
Duckweeds as Promising Food Feedstocks Globally
by
Débora Pagliuso
1, 
Adriana Grandis
1,
Janaina Silva Fortirer
1,
Plinio Camargo
2,
Eny Iochevet Segal Floh
3 and
Marcos Silveira Buckeridge
1,* 1
Laboratory of Plant Physiological Ecology, Department of Botany, Institute of Biosciences, University of São Paulo, São Paulo 05508-090, Brazil
2
Centro de Energia Nuclear na Agricultura, University of São Paulo, Piracicaba 13416-000, Brazil
3
Plant Cell Biology Laboratory, Department of Botany, Institute of Biosciences, University of São Paulo, São Paulo 05508-090, Brazil
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(4), 796; https://doi.org/10.3390/agronomy12040796
Submission received: 1 January 2022
/
Revised: 6 February 2022
/
Accepted: 17 February 2022
/
Published: 26 March 2022
(This article belongs to the Special Issue Using Our Agrobiodiversity: Plant-Based Solutions to Feed the World)
Abstract
:Duckweeds are the smallest flowering plants on Earth. They grow fast on water’s surface and produce large amounts of biomass. Further, duckweeds display high adaptability, and species are found around the globe growing under different environmental conditions. In this work, we report the composition of 21 ecotypes of fourteen species of duckweeds belonging to the two subfamilies of the group (Lemnoideae and Wolffioideae). It is reported the presence of starch and the composition of soluble sugars, cell walls, amino acids, phenolics, and tannins. These data were combined with literature data recovered from 85 publications to produce a compiled analysis that affords the examination of duckweeds as possible food sources for human consumption. We compare duckweeds compositions with some of the most common food sources and conclude that duckweed, which is already in use as food in Asia, can be an interesting food source anywhere in the world.
1. Introduction
Human growth is estimated to reach over 9 billion by 2050 [1], imposing anthropogenic pressure on our food system. This population growth, along with climate change and resource depletion, demands the development of a sustainable food system to secure nutritious food for future generations [2,3,4]. The International Assessment of Agricultural Knowledge, Science, and Technology for Development (IAASTD) highlights that current food systems increase intake of fats, meats, sugar, and salt, with a consumption reduction of fruits and vegetables [5], which is reflected in human health. New agendas and policies are modifying concepts in food production to increase food safety to ensure animal, plant, and human health [1,5]. The 17 Sustainable Development Goals (SDGs) adopted by the United Nations in 2015 are expected to impact each food system framework [6]. The pandemic of COVID-19 highlighted the importance of the SDG 2 goal (zero hunger) once malnutrition and food insecurity became a growing concern worldwide. The slowdown of the economy and the lockdowns adopted in different world regions disrupted the food supply chain, with loss of income and livelihoods, widening inequality, disrupting social protection programs, and altering food environments. These consequences are correlated with uneven food prices in localized contexts [7]. Therefore, improving the food system must help reduce climate change impacts, increase environmental efficiency, and increase the social and economic viability [2,3].
One of the biggest concerns regarding food production is the increase of urbanization and reducing land use for agriculture. In this way, aquaculture has emerged as a promising global food system [8], and duckweeds are a group of plants that can improve agriculture due to their lower impact on land use. Duckweeds are miniature aquatic plants that display high physiological fitness and everlasting life [9]. The Lemnaceae family contains 37 species classified in five genera and sub-grouped in Lemnoideae (Spirodela, Landoltia, and Lemna) and Wolffioideae (Wolffiella and Wolffia) [10,11]. These plants are globally distributed and are adapted to a diverse range of ecological conditions, being able to tolerate a wide range of pHs (from 3 to 10.5) and temperatures (from 17 to 30 °C) [12,13]. Duckweeds can duplicate their biomass within 96 h [14], making them an excellent feedstock for diverse applications, including food sources. Ducks, swans, and geese consume duckweeds naturally [15], and aquaculture also has been employed in animal feed (pigs, cattle, rams, sheep, horses, waterfowls, and fishes) since the 1960s [16].
Concerning human food, Asian countries such as Thailand, the genus Wolffia, especially Wolffia globosa (also named khai-nam, kai-pum, or kai-nhae), are easily found in the food markets [17]. On the other hand, the starch-based diet in India, Bangladesh, Pakistan, and other Asian countries needs protein supplementation due to the lack of a desirable amino acids balance required for human intake [15]. The availability of duckweeds as a food source in several markets led to the investigation of their amino acids contents, and several studies have reported a suitable composition for human ingestion [18,19,20]. Wolffia globosa was cultivated by an Israeli company in a greenhouse precision aquaculture, with a composition of 45% protein, 7% minerals, 37% carbohydrates, 8% fats, and 3% water and providing nine essential amino acids; iron; vitamins A, E, and B12; omega 3; zinc; potassium; and folate [21]. Nonetheless, several studies showed different compositions, revealing that some lineages and species are adequate for human consumption. Nevertheless, a lack of consensus on the composition still disables specific applications and the best nutritional advantage as food.
The potential of duckweed as feedstock is well-known but still unexplored. For these plants to gain recognition, the diverse information available needs to be concise, compiled, and summarized to be reported in the best way to the public utility. The systematic review is a methodology that performs a critical literature search involving a strategic plan designed a priori to reduce bias by identifying all relevant studies with further data analysis and quality evaluation to synthesize all available information [22,23,24,25]. In the present work, we compiled data regarding the concentrations of some key components that characterize plant materials as food sources. We report starch concentrations, soluble sugars, fiber, cell wall, amino acids, phenolics, and tannins from 14 species, which contemplate 21 ecotypes cultivated under laboratory conditions. These data are combined with a literature dataset compiled from a systematic review, using bibliometric analysis to generate an overview of duckweed as feedstock.
2. Materials and Methods
2.1. Plant Material
Duckweed species were obtained from the Rutgers Duckweed Stock Cooperative (RDSC) collection. Twenty-one duckweed specimens (Spirodela polyrhiza 7498, Spirodela polyrhiza 9346, Spirodela polyrhiza 9509, Spirodela polyrhiza 9264, Spirodela intermedia 7355, Landoltia punctata 7488, Landoltia punctata 7624, Landoltia punctata 7776, Lemna yungensis 9208, Lemna minor 8627, Lemna gibba DWC112, Lemna gibba DWC128, Lemna japonica 8695, Lemna obscura 9342, Wolffiella neotropica 8848, Wolffiella caudata 9139, Wolffiella gladiata 8261, Wolffia borealis 9144, Wolffia globosa 884, Wolffia globosa 8973, and Wolffia arrhiza 8853) were cultivated under axenic conditions in 100 mL of ½ Schenk-Hildebrandt medium (pH 6.5) (Sigma-Aldrich, St. Louis, MI, USA) supplemented with 0.5% of sucrose (Sigma-Aldrich, Steinheim, Germany. Plants were grown for 15 days at 25 °C with 16 h of light (20 µmoles m−2s−1). The biomass was recovered, frozen in liquid nitrogen, and lyophilized. The dried samples were pulverized in a grinding ball mill. All tests were performed using five replicates.
2.2. Soluble Sugar and Starch Extraction and Evaluation
Twenty milligrams of material were extracted exhaustively with ethanol 80% (v/v) (Synth, São Paulo, Brazil) at 80 °C for sugar solubilization. The soluble sugars were recovered by centrifugation at 14,000 rpm. The supernatant was freeze-dried (Thermo Scientific, Savant SC 250 EXP, Asheville, NC, USA) and resuspended in 1 mL of water. Sucrose, fructose, glucose, and raffinose (Sigma-Aldrich, Steinheim, Germany) were analyzed using a high-performance anion-exchange chromatography with pulsed amperometric detection (HPAEC-PAD- ICS 5000 system, Dionex-Thermo-Fischer Scientific, Sunnyvale, CA, USA) in a CarboPac PA1 column and eluted with 150-µM sodium hydroxide in an isocratic run of 27 min. The alcohol insoluble residue (AIR) was dried overnight at 45 °C to extract the starch.
The starch content was determined in AIR digested with 120 U·mL−1 of α-amylase (Bacillus licheniformis, Megazyme, Gatton, Australia) in 10-mM MOPS buffer, pH 6.5 at 75 °C for 1 h and 30 U·mL−1 of amyloglucosidase (Aspergillus niger, Megazyme, Gatton, Australia) in 100-mM sodium acetate, pH 4.5 at 50 °C for 1 h. The starch was quantified by a reaction of glucose-free with glucose oxidase (1100 U·mL−1), peroxidase (700 U·mL−1), 4-aminoantipirin (290 µmol·L−1), and 50 mM of phenol at pH 7.5 (Centerlab, Belo Horizonte, Brazil). The reactions were incubated for 15 min at 30 °C and measured at 490 nm. A calibration curve was prepared with glucose (Sigma-Aldrich, Steinheim, Germany) in the concentration range of 0.02–0.2 mg·mL−1.
2.3. Cell Wall Monosaccharides Composition
Cell walls were analyzed for their monosaccharides (apiose, arabinose, fucose, galactose, glucose, mannose, rhamnose, and xylose) by the hydrolysis of two mg of de-starched AIR (cell wall) 1 mL of 2-M trifluoroacetic acid (TFA) for 1 h at 100 °C. The hydrolysis reaction was dried under vacuum, resuspended in 1 mL of deionized water, and filtered on 0.22-µm (Merck Millipore, Darmstadt, Germany) filters. The released sugars were separated and analyzed isocratically with 99.2% water and 0.8% (v/v) sodium hydroxide (1 mL·min−1) by HPAEC-PAD (ICS 5000 system, Dionex-Thermo-Fischer Scientific, Sunnyvale, CA, USA) in a CarboPac SA10 column. We prepared standard curves of each monosaccharide (apiose, arabinose, fucose, galactose, glucose, mannose, rhamnose, and xylose—Sigma-Aldrich, Steinheim, Germany) using known concentrations to calibrate the instrument.
2.4. Amino Acids Extraction, Derivatization, and Quantification
The total amino acids were extracted with 6 mL of fresh 80% ethanol (v/v) (Synth, São Paulo, Brazil) of 20 mg of dry biomass samples. First, the samples were vortexed for 1 min and dried in a vacuum concentrator at 45 °C until reaching 0.6 mL of solution. The samples were resuspended with deionized water to a complete volume of 2 mL. Samples were centrifuged at 14,000× g at 4 °C and filtered at 0.2 μM (Sartorius Stedim Biotech, Göttingen, Germany). The amino acids were derivatized with o-phthalaldehyde (Sigma-Aldrich, St. Louis, MI, USA) and analyzed by HPLC (Shimadzu, Kyoto, Japan) on a C18 reverse-phase column (5 µm × 4.6 mm × 250 mm, Supelcosil LC-18, Sigma-Aldrich, St. Louis, MI, USA). The amino acids were separated with a 65% methanol with buffer solution (50-mM sodium acetate, 50-mM sodium phosphate, 20-mL·L−1 methanol, and 20-mL·L−1 tetrahydrofuran and adjusted to pH 8.1 with acetic acid) at 20% for the first 32 min, from 20% to 100% between 32 and 71 min and 100% between 71 and 80 min, with a flow rate of 1 mL·min−1 at 40 °C. The quantification was performed by a fluorescence detector (RF-20 A, Shimadzu, Kyoto, Japan) set at 250 nm and 480 nm.
2.5. Carbon, Nitrogen, and Protein Contents
Pulverized dried biomass (proximally 2 mg) was volatilized to CO2 and N2 in an elemental C and N equipment (Carlo Erba 1110, Bremmer Germany) and analyzed by comparing the concentration of standards in mass spectrometry (Finnegan Delta Plus, Thermo-Scientific, Waltham, MA, USA). The carbon and nitrogen concentrations in the biomass were expressed in percentage (%) concerning sugarcane leaves with known C and N concentrations as the standard, with a maximum error of 1 to 2% [26]. For protein quantification, the nitrogen content (N%) was multiplied by 6.25.
2.6. Total Phenolics and Tannins
The tannins and phenols were extracted from 50 mg of biomass with 1.5 mL of 60% ethanol (v/v) for 30 min under constant stirring. The supernatants were recovered by centrifugation (14,000× g) at 20 °C. The samples were vacuum dried and resuspended in 1-mL 80% ethanol. The total phenolics were determined by the Folin–Ciocalteu method using gallic acid as the standard described by Blainski et al. (2013) [27]. The samples were quantified at 720 nm. The tannins were determined by homogenization with bovine serum albumin (1 mg·mL−1), 1% SDS, 5% triethanolamine, 20% isopropanol, and 0.01-M ferric chloride, as described by Hagerman and Butler (1978) [28]. The quantification was performed at 510 nm, with tannic acid as the standard (0–0.8 mg·mL−1). All reagents were from Sigma-Aldrich (St. Louis, MI, USA).
2.7. Data Acquisition from Systematic Review and Bibliometric Analysis
The systematic review selected primary studies regarding a determined matter of unification evidence about what was already studied [22]. A literature search was conducted (articles and the keywords are shown in Box 1) on Scopus to obtain data on which guiding themes were duckweeds and nutrition. These article searches were performed on 31 August 2021. The data in the present article were analyzed and compared with the literature data. The systematic review retrieved 447 publications containing the keywords of the parameter search in the Scopus database (Box 1). After evaluating the recovered works, 85 were accepted for data extraction despite their relevance and the description of the duckweed species or genera studied (Supplemental Figure S1 and Supplemental Table S1). The collected data were plotted in boxplots by genera considering the following parameters: fiber, ash, fat, proteins, amino acids, starch, carbohydrates, minerals, cell wall, and monosaccharides. WebPlotDigitizer v4.2 [29] was used for data displayed only as figures to obtain the numeric data. The data were transformed in percentages to compare the quantity of each variable according to the species. The collected data of the amino acids levels were expressed in two distinct units (% protein and % biomass) due to the data unit found in the systematic review.
Besides the data produced in this work with some of the duckweed species and ecotypes, a complementary analysis including literature data is necessary to improve statistical and mathematical methods that analyze the contribution of different aspects using bibliometrics [30].
The present study examined the literature structure obtaining information about authors, citations, and keywords shared among papers and evaluated the impact of researchers and institutions in the academic world [30,31]. The bibliometrics analysis was applied to study the scientific production to understand the impacts of science on the state-of-the-art technology [32]. Quantitative characteristics were obtained through the bibliometric method, as the number of journal publications by countries and thematic categories for the research area on the duckweed nutrition content. In addition, this bibliometric analyzed the occurrence and distribution of the keyword in the world’s research that contributed to publications about duckweed feedstock.
Box 1. Research parameters for scientific publications used to search in the selected topics: keyword, title, abstract, and all. Source: Scopus (www.scopus.com, accessed on 31 August 2021).
Research Parameter: (TITLE-ABS-KEY (duckweed OR wolffia OR lemnaceae) AND ALL (food OR animal OR feed OR protein OR amino AND acids OR fatty AND acids OR aquatic AND farming OR fiber OR nutrition OR cytotoxicity OR agronomy))
3. Results and Discussion
Duckweeds are the smallest angiosperms with an outstanding growth capacity [10,14]. Their growth is 28 times faster than crops [33], with a biomass accumulation of up to 100-ton dry matter ha−1 year−1 [34]. The biomass production and growth capacity in aquaculture make duckweeds a promising alternative as a food supply, since their land-use effects are much lower than other terrestrial crops. On the other hand, a possible penalty associated with the cultivation of duckweeds as a food supply is that they display variable chemical composition, apparently dependent on the growth conditions and the nutrient availability [35]. The high content of fibers, ash, and fat with concomitant lower levels of proteins are found in duckweeds grown in low nutrient waters. The opposite is true when plants are grown in water containing high concentrations of ammonia and minerals. Under these conditions, the duckweed’s composition becomes high in protein and ash and low in fiber [13]. Therefore, the duckweed composition is crucial to food applications. The increase of duckweed consumption in Asia and the supporting literature regarding using these plants, as animals’ and humans’ feed are needed.
The present work combined the evaluation of 21 ecotypes of duckweeds (grown under laboratory conditions) concerning their fiber, monosaccharide, amino acids, soluble sugars, total phenolic, and tannin contents. The biochemical data were used for an integrative analysis with retrieved data from a systematic literature review focusing on the food supply described above.
3.1. Biochemical Analysis of the 21 Ecotypes
The contents of carbon, nitrogen, proteins, soluble sugars, and cell wall (expressed as fiber percentage) of 21 duckweeds ecotypes are shown in Table 1.
The carbon evaluation can be mostly related to the carbohydrates and the nitrogen to the protein. The carbon content was 37.8% among all genera, with no significant differences; the nitrogen and protein were 4.8 and 29.95%, respectively. Landoltia punctata 7488 had 8.3% more proteins than the other duckweeds evaluated. The total amino acids also distinguish among the species, being higher in Lemna gibba DWC 112 (18.2%) and Wolffiella caudata 9139 (18.6%). The overall average of soluble sugars was 50% of the biomass from the carbon derivatives, and the fibers were 49% (Table 1).
The nonstructural carbohydrates (glucose, fructose, sucrose, raffinose, and starch—Table 2) and structural carbohydrates (fucose, arabinose, galactose, rhamnose, glucose, xylose, mannose, and apiose—Table 3) were quantified. The glucose, fructose, sucrose, and raffinose were found in low proportions compared to starch (Table 2). Among the non-starch sugars was fructose dominates, being usually higher than glucose, sucrose, and raffinose. Wolffioideae displayed approximately twice (106.9 µg·mg−1) of the amount of starch compared to Lemnoideae (48 µg·mg−1). The exceptions are two Lemna species (Lemna minor 8627—97.5 µg·mg−1 and Lemna gibba DWC112—138.5 µg·mg−1) with similar starch contents to Wolffioideae (Table 2).
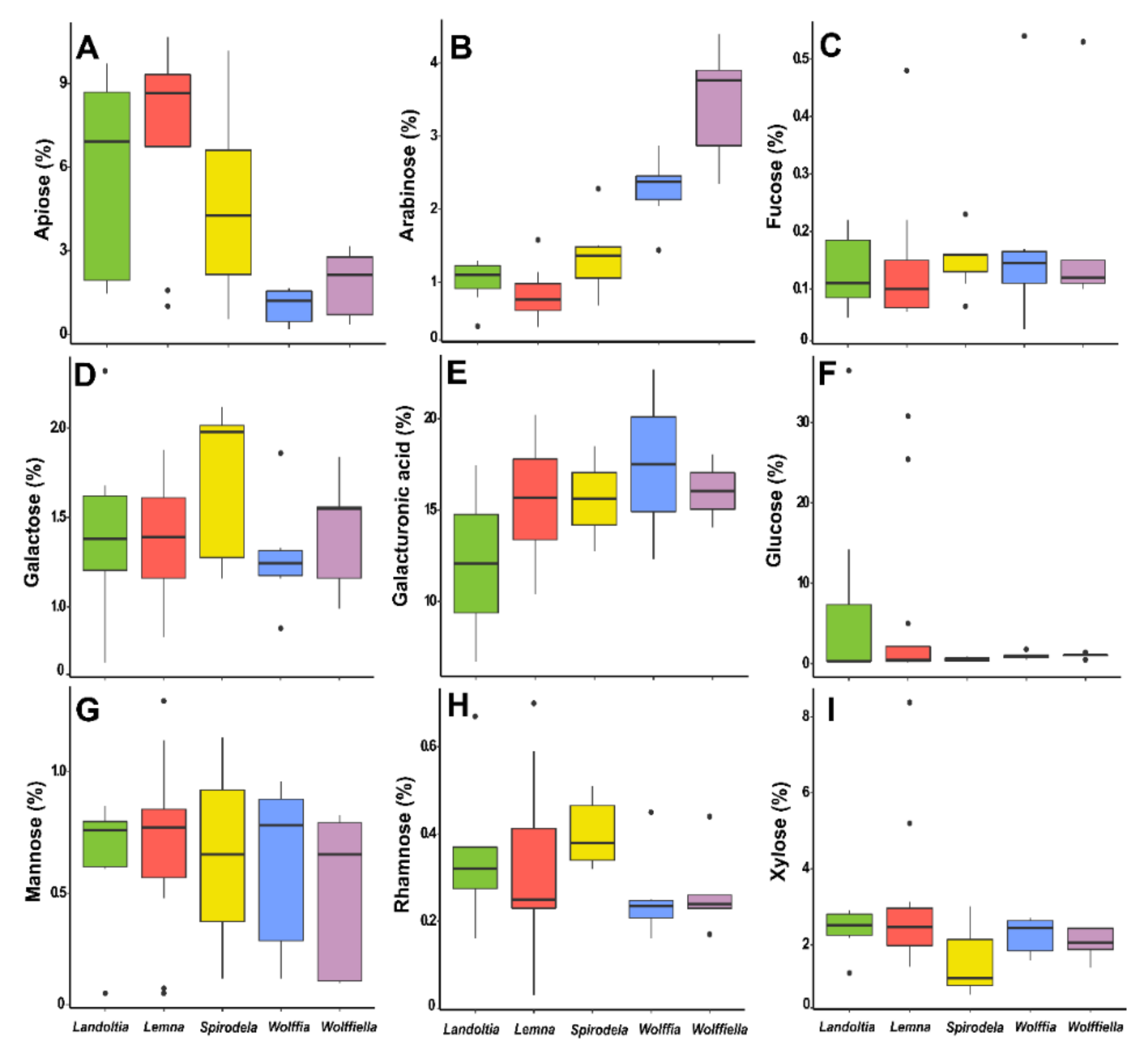
The structural carbohydrates are the constituents of the cell wall. It is well-known that the apiose levels are a trade-off between the sub-families. Here, on average, was detected 80.1 µg·mg−1 for Lemnoideae and 19.6 µg·mg−1 for Wolffioideae (Table 3). Among the genera, Lemna species had a higher apiose level (92.5 µg·mg−1) and Wolffia the lower (14.1 µg·mg−1). Another pentose, xylose, was also higher in the Landoltia (24.7 µg·mg−1) but also greater in Wolffia (23.1 µg·mg−1) and Wolffiella (21.2 µg·mg−1) (Table 3). Spirodela polyrhiza 7498 and 9624 had 2.5 times more xylose than the other Spirodela lineages (Table 3). Wolffia globosa 884 had 1.48 times less xylose than the other Wolffia species. Other pectin sugars, such as rhamnose and galactose, were different across the genera. Rhamnose was slightly higher in Spirodela (4.3 µg·mg−1) than in Landoltia, Lemna, Wolffiella, and Wolffia (3 µg·mg−1), except for Wolffiella neotropica 8848 (4.5 µg·mg−1) and Wolffia globosa 884 (4.6 µg·mg−1). Galactose was slightly higher in Spirodela (18.6 µg·mg−1) and Wolffiella (16.5 µg·mg−1). Spirodela polyrhiza 9346 had approximately half of the galactose compared to the other Spirodela species evaluated. Wolffia globosa 884 (18.6 µg·mg−1), Wolffiella gladiata 8261 (18.4 µg·mg−1), and Lemna gibba DWC112 (18.0 µg·mg−1) had more galactose than the average of the evaluated species. The arabinose levels had higher variability between the species of the Spirodela (ranging from 6.9 to 22.9 µg·mg−1) and Wolffiella (ranging from 23 to 44 µg·mg−1) genera. The glucose levels ranged from 2.2–12.2 µg·mg−1 across all genera, higher in Lemna gibba DWC112 and Wolffioideae (Table 3). All species had a considerable amount of mannose with 8.2 µg·mg−1, a sugar constituent of the hemicellulose mannans. Furthermore, the duckweeds displayed low fucosylation (see the fucose levels in Table 3—1.6 µg·mg−1) similarly to sweet corn (0.2 µg·mg−1), pumpkin (0.6 µg·mg−1), oats (0.2 µg·mg−1), green kiwifruit (1.4 µg·mg−1), golden kiwifruit (1.1 µg·mg−1), carrot (1 µg·mg−1), and blackcurrant (1.2 µg·mg−1) and differently from apple (3.6 µg·mg−1) [36].
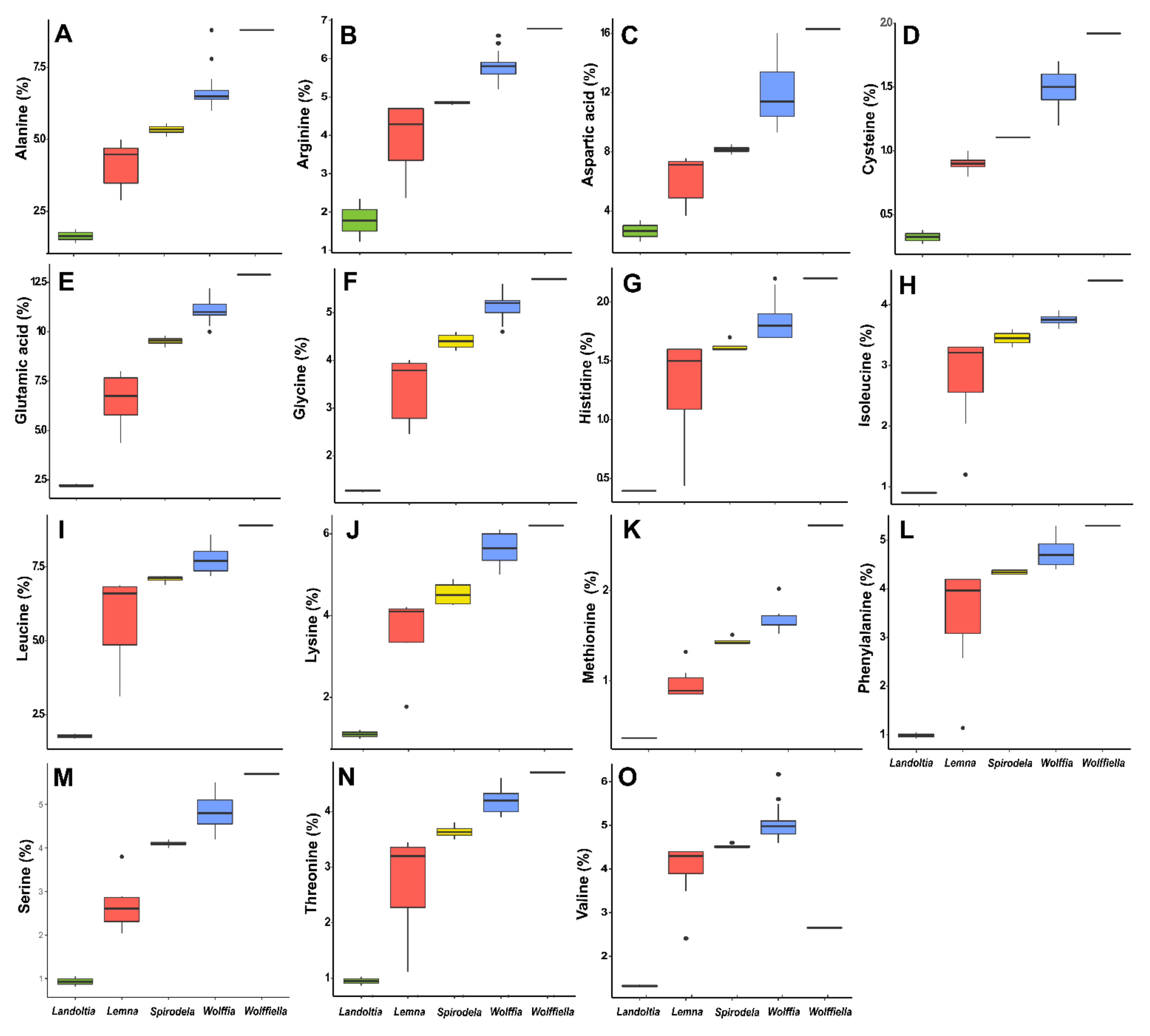
The total amino acids were higher in the Wolffia species (124 µg·mg−1 dry weight). Spirodela species have a reduction of 23%, Lemna 38%, Landoltia 48%, and Wolffiella 54% in comparison to Wolffia (Table 1). Thus, the quantity and diversity of duckweeds amino acids are related to the species’ ecotype. The amino acids evaluated were divided into essential (Table 4) and non-essential (Table 5) to facilitate the interpretation from a food point of view. The human body does not synthesize the essential amino acids, and their supply must come from food intake. Besides alanine and lysine, all the essential amino acids were found in low levels (0–2000 mg·mg−1 dry weight), while the nonessential amino acids displayed higher concentrations than the essential ones. The evaluated species have more than 70% as aspartic acid (Asp), glutamic acid (Glu), asparagine (Asn), 2% citrulline (Cit), 5% threonine (Thr), 6% alanine + tyrosine (Ala + Tyr), and 2% serine (Ser), and the remaining 15% are distributed into histidine (His), tryptophan (Trp), methionine (Met), valine (Val), isoleucine (Ile), leucine (Leu), lysine (Lys), serine (Ser), glutamine (Gln), Gamma-Aminobutyric Acid (GABA), and ornithine (Orn). Landoltia had proportionally higher amounts of arginine and asparagine. Wolffiella had more citrulline, tryptophan, methionine, valine, and phenylalanine, and Wolffia had more glycine and arginine (Table 4 and Table 5).
Tannins are secondary metabolites in plants that confer stringency and bitterness flavors to food [37]. In most cases, food containing tannins can decrease the quality of a particular food product [37]. In the 21 samples evaluated, along with the data found in the literature of four occurrences, Spirodela displayed lower levels of tannins in all genera (20 mg·mg−1) (Table 6), which represents less than 1% in the plant biomass. However, the phenolic content was distinct among the genera, with Spirodela = Lemna = Wolffia > Landoltia = Wolffiella. The low content of tannins in Spirodela might improve the organoleptic characteristics of this duckweed species due to the non-bitter flavor. This may be a critical factor in the acceptance of Spirodela as a food product.
3.2. Systematic Review and Bibliometric Analysis
One way to recover data from the literature is through a systematic review. Our revision of 85 studies aimed to evaluate the nutrients of different duckweed genera (Supplemental Figure S1). From the selected papers, 80 were found in the Scopus search, and five were added manually to analyze the cell walls. The genus Lemna had the higher frequency data on the publications (206), followed by Wolffia (165), Spirodela (117), Landoltia (47), and Wolffiella (34) (Table 7). In addition, the data of the amino acids (72), ash (48), carbohydrates (8), cell wall (13), energy (8), fat (81), fibers (58), minerals (24), protein (114), monosaccharides (37), starch (82), and tannin (24) were collected and evaluated.
A comprehensive systematic analysis of duckweed composition relevant for nutrition and food assessment with the unity of biochemical data to a bibliometric analysis enlightens the duckweed applications toward food. The systematic review includes using rigorous methodology according to current guidelines of conducting and reporting [22]. The bibliometric analysis showed the distribution of the countries that studied duckweed regarding the relevance for food applications. China is ranked in the first position in numbers of publications (47 papers), followed by the United States (19 papers) and India (18 papers) (Figure 1A). Asian countries were the significant contributors of scientific knowledge and are the largest duckweeds producers. The panel of author keywords demonstrates that most of the papers used here display words related to the fast growth of duckweeds, and the content of proteins, amino acids, and starch is the most recurrent and significant of human and animal feed (Figure 1B). The keyword panel corroborates the frequency of the recovered data, being the genus Lemna the most abundant variable in proteins (Table 7).
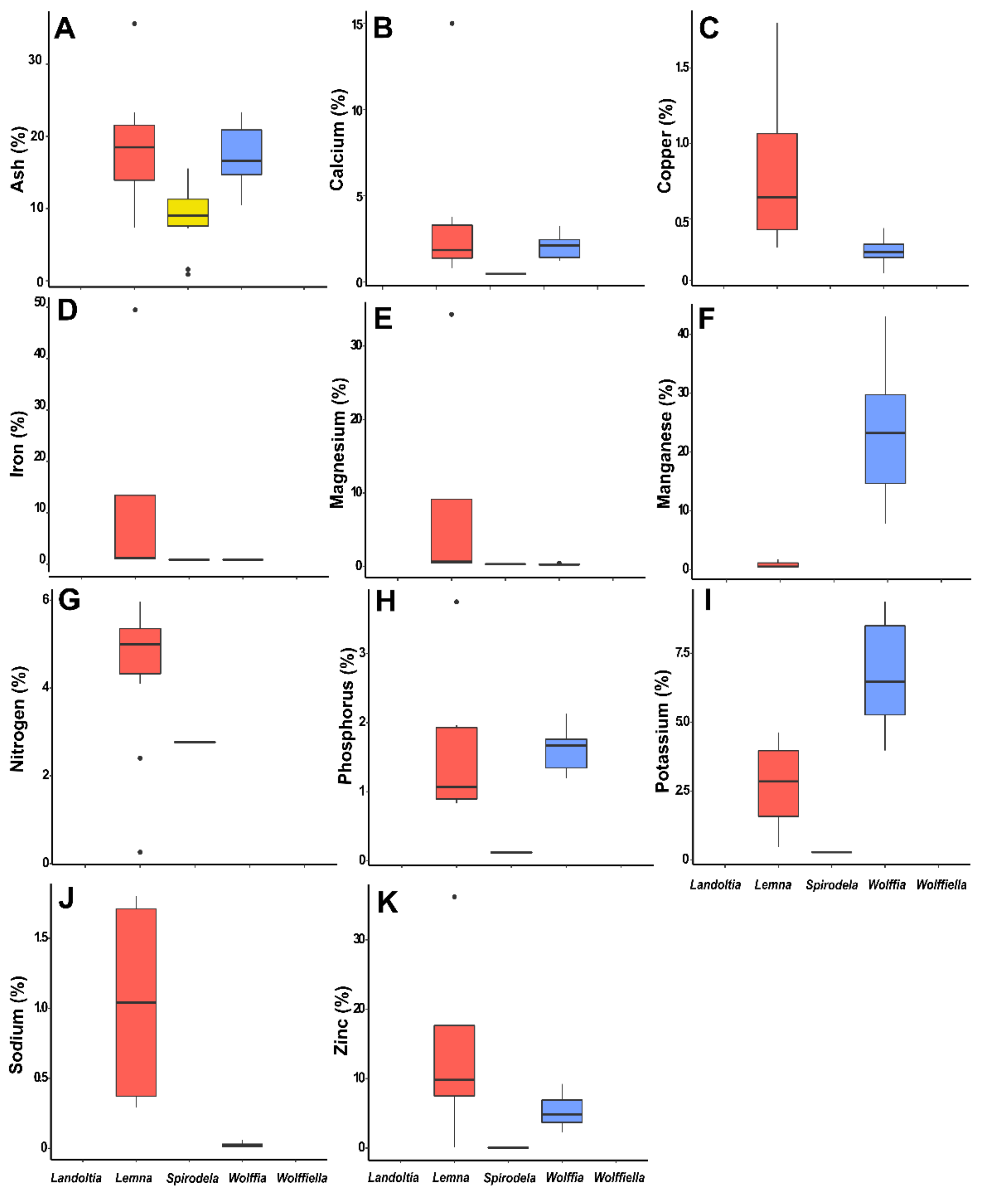
Fresh duckweed’s biomass is 92–94% water [35], and the ash content is considered high compared to other plants. The ash content of plant tissues is mostly inorganics and is species-dependent [38]. The ash data were collected from 20 publications with 48 occurrences, of which 12 were from Spirodela, 15 from Lemna, and 21 from Wolffia (Table 7). The ash content of the Spirodela biomass ranges from 1 to 16%, Lemna ranges from 7 to 36%, and Wolffia from 10 to 23% (Figure 2). The ash content is widely ranged in plants, being 0.1% for debarked woody chips, 3–12% for corn stover, 1–9% for Miscanthus straw, 8–26% rice straw, 1–15% sugarcane bagasse, 3–23% wheat straw, and 0.2–6% for woody residues [39]. Interestingly, the ash content of duckweeds is not influenced by the nutrient disponibility into the water body [35].
Duckweeds can accumulate minerals (macro and micronutrients) several hundred folds compared to the concentration of minerals in the water they grow in [40,41]. As a result, these plants have the capacity for phytoremediation [17]. The nutrients in duckweeds, obtained from 13 publications were calcium, copper, iron, potassium, magnesium, manganese, sodium, phosphorus, and zinc. Appenroth et al. (2018) evaluated the nutritional value of 11 species of the 16 ecotypes belonging to the genus Wolffia [17]. The mineral composition of Wolffia was rich in potassium and iron and poor in sodium, manganese, zinc, and copper (Figure 2). Spirodela polyrhiza has a low content of minerals when compared to Lemna and Wolffia, especially for zinc (0.5 mg·g−1), phosphorus (1.3 mg·g−1), and potassium (2.8 mg·g−1). Lemna has a distinct nutrient pattern, with elevated levels of iron, magnesium, zinc, copper, and sodium (Figure 2).
Plants are a rich source of minerals. Therefore, they can benefit the human diet and contain antinutrients that require processing before consumption [1]. Healthy diets contain macro- and micronutrients needed by the organism, but some contain anti-nutrients like phytates and oxalates that inhibit iron and zinc absorption [1]. The oxalate and phenolic compounds also are considered organoleptic inhibitory factors and interfere with digestion or metabolism [42]. Druse and raphide shapes of calcium oxalates are reported in Spirodela polyrhiza [43] and Lemna minor [44] which play a role in calcium regulation of these plants. Nevertheless, the Wolffioideae plants have oxalate in the free form favoring them for food consumption [17].
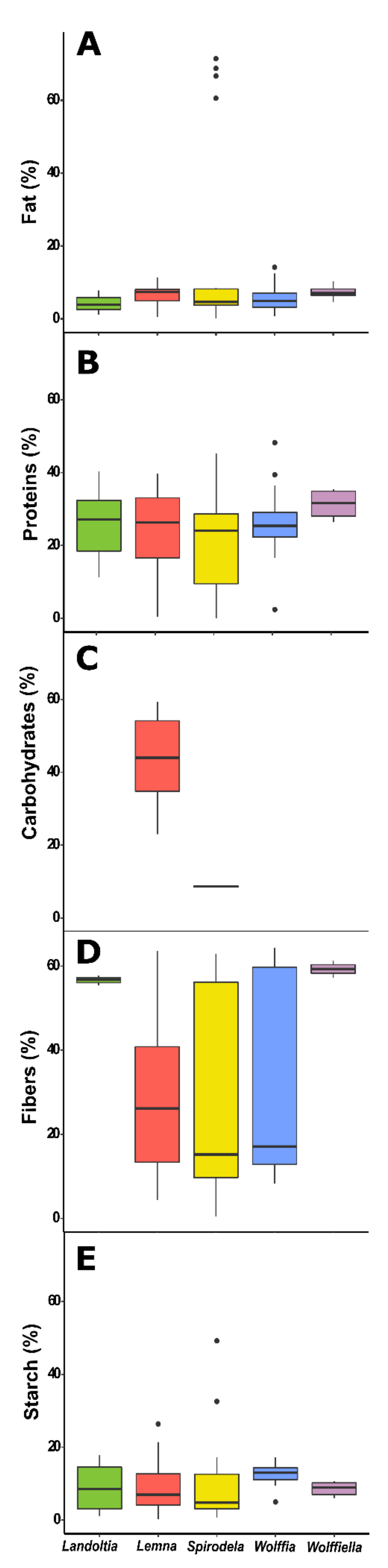
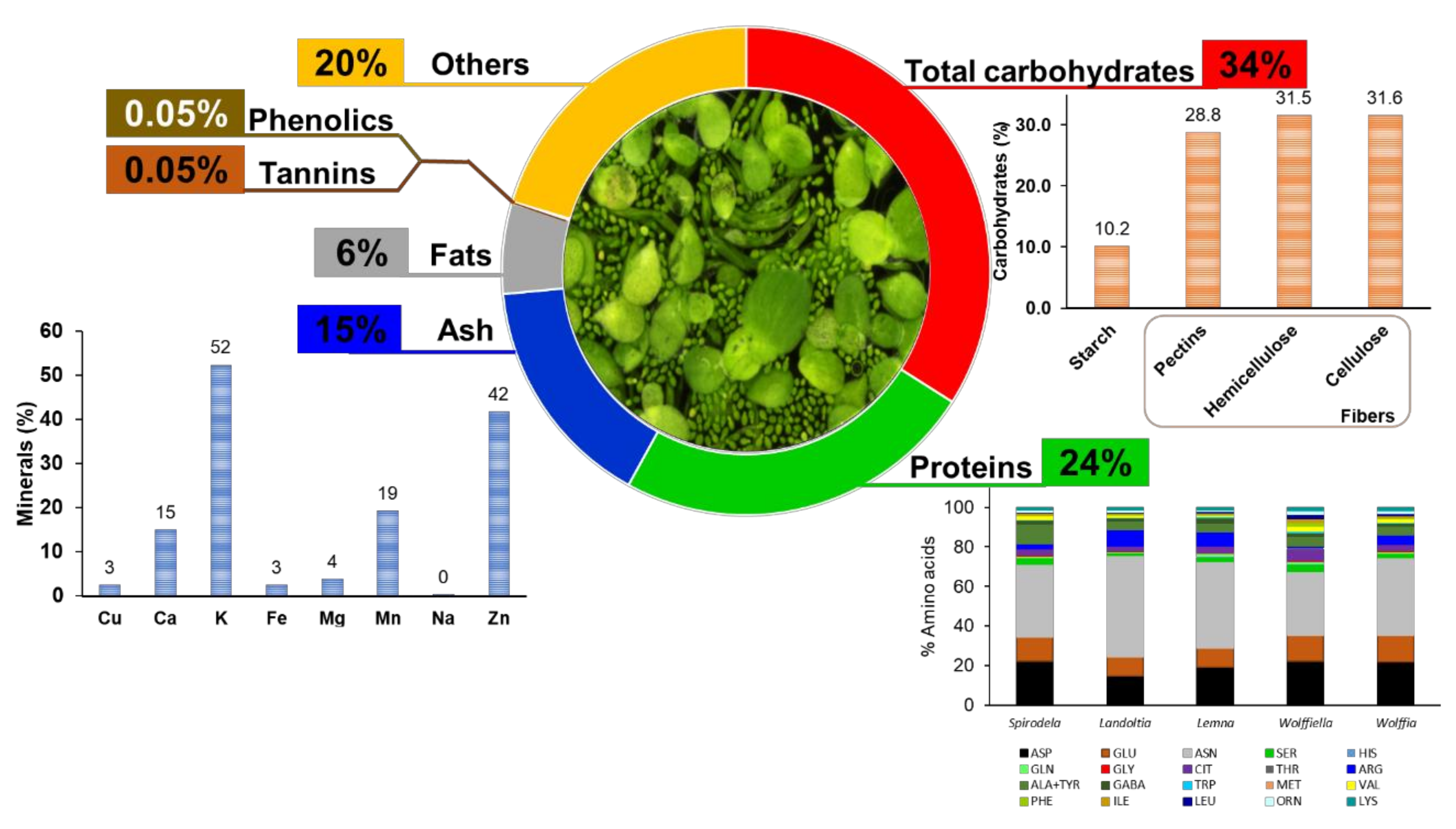
Proteins are macronutrients essential for the human diet once it is a source of amino acids. The protein content is variable among food sources, being 20–36% legumes, 8–25% nuts and seeds, 10–20% meat and fish, 15% eggs, 3.5% milk, and 1–3% vegetables [45]. Nowadays, a plant-based trend in diets is growing worldwide. Plant-based protein provides 65% of the world’s supply [45] and has the required amino acids for human intake [46]. Duckweed protein is considered high quality [19,47], which can substitute some protein intakes. The protein data was the most frequently found (43 papers with 114 occurrences) (Table 7). Variability among the duckweed genus was found, in which Spirodela was higher, ranging from 0.5 to 45% and Wolffia from 2 to 48%, followed by Landoltia (11 to 40%), Lemna (0.3–33%), and Wolffiella from 26 to 35% (Figure 3B). The protein content of duckweeds is related to the plants’ nutrition disponibility, temperature, and age [19]. The content of plants grown in natural water bodies varies from 7 to 20%, while the plants grown in mineral media or effluents have a high protein content ranging from 30 to 40% [19], which corroborates several studies.
A balanced diet requires daily protein to compensate for the nitrogen loss and provide protein for metabolism maintenance and growth [46], equivalent to 10–18% of the total energy [45]. Duckweed protein is highly digestible, which is important for diets due to the reuse of the amino acids ingested. The digestion was studied in fish [18,48,49,50,51,52,53,54,55], pigs [56], dogs [57], and turtles [58,59], with a range from 65 to 90%. The digestibility of Lemna minor was evaluated in comparison to green peas, revealing a lower capacity, but Lemna minor also led to lower plasma glucose and insulin levels in humans [60]. Thus, duckweeds can be a promising alternative for diabetics, reducing blood sugar and benefiting health.
The amino acids can be divided into essential (not produced by the human organism) and nonessentials (produced by the human organism). Furthermore, the essential amino acids are not fully available depending on the plant source, like cereals with low lysine and legumes with low sulfur-containing amino acids [46]. The amino acids lysine and arginine are essential for animal feed (~6% of the dietary protein), and the levels of these in duckweeds (up to 6%—Figure 4) are comparable to those found in alfalfa [40,61]. Arginine is conditionally essential during pregnancy, critical illness, and infancy [62], being necessary for human intake. The amino acid recovered data were analyzed as a percentage of the protein and biomass due to the nature of the data (Figure 4 and Supplemental Figure S2). The amino acids contents of the 21 ecotypes evaluated here had 72% of asparagine, glutamic acid, and aspartic acid (Table 5), and asparagine and glutamic acid corresponded to 2% of the biomass (Supplemental Figure S2). These amino acids play an essential role in protein synthesis and modification, the breakdown of ammonia, and the neurological system. Duckweed’s amino acid composition is similar to that found in other plants [15,19,63]. They are rich in leucine, threonine, valine, isoleucine, and phenylalanine and poor in methionine and tyrosine [40] (Figure 4 and Supplemental Figure S2). The cysteine + methionine was 22% higher than the WHO recommendation, and the levels of threonine, phenylalanine + tyrosine, and leucine also surpassed the intake recommendations [15]. Porath et al. (1979) found methionine and lysine levels comparable to milk and cheese in seven duckweeds species (Lemna gibba, Lemna minor, Spirodela polyrhiza, and Wolffia arrhiza) [47]. However, methionine was found in lower concentrations (100 mg·mg−1 in the 21 ecotypes evaluated biochemically and 0.3–2.7% of protein) than the recommended (Table 4 and Figure 4). In general, for duckweeds, protein is a good source of lysine (2% of dry matter). Besides methionine, the amino acids isoleucine, leucine, phenylalanine, threonine, valine, and tryptophan correspond to the FAO reference pattern, fulfilling the requirement for human nutrition [17], thus being an effective supplement in diets [19] (Figure 4 and Supplemental Figure S2).
Energy balance is essential to maintain health through a secure nutrient intake [20]. The metabolism of fatty acids generates twice the energy of the same amount of carbohydrate [20]. However, low amounts of fats are recommended in the diet. Plants have a distinct nutritional composition, and vegetables have low-fat contents. The fat content in Spirodela varies from 0.2 to 8%, Landoltia 1 to 8%, Lemna 0.5 to 11%, Wolffiela 6 to 10%, and Wolffia 0.7 to 14% (Figure 3A). This data were recovered in 21 papers with the lipid data from 1980 to the present, with 81 publications (Table 7). A wide range of fat contents was observed in Spirodela´s literature (0.2–71%) due to the evaluation of the cuticles of four Spirodela polyrhiza ecotypes by Borisjuk et al. (2018) [64]. The variance in the fat content of duckweed is related to the growing system, climatic conditions, light intensity, and nutrient availability [40,55]. A fatty acid survey of thirty duckweeds species was performed by Yan et al. (2013) [65]. The authors found that the total content varied between 5 and 14%, and the composition of the fats showed low variability with 80% palmitic acid (C16:0), linoleic acid (C18:2), and α-linolenic acid (C18:3) [65], which corroborates the one found in plants [15,17,40]. The main fatty acids from duckweeds (linoleic acid—C18:2 and α-linolenic acid—C18:3) are essential due to their role in retinal and brain development and the prevention of cardiovascular diseases [20]. Thus, the intake of food with these fatty acids is indispensable for human health during all life. The recommended intake level is 2.5% of the total energy of linoleic acid and 0.5% of the total energy of α-linolenic acid [20].
Saturated fatty food has greater relevance for human nutrition. In duckweeds, the high content of palmitic acid should be a hazard, but the levels of oleic acid, short-chain fatty acids, and medium-chain fatty acids are remarkably low [20]. The high omega-6 and omega-3 ratios are involved in the development of cardiovascular diseases, cancer, osteoporosis, and inflammatory and autoimmune diseases [17]. Therefore, it is recommended to consume food with a ratio between the two lipids below 5, which is not natural for some plants and additives [20]. Furthermore, the omega-6 and omega-3 ratios were 0.36 among the duckweed species that Yan et al. (2013) evaluated. Thus, adding duckweed to the human diet would be interesting to reduce the unfavorable omega-6 and omega-3 ratio [65].
Duckweeds display low-fat contents. Therefore, the energy of the intake of these plants comes from carbohydrates. Carbohydrates are the most abundant components of food, being polyhydroxy aldehydes; ketones; alcohols; acids; and their derivatives that are classified as sugars (glucose, galactose, fructose, sucrose, lactose, trehalose, sorbitol, and mannitol); oligosaccharides (maltodextrins, raffinose, stachyose, and fructooligosaccharides); and polysaccharides (starch, cellulose, hemicellulose, pectins, and hydrocolloids) [66]. The carbohydrate content of duckweeds is mainly fibers instead of starch, like in soy grains [52]. The total carbohydrate content in duckweeds was determined in eight publications with eight occurrences between Spirodela and Lemna (Table 7). The content in Lemna varies from 23 to 59%, and Spirodela contains 8% carbohydrates (Figure 3C). The highest report from Lemna minor (59%) was calculated based upon the difference of a total content with the mineral content, protein, and lipids [67]. A similar report was made by Zhao et al. (2014) with 51.2%, although the carbohydrate content seems to include the cell wall content [68].
The primary energy source in human diets is carbohydrates, which constitutes sugars, starch, and other polysaccharides [69]. Starch’s easy digestibility and the almost total absorbent capacity in the human trait make it a fast-energy assessment in the diet. Maize, rice, sorghum, wheat, barley, millet, rye, oats, potato, cassava, yam, and taro have 17–75% of starch and are the most consumed cereals and root crops [70]. Therefore, starchy plants are value-added for the human diet as a source of nutrients and energy and in several industrial applications like thickener, texturizer, gelling agent, paper and board production, biodegradable plastics, and packaging materials [71]. Due to the remarkable growth capacity, duckweed polysaccharides have importance for biorefinery applications. Starch in duckweeds can accumulate up to 70% into fronds [72] and have distinct morphological, crystallinity, physicochemical, and thermal properties [73]. The starch from these plants has a low peak of viscosity that is suitable for high-temperature processes and as materials for delivery systems [73]. The systematic review resulted in 22 papers from 1997 to 2021, with 82 occurrences among the five genera (Table 7). Spirodela had the higher variation of starch content of the evaluated works with a range of 0.7–49%, while the other genera varied as follows: Landoltia (1–17%), Lemna (0.2%–10%), Wolffiella (6–11%), and Wolffia (5–14%) of the biomass (Figure 3E). The growth system and conditions are essential for duckweeds biomass and starch production. Thus, the amplitude of the data reported in the literature must reflect this bias due to daytime and the light intensity that prevailed during the plant´s growth. Other factors might also have influenced the variety of the results of starch accumulation in duckweeds, such as temperature, light intensity, photoperiod, co-cultivation, and disponibility of nutrients [72,74,75,76,77]. Duckweeds starch properties and granules morphology and the capacity for high starch accumulation make these plants suitable for diverse applications in the food industry and as “starchy vegetables”, especially Spirodela, Landoltia, and Lemna species due to the small sizes of the granules [73].
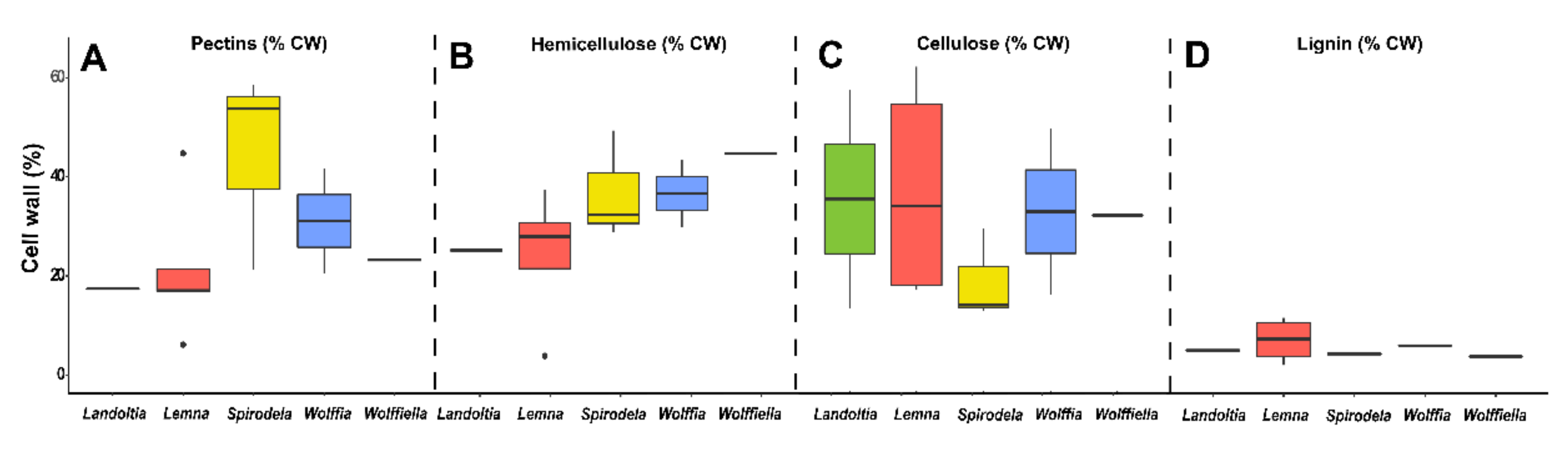
It is well-known that a diet shift of high fat and high sugars is needed to improve human health. The replacement of high-fat meats with beans, peas, lentils, and other fibers is a public health concern [70]. The diet fiber comprehends editable carbohydrates polymers mainly of plant origin with ten or more monomeric units that are not digested by endogenous enzymes and not absorbed by the small intestine of humans [78]. The diet fiber includes cellulose, hemicellulose, pectin, gums, mucilages, and lignin [79]. Thus, it is possible to consider food fiber as the plant cell walls. The latter is a complex structure of polysaccharides, phenolic compounds, and structural proteins arranged in a glucose core (cellulose) with hemicellulose, lignin, and structural proteins immersed into a pectin matrix [80]. Lignin is the phenolic compound responsible for polysaccharides binding, which is found in low proportion in duckweeds [68,81,82,83], assuming 5–10% of the cell wall (Figure 5D). Cell wall proteins are quantitatively irrelevant.
The proportion of cellulose in duckweeds (13–17%) is thought to be three times smaller than inland plants [84,85], although some publications reported ~40% of cellulose into the cell walls [68,83] (Figure 5C). This discrepancy might be related to the methodology employed during the cell wall fractionation process (Figure 5C). Cellulose can be used as a delivery system, food stabilizer, functional food ingredient (due to the low or indigestibility), and biodegradable packaging [86]. Hemicelluloses are composed of the monosaccharide’s glucose, arabinose, xylose, fucose, galactose, and mannose to form different classes of polysaccharides (e.g., xyloglucans, xylans, mannans, and β-glucans), assuming up to 29% of the duckweed cell wall (Figure 5B). Moreover, cellulose can be employed as a green film for food packing [87]. Mannans (polysaccharides made of mannose) are found in low proportions in most of the duckweeds (0.1–1% of dry mass), except for Lemna perpusilla, which can reach 13% [85] (Figure 6G). These polymers display several applications in the food industry as edible films, gel formation, stiffeners, viscosity, modifiers, stabilizers, texture improvers, water-absorbent, prebiotics in dairy products, and seasonings [88].
Pectins are acid polysaccharides whose backbones contain 70% of their structure as galacturonic acids interspaced with rhamnose. These are substituted with neutral polymers composed of fucose, galactose, apiose, xylose, and arabinose. The polymers formed are homogalacturonans, rhamnogalacturonans I and II, xylogalacturonans, and apiogalacturonans [89]. Pectins in the primary cell wall of dicots and non-graminaceous monocots represent, on average, 35%, while only 2–10% of grasses and commelinids and 5% of woody tissues [89]. Duckweeds are pectin-rich plants (20–54%) (Figure 5 and Figure 6) [57,68,83,84,85,90,91] with an evolution trend with high proportions of apiogalacturonans (see the apiose levels—Table 3 and Figure 6A) in Lemnoideae and xylogalacturonans (see the xylose levels—Table 3 and Figure 6I) in Wolffioideae [92].
Apiose is higher in Spirodela (0.5–10%), Landoltia (1.4–10%), and Lemna (1–11%) than in Wolffiella (0.3–3%), and Wolffia (0.1–2%) (Figure 6A). The xylose content in the whole cell wall is computed together with the residues of xylans, xyloglucans, and xylogalacturonans. Thus, the pattern of higher xylose contents in the Wolffioideae was not identified (Figure 6I). Spirodela had 0.7–3%, Landoltia 1–3%, Lemna 1.5–14%, Wolffiella 1–2%, and Wolffia 1.6–3% of xylose into the cell walls (Figure 6I). As previously mentioned, duckweeds have elevated pectins levels, so the concentration of galacturonic acid/uronic acids in this class of polysaccharides is 13–20% in Spirodela, 7–17% in Landoltia, 10–20% in Lemna, 14–18% in Wolffiella, and 12–23% in Wolffia (Figure 6E). The chemical properties of pectins make them suitable as a gelling agent, emulsifier, thickener, stabilizer, edible films or coating, and fat or sugar replacer in low-caloric food [93,94]. Such properties regarding the duckweed pectins remain to be studied.
A diet rich in fibers has several advantages, such as a reduction of appetite, lower blood sugar levels, reduced risk of heart disease, alleviating constipation, reduced risk of colorectal cancers, reducing the risk and symptoms of metabolic syndrome and diabetes, lowering cholesterol, balances intestinal pH, and stimulates intestinal fermentation [79]. The fiber levels were identified in 20 works with 58 occurrences, 15 of Spirodela, 3 of Landoltia, 27 of Lemna, 3 of Wolffiella, and 10 of Wolffia, ranging from 0.4 to 63%, 5 to 58%, 4 to 67%, 57 to 67%, and 8 to 64% respectively (Table 7 and Figure 3D). Thus, eating vegetables rich in the wall, like duckweeds, could help the fiber functions mentioned above and be advantageous for human health.
4. Advantages of Duckweeds as Food and Perspectives
Duckweeds display a remarkable capacity for growth and adaptation that can be useful as a food feedstock almost anywhere globally. These plants’ growth rate, composition, and macronutrients are strongly associated with nutrient availability, pH, light, and temperature [42]. Thus, the growth system must be considered when the purpose is to take advantage of the nutritional characteristics of duckweeds. The low dry matter of duckweeds has significant protein, fibers, and carbohydrates contents. Therefore, the nutritional profile is quite suitable, except for the tryptophan and methionine levels, requiring supplementation when duckweed is consumed. Besides that, the low content of lignin, total phenols, and tannins collaborate with human consumption due to the non-astringent and non-bitter flavor. Additionally, duckweeds do not have allergens and potentially benefit cardiometabolic states and reduce cardiovascular risk [95]. The whole food is converted into energy to maintain the body’s metabolism. Only seven works reporting the energy capacity on Spirodela and Lemna were found in the systematic review (Table 7). The energy/calorific value of duckweed was 1–19 KJ·g−1. This value is related to the photosynthetic capacity of the plant and its nutritional status [96]. Besides, the plant type, organs, and position in the forest influence the energy value of the plant [96]. The energy budget is a relationship between the food’s energy intake with the energy lost in feces and excreta, together with the energy deposited in growth [97]. The higher digestibility of duckweeds (65–90%) is directly related to the energy gain and the metabolic enhancement when consumed. In a study of carp feed with Lemna minor and Spirodela polyrhiza, carps’ growth and energy were lower compared to animal protein (tubeficid worm—Limnodulus hoffmeisteri), indicating a supplementation need [98].
A safety hazard is an important matter for human consumption, mainly the presence of heavy metals, phenols, pesticides, dioxins, and pathogens need to be considered [99]. Care is needed for heavy metals once duckweeds have clean-up water capacity with contaminants accumulation into their tissues [100], and if it enters the food chain, it may be transformed into food risk. Moreover, duckweed can uptake and transform phenols and pesticides, and also, dioxins were found in duckweed harvest due to the contamination with animal proteins from beetles and snakes [99]. Therefore, duckweeds as food consumption need monitoring during production and cannot be associated with clean-up systems [99]. The only known compound toxic to animals and humans in duckweeds is calcium oxalate. Calcium oxalate in high levels may cause kidney stones [101], hematuria, renal failure, and gastric hemorrhage [102]. Duckweed subfamilies accumulate calcium oxalate in different forms, in which crystals are not found in Wolffioideae. Thus, this subfamily has advantages for direct food intake due to the lack of processing required. Oxalate in plants is 50–75% water-soluble, leaching out during cooking [102]. Therefore, Spirodela, Landoltia, and Lemna intake are also suitable when cooked. Another safety concern is the cytotoxic effect and antiproliferative effects in human cells. Sree et al. (2019) tested seven duckweed species (Spirodela polyrhiza, Landoltia punctata, Lemna gibba, Lemna minor, Wolffiella hyalina, Wolffia globosaluence, and Wolffia microscopica) whole plant extracts and detected the absence of cytotoxic and antiproliferative effects in human cell lines [103].
Despite the several advantages of duckweeds, it is important to point out that the upscaling duckweed growth for human food intake needs to be controlled. The clean-water capacity cannot be associated with the food cultivation to avoid contamination with heavy metals, dioxins, phenols, and other chemicals with potential health hazards that can be stored into duckweeds tissues. Therefore, the cultivation for human consumption needs to be in clean water with farming control. Semi-indoor cultivation is possible nowadays in greenhouses and polytunnels, as used by the companies Hinoman and Rubisco, but improvements are needed [104]. Coughlan et al. (2021) reviewed the cultivation of duckweeds in bioreactors for large scale production, as in human food consumption. They stated six key challenges to surpass for indoor cultivation, which are a “design of cultivation structures (1), determination of operational conditions (2), choice of medium type (3), selection of the duckweed species and lineage (4), manipulation of the microbiome (5), development of harvesting technologies (6)” [104]. The cultivation structures can be addressed as batch (stationary), continuous (by flow-through), lagoon-style, and stashed systems [104]. Moreover, all of them can be optimized with new multifaced technologies that are realistic for several purposes that boost the circular economy [104]. Besides, vertical hydroponic farming and house cultivation systems are possible for duckweeds. Thus, large-scale duckweed production for human consumption is possible but still needs improvement. Wolffioideae species have an advantage over Lemnoideae concerning the calcium oxalate form, which is dismissal for cooking [102]. Furthermore, selection of the growth media is essential for duckweed’s chemical composition of sugars, minerals, proteins, and amino acids.
The overall nutrition composition of duckweeds is adequate for human intake as a plant-based source for proteins, starch, and fibers. Furthermore, even with a considerable content of sugars (~50%), these, compared to green peas, have a lower incidence of blood sugar [60], being an ally in diets to reduce diabetes levels. Additionally, duckweeds can accumulate essential minerals such as iron, phosphorus, and nitrogen, indispensable for human development. Furthermore, the fatty acid content has low variability among the genera and species with considerable amounts of C16:0, C18:2, and C18-3 and low incidences of short and medium-chain fatty acids that are undesirable for humans health. The amino acids composition is similar to those found in plants, being mainly asparagine, glutamic acid, and aspartic acid (70%) and also rich in leucine, threonine, valine, isoleucine, and phenylalanine but poor in methionine and tyrosine [40] (Figure 7). Thus, duckweeds are a promising feedstock that could help feed the world with the most required nutrients without the penalty of land use, which is one of the main issues related to global climate change.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy12040796/s1: Figure S1: Systematic reviews (PRISMA) flowchart with literature search and study selection. Figure S2: Amino acid levels in duckweeds by the percentage of the biomass. Supplemental Table S1: Retrieved works from the systematic review.
Author Contributions
M.S.B., D.P., A.G. and J.S.F. conceptualized the study; D.P., J.S.F. and P.C. performed the methodology; D.P., J.S.F. and A.G. performed the data analysis; and D.P., A.G., J.S.F., P.C., E.I.S.F. and M.S.B. were responsible for writing—original draft preparation and reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Instituto Nacional de Ciência e Tecnologia do Bioetanol—INCT do Bioetanol, grant numbers FAPESP 2014/50884-5 and CNPq 465319/2014-9 and Centro de Pesquisa e Inovação de Gases de Efeito Estufa—RCG2I (FAPESP/Shell 2020/15230-5). D.P. (CAPES 88882.377113/2019-1). A.G. (FAPESP 2019/13936-0). J.S.F. (CNPq 380198/2021-5).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data and scripts generated and/or analyzed during this study are available from the corresponding author on request.
Acknowledgments
We thank Amanda Macedo for the analyses of the amino acid in HPLC and Eglee S. Igarashi for analyzing the sugar in the DIONEX system. We also thank the Programa de Pós-Graduação em Interunidades em Biotecnologia from USP (São Paulo) for the finantial support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the study’s design, in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- HLPE. High Level Panel of Experts. Nutrition and food systems. Comm. World Food Secur. 2017, 44, 150. [Google Scholar]
- Drewnowski, A.; Finley, J.; Hess, J.M.; Ingram, J.; Miller, G.; Peters, C. Toward Healthy Diets from Sustainable Food Systems. Curr. Dev. Nutr. 2020, 4, nzaa083. [Google Scholar] [CrossRef]
- Herrero, M.; Thornton, P.K. Livestock and global change: Emerging issues for sustainable food systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20878–20881. [Google Scholar] [CrossRef] [Green Version]
- Porter, J.R.; Xie, L.; Challinor, A.J.; Cochrane, K.; Howden, S.M.; Iqbal, M.M.; Lobell, D.B.; Travasso, M.I.; Aggarwal, P.; Hakala, K.; et al. Food security and food production systems. In Climate Change 2014—Impacts, Adaptation and Vulnerability: Part A: Global and Sectoral Aspects; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2015; pp. 485–534. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, B.D.; Herren, H.R.; Wakhungu, J.; Watson, R.T. International Assessment of Agricultural Science and Technology for Development. In Agriculture at A Crossroads: International Assessment of Agricultural Knowledge; Island Press: Washington, DC, USA, 2009; ISBN 9781597265508. [Google Scholar]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development; UN General Assembly, A/RES/70/1; United Nations: New York, NY, USA, 2015; Volume 1833, pp. 271–287. [Google Scholar] [CrossRef]
- HLPE. Impacts of COVID-19 on food security and nutrition: Developing effective policy responses to address the hunger and malnutrition pandemic. HLPE Issues Pap. 2020, 1, 1–24. [Google Scholar] [CrossRef]
- Bush, S.R.; Belton, B.; Little, D.C.; Islam, M.S. Emerging trends in aquaculture value chain research. Aquaculture 2019, 498, 428–434. [Google Scholar] [CrossRef] [Green Version]
- Kutschera, U.; Niklas, K.J. Darwin-Wallace Demons: Survival of the fastest in populations of duckweeds and the evolutionary history of an enigmatic group of angiosperms. Plant Biol. 2014, 17, 24–32. [Google Scholar] [CrossRef]
- Landolt, E. Lemnaceae Duckweed Family. J. Arizona-Nevada Acad. Sci. 1992, 26, 10–14. [Google Scholar]
- Les, D.H.D.; Crawford, D.J.D.; Landolt, E.; Gabel, J.D.J.; Kimball, R.T.R. Phylogeny and Systematics of Lemnaceae, the Duckweed Family. Syst. Bot. 2002, 27, 221–240. [Google Scholar] [CrossRef]
- McLay, C.L. The effect of pH on the population growth of three species of duckweed: Spirodela oligorrhiza, Lemna minor, and Wolffia arrhiza. Freshw. Biol. 1976, 6, 125–136. [Google Scholar] [CrossRef]
- FAO. Duckweed: A Tiny Aquatic Plant with Enormous Potential for Agriculture and Environment; Animal Production and Health Div. University of Tropical Agriculture Foundation: Phnom Penh, Cambodia; Rome, Italy, 1999. [Google Scholar]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Relative in vitro growth rates of duckweeds (Lemnaceae)—The most rapidly growing higher plants. Plant Biol. 2014, 17, 1–9. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional value of duckweeds (Lemnaceae) as human food. Food Chem. 2017, 217, 266–273. [Google Scholar] [CrossRef]
- Cheng, J.J.; Stomp, A.M. Growing Duckweed to recover nutrients from wastewaters and for production of fuel ethanol and animal feed. Clean Soil Air Water 2009, 37, 17–26. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sowjanya Sree, K.; Bog, M.; Ecker, J.; Seeliger, C.; Böhm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Banasch, K.; et al. Nutritional value of the duckweed species of the Genus Wolffia (Lemnaceae) as human food. Front. Chem. 2018, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Van Dyke, J.M.; Sutton, D.L. Digestion of duckweed (Lemna spp.) by grass carp (Ctenopharyngodon idella). J. Fish Biol. 1977, 11, 273–278. [Google Scholar] [CrossRef]
- Rusoff, L.L.; Blakeney, E.W.; Culley, D.D. Duckweeds (Lemnaceae Family): A Potential Source of Protein and Amino Acids. J. Agric. Food Chem. 1980, 28, 848–850. [Google Scholar] [CrossRef]
- FAO. Fats and Fatty Acids in Human Nutrition; Report of an expert consultation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010; Volume 91. [Google Scholar]
- Hinoman—Cultivating Natures Wonder. Available online: https://www.hinoman.com/ (accessed on 10 March 2021).
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Health Care Interventions: Explanation and Elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef]
- Crowther, M.; Lim, W.; Crowther, M.A. Systematic review and meta-analysis methodology. Blood 2010, 116, 3140–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uman, L.S. Systematic Reviews and Meta-Analyses. J. Can. Acad. Child Adolesc. Psychiatry 2011, 20, 57. [Google Scholar] [PubMed]
- Ahn, E.; Kang, H. Introduction to systematic review and meta-analysis. Korean J. Anesthesiol. 2018, 71, 103. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, L.A.; Nardoto, G.B.; Perez, M.A.Z.; Junior, G.A.; Fracassi, F.C.; Oliveira, J.G.G.; Ottani, I.S.; Lima, S.H.; Mazzi, E.A.; Gomes, T.F.; et al. Carbon and Nitrogen Isotope Ratios of Food and Beverage in Brazil. Molecules 2020, 25, 1457. [Google Scholar] [CrossRef] [Green Version]
- Blainski, A.; Lopes, G.C.; De Mello, J.C.P. Application and Analysis of the Folin Ciocalteu Method for the Determination of the Total Phenolic Content from Limonium brasiliense L. Molecules 2013, 18, 6852. [Google Scholar] [CrossRef] [Green Version]
- Hagerman, A.E.; Butler, L.G. Protein Precipitation Method for the Quantitative Determination of Tannins. J. Agric. Food Chem. 1978, 26, 809–812. [Google Scholar] [CrossRef]
- Rohatgi, A. WebPlotDigitizer 4.2—Extract Data from Plots, Images, and Maps. Available online: https://automeris.io/WebPlotDigitizer/ (accessed on 10 December 2021).
- Pritchard, A. Statistical bibliography or bibliometrics? J. Doc. 1969, 24, 348–349. [Google Scholar]
- Zupic, I.; Čater, T. Bibliometric Methods in Management and Organization. Bibliometr. Methods Manag. Organ. 2014, 18, 429–472. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informer. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Lam, E.; Appenroth, K.J.; Michael, T.; Mori, K.; Fakhoorian, T. Duckweed in bloom: The 2nd International Conference on Duckweed Research and Applications heralds the return of a plant model for plant biology. Plant Mol. Biol. 2014, 84, 737–742. [Google Scholar] [CrossRef]
- Cao, H.X.; Fourounjian, P.; Wang, W. The Importance and Potential of Duckweeds as a Model and Crop Plant for Biomass-Based Applications and Beyond. In Handbook of Environmental Materials Management; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 1–16. [Google Scholar]
- Leng, R.A.; Stambolie, J.H.; Bell, R. Duckweed—A potential high-protein feed resource for domestic animals and fish. Livest. Res. Rural Dev. 1995, 7, 36. [Google Scholar]
- Parkar, S.G.; Frost, J.K.T.; Rosendale, D.; Stoklosinski, H.M.; Jobsis, C.M.H.; Hedderley, D.I.; Gopal, P. The sugar composition of the fiber in selected plant foods mod weaning infants’ gut microbiome composition and fermentation metabolites in vitro. Sci. Rep. 2021, 11, 9292. [Google Scholar] [CrossRef]
- Soares, S.; Brandão, E.; Guerreiro, C.; Soares, S.; Mateus, N.; De Freitas, V. Tannins in food: Insights into the molecular perception of astringency and bitter taste. Molecules 2020, 25, 2590. [Google Scholar] [CrossRef]
- Lacey, J.A.; Aston, J.E.; Thompson, V.S. Wear properties of ash minerals in biomass. Front. Energy Res. 2018, 6, 119. [Google Scholar] [CrossRef]
- Kenney, K.L.; Smith, W.A.; Gresham, G.L.; Westover, T.L. Understanding biomass feedstock variability. Biofuels 2013, 4, 111–127. [Google Scholar] [CrossRef]
- Chakrabarti, R.; Clark, W.D.; Sharma, J.G.; Goswami, R.K.; Shrivastav, A.K.; Tocher, D.R. Mass production of Lemna minor and its amino acid and fatty acid profiles. Front. Chem. 2018, 6, 479. [Google Scholar] [CrossRef] [PubMed]
- Vymazal, J. Constructed wetlands, surface flow. Encycl. Ecol. 2019, 1, 14–21. [Google Scholar]
- Goopy, J.P.; Murray, P.J. A review on the role of duckweed in nutrient reclamation and as a source of animal feed. Asian-Australasian J. Anim. Sci. 2003, 16, 297–305. [Google Scholar] [CrossRef]
- Jacobs, D.L. An ecological life-history of Spirodela polyrhiza (greater duckweed) with emphasis on the turions phase. Ecol. Monogr. 1947, 17, 439–469. [Google Scholar] [CrossRef]
- Mazen, A.M.A.; Zhang, D.; Franceschi, V.R. Calcium oxalate formation in Lemna minor: Physiological and ultrastructural aspects of high capacity calcium sequestration. New Phytol. 2004, 161, 435–448. [Google Scholar] [CrossRef]
- World of Cancer Research Fund; American Institute for Cancer Research Proteins. Food, Nutrition and the Prevention of Cancer: A Global Perspective; BANTA Book Group: Washington, DC, USA, 1997; pp. 394–397. ISBN 1-899533-05-2. [Google Scholar]
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Plant proteins as a high-quality nutritional source for the human diet. Trends Food Sci. Technol. 2020, 97, 170–184. [Google Scholar] [CrossRef]
- Porath, D.; Hepher, B.; Koton, A. Duckweed as an aquatic crop: Evaluation of clones for aquaculture. Aquat. Bot. 1979, 7, 273–278. [Google Scholar] [CrossRef]
- Nassar, H.F.; Shaban, A.M.; Bassem, S.M.; Abdel-Gawad, F.K. Utilization of duckweed (DW) in nutrient removal from agricultural waste water and producing alternative economic animal fodder. Pharma Chem. 2015, 7, 280–285. [Google Scholar]
- Sharma, J.G.; Kumar, A.; Saini, D.; Targay, N.L.; Khangembam, B.K.; Chakrabarti, R. In vitro digestibility study of some plant protein sources as aquafeed for carps Labeo rohita and Cyprinus carpio using pH-Stat method. Indian J. Exp. Biol. 2016, 54, 606–611. [Google Scholar]
- Bairagi, A.; Sarkar Ghosh, K.; Sen, S.K.; Ray, A.K. Duckweed (Lemna polyrhiza) leaf meal as a source of feedstuff in formulated diets for rohu (Labeo rohita Ham.) fingerlings after fermentation with a fish intestinal bacterium. Bioresour. Technol. 2002, 85, 17–24. [Google Scholar] [CrossRef]
- Da, C.T.; Lundh, T.; Lindberg, J.E. Digestibility of dietary components and amino acids in plant protein feed ingredients in striped catfish (Pangasianodon hypophthalmus) fingerlings. Aquac. Nutr. 2013, 19, 619–628. [Google Scholar] [CrossRef]
- El-Shafai, S.A.; El-Gohary, F.A.; Verreth, J.A.J.; Schrama, J.W.; Gijzen, H.J. Apparent digestibility coefficient of duckweed (Lemna minor), fresh and dry for Nile tilapia (Oreochromis niloticus L.). Aquac. Res. 2004, 35, 574–586. [Google Scholar] [CrossRef]
- Schneider, O.; Amirkolaie, A.K.; Vera-Cartas, J.; Editing, E.H.; Schrama, J.W.; Verreth, J.A.J. Digestibility, feces recovery, and related carbon, nitrogen, and phosphorus balances of five feed ingredients evaluated as fishmeal alternatives in Nile tilapia, Oreochromis niloticus L. Aquac. Res. 2004, 35, 1370–1379. [Google Scholar] [CrossRef]
- Cui, Y.; Chen, S.; Wang, S. Effect of ration size on the growth and energy budget of the grass carp, Ctenopharyngodon idella Val. Aquaculture 1994, 123, 95–107. [Google Scholar] [CrossRef]
- Hassan, M.R.; Chakrabarti, R. Use of Algae and Aquatic Macrophytes as Feed-In Small-Scale Aquaculture: A Review; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2009; Volume 531, pp. 1–123. [Google Scholar] [CrossRef]
- Dominguez, P.L.; Molinet, Y.; Ly, J. Ileal and in vitro digestibility in the pig of three floating aquatic macrophytes. Livest. Res. Rural Dev. 1996, 8, 37. Available online: http://www.lrrd.org/lrrd8/4/ly841.htm (accessed on 23 February 2022).
- Brown, W.Y.; Choct, M.; Pluske, J.R. Duckweed (Landoltia punctata) in dog diets dec digestibility but improves stool consistency. Anim. Prod. Sci. 2013, 53, 1188–1194. [Google Scholar] [CrossRef]
- Bouchard, S.S.; Murphy, A.K.; Berry, J.A. Non-additive dietary effects in juvenile slider turtles, Trachemys scriptarease. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, S.S.; Bjorndal, K.A. Nonadditive interactions between animal and plant diet items in an omnivorous freshwater turtle Trachemys scripta. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 144, 77–85. [Google Scholar] [CrossRef]
- Zeinstra, G.G.; Somhorst, D.; Oosterink, E.; Fick, H.; Klopping-Ketelaars, I.; Van Der Meer, I.M.; Mes, J.J. Postprandial amino acid, glucose and insulin responses among healthy adults after a single intake of Lemna minor in comparison with green peas: A randomised trial. J. Nutr. Sci. 2019, 8, e28. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.I.; Kayes, T.B.; Amundson, C.H. Requirements for lysine and arginine by rainbow trout (Oncorhynchus mykiss). Aquaculture 1992, 106, 333–344. [Google Scholar] [CrossRef]
- Morris, C.R.; Hamilton-Reeves, J.; Martindale, R.G.; Sarav, M.; Ochoa Gautier, J.B. Acquired Amino Acid Deficiencies: A Focus on Arginine and Glutamine. Nutr. Clin. Pract. 2017, 32, 30S–47S. [Google Scholar] [CrossRef] [PubMed]
- Jahreis, G.; Brese, M.; Leiterer, M.; Schäfer, U.; Böhm, V. Legume flours: Nutritionally important sources of protein and dietary fiber. Ernaehrungs Umsch. Int. 2016, 63, 36–42. [Google Scholar] [CrossRef]
- Borisjuk, N.; Peterson, A.A.; Lv, J.; Qu, G.; Luo, Q.; Shi, L.; Chen, G.; Kishchenko, O.; Zhou, Y.; Shi, J. Structural and biochemical properties of duckweed surface cuticle. Front. Chem. 2018, 6, 317. [Google Scholar] [CrossRef]
- Yan, Y.; Candreva, J.; Shi, H.; Ernst, E.; Martienssen, R.; Schwender, J.; Shanklin, J.; Landolt, E.; Les, D.; Hillman, W.; et al. Survey of the total fatty acid and triacylglycerol composition and content of 30 duckweed species and cloning of a Δ6-desaturase responsible for the production of γ-linolenic and stearidonic acids in Lemna gibba. BMC Plant Biol. 2013, 13, 201. [Google Scholar] [CrossRef] [Green Version]
- FAO. Carbohydrate in Human Nutrition; FAO: Rome, Italy, 1998. [Google Scholar]
- Ullah, H.; Gul, B.; Khan, H.; Zeb, U. Effect of salt stress on proximate composition of duckweed (Lemna minor L.). Heliyon 2021, 7, e07399. [Google Scholar] [CrossRef]
- Zhao, X.; Moates, G.K.; Wellner, N.; Collins, S.R.A.; Coleman, M.J.; Waldron, K.W. Chemical characterisation and analysis of the cell wall polysaccharides of duckweed (Lemna minor). Carbohydr. Polym. 2014, 111, 410–418. [Google Scholar] [CrossRef] [PubMed]
- World of Cancer Research Fund; American Institute for Cancer Research Carbohydrates. Food, Nutrition, and Prevention of Cancer: A Global Perspective; American Institute for Cancer Research Carbohydrates: Washington, DC, USA, 1997; pp. 376–383. [Google Scholar]
- Phillips, J.A. Dietary Guidelines for Americans, 2020–2025. Work. Health Saf. 2021, 69, 395. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell. Mol. Life Sci. 2016, 73, 2781. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Yu, C.; Yu, L.; Zhao, J.; Sun, C.; Ma, Y.; Zhou, G. The influence of light intensity and photoperiod on duckweed biomass and starch accumulation for bioethanol production. Bioresour. Technol. 2015, 187, 84–90. [Google Scholar] [CrossRef]
- Chen, L.; Yu, C.; Ma, Y.; Xu, H.; Wang, S.; Wang, Y.; Liu, X.; Zhou, G. Insights into the structural and physicochemical properties of small granular starches from two hydrophyte duckweeds, Spirodela oligorrhiza and Lemna minor. Carbohydr. Res. 2016, 435, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Li, J.M.; Du, A.P.; Liu, P.H.; Tian, X.P.; Jin, Y.L.; Yi, Z.L.; He, K.Z.; Fang, Y.; Zhao, H. High starch accumulation mechanism and phosphorus utilization efficiency of duckweed (Landoltia punctata) under phosphate starvation. Ind. Crops Prod. 2021, 167, 113529. [Google Scholar] [CrossRef]
- Xiao, Y.; Fang, Y.; Jin, Y.; Zhang, G.; Zhao, H. Culturing duckweed in the field for starch accumulation. Ind. Crops Prod. 2013, 48, 183–190. [Google Scholar] [CrossRef]
- Xu, J.; Cui, W.; Cheng, J.J.; Stomp, A.M. Production of high-starch duckweed and its conversion to bioethanol. Biosyst. Eng. 2011, 110, 67–72. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, F.; Daroch, M.; Tang, J. Positive effects of duckweed polycultures on starch and protein accumulation. Biosci. Rep. 2016, 36, e00380. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.M. CODEX-aligned dietary fiber definitions help to bridge the ‘fiber gap’. Nutr. J. 2014, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fiber in foods: A review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of the molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef]
- Pagliuso, D.; Grandis, A.; Igarashi, E.S.E.S.; Lam, E.; Buckeridge, M.S.M.S. Correlation of apiose levels and growth rates in duckweeds. Front. Chem. 2018, 6, 291. [Google Scholar] [CrossRef]
- Blazey, E.B.; McClure, J.W. The Distribution and Taxonomic Significance of Lignin in the Lemnaceae. Am. J. Bot. 1968, 55, 1240–1245. [Google Scholar] [CrossRef]
- Pagliuso, D.; Grandis, A.; Lam, E.; Buckeridge, M.S. High Saccharification, Low Lignin, and High Sustainability Potential Make Duckweeds Adequate as Bioenergy Feedstocks. Bioenergy Res. 2020, 14, 1082–1092. [Google Scholar] [CrossRef]
- Sowinski, E.E.; Gilbert, S.; Lam, E.; Carpita, N.C.; Carpita, C. Linkage structure of cell-wall polysaccharides from three duckweed species. Carbohydr. Polym. 2019, 223, 115119. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Bhaduri, S.K.; Sardar, D. Chemical characterization of pressed fibrous residues of four aquatic weeds. Aquat. Bot. 1991, 42, 81–85. [Google Scholar] [CrossRef]
- Mu, R.; Hong, X.; Ni, Y.; Li, Y.; Pang, J.; Wang, Q.; Xiao, J.; Zheng, Y. Recent trends and applications of cellulose nanocrystals in the food industry. Trends Food Sci. Technol. 2019, 93, 136–144. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, H.; Yang, B.; Weng, Y. Hemicellulose-Based Film: Potential Green Films for Food Packaging. Polymers 2020, 12, 1775. [Google Scholar] [CrossRef]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An overview of properties and application in food products. Int. J. Biol. Macromol. 2018, 119, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Zhao, X.; Moates, G.K.; Wilson, D.R.; Ghogare, R.J.; Coleman, M.J.; Waldron, K.W. Steam explosion pretreatment and enzymatic saccharification of duckweed (Lemna minor) biomass. Biomass Bioenergy 2015, 72, 206–215. [Google Scholar] [CrossRef]
- Souto, L.R.F.; da Silva, I.F.; Ninow, J.L.; Collins, S.R.A.; Elliston, A.; Waldron, K.W. Effect of hydrothermal pretreatment on duckweed (Landoltia punctata) biomass for simultaneous saccharification and fermentation process. Biomass Bioenergy 2019, 127, 105259. [Google Scholar] [CrossRef]
- Avci, U.; Peña, M.J.; O’Neill, M.A. Changes in the abundance of cell wall apiogalacturonan and xylogalacturonan and conservation of rhamnogalacturonan II structure during the diversification of the Lemnoideae. Planta 2018, 247, 953–971. [Google Scholar] [CrossRef]
- Thakur, B.R.; Singh, R.K.; Handa, A.K. Chemistry and uses of pectin—A review. Crit. Rev. Food Sci. Nutr. 2009, 37, 47–73. [Google Scholar] [CrossRef] [PubMed]
- Thibault, J.-F.; Ralet, M.-C. Physico-Chemical Properties of Pectins in the Cell Walls and After Extraction. Adv. Pectin Pectinase Res. 2003, 91–105. [Google Scholar] [CrossRef]
- Tsaban, G.; Yaskolka Meir, A.; Rinott, E.; Zelicha, H.; Kaplan, A.; Shalev, A.; Katz, A.; Rudich, A.; Tirosh, A.; Shelef, I.; et al. The effect of the green Mediterranean diet on cardiometabolic risk: A randomised controlled trial. Heart 2021, 107, 1054–1061. [Google Scholar] [CrossRef]
- Yan, P.; Xu, L.; He, N. Variation in the calorific values of different plants organs in China. PLoS ONE 2018, 13, e0199762. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, X.; Wang, S.; Chen, S. Growth and energy budget in young grass carp, Ctenopharyngodon idella Val., fed plant and animal diets. J. Fish Biol. 1992, 41, 231–238. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, J. Comparison of energy budget among six teleosts-II. Metabolic rates. Comp. Biochem. Physiol. Part A Physiol. 1990, 97, 169–174. [Google Scholar] [CrossRef]
- van der Spiegel, M.; Noordam, M.Y.; van der Fels-Klerx, H.J. Safety of novel protein sources (insects, microalgae, seaweed, duckweed, and rapeseed) and legislative aspects for their application in food and feed production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Oron, G.; Wildschut, L.R.; Porath, D. Waste water recycling by duckweed for protein production and effluent renovation. Water Sci. Technol. 1985, 17, 803–817. [Google Scholar] [CrossRef]
- Massey, L.K.; Roman-Smith, H.; Sutton, R.A.L. Effect of dietary oxalate and calcium on urinary oxalate and risk of formation of calcium oxalate kidney stones. J. Am. Diet. Assoc. 1993, 93, 901–906. [Google Scholar] [CrossRef]
- Noonan, S.C.; Savage, G.P. Oxalate content of foods and its effect on humans. Asia Pac. J. Clin. Nutr. 1999, 8, 64–74. [Google Scholar] [CrossRef]
- Sree, K.S.; Dahse, H.M.; Chandran, J.N.; Schneider, B.; Jahreis, G.; Appenroth, K.J. Duckweed for Human Nutrition: No Cytotoxic and No Anti-Proliferative Effects on Human Cell Lines. Plant Foods Hum. Nutr. 2019, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, N.E.; Walsh, É.; Bolger, P.; Burnell, G.; O’Leary, N.; O’Mahoney, M.; Paolacci, S.; Wall, D.; Jansen, M.A.K. Duckweed bioreactors: Challenges and opportunities for large-scale indoor cultivation of Lemnaceae. J. Clean. Prod. 2022, 336, 130285. [Google Scholar] [CrossRef]
Figure 1.
Bibliometric analysis of the occurrence and keyword distribution in the world’s research contributed to publications about duckweeds feedstocks. (A) Worldwide distribution network of publications among countries that have published papers related to nutritional feed-in duckweed species. The color intensity of the countries represents the number of articles found. (B) The panel of keywords that occur more often in recovered work from the bibliometric analysis. The larger the word size, the greater the word’s frequency in these studies (n = 80). Data extracted from 80 articles found in the Scopus database with the search described in Box 1. Accessed: 31 August 2021.
Figure 1.
Bibliometric analysis of the occurrence and keyword distribution in the world’s research contributed to publications about duckweeds feedstocks. (A) Worldwide distribution network of publications among countries that have published papers related to nutritional feed-in duckweed species. The color intensity of the countries represents the number of articles found. (B) The panel of keywords that occur more often in recovered work from the bibliometric analysis. The larger the word size, the greater the word’s frequency in these studies (n = 80). Data extracted from 80 articles found in the Scopus database with the search described in Box 1. Accessed: 31 August 2021.

Figure 2.
Ash and minerals found in duckweeds. (A) Ash, (B) Calcium, (C) Copper, (D) Iron, (E) Magnesium, (F) Manganese, (G) Nitrogen, (H) Phosphorus, (I) Potassium, (J) Sodium, and (K) Zinc.
Figure 2.
Ash and minerals found in duckweeds. (A) Ash, (B) Calcium, (C) Copper, (D) Iron, (E) Magnesium, (F) Manganese, (G) Nitrogen, (H) Phosphorus, (I) Potassium, (J) Sodium, and (K) Zinc.

Figure 3.
Fat, protein, carbohydrate, and fiber contents in duckweeds. (A) Fat (%) variability among the duckweed genera. (B) Proteins levels (%) variability in duckweeds. (C) Fibers (%) content. (D) Carbohydrates (%) variability in duckweed genera. (E) Starch (%) levels among duckweed genera.
Figure 3.
Fat, protein, carbohydrate, and fiber contents in duckweeds. (A) Fat (%) variability among the duckweed genera. (B) Proteins levels (%) variability in duckweeds. (C) Fibers (%) content. (D) Carbohydrates (%) variability in duckweed genera. (E) Starch (%) levels among duckweed genera.

Figure 4.
Amino acids levels in duckweeds by the percentage of protein. The data is shown in boxplots of the conversion of the percentage of protein of each amino acid recovered in the systematic review. (A) Alanine. (B) Arginine. (C) Aspartic acid. (D) Cysteine. (E) Glutamic acid. (F) Glycine. (G) Histidine. (H) Isoleucine. (I) Leucine. (J) Lysine. (K) Methionine. (L) Phenylalanine. (M) Serine. (N) Threonine. (O) Valine.
Figure 4.
Amino acids levels in duckweeds by the percentage of protein. The data is shown in boxplots of the conversion of the percentage of protein of each amino acid recovered in the systematic review. (A) Alanine. (B) Arginine. (C) Aspartic acid. (D) Cysteine. (E) Glutamic acid. (F) Glycine. (G) Histidine. (H) Isoleucine. (I) Leucine. (J) Lysine. (K) Methionine. (L) Phenylalanine. (M) Serine. (N) Threonine. (O) Valine.

Figure 5.
Cell wall polysaccharides and lignin levels of duckweeds. (A) Pectins (% cell wall). (B) Hemicellulose (% cell wall). (C) Cellulose (% cell wall). (D) Lignin (% cell wall).
Figure 5.
Cell wall polysaccharides and lignin levels of duckweeds. (A) Pectins (% cell wall). (B) Hemicellulose (% cell wall). (C) Cellulose (% cell wall). (D) Lignin (% cell wall).

Figure 6.
Monosaccharides from the cell wall of duckweeds. Data showed the intact cell wall (alcohol-insoluble residue—AIR). (A) Apiose (% biomass). (B) Arabinose (% biomass). (C) Fucose (% biomass). (D) Galactose (% biomass). (E) Galacturonic acid (% biomass). (F) Glucose (% biomass). (G) Mannose (% biomass). (H) Rhamnose (% biomass). (I) Xylose (% biomass).
Figure 6.
Monosaccharides from the cell wall of duckweeds. Data showed the intact cell wall (alcohol-insoluble residue—AIR). (A) Apiose (% biomass). (B) Arabinose (% biomass). (C) Fucose (% biomass). (D) Galactose (% biomass). (E) Galacturonic acid (% biomass). (F) Glucose (% biomass). (G) Mannose (% biomass). (H) Rhamnose (% biomass). (I) Xylose (% biomass).

Figure 7.
Overall duckweed composition.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General contents of carbon, nitrogen, protein, soluble sugar, and fiber in 21 ecotypes of duckweeds. Data represented by the mean ± standard error (n = 5).
Table 1.
General contents of carbon, nitrogen, protein, soluble sugar, and fiber in 21 ecotypes of duckweeds. Data represented by the mean ± standard error (n = 5).
| Species | %N | %C | C/N | Protein | % Soluble sugars | % Fiber | %Total aminoacids |
|---|---|---|---|---|---|---|---|
| Lemnoideae | |||||||
| Spirodela polyrhiza 7498 | 4.5 ± 0.0 | 38.1 ± 0.1 | 6.6 ± 0.0 | 28.4 ± 0.0 | 47.4 ± 0.5 | 52.6 ± 0.5 | 11.8 ± 6.8 |
| Spirodela polyrhiza 9346 | 5.3 ± 0.0 | 37.8 ± 0.2 | 6.6 ± 0.0 | 33.2 ± 0.0 | 46.4 ± 0.4 | 53.6 ± 0.4 | 9.2 ± 5.3 |
| Spirodela polyrhiza 9509 | 5.5 ± 0.0 | 36.8 ± 0.1 | 9.0 ± 0.0 | 34.7 ± 0.0 | 50.4 ± 0.3 | 49.6 ± 0.3 | 15.1 ± 8.7 |
| Spirodela polyrhiza 9264 | 4.0 ± 0.0 | 37.9 ± 0.2 | 7.9 ± 0.1 | 25.3 ± 0.0 | 46.1 ± 1.1 | 53.9 ± 1.1 | 2.5 ± 1.5 |
| Spirodela intermedia 7355 | 4.6 ± 0.0 | 38.1 ± 0.1 | 8.0 ± 0.0 | 28.6 ± 0.0 | 46.6 ± 0.4 | 53.4 ± 0.4 | 9.3 ± 5.4 |
| Landoltia punctata 7488 | 6.1 ± 0.0 | 39.8 ± 0.2 | 7.0 ± 0.1 | 38.3 ± 0.0 | 47.5 ± 0.9 | 52.5 ± 0.9 | 7.8 ± 4.5 |
| Landoltia punctata 7624 | 4.3 ± 0.0 | 39.0 ± 0.0 | 8.8 ± 0.0 | 27.1 ± 0.0 | 53.0 ± 0.4 | 47.0 ± 0.4 | 6.1 ± 3.5 |
| Landoltia punctata 7776 | 4.9 ± 0.0 | 36.2 ± 0.2 | 8.5 ± 0.1 | 30.4 ± 0.0 | 53.0 ± 0.8 | 47.0 ± 0.8 | 5.5 ± 3.2 |
| Lemna yungensis 9208 | 4.6 ± 0.1 | 36.4 ± 0.1 | 9.4 ± 0.0 | 28.8 ± 0.1 | 53.9 ± 0.5 | 46.1 ± 0.5 | 3.2 ± 1.9 |
| Lemna minor 8627 | 4.3 ± 0.0 | 38.3 ± 0.1 | 7.2 ± 0.0 | 26.8 ± 0.0 | 51.1 ± 1.0 | 48.9 ± 1.0 | 9.0 ± 5.2 |
| Lemna gibba DWC112 | 5.4 ± 0.0 | 37.5 ± 0.2 | 8.3 ± 0.0 | 33.5 ± 0.0 | 50.7 ± 0.6 | 49.3 ± 0.6 | 18.2 ± 10.5 |
| Lemna gibba DWC128 | 4.8 ± 0.0 | 37.4 ± 0.2 | 8.8 ± 0.0 | 29.9 ± 0.0 | 48.4 ± 1.0 | 51.6 ± 1.0 | 10.0 ± 5.8 |
| Lemna japonica 8695 | 4.2 ± 0.0 | 39.1 ± 0.2 | 8.8 ± 0.1 | 26.2 ± 0.0 | 50.7 ± 0.7 | 49.3 ± 0.7 | 3.5 ± 2.0 |
| Lemna obscura 9342 | 4.2 ± 0.0 | 37.1 ± 0.1 | 9.3 ± 0.0 | 26.3 ± 0.0 | 55.9 ± 0.7 | 44.1 ± 0.7 | 1.8 ± 1.0 |
| Average Lemnoideae | 4.8 | 37.8 | 8.2 | 29.8 | 50.1 | 49.9 | 8.1 |
| Wolffioideae | |||||||
| Wolffiella neotropica 8848 | 4.2 ± 0.0 | 37.2 ± 0.2 | 8.4 ± 0.1 | 26.5 ± 0.0 | 51.4 ± 0.6 | 48.6 ± 0.6 | 2.7 ± 1.5 |
| Wolffiella caudata 9139 | 5.7 ± 0.0 | 37.6 ± 0.1 | 7.4 ± 0.1 | 35.4 ± 0.0 | 54.5 ± 0.7 | 45.5 ± 0.7 | 18.6 ± 10.7 |
| Wolffiella gladiata 8261 | 4.6 ± 0.1 | 38.8 ± 0.1 | 7.4 ± 0.1 | 28.6 ± 0.1 | 48.6 ± 0.8 | 51.4 ± 0.8 | 16.1 ± 9.3 |
| Wolffia borealis 9144 | 4.6 ± 0.0 | 36.7 ± 0.1 | 7.8 ± 0.0 | 28.8 ± 0.0 | 50.7 ± 0.8 | 49.3 ± 0.8 | 2.8 ± 1.6 |
| Wolffia globosa 884 | 5.1 ± 0.0 | 36.9 ± 0.2 | 7.1 ± 0.1 | 31.9 ± 0.0 | 50.3 ± 0.9 | 49.7 ± 0.9 | 15.9 ± 9.2 |
| Wolffia globosa 8973 | 4.5 ± 0.0 | 39.5 ± 0.1 | 6.5 ± 0.0 | 28.2 ± 0.0 | 53.4 ± 0.5 | 46.6 ± 0.5 | 1.1 ± 0.7 |
| Wolffia arrhiza 8853 | 5.0 ± 0.0 | 36.9 ± 0.1 | 8.9 ± 0.1 | 31.1 ± 0.0 | 51.3 ± 1.6 | 48.7 ± 1.6 | 3.0 ± 1.7 |
| Average Wolffioideae | 4.8 | 37.6 | 7.7 | 30.1 | 51.5 | 48.5 | 8.6 |
| p-value | 2 × 10−16 | 2 × 10−16 | 2 × 10−16 | 2 × 10−16 | 2 × 10−16 | 2 × 10−16 | 1.42 × 10−15 |
Table 2.
Non-structural carbohydrates composition of 21 ecotypes of duckweeds. Data represented by the mean ± standard error (n = 5).
Table 2.
Non-structural carbohydrates composition of 21 ecotypes of duckweeds. Data represented by the mean ± standard error (n = 5).
| NON-STRUCTURAL CARBOHYDRATES (µg·mg−1 DW) | |||||
|---|---|---|---|---|---|
| Species | Glucose | Fructose | Sucrose | Raffinose | Starch |
| Lemnoideae | |||||
| Spirodela polyrhiza 7498 | 1.4 ± 0.2 | 6.3 ± 0.6 | 0.4 ± 0.1 | 0.2 ± 0.0 | 40.5 ± 2.3 |
| Spirodela polyrhiza 9346 | 0.6 ± 0.1 | 2.1 ± 0.3 | 0.6 ± 0.1 | 0.6 ± 0.0 | 30.4 ± 1.9 |
| Spirodela polyrhiza 9509 | 0.7 ± 0.1 | 3.5 ± 0.3 | 0.6 ± 0.1 | 0.0 ± 0.0 | 33.8 ± 3.2 |
| Spirodela polyrhiza 9264 | 0.7 ± 0.1 | 2.8 ± 0.2 | 1.0 ± 0.1 | 0.5 ± 0.0 | 16.8 ± 1.7 |
| Spirodela intermedia 7355 | 1.6 ± 0.2 | 3.6 ± 0.4 | 0.3 ± 0.1 | 0.6 ± 0.0 | 61.9 ± 1.5 |
| Landoltia punctata 7488 | 0.7 ± 0.1 | 1.3 ± 0.1 | 0.4 ± 0.0 | 1.0 ± 0.1 | 32.8 ± 2.2 |
| Landoltia punctata 7624 | 0.8 ± 0.1 | 2.9 ± 0.2 | 1.1 ± 0.1 | 1.1 ± 0.1 | 41.1 ± 1.9 |
| Landoltia punctata 7776 | 0.1 ± 0.0 | 0.2 ± 0.1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 11.4 ± 0.8 |
| Lemna yungensis 9208 | 1.1 ± 0.1 | 3.9 ± 0.2 | 1.2 ± 0.1 | 0.2 ± 0.1 | 55.8 ± 4.2 |
| Lemna minor 8627 | 5.6 ± 0.3 | 5.1 ± 0.3 | 0.1 ± 0.1 | 0.0 ± 0.0 | 97.5 ± 3.7 |
| Lemna gibba DWC112 | 0.8 ± 0.1 | 3.7 ± 0.3 | 1.7 ± 0.1 | 0.0 ± 0.0 | 138.5 ± 8.4 |
| Lemna gibba DWC128 | 0.5 ± 0.1 | 2.2 ± 0.3 | 0.7 ± 0.1 | 0.0 ± 0.0 | 30.4 ± 1.0 |
| Lemna japonica 8695 | 0.6 ± 0.0 | 2.7 ± 0.1 | 0.8 ± 0.1 | 0.0 ± 0.0 | 42.9 ± 4.1 |
| Lemna obscura 9342 | 0.9 ± 0.0 | 3.5 ± 0.2 | 1.5 ± 0.1 | 0.6 ± 0.0 | 38.5 ± 1.4 |
| Average Lemnoideae | 1.2 | 3.1 | 0.8 | 0.3 | 48.0 |
| Wolffioideae | |||||
| Wolffiella neotropica 8848 | 0.9 ± 0.1 | 3.1 ± 0.2 | 0.8 ± 0.0 | 0.1 ± 0.1 | 103.7 ± 1.2 |
| Wolffiella caudata 9139 | 0.6 ± 0.0 | 3.2 ± 0.7 | 3.3 ± 0.2 | 0.0 ± 0.0 | 97.2 ± 14.0 |
| Wolffiella gladiata 8261 | 3.3 ± 0.3 | 9.9 ± 0.8 | 1.1 ± 0.1 | 0.1 ± 0.1 | 81.9 ± 1.5 |
| Wolffia borealis 9144 | 1.1 ± 0.5 | 3.8 ± 1.4 | 0.5 ± 0.2 | 0.1 ± 0.0 | 111.2 ± 3.8 |
| Wolffia globosa 884 | 1.1 ± 0.1 | 4.0 ± 0.3 | 1.0 ± 0.1 | 0.4 ± 0.2 | 139.1 ± 3.8 |
| Wolffia globosa 8973 | 1.5 ± 1.0 | 3.4 ± 1.5 | 2.8 ± 2.4 | 0.0 ± 0.0 | 111.6 ± 1.7 |
| Wolffia arrhiza 8853 | 0.4 ± 0.0 | 1.9 ± 0.3 | 1.0 ± 0.1 | 0.0 ± 0.0 | 103.6 ± 3.9 |
| Average Wolffioideae | 1.3 | 4.2 | 1.5 | 0.1 | 106.9 |
| p-value | 2 × 10−16 | 4.83 × 10−15 | 0.01 | 2 × 10−16 | 2 × 10−16 |
Table 3.
Structural carbohydrates composition of 21 ecotypes of duckweeds. Data represented by the mean ± standard error (n = 5).
Table 3.
Structural carbohydrates composition of 21 ecotypes of duckweeds. Data represented by the mean ± standard error (n = 5).
| STRUCTURAL CARBOHYDRATES (µg·mg−1 DW) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Species | Fucose | Arabinose | Galactose | Rhamnose | Glucose | Xylose | Mannose | Apiose |
| Lemnoideae | ||||||||
| Spirodela polyrhiza 7498 | 1.6 ± 0.2 | 14.7 ± 1.2 | 21.3 ± 1.5 | 5.1 ± 0.5 | 4.9 ± 0.3 | 13.9 ± 0.7 | 11.5 ± 0.7 | 44.3 ± 4.3 |
| Spirodela polyrhiza 9346 | 0.8 ± 0.2 | 6.9 ± 0.6 | 11.8 ± 0.7 | 3.6 ± 0.2 | 3.4 ± 0.3 | 30.3 ± 0.3 | 6.7 ± 0.8 | 101.8 ± 17.1 |
| Spirodela polyrhiza 9509 | 2.3 ± 0.2 | 15.2 ± 0.9 | 20.3 ± 1.2 | 5.2 ± 0.3 | 6.3 ± 0.5 | 11.3 ± 0.5 | 8.9 ± 0.6 | 35.7 ± 2.3 |
| Spirodela polyrhiza 9264 | 1.6 ± 0.4 | 13.7 ± 2.1 | 20.0 ± 3.2 | 3.3 ± 0.7 | 3.5 ± 0.7 | 29.0 ± 1.4 | 9.8 ± 1.5 | 87.8 ± 12.8 |
| Spirodela intermedia 7355 | 1.7 ± 0.1 | 22.9 ± 0.4 | 19.9 ± 0.3 | 4.4 ± 0.1 | 7.8 ± 0.2 | 10.3 ± 0.1 | 5.8 ± 0.3 | 42.6 ± 1.4 |
| Landoltia punctata 7488 | 0.8 ± 0.1 | 11.0 ± 0.4 | 15.6 ± 0.5 | 2.8 ± 0.1 | 3.5 ± 0.1 | 21.8 ± 0.2 | 8.4 ± 0.8 | 69.4 ± 3.5 |
| Landoltia punctata 7624 | 2.0 ± 0.1 | 12.7 ± 0.4 | 16.9 ± 0.5 | 3.7 ± 0.1 | 5.1 ± 0.1 | 25.3 ± 0.2 | 7.7 ± 0.3 | 86.9 ± 3.1 |
| Landoltia punctata 7776 | 1.0 ± 0.2 | 13.0 ± 0.5 | 13.8 ± 0.6 | 2.7 ± 0.2 | 2.8 ± 0.3 | 27.1 ± 0.3 | 6.3 ± 0.5 | 97.2 ± 4.3 |
| Lemna yungensis 9208 | 1.6 ± 0.1 | 11.5 ± 0.2 | 13.9 ± 0.2 | 4.3 ± 0.1 | 7.6 ± 0.2 | 21.5 ± 0.1 | 8.4 ± 0.3 | 84.7 ± 1.6 |
| Lemna minor 8627 | 0.7 ± 0.1 | 6.3 ± 0.4 | 11.3 ± 0.6 | 2.5 ± 0.2 | 5.0 ± 0.4 | 27.9 ± 0.3 | 6.9 ± 0.5 | 106.8 ± 5.6 |
| Lemna gibba DWC112 | 1.3 ± 0.4 | 10.2 ± 1.4 | 18.1 ± 2.3 | 4.2 ± 0.7 | 12.2 ± 2.3 | 31.4 ± 1.0 | 11.4 ± 1.9 | 98.6 ± 12.9 |
| Lemna gibba DWC128 | 0.6 ± 0.1 | 7.7 ± 0.5 | 12.0 ± 0.8 | 2.3 ± 0.2 | 3.0 ± 0.3 | 23.6 ± 0.3 | 8.2 ± 0.2 | 91.6 ± 5.8 |
| Lemna japonica 8695 | 1.1 ± 0.1 | 7.8 ± 0.4 | 12.3 ± 0.5 | 2.4 ± 0.1 | 2.2 ± 0.1 | 24.8 ± 0.2 | 6.5 ± 0.3 | 88.0 ± 3.9 |
| Lemna obscura 9342 | 0.9 ± 0.2 | 9.7 ± 0.6 | 14.2 ± 0.8 | 2.6 ± 0.2 | 2.8 ± 0.2 | 25.5 ± 0.4 | 7.7 ± 0.4 | 85.3 ± 5.6 |
| Average Lemnoideae | 1.3 | 11.7 | 15.8 | 3.5 | 5.0 | 23.1 | 8.1 | 80.1 |
| Wolffioideae | ||||||||
| Wolffiella neotropica 8848 | 5.4 ± 0.3 | 23.6 ± 4.2 | 15.6 ± 2.8 | 4.5 ± 0.4 | 10.6 ± 2.2 | 18.9 ± 1.3 | 7.9 ± 0.5 | 21.4 ± 1.2 |
| Wolffiella caudata 9139 | 1.6 ± 0.1 | 37.7 ± 1.5 | 15.7 ± 0.5 | 2.6 ± 0.1 | 11.1 ± 0.4 | 20.6 ± 0.2 | 6.7 ± 0.3 | 27.7 ± 1.2 |
| Wolffiella gladiata 8261 | 1.2 ± 0.3 | 44.0 ± 5.7 | 18.5 ± 2.9 | 2.5 ± 0.5 | 11.2 ± 2.0 | 24.3 ± 1.3 | 8.3 ± 1.1 | 31.8 ± 4.8 |
| Wolffia borealis 9144 | 1.5 ± 0.1 | 24.8 ± 0.7 | 13.3 ± 0.7 | 2.1 ± 0.2 | 7.5 ± 0.2 | 25.9 ± 0.3 | 9.0 ± 0.6 | 9.1 ± 1.2 |
| Wolffia globosa 884 | 5.5 ± 0.4 | 28.8 ± 1.9 | 18.6 ± 1.1 | 4.6 ± 0.4 | 10.4 ± 0.6 | 16.9 ± 0.5 | 8.6 ± 0.4 | 15.8 ± 1.1 |
| Wolffia globosa 8973 | 1.1 ± 0.1 | 23.8 ± 1.2 | 12.2 ± 0.5 | 1.7 ± 0.1 | 10.4 ± 0.6 | 23.0 ± 0.2 | 9.7 ± 0.4 | 15.1 ± 1.3 |
| Wolffia arrhiza 8853 | 0.3 ± 0.1 | 20.5 ± 1.1 | 12.8 ± 0.8 | 2.6 ± 0.2 | 9.3 ± 0.6 | 26.6 ± 0.3 | 7.2 ± 0.7 | 16.6 ± 0.8 |
| Average Wolffioideae | 2.4 | 29.0 | 15.2 | 2.9 | 10.1 | 22.3 | 8.2 | 19.6 |
| p-value | 2 × 10−16 | 2 × 10−16 | 2 × 10−16 | 7.95 × 10−15 | 2 × 10−16 | 7.7 × 10−14 | 2.68 × 10−16 | 2 × 10−16 |
Table 4.
Essential amino acids composition of 21 duckweeds ecotypes. Data represented in g·mg−1 dry weight by average ± standard error (n = 3). HIS: histidine, TRP: tryptophan, MET: methionine, VAL: valine, PHE: phenylalanine, ILE: isoleucine, LEU: leucine, LYS: lysine, ALA: alanine, and TYR: tyrosine. Data represented by the mean ± standard error (n = 3).
Table 4.
Essential amino acids composition of 21 duckweeds ecotypes. Data represented in g·mg−1 dry weight by average ± standard error (n = 3). HIS: histidine, TRP: tryptophan, MET: methionine, VAL: valine, PHE: phenylalanine, ILE: isoleucine, LEU: leucine, LYS: lysine, ALA: alanine, and TYR: tyrosine. Data represented by the mean ± standard error (n = 3).
| Essential Amino Acids (mg·mg−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | HIS | TRP | MET | VAL | PHE | ILE | LEU | LYS | ALA+TIR |
| Lemnoideae | |||||||||
| Spirodela polyrhiza 7498 | 504.1 ± 291.1 | 449.9 ± 259.8 | 101.4 ± 58.5 | 3530.1 ± 2038.1 | 439.6 ± 253.8 | 1199.3 ± 692.4 | 613.8 ± 354.4 | 1123.4 ± 648.6 | 10486.5 ± 6054.4 |
| Spirodela polyrhiza 9346 | 399.1 ± 230.4 | 2044.5 ± 1180.4 | 138.3 ± 79.9 | 622.8 ± 359.6 | 105.6 ± 60.9 | 450.9 ± 260.3 | 237.3 ± 137.0 | 189.0 ± 109.1 | 4068.4 ± 2348.9 |
| Spirodela polyrhiza 9509 | 314.8 ± 181.8 | 259.0 ± 149.5 | 67.5 ± 39.0 | 3242.3 ± 1871.9 | 757.2 ± 437.2 | 1391.4 ± 803.3 | 1048.1 ± 605.1 | 3002.9 ± 1733.7 | 7830.4 ± 4520.9 |
| Spirodela polyrhiza 9264 | 338.6 ± 195.5 | 275.3 ± 159.0 | 305.9 ± 176.6 | 265.7 ± 153.4 | 406.1 ± 234.4 | 153.2 ± 88.5 | 92.3 ± 53.3 | 238.2 ± 137.5 | 986.6 ± 569.6 |
| Spirodela intermedia 7355 | 948.2 ± 547.5 | 616.9 ± 356.2 | 122.0 ± 70.4 | 1278.7 ± 738.3 | 863.8 ± 498.7 | 1465.2 ± 845.9 | 1126.3 ± 650.3 | 3206.5 ± 1851.3 | 2497.5 ± 1441.9 |
| Landoltia punctata 7488 | 250.9 ± 144.8 | 856.4 ± 494.5 | 304.0 ± 175.5 | 588.4 ± 339.7 | 564.7 ± 326.1 | 379.9 ± 219.4 | 311.2 ± 179.7 | 1760.1 ± 1016.2 | 2890.2 ± 1668.7 |
| Landoltia punctata 7624 | 314.6 ± 181.6 | 153.0 ± 88.3 | 15.0 ± 8.7 | 939.3 ± 542.3 | 341.7 ± 197.3 | 481.9 ± 278.2 | 409.3 ± 236.3 | 560.4 ± 323.5 | 4879.7 ± 2817.3 |
| Landoltia punctata 7776 | 269.4 ± 155.5 | 70.6 ± 40.8 | 36.4 ± 21.0 | 499.3 ± 288.3 | 150.7 ± 87.0 | 180.9 ± 104.4 | 518.6 ± 299.4 | 1372.3 ± 792.3 | 1272.1 ± 734.4 |
| Lemna yungensis 9208 | 485.4 ± 280.2 | 112.1 ± 64.7 | 96.3 ± 55.6 | 671.4 ± 387.6 | 167.8 ± 96.9 | 404.4 ± 233.5 | 266.2 ± 153.7 | 451.0 ± 260.4 | 2591.5 ± 1496.2 |
| Lemna minor 8627 | 0.0 ± 0.0 | 2114.5 ± 1220.8 | 26.6 ± 15.4 | 938.5 ± 541.8 | 629.7 ± 363.5 | 596.0 ± 344.1 | 674.7 ± 389.5 | 1657.2 ± 956.8 | 5096.8 ± 2942.6 |
| Lemna gibba DWC112 | 1590.8 ± 918.5 | 247.3 ± 142.8 | 141.7 ± 81.8 | 1661.9 ± 959.5 | 406.3 ± 234.6 | 764.1 ± 441.2 | 491.3 ± 283.7 | 3401.2 ± 1963.7 | 3963.6 ± 2288.4 |
| Lemna gibba DWC128 | 629.7 ± 363.5 | 610.6 ± 352.5 | 74.0 ± 42.7 | 1241.0 ± 716.5 | 1080.2 ± 623.6 | 881.6 ± 509.0 | 641.1 ± 370.1 | 1302.1 ± 751.8 | 6539.1 ± 3775.3 |
| Lemna japonica 8695 | 3.7 ± 2.1 | 282.7 ± 163.2 | 109.2 ± 63.0 | 937.1 ± 541.0 | 539.1 ± 311.2 | 172.9 ± 99.8 | 210.8 ± 121.7 | 1550.9 ± 895.4 | 2742.5 ± 1583.4 |
| Lemna obscura 9342 | 550.9 ± 318.1 | 281.6 ± 162.6 | 50.9 ± 29.4 | 989.0 ± 571.0 | 755.8 ± 436.4 | 668.2 ± 385.8 | 1345.5 ± 776.8 | 830.3 ± 479.4 | 3173.6 ± 1832.3 |
| Average Lemnoideae | 471.4 | 598.2 | 113.5 | 1243.2 | 514.9 | 656.4 | 570.5 | 1474.7 | 4215.6 |
| Wolffioideae | |||||||||
| Wolffiella neotropica 8848 | 469.8 ± 271.2 | 184.1 ± 106.3 | 46.5 ± 26.9 | 963.1 ± 556.1 | 363.6 ± 209.9 | 257.9 ± 148.9 | 391.1 ± 225.8 | 693.3 ± 400.3 | 1353.8 ± 781.6 |
| Wolffiella caudata 9139 | 1140.7 ± 658.6 | 778.7 ± 449.6 | 49.6 ± 28.6 | 802.2 ± 463.1 | 144.4 ± 83.4 | 499.7 ± 288.5 | 302.0 ± 174.4 | 2754.8 ± 1590.5 | 8648.1 ± 4993.0 |
| Wolffiella gladiata 8261 | 7046.1 ± 4068.1 | 2976.6 ± 1718.5 | 139.6 ± 80.6 | 2657.3 ± 1534.2 | 511.5 ± 295.3 | 1855.6 ± 1071.3 | 1882.9 ± 1087.1 | 2435.3 ± 1406.0 | 10885.6 ± 6284.8 |
| Wolffia borealis 9144 | 233.3 ± 134.7 | 147.7 ± 85.3 | 88.0 ± 50.8 | 242.0 ± 139.7 | 41.7 ± 24.1 | 226.1 ± 130.5 | 158.3 ± 91.4 | 880.0 ± 508.1 | 1098.7 ± 634.3 |
| Wolffia globosa 884 | 281.4 ± 162.4 | 156.0 ± 90.0 | 85.1 ± 49.1 | 1081.4 ± 624.3 | 367.5 ± 212.2 | 746.7 ± 431.1 | 729.5 ± 421.2 | 3166.6 ± 1828.2 | 6581.1 ± 3799.6 |
| Wolffia globosa 8973 | 335.7 ± 193.8 | 72.8 ± 42.0 | 19.0 ± 11.0 | 225.8 ± 130.4 | 156.5 ± 90.4 | 140.7 ± 81.2 | 339.0 ± 195.7 | 553.4 ± 319.5 | 588.9 ± 340.0 |
| Wolffia arrhiza 8853 | 602.8 ± 348.0 | 136.2 ± 78.6 | 50.6 ± 29.2 | 721.1 ± 416.3 | 71.6 ± 41.3 | 180.4 ± 104.2 | 143.1 ± 82.6 | 856.4 ± 494.4 | 5765.5 ± 3328.7 |
| Average Wolffioideae | 1444.3 | 636.0 | 68.3 | 956.1 | 236.7 | 558.2 | 563.7 | 1620.0 | 4988.8 |
| p-value | 5.5 × 10−4 | 2.1 × 10−16 | 8.4 × 10−5 | 1.6 × 10−7 | 4.6 × 10−12 | 6.1 × 10−7 | 2.3 × 10−4 | 6.2 × 10−7 | 3.2 × 10−15 |
Table 5.
Non-essential amino acids composition of 21 duckweeds ecotypes. Data represented in mg·mg−1 dry weight by average ± standard error (n = 3). ASP: aspartic acid, GLU: glutamic acid, ASN: asparagine, SER: serine, GLN: glutamine, CIT: citrulline, THR: threonine, GABA: Gamma-aminobutyric acid, and ORN: ornithine.
Table 5.
Non-essential amino acids composition of 21 duckweeds ecotypes. Data represented in mg·mg−1 dry weight by average ± standard error (n = 3). ASP: aspartic acid, GLU: glutamic acid, ASN: asparagine, SER: serine, GLN: glutamine, CIT: citrulline, THR: threonine, GABA: Gamma-aminobutyric acid, and ORN: ornithine.
| Non-Essential Amino Acids (mg·mg−1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | ASP | GLU | ASN | SER | GLN | GLY | CIT | THR | ARG | GABA | ORN |
| Lemnoideae | |||||||||||
| Spirodela polyrhiza 7498 | 2755.9 ± 1591.1 | 9083.5 ± 5244.4 | 22102.2 ± 12760.7 | 2071.6 ± 1196.0 | 418.2 ± 241.5 | 444.6 ± 256.7 | 4778.4 ± 2758.8 | 136.2 ± 78.7 | 1882.0 ± 1086.6 | 7812.6 ± 4510.6 | 759.7 ± 438.6 |
| Spirodela polyrhiza 9346 | 4708.9 ± 2718.7 | 15170.7 ± 8758.8 | 64239.2 ± 37088.5 | 2591.3 ± 1496.1 | 750.3 ± 433.2 | 155.0 ± 89.5 | 1608.2 ± 928.5 | 648.3 ± 374.3 | 3274.1 ± 1890.3 | 164.3 ± 94.8 | 326.6 ± 188.6 |
| Spirodela polyrhiza 9509 | 26923.3 ± 5217.2 | 10517.5 ± 6072.3 | 37448.3 ± 21620.8 | 2002.1 ± 1155.9 | 2271.6 ± 1311.5 | 336.3 ± 194.2 | 5299.8 ± 3059.8 | 602.5 ± 347.8 | 576.8 ± 333.0 | 1596.8 ± 921.9 | 603.8 ± 348.6 |
| Spirodela polyrhiza 9264 | 5523.1 ± 3188.7 | 2385.4 ± 1377.2 | 8076.5 ± 4663.0 | 933.4 ± 538.9 | 431.8 ± 249.3 | 131.1 ± 75.7 | 306.1 ± 176.7 | 637.5 ± 368.1 | 29.7 ± 17.1 | 357.5 ± 206.4 | 470.2 ± 271.5 |
| Spirodela intermedia 7355 | 13335.3 ± 7699.2 | 5619.6 ± 3244.5 | 35975.2 ± 20770.3 | 2640.4 ± 1524.4 | 1275.7 ± 736.5 | 551.2 ± 318.2 | 3193.2 ± 1843.6 | 765.2 ± 441.8 | 3907.3 ± 2255.9 | 3688.9 ± 2129.8 | 1784.2 ± 1030.1 |
| Landoltia punctata 7488 | 9216.0 ± 5320.9 | 9591.7 ± 5537.8 | 41039.1 ± 23693.9 | 979.2 ± 565.4 | 503.8 ± 290.9 | 465.9 ± 269.0 | 704.9 ± 407.0 | 775.0 ± 447.4 | 8699.8 ± 5022.8 | 1016.4 ± 586.8 | 717.1 ± 414.0 |
| Landoltia punctata 7624 | 16151.9 ± 9325.3 | 6412.3 ± 3702.2 | 21514.5 ± 12421.4 | 2649.4 ± 1529.6 | 664.8 ± 383.8 | 199.6 ± 115.2 | 3367.9 ± 1944.5 | 509.8 ± 294.3 | 645.7 ± 372.8 | 2148.9 ± 1240.7 | 601.7 ± 347.4 |
| Landoltia punctata 7776 | 9608.0 ± 5547.2 | 1709.7 ± 987.1 | 45128.4 ± 26054.9 | 216.1 ± 124.8 | 706.4 ± 407.9 | 624.9 ± 360.8 | 930.4 ± 537.2 | 368.9 ± 213.0 | 3180.4 ± 1836.2 | 691.3 ± 399.1 | 1412.5 ± 815.5 |
| Lemna yungensis 9208 | 9089.0 ± 5247.5 | 6993.6 ± 4037.8 | 6836.2 ± 3946.9 | 1268.0 ± 732.1 | 322.5 ± 186.2 | 214.1 ± 123.6 | 2075.2 ± 1198.1 | 63.5 ± 36.7 | 80.8 ± 46.7 | 1198.8 ± 692.2 | 101.8 ± 58.8 |
| Lemna minor 8627 | 4582.9 ± 2645.9 | 3632.9 ± 2097.5 | 41322.3 ± 23857.4 | 1152.9 ± 665.6 | 6158.5 ± 3555.6 | 123.4 ± 71.3 | 1530.9 ± 883.8 | 427.6 ± 246.9 | 12170.1 ± 7026.4 | 7802.0 ± 4504.5 | 1923.9 ± 1110.7 |
| Lemna gibba DWC112 | 29349.3 ± 16944.8 | 8453.2 ± 4880.4 | 42366.4 ± 24460.2 | 3694.7 ± 2133.2 | 883.4 ± 510.0 | 722.7 ± 417.3 | 1380.0 ± 796.7 | 670.7 ± 387.3 | 5795.1 ± 3345.8 | 5196.3 ± 3000.1 | 551.1 ± 318.2 |
| Lemna gibba DWC128 | 23730.2 ± 13700.6 | 8603.8 ± 4967.4 | 42427.7 ± 24495.6 | 1543.1 ± 890.9 | 241.3 ± 139.3 | 465.7 ± 268.8 | 2717.3 ± 1568.8 | 145.1 ± 83.8 | 11133.0 ± 6427.6 | 1224.7 ± 707.1 | 1418.3 ± 818.9 |
| Lemna japonica 8695 | 5909.2 ± 3411.7 | 6304.9 ± 3640.2 | 9904.4 ± 5718.3 | 427.5 ± 246.8 | 773.7 ± 446.7 | 547.6 ± 316.2 | 1507.8 ± 870.5 | 261.3 ± 150.9 | 645.1 ± 372.4 | 1616.9 ± 933.5 | 443.1 ± 255.8 |
| Lemna obscura 9342 | 7027.7 ± 4057.4 | 3596.1 ± 2076.2 | 5244.7 ± 3028.0 | 1067.1 ± 616.1 | 198.2 ± 114.4 | 241.9 ± 139.7 | 1103.2 ± 636.9 | 156.0 ± 90.1 | 799.0 ± 461.3 | 236.3 ± 136.4 | 82.4 ± 47.6 |
| Average Lemnoideae | 11993.6 | 7005.4 | 30258.9 | 1659.8 | 1114.3 | 373.2 | 2178.8 | 440.5 | 3772.8 | 2482.3 | 799.7 |
| Wolffioideae | |||||||||||
| Wolffiella neotropica 8848 | 4398.2 ± 2539.3 | 3989.8 ± 2303.5 | 9910.4 ± 5721.7 | 376.8 ± 217.5 | 129.2 ± 74.6 | 335.7 ± 193.8 | 873.5 ± 504.3 | 114.5 ± 66.1 | 2538.7 ± 1465.7 | 801.6 ± 462.8 | 660.8 ± 381.5 |
| Wolffiella caudata 9139 | 6228.7 ± 3596.1 | 6479.9 ± 3741.2 | 104530.3 ± 60350.6 | 2443.9 ± 1411.0 | 1146.9 ± 662.2 | 1165.1 ± 672.7 | 2486.5 ± 1435.6 | 547.6 ± 316.1 | 22032.5 ± 12720.5 | 9306.5 ± 5373.1 | 399.2 ± 230.5 |
| Wolffiella gladiata 8261 | 9452.0 ± 5457.1 | 5319.6 ± 3071.3 | 70743.1 ± 40843.5 | 4713.2 ± 2721.2 | 3109.7 ± 1795.4 | 703.1 ± 405.9 | 10242.5 ± 5913.5 | 1145.4 ± 661.3 | 16816.5 ± 9709.0 | 5412.2 ± 3124.7 | 2016.3 ± 1164.1 |
| Wolffia borealis 9144 | 5307.1 ± 3064.0 | 959.9 ± 554.2 | 15974.3 ± 9222.8 | 534.3 ± 308.5 | 243.6 ± 140.6 | 206.8 ± 119.4 | 506.3 ± 292.3 | 255.4 ± 147.4 | 845.2 ± 488.0 | 830.0 ± 479.2 | 1170.5 ± 675.8 |
| Wolffia globosa 884 | 16464.8 ± 9505.9 | 17920.2 ± 10346.2 | 28136.2 ± 16244.4 | 2026.1 ± 1169.8 | 770.7 ± 445.0 | 581.2 ± 335.6 | 954.8 ± 551.2 | 751.1 ± 433.6 | 4274.2 ± 2467.7 | 3009.0 ± 1737.2 | 476.6 ± 275.1 |
| Wolffia globosa 8973 | 2086.7 ± 1204.8 | 1827.2 ± 1055.0 | 7514.4 ± 4338.4 | 192.8 ± 111.3 | 62.8 ± 36.3 | 14.8 ± 8.6 | 245.6 ± 141.8 | 45.7 ± 26.4 | 292.5 ± 168.9 | 221.6 ± 127.9 | 226.8 ± 130.9 |
| Wolffia arrhiza 8853 | 3753.5 ± 2167.1 | 4399.4 ± 2540.0 | 15266.4 ± 8814.1 | 1937.8 ± 1118.8 | 453.4 ± 261.7 | 125.8 ± 72.6 | 820.8 ± 473.9 | 444.6 ± 256.7 | 671.9 ± 0.4 | 744.8 ± 430.0 | 186.5 ± 107.7 |
| Average Wolffioideae | 6813.0 | 5842.3 | 36010.7 | 1746.4 | 845.2 | 447.5 | 2304.3 | 472.0 | 6781.6 | 2903.7 | 733.8 |
| p-value | 9.4 × 10−16 | 2.5 × 10−13 | 2.0 × 10−16 | 3.5 × 10−10 | 2.0 × 10−16 | 7.6 × 10−10 | 7.2 × 10−6 | 2.8 × 10−8 | 2.0 × 10−16 | 3.9 × 10−8 | 1.4 × 10−13 |
Table 6.
Phenolics and tannin levels (mg·mg−1 dry weight) of 21 duckweeds species. Data represented by the mean ± standard error (n = 5).
Table 6.
Phenolics and tannin levels (mg·mg−1 dry weight) of 21 duckweeds species. Data represented by the mean ± standard error (n = 5).
| (mg·mg−1 DW) | ||
|---|---|---|
| Species | Total phenolics | Tannins |
| Lemnoideae | ||
| Spirodela polyrhiza 7498 | 36.1 ± 3.7 | 21.9 ± 2.0 |
| Spirodela polyrhiza 9346 | 29.2 ± 4.4 | 24.2 ± 0.9 |
| Spirodela polyrhiza 9509 | 48.8 ± 2.7 | 16.2 ± 0.9 |
| Spirodela polyrhiza 9264 | 79.2 ± 3.4 | 19.0 ± 1.1 |
| Spirodela intermedia 7355 | 76.0 ± 5.4 | 15.1 ± 0.8 |
| Landoltia punctata 7488 | 24.1 ± 3.6 | 29.1 ± 2.2 |
| Landoltia punctata 7624 | 38.2 ± 2.6 | 16.8 ± 1.6 |
| Landoltia punctata 7776 | 53.5 ± 4.2 | 24.3 ± 2.9 |
| Lemna yungensis 9208 | 45.9 ± 2.0 | 19.3 ± 1.1 |
| Lemna minor 8627 | 32.1 ± 2.9 | 2.4 ± 0.2 |
| Lemna gibba DWC112 | 41.1 ± 3.4 | 18.0 ± 0.6 |
| Lemna gibba DWC128 | 83.9 ± 2.8 | 27.4 ± 1.7 |
| Lemna japonica 8695 | 48.8 ± 5.8 | 21.0 ± 3.7 |
| Lemna obscura 9342 | 45.5 ± 1.4 | 18.3 ± 1.4 |
| Average Lemnoideae | 48.7 | 19.5 |
| Wolffioideae | ||
| Wolffiella neotropica 8848 | 35.5 ± 4.4 | 17.0 ± 1.7 |
| Wolffiella caudata 9139 | 24.0 ± 2.9 | 16.8 ± 1.0 |
| Wolffiella gladiata 8261 | 57.2 ± 4.2 | 21.2 ± 0.5 |
| Wolffia borealis 9144 | 29.4 ± 2.9 | 16.1 ± 0.7 |
| Wolffia globosa 884 | 55.2 ± 2.4 | 20.0 ± 1.4 |
| Wolffia globosa 8973 | 53.6 ± 2.7 | 17.2 ± 3.7 |
| Wolffia arrhiza 8853 | 45.3 ± 1.9 | 31.4 ± 6.7 |
| Average Wolffioideae | 42.9 | 20.0 |
| p-value | 2 × 10−16 | 9.96 × 10−11 |
Table 7.
Frequency of the recovery data from a systematic review among duckweed genera concerning feedstock thematic. Amino acids, ash, carbohydrates, cell wall, energy, fat, fiber, minerals, monosaccharides, protein, starch, and tannin were evaluated. The number of works recovered by the review is shown on the right of the table. The data content of each genus is shown in a box plot which, when the blue is darker, the higher the number of contemplated works. The occurrence refers to the number of entries of the variable. Variables marked with an asterisk (*) have included the data measured during the present work.
Table 7.
Frequency of the recovery data from a systematic review among duckweed genera concerning feedstock thematic. Amino acids, ash, carbohydrates, cell wall, energy, fat, fiber, minerals, monosaccharides, protein, starch, and tannin were evaluated. The number of works recovered by the review is shown on the right of the table. The data content of each genus is shown in a box plot which, when the blue is darker, the higher the number of contemplated works. The occurrence refers to the number of entries of the variable. Variables marked with an asterisk (*) have included the data measured during the present work.
| Variable | Works | Spirodela | Landoltia | Lemna | Wolffiella | Wolffia | Ocurrence |
|---|---|---|---|---|---|---|---|
| Amino acids* | 22 | 11 | 5 | 25 | 4 | 27 | 72 |
| Ash | 20 | 12 | 0 | 15 | 0 | 21 | 48 |
| Carbohydrate | 8 | 2 | 0 | 6 | 0 | 0 | 8 |
| Cell wall | 9 | 3 | 1 | 7 | 1 | 1 | 13 |
| Energy | 7 | 1 | 0 | 7 | 0 | 0 | 8 |
| Fat | 21 | 18 | 3 | 24 | 8 | 28 | 81 |
| Fiber* | 21 | 15 | 3 | 27 | 3 | 10 | 58 |
| Minerals | 13 | 1 | 0 | 7 | 0 | 16 | 24 |
| Monosaccharides* | 8 | 7 | 7 | 12 | 5 | 6 | 37 |
| Protein* | 43 | 25 | 9 | 47 | 4 | 29 | 114 |
| Starch* | 23 | 14 | 16 | 23 | 6 | 23 | 82 |
| Tannin* | 2 | 8 | 3 | 6 | 3 | 4 | 24 |
| Total | 197 | 117 | 47 | 206 | 34 | 165 | 569 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pagliuso, D.; Grandis, A.; Fortirer, J.S.; Camargo, P.; Floh, E.I.S.; Buckeridge, M.S. Duckweeds as Promising Food Feedstocks Globally. Agronomy 2022, 12, 796. https://doi.org/10.3390/agronomy12040796
AMA Style
Pagliuso D, Grandis A, Fortirer JS, Camargo P, Floh EIS, Buckeridge MS. Duckweeds as Promising Food Feedstocks Globally. Agronomy. 2022; 12(4):796. https://doi.org/10.3390/agronomy12040796
Chicago/Turabian StylePagliuso, Débora, Adriana Grandis, Janaina Silva Fortirer, Plinio Camargo, Eny Iochevet Segal Floh, and Marcos Silveira Buckeridge. 2022. "Duckweeds as Promising Food Feedstocks Globally" Agronomy 12, no. 4: 796. https://doi.org/10.3390/agronomy12040796
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.