Recovery of 2R.2Sk Triticale-Aegilops kotschyi Robertsonian Chromosome Translocations

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chromosome Preparation
2.3. Probe Preparation and Fluorescence in Situ Hybridization
2.4. Lr54 + Lr37 SSR Marker Screening
3. Results
3.1. Chromosome Segregation in 40 + M2R + M2Sk Plants of Triticale
3.2. Chromosome Segregation in 40 + M2RS + M2RL + M2Sk Plants of Triticale
3.3. Lr54 + Yr37 SSR Markers Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistics Database 1998. Available online: http://www.fao.org/faostat (accessed on 28 June 2019).
- McGoverin, C.M.; Snyders, F.; Muller, N.; Botes, W.; Fox, G.; Manley, M. A review of triticale uses and the effect of growth environment on grain quality. J. Sci. Food Agric. 2011, 91, 1155–1165. [Google Scholar] [CrossRef]
- Meale, S.J.; McAllister, T.A. Grain for Feed and Energy. In Triticale; Eudes, F., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 167–187. [Google Scholar]
- Pattison, A.L.; Trethowan, R.M. Characteristics of modern triticale quality: Commercially significant flour traits and cookie quality. Crop Pasture Sci. 2013, 64, 874–880. [Google Scholar] [CrossRef]
- Rakha, A.; Åman, P.; Andersson, R. Dietary fiber in triticale grain: Variation in content, composition, and molecular weight distribution of extractable components. J. Cereal Sci. 2011, 54, 324–331. [Google Scholar] [CrossRef]
- Woś, H.; Brzeziński, W. Triticale for Food—The Quality Driver. In Triticale; Eudes, F., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 213–232. [Google Scholar]
- Arseniuk, E. Triticale Diseases—A Review. In Triticale: Today and Tomorrow; Guedes-Pinto, H., Darvey, N., Carnide, V.P., Eds.; Springer: Dordrecht, The Netherlands, 1996; pp. 499–525. [Google Scholar]
- Arseniuk, E.; Góral, T. Triticale Biotic Stresses—Known and Novel Foes. In Triticale; Eudes, F., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 83–108. [Google Scholar]
- Tian, S.; Weinert, J.; Wolf, G.A. Infection of triticale cultivars by Puccinia striiformis: First report on disease severity and yield loss/Blatt-und Äenbefall sowie Ertragsverluste durch Puccinia striiformis in Triticale-Sorten. Z. Für Pflanzenkrankh. Und Pflanzenschutz/J. Plant Dis. Prot. 2004, 111, 461–464. [Google Scholar]
- McIntosh, R.; Yamazaki, Y.; Devos, K.M.; Dubcovsky, J.; Rogers, W.J.; Appels, R. Catalogue of Gene Symbols for Wheat; Akadémiai Kiadó: Budapest, Hungary, 2003; Volume 49. [Google Scholar]
- Adonina, I.G.; Orlovskaya, O.A.; Tereshchenko, O.Y.; Koren, L.V.; Khotyleva, L.V.; Shumny, V.K.; Salina, E.A. Development of commercially valuable traits in hexaploid triticale lines with Aegilops introgressions as dependent on the genome composition. Russ. J. Genet. 2011, 47, 453–461. [Google Scholar] [CrossRef]
- Kwiatek, M.; Belter, J.; Majka, M.; Wiśniewska, H. Allocation of the S-genome chromosomes of Aegilops variabilis Eig. carrying powdery mildew resistance in triticale (×Triticosecale Wittmack). Protoplasma 2016, 253, 329–343. [Google Scholar] [CrossRef]
- Kwiatek, M.; Majka, M.; Wiśniewska, H.; Apolinarska, B.; Belter, J. Effective transfer of chromosomes carrying leaf rust resistance genes from Aegilops tauschii Coss. into hexaploid triticale (X Triticosecale Witt.) using Ae. tauschii × Secale cereale amphiploid forms. J. Appl. Genet. 2015, 56, 163–168. [Google Scholar] [CrossRef]
- Salmanowicz, B.P.; Langner, M.; Wiśniewska, H.; Apolinarska, B.; Kwiatek, M.; Błaszczyk, L. Molecular, physicochemical and rheological characteristics of introgressive Triticale/Triticum monococcum ssp. monococcum lines with wheat 1D/1A chromosome substitution. Int. J. Mol. Sci. 2013, 14, 15595–15614. [Google Scholar]
- Kang, H.; Wang, H.; Huang, J.; Wang, Y.; Li, D.; Diao, C.; Zhu, W.; Tang, Y.; Wang, Y.; Fan, X.; et al. Divergent Development of Hexaploid Triticale by a Wheat–Rye–Psathyrostachys huashanica Trigeneric Hybrid Method. PLoS ONE 2016, 11, e0155667. [Google Scholar] [CrossRef]
- Schneider, A.; Molnár, I.; Molnár-Láng, M. Utilisation of Aegilops (goatgrass) species to widen the genetic diversity of cultivated wheat. Euphytica 2008, 163, 1–19. [Google Scholar] [CrossRef]
- Prażak, R.; Paczos-Grzęda, E. Gene transfer from Aegilops kotschyi Boiss. to Triticum aestivum L. Adv. Agric. Sci. Probl. 2010, 555, 613–620. [Google Scholar]
- Rawat, N.; Neelam, K.; Tiwari, V.K.; Randhawa, G.S.; Friebe, B.; Gill, B.S.; Dhaliwal, H.S. Development and molecular characterization of wheat-Aegilops kotschyi addition and substitution lines with high grain protein, iron, and zinc. Genome 2011, 54, 943–953. [Google Scholar] [CrossRef]
- Antonov, A.I.; Marais, G.F. Identification of leaf rust resistance genes in Triticum species for transfer to common wheat. S. Afr. J. Plant Soil 1996, 13, 55–60. [Google Scholar] [CrossRef]
- Marais, G.F.; McCallum, B.; Snyman, J.E.; Pretorius, Z.A.; Marais, A.S. Leaf rust and stripe rust resistance genes Lr54 and Yr37 transferred to wheat from Aegilops kotschyi. Plant Breed. 2005, 124, 538–541. [Google Scholar] [CrossRef]
- Heyns, I.; Pretorius, Z.A.; Marais, F. Derivation and Characterization of Recombinants of the Lr54/Yr37 Translocation in Common Wheat. Open Plant Sci. J. 2011, 5, 1–8. [Google Scholar] [CrossRef]
- Smit, C. Pyramiding of Novel Rust Resistance Genes in Wheat, Utilizing Marker Assisted Selection and Doubled Haploid Technology. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2013. RSA. [Google Scholar]
- Qi, L.; Friebe, B.; Zhang, P.; Gill, B.S. Homoeologous recombination, chromosome engineering and crop improvement. Chromosome Res. 2007, 15, 3–19. [Google Scholar] [CrossRef]
- Sears, E. Misdivision of univalents in common wheat. Chromosoma 1950, 4, 535–550. [Google Scholar] [CrossRef]
- Sánchez-Monge, E. Two Types of Misdivision of the Centromere. Nature 1950, 165, 80–81. [Google Scholar] [CrossRef]
- Friebe, B.; Zhang, P.; Linc, G.; Gill, B.S. Robertsonian translocations in wheat arise by centric misdivision of univalents at anaphase I and rejoining of broken centromeres during interkinesis of meiosis II. Cytogenet. Genome Res. 2005, 109, 293–297. [Google Scholar] [CrossRef]
- Robertson, W.R.B. Chromosome studies. I. Taxonomic relationships shown in the chromosomes of tettigidae and acrididae: V-shaped chromosomes and their significance in acrididae, locustidae, and gryllidae: Chromosomes and variation. J. Morph. 1916, 27, 179–331. [Google Scholar] [CrossRef]
- Lukaszewski, A.J.; Gustafson, J.P. Translocations andmodifications of chromosomes in triticale × wheat hybrids. Theor. Appl. Genet. 1983, 64, 239–248. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Further manipulation by centric misdivision of the 1RS.1BL translocation in wheat. Euphytica 1997, 94, 257–261. [Google Scholar] [CrossRef]
- Qi, L.L.; Pumphrey, M.O.; Friebe, B.; Zhang, P.; Qian, C.; Bowden, R.L.; Rouse, M.N.; Jin, Y.; Gill, B.S. A novel Robertsonian translocation event leads to transfer of a stem rust resistance gene (Sr52) effective against race Ug99 from Dasypyrum villosum into bread wheat. Theor. Appl. Genet. 2011, 123, 159–167. [Google Scholar] [CrossRef]
- Türkösi, E.; Darko, E.; Rakszegi, M.; Molnár, I.; Molnár-Láng, M.; Cseh, A. Development of a new 7BS.7HL winter wheat-winter barley Robertsonian translocation line conferring increased salt tolerance and (1,3;1,4)-β-D-glucan content. PLoS ONE 2018, 13, e0206248. [Google Scholar]
- Lukaszewski, A.J.; Curtis, C.A. Transfer of the Glu-D1 gene fromchromosome 1D of bread wheat to chromosome 1R in hexaploidtriticale. Plant Breed. 1992, 109, 203–210. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Manipulation of the 1RS.1BL translocation in wheat by induced homoeologous recombination. Crop Sci. 2000, 40, 216–225. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Cytogenetically engineered rye chromosomes 1R to improve bread-making quality of hexaploid triticale. Crop Sci. 2006, 46, 2183–2194. [Google Scholar] [CrossRef]
- Kwiatek, M.T.; Wiśniewska, H.; Ślusarkiewicz-Jarzina, A.; Majka, J.; Majka, M.; Belter, J.; Pudelska, H. Gametocidal Factor Transferred from Aegilops geniculata Roth Can Be Adapted for Large-Scale Chromosome Manipulations in Cereals. Front. Plant Sci. 2017, 8, 409. [Google Scholar] [CrossRef]
- Heckmann, S.; Jankowska, M.; Schubert, V.; Kumke, K.; Ma, W.; Houben, A. Alternative meiotic chromatid segregation in the holocentric plant Luzula elegans. Nat. Commun. 2014, 5, 4979. [Google Scholar] [CrossRef]
- Zwierzykowski, Z.; Zwierzykowska, E.; Taciak, M.; Jones, N.; Kosmala, A.; Krajewski, P. Chromosome pairing in allotetraploid hybrids of Festuca pratensis x Lolium perenne revealed by genomic in situ hybridization (GISH). Chromosome Res. 2008, 16, 575–585. [Google Scholar] [CrossRef]
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome 2013, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, M.T.; Kurasiak-Popowska, D.; Mikołajczyk, S.; Niemann, J.; Tomkowiak, A.; Weigt, D.; Nawracała, J. Cytological markers used for identification and transfer of Aegilops spp. chromatin carrying valuable genes into cultivated forms of Triticum. Comp. Cytogenet. 2019, 13, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Ardalani, S.; Mirzaghaderi, G.; Badakhshan, H. A Robertsonian translocation from Thinopyrum bessarabicum into bread wheat confers high iron and zinc contents. Plant Breed. 2016, 135, 286–290. [Google Scholar] [CrossRef]
- Tanaka, H.; Nabeuchi, C.; Kurogaki, M.; Garg, M.; Saito, M.; Ishikawa, G.; Nakamura, T.; Tsujimoto, H. A novel compensating wheat–Thinopyrum elongatum Robertsonian translocation line with a positive effect on flour quality. Breed. Sci. 2017, 67, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Danilova, T.V.; Friebe, B.; Gill, B.S.; Poland, J.; Jackson, E. Development of a complete set of wheat–barley group-7 Robertsonian translocation chromosomes conferring an increased content of β-glucan. Theor. Appl. Genet. 2018, 131, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Vega, J.M.; Feldman, M. Effect of the pairing gene Ph1 on centromere misdivision in common wheat. Genetics 1998, 148, 1285–1294. [Google Scholar] [PubMed]
- Steinitz-Sears, L.M. Somatic Instability of Telocentric Chromosomes in Wheat and the Nature of the Centromere. Genetics 1966, 54, 241–248. [Google Scholar]
- Ghazali, S.; Mirzaghaderi, G.; Majdi, M. Production of a novel Robertsonian translocation from Thinopyrum bessarabicum into bread wheat. Cytol. Genet. 2015, 49, 378–381. [Google Scholar] [CrossRef]
- Devos, K.M.; Atkinson, M.D.; Chinoy, C.N.; Francis, H.A.; Harcourt, R.L.; Koebner, R.M.D.; Liu, C.J.; Masojć, P.; Xie, D.X.; Gale, M.D. Chromosomal rearrangements in the rye genome relative to that of wheat. Theor. Appl. Genet. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Friebe, B.; Hatchett, J.H.; Sears, R.G.; Gill, B.S. Transfer of Hessian fly resistance from ‘Chaupon’ rye to hexaploid wheat via a 2BS/2RL wheat-rye chromosome translocation. Theor. Appl. Genet. 1990, 79, 385–389. [Google Scholar] [CrossRef]
- Rahmatov, M.; Rouse, M.N.; Nirmala, J.; Danilova, T.; Friebe, B.; Steffenson, B.J.; Johansson, E. A new 2DS•2RL Robertsonian translocation transfers stem rust resistance gene Sr59 into wheat. Theor. Appl. Genet. 2016, 129, 1383–1392. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
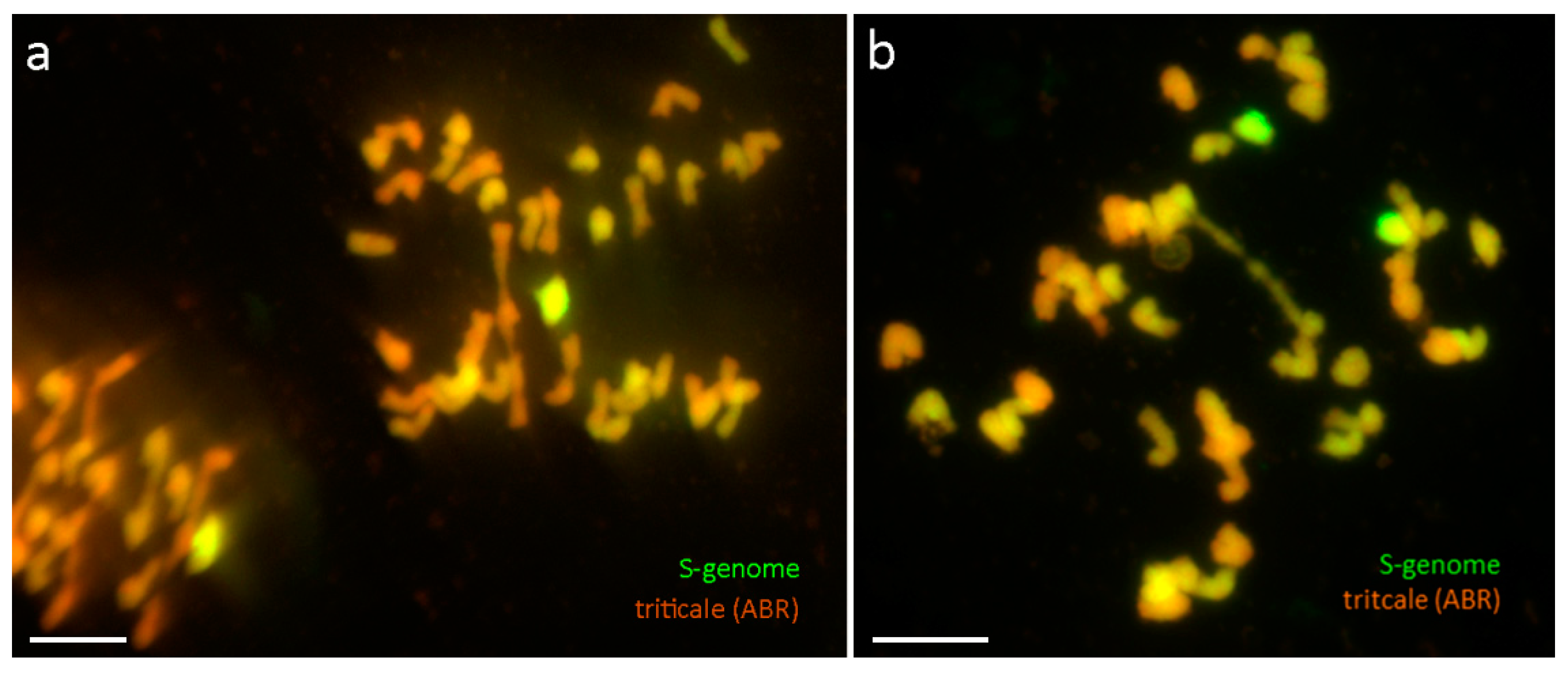{kind=link}
{kind=link}
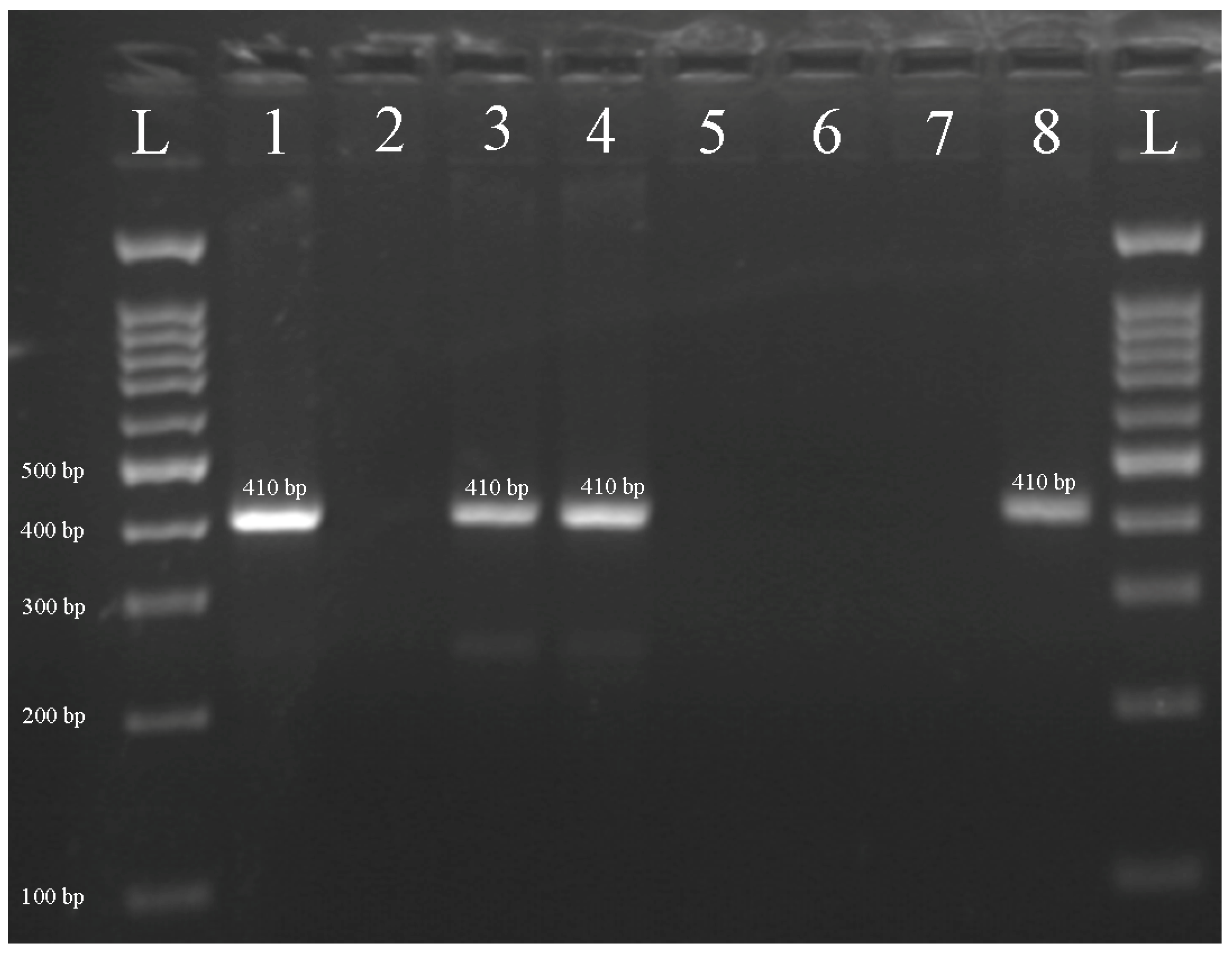{kind=link}
| Marker | Primer Sequence (5′ to 3′) | Amplification Temperature (°C) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| S14-297 | CATGCAGAAAACGACACACC | 60 | 297 | [21] |
| GGTAAGTGGTCAGGCGTTGT | ||||
| S14-410 | ACCAATTCAACTTGCCAAGAG | 61 | 410 | [22] |
| GAGTAACATGCAGAAAACGACA |
| Plant No. | Chromosome Constitution | Chromosome Number | Amplicon Size for S14-410 Marker (bp) | Amplicon Size for S14-297 Marker (bp) |
|---|---|---|---|---|
| 1 | 40 + M2RS + M2RL + M2SkS | 43 | null | null |
| 2 | 40 + T2RS.2SkL + M2RL | 42 | 410 | 297 |
| 3 | 40 + T2SkS.2RL | 41 | null | null |
| 4 | 40 + T2RS.2SkL | 41 | 410 | 297 |
| 5 | 40 + T2RS.2SkL + M2RL | 42 | 410 | 297 |
| 6 | 40 + T2RS.2SkL + t2SkS.2RL | 42 | 410 | 297 |
| 7 | 40 + T2SkS.2RL | 41 | null | null |
| 8 | 40 + T2RS.2SkL | 41 | 410 | 297 |
| 9 | 40 + M2RS + M2RL + M2SkS | 43 | null | null |
| 10 | 40 + T2RS.2SkL | 41 | 410 | 297 |
| 11 | 40 + T2SkS.2RL + M2RS | 42 | null | null |
| 12 | 40 + T2RS.2SkL + M2RL | 42 | 410 | 297 |
| 13 | 40 + T2SkS.2RL | 41 | null | null |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulaszewski, W.; Belter, J.; Wiśniewska, H.; Szymczak, J.; Skowrońska, R.; Phillips, D.; Kwiatek, M.T. Recovery of 2R.2Sk Triticale-Aegilops kotschyi Robertsonian Chromosome Translocations. Agronomy 2019, 9, 646. https://doi.org/10.3390/agronomy9100646
Ulaszewski W, Belter J, Wiśniewska H, Szymczak J, Skowrońska R, Phillips D, Kwiatek MT. Recovery of 2R.2Sk Triticale-Aegilops kotschyi Robertsonian Chromosome Translocations. Agronomy. 2019; 9(10):646. https://doi.org/10.3390/agronomy9100646
Chicago/Turabian StyleUlaszewski, Waldemar, Jolanta Belter, Halina Wiśniewska, Joanna Szymczak, Roksana Skowrońska, Dylan Phillips, and Michał T. Kwiatek. 2019. "Recovery of 2R.2Sk Triticale-Aegilops kotschyi Robertsonian Chromosome Translocations" Agronomy 9, no. 10: 646. https://doi.org/10.3390/agronomy9100646