerraz dos Santos, Lucas, Raner José Santana Silva, Loeni Ludke Falcão, Rafael Moyses Alves, Lucilia Helena Marcellino, and Fabienne Micheli. 2022. "Cupuassu (Theobroma grandiflorum [Willd. ex Sprengel] Schumann) Fruit Development: Key Genes Involved in Primary Metabolism and Stress Response" Agronomy 12, no. 4: 763. https://doi.org/10.3390/agronomy12040763