
Cultural Characterization and Antagonistic Activity of Cladobotryum virescens against Some Phytopathogenic Fungi and Oomycetes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain, Growth Media and Conditions
2.2. Culture Characterization of Cladobotryum virescens INIFAT C10/110
2.3. Ethyl Acetate Crude Extract
2.4. Antagonistic Assays
- Competition for substrate: Evaluations were made every 24 h by analysis of the diameter of the colonies in cm. All tests were carried out in quintuplicates, and results were analyzed with a bifactorial variance analysis using the Statgraphics 5 program. Differences between treatments were determined by a Tukey test at a 5% significance level;
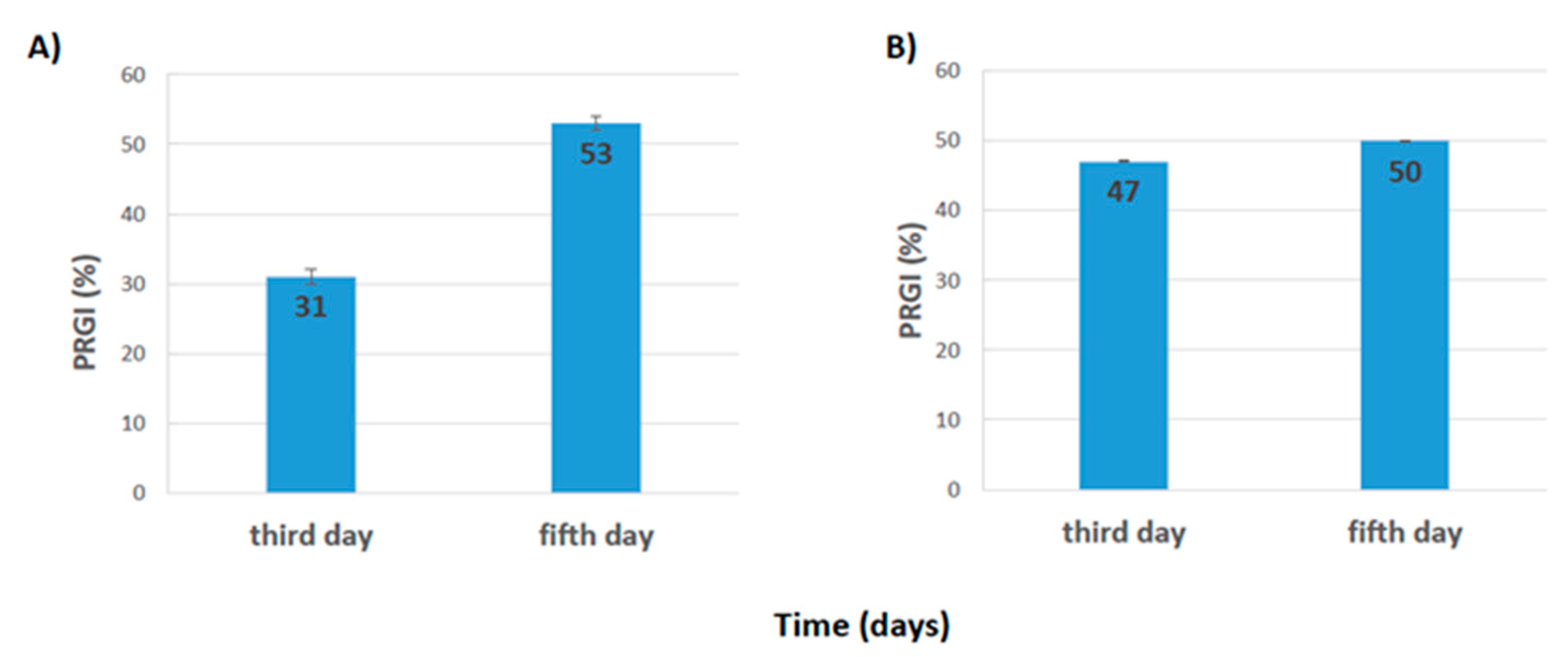
- Antibiosis: The percentage of radial growth inhibition (PRGI) was evaluated at three and five days after inoculation (before the colonies had contact) and was calculated as PRGI = (R1 − R2)/R1 * 100, where R1: radial growth of pathogen in control plates; R2: radial growth of pathogen in dual culture plates before the points of interaction [29];
- Mycoparasitism: Different types of mycoparasitism were evaluated: penetration (P), vacuolization (V), lysis (L), and coiling (C) [30]. From the area of interaction between both fungi (C. virescens and pathogen), three samples were taken. All the tests were carried out in triplicate. A drop of Lactofenol was added to the sample, which was analyzed with an optical microscope (Axioscop 40) at 100× and 400×.
2.5. Antifungal Screening of Ethyl Acetate Crude Extract
2.6. Statistical Analysis
3. Results
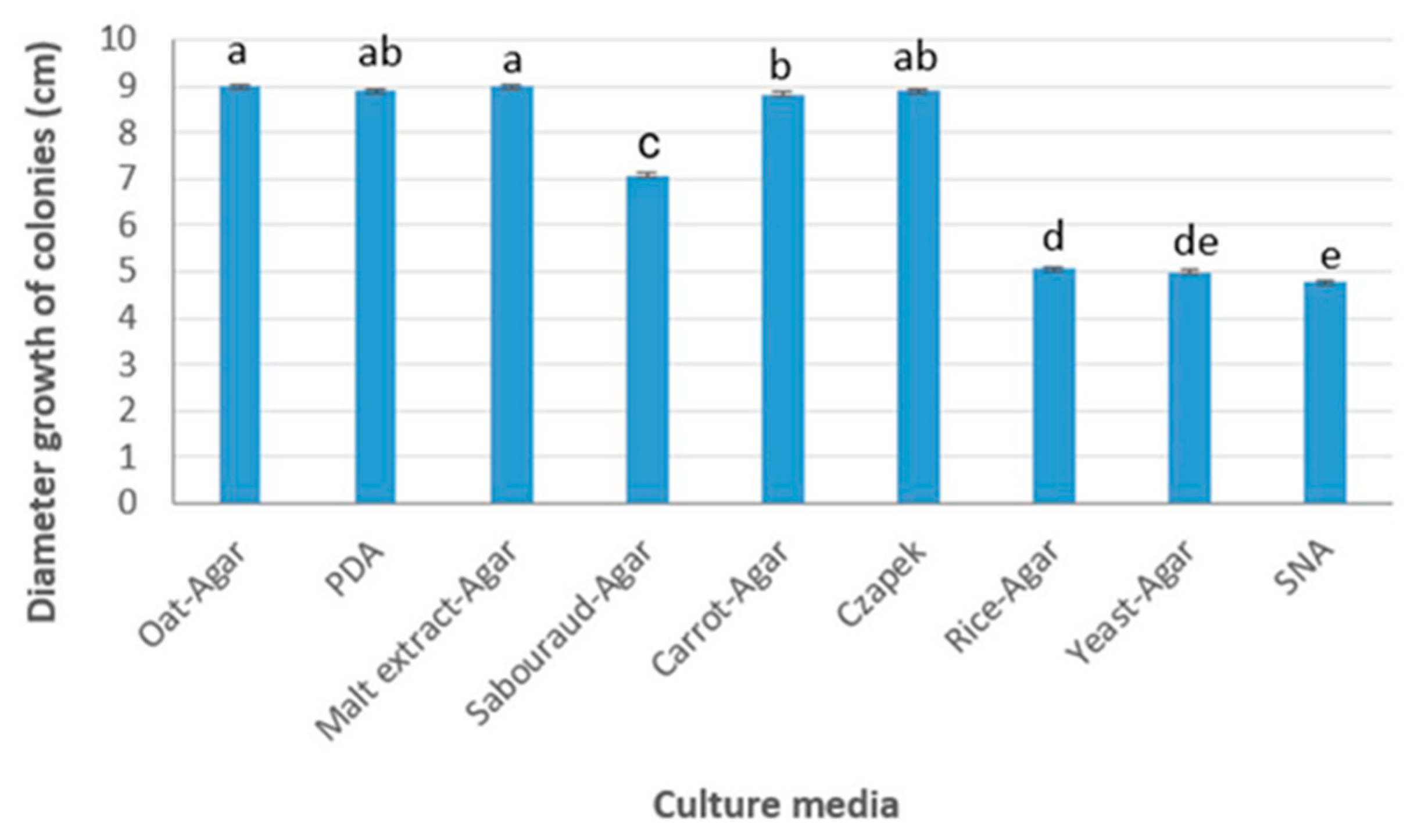
3.1. Culture Characterization
3.2. Antagonistic Assays
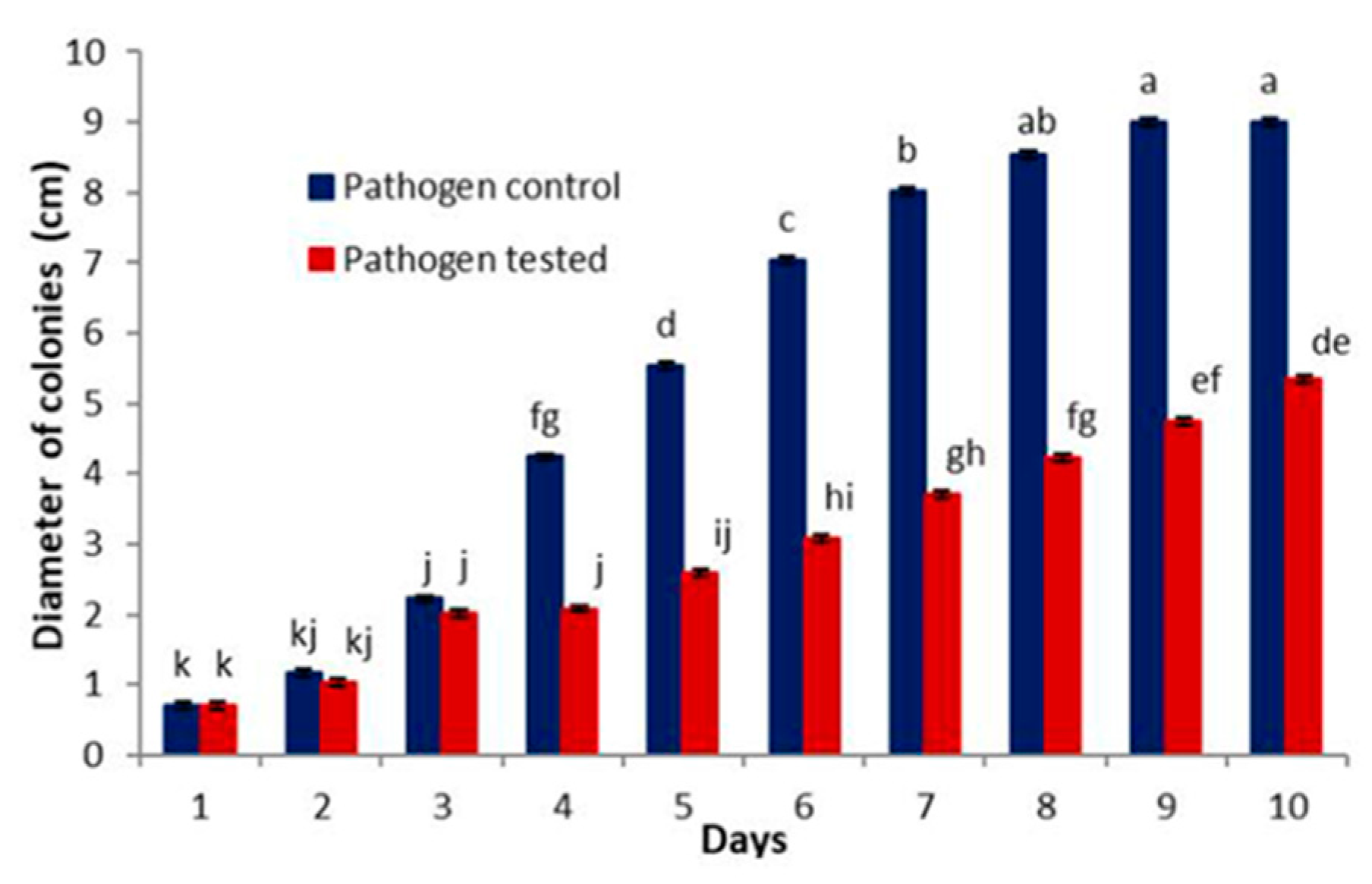
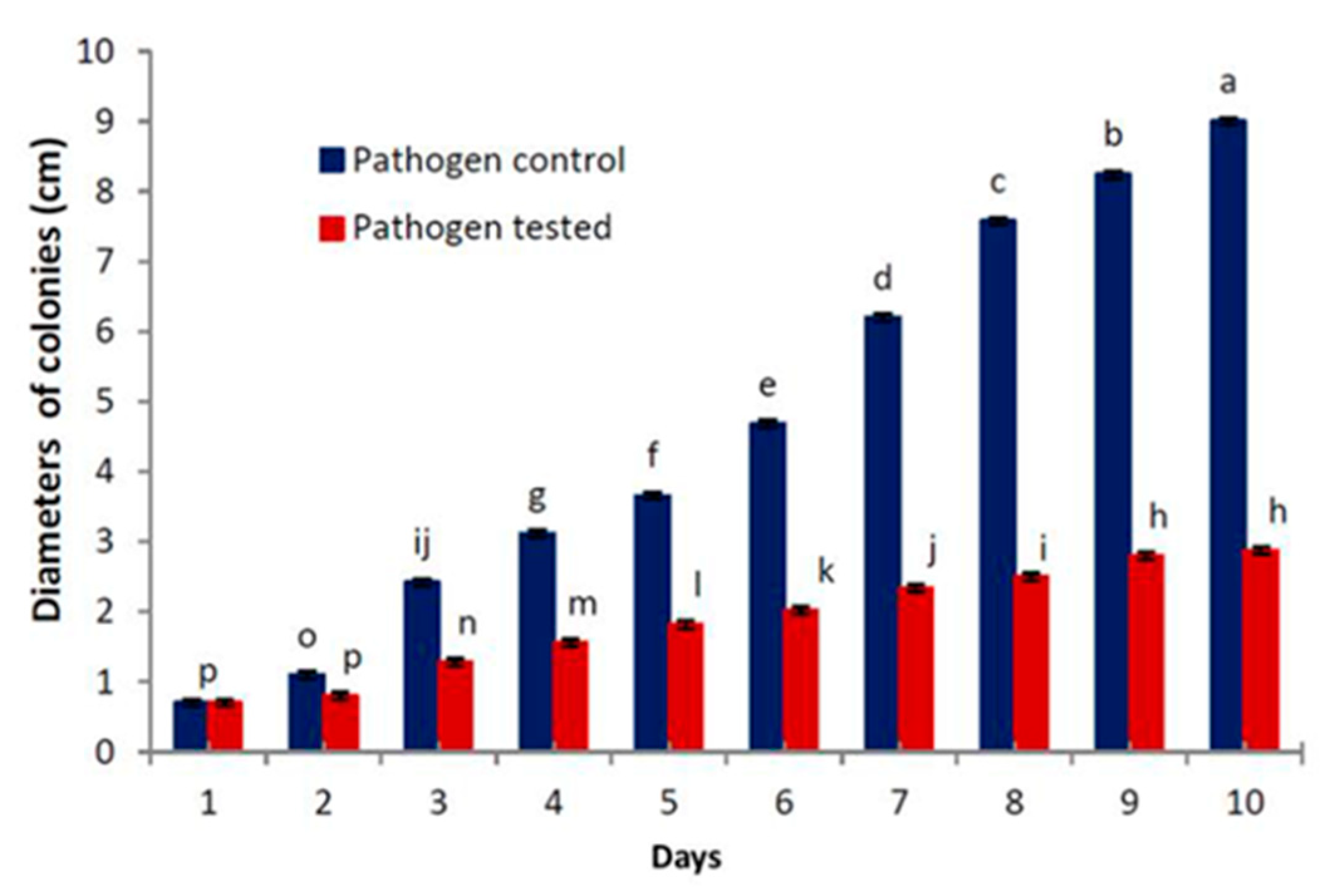
3.2.1. Competition for Substrate
3.2.2. Antibiosis
3.2.3. Mycoparasitism
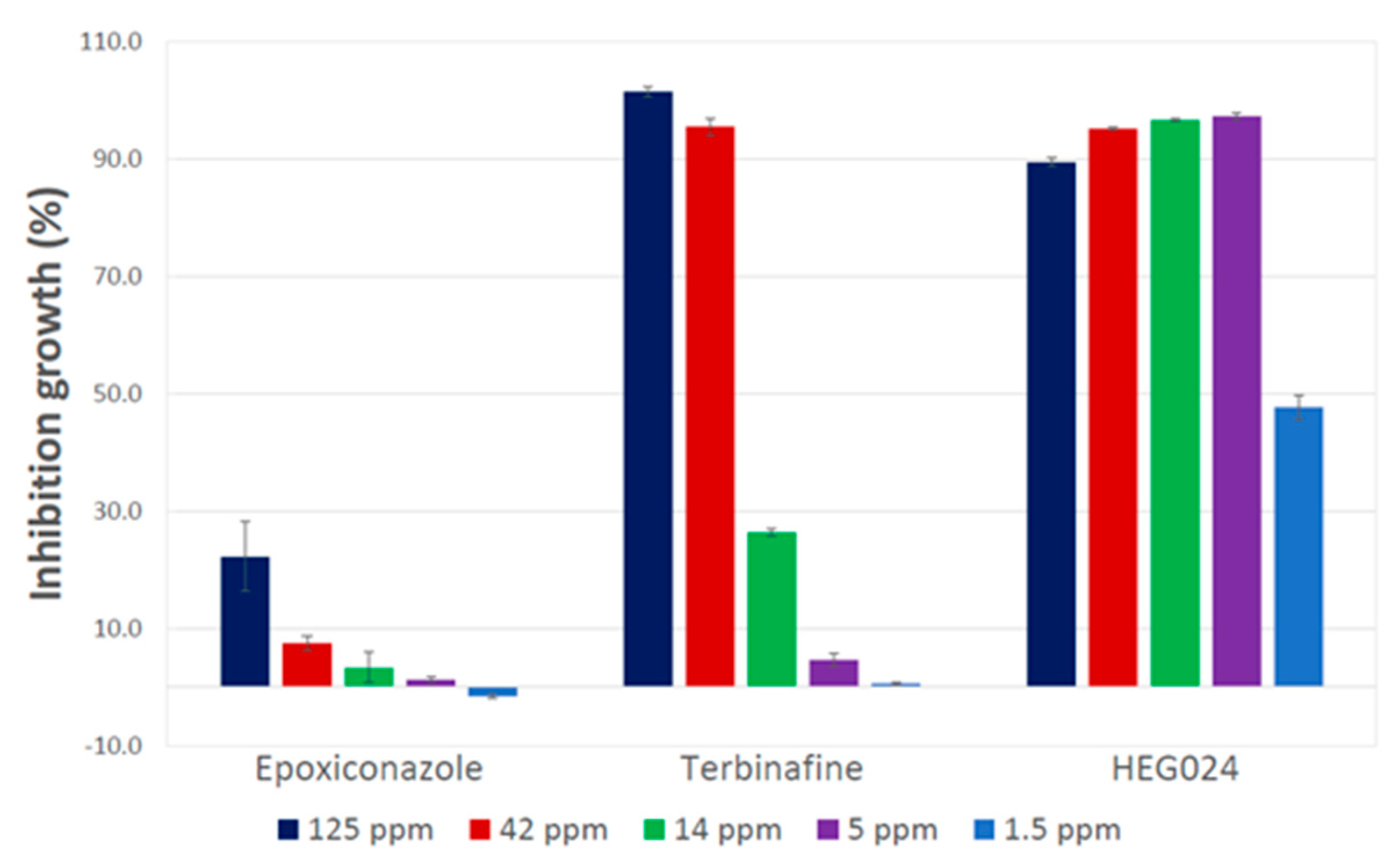
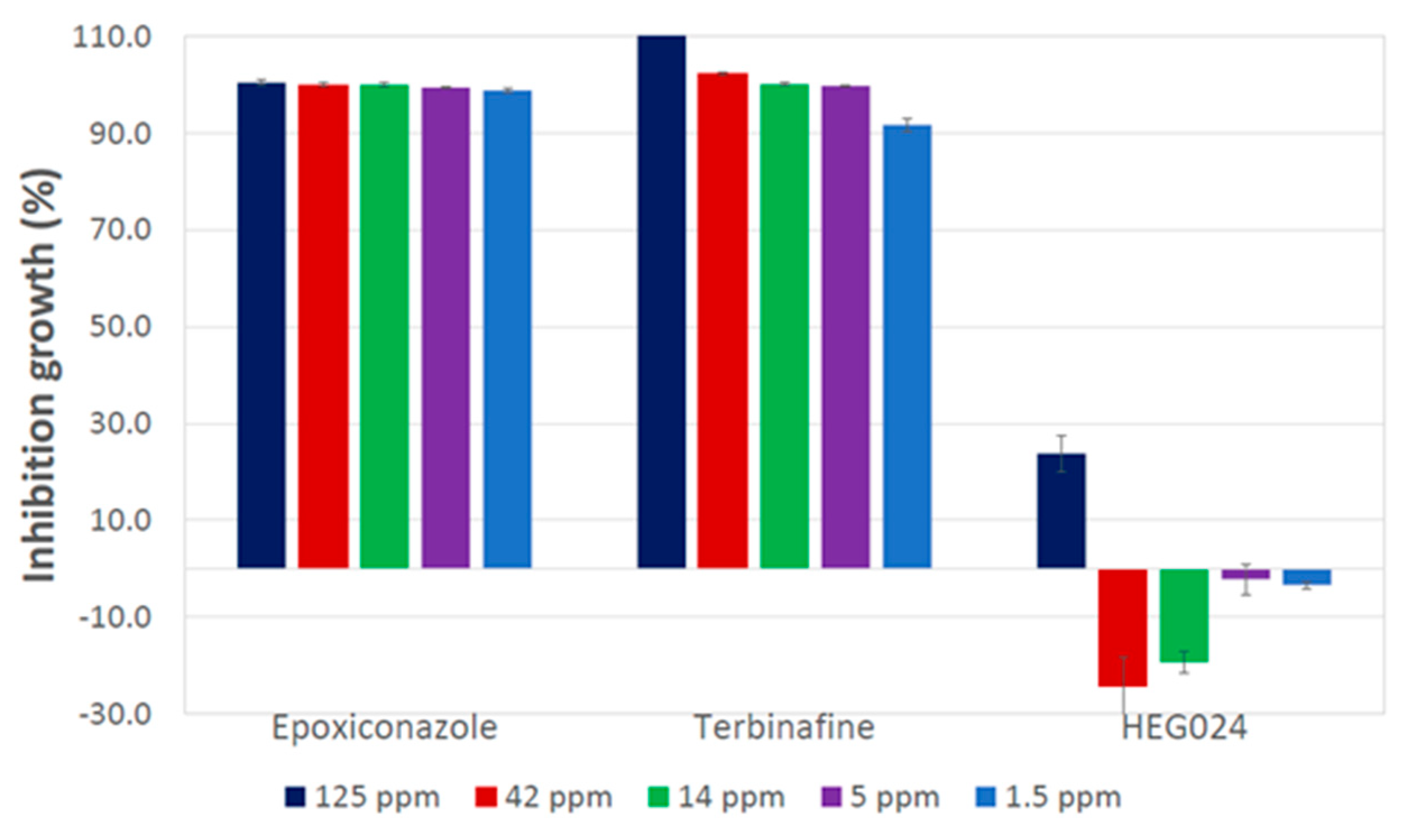
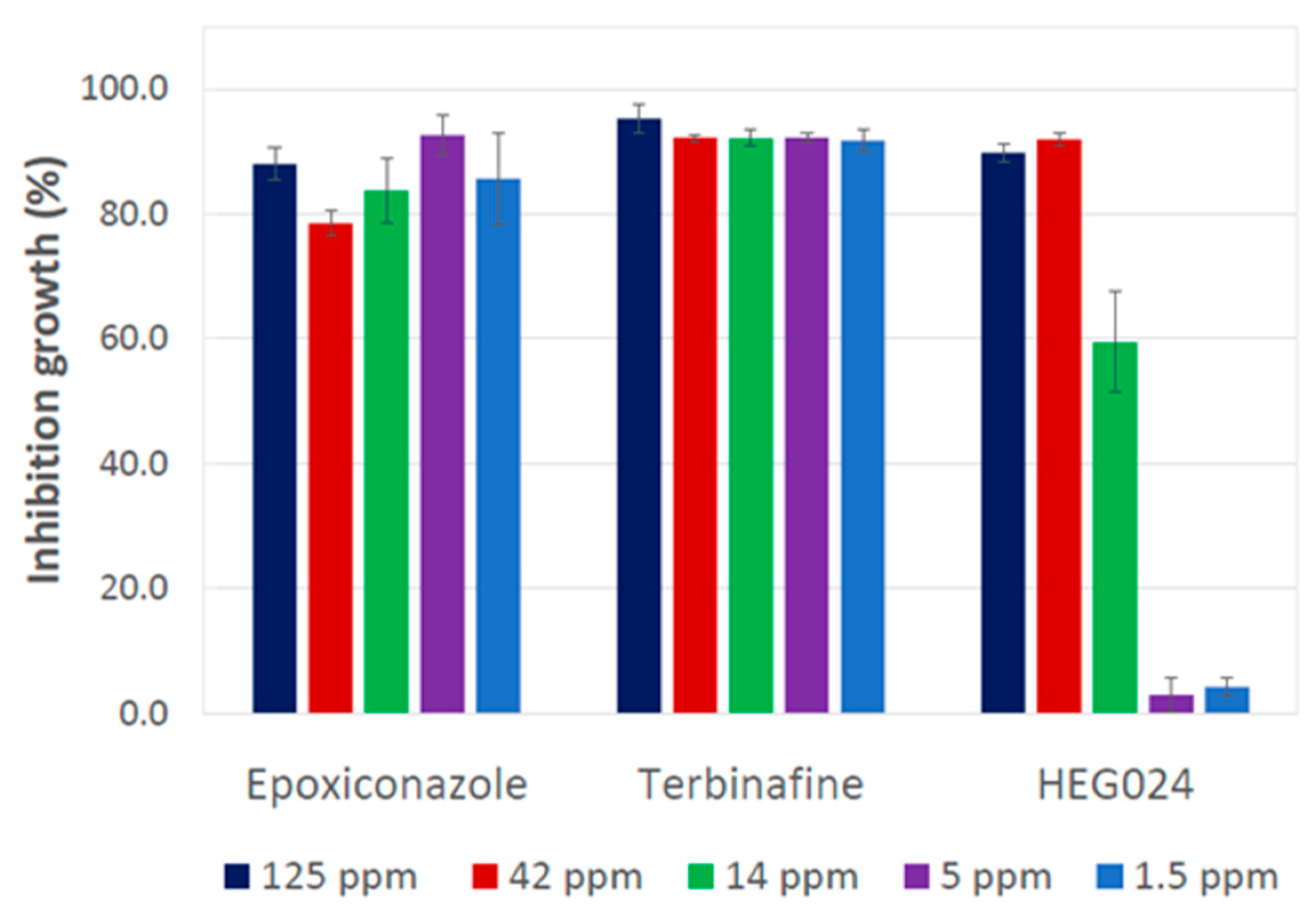
3.3. Testing of Ethyl Acetate Crude Extract from C. virescens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, K.; Han, G.; Ren, C.; Zhao, S.; Wu, X.; Bian, T. Fusarium solani infection depressed photosystem performance by inducing foliage wilting in apple seedlings. Front. Plant Sci. 2018, 9, 479. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.F.; Mohd, M.H.; Izham, N.M.; Zakaria, L. Pathogenicity of Fusarium semitectum and Fusarium chlamydosporum associated with pineapple fusariosis. Malays. J. Microbiol. 2016, 12, 164–170. [Google Scholar] [CrossRef]
- Macioszek, V.K.; Wielanek, M.; Morkunas, I.; Ciereszko, I.; Kononowicz, A.K. Leaf position-dependent effect of Alternaria brassicicola development on host cell death, photosynthesis and secondary metabolites in Brassica juncea. Physiol. Plant 2020, 168, 601–606. [Google Scholar] [CrossRef]
- Deep, S.; Sharma, P.; Behera, N.; Chowdappa, P. Diversity in India of Alternaria brassicicola (Schwein) Wiltsshire, causing black leaf spot diseases in Cauliflower. Plant Pathol. J. 2014, 13, 232–245. [Google Scholar] [CrossRef]
- Tomas-Grau, R.H.; Hael-Conrad, V.; Requena-Serra, F.J.; Perato, S.M.; Caro, M.d.P.; Salazar, S.M.; Díaz-Ricci, J.C. Biological control of strawberry grey mold disease caused by Botrytis cinerea mediated by Colletotrichum acutatum extracts. BioControl 2020, 65, 461–473. [Google Scholar] [CrossRef]
- Petrasch, S.; Knapp, S.J.; Van Kan, J.A.L.; Blanco-Ulate, B. Grey mould of strawberry, a devastating disease caused by the ubiquitous necrotrophic fungal pathogen Botrytis cinerea. Mol. Plant Path. 2019, 20, 877–892. [Google Scholar] [PubMed]
- Collins, F.; Bancal, P.; Spink, J.; Kock, P.; Smith, J.; Paveley, N.D.; Bancal, M.-O.; Foulkes, M.J. Wheat lines exhibiting variation in tolerance of Septoria tritici blotch differentiated by grain source limitation. Field Crops Res. 2018, 217, 1–10. [Google Scholar] [CrossRef]
- Brennan, C.J.; Benbow, H.R.; Mullins, E.; Doohan, F.M. A review of the known unknowns in the early stages of Septoria tritici blotch disease of wheat. Plant Pathol. 2019, 68, 1427–1438. [Google Scholar] [CrossRef]
- Morales, J.G.; Gaviria, A.E.; Gilchrist, E. Allelic variation and selection in effector genes of Phytophthora infestans (Mont.) de Bary. Pathogens 2020, 9, 551. [Google Scholar] [CrossRef]
- Seifu, Y.W. Reducing severity of late blight (Phytophthora infestans) and improving potato (Solanum tuberosum L.) tuber yield with pre-harvest application of calcium nutrients. Agronomy 2017, 7, 69. [Google Scholar] [CrossRef]
- Abbey, J.A.; Percival, D.; Lord, A.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)—Prospects and challenges. Biocontrol Sci. Technol. 2018, 29, 207–228. [Google Scholar] [CrossRef]
- Moura, R.D.; de Castro, L.A.M.; Culik, M.P.; Fernandes, A.A.R.; Fernandes, P.M.B.; Ventura, J.A. Culture medium for improved production of conidia for identification and systematic studies of Fusarium pathogens. J. Microbiol. Methods 2020, 173, 105915. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.F.; Platas, G.; Overy, J.; Fillola, A.; Jiménez, M.R.; Collado, J.; Vicente, F.; Martín, J.; Gonzalez, A.; Burzimmermann, J.; et al. Enhancement of antibiotic and secondary metabolite detection from filamentous fungi by growth on nutritional arrays. J. Appl. Microbiol. 2008, 104, 1644–1658. [Google Scholar] [CrossRef]
- Uzcátegui, N.M.C.; Serrano, J.A.; Panizo, M.M. Condiciones de cultivo que fomentan la producción de sustancias antimicrobianas en actinomicetos patógenos y del suelo. Rev. Soc. Venez. Microbiol. 2013, 33, 134–139. [Google Scholar]
- Back, C.G.; Lee, C.H.Y.; Seo, G.S.; Jung, H.Y. Characterization of species of Cladobotryum which cause cobweb disease in endible mushrooms grown in Korea. Mycobiology 2012, 40, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, J.; Jensen, H.C.; Kajaer, A.; Olsen, C.E.; Rassing, B.R.; Rosendahl, C.N.; Sotofte, I. Cladobotryal: A fungal metabolite with a novel ring system. Acta Chem. Scand. 1998, 52, 631–63417. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, Y.; Ramos, B.; Álvarez, M.E.; Valdés, R.; Lorenzo, Y.; Plana, L.; Marrero, I.; García, D.; Aguado, Y.; Ruenes, M.E.; et al. Actividad fungicida de cepas del género Cladobotryum sobre Corynespora cassiicola. Agrotec. Cuba 2016, 40, 71–83. [Google Scholar]
- Ramos, B. Actividad fungicida de cepas de Cladobotryum spp., para el control de hongos patógenos presentes en modalidades productivas de la agricultura urbana. Agrotec. Cuba 2018, 42, 98–99. [Google Scholar]
- Põldmaa, K. Tropical species of Cladobotryum and Hypomyces producing red pigments. Stud. Mycol. 2011, 68, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Index Fumgorum Fungal Names Search. Available online: http//:www.indexfungorum.org/names/Names.asp2021 (accessed on 24 September 2021).
- Centraalbureau Voor Schimmelcutures (CBS). List of Cultures; Fungal Biodiversity Center: Utrecht, The Netherlands, 2001; p. 687. [Google Scholar]
- Stammler, G.; Speakman, J. Microtiter Method to Test the Sensitivity of Botrytis cinerea to Boscalid. J. Phytopathol. 2006, 154, 508–510. [Google Scholar] [CrossRef]
- Stammler, G. Phytophthora infestans Microtiter Method with Sporangia. Available online: http://www.frac.info/docs/default-source/monitoring-methods/approved-methods/phytin-microtiter-method-sporangia-basf-2006-v1.pdf?sfvrsn=4. (accessed on 21 April 2021).
- Stammler, G.; Semar, M. Sensitivity of Mycosphaerella graminicola (anamorph: Septoria tritici) to DMI fungicides across Europe and impact on field performance. Bull. OEPP 2011, 41, 149–155. [Google Scholar] [CrossRef]
- Crous, P.W.; Verkley, G.J.M.; Groenewald, J.Z.; Samsom, R.A. Fungal Biodiversity; CBS Laboratory Manual Series; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2009; Volume 1, p. 269. [Google Scholar]
- Frey, D.; Oldfield, R.J.; Bridger, R.C. A Color Atlas of Pathogenic Fungi; Barry, G., Ed.; Wolfe Medical Publication Ltd London: London, UK, 1979; p. 168. Available online: https://catalogue.nla.gov.au/Record/2917010 (accessed on 31 May 2019).
- Bernal, A.; Martínez, B.; Castellanos, M.D. Caracterización Cultural de Tres Aislamientos de Cladosporium fulvum Cooke. Cent. Agríc. 2005, 32, 93–94. Available online: https://scholar.google.com/scholar?hl=es&as_sdt=0%2C5&q=Bernal%2C+A.%3B+Mart%C3%ADnez%2C+B.%3B+Castellanos%2C+M.D.+Caracterizaci%C3%B3n+cultural+de+tres+aislamientos+de+Cladosporium+fulvum+Cooke.+Centro+421+Agr%C3%ADcola+2005%2C+32%2C+93%E2%80%9394.&btnG=. (accessed on 31 May 2019).
- Martínez, B.; Reyes, Y.; Infante, D.; González, E.; Baños, H.; Cruz, A. Selección de aislamientos de Trichoderma spp. candidatos a biofungicidas para el control de Rhizoctonia sp. en arroz. Rev. Prot. Veg. 2008, 23, 118–125. [Google Scholar]
- Baiyee, B.; Ito, S.; Sunpapao, A. Trichoderma asperellum T1 mediated antifungal activity and induced defense response against leaf spot fungi in lettuce (Lactuca sativa L.). Physiol. Mol. Plant Pathol. 2019, 106, 96–101. [Google Scholar] [CrossRef]
- Reyes, Y.; Martínez, B.; Infante, D. Evaluación de la actividad antagónica de trece aislamientos de Trichoderma spp. sobre Rhizoctonia sp. Rev. Prot. Veg. 2008, 23, 112–117. [Google Scholar]
- Ren, G.C.; Wanasinghe, D.N.; Monkai, J.; Mortimer, P.E.; Hyde, K.D.; Xu, J.C.; Pang, A.; Gui, H. Novel saprobic Hermatomyces species (Hermatomycetaceae, Pleosporales) from China (Yunnan Province) and Thailand. MycoKeys 2021, 82, 57–79. [Google Scholar] [CrossRef]
- Sun, J.Z.; Liu, X.Z.; Jeewon, R.; Li, Y.L.; Lin, C.G.; Tian, Q.; Zhao, Q.; Xiao, X.P.; Hyde, K.D.; Nilthong, S. Fifteen fungicolous Ascomycetes on edible and medicinal mushrooms in China and Thailand. Asian J. Mycol. 2019, 2, 129–169. [Google Scholar] [CrossRef]
- Konopická, J.; Bohatá, A.; Zemek, R.; Čurn, V. The effects of natural substrates and artificial media on the production of conidiospores and blastospores of the entomopathogenic fungus Isaria fumosorosea, strain CCM 8367. Microb. Nematode Control. Invertebr. Pests IOBC-WPRS Bull. 2017, 129, 58–64. [Google Scholar]
- Gandarilla-Pacheco, F.L.; Arévalo-Niño, K.; Galán-Wong, L.J.; Sandoval-Coronado, C.F.; Quintero-Zapata, I. Evaluation of conidia production and mycelial growth in solid culture media from native strains of entomopathogenic fungi isolated from citrus-growing areas of México. Afr. J. Biotech. 2012, 11, 14453–14460. [Google Scholar] [CrossRef]
- Chithra, J.; Kulkarni, S.; Gururaj Sunkad, S.B.; Amresh, Y.S.; Shekhar, P. Effect of different media, temperature, pH and illumination on the growth of Fusarium oxysporum f.sp. zingiberi causing wet rot of ginger. J. Pharmacogn. Phytochem. 2020, 9, 786–790. [Google Scholar]
- Assefa, A.; Abate, D. Evaluation of nutritional requirements of medicinal fungus, Pyrofomes demidoffii under submerged fermentation. Biocatal. Agric. Biotech. 2020, 28, 101765. [Google Scholar] [CrossRef]
- Ulrich, S.; Schäfer, C. Toxin production by Stachybotrys chartarum genotype S on different culture media. J. Fungi 2020, 6, 159. [Google Scholar] [CrossRef]
- Shirouzou, T.; Hirose, D.; Tokumasu, S.; To-Anum, C.; Maekawa, N. Host affinity and phylogenetic position of new anamorphic fungus Beltraniella botryospora from living and fallen leaves of evergreen oaks. Fungal Divers. 2010, 43, 85–92. [Google Scholar] [CrossRef]
- Santos, M.; Diánez, F.; Moreno-Gavíra, A.; Sánchez-Montesinos, B.; Gea, F.J. Cladobotryum mycophilum as Potential Biocontrol Agent. Agronomy 2019, 9, 891. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Mohiddin, F.A. Trichoderma: Its multifarious utility in crop improvement. In Crop Improvement trough Microbial Biotechnology. New and Future Developments in Microbial Biotechnology and Bioengineering; Aligarh Muslim University: Aligarh, India; Srinagar, India, 2018; Chapter 13; pp. 263–291. [Google Scholar] [CrossRef]
- Khaledi, N.; Taheri, P. Biocontrol mechanisms of Trichoderma harziamam against soybean charcoal rot caused by Macrophomina phaseolina. J. Plant Prot. Res. 2016, 56, 21–31. [Google Scholar] [CrossRef]
- Fadhilah, N.; Kheng, Y.; Jiat, H.; Keng, Y.; Joo, K. Evaluation on the cultural characteristics and antagonistic activities of Cladobotryum semicirculare against Ganoderma boninense in vitro. J. Oil Palm Res. 2015, 27, 326–338. [Google Scholar]
- Zhou, T.; Katsuragawa, M.; Xing, T.; Fukaya, K.; Okuda, T.; Tokiwa, T.; Tashiro, E.; Imoto, M.; Oku, N.; Urabe, D.; et al. Cyclopeptides from the Mushroom Pathogen Fungus Cladobotryum varium. J. Nat. Prod. 2021, 84, 327–338. [Google Scholar] [CrossRef]
- Sakemi, S.; Bordner, J.; De Costa, D.L.; Dekker, K.A.; Hirai, H.; Inagaki, T.; Kim, Y.J.; Kojima, N.; Sims, C.J.; Siugie, Y.; et al. CJ-15,696 and its analogs, new furopyridine antibiotics from the fungus Cladobotryum varium: Fermentation, isolation, structural elucidation, biotransformation and antibacterial activities. J. Antibiot. 2002, 55, 6–18. [Google Scholar] [CrossRef]
- Mitova, M.I.; Lang, G.; Blunt, J.W.; Cummings, N.J.; Cole, A.L.; Robinson, W.T.; Munro, M.H. Cladobotric acids A-F: New cytotoxic polyketides from a New Zealand Cladobotryum sp. J. Org. Chem. 2006, 71, 492–497. [Google Scholar] [CrossRef]
- El Komy, M.H.; Saleh, A.A.; Eranthodi, A.; Molan, Y.Y. Characterization of Novel Trichoderma asperellum Isolates to Select Effective Biocontrol Agents Against Tomato Fusarium Wilt. Plant Pathol. J. 2015, 31, 50–60. [Google Scholar] [CrossRef]
- Infante, D.; Martínez, B.; González, N.; Reyes, Y. Mecanismos de acción de Trichoderma frente a hongos fitopatógenos. Rev. Prot. Veg. 2009, 24, 14–21. [Google Scholar]
- Singh, S.; Bhagat, S.; Singh, V.; Hashmi, S.; Bhagel, M. Evaluation of biocontrol agents against Phytophthora drechsleri f.sp. cajani (Leaf Blight) in Pigeonpea. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 3536–3543. [Google Scholar] [CrossRef]
- Savazzini, F.; Oliveira, C.M.; Pertot, I. Impact of the biocontrol agent Trichoderma atroviride SC1 on soil microbial communities of a vineyard in northern Italy. Soil Biol. Biochem. 2009, 41, 1457–1465. [Google Scholar] [CrossRef]
- Tezuka, Y.; Huang, Q.; Kikuchi, T.; Nishi, A.; Tubaki, K. Studies on the metabolites of mycoparasitic fungi. l. Metabolites of Cladobotryum varium. Chem. Pharm. Bull. 1994, 42, 2612–2617. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Number |
|---|---|
| Alternaria brassicicola Schwein | INIFAT 4145 |
| Botrytis cinerea Pers. | IPB KSH 892 |
| Cladobotryum virescens G.R.W. Arnold | INIFAT C10/110 |
| Fusarium chlamydosporum Wollenw and Reinking | INIFAT 4147 |
| Phytophthora infestans (Mont.) de Bary | IPB KSH 894 |
| Septoria tritici Rob. ex Desm. | IPB KSH 895 |
| Culture Media | Conidia Production Spores/mL |
|---|---|
| Oat agar | 28.2 × 106 |
| Potato dextrose agar | 25.0 × 106 |
| Malt extract agar | 23.7 × 106 |
| Sabouraud agar | 18.4 × 106 |
| Carrot agar | 27.8 × 106 |
| Czapek agar | 24.0 × 106 |
| Yeast agar | - |
| SNA agar | - |
| Rice agar | 8.2 × 106 |
| Strain | Culture Media | Coloration | Pellet Formation | Mycelium | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Day 5 | Day 10 | Day 15 | Day 5 | Day 10 | Day 15 | Day 5 | Day 10 | Day 15 | ||
| C. virescens INIFAT C10/110 | PDA | Light yellow | White | White | - | Low | Low | Low | Low | Low |
| Oat agar | Red | Red | Red | High | High | High | Abundant | Abundant | Abundant | |
| Days | C. virescens Control (Cc) | Pathogen Control (Pc) | Cladobotryum virescens Tested (CT) | Pathogen Tested (PT) | Media Proportion (Cc/Pc) | Media Proportion (CT/PT) |
|---|---|---|---|---|---|---|
| 1 | 0.7 | 0.7 | 0.7 | 0.7 | 1.00 | 1.00 |
| 2 | 1.34 | 1.16 | 1.46 | 1.04 | 1.16 | 1.40 |
| 3 | 2.18 | 2.22 | 2.36 | 2.02 | 0.98 | 1.17 |
| 4 | 2.9 | 4.24 | 2.86 | 2.08 | 0.68 | 1.38 |
| 5 | 3.76 | 5.54 | 3.74 | 2.58 | 0.68 | 1.45 |
| 6 | 4.68 | 7.04 | 4.64 | 3.08 | 0.66 | 1.51 |
| 7 | 5.42 | 8.02 | 5.5 | 3.7 | 0.68 | 1.49 |
| 8 | 6.5 | 8.54 | 6.74 | 4.22 | 0.76 | 1.60 |
| 9 | 7.7 | 9 | 7.6 | 4.7 | 0.86 | 1.62 |
| 10 | 8.94 | 9 | 9 | 5.34 | 0.99 | 1.69 |
| Days | Cladobotryum virescens Control (Cc) | Pathogen Control (Pc) | Cladobotryum virescens Tested (CT) | Pathogen Tested (PT) | Media Proportion (Cc/Pc) | Media Proportion (CT/PT) |
|---|---|---|---|---|---|---|
| 1 | 0.7 | 0.7 | 0.7 | 0.7 | 1.00 | 1.00 |
| 2 | 1.3 | 1.1 | 1.36 | 0.8 | 1.18 | 1.70 |
| 3 | 2.18 | 2.42 | 2.22 | 1.28 | 0.90 | 1.73 |
| 4 | 2.92 | 3.12 | 3.16 | 1.56 | 0.94 | 2.03 |
| 5 | 3.66 | 3.66 | 3.86 | 1.82 | 1.00 | 2.12 |
| 6 | 4.62 | 4.68 | 4.68 | 2.02 | 0.99 | 2.32 |
| 7 | 5.5 | 6.2 | 5.56 | 2.34 | 0.89 | 2.38 |
| 8 | 6.4 | 7.58 | 6.58 | 2.5 | 0.84 | 2.63 |
| 9 | 7.54 | 8.24 | 7.74 | 2.8 | 0.92 | 2.76 |
| 10 | 9 | 9 | 9 | 2.9 | 1.00 | 3.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández, G.; Ramos, B.; Sultani, H.N.; Ortiz, Y.; Spengler, I.; Castañeda, R.F.; Rivera, D.G.; Arnold, N.; Westermann, B.; Mirabal, Y. Cultural Characterization and Antagonistic Activity of Cladobotryum virescens against Some Phytopathogenic Fungi and Oomycetes. Agronomy 2023, 13, 389. https://doi.org/10.3390/agronomy13020389
Hernández G, Ramos B, Sultani HN, Ortiz Y, Spengler I, Castañeda RF, Rivera DG, Arnold N, Westermann B, Mirabal Y. Cultural Characterization and Antagonistic Activity of Cladobotryum virescens against Some Phytopathogenic Fungi and Oomycetes. Agronomy. 2023; 13(2):389. https://doi.org/10.3390/agronomy13020389
Chicago/Turabian StyleHernández, Giselle, Beatriz Ramos, Haider N. Sultani, Yarelis Ortiz, Iraida Spengler, Rafael F. Castañeda, Daniel G. Rivera, Norbert Arnold, Bernhard Westermann, and Yaneris Mirabal. 2023. "Cultural Characterization and Antagonistic Activity of Cladobotryum virescens against Some Phytopathogenic Fungi and Oomycetes" Agronomy 13, no. 2: 389. https://doi.org/10.3390/agronomy13020389